Dev Ds. 2008; 28(1): 62-77. doi: 10.1016 / j.dr.2007.08.003
Abstract
Adolessensie is 'n ontwikkelingsperiode wat gekenmerk word deur suboptimale besluite en aksies wat aanleiding gee tot 'n verhoogde voorkoms van onopsetlike beserings en geweld, alkohol- en dwelmmisbruik, onbedoelde swangerskap en seksueel oordraagbare siektes. Tradisionele neurobiologiese en kognitiewe verklarings vir adolessente gedrag het versuim om die nie-lineêre veranderinge in gedrag wat tydens adolessensie waargeneem word, in verhouding tot die kinderjare en volwassenheid te verduidelik. Hierdie hersiening bied 'n biologies-geloofwaardige konseptualisering van die neurale meganismes wat onderliggend aan hierdie nie-lineêre veranderinge in gedrag is, as 'n verhoogde reaksie op aansporings terwyl impulsbeheer nog relatief onvolwasse is gedurende hierdie tydperk. Onlangse menslike beeld- en dierstudies bied 'n biologiese basis vir hierdie siening, wat die differensiële ontwikkeling van limbiese beloningstelsels relatief tot top-down beheersisteme tydens adolessensie relatief tot kinder- en volwasse jare voorstel. Hierdie ontwikkelingspatroon kan in daardie adolessente vererger word met 'n aanleg tot risiko-neem, wat die risiko vir swak uitkomste verhoog.
Volgens die Nasionale Sentrum vir Gesondheidsstatistiek, is daar jaarliks meer as 13,000-adolessente sterftes in die Verenigde State. Ongeveer 70% van hierdie sterftes weens motorvoertuigongelukke, onopsetlike beserings, moord en selfmoord (Eaton et al., 2006). Resultate van die 2005 Nasionale Jeugrisikobestuursopname (YRBS) toon dat adolessente betrek word aan gedrag wat hul waarskynlikheid van dood of siekte verhoog deur 'n voertuig te bestuur nadat hulle gedrink het of sonder 'n veiligheidsgordel, wapens dra, onwettige stowwe gebruik en onbeskermde seks aangaan. wat lei tot onbedoelde swangerskappe en SOS'e, insluitend MIV-infeksieEaton et al., 2006). Hierdie statistieke beklemtoon die belangrikheid van die verstaan van riskante keuses en aksies in adolessente.
'N Aantal kognitiewe en neurobiologiese hipoteses is gepostuleer vir hoekom adolessente betrokke is by suboptimale keusgedrag. In 'n onlangse oorsig van die literatuur oor menslike adolessente breinontwikkeling, Yurgelun-Todd (2007) stel voor dat kognitiewe ontwikkeling deur die adolessente jare geassosieer word met progressief groter doeltreffendheid van kognitiewe beheerskapasiteite. Hierdie doeltreffendheid word beskryf as afhanklik van volwassenheid van die prefrontale korteks, soos blyk uit verhoogde aktiwiteit binne fokale prefrontale streke (Rubia et al., 2000; Tamm, Menon, & Reiss, 2002) en verminderde aktiwiteit in irrelevante breinstreke (Brown et al., 2005; Durston et al., 2006).
Hierdie algemene patroon, van verbeterde kognitiewe beheer met volwassenheid van die prefrontale korteks, dui op 'n lineêre toename in ontwikkeling vanaf die kinderjare tot volwassenheid. Tog is suboptimale keuses en aksies wat tydens adolessensie waargeneem word, 'n nie-lineêre verandering in gedrag wat van kinderjare en volwassenheid onderskei kan word, soos blyk uit die Nasionale Sentrum vir Gesondheidsstatistiek oor adolessente gedrag en sterftes. As kognitiewe beheer en 'n onvolwasse prefrontale korteks die basis was vir suboptimale keusgedrag, moet kinders opmerklik soortgelyk of selfs erger as adolessente lyk, gegewe hul minder ontwikkelde prefrontale korteks en kognitiewe vermoëns. So, onvolwasse prefrontale funksie alleen, kan nie rekening hou met adolessente gedrag nie.
'N Korrekte konseptualisering van kognitiewe en neurobiologiese veranderinge tydens adolessensie moet adolessensie as 'n oorgangsontwikkelingsperiode behandel (Spies, 2000), eerder as 'n enkele momentopname in die tyd (Casey, Tottenham, Liston en Durston, 2005). Met ander woorde, om hierdie ontwikkelingsperiode te verstaan, is oorgange in en uit adolessensie nodig om onderskeidende eienskappe van hierdie stadium van ontwikkeling te onderskei. Die vestiging van ontwikkelingsbane vir kognitiewe en neurale prosesse is noodsaaklik om hierdie oorgange te karakteriseer en interpretasies te beperk oor veranderinge in gedrag gedurende hierdie tydperk. Op 'n kognitiewe of gedragsvlak word adolessente gekenmerk as impulsiewe (dws ontbrekende kognitiewe beheer) en die neem van risiko's met hierdie konstrukte word sinoniem en sonder waardering vir duidelike ontwikkelingsbane van elk gebruik. Op 'n neurobiologiese vlak stel menslike beeld- en dierstudie verskillende neurobiologiese basisse en ontwikkelingsbane voor vir die neurale stelsels wat onderliggend is aan hierdie afsonderlike konstrukte van impulsbeheer en riskante besluite.
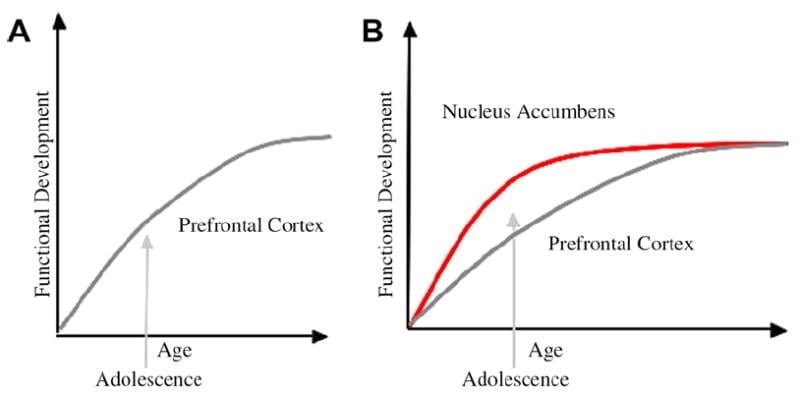
Ons het 'n neurobiologiese model ontwikkel van adolessente ontwikkeling binne hierdie raamwerk wat op knaagdiermodelle bou (Laviola, Adriani, Terranova en Gerra, 1999; Spies, 2000) en onlangse beeldstudie van adolessensie (Ernst et al., 2005; Galvan, Hare, Voss, Glover, & Casey, 2007; Galvan et al., 2006). Fig 1 hieronder wys hierdie model. Aan die linkerkant is die tradisionele karakterisering van adolessensie as verwant byna uitsluitlik aan die onvolwassenheid van die prefrontale korteks. Regs is ons voorgestelde neurobiologiese model wat illustreer hoe limbiese subkortikale en prefrontale top-down beheerstreke saam oorweeg moet word. Die spotprent illustreer verskillende ontwikkelingsbane vir hierdie stelsels, met limbiese stelsels wat vroeër ontwikkel as prefrontale beheerstreke. Volgens hierdie model word die individu meer bevoordeel deur funksioneel volwasse limbiese streke gedurende adolessensie (dws onbalans van limbies relatief tot prefrontale beheer), in vergelyking met kinders, vir wie hierdie sisteme (di limbies en voorfrontale) albei nog ontwikkel; en in vergelyking met volwassenes, vir wie hierdie stelsels volwasse is. Hierdie perspektief bied 'n basis vir nie-lineêre verskuiwings in gedrag oor ontwikkeling, as gevolg van vroeëre veroudering van hierdie limbiese relatief tot minder volwasse top-down prefrontale beheer streek. Met die ontwikkeling en ondervinding bied die funksionele konneksie tussen hierdie streke 'n meganisme vir top-down beheer van hierdie streke (Hare, Voss, Glover, & Casey, 2007a). Verder versoen die model die teenstrydigheid van gesondheidstatistiek van riskante gedrag tydens adolessensie, met die slim waarneming deur Reyna en Farley (2006) dat adolessente in staat is om risiko's van gedrag waarin hulle betrokke is, te redeneer en te verstaan. Volgens ons model, in emosionele waarskynlike situasies, sal die limbiese stelsel oor beheersisteme wen, gegewe sy volwassenheid relatief tot die prefrontale beheerstelsel. Bewyse van gedrags- en menslike beeldingstudies ter ondersteuning van hierdie model word verskaf in die konteks van aksies in belonende en emosionele kontekste (Galvan et al., 2006, 2007; Hare, Voss, Glover, & Casey, 2007b; Hare et al., 2007a). Daarbenewens spekuleer ons hoekom die brein op hierdie manier kan ontwikkel en hoekom sommige tieners groter risiko kan hê om suboptimale besluite te neem wat lei tot swakder langtermynuitkomste (Galvan et al., 2007; Hare et al., 2007b).
