ŞƏRHLƏR: piylənmə və qida asılılığı mövzusunda ən yaxşı tədqiqatçı tərəfindən nəzərdən keçirildi.
Həcmi 69, Sayı 4, 24 Fevral 2011, Səhifələr 664 – 679
http://dx.doi.org/10.1016/j.neuron.2011.02.016,
baxış
Paul J. Kenny1, ,
Davranış və molekulyar nevrologiya 1 Laboratoriyası, Molekulyar terapevtika şöbəsi, Scripps Araşdırma İnstitutu, Yupiter, FL 33458, ABŞ
________________________________________
Qidalanma balansında enerji balansını qorumaq üçün istehlak edilir. Ayrıca, istehlakçı qida da enerji statüsünden bağımsız hedonik özellikler üçün tüketilmektedir. Belə mükafatla əlaqəli istehlak istər artım tələb edənlərin kalorili istifadəsinə gətirib çıxara bilər və inkişaf etmiş ölkələrdə sürətlə artan obezitə nisbətlərində böyük bir günahkar hesab edilir. Bəslənmənin homeostatik mexanizmləri ilə müqayisədə, beyində hedonik sistemlərin qida qəbuluna necə təsir edəcəyi hələ az bilinir. Maraqlıdır ki, doyurucu yeməklərin həddən artıq istehlakı sui-istifadə dərmanlarına bənzəyən beyin mükafatlı dövrlərdə neuroadaptiv cavablar tetikleyebilir. Bundan əlavə, beyin mükafat sistemlərində oxşar genetik zəifliklər narkotik asılılığına və obeziteye meylli ola bilər. Burada bəslənmə davranışının hedonik aspektlərini tənzimləyən beyin dövriyyələri anlayışımızı əldə edən son inkişaflar nəzərdən keçiriləcəkdir. Həm də obezitenin və narkomaniyanın ümumi hedonik mexanizmləri paylaşa biləcəyinə dair ortaya çıxan dəlillər də nəzərdən keçiriləcəkdir.
________________________________________
Əsas mətn
"Yemək sevgisindən daha səmimi bir sevgi yoxdur."
-Ceorge Bernard Shaw
giriş
Bir bədən kütlə indeksi (BMI)> 30 olaraq təyin olunan piylənmə, piylənmənin anormal dərəcədə yüksək olduğu və hiperfagiya və ya metabolik nisbətin azalması ilə nəticələnə biləcəyi bir vəziyyətdir (O'Rahilly, 2009). Həddindən artıq piylənmə ürək-damar xəstəlikləri, xərçəng, tip 2 diabet və əhval-ruhiyyə ilə əlaqəli xəstəliklər üçün əsas risk faktorudur, obez insanlar tez-tez sosial damğalanma ilə üzləşirlər ([Bean və digərləri, 2008], [Xəstəliklərə Nəzarət və Profilaktika Mərkəzləri, 2009] və [Luppino et al., 2010]). Xəstəliklərə Nəzarət Mərkəzinə (CDC) görə, 1998-2000 arasında ABŞ-da obezliklə əlaqəli səhiyyə xərcləri təxminən 213 milyard dollar idi. Bundan əlavə, ABŞ-da hər il 300,000 ölüm həddindən artıq kilolu və piylənmə ilə əlaqəli xəstəliklərə aid edilə bilər (Allison və digərləri, 1999), piylənmə tütün istifadəsinin qarşısını alan ölümün ikinci səbəbidir. Buna baxmayaraq, Qərb cəmiyyətlərində obezliyin yayılması sürətlə artmağa davam edir, mövcud hesablamalar ABŞ-dakı yetkinlərin% 30-dan çoxunun obez olduğunu göstərir (Flegal və digərləri, 2010).
Qidalanma tənzimlənməsinin əksər konseptualizasiyaları iki paralel sistemin qida qəbuluna təsir göstərməsini təklif edir ([Hommel et al., 2006], [Lutter and Nestler, 2009] and [Morton et al., 2006]). Homeostatik sistem, enerji balansının lazımi səviyyəsini qorumaq üçün qidalanmanı stimullaşdıran və ya maneə törədən leptin, qrelin və insulin kimi aclıq, doyma və yağ səviyyəsini hormonal tənzimləyicilərdən ibarətdir. Anadangəlmə leptin çatışmazlığı kimi Homeostatik sistemin komponentlərindəki pozğunluq davamlı enerji balansının pozulmasına və obezliyin inkişafına səbəb ola bilər ([Campfield et al., 1995], [Halaas et al., 1995] və [Pelleymounter) et al., 1995]). Enerji homeostazını qorumaq üçün hipotalamik və beyin kök dövranlarında hormon tənzimləyicilərinin təsirini göstərən mexanizmlər başqa yerlərdə ətraflı təsvir edilmişdir və bu mövzu ilə maraqlanan oxucular bu mövzuda çox sayda mükəmməl rəylərə istinad etdilər (məsələn, [Abizaid) et al., 2006a] və [Gao and Horvath, 2007]).
