Skakel na abstrak - J Adolesc. 2019 Februarie 9; 72: 10-13. doi: 10.1016 / j.adolescence.2019.01.006.
Abstract
INLEIDING: Die fokus van hierdie kort literatuuroorsig is om te ondersoek of daar 'n verband bestaan tussen die unieke anatomiese en fisiologiese paradigmas van die adolessente brein en 'n verhoogde sensitiwiteit vir seksueel eksplisiete materiaal.
METODES: Die EBSCO-navorsingsdatabasisse is deursoek deur gebruik te maak van die volgende sleutelterme: adolessensie, adolessente breinontwikkeling, neuroplastisiteit, seksueel eksplisiete materiaal, seksualisering en pornografie.
RESULTATE: Die literatuur het verskeie komponente van die adolessente brein gemerk wat anders as die volwasse brein is. Dit sluit in: 'n onvolwasse prefrontale korteks en oormatige limbiese en striatale bane, verhoogde tydperk vir neuroplastisiteit, ooraktiewe dopamienstelsel, 'n uitgesproke HPA-as, verhoogde vlakke van testosteroon en die unieke impak van steroïedhormone. Die fisiologiese reaksie op seksueel eksplisiete materiaal word afgebaken. Die oorvleueling van sleutelareas wat verband hou met die unieke adolessente breinontwikkeling en seksueel eksplisiete materiaal, is opmerklik. 'N Werkmodel opsomming wat die reaksie van die volwasse en adolessente brein vergelyk met dieselfde seksueel eksplisiete stimulus, word uiteengesit.
GEVOLGTREKKINGS: Die literatuur dui daarop dat die adolessente brein wel meer sensitief is vir seksueel eksplisiete materiaal, maar weens 'n gebrek aan empiriese studies kan hierdie vraag nie finaal beantwoord word nie. Voorstelle vir toekomstige navorsing word gegee om die werk verder te bevorder in hierdie toepaslike veld van vandag.
SLEUTELWOORDE: Adolessensie; Adolessente breinontwikkeling; Neuroplasticity; pornografie; Sexualization; Seksueel eksplisiete materiaal
PMID: 30754014
DOI: 10.1016 / j.adolescence.2019.01.006
Unieke paradigmas van die adolessente brein
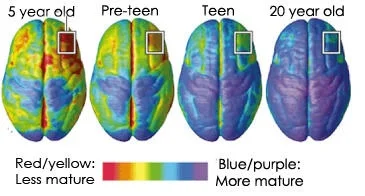
Die fokus van hierdie kort literatuuroorsig is om te ondersoek of daar 'n verband bestaan tussen die unieke anatomiese en fisiologiese paradigmas van die adolessente brein en 'n verhoogde sensitiwiteit vir seksueel eksplisiete materiaal. Die EBSCO-navorsingsdatabasis is deursoek met behulp van die volgende sleutelterme: adolessensie, adolessente breinontwikkeling, neuroplastisiteit, seksueel eksplisiete materiaal, seksualisering, pornografie. Adolessensie is die periode tussen kinderjare en volwassenheid wat deur veranderinge in fisieke, sielkundige en sosiale ontwikkeling omring word (Ernst, Pine, & Hardin, 2006).
Die unieke paradigmas van die adolessente brein sluit die volgende in: 1) 'n Onvolwasse prefrontale korteks en oorreaksiewe limbiese en striatale stroombane (Dumontheil, 2016; Somerville & Jones, 2010; Somerville, Hare, & Casey, 2011; Van Leijenhorst et al. , 2010; Vigil et al., 2011); 2) 'n Verhoogde periode vir neuroplastisiteit (McCormick & Mathews, 2007; Schulz & Sisk, 2006; Sisk & Zehr, 2005; Vigil et al., 2011); 3) Ooraktiewe dopamienstelsel (Andersen, Rutstein, Benzo, Hostetter, & Teicher, 1997; Ernst et al., 2005; Luciana, Wahlstrom, & White, 2010; Somerville & Jones, 2010; Wahlstrom, White, & Luciana, 2010) ; 4) 'n uitgesproke HPA-as (Dahl & Gunnar, 2009; McCormick & Mathews, 2007; Romeo, Lee, Chhua, McPherson, & McEwan, 2004; Walker, Sabuwalla, & Huot, 2004); 5) Verhoogde vlakke van testosteroon (Dorn et al., 2003; Vogel, 2008; Mayo Clinic / Mayo Medical Laboratories, 2017); en 6) Die unieke impak van steroïedhormone (kortisol en testosteroon) op breinontwikkeling tydens die organisatoriese venster van adolessensie (Brown & Spencer, 2013; Peper, Hulshoff Pol, Crone, Van Honk, 2011; Sisk & Zehr, 2005; Vigil et. al., 2011).
