YBOP-OPMERKINGS: Die Coolidge-effek is agter die krag van internetporno. Die Coolidge-effek is 'n verskynsel wat by soogdierspesies gesien word, waardeur mans (en in 'n mindere mate vroulike diere) hernude seksuele belangstelling toon as hulle aan nuwe ontvanklike seksmaats bekendgestel word, selfs nadat hulle seks van vorige, maar steeds beskikbare seksmaats geweier het. Seksuele nuwigheid oorheers hierdie gewoonte met hernude opgewondenheid wat veroorsaak word deur hoër dopamien. 'N Voortdurende stroom nuutheid is wat internetporn so anders maak as pornografie van die verlede.
Oorspronklike artikel, met grafieke
- Dennis F. Fiorino,
- Ariane Coury, en
- Anthony G. Phillips
- Die Joernaal van Neurowetenskap, 15 Junie 1997, 17 (12): 4849-4855;
Abstract
Die Coolidge-effek beskryf die herinitiasie van seksuele gedrag in 'n "seksueel versadigde" dier in reaksie op 'n nuwe ontvanklike maat. Gegewe die rol van die mesolimbiese dopamien (DA) stelsel in die aanvang en instandhouding van gemotiveerde gedrag, is mikrodialise gebruik om nukleus accumbens (NAC) DA-oordrag te monitor tydens copulasie, seksuele versadiging en die herinitiasie van seksuele gedrag. In ooreenstemming met vorige verslae is die aanbieding van 'n estrouse vrou agter 'n skerm en copulasie geassosieer met beduidende toenames in NAC DA efflux. Terugkeer van NAC DA konsentrasies tot basislynwaardes het saamgeval met 'n tydperk van seksuele versadiging, alhoewel konsentrasies van die DA metaboliete, dihydroksifenylaceticzuur en homovanilliensuur verhef het. Die aanbieding van 'n nuwe ontvanklike vrou agter 'n skerm het gelei tot 'n effense toename in NAC DA, wat aansienlik verhoog is tydens hernude copulasie met die nuwe vrou. Die huidige data dui daarop dat die stimulus eienskappe van 'n nuwe ontvanklike vrou kan dien om NAC DA-oordrag in 'n seksueel versadigde manlike rat te verhoog, en dit kan weer verband hou met die herinitiasie van seksuele gedrag.
Inleiding
'N Manlike rat wat tot versadiging gekopuleer het, kan aangewend word om weer te voed as die aanvanklike vrou vervang word met 'n nuwe ontvanklike vrou. Dit het bekend geword as die Coolidge-effek en is waargeneem in 'n aantal soogdierspesies (Wilson et al., 1963). Algemene faktore soos moegheid of motoriese depressie is nie voldoende om die oënskynlike toestand van seksuele versadiging te verduidelik nie, want stimuli van 'n nuwe vrou kan steeds kopulasie veroorsaak. Seksuele versadiging kan ook farmakologies in 'n wesenlike mate omgeskakel word deur die toediening van 'n verskeidenheid middels wat op verskillende neurotransmitterstelsels kan optree. Hierdie middels sluit in yohimbine, 8-OH-DPAT (Rodriguez-Manzo en Fernandez-Guasti, 1994, 1995a), nalaxone (Pfaus en Gorzalka, 1987; Rodriguez-Manzo en Fernandez-Guasti, 1995a,b), en apomorfien (Mas et al., 1995c). Alhoewel die perifere werking van hierdie middels nie uitgesluit kan word nie (bv. Adrenerge effekte op erektiele funksie), is die effekte op sentrale meganismes wat onderliggend aan seksuele versadiging is, voorgestel aan die hand van selektiewe sentrale noradrenergiese letsel eksperimente (Rodriguez-Manzo en Fernandez-Guasti, 1995a) en mikrodialise eksperimente wat dopaminerge metabolisme gemonitor het in die mediale preoptiese area (Mas et al., 1995a,b).
Aangesien sentrale meganismes die herinitiasie van seksuele gedrag kenmerkend van die Coolidge-effek kan bemiddel, is 'n waarskynlike kandidaat die mesolimbiese dopamien (DA) stelsel wat uit die ventrale tegmentale area na die NAC uitsteek. Mesolimbic DA lyk as 'n primêre modulator op te tree in komplekse integratiewe prosesse wat die evaluering van omgewingsprikkel insluit, soos leidrade van 'n seksueel ontvanklike vrou en die organisasie van doelgerigte gedrag, insluitend copulasie (Fibiger en Phillips, 1986; Blackburn et al., 1992; Phillips et al., 1992; LeMoal, 1995; Salamone, 1996).
Alhoewel middelbrein-DA neurone reageer op primêre belonings en leidrade wat voorspelbaar is van beloning, nuwe of onvoorspelbare omgewingsprikkels veroorsaak dat neuronale aktivering die sterkste oor herhaalde oefensessies (Fabre et al., 1983; Schultz, 1992; Mirenowicz en Schultz, 1994). Daar is baie bewyse wat 'n belangrike fasiliterende rol vir mesolimbiese DA ondersteun in die inisiëring en instandhouding van seksuele gedrag in die rat (Pfaus en Everitt, 1995), en 'n aantal mikrodialise-studieverslag verhoog toename in NAC DA-effek tydens die aptyt- en verbruiksfases van manlike seksuele gedrag (Pfaus et al., 1990; Pleim et al., 1990; Damsma et al., 1992; Wenkstern et al., 1993; Fumero et al., 1994; Mas et al., 1995b). Daar is egter relatief min data oor die neurochemiese korrelate van seksuele versadiging en die herinitiasie van seksuele gedrag. Die toepassing van in vivo mikrodialise om mesolimbiese DA neurotransmissie tydens die Coolidge-effek te monitor, bied 'n unieke geleentheid om die rol van NAC DA in kopulasie, seksuele versadiging en die herinitiasie van kopulasie te ondersoek.
'N Mikrodialise-eksperiment is uitgevoer om die volgende te bepaal: (1) of die aanvang van seksuele versadiging gepaard gaan met die terugkeer van ekstracellulêre DA konsentrasies in die NAC tot presopulasie waardes of hieronder, en (2) of die herinstelling van kopulatoriese gedrag in 'n " seksueel versadigde "manlike rat met 'n nuwe ontvanklike vrou is gekorreleer met toenames in NAC DA efflux.