Metabolik sistemlərə əlavə olaraq beyin mükafatlandırma sistemləri də qidalanma davranışında əhəmiyyətli rol oynayır ([Lutter and Nestler, 2009] və [Saper et al., 2002]). Ümumiyyətlə, yumşaq dadlı qidalar həddən artıq yeyilmir, dadlı qidalar isə enerji tələbləri ödənildikdən sonra da tez-tez istehlak olunur. Dadlı enerji ilə qidalanan yeməyin asanlığı obezlik üçün böyük bir ekoloji risk faktoru olaraq qəbul edilir (Volkow and Wise, 2005) və ləzzətli qidanın həddindən artıq istehlakı, son zamanlarda piylənmənin artmasına səbəb olan əsas amildir (Finkelstein et al., 2005], [Hill et al., 2003] və [Swinburn et al., 2009]). Həqiqətən, dadlı yeməyin xoş təsirlərini əldə etmək, müəyyən fərdlərdə homeostatik siqnalları ləğv edə biləcəyi güclü bir hərəkətverici qüvvədir ([Shomaker et al., 2010], [Sunday et al., 1983] və [Zheng et al., 2009]) . Bir seçim təqdim edildikdə, siçovullar çox miqdarda intravenöz kokain infüzyonlarını idarə etmək əvəzinə kalorisiz bir saxarin həllini istehlak etməyi üstün tuturlar (Lenoir və digərləri, 2007). Üstəlik, yaxşı bəslənmiş siçovullar könüllü olaraq şaxtalı, ətli pate, yer fıstığı yağı, Coca-Cola, M&M konfetləri, şokolad kimi dadlı qida məhsulları əldə etmək üçün həddindən artıq soyuqlara (−15 ° C), zərərli istilik ağrısına və ya zərərli ayaqaltıya məruz qalacaqlar. daha az dadlı standart çuxur sərbəst şəkildə mövcud olsa belə cips və ya qatıq damlaları ([Cabanac and Johnson, 1983], [Foo and Mason, 2005] və [Oswald et al., 2010]). Bu tapıntılar dadlı qida içərisindəki makronutrientlərin kalori dəyərindən asılı olmayaraq beyin mükafatlandırma sistemlərini nə qədər intensiv şəkildə stimullaşdıra biləcəyini və (dadlı qida istehlakı motivasiyasının nə qədər yüksək ola biləcəyini) göstərir. homeostatik enerji tələbləri olmadıqda belə. Kokain və ya nikotin kimi sui-istifadə dərmanları, kalori və qida dəyərlərindən məhrum olmasına baxmayaraq, eyni şəkildə yüksək səviyyədə istehlakçı davranışa səbəb ola bilər. Əslində, piylənmədə həddindən artıq yemək və asılılıqda həddindən artıq narkotik istifadəsi arasında çox oxşarlıqlar olduğundan (Volkow and Wise, 2004), piylənmənin beyin xəstəlikləri olaraq qəbul edilməli və qarşıdakı beşinci nəşrdə diaqnostik bir kateqoriya kimi göstərilməlidir. Ruhi Bozuklukların Diaqnostik və Statistik El Kitabının (DSM-V) ([Devlin, 2004] və [Volkow and O'Brien, 2005]). Qidalanma davranışının homeostatik mexanizmləri ilə müqayisədə, hedonik sistemlərin qida qəbuluna necə təsir göstərdiyi haqqında daha az bilinir. Eynilə, daxili və ya pəhrizlə əlaqəli dəyişikliklərin beyin mükafatlandırma sistemlərinin reaksiya qabiliyyətinə təsiri və bu təsirlərin həddindən artıq yeməyə və piylənməyə necə töhfə verməsi hələ də aydın deyil. Aşağıda xülasə edərək, kökəlmənin inkişafına töhfə verə biləcək beyin mükafatı fəaliyyətində yeməyin hedonik mexanizmləri və pəhriz səbəbi ilə dəyişikliklərimizdəki irəliləyişləri vurğulayan son məlumatlar verilmişdir.
Palitralı yeməyə cavab olaraq beyin mükafatlandırma sistemlərinin aktivləşdirilməsi: Enerji balansının hormonal tənzimləyiciləri ilə qarşılıqlı əlaqə
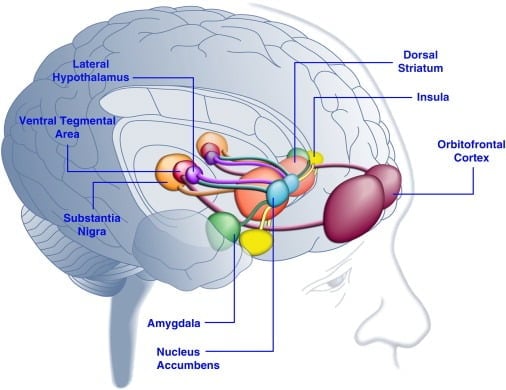
Ləzzətli qida istehlakı insanlarda əhval-ruhiyyəni artıra bilər ([Dallman et al., 2003] və [Macht and Mueller, 2007]) və laboratoriya heyvanlarında ([Imaizumi et al. 2001] və [ Sclafani et al., 1998]). Bu təsirlər, ehtimal ki, ləzzətli yeməklə beyin mükafatlandırma sistemlərinin stimullaşdırılması ilə əlaqədardır (Şəkil 1). Həqiqətən, insan beyninin görüntüləmə işləri qida və qida ilə əlaqəli vizual və ya qoxu ilə əlaqəli vizual və mükafatlandırılan kortikolimbik və mezo böyüdücü beyin dövranlarını, xüsusən də orbitofrontal korteks (OFC), insula, amigdala, hipotalamus, striatum və orta beyin bölgələrini aktivləşdirə biləcəyini göstərmişdir. ventral tegramal bölgə (VTA) və substantia nigra (SN) daxil olmaqla ([Bragulat et al., 2010], [Pelchat et al., 2004], [Schur et al., 2009] və [Simmons et al., 2005] ). Striatum, insula, ön cingüllü korteks və orta beyin quruluşları, hedonal təmsilçiliyin bu nöronal şəbəkə üçün roluna uyğun olaraq növlərindən (məsələn, qida, cinsiyyət, pul mükafatları) asılı olmayaraq mükafatların subyektiv dəyərini kodlaşdırır (Sescousse et al.) , 2010). Bunun əksinə olaraq, OFC, ləzzətli yemək ([Man et al., 2009], [Rolls, 2008] və [Sescousse et al., 2010) daxil olmaqla, xüsusi mükafat növlərinin dəyəri ilə əlaqəli təqdimatlarda xüsusilə məşhur rol oynayır. ). Aclıq insanlarda kortikolimbik və orta beyin bölgələrinin ləzzətli qida ilə təsirli aktivləşdirilməsini artıra