Blakemore en kollegas het die veld in adolessente breinontwikkeling gelei en het aangevoer dat die tienerjare as 'n sensitiewe tydperk beskou moet word as gevolg van die dramatiese breinreorganisasie wat plaasvind (Blakemore, 2012). Die areas van die brein wat die meeste verandering ondergaan tydens adolessensie sluit in interne beheer, multi-tasking en beplanning (Blakemore, 2012).
Blakemore en Robbins (2012) het adolessensie gekoppel aan risikobesluitende besluitneming en hierdie eienskap toegeskryf aan die dissosiasie tussen die relatief stadig, lineêre ontwikkeling van impulsbeheer en responsinhibisie tydens adolessensie teenoor die nie-lineêre ontwikkeling van die beloningstelsel, wat dikwels hiperresponsief is vir belonings in adolessensie.
Seksueel eksplisiete materiaal
Seksueel eksplisiete materiaal aktiveer die amygdala van die limbiese stelsel (Ferretti et al., 2005; Karama et al., 2002; Redoute et al., 2000; Walter et al., 2008). Aktivering van die amygdala begin die volgende: 1) die hipotalamus aktiveer neurone in die breinstam en rugmurg wat die simpatiese verdeling van die outonome senuweestelsel begin, wat lei tot sistemiese vrystelling van epinefrien en norepinefrien; 2) die hipotalamus stimuleer die pituïtêre klier, wat lei tot kortisol vrystelling via die hipotalamus-pituïtêre-bynier (HPA) -as, en testosteroon vrystelling via die hipotalamus-pituïtêre-gonadale (HPG) as (Viau, 2002); 3) word die nucleus accumbens geaktiveer via dopamien. Vir 'n omvattende oorsig van die amygdala en sy innervasies en regulering van somatiese prosesse, sien Mirolli, Mannella, en Baldassarre (2010). Die funksie van die prefrontale korteks verminder, en die funksie van die basale ganglia word verhoog as gevolg van die vrylating van neurotransmitters (Arnsten, 2009, Hanson et al., 2012; Radley, 2005).
Beide ongereelde en gereelde gebruik van pornografiese webwerwe was aansienlik geassosieer met sosiale wanaanpassing onder Griekse adolessente (Tsitsika et al., 2009). Die gebruik van pornografie het daartoe bygedra om afslag te vertraag, of die neiging van 'n individu om toekomstige uitkomste te verdiskonteer ten gunste van onmiddellike belonings (Negash, Sheppard, Lambert, & Fincham, 2016). Negash en kollegas het 'n steekproef met 'n gemiddelde ouderdom van 19 en 20 jaar gebruik, wat deur die skrywer beklemtoon word as biologies beskou as adolessente. Hulle het herhaal dat hul verslawing nie verslawend of dwangmatig is nie, maar dat daar steeds veranderinge in besluitnemingsprosesse getoon word.
Die gebruik van pornografie is gekoppel aan stimulus en neuroplastisiteit van die mesolimbiese dopaminerge beloningstelsel (Hilton, 2013). MRI-skanderings het 'n beduidende negatiewe verband gevind tussen gerapporteerde pornografie-ure per week en grysstofvolume in die regte caudate en funksionele verbinding met die dorsolaterale prefrontale korteks (Kuhn & Gallinat, 2014). Pornografie kan die oorsaak van hierdie neuroplastisiteit wees, maar 'n voorwaarde wat die gebruik van pornografie lonender maak, kan nie uitgesluit word nie.
Werkmodel opsomming
Ons stel 'n werkmodel opsomming voor, met inagneming van die unieke paradigmas van die adolessente brein en die eienskappe van seksueel eksplisiete materiaal. Die oorvleueling van sleutelareas wat met die unieke adolessente brein en seksueel eksplisiete materiaal verband hou, is opmerklik.
