- 1Centro Interdisciplinario de Neurociencias de Valparaiso, Elmlər Fakültəsi, Universidad de Valparaiso, Valparaiso, Çili
- 2Departamento de Neurociencia, Tibb Universiteti, Universidad de Çili, Santiaqo, Çili
- 3Núcleo Milenio NUMIND Nöropsikiyatrik Bozuklukların Biologiyası, Universidad de Valparaiso, Valparaiso, Çili
- 4Biologiya Elmləri Fakültəsi, Hüceyrə və Molekulyar Biologiya Bölümü, Pontifikiya Universidad Católica de Chile, Santiago, Çili
- 5Əczaçılıq şöbəsi və Kimyəvi Fakültələrarası Nevrologiya Mərkəzi, Pontifikiya Universidad Católica de Chile, Santiago, Çili
Məqsədyönlü davranışların gücü midbrain dopamin neyronları tərəfindən tənzimlənir. Narkomaniya və obsesif-kompulsif pozğunluqda dopaminergik sxemlərin pozğunluqları müşahidə olunur. Kompulsif davranış, hər iki pozğunluğun paylaşdığı bir xüsusiyyətdir, bu da yüksək dopamin nörotransmissiyası ilə əlaqələndirilir. Midbrain dopamin neyronlarının fəaliyyəti, əsas etibarilə neyronların atışını, həmçinin dopamin sintezini və sərbəst qalmasını azaldan D2 reseptorları (D2R) vasitəsilə dopaminin homeostatik təsiri ilə tənzimlənir. Dopamin ötürülməsi, digərləri arasında kappa opioid sistemi kimi heteroloji nörotransmitter sistemləri ilə də tənzimlənir. Kappa opioid sistemi və dopamin ötürülməsinə təsiri haqqında mövcud biliklərimizin çoxu beyin xəstəliklərinin preklinik heyvan modellərindən gəlir. 1988-ci ildə serebral mikrodializdən istifadə edərək Kappa Opioid Reseptorlarının (KOR) kəskin aktivləşməsi striatumdakı dopaminin sinaptik səviyyəsini azaldır. KOR-un bu inhibitor təsiri, KOR agonistlərinin məcburi dərman qəbulu üçün farmakoloji terapiya kimi istifadə edilməsinə səbəb olan sui-istifadə dərmanlarının dopamin sərbəst buraxılmasına təsirini asanlaşdırır. Təəccüblüdür ki, 30 il sonra KOR antaqonistlərinə narkotik asılılığını müalicə etmək təklif olunur. Bu illər ərzində paradiqmanın kəskin dəyişməsinə səbəb olan nə ola bilər? Toplanmış sübutlar KOR-un sinaptik dopamin səviyyəsinə təsirinin KOR aktivləşdirmə tezliyindən və dopamin neyronlarına daxil olan digər stimullarla vaxtından, cinsindən və növ fərqlərindən asılı olaraq mürəkkəb olduğunu irəli sürdü. Kəskin təsirinin əksinə olaraq, xroniki KOR aktivləşdirmə, dopamin nörotransmesi və dopamin vasitəçiliyi ilə davranışları asanlaşdırır. Kəskin və xroniki KOR aktivləşdirilməsinə qarşı olan əks hərəkətlər, sui-istifadə dərmanlarına məruz qaldıqda, ilk köməkçi və gecikmiş mükafatlandırıcı təsir ilə əlaqələndirilmişdir. D2R-nin təkrar aktivləşdirilməsi nəticəsində yaranan kompulsif davranışlar, eyni zamanda dopamin və həssaslaşmış D2R səviyyəsinin aşağı düşməsi ilə əlaqəli olan KOR-un davamlı aktivləşməsi ilə də təsir göstərir. Beləliklə, vaxtından asılı KOR aktivləşdirmə, əsaslandırılmış davranışların tənzimlənməsinə təsir edən birbaşa dofamin səviyyələrinə təsir göstərir. Bu araşdırmada kappa opioid sisteminin kompulsif davranışların dopaminergik əlaqələrinə verdiyi töhfə təhlil edilmişdir.
giriş
Kompulsiv Davranışlarda Dopaminerjik Sistem
Məcburiyyət, mənfi nəticələrə baxmayaraq, məlum nəticə ilə adi bir hərəkəti həyata keçirmək üçün özünü dayandırmağın mümkünsüzlüyüdür (Robbins et al., 2012). Kompulsif davranışlar digər psixiatrik xəstəliklər arasında obsesif-kompulsif pozğunluğun (OCD) və narkomaniyanın əlamətidir. Yoxlama davranışı, müəyyən bir rutinin daimi təkrarlanması, stereotip və ya ritualistik bir şəkildə xarakterizə olunan obsesif-kompulsif spektr pozğunluqlarında çox yaygındır (Williams et al., 2013). Normal davranışların geniş çeşidi (məsələn, yoxlama, təmizlik, əllərin yuyulması və s.) OKB xəstələrində kompulsiv hala çevrilə bilər və ümumiyyətlə narahatlıq doğuran obsesif və narahat düşüncələrə cavab olaraq ortaya çıxır. Eynilə, sui-istifadə dərmanlarını istəməsi və istehlak etməsi narkomanlarda məcburi hala gəlir. OKB-də olduğu kimi, təcrübəli narkotik istifadəçilərində də məcburi narkotik istehlakına səbəb olan narahatlıq əsas rol oynayır. Mükafat və cəza emalında eyni pozuntular hər iki şərtdə də müşahidə olunur (Figee və digərləri, 2016) bəzi müəlliflərin davranış asılılığı olaraq OKB-nin müzakirəsinə səbəb oldu (Holden, 2001).
Məcburi davranışa səbəb olan bir mexanizm bağımlılığın təşviqedici-həssaslaşdırma nəzəriyyəsi çərçivəsində qurulmuşdur ki, bu da narkotik üçün gücləndirilmiş bir motivasiya ("istəyən") asılılıq zamanı inkişaf etmiş xoşagəlməz ("bəyənmək") effekti yaratmadan inkişaf edir (Berridge və digərləri, 1989; Berridge və Robinson, 2016). Mükafat / motivasiya dövrünün davamlı həssaslaşması dərman axtaranlarla əlaqəli həvəsləndirici həssaslığın təşviqində iştirak edir. Mükafat / motivasiya dövrəsi orta beyin dopamin neyronlarından ibarətdir əsasən nigra (SN) və müvafiq olaraq striatumun dorsal və ventral səviyyələrini hədəf alan ventral tegramal bölgə (VTA). Vtrtral striatuma və ya nüvə böyüməsinə (NAc) aid olan dopamin neyronları ənənəvi olaraq məqsədyönlü davranışlarla əlaqəli olur, halbuki dorsal striatuma istiqamət verən dopamin neyronları vərdişlərin əldə edilməsi ilə əlaqələndirilmişdir (Everitt və Robbins, 2005; Müdrik, 2009; Yager və digərləri, 2015; Volkow və digərləri, 2017).
Potensial asılılıq dərmanı sabit dozanın təkrar qəbulu nəticəsində yaranan lokomotor fəaliyyətdə tədricən artım kimi mükafatlandırma / motivasiya dövrəsinin həssaslığı müşahidə olunur.Pierce və Kalivas, 1997; Robinson və Berridge, 2001). Lokomotor həssaslıq, dərman götürüldükdən bir neçə həftə sonra, bir neçə ay sonra və ya bir il sonra müşahidə edilə bilən bir fenomendir (Robinson və Berridge, 1993). Mükafat / motivasiya dövrünün həssaslığının məcburi dərman axtarmağa töhfə verdiyini əvvəlcədən söylədilər (Robinson və Berridge, 1993). Müvafiq olaraq, lokomotor həssaslığı bərpaedilməsini istəyən özünü idarəetmə kokainini asanlaşdırır (De Vries və digərləri, 2002). Bundan əlavə, kokainin özünü idarə etməsinə geniş imkanları olan siçovullar, məhdud girişi olan siçovullara nisbətən kokainə daha çox lokomotor reaksiya göstərir (Ferrario və digərləri, 2005). Bundan əlavə, məcburi dərman axtaranlarda lokomotor həssaslığın psixostimulyatorlara əsaslanan neyrokimyəvi dəyişiklikləri də müşahidə olunur (Steketee və Kalivas, 2011; Giuliano et al., 2019). Bu məlumatlar, insanlarda müşahidə edilən lokomotor həssaslıq və kompulsif dərman axtarma arasındakı erkən təklif olunan yazışmanı dəstəkləyir (Robinson və Berridge, 1993; Vanderschuren və Kalivas, 2000). Sui-istifadə dərmanlarının mexaniki olaraq təkrar qəbul edilməsi, dopaminergik nörotransmissiyanı artıran mesolimbik dopamin dövranlarını həssaslaşdırır. Plazma membran dopamin nəqlini (DAT) əngəlləyən kokain və ya amfetaminlər kimi psixostimulyatorlar, striatumdakı və NAcdakı sinaptik məkanda dopamin miqdarının artmasına səbəb olur və beləliklə lokomotivi aktivləşdirir (Steketee və Kalivas, 2011). Narkomaniyada olduğu kimi, dopamin mükafatlandırma / motivasiya dövrünün həssaslığı OKB-də görülən məcburi davranışlara kömək edir. Həqiqətən, dopamin D2 reseptorlarının (D2Rs) təkrar aktivləşdirilməsi həm siçovullarda, həm də siçanlarda lokomotor həssaslığı və davranışı yoxlamaq üçün kifayətdir (Szechtman et al., 1998; Szechtman et al., 1999; Sun və digərləri, 2019). DinR / D2R agonisti olan quinpirolun təkrar administrasiyası, serotoninin bərpası ilə xroniki müalicədən sonra məcburi davranışların azalması ilə görülən kompulsif yoxlama və stereotipli davranış, proqnozlaşdırılan etibarlılıq artması ilə üzün etibarlılığını təkrarladığı üçün OKB-nin qəbul edilmiş bir modelidir. inhibitorları (SRI) və etibarlılıq qurur, çünki bu modeldə iştirak edən beyin strukturları patoloji olanlarla bölüşülür (Stuchlik et al., 2016; Szechtman et al., 2017). Xülasə, ya pre-sinaptik (dopamin sərbəst buraxılması) və ya post-sinaptik (D2R-in aktivləşdirilməsi) mexanizmlər tərəfindən dopamin ötürülməsinin təkrar aktivləşdirilməsi lokomotor həssaslığa və məcburi davranışlara səbəb olur.
Kappa opioid sistemi mükafat / motivasiya dövrəsində dopamin ötürülməsini tənzimləyən ən üstün sistemlərdən biridir. Sübutlar göstərir ki, kappa-opioid ötürülməsi dopamin təsirinə qarşı çıxır; kappa opioid reseptorlarının (KORs) kəskin aktivləşməsi psixostimulyatorların hərəkətə gətirdiyi lokomotor fəaliyyətə təsir göstərir. (Grey və digərləri, 1999). Əksinə təkrarlanan KOR aktivləşdirmə dərmanı məcburi və adət axtarışını saxlayır və artırır (Koob, 2013). Sui-istifadə dərmanlarının istehlakı, ehtimal ki, disforiyanın mənfi emosional vəziyyətinə töhfə verən bir homeostatik inkişaf etmiş kappa opioid ötürülməsinə səbəb olur (Koob, 2013) məcburi narkotik istifadəsinə səbəb olan (Chavkin və Koob, 2016). Əslində, KORun blokadası, nikotinin dərmanla əlaqəli deyil, stresin qarşısını aldı (Jackson və digərləri, 2013), kokain (Beardsley et al., 2005) və etanol (Sperling və başqaları, 2010). Bu tapıntıya uyğun olaraq, KOR blokadası, amfetamin həssaslığı olan siçovulların dorsolateral striatumundakı dərmanlara qarşı inkişaf etmiş lokomotor cavabını dəyişdirmədən dopaminergik dəyişiklikləri geri qaytarır (Azocar və digərləri, 2019). Beləliklə, KOR sistemi dərman dəyəri artan mənfi möhkəmləndirməni artırır. OKB-də mənfi möhkəmləndirmə, bu vəsvəsə etməmək üçün müəyyən bir məcburiyyəti gücləndirən vəsvəsələrə səbəb olur. Birbaşa sınaqdan keçməməsinə baxmayaraq, mənfi möhkəmləndirici quinpirole həssaslığına təsir göstərə bilər. Həqiqətən, D2R mənfi möhkəmləndirmənin yaranmasında iştirak edir. Məsələn, D2R uzun izoformu olmayan siçanlarda morfin-çəkilmə cütləşmiş əraziyə yol verilməməsi inkişaf etdirilməmişdir (Smith və digərləri, 2002) və abstinasiya dövründə təkrarlanan quinpirol müalicəsi, kokain və eroin, avtomatik idarəetmə paradiqmasında, quinpirole həssas lokomotivlə əlaqəli bir təsiri bərpa edir (De Vries və digərləri, 2002), psixostimulyator və quinpirolun təsirli həssaslıq arasında ortaq mexanizmləri təklif edir. Üstəlik, yeni bir qəfəs deyil, ev qəfəsinin açıq sahəyə gətirilməsi lokomotor həssaslığı və məcburi yoxlama davranışını azaldır (Szechtman et al., 2001), təhlükəsizlik / tanış işarələrin həssaslığa üstünlük verən mənfi ekoloji cəhətlərlə rəqabət edə biləcəyini göstərir. Psikostimulanta səbəb olan həssaslaşma kimi, KOR-un təkrar aktivləşdirilməsi lokomotor həssaslığı asanlaşdırır (Escobar və başqaları, 2017) və məcburi yoxlama davranışı (Perreault və digərləri, 2007) quinpirolun təkrar administrasiyası ilə əlaqələndirilir. Bu gücləndirici təsirin inkişaf etmiş mənfi möhkəmlənmənin nəticəsi olub olmadığını izah etmək qalır.