bilər (LaBar et al., 2001). Məsələn, yüksək kalorili ləzzətli yeməyə cavab olaraq ventral striatum, amigdala, insula və OFC-nin aktivləşmə intensivliyi insan subyektləri yaxşı bəslənmədən çox ac olduqda daha yüksək idi (Goldstone et al. 2009). Bu, aclıq və pəhriz dövrlərinin "cazibədar" yeməyə ([Hofmann et al., 2010] və [Rolls et al. 1983) ləzzətli yeməklərin və istəklərin "güc" inin özünün bildirilmiş qiymətlərini artırması ilə uyğundur. ). Əksinə, həddindən artıq bəslənmə, xüsusən də insulinq korteksində və hipotalamusda xoşagəlməz qidaya neyron cavablarını azalda bilər (Cornier et al., 2009). Beləliklə, yemin hedonik dəyəri metabolik vəziyyətə təsir edir, belə ki, leptin və qrelin kimi maddələr mübadiləsinin tənzimləyicilərinin beyindəki hedonik sistemlərin fəaliyyətinə təsir göstərə bilər. Leptin və ya bağırsaqdan alınan postprandial amil peptidi YY3-36 (PYY) ([Batterham et al., 2007] və [Farooqi et al., 2007]) və ya mədə pozğunluğundan əziyyət çəkənlər ilə əlaqəli bu mövzuya uyğundur. təqlid etmə ilə əlaqəli beyin bölgələrində aktivliyi azaldıb (Wang et al., 2008). Əksinə, anadangəlmə leptin çatışmazlığı olan hiperfagik insan xəstələri qida şəkillərinə cavab olaraq ([Baicy et al., 2007] və [Farooqi et al., 2007]) insulent korteksində və striatumda artan aktivlik nümayiş etdirir. Bu şəxslərdə leptin əvəzedici terapiya gücləndirilmiş insuler və striatal aktivliyi azaldır və yeməyin özünəməxsus bildirilməsini azaldır ([Baicy et al., 2007] və [Farooqi et al., 2007]). Leptin müalicəsi eyni zamanda dopamin reseptor antaqonisti α-flupenthixol (Figlewicz et al., 2001) ilə bənzər qida məhdudlaşdıran siçovullarda saxarozanın mükafatlandırıcı xüsusiyyətlərini də maneə törədir. Bundan əlavə, leptin reseptorları VTA və SN (Figlewicz et al., 2003) arasında orta beyin dopamin neyronlarında ifadə edilir, leptinin mesostriatal dopamin ötürülməsinin modulyasiyası yolu ilə qidalanma davranışının hedonic aspektlərinə təsir göstərə biləcəyini göstərir. Bu ehtimalı təsdiqləyərək, VTA-ya leptin infüzyonları dopamin neyronlarının fəaliyyətini və siçovullarda qida qəbulunu azaltdı (Hommel et al., 2006; həmçinin Krügel et al., 2003). Əksinə, VTA-da leptin reseptorlarının yıxılması qida qəbulunu artırdı, lokomotor aktivliyi artırdı və siçovullarda ləzzətli qidaya üstünlük artdı (Hommel et al. 2006). Buna görə Leptin, mesoaccumbens dopamin ötürülməsinə, mükafat və motivasiyada çox təsir edən, lakin enerji homeostazında daha az təsir göstərən bir neyrotransmitter sisteminə təsir göstərir ([de Araujo et al. 2010] və [Vucetic and Reyes, 2010]). Bu yaxınlarda, aclıq ilə əlaqəli hormon ghrelin ([Kojima et al., 1999] və [Nakazato et al., 2001]), beyində hedonik sistemlərin qidalanma səbəblərinə cavab olaraq aktivləşdirilməsini gücləndirmək üçün göstərilmişdir (Malik et al.) , 2008). Xüsusilə, grelin, obez insanlarda yüksək ləzzətli yeməklərin şəkillərinə cavab olaraq OFC, amigdala, insula, striatum, VTA və SN-nin aktivləşdirilməsini artırdı (Malik et al., 2008). Siçovullarda ghrelin orta beyin dopamin sistemlərinə ([Abizaid et al., 2006b], [Jerlhag et al., 2006] and [Jerlhag et al., 2007]) stimullaşdırıcı təsir göstərir və ləzzətli yeməyin (Perello) mükafatlandırıcı dəyərini artırır. et al., 2010).
Şəkil 1. Dadlı Yemək və ya Yeməklə əlaqəli işarələrə cavab olaraq insan beyininin aktiv olduğu sahələr. Orbitofrontal korteks və amigdalanın yeməyin mükafat dəyəri ilə əlaqəli məlumatları kodladığı düşünülür ([Baxter and Murray, 2002], [Holland and Gallagher, 2004], [Kringelbach et al., 2003], [O'Doherty et al. , 2002] və [Rolls, 2010]). İnsula yeməyin dadı və hedonik qiymətləndirilməsi ilə əlaqəli məlumatları işləyir ([Ballein and Dickinson, 2000] and [Small, 2010]). Ventral tegmental nahiyədən və substansiya nigradan dopaminerjik giriş alan nüvə akumbens və dorsal striatum qidanın motivasiya və stimullaşdırıcı xüsusiyyətlərini tənzimləyir ([Baicy və digərləri, 2007], [Berridge, 1996], [Berridge, 2009], [Farooqi et al., 2007], [Malik et al., 2008] və [Söderpalm and Berridge, 2000]). Yanal hipotalamus dadlı qidaya mükafatlandırıcı cavabları tənzimləyə bilər və qida axtaran davranışları idarə edə bilər (Kelley və digərləri, 1996). Bu beyin strukturları qidanın hedonik xüsusiyyətləri haqqında öyrənməyi tənzimləmək, diqqət və səyləri qida mükafatları almağa yönəltmək və qida mükafatlarının mövcudluğunu proqnozlaşdıran ətraf mühit stimullarının təşviq dəyərini tənzimləmək üçün uyğun bir şəkildə hərəkət edirlər (Dagher, 2009). Aydınlıq üçün bu strukturlar arasındakı bütün əlaqələr göstərilmir.