By blootstelling aan seksueel eksplisiete materiaal sou die stimulasie van die amygdala en die HPA-as by die adolessent verbeter word, vergeleke met die volwassene. Dit sou lei tot 'n meer uitgesproke inkorting van die prefrontale korteks en verhoogde aktivering van die basale ganglia by die adolessent. Hierdie toestand sal die uitvoerende funksie, wat inhibisie en selfbeheersing insluit, in gevaar stel en die impulsiwiteit verhoog. Omdat die adolessent se brein nog ontwikkel, is dit meer bevorderlik vir neuroplastisiteit. Die prefrontale korteks wat 'off-line' gaan, so te sê, dryf die subtiele herbedrading aan wat die ontwikkeling van subkortikale bevoordeel. As die neuroplastisiteitswanbalans met verloop van tyd voortduur, kan dit lei tot 'n relatief verswakte kortikale stroombaan ten gunste van 'n meer dominante subkortikale stroombaan, wat die adolessent kan voorspel vir voortgesette selfbevrediging en impulsiwiteit. Die kern van die adolessent, of die plesier sentrum van die brein, het 'n oordrewe stimulasie in vergelyking met die volwassene. Die verhoogde vlakke van dopamien kan vertaal word in aangevulde emosies wat verband hou met dopamien, soos plesier en drang (Berridge, 2006; Volkow, 2006).
As gevolg van die puberteitstuwing van testosteroon, sal die vlak daarvan ook vergroot word in vergelyking met die volwassene. Hierdie toename in testosteroon kan lei tot hoër neigings van aggressie (Banks & Dabbs, 1996; Goetz et al., 2014; Nelson, Leibenluft, McClure, & Pine, 2005; Schulz & Sisk, 2006) en seksuele afwagting (Amstislavskaya & Popova, 2004; Bonilla – Jaime, Vazquez-Palacios, Arteaga-Silva, & Retana-Marquez, 2006; Exton et al., 1999; Redoute et al., 2000; Stoleru et al., 1999;).
Vanweë die organisatoriese ontwikkelingsvenster tydens adolessensie, sal kortisol en testosteroon 'n unieke invloed hê op die breinorganisasie of die inherente lewensvatbaarheid van verskillende neurale bane. Hierdie effek sal nie by volwassenes gevind word nie, omdat hierdie spesifieke organisasievenster gesluit is. Chroniese blootstelling aan kortisol het die potensiaal om gedurende die adolessente organisasietydperk neuroplastisiteit aan te dryf wat tot gevolg het dat kognitiewe funksies en stresbestendigheid tot in volwassenheid lei (McEwen, 2004; Tsoory & Richter-Levin, 2006; Tsoory, 2008; McCormick & Mathews, 2007; 2010). Die robuustheid van die amygdala post puberteit, hang ten minste gedeeltelik af van die omvang van blootstelling aan testosteroon tydens die kritieke ontwikkelingsvenster vir adolessente (De Lorme, Schulz, Salas-Ramirez, & Sisk, 2012; De Lorme & Sisk, 2013; Neufang et. al., 2009; Sarkey, Azcoitia, Garcia- Segura, Garcia-Ovejero, & DonCarlos, 2008). 'N Robuuste amygdala word gekoppel aan verhoogde vlakke van emosionaliteit en gekompromitteerde selfregulering (Amaral, 2003; Lorberbaum et al., 2004; De Lorme & Sisk, 2013).
Bespreking en toekomstige rigting
Hierdie vraestel wou die akademiese gesprek begin: Kan adolessente meer sensitief wees vir seksueel eksplisiete materiaal as gevolg van die unieke anatomiese en fisiologiese paradigmas van die adolessente brein? Die huidige literatuur dui daarop dat die adolessente brein wel meer sensitief is vir seksueel eksplisiete materiaal, maar weens die gebrek aan empiriese studies kan hierdie vraag nie finaal beantwoord word nie. Die uitdaging om deur etiese oorwegings vir gekontroleerde studies te werk, is ook 'n beduidende, maar verstaanbare, hindernis vir wetenskaplike vordering in hierdie veld.
As 'n aanvang beveel ons aan om bevolkingsstudies te doen met behulp van selfassesseringsopnames wat navraag doen oor gedragstendense voor die aanvanklike blootstelling aan seksueel eksplisiete materiaal en na verskillende grade van blootstelling. Opnames kan ook aan ouers gegee word om vas te stel of die ouer-kind verhouding 'n beduidende faktor is teenoor die self-effektiwiteit van kinders se gesondheid (en skolastiese prestasie).