MATERIAAL EN METODES
Vakke. Manlike Sprague Dawley-rotte, verkry uit die Diereversorgingsentrum (aan die Universiteit van British Columbia), en vroulike Long Evans-rotte, verkry uit Charles River Canada (St Constant, Quebec, Kanada), is gehuisves in gaashokke (18 × 25 × 65 cm, vyf per hok) in aparte kolonie kamers. Koloniekamers is gehandhaaf teen 'n temperatuur van ~20 ° C op 'n omgekeerde 12 uur lig / donker siklus. Rotte het onbeperkte toegang tot kos (Purina Rat Chow) en water.
Chirurgie en gedragstoetsing voor brein mikrodialise.Vroulike rotte was ten minste 4 weke voor toetsing tweedens 10 weke bilateraal onder halotaan-gas-narkose (Fluothane, Ayerst Laboratories). Seksuele ontvanklikheid in die stimulusvroue is voor elke toets sessie geïnduceerd deur subkutane inspuitings van estradiolbenzoaat (500 μg) en progesteroon (48 μg), 4 en 4 uur. Manlike rotte is by twee geleenthede gesif vir seksuele gedrag, 35 d uitmekaar, in pleksuskamerkamers (35 × 40 × 5 cm) met gaasvloere. Slegs manlike rotte wat 'n prestasie maatstaf bereik het, wat intromissie binne 15 insluit, min van die voorkoms van die vroulike en ejakulasie binne XNUMX min van die eerste intromissie, tydens die twee siftingstoetse, is geïmplanteer met mikrodialise sonde geleidingskanale.
Manlike rotte (n = 5) was narkose met ketamienhidrochloried (100 mg / kg, ip) en xylazine (10 mg / kg, ip) voor stereotaksiese chirurgie. Mikrodialise sonde geleide kanule (19 gauge) is geïmpareer bilateraal oor die NAC (koördinate van bregma: anterior, + 1.7 mm, mediaal, ± 1.1 mm, ventraal, -1.0 mm, plat skedel) en is aan die skedel vasgemaak met tandheelkundige akriel en juwelier se skroewe. Bilaterale geleek kanulale inplantings is gebruik om die geleentheid vir 'n suksesvolle mikrodialise-eksperiment te maksimeer. Gelukkig, in die huidige eksperiment, was slegs een kanul nodig vir elke rat. Manlike rotte is individueel in groot plastiekhokke met koringbeddegoed gehuisves vir die res van die eksperiment. Een week na die operasie is rotte getoets vir seksuele gedrag. Gedurende hierdie gedeelte van die opleiding was die toetskamer toegerus met 'n glyplastiglasskerm wat die kamer in groot en klein kompartemente verdeel het. Manlike rotte is later in die groot kompartement ingevoer en 15 min later is 'n wyfie agter die skerm geplaas. Na 'n 15 min voorbereidende tydperk, is die skerm verwyder, en die rotte mag vir 30 min kopieer. Drie oefensessies is uitgevoer, een elke 4 d. Alle rotte het gedurende elke sessie die prestasiekriterium bereik.
Coolidge effek eksperiment. Rotte is eenmalig geïmplanteer met mikrodialise probes 12-18 uur voor die Coolidge effek-eksperiment en in die groot kompartement van die toetskamer geplaas met vrye toegang tot voedsel en water. Op die oggend van die eksperiment is mikrodialise monsters elke 15 min versamel. Die eksperiment het bestaan uit die volgende sewe opeenvolgende fases: (1) basislyn (ten minste 60 min); (2) vroulike 1 agter die skerm (15 min); (3) kopulasie met vroulike 1 tot 'n 30-minperiode sonder 'n berg geslaag het; (4) herinvoering van vroulike 1 agter die skerm (15 min); (5) toegang tot vroulike 1 vir 'n 15-tydperk, mits daar geen montering was nie (indien montering plaasgevind het, is hierdie fase behandel as fase 3); (6) bekendstelling van vroulike 2 agter die skerm (15 min); 7) kopulasie met vroulike 2 vir 60 min.
Gedrag is verfilm onder lae verligting met 'n JVC-videosisteem en waargeneem op 'n videomonitor buite die toetskamer. Standaard maatreëls van seksuele gedrag is aangeteken met behulp van 'n rekenaar en toepaslike sagteware (Holmes et al., 1987).
Na die mikrodialise-eksperiment het diere 'n oordosis chloorhidraat gegee en intrakardiaal met sout en formalien (4%) geperuseer. Breine is gesny en bevrore, en daarna is koronale afdelings gevlek met cresylviolet om die plasing van mikrodialise probes te bepaal. Slegs rotte met sondeplasings binne die NAC is gebruik vir gedrags- en neurochemiese ontledings.
Mikrodialise en HPLC-elektrochemiese opsporing. Mikrodialise probes was konsentries in ontwerp met 'n semipermeabele holveselmembraan (2 mm membraan blootgestel, 340 μm buitenste deursnee, 65000 molekulêre gewig cutoff, Filtral 12, Hospal) aan die distale einde. Probes is geperfumeer teen 1.0 μl / min met 'n gemodifiseerde Ringer se oplossing (0.01 m natriumfosfaatbuffer, pH 7.4, 1.3 mmCaCl2, 3.0 mm KCl, 1.0 mmMgCl2, 147 mm NaCl) met behulp van 'n gastight spuit (Hamilton, Reno, NV) en 'n spuitpomp (model 22, Harvard Apparatus, South Natick, MA). 'N Mikrodialise sonde geleiding kraag is gebruik om die microdialysis sonde in die gids cannula te beveilig. 'N Staalspoel, vasgemaak aan 'n vloeibare draaibank (Instech 375s) wat bo-op die toetskamer gemonteer is, is gebruik om die sondebuis te beskerm (Fiorino et al., 1993).
Mikrodialisaat analise, wat DA en sy metaboliete dihyroxifenylacetic acid (DOPAC) en homovanilliensuur (HVA) ingesluit het, is geskei deur omgekeerde fase chromatografie (Ultrasphere kolom; Beckman, Fullerton, CA, ODS 5 μm, 15 cm, 4.6 mm, binnendiameter ) met 'n 0.083m natriumasetaatbuffer, pH 3.5 (5% methanol). Analise konsentrasies is gekwantifiseer deur elektrochemiese (EC) opsporing. Die toestel bestaan uit 'n Bio-Rad (Richmond, CA) pomp, 'n Valco Instruments (Houston, TX) EC10W twee-posisie inspuiting, 'n ESA (Bedford, MA) Coulochem II EC detektor en 'n tweekanaal-kaartopnemer en Zonen, Bohemia, NY). Elektrochemiese detektor parameters was die volgende: elektrode 1, + 450 mV; elektrode 2, -300 mV; en wag sel, -450 mV. Tipiese sonde herwinnings, uitgevoer vitro en by kamertemperatuur was 22% vir DA, 18% vir DOPAC, en 18% vir HVA.