Bənzər beyin bölgələri, c-fos, qövs və ya zif268 kimi dərhal erkən genlərin (IEG) ifadəsi ilə ölçülən kimi, insanlarda aktivləşdirilənlər kimi siçovul beyində xoş ovqat ilə aktivləşdirilir. Həqiqətən, ləzzətli yemək dorsal və ventral striatumu, VTA, yanal hipotalamusu (LH) və siçovullarda amigdala və mükafatla əlaqəli kortikal quruluşların mərkəzi və bazolateral nüvələrini aktivləşdirir ([Angeles-Castellanos et al., 2007], [Park və Carr, 1998] və [Schiltz et al., 2007]). Maraqlıdır ki, Fos immunoreaktivliyi, ləzzətli qida istehlakından (LHb) (Park and Carr, 1998) sonra siçovullarda yanal və medial habenulada həqiqətən azalmışdır. Qeyri-insani primatlarda LHb aversiv stimullaşdırma və ya gözlənilən mükafatların verilməməsi ilə aktivləşdirilir və ləzzətli bir şirəli mükafat (Matsumoto və Hikosaka, 2007) təqdim edilməsinə mane olur. Bundan əlavə, LHb fəaliyyəti, mükafatla əlaqəli mesoaccumbens dopamin ehtiva edən neyronları, rostromedial teimental nüvəni (RMTg) (Jhou et al., 2009) əhatə edən dolayı bir yol ilə maneə törədir. Buna görə habenular fəaliyyət, yemək hedonikası ilə tərs olaraq əlaqələndirilir, bu da habenular kompleksin nonhomeostatik yeməyə təsir göstərə biləcəyini göstərir. Həqiqətən, LHb'nin aktivləşdirilməsi bu yaxınlarda siçovullarda saxaroza istehlakının azaldığı göstərildi, halbuki LHb-nin lezyonları saxarozanın davranışını artırır (Friedman et al., 2011). Xabenular kompleksinin kiçik və çətin olduğunu və insanlarda funksional görüntünü (Salas et al., 2010) nəzərə alsaq, bu, yemək yeməyinə cavab olaraq insan görüntüləmə işlərində xaban fəaliyyətindəki dəyişikliklərin niyə bildirilmədiyini izah edə bilər.
Hedonik yeməyi tənzimləyən beyin dövranları: Midbrain Dopamin Sistemləri
Mezoakkumbens dopamin yolu insanlarda və laboratoriya heyvanlarında dadlı qidaya və ya iştahaaçan qida ilə əlaqəli işarələrə və leptin, qrelin və bu sistemdəki iştaha təsir göstərən digər tənzimləyicilərə cavab olaraq aktivləşdirilir. Bu, orta beyin dopamin sistemlərinin dadlı qida istehlakında əhəmiyyətli bir rol oynadığını göstərir. Bəlkə də orta beyin dopamin ötürülməsinin insanlarda xoşagələn qida qəbuluna təsir göstərməsinin ən açıq göstəricisi, orta beyində dopamin ehtiva edən neyronların dejenerasiyası olduğu Parkinson xəstəliyi (PD) xəstələrinin təsirlənməyən şəxslərdən daha az qida istehlak etməsi (Nirenberg və Sular, 2006). Üstəlik, PD xəstələrinin dopamin reseptor agonistləri ilə müalicəsi dadlı qidanın kompulsiv şəkildə istifadəsinə səbəb ola bilər ([Dagher and Robbins, 2009] və [Nirenberg and Waters, 2006]). Əslində, dopamin reseptor agonistləri, PD olmayan şəxslərdə də hedonik aşırı yeməyə səbəb ola bilər (Cornelius və digərləri, 2010). Heyvanlarda dadlı saxaroza məhlulları NAc-da dopamin ötürülməsini stimullaşdırır (Hernandez və Hoebel, 1988), bu insan beyin görüntüləmə işlərinə uyğun bir təsirdir (Small et al., 2003). Sürətli taramalı tsiklik voltammetrini istifadə edərək, saxaroza mükafatının çatdırılmasını və ya saxarozanın gözlənilmədən çatdırılmasını NAc-da dopamin ötürülməsini təhrik edən işarələrin göstərildiyi göstərildi ([Roitman və digərləri, 2004] və [Roitman və digərləri, 2008]). Bundan əlavə, zərərli xinin məhlullarının gözlənilmədən çatdırılması əks təsir göstərərək akupbal dopamin ötürülməsini azaldıb (Roitman və digərləri, 2008). Nəhayət, tirozin hidroksilaz (TH) fermentinin təsirsiz hala gətirildiyi və bunların dopamin çatışmazlığına səbəb olduğu siçanlar, hələ də sükroz (və ya sakarin) məhlullarına su ilə müqayisədə nəzərəçarpacaq dərəcədə üstünlük verdiyini göstərir, lakin saxarozanı nəzarət siçanlarından daha az miqdarda istehlak edirlər ( Cannon and Palmiter, 2003). Bu, dopamin çatışmazlığı olan siçanların hələ də sükrozun dadlılığını aşkar edə biləcəyini və bu həlləri sudan üstün tutduğunu, lakin dadlı həllərin istehlakını davam etdirə bilmədiyini göstərir. Bu səbəbdən mezoakkumbens dopamin ötürülməsinin qida tədarükü ilə əlaqəli bəslənmə davranışının motivasiya aspektlərini tənzimlədiyi və digər nörotransmitter sistemlərinin dadlı qida istehlakının hedonik aspektlərini tənzimlədiyi irəli sürülmüşdür.