Nog 'n navorsingsveld wat oorweeg moet word, is die rol van tegnologie as 'n poort vir adolessente om blootgestel te word aan seksueel eksplisiete materiaal. Aangesien werklike sosiale media gebruik opgespoor kan word en vergelyk kan word met waargenome gebruik, sal opnames wat die deelnemers vra om hul tegnologieverbruik en blootstelling aan seksueel eksplisiete materiaal self te evalueer, 'n redelik reguit studie wees.
Uiteindelik kan 'n belangrike bydrae tot hierdie veld 'n longitudinale studie wees wat sou behels dat 'n groep kinders deur adolessensie en volwassenheid gevolg word, tesame met gedokumenteerde mediese geskiedenis en die verkryging van anatomiese, fisiologiese en sielkundige data van gereelde geskeduleerde strukturele en funksionele MRI, en / of PET beeldvorming.
Die ontwerp van noukeurige etiese studies om die effek van seksueel eksplisiete materiaalblootstelling op die adolessente brein te ondersoek, is 'n noodsaaklike stap om die veranderlikheid van volwasse ervarings met seksueel eksplisiete materiaal te verstaan.
Verwysings
- Amaral, DG (2003). Die amygdala, sosiale gedrag en gevaaropsporing. Annale van die New York Akademie van Wetenskappe, 1000, 337-347. https://doi.org/10.1196/
annals.1280.015. - Amstislavskaya, TG, & Popova, NK (2004). Vroulike geïnduseerde seksuele opwinding by manlike muise en rotte: Gedrags- en testosteroonreaksie. Hormone en gedrag, 46,
544-550. - Andersen, SL, Rutstein, M., Benzo, JM, Hostetter, JC, & Teicher, MH (1997). Seksverskille in oorproduksie en eliminasie van dopamienreseptore. NeuroReport,
8, 1495–1498. https://doi.org/10.1097/00001756-199704140-00034. - Arnsten, AFT (2009). Spanningsaanwysingspaaie wat die prefrontale korteksstruktuur en funksie benadeel. Natuur Resensies Neurowetenskap, 10 (6), 410-422. https://doi.org/
10.1038 / nrn2648. - Banks, T., & Dabbs, JM, Jr. (1996). Speeksel testosteroon en kortisol in 'n misdadige en gewelddadige stedelike subkultuur. The Journal of Social Psychology, 136 (1), 49–56.
https://doi.org/10.1080/00224545.1996.9923028. - Berridge, KC (2006). Die debat oor dopamien se rol in beloning: Die saak vir aansporing van aansporing. Psigofarmakologie, 191, 391–431. https://doi.org/10.1007/
s00213-006-0578-x. - Blakemore, S. (2012). Ontwikkeling van die sosiale brein in adolessensie. Tydskrif van die Royal Society of Medicine, 105, 111-116. https://doi.org/10.1258/jrsm.2011.
110221. - Blakemore, S., & Robbins, TW (2012). Besluitneming in die adolessente brein. Nature Neuroscience, 15 (9), 1184–1191. https://doi.org/10.1038/nn.3177.
- Bonilla-Jaime, H., Vazquez-Palacios, G., Arteaga-Silva, M., & Retana-Marquez, S. (2006). Hormonele reaksies op verskillende seksueel verwante toestande by manlike rotte.
Hormone en Gedrag, 49, 376-382. - Brown, GR, & Spencer, KA (2013). Steroïedhormone, spanning en die adolessente brein: 'n vergelykende perspektief. Neurowetenskap, 249, 115–128. https://doi.org/10.
1016 / j.neuroscience.2012.12.016. - Dahl, RE, & Gunnar, MR (2009). Verhoogde stresresponsiwiteit en emosionele reaktiwiteit tydens puberteitrypheid: Implikasies vir psigopatologie.