RESULTATE
Gedrag
Gedragsmaatreëls uit die Coolidge effek-eksperiment word in Tabel aangebied 1. Latsies aan die berg, intromiet, en ejakulasie sowel as die postejaculatoriese interval na die eerste ejakulasie was soortgelyk aan dié van die vorige oefensessie (data nie getoon nie). Dit dui daarop dat die mikrodialise prosedure nie normale seksuele gedrag verander het nie. Die ontwikkeling van seksuele versadiging, soos aangedui deur die gemiddelde aantal ejakulasies voor die kriterium, is nagekom (7.8 ± 0.5), 'n progressiewe afname in die aantal intromissies wat voorafgaan aan elke ejakulasie en 'n progressiewe toename in die postjakulatoriese interval (data nie getoon nie) , was soortgelyk aan dié wat in vorige studies gerapporteer is (Strand en Jordanië, 1956; Fowler and Whalen, 1961; Fisher, 1962; Bermant et al., 1966; Rodriguez-Manzo en Fernandez-Guasti, 1994; Mas et al., 1995d). Individuele veranderlikheid is waargeneem met betrekking tot die aantal ejakulasies wat met vroulike 1 behaal is, die tyd wat bestee word om met vroulike 1 te kopieer, en die aantal aanbiedings van vroulike 1 wat nodig is om die versadigingskriterium te bereik (tabel 1, onder). Sommige rotte benodig talle herinvoerings van vroulike 1 tot fase 5 voltooi is (n = 3). Die dade van die plasing van vroulike 1 agter die skerm en die verwydering van die partisie mag as primêre appetitiewe leidrade gedien het wat tot copulasie lei. Daar moet ook op gelet word dat 'n versadigingskriterium van 30 min sonder 'n berg, hoewel dit voorheen gebruik is (Beach and Jordan, 1965; Mas et al., 1995b), is arbitrêr en waarborg nie dat 'n rot nie gemonteer sal wees nie, aangesien meer tyd gegee word. Tog het vertragings of verwydering en vervangingsprosedures nie betroubaar in hernude copulasie met vroulike 1 (bv. Fases 4 en 5) opgelewer nie.

Gedrag tydens die Coolidge effek eksperiment
Alle rotte vertoon die Coolidge-effek. Die aktiwiteit wat verband hou met die plasing van vroulike 2 agter die skerm, en veral die verwydering van die partisie, kan bygedra het tot hierdie uitslag, maar weer was hierdie gebeurtenisse nie voldoende om kopering vroeger in die eksperiment te vernuwe nie. Vergelykings tussen maatreëls van seksuele gedrag met vroulike 1 en vroulike 2 is gemaak deur gebruik te maak van t toetse met 'n Bonferroni-regstelling. Alhoewel die berg- en intromissie-laatstande in reaksie op vroulike 2 nie betekenisvol verskil van dié van die eerste copulatoriese bout met vroulike 1 nie, was seksuele gedrag met vroulike 2 in die algemeen minder robuust, soos aangedui deur aansienlik minder ejakulasies (gemiddelde 0.6 vs 4.2; F = 49.86;p <0.01) en intromsies (gemiddelde = 11.2 vs 37.0;F = 20.17; p <0.05) gedurende die eerste uur. Die aantal monteerders in die eerste uur met wyfies 1 en 2 was nie betekenisvol nie.
Dit is belangrik om daarop te let dat wyfies wat tydens die versadigingsgedeelte van die eksperiment gebruik is (bv. Vroulike 1) steeds sterk voornemende (dws hopping en darting) en ontvanklike (dws lordose) gedrag vir die volledige duur van hul kontak met die mannetjie .
neurochemie
Basale nanomolêre konsentrasies van DA en sy metaboliete in mikrodialisate, aangebied as die gemiddelde ± SEM, van die eerste drie baseline monsters was: DA, 3.0 ± 0.7; DOPAC, 619.1 ± 77.7; en HVA, 234.2 ± 49.0 (ongeregverdig vir sonde herstel;n = 5). Hierdie waardes verteenwoordig 100% basislyn tellings.
Gedrags-gedefinieerde data punte, wat ooreenstem met elke fase van die eksperiment en algemeen by elke rot, is gebruik vir neurochemiese ontledings. Dit sluit die volgende in: (1) sewe monsters na die eerste bekendstelling van vroulike 1, (2) vier monsters wat die afwesigheid van kopulatoriese gedrag met vroulike 1, en (3) vyf monsters vergesel na die aanbieding van vroulike 2. Figuur 1 illustreer veranderinge in konsentrasies van DA (lyngrafiek, middel) en DA metaboliete (lyngrafiek, Top) parallel copulatoriese gedrag (staafgrafiek, onderkant) tydens die toets vir die Coolidge-effek.
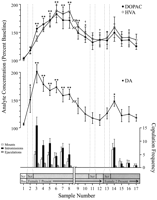
Bekyk groter weergawe:
Fig. 1.