Hedonik yeməyi tənzimləyən beyin dövranları: Striatohipotalamik sistemlər
NM-ə mikro-opioid reseptor agonistlərinin, məsələn, yem sərbəst buraxılması (yəni nonhomeostatik qidalanma) olan siçovullarda qidalanma davranışını stimullaşdırır (D-Ala2-N-Me-Phe4-gly-ol5) -enkefalin (DAMGO). [Peciña və Berridge, 2005] və [Zhang et al., 1998]). Əksinə, NAc-a yoluxmuş opioid reseptor antaqonistləri, daha az dadlı alternativlərin qəbuluna təsir etmədən üstünlük verilən yemək istehlakını azaldır (Kelley et al., 1996). Bu məlumatlar striatal opioid sistemlərinin ləzzətli yeməyin hedonik xüsusiyyətlərini tənzimlədiyi qənaətinə uyğundur. NAcın qabıq bölgəsi və xüsusən medial qabığın rostrodorsal bölgəsindəki hedonik "isti nöqtələr" ([Peciña və Berridge, 2005] və [Peciña et al., 2006b]) nonhomeostatik qidalanmada xüsusilə vacib rol oynayır. Μ-opioid reseptorunun aktivləşdirilməsi NAc-da orta spiny neyron fəaliyyətinin inhibe edilməsi ilə nəticələndiyindən, NAc qabığının ləzzətli qida istehlakına tonik inhibitor təsir göstərməsi təklif edilmişdir (Kelley et al. 2005). Bu görünüşə uyğun olaraq, inhibe GABAA və ya GABAB reseptorlarının stimullaşdırılması ([Basso və Kelley, 1999] və [Stratford və Kelley, 1997]) və ya NAc qabığındakı həyəcanverici ionotrop glutamat reseptorlarının (Maldonado-Irizarry et al. 1995) blokadası. qida istehlakını artırır. Eynilə, NAc qabığının excitotoksik lezyonu da qida istehlakını artırır və qida mükafatına həssaslığı artırır ([Johnson et al., 1996] və [Maldonado-Irizarry and Kelley, 1995]). Xüsusilə, enerji sıx olan ləzzətli qidaların istehlakı bu manipulyasiyalarla ([Basso və Kelley, 1999], [Kelley et al., 2005] və [Zhang et al., 1998]) üstünlük təşkil edir.
Accumbal siqnalın hedonik qidalanmaya əsas təsirini nəzərə alaraq, Thompson və Swanson (2010), NAcın ləzzətli qida istehlakına təsir göstərə biləcəyi dəqiq anatomik şəbəkələri dəqiq müəyyənləşdirmək üçün bir dövrə izləmə prosedurundan istifadə etdilər. Bu zərif tədqiqatlarda siçovullar, qida istehlakına güclü təsir göstərən NAc qabığının yerlərinə iki anterograd / retrograd tracers (COIN adlanır) inyeksiyasını aldılar və afferent / efferent bağlantılar təyin olundu. NAc-da qidalanma ilə əlaqəli saytların əsasən ön LH və ventral pallidum (VP) (Thompson and Swanson, 2010) inhibitor proqnozlarını genişləndirdiyi göstərildi. VTA ilə sıx birləşən NAc-dan fərqli olaraq, NAc qabığındakı qida ilə əlaqəli hedonic isti nöqtələr, interastrikulyar nüvəyə (IFN), VTA-ya bitişik bir quruluşa qarşılıqlı şəkildə dopaminergik proqnozları geri qarşı tərəfə uzadır. NAc qabığı (Thompson və Swanson, 2010). Bundan əlavə, LHb-ə (Tompson və Swanson, 2010) gələn LH layihələri, habenular kompleksinin qida hedonikalarında rol oynaya biləcəyini göstərir (Friedman et al., 2011).
Yuxarıda göstərilən məlumatlar göstərir ki, LH, NAc'ın ləzzətli qida istehlakına tonik bir inhibe təsirini göstərən saytlardan əhəmiyyətli bir inhibitor girişini alır. LH eyni zamanda OFC, insula və amigdala kimi ləzzətli yemək əldə etmək üçün davranışın təşkili və istiqamətləndirilməsi ilə əlaqəli digər kortikal və limbik beyin sahələri ilə funksional əlaqəyə malikdir. Əhəmiyyətli olan, LH-nin inaktivasiyası, NAc manipulyasiyalarının qida qəbuluna verdiyi stimullaşdırıcı təsirləri ([Maldonado-Irizarry et al., 1] və [Will et al., 1995]) ləğv edir. Bundan əlavə, NAc qabığının inaktivasiyası FH immunoreaktivliyi ([Baldo et al., 2003] və [Stratford və Kelley, 2004) ilə ölçülən nöropeptid hipokretini (orexin kimi də tanınır) sintez edən LH neyronlarının fəaliyyətini artırır. ]). Həqiqətən, μ-opioid reseptor agonist DAMGO-nun NAc qabığına infuziyası hipotalamusdakı hipokretin ehtiva edən neyronları aktivləşdirir (Zheng et al., 1999) və VTA-da hipokretin ötürülməsinin pozulması, daxili NAc DAMGO infuziyalarının təsirli olan qida qəbulunu ləğv edir. (Zheng et al., 2007). Beləliklə, NAc qabığındakı hedonic isti nöqtələr LH neyronlarına və xüsusən hipokretin tərkibli neyronlara (Louis et al., 2007) tonik bir inhibitor təsir göstərir və bununla da ləzzətli yemək istehlakını məhdudlaşdırır. Məsələn, gücləndirilmiş opioid reseptor siqnalının köməyi ilə artan bu "dayandırma siqnalının" pozulması, ləzzətli yeməyin nonomehomatik istehlakına səbəb olan inkişaf etmiş LH fəaliyyətinə səbəb olur (Şəkil 2010).