Bu yaxınlarda aparılmış hərtərəfli analiz kappa-opioid sisteminin dopaminergik ötürülməsinə təsirinin mürəkkəb olduğunu göstərir: cəlb olunan dopamin yolundan asılıdır (Margolis et al., 2006; Margolis et al., 2008) və KOR reseptorunun aktivləşdirilməsi ilə dopamin reseptorunun aktivləşdirilməsi arasındakı vaxt barədə (Chartoff et al., 2016). Bu mürəkkəbliyə uyğun olaraq, KOR ligandlarının potensial terapevtik istifadəsi geniş müzakirə edilmişdir. Dərman istifadəsi mərhələsində hiperdopaminergiya pozan KOR agonistinin dərman istifadəsi dövründə klinik baxımdan faydalı ola biləcəyi təklif edilmişdir (Shippenberg et al., 2007). Digər tərəfdən, KOR antaqonistinin təkrar narkotik istehlakından sonra dynorfin ifadəsinin artması ilə ortaya çıxan çəkilmə sindromunu müalicə etməkdə faydalı ola bilər (Wee və Koob, 2010). Buna görə, KOR qismən agonist olması təklif edildi (Béguin və digərləri, 2012) asılısı şəxslərdə həm kompulsif dərman qəbulunu, həm də çəkilmə simptomlarını müalicə etmək üçün terapevtik bir seçim ola bilər (Chartoff et al., 2016; Callaghan et al., 2018). Bu araşdırmada davranış həssaslığının və kompulsivliyin dopaminergik korrelyatlarının vaxt / kontekstdən asılı modulyasiyasını təhlil edirik.
Striatal və Midbrain Bölgələrində Kappa Opioid və Dopaminergik Sistemlər arasındakı anatomik və funksional krosstalk
Striatal bölgələr
KORlar orta beyin dopamin sistemində yüksək dərəcədə ifadə olunan Gi / o proteinlə əlaqəli reseptorlardır (Mansour və s., 1996). Bu reseptorlar mu (MOR), delta (DOR) və kappa (KORs) tərəfindən qurulmuş opioid reseptorları ailəsinə aiddir. Bu reseptorlar üçün endogen agonistlər müvafiq olaraq endorfinlər, enkefalin və dynorfindir. Striatumda, dynorphin, eyni nüvədən KOR-ları aktivləşdirən təkrarlayan aksonlara sahib olan orta ölçülü neyron (MSN) olan dopamin D1receptor (D1R) tərəfindən sintez olunur (Mansour və s., 1995). Siçovulların NAc elektron mikroskopiya görüntüləri KORların DAT tərkibli presinaptik quruluşlarda aşkar edildiyini göstərir, KORların az hissəsi isə DAT-a uyğunlaşma zamanı dendritlərdə lokallaşdırıldığını göstərir (Svingos və digərləri, 2001; Kivell və digərləri, 2014). NAc-dan presinaptik-sinaptosomal hazırlıqları xarakterizə edən immunofluoresan tədqiqatları göstərir ki, KOR və D2Rs, dopamin sintez edən fermenti, tirozin hidroksilazasını (TH) ehtiva edən sinaptozomlarda üstünlük təşkil edir (Escobar və başqaları, 2017). Bundan əlavə, KORs NAc və striatumun hüceyrə orqanlarında çoxdur və bir hüceyrə subpopulyasiyasında D2R ilə kolokalize olunur (Escobar və başqaları, 2017). Genetik və molekulyar anlayışlarla, striatumdakı KOR-un bağlanmasının 20% -nin DA terminallarında müşahidə olunduğu irəli sürüldü (Van't Veer et al., 2013). Bundan əlavə, Tejeda və başqaları. (2017) göstərdi ki, həm D1R, həm də D2R MSN-ləri MSN ehtiva edən D1R-lərə daha çox üstünlük verirlər (Tejeda və başqaları, 2017). Bu anatomik məlumatlar KOR-ların mükafat / motivasiya dövrəsində dopamin nörotransmissiyasını tənzimləyəndən əvvəl və postinaptik olaraq mövcud olduğunu göstərir.
Bir neçə təcrübi yanaşma göstərir ki, KORların aktivləşdirilməsi dopamin releasiyasını inhibə edire. Sistemli bir enjeksiyon və ya agonistlərin yerli infuziyası ilə KOR-ların kəskin aktivləşməsi NAc-da dopamin hüceyrədənkənar səviyyəsini azaldır (Di Chiara və Imperato, 1988; Spanagel və digərləri, 1992; Fuentealba və digərləri, 2006) və dorsal striatum (Gehrke və digərləri, 2008). KORların dopamin nörotransmissiyası üzərində tonik bir inhibitor təsirini dəstəkləyən, uzunmüddətli və seçici KOR antaqonistinin və qeyri-binaltorfiminin birbaşa infuziyası (nor-BNI) (Broadbear et al., 1994) NAcdakı dopamin səviyyəsini artırır (Spanagel və digərləri, 1992) və dorsal striatumda dopamin sərbəst buraxılması (Azocar və digərləri, 2019). Dopamin KOR tonik inhibisyonunun son dəlili, striatumda və NAcda dopamin həddindən artıq hüceyrə səviyyəsini göstərən KOR nokaut siçanlarında göstərildi (Chefer et al., 2005). Dopamin salınmasının KOR inhibisyonundan məsul olan mexanizmlər tamamilə aydınlaşdırılmamışdır. Bununla birlikdə, KOR-lərin aktivləşdirilməsinin K + artmasına və Ca2 + keçiriciliyinin azalmasına səbəb olduğu, hüceyrə hiperpolarizasiyasına və veziküler nörotransmitterin sərbəst buraxılmasına səbəb olduğu yaxşı bilinir (Bruchas və Chavkin, 2010-cu il; Margolis və Karhanis, 2019).
Bundan əlavə,, in vitro və vivo ilə funksional məlumatlar KORların DAT fəaliyyətini modulyasiya edərək dopamin hüceyrədənkənar səviyyələrini dəyişdirməsini təklif edir. Məsələn, EM4 hüceyrələrində KOR və DAT-ı ifadə edən hüceyrələrdə KOR-ların aktivləşməsi voltametriya ilə ölçülən dopamin artmasına səbəb olur (Kivell və digərləri, 2014). Bir ex vivo Ayrılan toxumalarda voltametriyadan istifadə edərək aparılan analiz, KOR agonist U-69593 sistemli bir enjeksiyonun NAc-da dopamin qəbulunu artırdığını göstərdi (Thompson və digərləri, 2000). Bənzər son bir məqalə göstərir ki, nor-BNI qarışıq Kappa / Delta opioid reseptor agonisti olan MP1104-in kəskin sistemli bir enjeksiyonu ilə törədilən ventral və dorsal striatumda dopamin qəbulunun artmasına mane olur (Atigari və başqaları, 2019). Buna baxmayaraq, KOR aktivləşdirməsinin dopamin qəbuluna təsiri hələ tam izah edilməmişdir. KOR qismən agonist nalmefenin sistemli rəhbərliyi, sürətli tarama tsiklik voltametriya (FSCV) ilə ölçülən, striatal dopamin qəbuledici dozasını asılı olaraq azaltdı (Gül et al., 2016). Yetkin kişi siçovullarında şəbəkəsiz axın mikdializindən istifadə edərək, KOR-un bloklanması dopamin alışının dolayı ölçüsü olan hasilat fraksiyasının (Ed) artması ilə müşayiət olundu (Chefer et al., 2006; Azocar və digərləri, 2019), KOR-un tonik aktivləşdirilməsinin DAT fəaliyyətinə (dopamin qəbulu) maneə törətdiyini göstərir. Bu nəticələr, dopamin hüceyrədənkənar səviyyələrini idarə etmək üçün dopamin qəbul etməsində endogen KOR fəaliyyətinin kompleks rolunu vurğulayır. FSCV kimi yüksək müvəqqəti qətnamə yanaşmaları KOR'un dopamin qəbuluna təsirini göstərə bilmədi (Ebner və başqaları, 2010; Ehrich və başqaları, 2015; Hoffman et al., 2016), striatal bölgələrdə DAT fəaliyyətini artıran bir inkubasiya dövrünə ehtiyac olduğunu irəli sürdü. DAT fəaliyyətinin KOR vasitəçiliyi ilə inkişaf etdirilməsi, striatal sinaptosomalarda və hüceyrədə bildirildiyi kimi KOR aktivləşməsi nəticəsində meydana gələn hüceyrə membranlarında DAT sayının artması ilə izah edilə bilər. xətlər (Kivell və digərləri, 2014).
Midbrain Regionlar
Siçovulların orta beyində aparılmış avtoreqrafik təhlillər, SN və VTA-nın rostrocaudal oxlarında KORlar üçün əhəmiyyətli bağlantı olduğunu göstərir (Speciale və digərləri, 1993). Digər tərəfdən, elektron mikroskopiya məlumatları göstərir ki, dynorfin tərkibli terminallar SN və VTA-da TH müsbət dendritlərə birbaşa sinaps edir (Sesack və Pickel, 1992), KORların dopamin neyronlarının somatodendritik hissələrində lokalizasiyasını təklif edir. Striatal D1R tərkibli MSN, midbrain dopamin neyronlarına daxil olan dynorfin girişlərindən biridir. Maraqlıdır ki, KORların tıxanması D1R-MSN-lərin VTA dopamin neyronlarına təsirini dəyişdirmir və bu inhibisyonun GABA vasitəçiliyini göstərir (Edwards və digərləri, 2017). KORlar dopamin orta beyin neyronlarının somatodendritik cavablarını modulyasiya edir. Elektrofizyolojik tədqiqatlar göstərir ki, VTA-da KORların aktivləşməsi dopamin neyronlarının spontan atəş dərəcəsini azaldır (Margolis et al., 2003). Nəticə etibarilə KOR agonistlərinin infuziyası somatodendritik dopamin effektini azaldır (Smith və digərləri, 1992; Dalman və O'Malley, 1999). Bununla birlikdə, KORların dopamin neyronlarına təsirini bu dövrə bağlı olduğu görünür. VTA-da kappa-opioid agonistlərinin infuziyası medial prefrontal korteksdə dopamin ifrazını azaldır (mPFC) (Margolis et al., 2006) lakin NAc-da deyil (Devine et al., 1993; Margolis və digərləri, 2006). Bundan əlavə, Margolis və başqaları. (2006) KOR-ların, NAc-a aid olan layihələri deyil, mPFC və bazolateral amigdala istiqamətləndirən VTA dopamin neyronlarını inhibə etdiyini tapdı. Elə həmin il, Ford və s. (2006) KOR agonistlərinin siçan VTA dilimlərində vanna tətbiq etməsi, NAc-a proyekt edən dopamin neyronlarında NAc-a nisbətən daha yüksək bir cərəyanı meydana gətirdiyini göstərdi, bu da KOR-ların NAc-a daha çox olan dopamin neyronlarını daha çox inhibə etdiyini göstərir. amiqdala. Bundan əlavə, KOR-un aktivləşməsi həyəcan amplitüdünü azaldır (Margolis et al., 2005) və inhibe (Ford və digərləri, 2007) midbrain dopamin neyronlarına postsinaptik cərəyanlar. VTA'nın mPFC və NAc'a olan kompleks efferents proyeksiyaları arasında növlər və fərqlər (Van Bockstaele və Pickel, 1995; Carr və Sesack, 2000) KOR-ların VTA neyron dopamin populyasiyasının bir hissəsini seçici şəkildə maneə törətməyini çətinləşdirir. Buna baxmayaraq, burada ümumiləşdirilmiş məlumatlar göstərir ki, KORs dofamin neyronlarının soma və terminallarında, həmçinin onları tənzimləyən girişlərdə olur və beləliklə midbrain dopamin neyronlarının sinaptik fəaliyyətini idarə etmək üçün əla mövqedədir.