Kern verleen neurochemiese korrelate van seksuele gedrag tydens die Coolidge-effek. Die eerste agt monsters verteenwoordig chronologies deurlopende data punte vanaf fases 1 tot 3. Voorbeeld 1 is die vierde en laaste presopulasie basislyn monsters (Bas). Voorbeeld 2 verteenwoordig die bekendstelling van vroulike 1 agter die skerm (BSG). Na 15 min is die skerm verwyder, en rotte word toegelaat om te kopieer (monsters 3-8). Diebreek op die x-Axis stem ooreen met die uitsluiting van data van drie rotte wat vir lang periodes saam met die aanvanklike vroulik gekopuleer het. Die laaste nege monsters was ook deurlopend chronologies. Monsters 9 en 10 stem ooreen met die versadigingsperiode van fase 3 (dws 30 min sonder 'n berg). Vroulike 1 is dan weer agter die skerm ingeplaas (monster 11) en, 15 min later, is die skerm verwyder (monster 12). Na 15 min sonder copulasie is vroulike 2 agter die skerm geplaas (monster 13). Monsters 14-17 stem ooreen met kopulasie met vroulike 2. Die aantal mounts, intromissies of ejakulasies wat met elke 15 min mikrodialise-monster verband hou, word in die onderste staafgrafiek. Neurochemiese data word uitgedruk in terme van persentasie baseline konsentrasies. Veranderinge in NAK DA (geslote vierkante), DOPAC (geslote sirkels), en HVA (oop kringe) efflux word aangebied as lyngrafieke. Die volgende vergelykings is gemaak: basislynmonster 1 teenoor monsters 2-10; Nuwe basislynmonster 10 teenoor monsters 11 en 12; nuwe basislynmonster 12 teenoor monsters 13-17 (*p <0.05; ** p <0.01). Onafhanklik t toetse is gemaak tussen basislynwaardes (monsters 1, 10, en 12). Vir beduidende verskille vanaf die eerste basislyn (monster 1), †p <0.05.
Afsonderlike eenrigting, herhaalde-maat ANOVA's, is uitgevoer op neurochemiese data wat geassosieer word met vroulike 1 (monsters 1-12) en vroulike 2 (monsters 12-17). A priori vergelykings is gemaak met behulp van Dunn se meervoudige vergelykingstoets (Bonferroni t). Die volgende drie hoofvergelykings is gemaak: (1) aanvanklike basislyn (monster 1) teenoor monsters 2-10 (eerste blootstelling aan vroulike 1), (2) tweede basislyn (monster 10) teenoor monsters 11 en 12 (herexposure na vroulike 1) , en (3) derde basislyn (monster 12) teenoor monsters 13-17 (blootstelling aan vroulike 2).
Daar was 'n beduidende algehele verandering in DA-effek in reaksie op vroulike 1 [F (11,44) = 8.48; p <0.001] en vroulik 2 [F (5,20) = 2.83;p <0.05]. 'N Beduidende toename in DA-uitvloei is gevind toe vroulike 1 agter die skerm was (+ 44%,p <0.05; monster 2). Tydens kopulasie het DA-konsentrasies verder toegeneem en 'n maksimum waarde bereik (+ 95%;p <0.01) tydens die eerste kopulatoriese aanval (voorbeeld 3). DA het gedurende die hele kopulasie verhoog gebly en het eers teruggekeer na die basislynkonsentrasies in die 30 minute periode waarin geen toename plaasgevind het nie (monsters 9 en 10). Die herinvoering van vroulike 1 agter die skerm (monster 11) of die geleentheid om fisies te kommunikeer, maar sonder montering (monster 12), verhoogde DA-konsentrasies in verhouding tot die tweede basislyn (monster 10). Die teenwoordigheid van vroulike 2 agter die skerm (monster 13) het gelei tot 'n klein toename in DA-uitvloei (12%) vanaf die derde basislynwaarde (monster 12) wat nie statistiese betekenisvolheid bereik het nie. Hernuwe kopulasie met vroulike 2 het gelei tot 'n beduidende toename (34%) (p <0.05) in DA-uitvloei tydens die eerste kopulasiemonster (monster 14). Alhoewel swak copulatoriese gedrag gedurende die volgende drie monsters voortgesit het, het DA-konsentrasies afgeneem tot basiswaardes (monsters 15-17). Onafhanklik t toetse wat onder "basislyn" -monsters uitgevoer is (dws 1, 10, en 12) het getoon dat hierdie waardes nie beduidend anders was nie.
In die drie rotte wat copulasie hervat het toe die vroulike 1 weer ingestel is, het NAC DA-konsentrasies toegeneem toe vroulike 1 agter die skerm (reeks 25-47%) en tydens samestelling (reeks 13-37%) teen die monster gewoon was voor die herintroduksie van die vrou. Hierdie verhogings het egter slegs plaasgevind toe seksuele gedrag sterk was en tot ejakulasie gelei het.
Beduidende algehele veranderinge in DOPAC [F (11,44) = 9.57; p <0.001] en HVA [F (11,44) = 12.47; p <0. 001] konsentrasies is gevind in reaksie op vroulike 1, maar nie vroulike 2. Metabolietkonsentrasies het effens toegeneem (+ 15% in beide gevalle) tydens die aanbieding van vroulike 1 agter die skerm (monster 2), maar dit was nie betekenisvol nie statisties. Daar was egter beduidende toenames in die konsentrasies van DOPAC en HVA tydens kopulasie (monsters 3-8), en bereik maksimum waardes (onderskeidelik +80 en + 86%; p <0.01) na 60 minute (monster 6 in beide gevalle). Alhoewel metabolietkonsentrasies afgeneem het gedurende die periode van seksuele onaktiwiteit aan die einde van kontak met vroulike 1 (monsters 9 en 10), het die konsentrasies steeds verhoog met betrekking tot die basislyn (p <0.05 in beide gevalle). Herinvoering van wyfie 1 agter die skerm (monster 11), toegang tot wyfie 1 na verwydering van die skerm (monster 12) en die bekendstelling van wyfie 2 (monster 13) het geen veranderinge in metabolietkonsentrasies tot gevolg gehad nie. Ligte, maar statisties onbeduidende, toenames in DOPAC- en HVA-konsentrasies (+ 23% in beide gevalle) relatief tot die basislyn (monster 12) stem ooreen met die eerste samestelling met vroulike 2 (monster 14). Hierdie toename was egter van korte duur en het afgeneem tot basiswaardes vir die oorblywende drie monsters (15-17). Onafhanklik ttoetse wat onder "basislyn" monsters uitgevoer is (dws 1, 10 en 12) het aangedui dat die tweede en derde basislynwaardes (monsters 10 en 12 respektiewelik), hoewel nie van mekaar verskil nie, aansienlik verhef is in vergelyking met die eerste basislynmonster vir DOPAC en HVA (p <0.05 in beide gevalle).
histologie
Mikrodialise probes was in die NAC geleë (Fig.2) in 'n reeks wat + 1.20 na + 1.70 mm van bregma (plat skedel) uitsteek. Daar was ook wisselvalligheid in die mediolaterale vlak; data weerspieël steekproefneming van die dop en kern substreke van die NAK.