Ontwikkeling en Psigopatologie, 21, 1-6. https://doi.org/10.1017/S0954579409000017. - De Lorme, KC, Schulz, KM, Salas-Ramirez, KY, & Sisk, CL (2012). Pubertale testosteroon organiseer die plaaslike volume en die neuronale getal binne die mediale
amygdala van volwasse manlike Siriese hamsters. Breinnavorsing, 1460, 33-40. https://doi.org/10.1016/j.brainres.2012.04.035. - De Lorme, KC, & Sisk, CL (2013). Pubertale testosteroon programmeer konteks-toepaslike agonistiese gedrag en gepaardgaande neurale aktiveringspatrone in Siriese mans
hamsters. Sielkunde en gedrag, 112–113, 1–7. https://doi.org/10.1016/j.physbeh.2013.02.003. - Dorn, LD, Dahl, RE, Williamson, DE, Birmaher, B., Axelson, D., Perel, J., et al. (2003). Ontwikkelingsmarkers in adolessensie: Implikasies vir studie van pubertal
prosesse. Joernaal van Jeug en Adolessensie, 32 (5), 315-324. - Dumontheil, I. (2016). Adolessente breinontwikkeling. Huidige opinie in Gedragswetenskappe, 10, 39-44. https://doi.org/10.1016/j.cobeha.2016.04.012.
- Ernst, M., Nelson, EE, Jazbec, S., McClure, EB, Monk, CS, Leibenluft, E., et al. (2005). Amygdala en nukleus kom in reaksie op ontvangs en weglating van
winste by volwassenes en adolessente. NeuroImage, 25, 1279-1291. https://doi.org/10.1016/j.neuroimage.2004.12.038. - Ernst, M., Pine, DS, & Hardin, M. (2006). Triadiese model van die neurobiologie van gemotiveerde gedrag in adolessensie. Psychological Medicine, 36 (3), 299–312.
- Exton, MS, Bindert, A., Kruger, T., Scheller, F., Hartmann, U., & Schedlowski, M. (1999). Kardiovaskulêre en endokriene veranderinge na masturbasie
orgasme in vroue. Psigosomatiese Geneeskunde, 61, 280-289. - Ferretti, A., Caulo, M., Del Gratta, C., Di Matteo, R., Merla, A., Montorsi, F., et al. (2005). Dinamiek van manlike seksuele opwinding: Duidelike komponente van breinaktivering
geopenbaar deur fMRI. NeuroImage, 26, 1086-1096. https://doi.org/10.1016/j.neuromiage.2005.03.025. - Goetz, SMM, Tang, L., Thomason, ME, Diamond, MP, Hariri, AR, & Carre, JM (2014). Testosteroon verhoog vinnig die neurale reaktiwiteit teen bedreigings by gesonde
mans: 'n Nuwe twee-stap farmakologiese uitdagingsparadigma. Biologiese Psigiatrie, 76, 324-331. - Hanson, JL, Chung, MK, Avants, BB, Rudolph, KD, Shirtcliff, EA, Gee, JC, et al. (2012). Strukturele variasies in prefrontale korteks bemiddel die verhouding
tussen vroeë kinderjare stres en ruimtelike werkgeheue. Die Tydskrif vir Neurowetenschappen, 32 (23), 7917-7925. https://doi.org/10.1523/jneurosci.0307-12.2012. - Hilton, DL (2013). Pornografieverslawing - 'n supranormale stimulus wat beskou word in die konteks van neuroplastisiteit. Sosio-affektiewe neurowetenskap en sielkunde, 3, 20767.
https://doi.org/10.3402/snp.v3i0.20767. - Karama, S., Lecours, AR, Leroux, J., Bourgouin, P., Beaudoin, G., Joubert, S., et al. (2002). Gebiede van breinaktivering by mans en vroue tydens die besigtiging van erotiese
film uittreksels. Menslike breinkaarte, 16, 1-13. https://doi.org/10.1002/hbm.10014. - Kuhn, S., & Gallinat, J. (2014). Breinstruktuur en funksionele konneksie wat verband hou met die verbruik van pornografie. JAMA Psigiatrie. https://doi.org/10.1001/
jamapsychiatry.2014.93. - Lorberbaum, JP, Kose, S., Johnson, MR, Arana, GW, Sullivan, LK, Hamner, MB, et al. (2004). Neurale korrelate van spraakvoorkomende angs in algemene
sosiale fobie. NeuroReport, 15 (18), 2701-2705. - Luciana, M., Wahlstrom, D., & White, T. (2010). Neurobehavioral bewyse vir veranderinge in dopamienstelselaktiwiteit tydens adolessensie. Neurowetenskap en biogedrag
Resensies, 34 (5), 631-648. https://doi.org/10.1016/j.neubiorev.2009.12.007. - Mayo Clinic (2017). Mayo mediese laboratoriums. Toets ID: TTFB testosteroon, totaal, biobeskikbaar, en vry, serum. Ontsluit van http://www.mayomedicallaboratories.com/
toets-katalogus / Kliniese + en + Interpretief / 83686. - McCormick, CM, & Mathews, IZ (2007). HPA-funksie in adolessensie: rol van geslagshormone in die regulering daarvan en die blywende gevolge van blootstelling aan stressors. Farmakologie, biochemie en gedrag, 86, 220–233. https://doi.org/10.1016/j.pbb.2006.07.012.