Psixostimulyatorlarla induksiya olunan həssaslaşma və kompulsiv davranışlarda dopamin nörotransmissiyasına nəzarət edən KOR-ların rolu
Narkomanlıq, əvvəlcə müsbət gücləndirici təsiri ilə əlaqəli impulsiv dərman axtaran bir prosesdir. Digər tərəfdən, kompulsivlik narkomanlarda müşahidə olunan bir şəxsiyyət əlamətidir. Dopaminergik yollarda bir neçə nöroadaptasiya, məcburi narkotik axtarma və sui-istifadə dərmanlarına təkrar məruz qaldıqdan sonra qəbul edilməsi təklif edilmişdir (Everitt və Robbins, 2005; Koob və Volkow, 2016). Kompulsif dərman qəbulunu təklif edən fərziyyələrdən biri də onun mənfi gücləndirici təsirinin həssaslaşmasıdır (Koob, 2013). Kopa opioid sisteminin dopamin sərbəst buraxılmasına inhibitor nəzarəti sui-istifadə dərmanlarının mənfi gücləndirici xüsusiyyətlərinə kömək edə bilər. Bununla birlikdə, KOR aktivləşdirməsinin dopamin nörotransmesi və məcburi dərman axtaran dərmanlar üzərində təsiri mürəkkəb və zahirən zidd görünür. Həqiqətən, amfetamin və kokainin təsir etdiyi dopamin sərbəst buraxılması KOR agonistlərinin müşayiətedici administrasiyası tərəfindən pozulur (Heidbreder və Shippenberg, 1994-cü il; Maisonneuve və digərləri, 1994; Thompson və digərləri, 2000) və hətta kokainin özünü idarə etməsini azaldır (Negus və digərləri, 1997). Bundan əlavə, KORs, psixostimulyatorlara təkrar məruz qalması ilə meydana gələn post-sinaptik D1R-in davamlı aktivləşməsinə cavab olaraq mesolimbik yolun dopamin buraxması ilə əlaqədar bir inhibitor rəy verirlər (Cole və ark., 1995; Nestler, 2001). Paradoksal olaraq, KORların aktivləşdirilməsi mükafat / motivasiya yolunda dopamin buraxılmasını da asanlaşdıra bilər (Fuentealba və digərləri, 2006; Fuentealba və digərləri, 2007) və psixostimulyatorların istehlakı (Wee et al., 2009). Fuentealba və başqaları. (2007) U69593, KOR agonisti tətbiq edildiyi dörd gündən sonra, NAc-də amfetamin təsirli dofamin ifrazının artdığını göstərdi. Bu yaxınlarda, KOR-ların bloklanması amfetamin səbəb olduğu lokomotor həssaslığı zamanı baş verən dorsamin striatumundakı dopamin ifrazatındakı dəyişiklikləri geri aldığını göstərdi (Azocar və digərləri, 2019). Ümumilikdə, bu məlumatlar KOR-ların aktivləşdirilməsinin narkotik maddələrin müsbət gücləndirici xüsusiyyətlərinə də kömək edə biləcəyini göstərir (Chartoff et al., 2016).
Bundan əlavə, KOR-ların aktivləşdirilməsi də məcburi narkotik axtarışına kömək edir; KOR blokadası kokaini azaldır (Wee et al., 2009), heroin (Schlosburg və başqaları, 2013) və metamfetamin (Whitfield vd., 2015) Dərmana məhdudiyyətsiz çıxışı olan siçovulların qəbulu (Wee və digərləri, 2009). Bu təsir stresə səbəb olan dərman axtaranlarda da sübut olunur. Məsələn, KOR nokaut siçanları məcburi üzgüçülük stressindən sonra kokainə üstünlük vermədilər (McLaughlin və digərləri, 2006a). KORların bloklanması, məcburi üzmə stresinə məruz qalma səbəb olan nikotin yerini üstün tutur (Smith və digərləri, 2012). Maraqlısı budur ki, KOR-un bloklanması kokaini və nikotini stresdən yayındırmaq istəyini azaldır, lakin bir dərman problemi ilə əlaqəli axtarmağa təsir etmədi (Beardsley et al., 2005; Jackson və digərləri, 2013). Stressdən qaynaqlanan bu köməkçi KOR effekti mükafat / motivasiya sxemi ilə vasitəçi olduğu görünür (Shippenberg et al., 2007; Wee və Koob, 2010). Doktor Kauer və qrupunun apardığı zərif bir araşdırmada, VTA-da KORların bloklanması, ya əvvəllər ya da kəskin bir stresdən sonra, kokain axtaran şəxslərin bərpasına mane olduğunu, uzunmüddətli- dopamin neyronlarında inhibitor sinapsların potensiallaşdırılması (Graziane və digərləri, 2013; Polter və digərləri, 2014).
KORs tərəfindən tətbiq olunan psixostimulyatorların qəbulunun asanlaşdırılması, dərmanlara məruz qalma ilə bağlı zaman pəncərəsindən asılı görünür. KOR agonistinin U50488 rəhbərliyindən 1 saat əvvəl kokain ifraz etməsi həm kokain yerini üstün tutur, həm də NAc-da kokain tərəfindən atılan nisbi dopamin ifrazını 15 dəqiqə əvvəl verildiyi zaman əks təsir göstərir (McLaughlin və digərləri, 2006a; Ehrich və başqaları, 2014). İntrakranial özünü stimullaşdırmaqdan istifadə Chartoff və s. (2016) KOR agonisti Salvinorin A'nın NAc-da azalması və stimullaşdırılmış dopamin salınmasının artması ilə müşayiət olunan ilkin aversiv və gecikmiş mükafatlandırıcı təsir göstərdiyini müşahidə etdi. Bütün bu məlumatlar KOR aktivləşdirməsinin kokainin mükafatlandırıcı xüsusiyyətlərinə zamanla bağlı təsir göstərdiyini göstərir və məcburi dərman axtaran insanın inkişafı üçün əsas rol oynayan stress vasitəçisi KOR aktivləşdirilməsini göstərir.
Quinpirole ilə əlaqəli lokomotor həssaslıq və kompulsiv davranış
Dopamin sisteminin həssaslaşma və kompulsivlik meydana gəlməsində iştirak etdiyi faktlar, D2R agonisti, quinpirole ilə müalicə olunan kemiricilərdə müşahidə olunan davranışla güclənir. Qısaca, D2Rs mükafat / motivasiya dövrəsində geniş ifadə olunan Gi qoşulmuş reseptorlardır; onlar somatodendritik olaraq və dopamin neyronlarının akson terminallarında ifadə edilir (Sesack və digərləri, 1994) və onun aktivləşdirilməsi dopamin hüceyrədənkənar səviyyəsini azaldır (Imperato və Di Xiara, 1988). Striatumda D2Rs də orta spiny neyronların üzərində postinaptik olaraq yerləşirlər (Sesack və digərləri, 1994) və onun aktivləşdirilməsi lokomotor fəaliyyətə imkan verən dolayı yolu maneə törədir.
Dr. Henry Szechtman, kinpirolun 1980-ci ilin onilliyində bitən siçovulların davranışına təsirlərini öyrənməyə başladı. İlkin tapıntılar quinpirolun kəskin tətbiqinin lokomotor fəaliyyətə doza bağlı təsir göstərdiyini göstərdi. Aşağı dozalarda (0.03 mq / kq) lokomotor aktivlik azalır, daha yüksək dozalarda (> 0.5 mq / kq) artar. (Eilam və Szechtman, 1989). Bu effektlər müvafiq olaraq yüksək yaxınlıqdakı presinaptik D2R və aşağı yaxınlıq postsinaptik D2R-lərin aktivləşməsi ilə əlaqələndirilir (Usiello et al., 2000). Gözlənilmədən, quinpirolun təkrarlanan (hər gün) təkrar qəbul edilməsi, psixostimulyatorların cəlb etdiyi lokomotor həssaslığa bənzəyən lokomotivin tədricən və davamlı artmasına səbəb olur (Szechtman et al., 1993; Szechtman et al., 1994). Lokomotor həssaslaşdırıcı effektin D2R-lə bağlı olduğu göstərildi, çünki bu reseptor üçün çatışmazlığı olan siçanlar quinpirole lokomotor həssaslığını inkişaf etdirmir (Escobar və başqaları, 2015).
90-cı illərin əvvəllərində Szechtman və Eilam, lokomotor həssaslaşma ilə yanaşı, siçovulların quinpirolun hər tətbiqi ilə gücləndirilən stereotip bir davranış inkişaf etdirdiyini bildirdilər (Eilam və Szechtman, 1989; Szechtman et al., 1993). Bu gün quinpirole təkrar administrasiyası OKB üçün təsdiq edilmiş bir modeldir (Szechtman et al., 1999; Szechtman et al., 2001; Eilam və Szechtman, 2005; Stuchlik et al., 2016; Szechtman et al., 2017), siçovulların davranışının getdikcə daha da quruluşlu və çevik hala gəldiyini müşahidə edərək, məcburi yoxlama davranışının xarakterik ritual davranışını xatırladır (Szechtman et al., 1998; Szechtman et al., 2017). Son tədqiqatlar göstərir ki, təkrarlanan quinpirole siçanlarda məcburi yoxlama kimi məcburi davranışlara da yol açır (Sun və digərləri, 2019), davranış dəyişkənliyi və məcburi çeynəmə (Asaoka və digərləri, 2019), ikincisi striatumdakı D2Rs blokadası ilə geri qaytarıldı və kompulsiv davranışları inkişaf etdirmək üçün təkrar D2R-lərin aktivləşdirilməsinin lazım olduğunu dəstəklədi. Birlikdə məlumatlar lokomotor həssaslığı və kompulsivliyi stimullaşdırmaq üçün orta beyin dopamin yolları içərisində D2R-lərin mühüm rolunu göstərir. Təkrar quinpirole administrasiyası kokain səbəb olan stereotipli davranışa meyl göstərir (Thompson və digərləri, 2010) və amfetaminin lokomotor təsirləri (Cope et al., 2010), D2R-lərin aktivləşdirilməsinin psixostimulyasiya edən sensibilizasiyanın əsaslandığı və quinpirole ilə psixostimulyantların təsirli həssaslaşma arasında ortaq bir mexanizm təklif etdiyi fikrini gücləndirir. Maraqlıdır ki, təkrar D2R-lərin aktivləşdirilməsinin həssaslaşdırıcı təsiri psixostimulyatorların təsirindən daha güclü görünür, çünki quinpirole ilə müalicə olunan hər bir siçovan lokomotor həssaslığı inkişaf etdirir (Escobar və başqaları, 2015), siçovulların altmış faizi amfetamin həssas olduqda (Escobar və başqaları, 2012; Casanova və başqaları, 2013).
D2R-lərin təkrar aktivləşdirilməsi ilə nəticələnən davranış həssaslığı mükafat / motivasiya dövrəsində uyğunlaşmalarla müşayiət olunur. Quinpirole ilə həssaslaşan siçovulların NAc-da aşağı dopaminergik tonu var, azalmış bazal olaraq müşahidə olunur (Koeltzow və başqaları, 2003) və stimullaşdırılmış tonik və phasic dopamin azadEscobar və başqaları, 2015), dopamin orta beyin dövranının dopamin buraxma qabiliyyətinin azaldığını göstərir. NAc-dakı sinaptik dopamin səviyyələri, həm DAT, həm də dopamin neyronlarının fəaliyyəti ilə idarə olunur (Goto və Grace, 2008), hansı vivo ilə tonik və partlayış atəşindən ibarətdir (Wilson və digərləri, 1977; Grace və Bunney, 1980). Əvvəlki hesabatlar quinpirole həssas olan siçovulların daha az sayda dopamin neyronını tonik və VTA-da atəşə tutduğunu göstərir (Sesia və digərləri, 2013). Birlikdə bu məlumatlar, quinpirole həssaslığından sonra görülən dopamin ifrazının azalmasının dopamin neyronlarının ümumi fəaliyyətinin azalmasının nəticəsi olduğunu göstərir. Quinpirole ilə təkrar müalicə nəticəsində yaranan kompulsif davranış və həssas lokomotor fəaliyyət, NAc-də dopaminergik tonun azalması səbəbindən D2Rlərin həssaslaşmasının nəticəsi ola bilər. Həqiqətən, quinpirole həssas olan siçovullar dopamin D2R-in bağlanmasında artım göstərir (Culver və digərləri, 2008) və bu reseptorların yaxınlıq vəziyyətinin artması (Perreault və digərləri, 2007), bu fərziyyəni dəstəkləyir.
Quinpirole ilə əlaqəli kompulsif davranışlarda KOR-Dopamin qarşılıqlı təsiri
DOR ilə əlaqəli kompulsif davranışlarda KOR-un rolu ilə bağlı ilk tədqiqatlar Szechtman laboratoriyasından da gəldi. Bu qrup KOR agonisti U2-ün lokomotor aktivliyə xinpirol ilə eyni vaxtda verilməsini araşdırdı. Konkret olaraq, müəlliflər, U69593 qarışığı və quinpirol ilə siçovullara subkutan enjeksiyonlar, 69593 ilə 8 iynə tamamlanana qədər tətbiq etdilər. Yalnız U10’ün hipolokomotor təsirindən fərqli olaraq, kinpirolun aşağı (presinaptik) və yüksək (postsinaptik) dozaları ilə eyni vaxtda tətbiq edildikdə hiperlokomotion müşahidə edildi. U69593, xinpirolun presinaptik dozasının hipolokomotor təsirini hiperlokomiyasiyaya dəyişdirdi və kinpirolun postsinaptik dozasının hiperlokomotor təsirini artırdı (Perreault və digərləri, 2006). KOR-ların birgə aktivləşdirilməsi də lokomotor həssaslığın induksiyasını sürətləndirdi və D2R-lərin aktivləşməsinin təsirini gücləndirdi, çünki ikiqat müalicənin əldə etdiyi maksimal lokomotiv təkcə quinpirole tərəfindən yaranan lokomotor təsirini artırır (Perreault və digərləri, 2006; Escobar və başqaları, 2017). KOR-ların birgə aktivləşdirilməsi də məcburi yoxlama davranışının əldə edilməsini sürətləndirir (Perreault və digərləri, 2007). KOR-ların quinpirole ilə əlaqəli davranışlara təsir göstərən bu təsirləri KORların təkrar aktivləşdirilməsini tələb edir. Əslində, KOR agonist U69593-nin kəskin inyeksiyası quinpirole ilə həssas olan siçovullarda lokomotor fəaliyyətini daha da dəyişdirmədi (Escobar və başqaları, 2017). D2R ilə əlaqəli həssaslaşma təmin edən KOR mexanizmi məlum deyil. Bir ehtimal, endogen kappa opioid sisteminin özü D2R-dən asılı olan həssaslığın vasitəçiliyi olmasıdır. Bununla birlikdə, bu ehtimal norBNI-nin əvvəlcədən idarə edilməsinin lokomotor həssaslığını quinpirole dəyişdirmədiyini göstərərək dynorfinin D2R aktivləşdirilməsinin aşağı axınına səbəb olmadığını göstərir (Escobar və başqaları, 2017). Bu məlumatlar dynorfinin kompulsif davranışları həssaslaşdırmaqda bir rol oynaya biləcəyini istisna etmir, məsələn, stres dynorfinin sərbəst buraxılmasına və kompulsif davranışları asanlaşdıran KORların aktivləşməsinə səbəb olur (McLaughlin və digərləri, 2003; McLaughlin və digərləri, 2006a; McLaughlin və digərləri, 2006b).