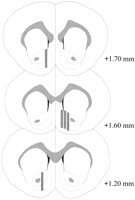
Bekyk groter weergawe:
Fig. 2.
Ligging van mikrodialise probes binne die NAC van manlike rotte wat gebruik word in die Coolidge effek eksperiment. Gekleurde reghoeke stem ooreen met die blootgestelde membraan area van die mikrodialise probes. Seriale koronale breinafdelings is teruggetrekPaxinos en Watson (1986).
BESPREKING
In ooreenstemming met vorige verslae, toon die huidige resultate verbeterde mesolimbiese DA-oordrag wat verband hou met eet- en eetkomponente van manlike rotse seksuele gedrag soos beoordeel deurin vivo mikrodialise (Mas et al., 1990; Pfaus et al., 1990;Pleim et al., 1990; Damsma et al., 1992; Wenkstern et al., 1993; Fumero et al., 1994; Mas et al., 1995a,b,d). Daarbenewens bied hierdie resultate 'n neurochemiese korrelaat vir seksuele versadiging en die daaropvolgende herinitiasie van copulasie in reaksie op 'n nuwe ontvanklike vrou (die Coolidge-effek). Die huidige data dui daarop dat die stimulus eienskappe van 'n nuwe ontvanklike vrou kan dien om NAC DA-oordrag te verhoog in 'n seksueel versadigde manlike rat wat op sy beurt verband hou met die herinitiasie van seksuele gedrag. Dit is die eerste keer in die ligte toename in NAC DA tydens die aanbieding van 'n nuwe vrou agter die skerm en vind die meeste oortuigend plaas as 'n groter toename tydens hernude copulasie met vroulike 2 (Fig.1).
Die teenwoordigheid van die eerste ontvanklike vrou agter die skerm het gelei tot 'n robuuste aptytverhoging in NAC DA efflux (44% vanaf basislyn) soortgelyk in grootte as wat in vorige eksperimente gerapporteer is deur 'n soortgelyke ontwerp te gebruik (30% Pfaus et al., 1990; 35%,Damsma et al., 1992). In ooreenstemming met hierdie studies was ook die waarneming dat die NAC DA-uitvloei tydens die samestelling verder verbeter is (tot> 95% bo die basislyn in die huidige eksperiment). Alhoewel ons die voltooide gedrag kan geassosieer met 'n verbeterde vrystelling van DA-DA (Wenkstern et al., 1993; Wilson et al., 1995), is dit belangrik om die terme "appetitief" en "verbruikend" in die konteks van seksuele gedrag te ondersoek. Terwyl die fase waarin die wyfie agter die skerm teenwoordig is, uitsluitlik aptyt of voorbereidend is, kan die gedrag gedurende die copulasiefase nie suiwer verbruiksmatig beskou word nie. Aangesien "appetitief" gebruik kan word om alle gedrag te beskryf wat tot die voortsetting van 'n gemotiveerde gedrag lei (kopulasie), word die primêre gedrag wat die mannetjie uitoefen terwyl dit aktief is in die "consummatic" fase die beste beskryf as 'n aptyt die man spandeer die meeste van sy tyd en moeite om die vrou te volg om te copuleer. In hierdie verband kan ons die maksimum NAC DA-oordrag met verbruikswaarde korreleer sowel as intense appetitiewe komponente van manlike rot seksuele gedrag.
Toegang tot die tweede, nuwe vrou het gelei tot vernuwing in elke vak. Vorige studies het getoon dat die meerderheid rotte toegelaat word om na versadiging te kopieer, met behulp van 'n soortgelyke gedragsprotokol vir die een wat in die huidige eksperiment gebruik is, het nie paring hervat wanneer dit later 24 uur getoets is nie (Strand en Jordanië, 1956). Dit is waarskynlik dat die teenwoordigheid van die nuwe stimulus eienskappe van vroulike 2, wat oraal sowel as visuele en ouditiewe leidrade ingesluit het, tot hernude samestelling gelei het. 'N Interessante vraag wat nog beantwoord moet word, is deur watter meganisme 'n manlike rot onderskei van 'n nuwe vrou van 'n vrou wat hy onlangs ontmoet het. 'N Plek vir daardie meganisme kan in die hoof-olfaktoriese stelsel lê. Daar is gerapporteer dat die integriteit van hierdie stelsel noodsaaklik is vir die Coolidge-effek in hamsters (Johnston en Rasmussen, 1984). Die vomeronasal-toebehore-olfaktoriese stelsel, egter, waarin 'n pheromonale geheueproses onlangs in muise beskryf is (Kaba et al., 1994), is ook 'n toonaangewende kandidaat. In hierdie verband is dit opmerklik dat die toename in NAC DA-oordrag gemeet is deur gebruik te maak in vivo voltammetrie in manlike rotte aangebied met beddegoed wat aan vroulike rotte in estrus blootgestel is (Louillot et al., 1991; Mitchell en Gratton, 1992). Verder, die toepassing van K+ direk na die vomeronasale senuweelag van die toebehore-olfaktoriese gloeilamp, sowel as die toebehore-olfaktoriese gloeilamp self, was voldoende om NAC DA-oordrag te verhoog (Mitchell en Gratton, 1992).
Die eerste 15 min van copulasie met vroulike 2 was geassosieer met 'n beduidende toename in NAC DA. In teenstelling met die vroulike 1, het interaksie met vroulike 2 nie toenames in NAC DA van dieselfde grootte veroorsaak gedurende die aptyt (12%) of consummate (34%) fases nie. Hierdie klein stygings in NAC DA korreleer egter goed met die verminderde seksuele gedrag wat met vroulike 2 vertoon word, in vergelyking met die vroulike 1. Metabolietkonsentrasies het gedurende die versadigingsfase verhef, wat lei tot nuwe baseline konsentrasies (monsters 10 en 12) wat aansienlik vermeerder is vanaf die aanvanklike basislynwaarde (monster 1).
Die temporale vertraging in die toename in DOPAC- en HVA-konsentrasies tydens samestelling is in ooreenstemming met hul vorming as metaboliete van die moederverbinding, DA. Daar is voorgestel dat mikrodialise metaboliet konsentrasies, ten minste tydens 'n natuurlike gedrag wat nie farmakologies aangedryf word nie, 'n nuttige indeks van neurale aktiwiteit bied (Damsma et al., 1992; Fumero et al., 1994). Die feit dat metabolietkonsentrasies selfs gedurende periodes van seksuele onaktiwiteit in hierdie eksperiment verhoed het, toe DA-konsentrasies teruggekeer het om die basislynwaardes te verbeel, twyfel aan hierdie voorstel.