Die tradisionele verklaring van adolessente gedrag word voorgestel as gevolg van die langdurige ontwikkeling van die prefrontale korteks (A). Ons model neem die ontwikkeling van die prefrontale korteks in ag, saam met subkortiese limbiese streke (bv. Nucleus accumbens) wat by riskante keuses en aksies betrokke was (B).
Ontwikkeling van doelgerigte gedrag
'N Hoeksteen van kognitiewe ontwikkeling is die vermoë om onvanpaste gedagtes en aksies ten gunste van doelgerigte kinders te onderdruk, veral in die teenwoordigheid van dwingende aansporings (Casey, Galvan, & Hare, 2005; Casey et al., 2000b; Casey, Thomas, David-son, Kunz, & Franzen, 2002a; Casey, Tottenham, & Fossella, 2002b). 'N Aantal klassieke ontwikkelingsstudies het getoon dat hierdie vermoë ontwikkel word deur die kinderjare en adolessensie (Geval, 1972; Flavell, Feach en Chinsky, 1966; Keating & Bobbitt, 1978; Pascual-Leone, 1970). Verskeie teoretici het aangevoer dat kognitiewe ontwikkeling te wyte is aan toenames in verwerkingsspoed en doeltreffendheid en nie as gevolg van 'n toename in verstandelike kapasiteit nie (bv. Bjorkland, 1985; Bjorkland, 1987; Geval, 1985). Ander teoretici het die konstruksie van "inhibitiewe" prosesse in hul rekening van kognitiewe ontwikkeling ingesluit (Harnishfeger & Bjorkland, 1993). Volgens hierdie rekening word onvolwasse kognisie gekenmerk deur vatbaarheid vir inmenging van mededingende bronne wat onderdruk moet word (bv. Brainerd & Reyna, 1993; Casey, Thomas, Davidson, Kunz, & Franzen, 2002a; Dempster, 1993; Diamant, 1985; Munakata & Yerys, 2001). Dus doelgerigte gedrag vereis die beheer van impulse of vertraging van bevrediging vir die optimalisering van uitkomste, en hierdie vermoë blyk te wees oor volwassenheid oor die kind en adolessensie.
Adolessente gedrag is beskryf as impulsief en riskant, byna sinoniem, maar hierdie konstrukte maak staat op verskillende kognitiewe en neurale prosesse, wat verskillende konstrukte met verskillende ontwikkelingsbane aandui. Spesifiek, 'n oorsig van die literatuur dui daarop dat impulsiwiteit verminder met ouderdom oor die kinderjare en adolessensie (Casey et al., 2002a; Casey, Galvan et al., 2005; Galvan et al., 2007) en word geassosieer met uitgerekte ontwikkeling van die prefrontale korteks (Casey, Galvan et al., 2005), hoewel daar verskille is in die mate waarin 'n gegewe individu impulsief is of nie, ongeag ouderdom.
In teenstelling met impuls / kognitiewe beheer, blyk dit dat die neem van risiko's gedurende adolessensie relatief tot kinderjare en volwassenheid verband hou met subkortikale stelsels wat bekend is om by die evaluering van belonings betrokke te wees. Menslike beeldingstudies wat hersien gaan word, dui op 'n toename in subkortiese aktivering (bv. Accumbens) by die maak van riskante keuses (Kuhnen & Knutson, 2005; Matthews & et al., 2004; Montague & Berns, 2002) wat in adolessente oordrewe is, relatief tot kinders en volwassenes (Ernst et al., 2005; Galvan et al., 2006). Hierdie bevindings dui op verskillende trajecten vir beloning of incentive-based gedrag, met die vroeëre ontwikkeling van hierdie stelsels in verhouding tot die controlesystemen wat 'n langdurige en lineaire ontwikkelingscursus toon, in terme van die oorheersing van onvanpaste keuses en aksies ten gunste van doelgerigte.
Bewyse van neuroimaging studies van menslike brein ontwikkeling
Onlangse ondersoeke na die ontwikkeling van adolessente brein is gebaseer op vooruitgang in neuroimaging metodologieë wat maklik gebruik kan word met die ontwikkeling van menslike populasies. Hierdie metodes is gebaseer op magnetiese resonansiebeeldvorming (MRI) metodes (sien Fig 2) en sluit in: strukturele MRI, wat gebruik word om die grootte en vorm van strukture te meet; funksionele MRI wat gebruik word om patrone van breinaktiwiteit te meet; en diffusie tensor imaging (DTI) wat gebruik word om die verbindings van wit materie veselkanale te indekseer. Bewyse vir ons ontwikkelingsmodel van mededinging tussen kortikale en subkortiese streke word ondersteun deur onvolwasse strukturele en funksionele konnektiwiteit soos gemeet deur onderskeidelik DTI en fMRI.
MRI studies van menslike brein ontwikkeling
Verskeie studies het strukturele MRI gebruik om die anatomiese verloop van normale breinontwikkeling te karteer (sien oorsig deur Durston et al., 2001). Alhoewel die totale breingrootte op ouderdom ses ongeveer 90% van sy volwasse grootte is, bly die grys en wit materie-subkomponente van die brein dinamiese veranderinge gedurende adolessensie. Data uit onlangse longitudinale MRI-studies dui aan dat grysstofvolume 'n omgekeerde U-vormpatroon het, met groter streeksvariasie as wit materie (Giedd, 2004; Gogtay et al., 2004; Sowell et al, 2003; Sowell, Thompson, & Toga, 2004). Oor die algemeen, streke wat primêre funksies onderhou, soos motoriese en sensoriese stelsels, wat die vroegste is; hoër-assosiasie areas, wat hierdie primêre funksies integreer, later volwassenes (Gogtay et al., 2004; Sowell, Thompson, & Toga, 2004). Byvoorbeeld, studies wat MRI-gebaseerde maatreëls gebruik, toon dat kortikale grysstofverlies die vroegste in die primêre sensorimotoriese gebiede voorkom en die laaste in die dorsolaterale prefrontale en laterale tydelike kortikale (Gogtay et al., 2004). Hierdie patroon is in ooreenstemming met niehuman primate en menslike postmortem studies wat toon dat die prefrontale korteks een van die laaste breinstreke is om te volwasse (Bourgeois, Goldman-Rakic, & Rakic, 1994; Huttenlocher, 1979). In teenstelling met grys materie, groei die wit materie volume in 'n rowwe lineêre patroon, wat deurlopend ontwikkel tot in volwasse jare (Gogtay et al., 2004). Hierdie veranderinge weerspieël vermoedelik deurlopende myelering van aksone deur oligodendrocytes wat neuronale geleiding en kommunikasie versterk.