- McCormick, C., M., & Mathews, IZ (2010). Adolessente ontwikkeling, hipotalamus-pituïtêre-bynierfunksie en programmering van leer en geheue vir volwassenes.
- Vordering in neuro-psigofarmakologie en biologiese psigiatrie, 34, 756–765. https://doi.org/10.1016/j.pnpbp.2009.09.019.
- McEwen, B. (2004). Beskerming en beskadiging van akute en chroniese stres. Annale van die New York Akademie van Wetenskappe, 1032, 1-7. https://doi.org/10.1196/annals.
1314.001. - Mirolli, M., Mannella, F., & Baldassarre, G. (2010). Die rolle van die amygdala in die affektiewe regulering van liggaam, brein en gedrag. Connection Science, 22, 215-245.
https://doi.org/10.1080/09540091003682553. - Negash, S., Sheppard, N., Lambert, NM, & Fincham, FD (2016). Later beloning vir huidige plesier: verbruik van pornografie en vertraagde afslag. Die
Tydskrif van Geslagsnavorsing, 53 (6), 689-700. https://doi.org/10.1080/00224499.2015.1025123. - Nelson, EE, Leibenluft, E., McClure, EB, & Pine, DS (2005). Die sosiale heroriëntering van adolessensie: 'n Neurowetenskaplike perspektief op die proses en die verband daarvan met
psigopatologie. Sielkundige Geneeskunde, 35, 163-174. https://doi.org/10.1017/S0033291704003915. - Neufang, S., Specht, K., Haus Mann, M., Gunturkun, O., Herpertz-Dahlmann, B., Fink, GR, et al. (2009). Seksverskille en die impak van steroïedhormone op die
ontwikkelende menslike brein. Serebrale Cortex, 19, 464-473. https://doi.org/10.1093/cercor/bhn100. - Peper, JS, Hulshoff Pol, HE, Crone, EA, & Van Honk, J. (2011). Geslagsteroïede en breinstruktuur by puberteitseuns en -dogters: 'n mini-oorsig van neuro-beeldingstudies.
Neurowetenschappen, 191, 28-37. - Radley, J. (2005). Herhaalde stres en strukturele plastisiteit in die brein. Aging Research Reviews, 4, 271-287. https://doi.org/10.1016/j.arr.2005.03.004.
- Redoute, J., Stoleru, S., Gregoire, M., Costes, N., Cinotti, L., Lavenne, F., et al. (2000). Breinverwerking van visuele seksuele stimuli by menslike mans. Menslike breinkaarte,
11, 162-177. - Romeo, RD, Lee, SJ, Chhua, N., McPherson, CR, & McEwen, BS (2004). Testosteroon kan nie 'n volwasse-agtige stresrespons by prepubertale manlike rotte aktiveer nie.
Neuroendocrinology, 79, 125-132. https://doi.org/10.1159/000077270. - Sarkey, S., Azcoitia, I., Garcia-Segura, LM, Garcia-Ovejero, D., & DonCarlos, LL (2008). Klassieke androgeen reseptore op nie-klassieke plekke in die brein. Hormone
en Gedrag, 53, 753-764. - Schulz, KM, & Sisk, CL (2006). Pubertehormone, die adolessente brein en die rypwording van sosiale gedrag: lesse uit die Siriese hamster. Molekulêre en
Sellulêre Endokrinologie, 254-256, 120-126. https://doi.org/10.1016/j.mce.2006.04.025. - Sisk, CL, en Zehr, JL (2005). Puberteitse hormone organiseer die adolessente brein en gedrag. Frontiers in Neuroendocrinology, 26, 163–174. https://doi.org/10.1016/
j.yfrne.2005.10.003. - Somerville, LH, Hare, T., & Casey, BJ (2011). Frontostriatale rypwording voorspel kognitiewe beheermaatreëls tot aptytlike leidrade by adolessente. Tydskrif vir kognitiewe
Neurowetenschappen, 23, 2123-2134. https://doi.org/10.1162/jocn.2010.21572. - Somerville, LH, & Jones, R. (2010). Tyd van verandering; gedrags- en neurale korrelasies van adolessente sensitiwiteit vir aptytwekkende en aversiewe omgewingsaanwysings. Brein
en Cognition, 72 (1), 124-133. https://doi.org/10.1016/j.bandc.2009.07.003. - Stoleru, S., Gregoire, MC, Gerard, D., Decety, J., Lafarge, E., Cinotti, L., et al. (1999). Neuroanatomiese korrelate van visueel opgewekte seksuele opwekking in menslike mans.