D2R və KOR arasındakı crosstalk mürəkkəbdir və hər iki reseptorun aktivləşməsinin təsadüf və ya müvəqqəti olaraq ayrıldığından asılı görünür. Anatomik məlumatlar göstərir ki, D2Rs və KORs arasındakı qarşıdurma presinaptik olaraq axon və soma dopamin neyronlarında, habelə striatumun MSN-lərində postinaptik olaraq baş verə bilər. Digər neyrokimyəvi sistemlərin aksonlarında yerləşən KORlar üçün bir rolun olacağını istisna etməsə də, anatomik məlumatlar D2R-ləri tənzimləyən KORların birbaşa roluna güclü işarə edir. Kəskin və ya təkrarlanan, KORların aktivləşməsi dopamin neyronlarında inhibe D2Rs funksiyasını azaldır. Elektrofiziologiya tədqiqatları göstərdi ki, VTA və SN-nin dopamin neyronlarında KOR-un kəskin aktivləşməsi D2R ilə əlaqəli inhibitor postsinaptik cərəyanı inhibə edir, KOR dopamin buraxmasını azaldır və dynorfin vanna tətbiq olunan dopamin inhibe təsirini bloklayır, çünki pre və postinaptik mexanizmlərin vasitəçilik etdiyi bir təsir.Ford və digərləri, 2007). Neyrokimyəvi tədqiqatlar KORların təkrar aktivləşməsini NAc-da dopamin sərbəst buraxılmasının D2R-inhibə etdiyi inhibe etdiyini göstərdi (Fuentealba və digərləri, 2006). Üstəlik, təsadüf olunan D2R və KOR-ların kəskin aktivləşməsi təkcə hər bir reseptorun təsiri ilə müqayisədə NA-dopamin azad edilməsinin azalmasını azaldır (Escobar və başqaları, 2017). Beləliklə, presinaptik KOR-lar əlavə və ya presinaptik D2R-lərlə sinergiyada hərəkət etmir, əksinə KORlar ya D2R inhibe effektini ya da maneə törədirlər. Bu mexanizm, KOR agonistlərinin kəskin bir dozasının aşağı dozalı quinpirol ilə əlaqələndirici lokomotor aktivləşdirici təsirini izah edə bilər (Perreault və digərləri, 2006).
Son bir araşdırma göstərir ki, VTA-da KOR aktivləşməsi davranış inhibe və mərmər basdırması kimi ölçülən kompulsif davranışları vasitəçilik edir (İbrahim və digərləri, 2017), KORların aktivləşdirilməsinin həqiqətən kompulsivlik üçün bir tətik olduğu fikrini gücləndirir. Məlumat yayımladı Margolis et al. (2006); 2008) göstərir ki, KORs və D2Rs qarşılıqlı təsiri mPFC-ni hədəf alan dopamin neyronlarında baş verməlidir (Margolis et al., 2006; Margolis et al., 2008). Baxmayaraq, Ford və s. (2006); 2007) D2R vasitəçi IPSC'nin KORların inhibe edilməsinin, NAc'ı hədəf alan dopamin nöronlarında reallaşdığını tapdı (Ford və digərləri, 2006; Ford və digərləri, 2007). Birlikdə bu məlumatlar göstərir ki, Dopamin neyronlarının somatodendritik bölməsində DOR ilə D2R ilə qarşılıqlı təsir eyni dopamin neyronundakı crosstalk nəticəsində yarana bilər. Bunun mezolimbikdə və ya mezokortikal proqnozlarda olub olmaması hələ mübahisəlidir.
Xatırladaq ki, KOR, NAc'ın MSN'lərində tapıldı (Escobar və başqaları, 2017; Tejeda və başqaları, 2017), beləliklə D2R ilə əlaqəli kompulsif davranışın potensialının dopamin neyronlarının hədəf hüceyrələrinə birbaşa hərəkətləri ilə də yarana biləcəyini göstərir. Bununla əlaqədar olaraq qeyd etmək lazımdır ki, U69593-in təkrar administrasiyası yüksək yaxınlıq vəziyyətində D2R miqdarını artırır (Perreault və digərləri, 2007). Neyrokimyəvi məlumatlar göstərir ki, dopamin hüceyrədənkənar səviyyəsinin azalması D2Rs həssaslığı ilə əlaqələndirilir. KOR-ların aktivləşməsi, D2R-lərin təkrar aktivləşdirilməsi ilə azalmış NAc-dakı hüceyrədənkənar dopamin səviyyəsini daha da azaldır (Escobar və başqaları, 2017), bu mexanizm vasitəsilə NAcdakı D2R-lərin həssaslığını sürətləndirən və ya artıran presinaptik KORların rolunu istisna etmək. Buna görə, KORs, D2R-lərin neyrokimyəvi və davranış təsirlərini daha da həssaslaşdıran yavaş molekulyar mexanizmləri işə salır, lokomotor həssaslığın artırılmasının presinaptik effektdən daha çox uyğunlaşan postsinaptik ola biləcəyini göstərir. Bu baxımdan KORların təkrar aktivləşdirilməsi D2R / D1R tarazlığının D2R induksiya edən kompulsivliyə keçid D1R dolayı striatal yolunun inhibisyonuna səbəb ola bilər (Şəkil 1).
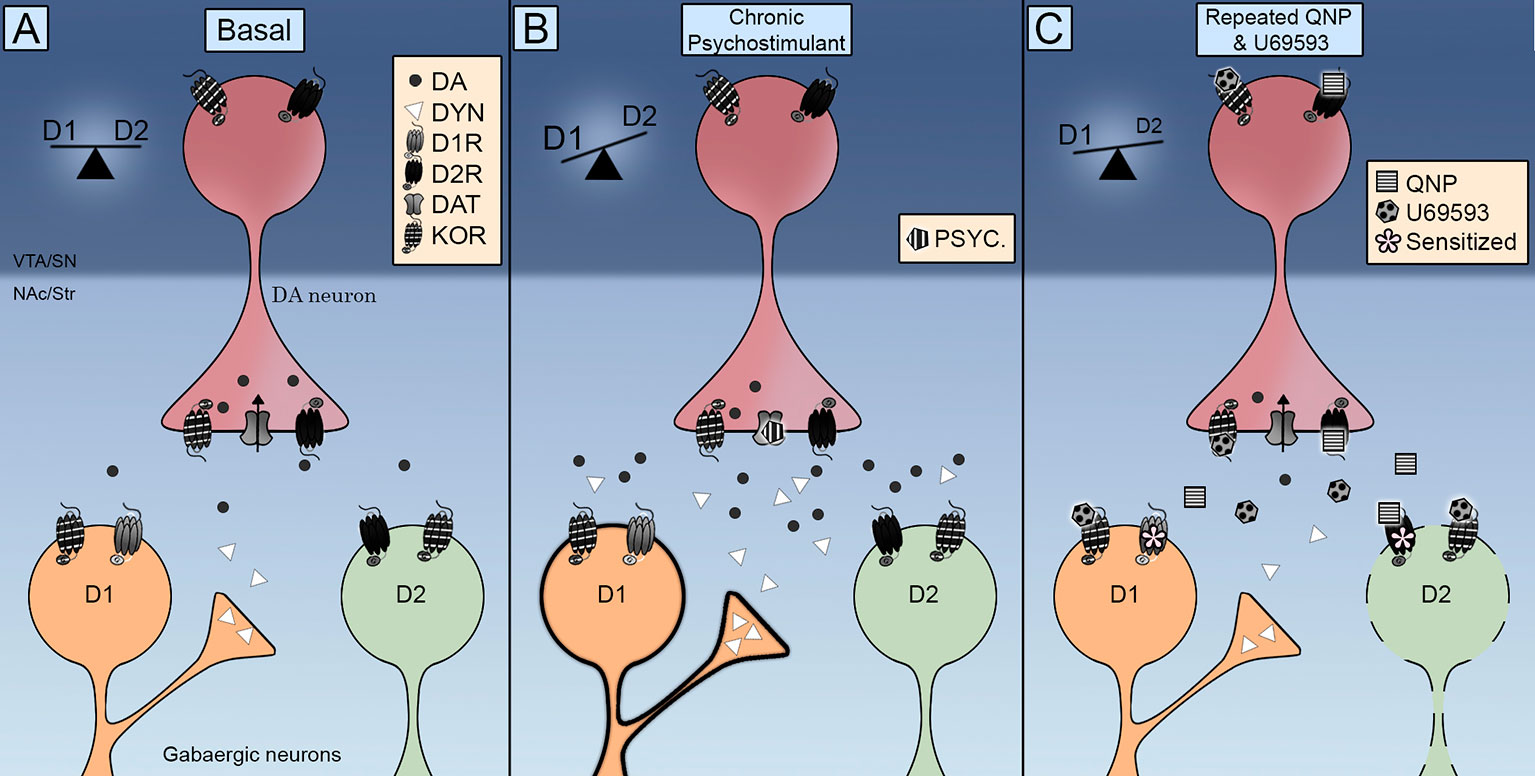 Şəkil 1 Birbaşa (D1R) və dolayı (D2R) striatal phatways üzərində Kappa Opioid Reseptorları (KOR) nəzarətinin inteqrativ sxemi. (A) KOR, əvvəlcədən sinaptik olaraq dopamin terminallarında yerləşdirilir və orta ölçülü neyronlarda (MSN) sinaptik şəkildə yerləşdirilir. Onun aktivləşdirilməsi dopamin hüceyrədənkənar səviyyələrini idarə edir və onun lokalizasiyası dopamin daşıyıcısı (DAT) və dopamin D2 reseptorları ilə qarşılıqlı əlaqəni inkişaf etdirir. (B) Psixostimulyatora təkrar məruz qalma həm dopamin hüceyrədənkənar səviyyəsində, həm də dynorfində artımla müşayiət olunur. D1 və D2 reseptorlarının aktivləşdirilməsi tarazlığı lokomotor həssaslığı təşviq edən D1R birbaşa yoluna keçir. (C) Quinpirole və U69593-in birgə administrasiyası dopamin hüceyrədənkənar səviyyəsində azalma ilə müşayiət olunur. KOR və D2 reseptorlarının eyni vaxtda aktivləşdirilməsi kompulsif davranışa səbəb olan D2 dolayı yolunu azaldır.
Şəkil 1 Birbaşa (D1R) və dolayı (D2R) striatal phatways üzərində Kappa Opioid Reseptorları (KOR) nəzarətinin inteqrativ sxemi. (A) KOR, əvvəlcədən sinaptik olaraq dopamin terminallarında yerləşdirilir və orta ölçülü neyronlarda (MSN) sinaptik şəkildə yerləşdirilir. Onun aktivləşdirilməsi dopamin hüceyrədənkənar səviyyələrini idarə edir və onun lokalizasiyası dopamin daşıyıcısı (DAT) və dopamin D2 reseptorları ilə qarşılıqlı əlaqəni inkişaf etdirir. (B) Psixostimulyatora təkrar məruz qalma həm dopamin hüceyrədənkənar səviyyəsində, həm də dynorfində artımla müşayiət olunur. D1 və D2 reseptorlarının aktivləşdirilməsi tarazlığı lokomotor həssaslığı təşviq edən D1R birbaşa yoluna keçir. (C) Quinpirole və U69593-in birgə administrasiyası dopamin hüceyrədənkənar səviyyəsində azalma ilə müşayiət olunur. KOR və D2 reseptorlarının eyni vaxtda aktivləşdirilməsi kompulsif davranışa səbəb olan D2 dolayı yolunu azaldır.
Kompulsif davranışlarda KOR-Dopamin qarşılıqlı təsirlərinin cins fərqləri
Klinik tədqiqatlar, kompulsif dərman axtaran da daxil olmaqla, məcburi davranışda cins fərqliliyini göstərdi. OKB simptomlarının daha erkən başlanğıcı qadınlarla müqayisədə kişilərdə müşahidə olunur (Mathis və digərləri, 2011), çirklənmə və təmizləyici simptomların daha çox yayıldığını göstərən qadınlarla (Labad və digərləri, 2008). Narkomaniya ilə cins fərqliliyinə gəldikdə, klinik dəlillər narkotik istifadəsinin kişilərdə daha çox olduğu bir vaxtda qadınların məcburi narkotik axtaran kişilərə nisbətən daha sürətli inkişaf etdiklərini göstərir (Hernandez-Avila və digərləri, 2004; Fattore və Melis, 2016).