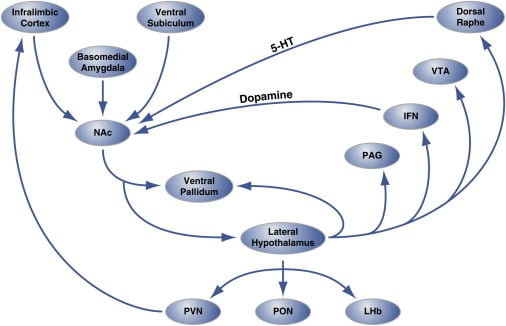
Şəkil 2. Hedonik yeməyi tənzimləyən Nukleus Accumbens Shell-də Hedonik "Qaynar nöqtələrin" dövrə səviyyəli təşkili
Nüvə accumbensinin (NAc) qabıq bölgəsi kortikal və limbik beyin sahələrindən və layihələrdən yan hipotalamusa və ventral palliduma qədər innervasiya alır. Öz növbəsində yanal hipotalamus ventral palliduma və həmçinin PAG, IFN, VTA və dorsal nəcis nüvəsinə təsir göstərir. IFN və dorsal raf, müvafiq olaraq, dopaminergik və serotonergik proqnozları NAc-a qaytarır. Yanal hipotalamus da talamik (PVN və PON) və epitalamik (LHb) quruluşları innervasiya edir. Yanal hipotalamusdan septal beyin sahələrinə qədər kiçik proqnozlar göstərilmir. 5-HT, serotonin; IFN, interfasikulyar nüvə; LHb, yanal habenula; PON, preoptik nüvə; PVN, talamusun paraventrikulyar nüvəsi; VTA, ventral tegramal bölgə. Şəkil Thompson və Swanson (2010) icazəsi ilə uyğunlaşdırılmışdır.
Hedonik yeməyi tənzimləyən beyin dövranları: Striatopallidal sistemlər
LH-dən əlavə, NAc qabıq neyronları da VP-yə istiqamət verir (Şəkil 2). Maraqlı bir təcrübə seriyasında, VP və LH'nin uyğunlaşma proqnozlarının qeyri -omeostatik yeməyin ayrılan aspektlərini (Smith and Berridge, 2007) tənzimləyə biləcəyi göstərildi. Siçovullarda "bəyənmə" reaksiyalarını (yəni ləzzətlilik cavabı) əks etdirmək üçün hipotezasiya edilmiş saxaroza məhlullarına və ya NAc qabığına və ya VP-yə DAMGO infuziyaları artmış qida istehlakı (Smith and Berridge, 2007). Naloksonun ya NAc ya da VP-yə yoluxması, saxaroza reaksiya göstərən üz bənzər reaksiyalarının azalması, NAc və VP-də sinxron opioid ötürülməsinin qida rahatlığı ilə əlaqəli məlumatları emal etmək üçün lazım olduğunu göstərir. Bununla birlikdə, NAc'a NAc daxil edilmiş, lakin VP deyil, qeyri -omeostatik yemək azalmışdır (Smith and Berridge, 2007), qeyri -omeostatik yemək bu NAc → VP bağlantısından asılı olaraq baş verir və bunun əvəzinə NAc → LH yoluna güvənir ([Smith və Berridge, 2007] və [Taha et al., 2009]). Qeyri -omeostatik yeməyin aspektlərinin dağıla biləcəyi anlayışına uyğundur, vahid qeydlər, NAc neyronlarının bir populyasiyasının, qidanın nisbi gücləndirici xüsusiyyətləri (yəni ləzzətlilik) ilə əlaqəli məlumatları seçərək kodlaşdırdığı görünür (Taha və Fields, 2005). . Eyni siçovullarda NAc neyronlarının ikinci bir populyasiyasının fəaliyyətindəki dəyişikliklər qidalanma davranışının başlaması ilə üst-üstə düşdü (Taha and Fields, 2005).
Hedonik yeməyi tənzimləyən beyin dövranları: Amygdalar sistemləri
Qeyri -omeostatik yeməyin aspektlərinin NAc qabığına və ya VP-yə bölünən, nalokson infuziyalarının olması, ancaq bazolateral amigdala (BLA) deyil, saxaroza məhlullarının dadını azaldır (Wassum et al., 2009). Bununla birlikdə, μ-opioid reseptor antagonistləri nalokson və ya CTOP BLA'ya daxil olunduqda, NAc qabığı və ya VP deyilsə, ümumiyyətlə ac vəziyyətdə görülən saxaroza məhlullarına cavab vermək üçün artan motivasiyanın azalması müşahidə edildi. ([Wassum et al ., 2011] və [Wassum et al., 2009]). Bu, saxarozanın təşviqedici xüsusiyyətlərinin amigdalar dövrə ilə tənzimləndiyini göstərir. Ümumiyyətlə, yuxarıdakı tapıntılar, yemli yeməyin fərqli cəhətləri, məsələn, qida rahatlığı ilə əlaqəli məlumatların emalı, yaxınlaşan davranışlar və ac heyvanlarda ləzzətli yeməyin stimullaşdırıcı dəyərinin artması, ayrı bir kontekstdə diskret mikrosxemlər tərəfindən fərqli şəkildə tənzimləndiyini göstərir. daha böyük kortikolimbik-striatopallidal-hipotalamik-talamokortikal dövrə (Şəkil 2).
Beyin Hedonik Dövrlərindəki Uyğunlaşmalar Kompulsif Yeməyə səbəb olurmu?
NAc qabığındakı hedonic isti nöqtələrin funksional aktuallığı və beyindəki daha geniş qidalanma ilə əlaqəli dövrə təsiri Kelley et al. (2005). NAc qabığı → LH yolunun yuxarı və aşağı tənzimlənən beyin bölgələri (Şəkil 2) ilə birlikdə "sentinel" bir məqsədə xidmət etdiyini fərz edirlər (Kelley et al. 2005). Xüsusilə, təklif edirlər ki, hətta ac heyvanlarda yemək sürücüsü güclü olduqda, ətraf mühitdən gələn təhdidlər halında qidalanma davranışını dayandırmaq lazımdır (Kelley et al. 2005). Beləliklə, NAc qabığı neyronlarının aktivləşdirilməsi və LH neyronlarının bir arada inhibe edilməsi davam edən qidalanmanı poza bilər və davranışın donma və ya qaçma kimi daha uyğun uyğunlaşma reaksiyalarına keçməsinə kömək edə bilər (Kelley et al. 2005). Əgər bu həqiqətən belədirsə, onda bu NAc qabığı → LH idarəetmə yolunun ləzzətli yeməyin həddindən artıq istehlakı və ya piylənməyə həssaslığı təsir edən genetik amillər tərəfindən pozulduğunu araşdırmaq vacibdir. Bunu nəzərə alaraq laboratoriyamız və başqalarımız bu yaxınlarda bildirdilər ki, ləzzətli kalorili sıx qidaların həddindən artıq istehlakı siçovullarda kompulsif xarakterli qidalanma davranışının yaranması ilə əlaqədardır ([Johnson and Kenny, 2010], [Latagliata et al., 2010] və [Oswald et al., 2010]). Xüsusilə, obez siçovullarda ləzzətli qida istehlakının mənfi nəticəni (elektrik ayaqları) proqnozlaşdıran tərs şərtli bir stimul tərəfindən pozulmağa davamlı olduğunu gördük (Johnson and Kenny, 2010). Beləliklə, həddindən artıq kilolu və piylənmiş şəxslərin yemək istehlakının normallaşdırılması üçün istehlakçı davranışlarının zərərli nəticələri ilə bağlı məlumatlardan istifadə etmək üçün ən azı qismən həddindən artıq köklənmədən yaranan NAc qabığı → LH nəzarət yolundakı çatışmazlıqların yoxlanılmasının vacibliyini yoxlamaq lazımdır. .