Alhoewel minder aandag aan subkortiese streke gegee word wanneer strukturele veranderinge ondersoek word, word sommige van die grootste veranderinge in die brein oor ontwikkeling in hierdie streke gesien, veral in die basale ganglia (Sowell et al., 1999, Sien Fig 3) en veral by mans (Giedd et al., 1996). Ontwikkelingsveranderinge in strukturele volume binne basale ganglia en prefrontale streke is interessant in die lig van bekende ontwikkelingsprosesse (bv. Dendritiese arborisering, selood, sinaptiese snoei, miellinasie) wat tydens die kinder- en adolessensie voorkom. Hierdie prosesse maak voorsiening vir fyn afstemming en versterking van verbindings tussen prefrontale en subkortiese streke met ontwikkeling en leer wat saamval met groter kognitiewe beheer. Hoe hou hierdie strukturele veranderinge verband met kognitiewe veranderinge? 'N aantal studies het verwante frontale lobstruktuurveroudering en kognitiewe funksie met behulp van neuropsigologiese en kognitiewe maatreëls (bv. Sowell et al., 2003). Spesifiek, is verenigings gerapporteer tussen MRI-gebaseerde prefrontale kortikale en basale ganglia streeksvolumes en maatstawwe van kognitiewe beheer (dws die vermoë om 'n onvanpaste respons ten gunste van 'n ander te oorkom of om aandag te skenk aan irrelevante stimulus-eienskappe ten gunste van relevante stimulus-eienskappe (Casey, Trainor et al., 1997). Hierdie bevindings dui daarop dat kognitiewe veranderinge weerspieël word in strukturele breinveranderinge en onderstreep die belangrikheid van subkortiese (basale ganglia) sowel as kortikale (bv. Prefrontale korteks) ontwikkeling.

DTI studies van menslike brein ontwikkeling
Die MRI-gebaseerde morfometrie studies het aanbeveel dat corticale verbindings fyn ingestel word met die uitskakeling van 'n oorvloed van sinapse en die versterking van relevante verbindings met ontwikkeling en ondervinding. Onlangse vordering in MRI-tegnologie, soos DTI, bied 'n potensiële instrument om die rol van spesifieke wit materie-areas na die ontwikkeling van die brein en gedrag met meer besonderhede te ondersoek. Relevant vir hierdie vraestel is die neuroimaging studies wat die ontwikkeling van veselkanale verbind het met verbeterings in kognitiewe vermoë. Spesifiek, verenigings tussen DTI-gebaseerde maatreëls van prefrontale wit materie ontwikkeling en kognitiewe beheer by kinders is getoon. In een studie is die ontwikkeling van hierdie kapasiteit positief gekorreleer met prefrontale pariëtale veselkanale (Nagy, Westerberg, & Klingberg, 2004) in ooreenstemming met funksionele neuroimaging studies wat die differensiële werwing van hierdie streke toon in kinders relatief tot volwassenes.
Met 'n soortgelyke benadering, Liston et al. (2005) het getoon dat wit materiaalkanale tussen prefrontale basale ganglia en postosterale veselkanale steeds oor die kinderjare tot in volwassenheid ontwikkel, maar slegs die dele tussen die prefrontale korteks en basale ganglia word gekorreleer met impulsbeheer, soos gemeet deur prestasie op 'n poging taak. Die prefrontale veselkanale is gedefinieer deur streke van belange wat in 'n fMRI-studie geïdentifiseer is, met dieselfde taak. Tussen beide ontwikkelings-DTI-studies is vezelwegmaatreëls met ontwikkeling verband gehou, maar spesifisiteit van spesifieke veselkanale met kognitiewe prestasie is getoon deur dissosiasie van die betrokke kanaal (Liston et al., 2005) of kognitiewe vermoë (Nagy et al., 2004). Hierdie bevindinge onderstreep die belangrikheid van die ondersoek van nie net streeks-, maar kringverwante veranderings wanneer eise oor ouderdomsafhanklike veranderinge in neurale substrates van kognitiewe ontwikkeling gemaak word nie.
Funksionele MRI-studies van gedrags- en breinontwikkeling
Alhoewel strukturele veranderinge gemeet deur MRI en DTI geassosieer is met gedragsveranderings tydens ontwikkeling, is 'n meer direkte benadering vir die ondersoek van struktuur-funksie assosiasie gelyktydig veranderinge in die brein en gedrag, soos by fMRI. Die vermoë om funksionele veranderings in die ontwikkelende brein met MRI te meet, het 'n belangrike potensiaal vir die veld van ontwikkelingswetenskap. In die konteks van die huidige vraestel bied fMRI 'n middel om interpretasies van adolessente gedrag te beperk. Soos voorheen gesê, word die ontwikkeling van die prefrontale korteks geglo dat dit 'n belangrike rol speel in die veroudering van hoër kognitiewe vermoëns soos besluitneming en kognitiewe beheer (Casey, Tottenham en Fossella 2002b; Casey, Trainor et al., 1997). Baie paradigmas is saam met fMRI gebruik om die neurobiologiese basis van hierdie vermoëns te assesseer, insluitend flank, Stroop en go / nogo take (Casey, Castellanos et al., 1997; Casey, Giedd, & Thomas, 2000a; Durston et al., 2003). Gesamentlik toon hierdie studies dat kinders afsonderlike maar dikwels groter, meer diffuse prefrontale streke werf wanneer hulle hierdie take uitvoer as volwassenes. Die patroon van aktiwiteit binne breinstreke sentraal tot taakprestasie (dws wat korreleer met kognitiewe prestasie) word meer gefokus of verfyn met ouderdom, terwyl streke nie korreleer met taakverrigting verminder in aktiwiteit met ouderdom nie. Hierdie patroon is waargeneem oor beide dwarsdeursnee (Brown et al., 2005) en longitudinale studies (Durston et al., 2006) en oor 'n verskeidenheid paradigmas. Alhoewel neuroimaging studies nie die meganisme van sulke ontwikkelingsveranderinge definitief kan karakteriseer nie (bv. Dendritiese arborisering, sinaptiese snoei), weerspieël die bevindinge ontwikkeling binne en verfyning van projeksies na en van geaktiveerde breinstreke met rijping en stel voor dat hierdie veranderinge oor 'n langdurige periode van tyd (Brown et al., 2005; Bunge, Dudukovic, Thomason, Vaidya, & Gabrieli, 2002; Casey, Trainor et al., 1997; Casey et al., 2002a; Crone, Donohue, Honomichl, Wendelken, & Bunge, 2006; Luna et al., 2001; Moses et al., 2002; Schlaggar et al., 2002; Tamm et al., 2002; Thomas et al., 2004; Turkeltaub, Gareau, Flowers, Zeffiro, en Eden, 2003).
Hoe kan hierdie metodologie ons inlig oor die vraag of adolessente inderdaad onvoldoende kognitiewe beheer (impulsiewe) ontbreek of riskant is in hul keuses en aksies? Impulskontrole soos gemeet deur kognitiewe beheerstake soos die taak / taak toon 'n lineêre patroon van ontwikkeling oor kinderjare en adolessensie soos hierbo beskryf. Die onlangse neuroimaging studies het egter begin met die ondersoek van beloningsverwante prosessering spesifiek vir die risiko-opname in adolessente (Bjork et al., 2004; Ernst et al., 2005; Mei et al., 2004). Hierdie studies het hoofsaaklik gefokus op die streek van die pasgeborenes, 'n gedeelte van die basale ganglia wat betrokke is by die voorspelling van beloning, eerder as die karakterisering van die ontwikkeling van hierdie streek in kombinasie met top-down beheerstreke (prefrontale korteks). Alhoewel 'n onlangse verslag van minder ventrale prefrontale aktiwiteit in adolessente relatief tot volwassenes tydens 'n monetêre besluitneming op risiko-gedrag is getoon (Eshel, Nelson, Blair, Pine, & Ernst, 2007).