Son zamanlarda klinikadan əvvəl dəlil klinik araşdırmalarda müşahidə edilən narkotikdən sui-istifadə hallarında cins fərqlərini əsas gətirən neyrobioloji əsasları vurğuladı (Becker və Chartoff, 2019). Təmiz olmayan axın mikdializindən istifadə edərək aparılan erkən müşahidələr göstərdi ki, dorsal striatumdakı dopamin hüceyrədənkənar konsentrasiyası, estrogen dövrü ərzində prostrus və estrusda daha yüksək səviyyədə olur. Bundan əlavə, ovariektomiya qadın siçovullarda striatal dopamin hüceyrədənkənar konsentrasiyasını azaldaraq, kişi siçovulların kastrasiyası dopamin striatal hüceyrədənkənar konsentrasiyasını dəyişdirmir (Xiao və Becker, 1994), yumurtalıq hormonlarının dopamin fəaliyyətində əhəmiyyətli rolunu təklif edir. Bundan əlavə, qadın hormonları psixostimulyatorların reaksiyasını tənzimləyir. Erkən vitro Təcrübələr estradiol plus progesteronun ovariektomiyalı qadın siçovullarından alınan striatal toxumadan amfetaminlə əlaqəli dofamin ifrazını bərpa etdiyini göstərdi (Becker və Ramirez, 1981). Bu yaxınlarda, sürətli tarama dövriyyəli voltametriya tədqiqatları, qadınların kişilərə nisbətən daha çox elektrikli stimullaşdırılmış dopamin sərbəst buraxma və alışma nümayiş etdirdiyini göstərdi (Walker və digərləri, 2000). Dopamin nörotransmisiyasındakı bu cinsi fərqlər qadınlarda müşahidə olunan daha yüksək kokain və amfetamin ehtiva edə bilər. (Roberts və digərləri, 1989; Cox et al., 2013).
KOR'un dopamin hüceyrədənkənar səviyyələrdə tənzimlənməsi də cins fərqliliyini göstərir (Chartoff və Mavrikaki, 2015). İntrakranial özünü stimullaşdırma və tsiklik voltametriyadan istifadə edərək, Conway və s. (2019) qadın siçovullarda müşahidə olunan KOR agonistinin kəskin anhedonik təsirə qarşı həssaslığının NAc-də stimullaşdırılmış dopamin sərbəst buraxılmasının dayandırılması ilə müşayiət olunduğunu göstərdi (Conway et al., 2019). Estradiolun KOR aktivləşdirilməsindən sonra qadın siçovullarda müşahidə olunan dopamin ifrazının kəskin inhibisiyasına töhfə verməsi təklif edilmişdir (İbrahim və digərləri, 2018). KOR və dopamin siqnalları arasındakı crosstalk, kişilərdə öyrənilmişdir (Tejeda və Bonci, 2019), bu qarşılıqlı təsir və qadınlarda bağımlılık prosesindəki təsiri mövzusunda tədqiqat yoxdurChartoff və Mavrikaki, 2015). Qadın siçovullarında, KOR agonist U69593-in kəskin idarəsi, həm nəzarət, həm də ovariektomiya edilmiş siçovullarda kokain səbəb olan hiperlokomozasiya. Maraqlıdır ki, U69593 təkrar administrasiyası estradioldan asılı şəkildə kokainə səbəb olan hiperlokomozu (Puig-Ramos və başqaları, 2008). Bu məlumatlar, qadın siçovullarındakı estradiol prormlarının KOR hərəkətlərini, stres reaksiyasında cins fərqliliyi ilə əlaqəli ola biləcəyini göstərir (Puig-Ramos və başqaları, 2008). Dişi siçovullarda KOR-ların təkrar aktivləşdirilməsi kişilərdə müşahidə olunduğu kimi striatal dopamin ifrazını asanlaşdırır.
Kişi siçovullarına nisbətən qadınlarda psixostimulyatorlu dofamin sərbəst buraxılmasının asanlaşdırılması müşahidə olunsa da, amfetamin lokomotor həssaslığının altındakı dopamin mexanizmlərindəki cins fərqləri tam izah edilməmişdir (Becker, 1999). Amfetaminə dəfələrlə məruz qalması həm ikisi, həm də yeniyetmədə daha çox lokomotor fəaliyyətə səbəb olur (Mathews və McCormick, 2007) və yetkin qadın siçovulları (Milesi-Hallé və digərləri, 2007), amfetaminə təkrar məruz qaldıqdan sonra daha güclü bir lokomotor həssaslığı göstərən qadın yetkin siçovullarla. D2 reseptorunun neonatal aktivləşdirilməsi amfetamin təsir edən davranış həssaslığını yalnız qadın siçovullarda artırdı (Brown və ark., 2011). Daha əvvəl də qeyd edildiyi kimi, kişi siçovullarında D2 agonistinə təkrar məruz qalma lokomotor həssaslığı və kompulsiv xarakterli davranışa səbəb olur (Dvorkin və başqaları, 2006). Bundan əlavə, KOR-un aktivləşdirilməsi, NAc-da DA sərbəst buraxılması zamanı D2 reseptorlarının inhibitor nəzarətini asanlaşdıraraq quinpirolun təkrar məruz qalması nəticəsində yaranan lokomotor həssaslığı artırır (Escobar və başqaları, 2017). KOR-un qadınlarda dopamin salınmasına təsir edən KOR-un inhibe təsirinə müşahidə olunan həssaslıq kimi cins fərqləri (Conway et al., 2019) KOR-un məcburi dərman axtarmağa fərqli töhfəsini verə bilər.
Nəticələr
KORs, motivasiya edilmiş davranışları inkişaf etdirmək üçün dofamin siqnalını necə modulyasiya edir və həssas bir kompulsif davranış nə ilə nəticələnir? Anatomik məlumatlar KOR-ların orta beyin dopamin neyronlarının sinaptik fəaliyyətini idarə etmək üçün əla mövqedə olduğunu göstərir. Funksional məlumatlar göstərir ki, KORlar DAT və D2R fəaliyyətini, həmçinin dopamin neyronlarının atəş sürətini idarə edir. KOR-ların kəskin aktivləşməsinin, narkotik maddələrin təsirindən yaranan dopamin ifrazının azaldığını göstərən ilkin sübutlar, KOR-un təkrar aktivləşdirilməsinin dopamin sərbəst buraxılmasına və məcburi narkotik axtarmağa kömək etdiyini göstərən məlumatlarla tamamlandı. Dopamin siqnal balansı striatal bölgələrdən birbaşa və dolayı çıxış yolları (Şəkil 1A). Həm D1R həm də D2R aktivləşdirən dopamin ifrazını artıran psixostimulyantlarla ya da xroniki stimullaşdırma (Şəkil 1B) və ya yalnız D2R aktivləşdirən quinpirole (Şəkil 1C) lokomotor həssaslaşma və məcburi davranışların pozulmuş D2R dolayı yolu ilə nəticələnməsi və bununla da tarazlığın D1R birbaşa yoluna keçməsi. KOR ötürülməsi striatal D1 neyronlarında dynorfinin artması ilə xroniki psixostimulyator qəbulu zamanı güclənir (Şəkil 1B). Genişləndirilmiş KOR ötürülməsi U69593 tətbiq etməklə OKB-nin farmakoloji modelində yayılmışdır. Bu KOR aktivləşdirmə D2 dolayı yolunu daha da zəiflədir (Şəkil 1C). KOR aktivləşdirməsinin DAT fəaliyyətindəki nəticələrini tam aydınlaşdırmaq, quinpirolun meydana gətirdiyi kompulsivliyindəki endogever KOR sisteminin rolunu anlamaq və KOR sisteminin kompulsif davranışlarda müşahidə olunan cinsiyyət fərqlərinə töhfəsini təyin etmək üçün gələcək tədqiqatlar aparılmalıdır.
Müəllif iştirakları
AE, MA və JF əlyazmanın konsepsiyasına kömək etdi. AE və JF əlyazmanın ilk layihəsini MA-nın girişi ilə yazdılar. MA və JC əlyazmanın tənqidi nəzərdən keçirilməsinə və redaktəsinə kömək etdi. Bütün müəlliflər dərc üçün təsdiq etdilər.
Maliyyələşdirmə
Bu rəydə göstərilən müəlliflərin işi FONDECYT qrant nömrələri ilə dəstəklənmişdir: MA-ya 1110352 və 1150200; 1141088-dən JF-ə; JF-yə 391340281 DIPOG qrantı; FONDECYT postdoctoral həmkarı 3170497 JC-yə və 3190843-ü AE-yə.
Maraqların münaqişəsi
Müəlliflər bildirirlər ki, tədqiqat potensial münaqişələr kimi başa düşülə bilən hər hansı bir kommersiya və ya maliyyə əlaqəsi olmadıqda həyata keçirilir.
İşləmə redaktoru hazırda müəlliflərdən biri JF ilə bir araşdırma mövzusu təşkil edir və başqa bir əməkdaşlığın olmamasını təsdiqləyir.
References
Abraham, AD, Fontaine, HM, Song, AJ, Andrews, MM, Baird, MA, Kieffer, BL və s. (2017). Dopamin neyronlarında Kappa opioid reseptorunun aktivləşdirilməsi davranış inhibisyonunu pozur. Neuropsychopharmacology 43 (2), 362–372. doi: 10.1038 / npp.2017.133
Abraham, AD, Schattauer, SS, Reichard, KL, Cohen, JH, Fontaine, HM, Song, AJ, və s. (2018). GRK2-nin estrogen tənzimlənməsi vasitəçi analjeziyanı siqnal edən kappa opioid reseptorunu təsirsiz hala gətirir, lakin əksinə deyil. J. Neurosci. 38 (37), 8031–8043. doi: 10.1523 / JNEUROSCI.0653-18.2018
Asaoka, N., Nishitani, N., Kinoshita, H., Nagai, Y., Hatakama, H., Nagayasu, K., et al. (2019). Bir adenozin A2A reseptor antaqonisti təkrar quinpirol ilə əlaqəli psixozun çoxsaylı simptomlarını yaxşılaşdırır. eNeuro 6 (1), 1-16. ENEURO.0366-18.2019. doi: 10.1523 / ENEURO.0366-18.2019
Atigari, DV, Uprety, R., Pasternak, GW, Majumdar, S., Kivell, BM (2019). MP1104, qarışıq kappa-delta opioid reseptor agonist, siçovullarda azalmış yan təsirləri olan anti-kokain əleyhinə xüsusiyyətlərə malikdir. Neurofarmakoloji 150, 217-228. doi: 10.1016 / j.neuropharm.2019.02.010
Azocar, VH, Sepúlveda, G., Ruiz, C., Aguilera, C., Andres, ME, Fuentealba, JA (2019). Kappa-opioid reseptorunun bloklanması amfetamin həssaslığı zamanı dorsolateral striatum dopamin dinamikasında dəyişiklikləri bərpa edir. J. Neurochem. 148, 348–358. doi: 10.1111 / jnc.14612
Béguin, C., Potuzak, J., Xu, W., Liu-Chen, LY, Streicher, JM, Groer, CE və s. (2012). 12-epi-salvinorin A və onun analoqlarının kappa opioid reseptorunda diferensial siqnal xüsusiyyətləri. Bioorg. Med. Kimya. Lett. 15; 22 (2), 1023–1026. doi: 10.1016 / j.bmcl 2011.11.128
Beardsley, PM, Howard, JL, Shelton, KL, Carroll, FI (2005). Yeni kappa opioid reseptor antaqonisti JDTic-in, kokain axtaran insanları kokain praksiyaları və siçovulların antidepresan təsirinə qarşı təsir edən kokainin bərpasına fərqli təsirləri. Psixofarmakol. (Berl) 183, 118–126. doi: 10.1007/s00213-005-0167-4
Becker, JB, Chartoff, E. (2019). Mükafat və asılılıq vasitəçilik edən sinir mexanizmlərindəki cins fərqləri Neuropsychopharmacology 44, 166–183. doi: 10.1038/s41386-018-0125-6