Die aanhoudende verheffing van DA metaboliet konsentrasies gesien in hierdie eksperiment weerspieël die mediale preoptiese area (mPOA) profiel van DA metaboliete waargeneem in rotte die eerste dag nadat hulle na versadiging (Mas et al., 1995a,b). Volgehoue verhogings van DOPAC- en HVA-konsentrasies in die NAC of mPOA word nie altyd waargeneem wanneer die paringstydperk van 'n vaste duur is nie, veel korter as die tyd wat nodig is om saturasie te bereik. Byvoorbeeld, baie studies het getoon dat DOPAC konsentrasies toegeneem het en gedurende die kopulasie verhef bly, maar geweier het om basislynwaardes te verval kort nadat die wyfie verwyder is (Pfaus et al., 1990; Pleim et al., 1990; Damsma et al., 1992;Hull et al., 1993; Wenkstern et al., 1993; Hull et al., 1995). In die studie deur Mas et al. (1995b), was basale ekstrasellulêre konsentrasies van DOPAC en HVA in die mPOA verhef oor 4 opeenvolgende dae wat ooreenstem met 'n tydperk van seksuele onaktiwiteit. Teen die vierde dag het die basale konsentrasies van die metaboliete net voor die diere kopulasie hervat, naby aan presatiwiteitswaardes. Die skrywers het die patroon van neurochemiese veranderinge vergelyk met dié wat gesien is na die toediening van DA-reseptorblokkers (Zetterström et al., 1984; Imperato en DiChiara, 1985) en het voorgestel dat die staat van seksuele onaktiwiteit bemmed kan word via prolactienvrystelling, wat kan optree as 'n "endogene neuroleptiese" (Mas et al., 1995a,b,d). Dit is duidelik dat neuroleptiese toediening gepaard gaan met toenames in ekstrasellulêre metaboliet konsentrasies en DA efflux (Zetterström et al., 1984; Imperato en DiChiara, 1985). Ongelukkig, Mas et al. (1995a,b) kon nie mPOA DA konsentrasies opspoor nie. In die huidige studie het DA konsentrasies in die NAC teruggekeer na presopulasie waardes, terwyl DOPAC en HVA konsentrasies verhef. Hierdie patroon is teenstrydig met 'n rol vir 'n endogene neuroleptiese wat in die NAC optree om seksuele versadiging te bewerkstellig.
Gegewe die betrokkenheid van mesolimbiese DA neurone in gemotiveerde gedrag (Fibiger en Phillips, 1986; Blackburn et al., 1992; Kalivas et al., 1993; LeMoal, 1995) en hul sensitiwiteit vir nuwe omgewingstimuli (Fabre et al., 1983; Schultz, 1992; Mirenowicz en Schultz, 1994), die waargenome toenames in ekstrasellulêre konsentrasies van NAC DA in reaksie op die nuwe vrou is in ooreenstemming met die hipotese dat aktiwiteit in hierdie DA-stelsel belangrik is vir die herinitiasie van seksuele gedrag. Daarbenewens is verslae van eetlustige en verbruiksverhogings in DA-oordrag (Hull et al., 1993, 1995;Mas et al., 1995b; Sato et al., 1995) en neuronale aktiwiteit (Shimura et al., 1994) in die mPOA van manlike rotte tydens seksuele gedrag dui daarop dat hierdie struktuur ook kan bydra tot hernude kopulasie eienskappe van die Coolidge effek.
In ooreenstemming met 'n algemene rol vir die mesolimbiese DA-stelsel in gemotiveerde gedrag, is dit goed gevestig dat ekstrasellulêre konsentrasies van DA ook voor, tydens en dadelik na die gebruik van 'n maaltyd verhef word, met 'n terugkeer na basislynwaardes ~30 min later (Wilson et al., 1995). Dit is bekend dat versadiging deur voedsel beïnvloed word deur sy sensoriese eienskappe. Mense en diere verwerp die voedsel waarop hulle versadig is en versadig ander kosse wat nie geëet is nie (Rolls, 1986). Dit stel die vraag of ekstrasellulêre DA-effek in die NAK selektief verhoog moet word deur die aanbieding van 'n nuwe soort kos, maar nie deur voedsel wat onlangs versadig is op versadiging op 'n manier wat analoog is aan wat in die huidige studie gerapporteer is in die konteks van seksuele motivering. As dit bevestig word, sal hierdie algemene verwantskap tussen die sensoriese eienskappe van natuurlike belonings, versadiging en mesolimbiese DA-oordrag 'n kritieke rol vir hierdie neurale stelsel in die regulering van motiveringsprosesse impliseer, wat die ontwrigting van ernstige eetversteurings en seksuele funksie kan veroorsaak .
Verwysings
- ↵
- Beach FA,
- Jordanië L
(1956) Seksuele uitputting en herstel in die manlike rat. QJ Exp Psychol 8: 121-133.
- ↵
- Bermant G,
- Lott DF,
- Anderson L
(1966) Temporale eienskappe van die Coolidge-effek in manlike ratkopulatoriese gedrag. J Comp Fisioloog Psigiatrie 65: 447-452.
- ↵
- Blackburn JR,
- Pfaus JG,
- Phillips AG
(1992) Dopamienfunksies in eetlus en verdedigende gedrag. Prog Neurobiol 39: 247-279.
- ↵
- Damsma G,
- Pfaus JG,
- Wenkstern D,
- Phillips AG,
- Fibiger HC
(1992) Seksuele gedrag verhoog dopamien oordrag in die nukleus accumbens en striatum van manlike rotte: vergelyking met nuwigheid en voortbeweging. Behav Neurosci 106: 181-191.
- ↵
- Fabre M,
- Rolls ET,
- Ashton JP,
- Williams G
(1983) Aktiwiteit van neurone in die ventrale tegmentale streek van die gedragende aap. Behav Brain Res 9: 213-235.
- ↵
- Fibiger HC,
- Phillips AG
(1986) Beloning, motivering, kognisie: psigobiologie van mesotelenfale dopamienstelsels. in die handboek van fisiologie: die senuweestelsel IV, eds Bloom FE, Geiger SD (American Physiology Society, Bethesda, MD), pp 647-675.