Oor die algemeen het min studies ondersoek hoe die ontwikkeling van beloningskringe in subkortiese streke (bv. Akkumbens) verander in samewerking met die ontwikkeling van kortikale prefrontale streke. Verder, hoe hierdie neurale veranderinge saamval met beloningsoekende, impulsiwiteits- en risiko-optrede, bly relatief onbekend. Ons neurobiologiese model stel voor dat die kombinasie van verhoogde responsiwiteit vir belonings en onvolwassenheid in gedragsbeheerareas adolessente kan vooroordeel om onmiddellike, eerder as langtermynwinste te soek, miskien hul toename in riskante besluitneming en impulsiewe gedrag te verduidelik. Die opsporing van subkortiese (bv. Toevallige) en kortikale (bv. Prefrontale) ontwikkeling van besluitneming oor die kinderjare deur volwassenheid, bied addisionele beperkinge op of veranderinge wat in adolessensie gerapporteer is spesifiek is vir hierdie ontwikkelingstydperk, of weerspieëling van die volwassenheid wat in 'n ietwat lineêre patroon van kinderjare tot volwassenheid.
Empiriese bewyse uit 'n onlangse fMRI-studie help om ons neurobiologiese model te ondersteun en neem 'n oorgangsbenadering om adolessensie te verstaan deur veranderinge voor en na adolessensie te ondersoek. In hierdie studie (Galvan et al., 2006), het ons gedrags- en neurale reaksies ondersoek om manipulasies oor ontwikkeling te beloon, met fokus op breinstreke wat betrokke is by beloningverwante leer en gedrag by diere (Hikosaka & Watanabe, 2000; Pecina, Cagniard, Berridge, Aldridge, & Zhuang, 2003; Schultz, 2006) en volwasse beeldvormingstudies (bv. Knutson, Adams, Fong, & Hommer, 2001; O, Doherty, Kringelbach, Rolls, Hornak, Andrews, 2001; Zald et al., 2004) en in studies van verslawing (Hyman & Malenka, 2001; Volkow & Li, 2004). Gebaseer op knaagdiermodelle (Laviola et al., 1999; Spies, 2000) en vorige beeldwerkwerk (Ernst et al., 2005), het ons vermoed dat relatief tot kinders en volwassenes, sou adolessente oordrewe aktivering van die accumbens toon, in kombinasie met minder volwasse werwing van top-down prefrontale beheerstreke. Onlangse werk wat vertraagde funksionele konnektiwiteit tussen hierdie prefrontale en limbiese subkortikale streke in adolessensie teenoor volwassenes toon, bied 'n meganisme vir die gebrek aan top-down beheer van hierdie streke (Hare et al., 2007a).
Ons bevindings was in ooreenstemming met knaagdiermodelle (Laviola, Macri, Morley-Fletcher, & Adriani, 2003) en vorige beeldvormingstudies (Ernst et al., 2005) wat verbeterde aktiwiteite vir beloning tydens adolessensie voorstel. Inderdaad, in verhouding tot kinders en volwassenes, het adolessente 'n oordrewe reaksie in reaksie op beloning getoon. Beide kinders en adolessente het egter 'n minder volwasse respons in prefrontale beheerstreke as volwassenes getoon. Hierdie bevindinge dui op verskillende ontwikkelingsraamwerke vir hierdie streke kan die verbetering in die toonaangewende aktiwiteit, relatief tot kinders of volwassenes, onderliggend maak, wat op sy beurt verband hou met die verhoogde impulsiewe en riskante gedrag wat gedurende hierdie ontwikkelingstydperk waargeneem word (kyk Fig 4).

Differensiële werwing van prefrontale en subkortiese streke is oor 'n aantal ontwikkelingsfMRI-studies gerapporteer (Casey et al., 2002b; Monk et al., 2003; Thomas et al., 2004). Hierdie bevindings is tipies geïnterpreteer in terme van onvolwasse prefrontale streke eerder as 'n wanbalans tussen prefrontale en subkortikale streeksontwikkeling. Bewyse gegee van prefrontale streke om toepaslike aksies in verskillende kontekste te lei (Miller & Cohen, 2001) Onvolwasse prefrontale aktiwiteit kan gepaste beraming van toekomstige uitkomste en beoordeling van riskante keuses belemmer, en kan dus minder invloedryk wees op beloningwaardasie as die accumbens. Hierdie patroon is in ooreenstemming met vorige navorsing wat verhoogde subkortikale, relatief tot kortikale aktiwiteit toon, wanneer besluite bevoordeel word deur onmiddellike oor langtermyn-winste (McClure, Laibson, Loewenstein & Cohen, 2004). Verder is die aktiwiteit van accumbens getoon met fMRI om positief te korreleer met die daaropvolgende risiko-take gedrag (Kuhnen & Knutson, 2005). Tydens adolessensie, relatief tot kinder- of volwassenheid, kan onvolwasse ventrale prefrontale korteks nie voldoende boonste beheer van robuust geaktiveerde beloningsverwerkingsgebiede (bv. Accumbens) verskaf nie, wat minder invloed op prefrontale stelsels (orbitofrontale korteks) in verhouding tot die accumbens in beloning tot gevolg het. waardasie.
Waarom sal die brein geprogrammeer word om hierdie manier te ontwikkel?
Adolessensie is die oorgangstydperk tussen die kinderjare en volwassenheid wat dikwels by puberteit voorkom. Puberteit dui op die begin van seksuele volwassenheid (Graber & Brooks-Gunn, 1998) en kan deur biologiese merkers gedefinieer word. Adolessensie kan beskryf word as 'n progressiewe oorgang tot volwassenheid met 'n onstuimige ontogenetiese tydskursus (Spies, 2000). Evolusionêre praat is adolessensie die tydperk waarin onafhanklikheidsvaardighede aangeleer word om sukses te behaal by skeiding van die beskerming van die gesin, alhoewel dit die kans op skadelike omstandighede verhoog (bv. Besering, depressie, angs, dwelmgebruik en verslawingKelley, Schochet, & Landry, 2004). Onafhanklik-soekende gedrag is algemeen oor spesies, soos toenames in maatskaplike interaksies en intensifikasies in nuwigheids- en risiko-optrede. Psigososiale faktore beïnvloed adolessente geneigdheid vir riskante gedrag. Risikovolle gedrag is egter die produk van 'n biologies gedrewe wanbalans tussen verhoogde nuwigheids- en sensasie-soek in kombinasie met onvolwasse selfregulerende bevoegdheid (Steinberg, 2004). Ons neurobiologiese data stel voor dat dit plaasvind deur die differensiële ontwikkeling van hierdie twee stelsels (limbies en beheer).
Spekulasie sou daarop dui dat hierdie ontwikkelingspatroon 'n evolusionêre funksie is. Jy moet hoërisikogedrag betree om jou familie en dorp te verlaat om 'n mate en risiko te neem op dieselfde tyd as hormone adolessente bestuur om seksuele vennote te soek. In die hedendaagse samelewing wanneer adolessensie onbepaald kan strek, met kinders wat saam met ouers woon en finansiële afhanklikheid het en die ouers later in die lewe kies, kan hierdie evolusie as onvanpas beskou word.