Becker, JB, Ramirez, VD (1981). Amfetamin cinsi fərqləri in vitro-da katekolaminlərin siçovul striatal toxumasından sərbəst buraxılmasını stimullaşdırdı. Brain Res. 204, 361–372. doi: 10.1016/0006-8993(81)90595-3
Becker, JB (1999). Striatum və nüvə böyüdülməsində dopaminergik funksiyada cins fərqləri. Pharmacol. Biochem. Behav. 64, 803–812. doi: 10.1016/S0091-3057(99)00168-9
Berridge, KC, Robinson, TE (2016). Bəyənmə, istəmək və asılılığın stimullaşdırıcı-həssaslaşdırma nəzəriyyəsi. Am. Psychol. 71, 670–679. doi: 10.1037 / amp0000059
Berridge, KC, Venier, IL, Robinson, TE (1989). 6-hidroksidopamin ilə əlaqəli afaqiyanın dad reaktivliyi təhlili: dopamin funksiyasının oyanış və anhedoniya fərziyyələrinə təsiri. Behav. Neurosci. 103, 36-45. doi: 10.1037 / 0735-7044.103.1.36
Broadbear, JH, Negus, SS, Butelman, ER, de Costa, BR, Woods, JH (1994). Siçan dartma analizində kappa-opioid agonistlərinə sistemli idarə olunan nor- binaltorfiminin (nor-BNI) fərqli təsirləri. Psychopharmacology 115, 311-319. doi: 10.1007 / BF02245071
Brown, RW, Perna, MK, Noel, DM, Whittemore, JD, Lehmann, J., Smith, ML (2011). Neonatally quinpirole ilə müalicə olunan yeniyetmə kişi və qadın siçovullarda amfetamin lokomotor həssaslığı və şərtli yerə üstünlük. Behav. Pharmacol. 22, 374–378. doi: 10.1097/FBP.0b013e328348737b
Bruchas, MR, Chavkin, C. (2010). Kinaz kaskadları və kappa opioid reseptorunda ligand yönümlü siqnal. Psixofarmakol. (Berl) 210, 137–147. doi: 10.1007/s00213-010-1806-y
Callaghan, CK, Rouine, J., O'Mara, SM (2018). Motivasiya və böyük depresif xəstəliklərdə opioid reseptorları üçün potensial rollar. Prog. Brain Res. 239, 89–119. doi: 10.1016 / bs.pbr.2018.07.009
Carr, DB, Sesack, SR (2000). Prefrontal korteksə siçovul ventral tegramal bölgə layihəsində GABA ehtiva edən neyronlar. Synapse 38 (2), 114–123. doi: 10.1002/1098-2396(200011)38:2<114::AID-SYN2>3.0.CO;2-R
Casanova, JP, Velis, GP, Fuentealba, JA (2013). Amfetamin lokomotor həssaslığı siçovul medial prefrontal korteksdə inkişaf etmiş yüksək K + -stimulyasiya olunmuş Dopamin sərbəst buraxılması ilə müşayiət olunur. Behav. Brain Res. 237, 313-317. doi: 10.1016 / j.bbr.2012.09.052
Chartoff, EH, Mavrikaki, M. (2015). Kappa opioid reseptor funksiyasındakı cins fərqləri və asılılığa potensial təsiri. Cəbhə. Neurosci. 9, 466. doi: 10.3389 / fnins.2015.00466
Chartoff, EH, Ebner, SR, Sərçə, A., Potter, D., Baker, PM, Ragozzino, ME, et al. (2016). Kappa opioid reseptorunun aktivləşdirilməsi və kokain arasındakı nisbi vaxt, mükafat və dopamin sərbəst buraxılmasına təsirini müəyyənləşdirir. Neuropsychopharmacology 41, 989-1002. doi: 10.1038 / npp.2015.226
Chavkin, C., Koob, GF (2016). Dynorfin, disforiya və asılılıq: asılılığın stresi. Neuropsychopharmacology 41, 373-374. doi: 10.1038 / npp.2015.258
Chefer, VI, Czyzyk, T., Bolan, EA, Moron, J., Pintar, JE, Shippenberg, T. S. (2005). Endogen kappa-opioid reseptor sistemləri mezoakkumbal dopamin dinamikasını və kokainə qarşı həssaslığını tənzimləyir. J. Neurosci. 25, 5029-5037. doi: 10.1523 / JNEUROSCI.0854-05.2005
Chefer, VI, Zapata, A., Shippenberg, TS, Bungay, PM (2006). Kəmiyyətsiz təmiz flux mikdializ, siçan nüvəsinin böyüdülməsində dopamin alma səviyyəsində artım və azalmanın aşkarlanmasına imkan verir. J. Neurosci. Metodlar 155, 187-193. doi: 10.1016 / j.jneumeth.2005.12.018
Cole, RL, Konradi, C., Duglass, J., Hyman, SE (1995). Amfetamin və dopaminə neyron uyğunlaşma: siçovul striatumundakı prodinorfin geninin tənzimlənməsinin molekulyar mexanizmləri. Neyron 14, 813–823. doi: 10.1016/0896-6273(95)90225-2
Conway, SM, Puttick, D., Russell, S., Potter, D., Roitman, MF, Chartoff, E. H. (2019). Qadınlar kappa opioid reseptorlarının aktivasiyasının motivasiya və dopamin basdırıcı təsirlərinə nisbətən kişilərə nisbətən daha az həssasdırlar. Neurofarmakoloji 146, 231-241. doi: 10.1016 / j.neuropharm.2018.12.002
Cope, ZA, Huggins, KN, Sheppard, AB, Noel, DM, Roane, DS, Brown, RW (2010). Neonatal quinpirole müalicəsi yetkin yaşlarda amfetamin müalicəsinə cavab olaraq lokomotor aktivləşdirməni və nüvədəki dopamin ifrazını artırır. Synapse 64, 289-300. doi: 10.1002 / syn.20729
Cox, BM, Gənc, AB, Bax, RE, Reichel, CM (2013). Siçovullarda axtaran metamfetamin cinsi fərqləri: oksitosinin təsiri. Psikoneuroendokrinologiya 38, 2343-2353. doi: 10.1016 / j.psyneuen.2013.05.005
Culver, KE, Szechtman, H., Levant, B. (2008). Xinpirolun davranış həssaslığı olan siçovullarda Dopamin D2 kimi reseptorun bağlanması: Ro 41-1049 ilə əvvəlcədən müalicənin təsiri. Avro. J. Pharmacol. 592, 67-72. doi: 10.1016 / j.ejphar.2008.06.101
Dalman, FC, O'Malley, KL (1999). dopaminerjik orta beyin neyronlarının mədəniyyətlərində kappa-opioid tolerantlığı və asılılığı. J. Neurosci. 19, 5750–5757. doi: 10.1523/JNEUROSCI.19-14-05750.1999
De Vries, TJ, Schoffelmeer, AN, Binnekade, R., Raasø, H., Vanderschuren, LJ (2002). Dopamin D2 reseptorlarının vasitəçiliyi ilə kokain və eroin axtaran davranışa dönüş vaxtdan asılıdır və davranış həssaslığı ilə əlaqədardır. Neuropsychopharmacology 26, 18–26. doi: 10.1016/S0893-133X(01)00293-7
Devine, DP, Leone, P., Pocock, D., Wise, RA (1993). Bazal mezolimbik dopamin salınmasının modulyasiyasında ventral tegramal mu, delta və kappa opioid reseptorlarının diferensial iştirakı: in vivo mikrodializ tədqiqatlarında. J. Pharmacol. Exp. Ther. 266, 1236-1246.
Di Chiara, G., Imperato, A. (1988). Mu və kappa tiryəkli agonistlərin nüvələrin böyüdülməsində və sərbəst hərəkət edən siçovulların dorsal kaudatında dopamin buraxılmasına əks təsirləri. J. Pharmacol. Exp. Ther. 244, 1067-1080. doi: 10.1073 / pnas.85.14.5274
Dvorkin, A., Perreault, ML, Szechtman, H. (2006). Obsesif-kompulsif pozğunluğun bir heyvan modelində dopamin agonist quinpirole'nin təkrar inyeksiyası ilə nəticələnən məcburi yoxlamanın inkişafı və müvəqqəti təşkili. Behav. Brain Res. 169, 303-311. doi: 10.1016 / j.bbr.2006.01.024
Ebner, SR, Roitman, MF, Potter, DN, Rachlin, AB, Chartoff, EH (2010). Kappa opioid reseptor agonist salvinorin A-nin depresif bənzər təsirləri, nüvələrin böyüdülməsində phasic dopamin salınmasının azalması ilə əlaqələndirilir. Psixofarmakol. (Berl) 210, 241–252. doi: 10.1007/s00213-010-1836-5
Edvards, NJ, Tejeda, HA, Pignatelli, M., Zhang, S., McDevitt, RA, Wu, J., et al. (2017). VTA-nın inhibe arxitekturasında dövrə spesifikliyi kokainlə əlaqəli davranışı tənzimləyir. Nat. Neurosci. 20 (3), 438–448. doi: 10.1038 / nn.4482
Ehrich, JM, Phillips, PEM, Chavkin, C. (2014). Kappa opioid reseptorlarının aktivləşdirilməsi, siçan nüvəsinin böyüdülməsində in vivo ilə qeydə alınmış evakuasiya olunmuş dopamin ifrazının kokain səbəbi artmasını gücləndirir. Neuropsychopharmacology 39, 3036-3048. doi: 10.1038 / npp.2014.157
Ehrich, JM, Messinger, DI, Knakal, CR, Kuhar, JR, Schattauer, SS, Bruchas, MR və s. (2015). Kappa Opioid Reseptoru ilə əlaqəli bir Vvers VTA Dopamin Neyronlarında p38 MAPK aktivləşdirilməsini tələb edir. J. Neurosci. 35, 12917-12931. doi: 10.1523 / JNEUROSCI.2444-15.2015
Eilam, D., Szechtman, H. (1989). D-2 agonist quinpirolun lokomotivə və hərəkətlərə bifazik təsiri. Avro. J. Pharmacol. 161, 151–157. doi: 10.1016/0014-2999(89)90837-6
Eilam, D., Szechtman, H. (2005). Obsessiv-kompulsif pozğunluğun bir heyvan modeli kimi psixostimulyasiya edən davranış: məcburi rituallar formasına etoloji bir yanaşma. CNS Spectr. 10, 191–202. doi: 10.1017 / S109285290001004X
Escobar, AP, Cornejo, FA, Andres, ME, Fuentealba, JA (2012). Kappa opioid reseptor agonisti U69593 ilə təkrarlanan müalicə ampulada həssas olan siçovullarda lokomotor həssaslığının ifadəsi deyil, güclənmiş K + induksiyalı dopamin ifrazını bərpa edir. Neurochem. Int. 60 (4), 344–349. doi: 10.1016 / j.neuint.2012.01.014
Escobar, AP, Cornejo, FA, Olivares-Costa, M., González, M., Fuentealba, JA, Gysling, K., et al. (2015). Quinpirole həssas siçovulların nüvəsindəki azaldılmış dopamin və glutamat nörotransmissiya, D2 autorepeptor funksiyasının inhibe edilməsinə işarə edir. J. Neurochem. 134, 1081–1090. doi: 10.1111 / jnc.13209
Escobar, AP, González, MP, Meza, RC, Noches, V., Henny, P., Gysling, K., et al. (2017). Siçovullarda quinpirol ilə əlaqəli lokomotor həssaslıqda dopamin D2 reseptor funksiyasının kappa opioid reseptoru potensiallaşdırma mexanizmləri. Int. J. Neuropsychopharmacol. 20, 660–669. doi: 10.1093 / ijnp / pyx042
Everitt, BJ, Robbins, TW (2005). Narkomaniya üçün sinir gücləndirmə sistemləri: hərəkətlərdən vərdişlərə qədər. Nat. Neurosci. 8, 1481-1489. doi: 10.1038 / nn1579
Fattore, L., Melis, M. (2016). Dürtüsel və məcburi davranışlarda cins fərqləri: narkomaniyaya bir diqqət. Addict. Biol. 21 (5), 1043–1051. doi: 10.1111 / adb.12381
Ferrario, CR, Gorny, G., Crombag, HS, Li, Y., Kolb, B., Robinson, TE (2005). İdarə olunandan kokain istifadəsinə keçidlə əlaqəli sinir və davranış plastikliyi. Biol. Psixi. 58 (9), 751-9.