- ↵
- Fiorino DF,
- Coury AG,
- Fibiger HC,
- Phillips AG
(1993) Elektriese stimulasie van beloningsterreine in die ventrale tegmentale area verhoog dopamien-oordrag in die nukleusbuis van die rat. Behav Brain Res 55: 131-141.
- ↵
- Fisher A
(1962) Effekte van stimulusvariasie op seksuele versadiging in die manlike rat. J Comp Fisioloog Psigiatrie 55: 614-620.
- ↵
- Fowler H,
- Whalen RE
(1961) Variasie in aansporingstimulus en seksuele gedrag in die manlike rat. J Comp Fisioloog Psigiatrie 54: 68-71.
- ↵
- Fumero B,
- Fernendez-Vera JR,
- Gonzalez-Mora JL,
- Mas M
(1994) Veranderinge in monoamienomset in voorhoedgebiede wat geassosieer word met manlike seksuele gedrag: 'n mikrodialise-studie. Brain Res 662: 233-239.
- ↵
- Holmes GM,
- Holmes DG,
- Sachs BD
(1987) 'n databasis-databasisstelsel vir die opneem van knaagdiere en vir algemene gebeurtenisopname. Physiol Behav 44: 825-828.
- ↵
- Hull EM,
- Eaton RC,
- Moses J,
- Lorrain DS
(1993) Kopulasie verhoog dopamienaktiwiteit in die mediale preoptiese area van manlike rotte. Life Sci 52: 935-940.
- ↵
- Hull EM,
- Jianfang D,
- Lorrain DS,
- Matuszewich L
(1995) Ekstracellulêre dopamien in die mediale preoptiese area: implikasies vir seksuele motivering en hormonale beheer van kopulasie. J Neurosci 15: 7465-7471.
- ↵
- Imperato A,
- DiChiara G
(1985) Dopamien vrylating en metabolisme in wakker rotte na sistemiese neuroleptika soos bestudeer deur transstriatale dialise. J Neurosci 5: 297-306.
- ↵
- Johnston RE,
- Rasmussen K
(1984) Individuele herkenning van vroulike hamsters deur mans: rol van chemiese leidrade en van die olfaktoriese en vomeronasale stelsels. Physiol Behav 33: 95-104.
- ↵
- Kaba H,
- Hayashi Y,
- Higuchi T,
- Nakanishi S
(1994) Induksie van 'n olfaktoriese geheue deur die aktivering van 'n metabotropiese glutamaatreseptor. Wetenskap 265: 262-264.
- ↵
- Kalivas PW,
- Sorg BA,
- Hake MS
(1993) Die farmakologie en neurale stroombaan van sensitiwiteit vir psigostimulante. Behav Pharmacol 4: 315-334.
- ↵
- LeMoal M
(1995) Mesokortikolimbiese dopaminerge neurone. Funksionele en regulatoriese rolle. in Psigofarmakologie: die vierde generasie van vordering, eds Bloom FE, Kupfer DJ (Raven, New York), pp 283-294.
- ↵
- Louillot A,
- Gonzalez-Mora JL,
- Guadalupe T,
- Mas M
(1991) Seksverwante olfaktoriese stimuli veroorsaak 'n selektiewe toename in dopamien vrylating in die kernkloppers van manlike rotte. Brain Res 553: 313-317.
- ↵
- Mas M,
- Gonzalez-Mora JL,
- Louillot A,
- Sole C,
- Guadalupe T
(1990) Verhoogde dopamien vrylating in die nukleus accumbens van copulerende manlike rotte soos blyk uit in vivo voltammetry. Neurosci Lett 110: 303-308.
- ↵
- Mas M,
- Fumero B,
- Fernandez-Vera JR,
- Gonzalez-Mora JL
(1995a) Neurochemiese korrelate van seksuele uitputting en herstel soos beoordeel deur in vivo mikrodialise. Brain Res 675: 13-19.
- ↵
- Mas M,
- Fumero B,
- Gonzalez-Mora JL
(1995b) Voltammetriese en mikrodialise monitering van brein monoamien neurotransmitter vrystelling tydens sosio-seksuele interaksies. Behav Brain Res 71: 69-79.
- ↵
- Mas M,
- Fumero B,
- Perez-Rodriguez I
(1995c) Induksie van paringsgedrag deur apomorfien by seksueel-gesonde rotte. Eur J Pharmacol 280: 331-334.
- ↵
- Mas M,
- Fumero B,
- Perez-Rodriguez Ek,
- Gonzalez-Mora JL
(1995d) Die neurochemie van seksuele versadiging. 'N eksperimentele model van geïmmiteerde begeerte. in die farmakologie van seksuele funksie en disfunksie, ed Bancroft J (raaf, New York), pp 115-126.
- ↵
- Mirenowicz J,
- Schultz W
(1994) Belangrikheid van onvoorspelbaarheid vir beloningsresponse in primate dopaminerge neurone. J Neurophysiol 72: 1024-1027.
- ↵
- Mitchell JB,
- Gratton A
(1992) Mesolimbiese dopamien vrylating verkry deur aktivering van die toebehore-olfaktoriese stelsel: 'n hoë spoed chrono -perametriese studie. Neurosci Lett 140: 81-84.
- ↵
- Paxinos G,
- Watson C
(1986) Die rotbrein in stereotaksiese koördinate (2ed ed). (Akademies, San Diego).
- ↵
- Pfaus JG,
- Damsma G,
- Nomikos GG,
- Wenkstern D,
- Blaha CD,
- Phillips AG,
- Fibiger HC
(1990) Seksuele gedrag verhoog sentrale dopamien-oordrag in die manlike rat. Brain Res 530: 345-348.
- ↵
- Pfaus JG,
- Everitt BJ
(1995) Die psigofarmakologie van seksuele gedrag. in Psigofarmakologie: die vierde generasie van vordering, eds Bloom FE, Kupfer DJ (Raven, New York), pp 743-758.
- ↵
- Pfaus JG,
- Gorzalka BB
(1987) Opioïede en seksuele gedrag. Neurosci Biobehav Ds 11: 1-34.
- ↵
- Phillips AG,
- Blaha CD,
- Pfaus JG,
- Blackburn JR
(1992) Neurobiologiese korrelate van positiewe emosionele toestande: dopamien, afwagting en beloning. in Internasionale oorsig van studies oor emosie, Ed Strongman (Wiley, New York), pp 31-50.