Daar is bewyse oor spesies vir verhoogde nuwigheids- en risiko-opnames gedurende die adolessente jare. Om dieselfde eweknie-eweknieë te soek en om met ouers te veg, wat almal help om die adolessent weg van die tuisgebied te kry vir paring, word gesien in ander spesies, insluitende knaagdiere, nie-menslike primate en sommige voëls (Spies, 2000). In verhouding tot volwassenes toon periadolessente rotte verhoogde nuwigheidsoptrede in 'n vrye keuse-nuwigheidsparadigma (Laviola et al., 1999). Neurochemiese bewyse dui daarop dat die balans in die adolessente brein tussen kortikale en subkortiese dopamien stelsels begin beweeg na groter kortikale dopamienvlakke gedurende adolessensie (Spies, 2000). Soortgelyke uitgerekte dopaminerge enerverwaring deur adolessensie tot volwassenheid is ook in die nie-menslike primaat prefrontale korteks getoon (Rosenberg & Lewis, 1995). Dus blyk dit verhoogde skynbare risiko's oor spesies en het belangrike aanpassingsdoeleindes.
Biologiese predisposisies, ontwikkeling en risiko
Individuele verskille in impulsbeheer en risiko's word vir 'n geruime tyd in die sielkunde erken (Benthin, Slovic en Severson, 1993). Miskien is een van die klassieke voorbeelde van individuele verskille wat gerapporteer word in hierdie vermoëns in die sosiale, kognitiewe en ontwikkelingsielkunde-literatuur, vertraging van bevrediging (Mischel, Shoda, & Rodriguez, 1989). Vertraging van bevrediging word tipies geassesseer in 3- tot 4-jarige kleuters. Die kleuter word gevra of hulle 'n klein beloning (een koekie) of 'n groot beloning (twee koekies) sal verkies. Die kind word dan vertel dat die proefpersoon die kamer sal verlaat om voor te berei vir opkomende aktiwiteite en verduidelik aan die kind dat as sy in haar sitplek bly en nie 'n koekie eet nie, sy die groot beloning sal ontvang. As die kind ook nie of nie kan wag nie, moet sy 'n klok bel om die eksperiment te ontbied en sodoende die kleiner beloning te ontvang. Sodra dit duidelik is, verstaan die kind die taak, sy sit by die tafel met die twee belonings en die bel. Afleiding in die vertrek word geminimaliseer, sonder speelgoed, boeke of prente. Die eksperimenteerder kom na 15 min terug, of nadat die kind die bel geroep het, die beloning geëet het, of enige tekens van nood aangedui het. Mischel het getoon dat kinders tipies op een van twee maniere optree: (1) óf hulle skakel die klokkie byna dadelik om die koekie te kry, wat beteken dat hulle net een kry; of (2) hulle wag en optimaliseer hul winste, en ontvang albei koekies. Hierdie waarneming dui daarop dat sommige individue beter is as ander in hul vermoë om impulse te beheer in die gesig van hoogs belangrike aansporings en hierdie vooroordeel kan vroeg in die kinderjare opgespoor word (Mischel et al., 1989) en hulle blyk te bly deur adolessensie en jong volwassenheid (Eigsti et al., 2006).
Wat kan individuele verskille in optimale besluitneming en gedrag verklaar? Sommige teoretici het geposuleer dat dopaminerge mesolimbiese kringe, wat betrokke is by die vergoeding van prosesse, onderliggend is aan riskante gedrag. Individuele verskille in hierdie stroombaan, soos alleliese variante in dopamienverwante gene, wat te min of te veel dopamien in subkortiese streke tot gevolg het, kan verband hou met die geneigdheid om aan riskante gedrag deel te neem (O'Doherty, 2004). Die kernwapens is getoon om onmiddellik in aktiwiteit te styg voordat risikobesluitings op monitêre-risiko-paradigmas gemaak word (Kuhnen & Knutson, 2005; Matthews et al., 2004; Montague & Berns, 2002) en soos voorheen beskryf, toon adolessente oordrewe accumbens-aktiwiteit om uitkomste te vergeleke met kinders of volwassenes (Ernst et al., 2005; Galvan et al., 2006). Hierdie data dui gesamentlik daarop dat adolessente as groep meer geneig is tot riskante keuses (Gardener & Steinberg, 2005), maar sommige adolessente sal meer geneig wees as ander om met riskante gedrag aan te pak, wat hulle op 'n potensiële groter risiko vir negatiewe uitkomste kan hê. Daarom is dit belangrik om individuele veranderlikheid te oorweeg wanneer komplekse brein-gedrag verhoudings met betrekking tot risiko-opname en beloning van verwerking in ontwikkelingsbevolkings ondersoek word.
Om individuele verskille in risiko-gedrag te ondersoek, Galvan et al. (2007) het onlangs die verband tussen aktiwiteit in beloningsverwante neurale kringe ondersoek in afwagting van 'n groot monetêre beloning met persoonlikheidseienskappe van risiko-neem en impulsiwiteit in adolessensie. Funksionele magnetiese resonansie beelding en anonieme selfverslaggraderingskale van riskante gedrag, risiko persepsie en impulsiwiteit is verkry in individue tussen die ouderdomme van 7 en 29 jaar. Daar was 'n positiewe assosiasie tussen accumbens-aktiwiteit en die waarskynlikheid om in riskante gedrag oor die ontwikkeling betrokke te raak. Hierdie aktiwiteit het gewissel as 'n funksie van individue se graderings van verwagte positiewe of negatiewe gevolge van sulke gedrag. Dié individue wat riskante gedrag beskou het as gevolg van ernstige gevolge, het die accumbens minder te beloon. Hierdie vereniging is grotendeels deur die kinders gedryf, met die volwassenes wat die gevolge van sulke gedrag as moontlik beskou. Impulsiwiteitsgraderings is nie geassosieer met accumbens-aktiwiteit nie, maar eerder met ouderdom. Hierdie bevindings dui daarop dat sommige individue tydens adolessensie meer geneig is om risiko-gedrag te betree as gevolg van ontwikkelingsveranderings in ooreenstemming met veranderlikheid in die voorkoms van 'n gegewe individu om risiko-optrede te betree, eerder as om eenvoudige impulsiwiteitsveranderinge te doen (kyk Fig 5).

Adolessente gedrag is herhaaldelik as impulsief en riskant gekenmerk (Steinberg, 2004, 2007), maar hierdie resensie van die beeldende literatuur dui op verskillende neurobiologiese substrate en verskillende ontwikkelingsraamwerke vir hierdie gedrag. Spesifiek, impulsiwiteit word geassosieer met onvolwasse ventrale prefrontale ontwikkeling en verminder geleidelik van kleins af tot volwassenheid (Casey, Galvan et al., 2005). Die negatiewe korrelasie tussen impulsiwiteitsgraderings en ouderdom in die studie deur Galvan et al. (2007) ondersteun hierdie begrip verder. In teenstelling hiermee word risiko-opname geassosieer met 'n toename in akkumulasieaktiwiteit (Kuhnen & Knutson, 2005; Matthews et al., 2004; Montague & Berns, 2002), dit is oordrewe in adolessente, relatief tot kinders en volwassenes (Ernst et al., 2005; Galvan et al., 2006). So adolessente keuses en gedrag kan nie verklaar word deur impulsiwiteit of uitgerekte ontwikkeling van die prefrontale korteks alleen nie, aangesien kinders dan voorspel word om groter risikotakers te wees. Die bevindings bied 'n neurale basis vir waarom sommige adolessente groter risiko as ander het, maar bied verder 'n basis vir hoe adolessente gedrag verskil van kinders en volwassenes in risikobepaling.
Gesamentlik, hierdie data dui daarop dat alhoewel adolessente as 'n groep as risikokopers beskou word (Gardener & Steinberg, 2005), sal sommige adolessente meer geneig wees as ander om met riskante gedrag betrokke te raak, sodat hulle op 'n groter potensiaal vir negatiewe uitkomste kan verkeer. Hierdie bevindinge onderstreep die belangrikheid om individuele veranderlikes te oorweeg wanneer komplekse breingedragverhoudings met betrekking tot risikobepaling en beloningverwerking in ontwikkelingsbevolkings ondersoek word. Verder kan hierdie individuele en ontwikkelingsverskille help om die kwesbaarheid van sommige individue te verklaar om risiko-aanneming wat verband hou met substansgebruik, en uiteindelik verslawing.