Piylənmə zamanı dəyişdirilən beyin mükafatlandırma fəaliyyəti: İnsan beyin görüntüləmə işləri
Zövqlü yeməklərin beyin mükafatlandırma sistemlərinə stimullaşdırıcı təsirini əldə etmək, həddindən artıq qidalanmaya töhfə verən mühüm həvəsləndirici amildir. Beləliklə, vacib bir sual beyin mükafatlandırma funksiyasındakı dəyişikliklərin piylənmənin inkişafına kömək edə biləcəyidir. İntuitiv bir proqnoz beyin mükafatlandırma sistemlərinin ləzzətli qidaya inkişaf etmiş konstitusional reaksiyasının həddindən artıq dərəcədə artması və kilo verməsi ilə nəticələnməsidir. Bu fərziyyəyə uyğun olaraq, yüksək səviyyəli mükafat həssaslığına sahib olan insanlar, şokolad tortu və pizza kimi ləzzətli yeməklərə məruz qaldıqda, NAc, amigdala, OFC və VP daxil olmaqla, qida mükafatına təsir edən beyin bölgələrində inkişaf etmiş bir fəaliyyət göstərir (Beaver et al.) , 2006). Şişman fərdlər arıq idarələrə ([Gautier et al., 2000], [Karhunen et al., 1997] və [Rothemund et al., 2007) nisbətən xoşagəlməz qida və ya qida ilə əlaqəli istəklərə cavab olaraq beyin mükafatı dövranlarının artan aktivliyini nümayiş etdirir. ]). Təcrübə mükafatlarına həssaslığın yüksək səviyyəsi, insan mövzularında artan bədən çəkisi ilə də əlaqələndirilmişdir ([Davis et al., 2004] və [Franken and Muris, 2005]). Bununla birlikdə, obez qadında (BMI> 30) anhedoniyanın səviyyəsi yüksək idi (yəni mükafat üçün başlanğıc həssaslığı azalmışdır), kilolu qadından (BMI> 25 <30) daha yüksək idi (Davis və ark., 2004). Eynilə, Stice və iş yoldaşları (2008b), obez yetkin qızların, yağsız nəzarət subyektləri ilə müqayisədə xoşagəlməz qida və ya qida ilə əlaqəli istəklərə cavab olaraq insula və digər kortikal beyin bölgələrinin aktivləşməsini artırdıqlarını, ancaq kaudat bölgəsinin aktivləşdiyini göstərdi. ləzzətli qidaya cavab olaraq striatum, obez mövzularda BMI ilə tərs əlaqə oldu. Üstəlik, bir 6 aylıq dövr ərzində kilo verən qadınlar, kilo almamış qadınlarla müqayisədə bu müddət ərzində ləzzətli qidaya cavab olaraq striatal fəaliyyətdə nəzərəçarpacaq dərəcədə azalma müşahidə etdilər (Stice et al., 2010a). Bütün bunları birlikdə götürdükdə, mükafat dövrə həssaslığının bir insanın həddindən artıq kökəlməyə və çəki artmasına meyl göstərə biləcəyi görünür (Stice et al., 2010b). Ancaq kilo artdıqca beyin mükafatlandırma sisteminin müəyyən komponentlərinin, xüsusən striatumun fəaliyyətində çatışmazlıqlar yaranmağa başlaya bilər. Bu mükafat hiposensitivliyinin vəziyyətinin ortaya çıxması, bu cür mükafat çatışmazlıqlarını aradan qaldırmaq üçün ləzzətli yeməklərin həddən artıq istehlak edilməsinə səbəb ola biləcəyi təklif edilmişdir ([Stice et al., 2008a] and [Wang et al., 2002]). Beləliklə, çox az və ya çox miqdarda qida mükafatı, overeating və piylənməyə qarşı həssaslığı artır (Stoeckel, 2010). Bu açıq-aydın bir-birinə zidd olan fikirləri uzlaşdırmaq üçün cəlbedici konseptual çərçivə odur ki, qida mükafatlarını əldə etmək üçün davranışın təşkili və gözlənilən gələcək ərzaq mükafatı barədə proqnozlar verməklə məşğul olan kortikolimbik bölgələr kilolu insanlarda və piylənməyə meylli olanlarda hiperaktiv ola bilər. Əksinə, hedonik yeməkdən həzz almağın həqiqi təcrübəsini emal edən striatal beyin saytları həmin şəxslərdə tədricən az işləyə bilər. Buna görə ləzzətli qidanın nisbi motivasiya dəyərinin, obezliyin inkişafı zamanı eyni zamanda ləzzətli qida istehlakından əldə edilən hedonik dəyərin azalması ilə artacağı gözlənilir.