Figee, M., Pattij, T., Willuhn, I., Luigjes, J., van den Brink, W., Goudriaan, A., et al. (2016). Obsesif-kompulsif pozğunluq və asılılıqlarda kompulsivlik. Avro. Neuropsychopharmacol. 26, 856-868. doi: 10.1016 / j.euroneuro.2015.12.003
Ford, CP, Mark, GP, Williams, JT (2006). Mezolimbik dopamin neyronlarının xüsusiyyətləri və opioid inhibisiyası hədəf yerə görə dəyişir. J. Neurosci. 26, 2788-2797. doi: 10.1523 / JNEUROSCI.4331-05.2006
Ford, CP, Beckstead, MJ, Williams, JT (2007). Somatodendritik dopamin inhibe postsinaptik cərəyanların Kappa opioid inhibe. J. Neurophysiol. 97, 883-891. doi: 10.1152 / jn.00963.2006
Fuentealba, JA, Gysling, K., Magendzo, K., Andres, ME (2006). Selektiv kappa-opioid reseptor agonist U-69593 təkrar tətbiq edilməsi, siçovul nüvəsinin böyüdülməsində stimullaşdırılmış dopamin hüceyrədənkənar səviyyəsini artırır. J. Neurosci. Res. 84, 450-459. doi: 10.1002 / jnr.20890
Fuentealba, JA, Gysling, K., Andres, ME (2007). Seçici kappa-opioid reseptor agonist U-69593-in təkrar administrasiyası tərəfindən törədilən amfetamin artan lokomotor reaksiya. Synapse 61, 771-777. doi: 10.1002 / syn.20424
Gehrke, BJ, Chefer, VI, Shippenberg, TS (2008). Salvinorinin A-nin kəskin və təkrar tətbiq edilməsinin siçovul dorsal striatumundakı dopamin funksiyasına təsiri. Psixofarmakol. (Berl) 197, 509–517. doi: 10.1007/s00213-007-1067-6
Giuliano, C., Belin, D., Everitt, BJ (2019). Kompulsif spirtli davranış davranış üzərində dorsolateral striatal nəzarəti dayandıra bilməməsi nəticəsində yaranır. J. Neurosci. 39 (9), 1744–1754. doi: 10.1523 / JNEUROSCI.2615-18.2018
Goto, Y., Grace, AA (2008). Nüvə accumbensində limbik və kortikal məlumatların işlənməsi. Trends Neurosci. 31, 552-558. doi: 10.1016 / j.tins.2008.08.002
Grace, AA, Bunney, BS (1980). Nigral dopamin neyronları: hüceyrədaxili qeyd və L-dopa enjeksiyonu və histofluoresans ilə eyniləşdirmə. Elm 210, 654-656. doi: 10.1126 / science.7433992
Grey, AM, Rawls, SM, Shippenberg, TS, McGinty, JF (1999). Ü-opioid agonisti olan U-69593, kəskin amfetamin təsirli davranışlarını və ventral striatumdakı dopamin və qlutamat səviyyəsini kalsiuma bağlı dializat səviyyələrini azaldır. J. Neurochem. 73, 1066-1074. doi: 10.1046 / j.1471-4159.1999.0731066.x
Graziane, NM, Polter, AM, Briand, LA, Pierce, RC, Kauer, JA (2013). Kappa opioid reseptorları stresə səbəb olan kokain axtaran və sinaptik plastikliyi tənzimləyir. Neyron 77, 942-954. doi: 10.1016 / j.neuron.2012.12.034
Heidbreder, CA, Shippenberg, TS (1994). U-69593 bazal accumbens dopamin normallaşdıraraq kokainin həssas olmasının qarşısını alır. Neuroreport 5, 1797–1800. doi: 10.1097/00001756-199409080-00028
Hernandez-Avila, CA 1., Rounsaville, BJ, Kranzler, HR (2004). Opioid-, sirr və spirtdən asılı qadınlar, maddə asılılığı müalicəsində daha sürətli inkişaf göstərir. Drug Alkoqolundan asılıdır. 74 (3), 265–272. doi: 10.1016 / j.drugalcdep.2004.02.001
Hoffman, AF, Spivak, CE, Lupica, CR (2016). Dopamin nəqliyyat inhibitorları tərəfindən məhdud bir diffuziya modeli və sürətli taranan tsiklik voltametriya ilə izah edilən genişləndirilmiş dopamin sərbəst buraxılması. ACS Chem. Neurosci. 7, 700-709. doi: 10.1021 / acschemneuro.5b00277
Holden, C. (2001). "Davranış" asılılığı: varmı? Elm 294, 980-982. doi: 10.1126 / science.294.5544.980
Imperato, A., Di Chiara, G. (1988). Yerli tətbiq olunan D-1 və D-2 reseptor agonistlərinin və antaqonistlərin beyin dializi ilə təsirləri. Avro. J. Pharmacol. 156, 385–393. doi: 10.1016/0014-2999(88)90284-1
Cekson, KJ, McLaughlin, JP, Carroll, FI, Damaj, MI (2013). Kappa opioid reseptor antaqonistinin, norbinaltorfiminin, siçanlarda nikotinli şərtli yer seçiminin stresə və dərmanla əlaqəli bərpa vəziyyətinə təsiri. Psixofarmakol. (Berl) 226, 763–768. doi: 10.1007/s00213-012-2716-y
Kivell, B., Uzelac, Z., Sundaramurthy, S., Rajamanickam, J., Ewald, A., Chefer, V., et al. (2014). Salvinorin A, kappa opioid reseptoru və ERK1 / 2-dən asılı mexanizm vasitəsi ilə dopamin ötürücü funksiyasını tənzimləyir. Neurofarmakoloji 86, 228-240. doi: 10.1016 / j.neuropharm.2014.07.016
Koeltzow, TE, Austin, JD, Vezina, P. (2003). Quinpirole üçün davranış həssaslığı artan nüvə accumbens dopamin daşması ilə əlaqələndirilmir. Neurofarmakoloji 44, 102–110. doi: 10.1016/S0028-3908(02)00328-3
Koob, GF, Volkow, ND (2016). Asılılığın neyrobiologiyası: bir neyrokirkulyasiya analizi. Lancet Psixiatriya 3, 760–773. doi: 10.1016/S2215-0366(16)00104-8
Koob, GF (2013). Asılılıq bir mükafat çatışmazlığı və stres səthi pozğunluqdur. Cəbhə. Psixiatriya 4, 72. doi: 10.3389 / fpsyt.2013.00072
Labad, J. 1., Menchon, JM, Alonso, P., Segalas, C., Jimenez, S., Jaurrieta, N., et al. (2008). Obsesif-kompulsif simptom ölçülərindəki cins fərqləri. Anksiyete qazanmaq. 25 (10), 832-838. doi: 10.1002 / da.20332
Maisonneuve, IM, Archer, S., Glick, SD (1994). U50,488, kappa opioid reseptor agonisti, siçovulların nüvəsindəki hüceyrədaxili dopamin artımını kokain ilə əlaqələndirir. Neurosci. Lett. 181, 57–60. doi: 10.1016/0304-3940(94)90559-2
Mansour, A., Fox, CA, Akil, H., Watson, SJ (1995). Siçovul CNS-də opioid-reseptor mRNA ifadəsi: anatomik və funksional təsir. Trends Neurosci. 18 (1), 22–29. doi: 10.1016/0166-2236(95)93946-U
Mansour, A., Burke, S., Pavlic, RJ, Akil, H., Watson, SJ (1996). Siçovul CNS və hipofizdə klonlanmış kappa 1 reseptorunun immunohistokimyəvi lokalizasiyası. Nevrologiyada 71, 671–690. doi: 10.1016/0306-4522(95)00464-5
Margolis, EB, Karhanis, AN (2019). Kappa opioid reseptoruna edilən dopaminergik hüceyrə və dövrə töhfələri vasitəçilik amili. Neurochem. Int. 129, 104504. doi: 10.1016 / j.neuint.2019.104504
Margolis, EB, Hjelmstad, GO, Bonci, A., Fields, HL (2003). Kappa-opioid agonistləri orta beyin dopaminergik neyronları birbaşa inhibə edirlər. J. Neurosci. 23, 9981–9986. doi: 10.1523/JNEUROSCI.23-31-09981.2003
Margolis, EB, Hjelmstad, GO, Bonci, A., Fields, HL (2005). Həm Kappa, həm də Mu Opioid Agonistləri, Ventral Tegmental Bölgə Neyronlarına Glutamatergic Girişini maneə törədirlər. J. Neurophysiol. 93, 3086-3093. doi: 10.1152 / jn.00855.2004
Margolis, EB, Lock, H., Chefer, VI, Shippenberg, TS, Hjelmstad, GO, Fields, HL (2006). Kappa opioidləri prefrontal korteksə yönəlmiş dopaminergik neyronları seçici şəkildə idarə edir. Proc. Natl. Acad. Sci. ABŞ 103, 2938-2942. doi: 10.1073 / pnas.0511159103
Margolis, EB, Mitchell, JM, Ishikawa, J., Hjelmstad, GO, Fields, HL (2008). Midbrain dopamin neyronları: proyeksiya hədəfi fəaliyyət potensial müddəti və dopamin D (2) reseptorun inhibisyonunu təyin edir. J. Neurosci. 28, 8908-8913. doi: 10.1523 / JNEUROSCI.1526-08.2008
Mathews, IZ, McCormick, CM (2007). Gec yetkinlik dövründə qadın və kişi siçovullar amfetamin təsirli lokomotor fəaliyyətində yetkinlərdən fərqlənir, lakin amfetamin üçün şərtli yer seçimində deyil. Behav. Pharmacol. 18, 641–650. doi: 10.1097/FBP.0b013e3282effbf5
Mathis, MA 1., Pd, A., Funaro, G., RC, T., Moraes, I., AR, T., et al. (2011). Obsesif-kompulsif pozğunluqdakı cins fərqləri: bir ədəbiyyat icmalı. Braz. J. Psixiatriya 33 (4), 390-399. doi: 10.1590 / S1516-44462011000400014
McLaughlin, JP, Marton-Popovici, M., Chavkin, C. (2003). Kappa opioid reseptorlarının antaqonizmi və prodynorfin geninin pozulma bloku stresə səbəb olan davranış reaksiyaları. J. Neurosci. 23 (13), 5674–5683. doi: 10.1523 / JNEUROSCI.23-13-05674.2003
McLaughlin, JP, Land, BB, Li, S., Pintar, JE, Chavkin, C. (2006a). Kappa opioid reseptorlarının U50,488 mimikası tərəfindən aktivləşdirilməsindən əvvəl kokain yerini üstünlük təşkil edən kondensatı gücləndirmək üçün məcburi üzmə stressini təkrarladı. Neuropsychopharmacology 31, 787-794. doi: 10.1038 / sj.npp.1300860
McLaughlin, JP, Li, S., Valdez, J., Chavkin, TA, Chavkin, C. (2006b). Sosial məğlubiyyət stresə səbəb olan davranış reaksiyaları endogen kappa opioid sistemi ilə vasitəçilik olunur. Neuropsychopharmacology 31 (6), 1241–1248. doi: 10.1038 / sj.npp.1300872
Milesi-Hallé, A., McMillan, DE, Laurenzana, EM, Birnes-Bleyk, KA, Ouens, SM (2007). (+) - Amfetamin və (+) - kişi və qadın Sprague-Dawley siçovullarında metamfetamin təsirli davranış reaksiyalarında cins fərqləri. Pharmacol. Biochem. Behav. 86, 140-149. doi: 10.1016 / j.pbb.2006.12.018
Negus, SS, Mello, NK, Portoghese, PS, Lin, CE (1997). Kappa opioidlərinin rezus meymunları tərəfindən kokainin özünü idarə etməsinə təsiri. J. Pharmacol. Exp. Ther. 282, 44-55.
Nestler, EJ (2001). Bağımlılığa əsaslanan uzun müddətli plastisiyanın molekulyar əsasları. Nat. Rev. Neurosci. 2, 119-128. doi: 10.1038 / 35053570
Perreault, ML, Graham, D., Bisnaire, L., Simms, J., Hayton, S., Szechtman, H. (2006). Kappa-opioid agonist U69593, lokomotor həssaslığı D2 / D3 agonist quinpirole-a kömək edir: pre-və postsinaptik mexanizmlər. Neuropsychopharmacology 31, 1967-1981. doi: 10.1038 / sj.npp.1300938
Perreault, ML, Seeman, P., Szechtman, H. (2007). Kappa-opioid reseptorun stimullaşdırılması obsesif-kompulsif pozğunluğun (OCD) quinpirole həssaslaşma modelində kompulsif yoxlanmanın patogenezini sürətləndirir. Behav. Neurosci. 121, 976-991. doi: 10.1037 / 0735-7044.121.5.976
Pirs, RC, Kalivas, PW (1997). Amfetamin bənzər psixostimulyatorlara davranış həssaslığının ifadəsinin dövrə modeli. Brain Res. Brain Res. Rev. 25, 192–216. doi: 10.1016/S0165-0173(97)00021-0
Polter, AM, Bishop, RA, Briand, LA, Graziane, NM, Pierce, RC, Kauer, JA (2014). Kappa Opioid Reseptorlarının Poststress Bloku İnhibe Sinapslarının Uzunmüddətli Potensiasiyasını xilas edir və Kokain axtarmağın bərpa olunmasına mane olur. Biol. Psixiatriya 76, 785-793. doi: 10.1016 / j.biopsych.2014.04.019
Puig-Ramos, A., Santiago, GS, Segarra, AC (2008). Bir kappa opioid reseptor agonisti olan U-69593, qadın siçovullarında kokainlə əlaqəli davranış həssaslığını azaldır. Behav. Neurosci. 122, 151-160. doi: 10.1037 / 0735-7044.122.1.151
Robbins, TW, Gillan, CM, Smith, DG, de Wit, S., Ersche, KD (2012). Dürtüselliyin və kompulsivliyin neyrokoqnitiv endofenotipləri: ölçülü psixiatriya. Trends Cognit. Sci. 16, 81-91. doi: 10.1016 / j.tics.2011.11.009
Roberts, DCS, Bennett, SAL, Vickers, GJ (1989). Estrogen dövrü siçovullarda mütərəqqi nisbət cədvəlində kokainin özünü idarə etməsinə təsir göstərir. Psixofarmakol. (Berl) 98, 408-411. doi: 10.1007 / BF00451696
Robinson, TE, Berridge, KC (1993). Narkotik istəklərinin neyron əsasları: asılılığın stimullaşdırıcı-həssaslaşdırma nəzəriyyəsi. Brain Res. Brain Res. Rev. 18, 247–291. doi: 10.1016/0165-0173(93)90013-P
Robinson, TE, Berridge, KC (2001). Həvəsləndirici-həssaslıq və asılılıq. Narkomaniya 96, 103-114. doi: 10.1046 / j.1360-0443.2001.9611038.x
Gül, JH, Karxanis, AN, Steiniger-Brach, B., Jones, SR (2016). Nalmefenin dopamin qəbul etmə nisbətinə və nüvədəki kappa opioid reseptor fəaliyyətinə xroniki aralıqlı etanol məruz qaldıqdan sonra artan təsiri. Int. J. Mol. Sci. 17, 1216. doi: 10.3390 / ijms17081216
Schlosburg, JE, Whitfield, TW, Jr., Park, PE, Crawford, EF, George, O., Vendruscolo, LF, et al. (2013). Opioid reseptorlarının uzunmüddətli antaqonizmi, eroin qəbulunun artması və motivasiyasının qarşısını alır. J. Neurosci. 33 (49), 19384–19392. doi: 10.1523 / JNEUROSCI.1979-13.2013
Sesack, SR, Pickel, VM (1992). Siçovul ventral tegramal bölgədəki enkefalin və tirozin hidroksilaza immunoreaktivliyinin ikiqat ultrastruktur quruluşu: opiat-dopamin qarşılıqlı təsiri üçün çoxlu substrat. J. Neurosci. 12, 1335–a1350. doi: 10.1523/JNEUROSCI.12-04-01335.1992
Sesack, SR, Aoki, C., Pickel, VM (1994). Orta beyin dopamin neyronlarında D2 reseptoruna bənzər immunoreaktivliyin ultrastruktur quruluşu və onların striatal hədəfləri. J. Neurosci. 14, 88–106. doi: 10.1523/JNEUROSCI.14-01-00088.1994
Sesia, T., Bizup, B., Grace, AA (2013). Obsesif-kompulsif bozuklukun heyvan modellərinin qiymətləndirilməsi: phasic dopamin neyron fəaliyyətinin əlaqəsi. Int. J. Neuropsychopharmacol. 16, 1295–1307. doi: 10.1017 / S146114571200154X
Shippenberg, TS, Zapata, A., Chefer, VI (2007). Dynorfin və narkomaniyanın patofiziologiyası. Farmakol. Ther. 116, 306-321. doi: 10.1016 / j.pharmthera.2007.06.011
Smith, JA, Loughlin, SE, Leslie, FM (1992). kappa-Opioid inhibe [3H] siçovul ventral mesencephalic dağılmış hüceyrə mədəniyyətlərindən azad. Mol. Pharmacol. 42.