Argiewe van seksuele gedrag, 28, 1-21. - Tsitsika, A., Critselis, E., Kormas, G., Konstantoulaki, E., Constantopoulos, A., & Kafetzis, D. (2009). Adolessente pornografiese webwerf gebruik: 'n multivariaat
regressie-analise van die voorspellende faktore van gebruik en psigososiale implikasies. Cyberpsigologie en Gedrag, 12 (5), 545-550. https://doi.org/10.1089/cpb.
2008.0346. - Tsoory, M. (2008). Blootstelling aan stressors tydens jeugdigheid ontwrig ontwikkelingsverwante veranderinge in die PSA-NCAM na NCAM uitdrukkingsverhouding: Potensiële relevansie vir
bui en angsversteurings. Neuropsychopharmacology, 33, 378-393. https://doi.org/10.1038/sj.npp.1301397. - Tsoory, M., & Richter-Levin, G. (2006). Leer onder spanning by die volwasse rot word op verskillende maniere beïnvloed deur 'jeugdige' of 'adolessente' spanning. Internasionale Tydskrif vir
Neuropsigofarmakologie, 9 (6), 713-728. https://doi.org/10.1017/S1461145705006255. - Van Leijenhorst, L., Zanolie, K., Van Meel, CS, Westenberg, PM, Rombouts, SARB, & Crone, EA (2010). Wat motiveer die adolessent? Breinstreke
bemoedigende sensitiwiteit oor adolessensie. Serebrale Cortex, 20, 61-69. https://doi.org/10.1093/cercor/bhp078. - Viau, V. (2002). Funksionele kruisbespreking tussen die hipotalamus-pituïtêre-gonadale en bynierasse. Blaar van Neuroendokrinologie, 14, 506-513.
- Vigil, P., Orellana, RF, Cortes, ME, Molina, CT, Switzer, BE, & Klaus, H. (2011). Endokriene modulasie van die adolessente brein: 'n oorsig. Tydskrif vir Pediatriese en
Adolessente Ginekologie, 24 (6), 330-337. https://doi.org/10.1016/j.jpag.2011.01.061. - Vogel, G. (2008). Tyd om groot te word. Wetenskap Nou, 2008 (863), 1.
- Volkow, N. (2006). Kokaïen leidrade en dopamien in dorsale striatum: Meganisme van drang in kokaïenverslawing. Die Tydskrif vir Neurowetenschappen, 26 (24), 6583-6588.
https://doi.org/10.1523/JNEUROSCI.1544-06.2006. - Wahlstrom, D., White, T., & Luciana, M. (2010). Neurobehavioral bewyse vir veranderinge in dopamienstelselaktiwiteit tydens adolessensie. Neurowetenskap en biogedrag
Resensies, 34, 631-648. https://doi.org/10.1016/j.neubiorev.2009.12.007. - Walker, EF, Sabuwalla, Z., & Huot, R. (2004). Pubertale neuromaturasie, stresgevoeligheid en psigopatologie. Ontwikkeling en Psigopatologie, 16, 807–824.
https://doi.org/10.1017/S0954579404040027. - Walter, M., Bermpohl, F., Mouras, H., Schiltz, K., Tempelmann, C., Rotte, M., et al. (2008). Onderskeidende spesifieke seksuele en algemene emosionele effekte in fMRI-subkortiese en kortikale opwinding tydens erotiese beeldbesigtiging. NeuroImage, 40, 1482-1494. https://doi.org/10.1016/j.neuroimage.2008.01.040.