Gevolgtrekkings
Menslike beeldingstudies toon strukturele en funksionele veranderinge in frontostriatale streke (Giedd et al., 1996, 1999; Jernigan et al., 1991; Sowell et al., 1999; Vir hersiening, Casey, Galvan et al., 2005) wat blykbaar parallelle toenames in kognitiewe beheer en selfregulering (Casey, Trainor et al., 1997; Luna & Sweeney, 2004; Luna et al., 2001; Rubia et al., 2000; Steinberg, 2004; sien ook Steinberg, 2008, hierdie kwessie). Hierdie veranderinge blyk 'n verskuiwing in die aktivering van prefrontale streke te toon van diffuse na meer fokale werwing oor tyd (Brown et al., 2005; Bunge et al., 2002; Casey, Trainor et al., 1997; Durston et al., 2006; Moses et al., 2002) en verhoogde werwing van subkortiese streke gedurende adolessensie (Casey et al., 2002a; Durston et al., 2006; Luna et al., 2001). Alhoewel neuroimaging studies nie die meganisme van sulke ontwikkelingsveranderings finaal kan kenmerk nie, kan hierdie veranderinge in volume en struktuur die ontwikkeling binne en verfyning van projeksies na en van hierdie breinstreke weerspieël tydens die volwassing wat aandui dat die stelsel met ontwikkeling ontwikkel word.
Samevattend dui die bevindinge wat hier gesintetiseer word daarop dat verhoogde risikogedragsgedrag in adolessensie geassosieer word met verskillende ontwikkelingsbane van subkortiese genot en kortikale beheerstreke. Hierdie ontwikkelingsveranderings kan vererger word deur individuele verskille in aktiwiteit van beloningstelsels. Alhoewel adolessensie onderskei word as 'n tydperk wat gekenmerk word deur beloningsoekende en risiko-optredende gedrag (Gardener & Steinberg, 2005; Spies, 2000) Individuele verskille in neurale reaksies om te beloon, stel sommige adolessente voor om meer risiko's te neem as ander, en stel hulle groter risiko vir negatiewe uitkomste. Hierdie bevindings bied belangrike grondslag deur die verskillende bevindings wat verband hou met die gedrag van risiko's in adolessensie te sintetiseer en individuele verskille en ontwikkelingsmerkers te verstaan vir geneigdhede om negatiewe gedrag aan te pak.
Erkennings
Hierdie werk is gedeeltelik ondersteun deur toekennings van die Nasionale Instituut vir Dwelmmisbruik R01 DA18879 en die Nasionale Instituut van Geestesgesondheid 1P50 MH62196.
Verwysings
- Benthin A, Slovic P, Severson H. 'n Psigometriese studie van adolessente risiko persepsie. Joernaal van Adolessensie. 1993;16: 153-168. [PubMed]
- Bjork JM, Knutson B, Fong GW, Caggiano DM, Bennett SM, Hommer DW. Incentive-opgewekte breinaktivering by adolessente: ooreenkomste en verskille van jong volwassenes. Blaar van Neurowetenskap. 2004;24: 1793-1802. [PubMed]
- Bjorkland DF. Die rol van konseptuele kennis in die ontwikkeling van organisasie in kinders se geheue. In: Brainerd CJ, Pressley M, redakteurs. Basiese prosesse in geheue-ontwikkeling: Vordering in kognitiewe ontwikkelingsnavorsing. New York: Springer-Verlag; 1985. pp. 103-142.
- Bjorkland DF. Hoe ouderdom verander in kennisbasis dra by tot die ontwikkeling van kinders se geheue: 'n Interpretatiewe oorsig. Ontwikkelingsoorsig. 1987;7: 93-130.
- Bourgeois JP, Goldman-Rakic PS, Rakic P. Synaptogenese in die prefrontale korteks van rhesus ape. Serebrale korteks. 1994;4: 78-96. [PubMed]
- Brainerd CJ, Reyna VF. Geheue onafhanklikheid en geheue inmenging in kognitiewe ontwikkeling. Sielkundige Oorsig. 1993;100: 42-67. [PubMed]
- Brown TT, Lugar HM, Coalson RS, Miezin FM, Petersen SE, Schlaggar BL. Ontwikkelingsveranderinge in menslike serebrale funksionele organisasie vir woordgenerering. Serebrale korteks. 2005;15: 275-290. [PubMed]
- Bunge SA, Dudukovic NM, Thomason ME, Vaidya CJ, Gabrieli JD. Onvolwasse frontale lob bydraes tot kognitiewe beheer by kinders: Bewyse van fMRI. Neuron. 2002;33: 301-311. [PubMed]
- Geval R. Validasie van 'n neo-Piagetiese kapasiteitsbou. Tydskrif van Eksperimentele Kinderpsigologie. 1972;14: 287-302.
- Geval R. Intellektuele ontwikkeling: Geboorte tot volwassenheid. New York: Academic Press; 1985.
- Casey BJ, Castellanos FX, Giedd JN, Marsh WL, Hamburger SD, Schubert AB. et al. Implikasie van regte frontostriatale kringe in reaksie inhibisie en aandag-tekort / hiperaktiwiteitsversteuring. Tydskrif van die Amerikaanse Akademie vir Kinder- en Adolessente Psigiatrie. 1997;36: 374-383. [PubMed]
- Casey BJ, Galvan A, Hare TA. Veranderinge in serebrale funksionele organisasie tydens kognitiewe ontwikkeling. Huidige Advies in Neurobiologie. 2005;15: 239-244. [PubMed]
- Casey BJ, Giedd JN, Thomas KM. Strukturele en funksionele breinontwikkeling en sy verhouding tot kognitiewe ontwikkeling. Biologiese Sielkunde. 2000a;54: 241-257. [PubMed]
- Casey BJ, Thomas KM, Davidson MC, Kunz K, Franzen PL. Dissociatie van striatale en hippocampale funksie ontwikkelend met 'n stimulus-respons-verenigbaarheidstaak. Blaar van Neurowetenskap. 2002a;22: 8647-8652. [PubMed]
- Casey BJ, Thomas KM, Walliese TF, Badgaiyan RD, Eccard CH, Jennings JR, et al. Dissociation of response conflict, attentional selection and expectancy with functional magnetic resonance imaging. Verrigtinge van die Nasionale Akademie vir Wetenskap. 2000b;97: 8728-8733.
- Casey BJ, Tottenham N, Fossella J. Kliniese, beeldvorming, letsel en genetiese benaderings tot 'n model van kognitiewe beheer. Ontwikkelingspsigologie. 2002b;40: 237-254. [PubMed]
- Casey BJ, Tottenham N, Liston C, Durston S. Imaging die ontwikkelende brein: Wat het ons geleer oor kognitiewe ontwikkeling? Neigings in Kognitiewe Wetenskap. 2005;9: 104-110.
- Casey BJ, Trainor RJ, Orendi JL, Schubert AB, Nystrom LE, Giedd JN, et al. 'N Ontwikkelings funksionele MRI studie van prefrontale aktivering tydens die uitvoering van 'n go-no-go-taak. Tydskrif van Kognitiewe Neurowetenskap. 1997;9: 835-847.
- Crone E, Donohue S, Honomichl R, Wendelken C, Bunge S. Breinstreke wat buigbare reël gebruik tydens ontwikkeling. Blaar van Neurowetenskap. 2006;26: 11239-11247. [PubMed]
- Dempster FN. Weerstand teen inmenging: Ontwikkelingsveranderinge in 'n basiese verwerkingsmeganisme. In: Howe ML, Pasnak R, redakteurs. Opkomende temas in kognitiewe ontwikkeling Volume 1: Grondslae. New York: Springer; 1993. pp. 3-27.