- ↵
- Pleim ET,
- Matochik JA,
- Barfield RJ,
- Auerbach SB
(1990) Korrelasie van dopamien vrylating in die kernklemme met manlike seksuele gedrag by rotte. Brain Res 524: 160-163.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1994) Terugkeer van seksuele uitputting deur serotonergiese en noradrenerge middels. Behav Brain Res 62: 127-134.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1995a) Deelname van die sentrale noradrenerge stelsel in die herstigting van kopulatoriese gedrag van seksueel uitgeputte rotte deur yohimbien, naloksoon en 8-OH-DPAT. Brein Res Bull 38: 399-404.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1995b) Opioïede antagoniste en die seksuele versadigingsverskynsel. Psychopharmacol 122: 131-136.
- ↵
- Rolls BJ
(1986) Sensories-spesifieke versadiging. Nutr Ds 44: 93-101.
- ↵
- Salamone JD
(1996) Die gedrags-neurochemie van motivering: metodologiese en konseptuele kwessies in die studie van die dinamiese aktiwiteit van nukleus volg dopamien. J Neurosci Metodes 64: 137-149.
- ↵
- Sato Y,
- Wada H,
- Horita H,
- Suzuki N,
- Shibuya A,
- Adachi H,
- Kato R,
- Tsukamoto T,
- Kumamoto Y
(1995) Dopamien vrystelling in die mediale preoptiese area tydens manlike kopulatoriese gedrag by rotte. Brain Res 692: 66-70.
- ↵
- Schultz W
(1992) Aktiwiteit van dopamienneurone in die optredende primaat. Semin Neurosci 4: 129-138.
- ↵
- Shimura T,
- Yamamoto T,
- Shimokochi M
(1994) Die mediale preoptiese area is betrokke by beide seksuele opwinding en prestasie by manlike rotte: herbeoordeling van neuronaktiwiteit in vrybewegende diere. Brain Res 640: 215-222.
- ↵
- Wenkstern D,
- Pfaus JG,
- Fibiger HC
(1993) Dopamien-oordrag styg in die kernkloppers van manlike rotte tydens hul eerste blootstelling aan seksueel ontvanklike vroulike rotte. Brain Res 618: 41-46.
- ↵
- Wilson C,
- Nomikos GG,
- Collu M,
- Fibiger HC
(1995) Dopaminerge korrelate van gemotiveerde gedrag: belangrikheid van ry. J Neurosci 15: 5169-5178.
- ↵
- Wilson JR,
- Kahn RE,
- Strand FA
(1963) Modifikasie in die seksuele gedrag van manlike rotte wat geproduseer word deur die stimulusvroulike te verander. J Comp Fisioloog Psigiatrie 56: 636-644.
- ↵
- Zetterström T,
- Skerp T,
- Ungerstedt U
(1984) Effek van neuroleptiese middels op striatale dopamien vrystelling en metabolisme in die wakker rot, bestudeer deur intrakerebrale dialise. Eur J Pharmacol 106: 27-37.
Artikels wat verwys na hierdie artikel
- Endogene Opioïed-geïnduceerde Neuroplasticiteit van Dopaminerge Neuronen in die Ventrale Tegmentale Gebied Invloede Natuurlike en Opiate Beloning Blaar van Neurowetenschappen, 25 Junie 2014, 34 (26): 8825-8836
- Onbalans in die sensitiwiteit vir verskillende soorte belonings in patologiese dobbelary Brein, 1 Augustus 2013, 136 (8): 2527-2538
- Wet op Natuurlike en Dwelmbelonings oor gemeenskaplike neurale plastisiteitsmeganismes met (Delta) FosB as 'n sleutelmediator Blaar van Neurowetenskap, 20 Februarie 2013, 33 (8): 3434-3442
- Neuronale Responses in die Nucleus Accumbens Shell tydens seksuele gedrag in manlike rotte Blaar van Neurowetenskap, 1 Februarie 2012, 32 (5): 1672-1686
- Romantiese liefde: 'n soogdier breinstelsel vir maat keuse Filosofiese Transaksies van die Koninklike Genootskap B: Biologiese Wetenskappe, 29 Desember 2006, 361 (1476): 2173-2186
- Hedoniese warm kolle in die brein Die Neuroscientist, 1 Desember 2006, 12 (6): 500-511
- Modulasie deur sentrale en basolaterale Amygdalar Kern van Dopaminerge Korrelate van Voeding tot Satiety in die Rat Nucleus Accumbens en Mediale Prefrontale Cortex Blaar van Neurowetenschappen, 15 Desember 2002, 22 (24): 10958-10965
- Frekwensie van Dopamien Konsentrasie Transiente Verhogings in Dorsale en Ventrale Striatum van Manlike Rotte tydens Inleiding van Conspecifics Blaar van Neurowetenschappen, 1 Desember 2002, 22 (23): 10477-10486
- Boekoorsig: Dinamika van Ekstracellulêre Dopamien in die Akute en Chroniese Aksies van Kokaïen Die Neuroscientist, 1 Augustus 2002, 8 (4): 315-322
- Differensiële Betrokkenheid van NMDA, AMPA / Kainate, en Dopamien Receptors in die Nucleus Accumbens Kern in die Verkryging en Uitvoering van Pavlovian Benadering Gedrag Blaar van Neurowetenschappen, 1 Desember 2001, 21 (23): 9471-9477
- Beheer van kokaïen-soekgedrag deur dwelmverwante stimuli by rotte: Effekte op die herstel van blus operante reaksie en ekstrasellulêre dopamienvlakke in amygdala en nucleus accumbens PNAS, 11 April 2000, 97 (8): 4321-4326
- Fasilitering van seksuele gedrag en verbeterde dopamien Efflux in die Nucleus Accumbens van Manlike Rotte na D-Amfetamien-Geïnduceerde Gedrags Sensitisering Blaar van Neurowetenschappen, 1 Januarie 1999, 19 (1): 456-463
- NEUROWETENSKAP: die aandag van die brein kry Wetenskap, 3 Oktober 1997, 278 (5335): 35-37
- Dopaminerge Korrelate van Sensoriespesifieke Satiety in die Mediale Prefrontale Cortex en Nucleus Accumbens van die Rat Blaar van Neurowetenschappen, 1 Oktober 1999, 19 (19): RC29