Smith, JW, Fetsko, LA, Xu, R., Wang, Y. (2002). Dopamin D2L reseptoru nokaut edən siçanlar, morfinin müsbət və mənfi gücləndirici xüsusiyyətlərində və öyrənmənin qarşısını alır. Nevrologiyada 113 (4), 755–765. doi: 10.1016/S0306-4522(02)00257-9
Smith, JS, Schindler, AG, Martinelli, E., Gustin, RM, Bruchas, MR, Chavkin, C. (2012). Amigdala dynorphin / κ-opioid reseptor sisteminin stresdən qaynaqlanan aktivləşdirilməsi nikotinlə şərtlənmiş yerə üstünlük verir. J. Neurosci. 32, 1488-1495. doi: 10.1523 / JNEUROSCI.2980-11.2012
Spanagel, R., Herz, A., Shippenberg, TS (1992). Qarşı tonikli aktiv endogen opioid sistemlər mezolimbik dopaminergik yolunu modullaşdırır. Proc. Natl. Acad. Sci. 89, 2046-2050. doi: 10.1073 / pnas.89.6.2046
Speciale, SG, Manaye, KF, Sadeq, M., Alman, DC (1993). Siçovulun orta beyin dopaminergik bölgələrində opioid reseptorları. II. Kappa və delta reseptoru avtoreqrafiyası. J. Neyral Transm. General təriqət. 91, 53-66. doi: 10.1007 / BF01244918
Sperling, RE, Gomes, SM, Sypek, EI, Carey, AN, McLaughlin, JP (2010). Etanol şərtli yer üstünlüyü və özünü idarəetmə ilə stresdən qaynaqlanan potensiasiyanın endogen kappa-opioid vasitəçiliyi. Psixofarmakol. (Berl) 210 (2), 199–209. doi: 10.1007/s00213-010-1844-5
Steketee, JD, Kalivas, PW (2011). Narkotik istənən: davranış həssaslığı və narkotik axtaran davranışa reabilitasiya sibley DR, ed. Pharmacol. Rev. 63, 348–365. doi: 10.1124 / pr.109.001933
Stuchlik, A., Radostová, D., Hatalova, H., Vales, K., Nekovarova, T., Koprivova, J., et al. (2016). OKB-nin quinpirole həssaslaşdırma siçovul modelinin etibarlılığı: heyvan və klinik tədqiqatlardan sübutları əlaqələndirmək. Cəbhə. Behav. Neurosci. 10, 209. doi: 10.3389 / fnbeh.2016.00209
Günəş, T., Song, Z., Tian, Y., Tian, W., Zhu, C., Ji, G., et al. (2019). Medial prefrontal korteksə basolateral amigdala girişi obsesif-kompulsif pozğunluq kimi yoxlanış davranışını idarə edir. Proc. Natl. Acad. Sci. ABŞ 116, 3799-3804. doi: 10.1073 / pnas.1814292116
Svingos, AL, Chavkin, C., Colago, EEO, Pickel, VM (2001). Opioid reseptorları və nüvədəki dopamin daşıyıcısı aksonal profilləri artır. Synapse 42, 185-192. doi: 10.1002 / syn.10005
Szechtman, H., Talangbayan, H., Eilam, D. (1993). Dopamin agonist quinpirole ilə əlaqəli həssaslığın ətraf mühit və davranış komponentləri. Behav. Pharmacol. 4, 405–410. doi: 10.1097/00008877-199308000-00014
Szechtman, H., Talangbayan, H., Canaran, G., Dai, H., Eilam, D. (1994). Dopamin agonist quinpirole və təklif olunan mərkəzi enerjiyə nəzarət mexanizmi tərəfindən tətbiq olunan davranış həssaslığının dinamikası. Psixofarmakol. (Berl) 115, 95-104. doi: 10.1007 / BF02244757
Szechtman, H., Sulis, W., Eilam, D. (1998). Quinpirole siçovullarda məcburi yoxlama davranışına səbəb olur: obsesif-kompulsif pozğunluğun (OCD) potensial heyvan modeli. Behav. Neurosci. 112, 1475-1485. doi: 10.1037 / 0735-7044.112.6.1475
Szechtman, H., Culver, K., Eilam, D. (1999). Obsesif-kompulsif pozğunluqda (OKB) dopamin sistemlərinin rolu: yeni bir psixostimulyasiya edən heyvan modelinin təsirləri. Pol. J. Farmakol. 51, 55-61.
Szechtman, H., Eckert, MJ, Tse, WS, Boersma, JT, Bonura, C a, JZ, M., et al. (2001). Obsesif-Kompulsif Bozukluk (OCD) bir heyvan modeli kimi quinpirole həssas siçovulların kompulsif yoxlanış davranışı: forma və nəzarət. BMC Neurosci. 2, 4. doi: 10.1186/1471-2202-2-4
Szechtman, H., Ahmari, SE, Beninger, RJ, Eilam, D., Harvey, BH, Edemann-Callesen, H., et al. (2017). Obsesif-kompulsif pozğunluq: Heyvan modellərindən anlayışlar. Neurosci. Biobehav. Rev. 76, 254-279. doi: 10.1016 / j.neubiorev.2016.04.019
Tejeda, HA, Bonci, A. (2019). Dopamin dinamikasının dinorfin / kappa-opioid reseptoru nəzarəti: mənfi affektiv vəziyyətlər və psixiatrik xəstəliklər üçün təsir. Brain Res. 1713, 91-101. doi: 10.1016 / j.brainres.2018.09.023
Tejeda, HA, Wu, J., Kornspun, AR, Pignatelli, M., Kashtelyan, V., Krashes, M. J., et al. (2017). Yol və hüceyrəyə xas spesifik kappa-opioid reseptor modulyasiyası, həyəcan-inhibisiya balansının diferensial olaraq qapıları d1 və d2 akumbens neyron aktivliyini. Neyron 93 (1), 147-163. doi: 10.1016 / j.neuron.2016.12.005
Thompson, AC, Zapata, A., Ədalət, JB, Vaughan, RA, Sharpe, LG, Shippenberg, TS (2000). Kappa-opioid reseptorunun aktivləşdirilməsi, nüvədəki dopamin qəbulunu dəyişdirir və kokainin təsirinə qarşı çıxır. J. Neurosci. 20, 9333–9340. doi: 10.1523/JNEUROSCI.20-24-09333.2000
Thompson, D., Martini, L., Whistler, JL (2010). Siçan striatumundakı D1 və D2 dopamin reseptorlarının dəyişdirilmiş nisbəti kokainə qarşı davranış həssaslığı ilə əlaqələndirilir. PloS One 5, e11038. doi: 10.1371 / jurnal.pone.0011038
Usiello, A., Baik, J.-H., Rougé-Pont, F., Picetti, R., Dierich, A., LeMeur, M., et al. (2000). Dopamin D2 reseptorlarının iki izoformunun fərqli funksiyaları. təbiət 408, 199-203. doi: 10.1038 / 35041572
Van Bockstaele, EJ, Pickel, VM (1995). Siçovul beyinindəki nüvələrə vurulan ventral tegramal bölgədəki GABA tərkibli neyronlar. Brain Res. 682 (1-2), 215–221. doi: 10.1016/0006-8993(95)00334-M
Van't Veer, A., Bechtholt, AJ, Onvani, S., Potter, D., Wang, Y., Liu-Chen, LY, et al. (2013). Beyin dopamin neyronlarından kappa-opioid reseptorlarının ləğvi anksiyolitik oxşar təsirə malikdir və kokainə səbəb olan plastisiyanı artırır. Neuropsychopharmacology 38 (8), 1585–1597. doi: 10.1038 / npp.2013.58
Vanderschuren, LJ, Kalivas, PW (2000). Davranış həssaslığının induksiyasında və ifadəsində dopaminergik və glutamaterjik ötürülmə ilə əlaqəli dəyişikliklər: preklinik tədqiqatlara kritik bir baxış. Psixofarmakol. (Berl) 151 (2-3), 99-120. doi: 10.1007 / s002130000493
Volkow, ND, Aqil, RA, Baler, R. (2017). Dopamin motiv sistemi: narkotik və qida asılılığı üçün təsirlər. Nat. Rev. Neurosci. 18, 741–752. doi: 10.1038 / nrn.2017.130
Walker, QD, Rooney, MB, Wightman, RM, Kuhn, CM (2000). Sürətli tsiklik voltametriya ilə ölçüldüyü kimi, qadınlarda dofaminin sərbəst buraxılması və tutulması qadın siçan striatumundan daha çoxdur. Nevrologiyada 95, 1061–1070. doi: 10.1016/S0306-4522(99)00500-X
Wee, S., Koob, GF (2010). Dynorphin-opioid sisteminin sui-istifadə dərmanlarının gücləndirici təsirində rolu. Psixofarmakol. (Berl) 210, 121–135. doi: 10.1007/s00213-010-1825-8
Wee, S., Orio, L., Ghirmai, S., Cashman, JR, Koob, GF (2009). Kappa opioid reseptorlarının maneə törədilməsi siçovullarda kokainə geniş giriş imkanı olan kokain qəbulunu artırdı. Psixofarmakol. (Berl) 205, 565–575. doi: 10.1007/s00213-009-1563-y
Whitfield, TW, Jr., Schlosburg, JE, Wee, S., Gould, A., George, O., Grant, Y., et al. (2015). the Nüvədəki opioid reseptorları metamfetamin qəbulunun artmasına vasitəçilik edir. J. Neurosci. 35 (10), 4296–4305. doi: 10.1523 / JNEUROSCI.1978-13.2015
Williams, MT, Mugno, B., Franklin, M., Faber, S. (2013). Obsesif-kompulsif pozğunluqda simptom ölçüləri: məruz qalma və ritual qarşısının alınması ilə fenomenologiya və müalicə nəticələri. Psixopatologiya 46, 365-376. doi: 10.1159 / 000348582
Wilson, CJ, Young, SJ, Groves, PM (1977). Təzyiq nigrasında neyron sünbül qatarlarının statistik xüsusiyyətləri: hüceyrə növləri və onların qarşılıqlı əlaqələri. Brain Res. 136, 243–260. doi: 10.1016/0006-8993(77)90801-0
Aqil, RA (2009). Nigrostriatal üçün rollar - yalnız mezokortikolimbik deyil, mükafat və asılılıqdakı dopamin. Trends Neurosci. 32, 517-524. doi: 10.1016 / j.tins.2009.06.004
Xiao, L., Becker, JB (1994). Kişi və qadın siçovullarda hüceyrədənkənar striatal dopamin konsentrasiyasının kəmiyyət mikdializ təyini: estrogen dövrü və gonadektomiyanın təsiri. Neurosci. Lett. 180, 155–158. doi: 10.1016/0304-3940(94)90510-X
Açar sözlər: kappa opioid reseptoru, dopamin, kompulsivlik, amfetamin, quinpirol, lokomotor həssaslıq
Sitat: Escobar AdP, Casanova JP, Andrés ME və Fuentealba JA (2020) Kompulsif Davranışlarda Kappa Opioid və Dopamin Sistemləri arasındakı Crosstalk. Cəbhə. Pharmacol. 11: 57. doi: 10.3389 / fph.2020.00057
Alındı: 16 Oktyabr 2019; Qəbul edildi: 22 Yanvar 2020;
Yayınlandı: 18 Fevral 2020.
Düzenleyen:
Gonzalo E. Yevenes, Konsepsiya Universiteti, Çili
Tərəfindən nəzərdən:
Luis Gerardo Aquayo, Konsepsiya Universiteti, Çili
Hüqo Tejeda, Narkomanlığa qarşı Milli İnstitut (NIDA), Amerika Birləşmiş Ştatları
Cecilia Scorza, İnstitut İnstitutu Biológicas Clemente Estable (IIBCE), Uruqvay
Müəllif hüquqları © 2020 Escobar, Casanova, Andrés və Fuentealba. Bu, şərtlərə əsasən paylanmış açıq bir məqalədir Creative Commons Attribution Lisenziyası (CC BY). Müəllif (lər) və müəllif hüququ sahibi (lər) ini kredit verildiyi və qəbul edilmiş akademik praktikaya uyğun olaraq bu jurnalın əsli dərcinə istinad edildiyi halda, digər forumlarda istifadəsi, bölüşdürülməsi və ya bərpasına icazə verilir. Bu şərtlərə uyğun gəlməyən istifadəsi, paylanması və ya təkrar istifadəsinə icazə verilmir.
* Yazışma: Xose Antonio Fuentealba, [e-poçt qorunur]
†ORCID: Xose Antonio Fuentealba, orcid.org/0000-0003-0775-0675