- Diamant A. Ontwikkeling van die vermoë om herroeping te gebruik om aksie te rig, soos aangedui deur kinders se prestasie op AB. Kinderontwikkeling. 1985;56: 868-883. [PubMed]
- Durston S, Davidson MC, Thomas KM, Word MS, Tottenham N, Martinez A, et al. Parametriese manipulasie van konflik- en reaksie-kompetisie deur gebruik te maak van vinnige, gemengde-proefgebeurtenis-verwante fMRI. Neuro Image. 2003;20: 2135-2141. [PubMed]
- Durston S, Davidson MC, Tottenham N, Galvan A, Spicer J, Fossella J et al. 'N Skuif van diffuse na fokale kortikale aktiwiteit met ontwikkeling. Ontwikkelingskunde. 2006;1: 18-20. [PubMed]
- Durston S, Hulshoff Pol HE, Casey BJ, Giedd JN, Buitelaar JK, van Engeland H. Anatomiese MRI van die ontwikkelende menslike brein: Wat het ons geleer? Tydskrif van die Amerikaanse Akademie vir Kinderadolente Psigiatrie. 2001;40: 1012-1020.
- Eaton LK, Kinchen S, Ross J, Hawkins J, Harris WA, Lowry R, et al. Jeug risiko gedrag toesig-Verenigde State, 2005, toesig opsommings. Weeklikse verslag oor morbiditeit en mortaliteit. 2006;55: 1-108. [PubMed]
- Eigsti IM, Zayas V, Mischel W, Shoda Y, Ayduk O, Dadlani MB, et al. Voorspelling van kognitiewe beheer vanaf voorskoolse tot laat adolessensie en jong volwassenheid. Sielkundige Wetenskap. 2006;17: 478-484. [PubMed]
- Ernst M, Nelson EE, Jazbec S, McClure EB, Monk CS, Leibenluft E, et al. Amygdala en kern sluit in reaksies op die ontvangs en weglating van winste by volwassenes en adolessente. Neuro Image. 2005;25: 1279-1291. [PubMed]
- Eshel N, Nelson EE, Blair RJ, Pine DS, Ernst M. Neurale substraten van keuse keuse by volwassenes en adolessente: Ontwikkeling van die ventrolaterale prefrontale en anterior cingulate kortikale. Neuropsychologia. 2007;45: 1270-1279. [PMC gratis artikel] [PubMed]
- Flavell JH, Feach DR, Chinsky JM. Spontane verbale repetisie in 'n geheue taak as 'n funksie van ouderdom. Kinderontwikkeling. 1966;37: 283-299. [PubMed]
- Galvan A, Hare TA, Parra CE, Penn J, Voss H, Glover G, et al. Vroeër ontwikkeling van die pasiënte met betrekking tot orbitofrontale korteks kan onderliggend wees aan die neem van risiko's in adolessente. Blaar van Neurowetenskap. 2006;26: 6885-6892. [PubMed]
- Galvan A, Hare T, Voss H, Glover G, Casey BJ. Risikobepaling en die adolessente brein: Wie is in gevaar? Ontwikkelingskunde. 2007;10: F8-F14. [PubMed]
- Gardener M, Steinberg L. Peer invloed op risikobepaling, risiko voorkeur en riskante besluitneming in adolessensie en volwassenheid: 'n eksperimentele studie. Ontwikkelingsielkunde. 2005;41: 625-635. [PubMed]
- Giedd JN. Strukturele magnetiese resonansie beelding van die adolessente brein. Annale van die New York Academy of Sciences. 2004;1021: 77-85. [PubMed]
- Giedd JN, Blumenthal J, Jeffries NO, Castellanos FX, Liu H, Zijdenbos A, et al. Brein ontwikkeling tydens kinderjare en adolessensie: 'n MRI-longitudinale MRI-studie. Natuur Neurowetenskap. 1999;2: 861-863.
- Giedd JN, Snell JW, Lange N, Rajapakse JC, Casey BJ, Kozuch PL, et al. Kwantitatiewe magnetiese resonansie beelding van menslike brein ontwikkeling: Ouderdomme 4-18. Serebrale korteks. 1996;6: 551-560. [PubMed]
- Gogtay N, Giedd JN, Lusk L, Hayashi KM, Greenstein D, Vaituzis AC, et al. Dinamiese kartering van menslike kortikale ontwikkeling gedurende die kinderjare deur vroeë volwasse jare. Verrigtinge van die Nasionale Akademie van Wetenskappe van die Verenigde State van Amerika. 2004;101: 8174-8179. [PMC gratis artikel] [PubMed]
- Graber JA, Brooks-Gunn J. Puberty. In: Blechman EA, Brownell KD, redakteurs. Gedragsleer en vroue 'n omvattende handboek. New York, NY: Guilford Press; 1998. pp. 51-58.
- Hare TA, Voss HU, Glover GH, Casey BJ. Die adolessente brein en potensiële risiko vir angs en depressie. 2007a Inhandig vir publikasie.
- Hare TA, Voss HU, Glover GH, Casey BJ. Mededinging tussen prefrontale en subkortiese limbiese stelsels lê onderliggend aan emosionele reaktiwiteit tydens adolessensie. 2007b Inhandig vir publikasie.
- Harnishfeger KK, Bjorkland F. Die ontogenie van inhibisie meganismes: 'n Hernude benadering tot kognitiewe ontwikkeling. In: Howe ML, Pasnek R, redakteurs. Opkomende temas in kognitiewe ontwikkeling. Vol. 1. New York: Springer-Verlag; 1993. pp. 28-49.
- Hikosaka K, Watanabe M. Vertraag aktiwiteit van orbitale en laterale prefrontale neurone van die aap wat wissel met verskillende belonings. Serebrale korteks. 2000;10: 263-271. [PubMed]
- Huttenlocher PR. Sinaptiese digtheid in menslike frontale korteks-Ontwikkelingsveranderinge en effekte van veroudering. Breinnavorsing. 1979;163: 195-205. [PubMed]
- Hyman SE, Malenka RC. Verslawing en die brein: Die neurobiologie van dwang en sy volharding. Natuur Resensies Neurowetenskap. 2001;2: 695-703.
- Jernigan TL, Zisook S, Heaton RK, Moranville JT, Hesselink JR, Braff DL. Magnetic resonance imaging abnormalities in lentikulêre kerne en serebrale korteks in skisofrenie. Argiewe van Algemene Psigiatrie. 1991;48: 811-823.
- Keating DP, Bobbitt BL. Individuele en ontwikkelingsverskille in kognitiewe verwerkingskomponente van verstandelike vermoë. Kinderontwikkeling. 1978;49: 155-167.
- Kelley AE, Schochet T, Landry C. Annale van die New York Academy of Sciences. 2004;1021: 27-32. [PubMed]
- Knutson B, Adams CM, Fong GW, Hommer D. Die verwagting van toenemende monetêre beloning werf selektief kernkwartiere. Blaar van Neurowetenskap. 2001;21: RC159. [PubMed]
- Kuhnen CM, Knutson B. Die neurale basis van finansiële risiko's. Neuron. 2005;47: 763-770. [PubMed]
- Laviola G, Adriani W, Terranova ML, Gerra G. Psigobiologiese risikofaktore vir kwesbaarheid vir psigostimulante in menslike adolessente en diermodelle. Neurowetenschappen en Biobehavioral Reviews. 1999;23: 993-1010. [PubMed]
- Laviola G, Macri S, Morley-Fletcher S, Adriani W. Opsomming van risiko-gedrag in adolessente muise: Psigobiologiese determinante en vroeë epigenetiese invloed. Neurowetenschappen en Biobehavioral Reviews. 2003;27: 19-31. [PubMed]
- Liston C, Watts R, Tottenham N, Davidson MC, Niogi S, Ulug AM, et al. Frontostriatale mikrostruktuur moduleer doeltreffende werwing van kognitiewe beheer. Serebrale korteks. 2005;16: 553-560. [PubMed]
- Luna B, Sweeney JA. Die opkoms van gesamentlike breinfunksie: FMRI-studies van die ontwikkeling van reaksie-inhibisie. Annale van die New York Academy of Sciences. 2004;1021: 296-309. [PubMed]
- Luna B, Thulborn KR, Munoz DP, Merriam EP, Garver KE, Minshew NJ, et al. Volwasse verdeling van breinfunksie ondergaan kognitiewe ontwikkeling. Neuro Image. 2001;13: 786-793. [PubMed]
- Matthews SC, et al. Selektiewe aktivering van die nucleaire accumbens tydens die besluitvorming op die gebied van die neem van risiko's. Neuroreport. 2004;15: 2123-2127. [PubMed]
- Mei JC, Delgado MR, Dahl RE, Stenger VA, Ryan ND, Fiez JA, et al. Gebeurtenisverwante funksionele magnetiese resonansiebeeldvorming van beloningsverwante breinkringe by kinders en adolessente. Biologiese Psigiatrie. 2004;55: 359-366. [PubMed]
- McClure SM, Laibson DI, Loewenstein G, Cohen JD. Afsonderlike neurale stelsels waardeer onmiddellike monetêre belonings. Wetenskap. 2004;306: 503-507. [PubMed]
- Miller EK, Cohen JD. 'N Integratiewe teorie van prefrontale korteksfunksie. Jaarlikse oorsig van Neurowetenskap. 2001;24: 167-202.
- Mischel W, Shoda Y, Rodriguez MI. Vertraging van bevrediging by kinders. Wetenskap. 1989;244: 933-938. [PubMed]
- Monnik CS, McClure EB, Nelson EE, Zarahn E, Bilder RM, Leibenluft E, et al. Adolessente onvolwassenheid in aandagverwante breinbetrokkenheid by emosionele gesigsuitdrukkings. Neuro Image. 2003;20: 420-428. [PubMed]
- Montague PR, Berns GS. Neurale ekonomie en die biologiese substraten van waardering. Neuron. 2002;36: 265-284. [PubMed]
- Moses P, Roe K, Buxton RB, Wong EC, Frank LR, Stiles J. Funksionele MRI van globale en plaaslike verwerking by kinders. Neuro Image. 2002;16: 415-424. [PubMed]
- Munakata Y, Yerys BE. Al nou saam: Wanneer dissosiasies tussen kennis en aksie verdwyn. Psikologiese Wetenskap. 2001;12: 335-337.
- Nagy Z, Westerberg H, Klingberg T. Volwassenheid van wit materie word geassosieer met die ontwikkeling van kognitiewe funksies gedurende die kinderjare. Tydskrif van Kognitiewe Neurowetenskap. 2004;16: 1227-1233. [PubMed]
- O'Doherty J, Kringelbach ML, Rolls ET, Hornak J, Andrews C. Abstracte beloning en strafvoorstellings in die menslike orbitofrontale korteks. Natuur Neurosci. 2001;4: 95-102. [PubMed]
- O'Doherty JP. Beloon voorstellings en beloningsverwante leer in die menslike brein: Insigte van neuroimaging. Huidige opinies in Neurobiologie. 2004;14: 769-776.
- Pascual-Leone JA. 'N Wiskundige model vir oorgang in Piaget se ontwikkelingsfases. Acta Psychologica. 1970;32: 301-345.
- Pecina S, Cagniard B, Berridge KC, Aldridge JW, Zhuang X. Hyperdopaminergiese mutante muise het 'n hoër "wil" maar nie "hou" vir soveel belonings nie. Blaar van Neurowetenskap. 2003;23: 9395-9402. [PubMed]
- Reyna VF, Farley F. Risiko en rasionaliteit in adolessente besluitneming: Implikasies vir teorie, praktyk en openbare beleid. Sielkundige Wetenskap in die Publieke Belang. 2006;7: 1-44.
- Rosenberg DR, Lewis DA. Postnatale veroudering van die dopaminerge innervasie van aap prefrontale en motoriese kortikale: 'n Tyrosien hidroksilase immunohistochemiese analise. Die Tydskrif van Vergelykende Neurologie. 1995;358: 383-400. [PubMed]
- Rubia K, Overmeyer S, Taylor E, Brammer M, Williams SC, Simmons A, et al. Funksionele frontalisasie met ouderdom: Mapping neurodevelopmental trajectories with fMRI. Neurowetenschappen en Biobehavioral Reviews. 2000;24: 13-19. [PubMed]
- Schlaggar BL, Brown TT, Lugar HM, Visscher KM, Miezin FM, Petersen SE. Funksionele neuroanatomiese verskille tussen volwassenes en skoolgaande kinders in die verwerking van enkele woorde. Wetenskap. 2002;296: 1476-1479. [PubMed]
- Schultz W. Gedragsteorieë en die neurofisiologie van beloning. Jaarlikse oorsigte van sielkunde. 2006;57: 87-115.
- Sowell ER, Peterson BS, Thompson PM, Welkom SE, Henkenius AL, Toga AW. Mapping van kortikale verandering oor die menslike lewensduur. Natuur Neurowetenskap. 2003;6: 309-315.
- Sowell ER, Thompson PM, Holmes CJ, Jernigan TL, Toga AW. In vivo getuienis vir post-adolessente breinveroudering in frontale en striatale gebiede. Natuur Neurowetenskap. 1999;2: 859-861.
- Sowell ER, Thompson PM, Toga AW. Mapping veranderinge in die menslike korteks regdeur die lewensduur. Neurowetenskaplike. 2004;10: 372-392. [PubMed]
- Spies LP. Die adolessente brein en ouderdomverwante gedrags manifestasies. Neurowetenschappen en Biobehavioral Reviews. 2000;24: 417-463. [PubMed]
- Steinberg L. Risiko-opname in adolessensie: Wat verander, en hoekom? Annale van die New York Academy of Sciences. 2004;1021: 51-58. [PubMed]
- Steinberg L. Risiko-opname in adolessensie: Nuwe perspektiewe van brein- en gedragswetenskappe. Huidige aanwysings in Sielkundige Wetenskap. 2007;16: 55-59.
- Steinberg L. 'n Sosiale neurowetenskapperspektief op adolessente risikobepaling. Ontwikkelingsoorsig. 2008;28: 78-106. [PMC gratis artikel] [PubMed]
- Tamm L, Menon V, Reiss AL. Volwasse breinfunksie geassosieer met reaksie-inhibisie. Tydskrif van die Amerikaanse Akademie vir Kinder- en Adolessente Psigiatrie. 2002;41: 1231-1238. [PubMed]
- Thomas KM, Hunt RH, Vizueta N, Sommer T, Durston S, Yang Y, et al. Bewyse van ontwikkelingsverskille in implisiete volgorde leer: 'n FMRI studie van kinders en volwassenes. Tydskrif van Kognitiewe Neurowetenskap. 2004;16: 1339-1351. [PubMed]
- Turkeltaub PE, Gareau L, Blomme DL, Zeffiro TA, Eden GF. Ontwikkeling van neurale meganismes vir lees. Natuur Neurowetenskap. 2003;6: 767-773.
- Volkow ND, Li TK. Dwelmverslawing: Die neurobiologie van gedrag het verkeerd gegaan. Natuur Resensies Neurowetenskap. 2004;5: 963-970.
- Yurgelun-Todd D. Emosionele en kognitiewe veranderinge tydens adolessensie. Huidige Advies in Neurobiologie. 2007;17: 251-257. [PubMed]
- Zald DH, Boileau I, El-Dearedy W, Gunn R, McGlone F, Dichter GS, et al. Dopamien-oordrag in die menslike striatum tydens monetêre beloningstake. Blaar van Neurowetenskap. 2004;24: 4105-4112. [PubMed]