Piylənmə zamanı dəyişdirilmiş beyin mükafatlandırma fəaliyyəti: gəmirici tədqiqatları
Zövqlü yemək istehlakının beyin mükafatlandırma sistemlərinə təsiri laboratoriya heyvanlarında beyin stimullaşdırılması mükafatı (BSR) prosedurundan istifadə edərək birbaşa qiymətləndirilmişdir. Məlumdur ki, accumbal hedonic isti nöqtələrdən tonik inhibitor girişini alan LH-nin elektrikli stimullaşdırılması (Şəkil 2) yüksək dərəcədədir və siçovullar bu beyin bölgəsini, məsələn (Markou və Frank, 1987) öz-özünə stimullaşdırmaq üçün çox çalışacaqdır. ). Öz-özünə stimullaşdırma davranışına əlavə olaraq, LH-nin elektrikli stimullaşdırılması da qidalanma davranışının (Margules and Olds, 1962) güclü sıxışmalarına səbəb ola bilər və LH stimullaşdırmasının mükafatlandırıcı xüsusiyyətlərinin bunun daxili rolu ilə əlaqəli ola biləcəyi təklif edilmişdir. Yeməyin iştahaaçan və təşviqedici xüsusiyyətlərində beyin saytı (Margules and Olds, 1962). Bu görüşə uyğun olaraq, aclıq və kilo itkisi siçovulların mükafatlandırıcı dəyəri LH-nin özünü stimullaşdırmasına ([Blundell və Herberg, 1968], [Carr və Simon, 1984] və [Margules and Olds, 1962]) həssaslığı artırır leptinin (Fulton et al., 2000) intracerebroventrikulyar infuziyası ilə tıxanacaq. Əksinə, LH-nin elektrikli özünü stimullaşdırması doymuş heyvanlarda (Wilkinson və Peele, 1962) inhibe olunur. Həqiqətən, siçovulların intraqastral qidalanma borusu (Hoebel və Teitelbaum, 1962), həddindən artıq miqdarda bağırsaq damarlığı və ya venadaxili qlükaqon infuziyası postprandial doymuşluğu təqlid edən ([Hoebel, 1969], [Hoebel və Balagura, 1967] və [Mount and Hoebel, 1967]) ), hamısı LH stimullaşdırılmasına cavab verir. Əslində, LH stimullaşdırılması üçün əvvəlcədən güclü cavab verən siçovullar, qida qəbulundan və ya piylənmənin inkişafından sonra (Hoebel və Thompson, 1969) bu stimullaşdırma pis olardı. Beləliklə, xroniki qida məhdudlaşdırılması və kilo itkisi artır, həddindən artıq qidalanma azalsa, LH-də mükafatla əlaqəli saytların həssaslığı artır. LH neyronlarının mükafatlandırıcı elektrikli öz-özünə stimullaşdırılmasına həssaslığı, qida hedonik reaksiyalarını tənzimləyən beyin dövranının işləməsi ilə bağlı vacib məlumatlar verə bilər.
Enerji sıx olan dadlı qidaya daxil olmaq asanlığı və nəticədə həddən artıq istehlak piylənməyə səbəb olan əsas ekoloji amil hesab olunur (Volkow və Wise, 2005), laboratoriyamız bu yaxınlarda ləzzətli yeməklərə geniş çıxışı olan siçovullarda beyin mükafatı fəaliyyətini qiymətləndirmək üçün BSR prosedurundan istifadə etdi. . Xüsusilə, 18-23 saatlıq xoş bir pəhrizə gündəlik girişi ilə birlikdə qidalanma çowuna və ya çowa adit hüququ verən siçovullarda LH-nin elektrik stimullaşdırılmasına cavab verdik. Bu pəhriz pendir, donuz, kolbasa və digər iştahaaçan yemək maddələrindən ibarət idi (Johnson and Kenny, 2010). Ləzzətli qidaya geniş çıxışı olan siçovulların sürətlə əhəmiyyətli miqdarda çəki qazandığını və tədricən pisləşən beyin mükafat çatışmazlığını (LH stimullaşdırılmasına azaldılmış cavab kimi əks olundu) göstərdik (Johnson və Kenny, 2010; Şəkil 3). Bu, diyetə bağlı piylənmənin inkişafının LH-də mükafat saytlarının reaksiyasının tədricən azalması ilə əlaqəli olduğunu göstərir (Johnson and Kenny, 2010). Mükafat siqnalında çatışmazlıqlar, əvvəllər yetkinlik dövründə saxaroza və ya yüksək yağlı qidaya məhdudiyyətsiz çıxışı olan yetkin siçovullarda da bildirilmişdir ([Teegarden et al., 2009], [Vendruscolo et al., 2010a] və [Vendruscolo et al.,) 2010b]). Siçovullardakı bu təsirlər, bir 6 aylıq dövr ərzində kilo aldıqları üçün insanlarda yuxarıda təsvir edilən qida mükafatına cavab olaraq azalmış striatal aktivləşməsini xatırladır (Stice et al., 2010a; Şəkil 4). Aşırı kilolu siçovullarda və bəlkə də kilo alan insanlarda bu cür pəhriz səbəbli mükafat çatışmazlığı, qida mükafatları dövrlərindəki ləzzətli qida (Johnson və Kenny, 2010) tərəfindən həddindən artıq dərəcədə artmasına qarşı olan əks reaksiya əksini tapır. Bu tapıntının vacib tərəfi odur ki, mükafat funksiyasındakı oxşar çatışmazlıqlar kokain və ya eroin həddindən artıq istehlak edən siçovullarda da aşkar olunur ([Əhməd et al. 2002], [Kenny et al., 2006] və [Markou and Koob, 1991]; Şəkil 3). Əslində, dərmanla əlaqəli mükafat disfonksiyonunun azalmış mükafatın davamlı vəziyyətini yüngülləşdirmək üçün dərmanı istehlak etmək üçün yeni bir motivasiya mənbəyi təmin etməklə, idarədən narkotikə keçilməyə kömək edə biləcəyi ehtimal edilir ([Əhməd və Koob) , 2005] və [Koob və Le Moal, 2008]). Buna görə, həddindən artıq kökəlmə ilə nəticələnən LH-də mükafat sahələrinin həssaslığının çatışmazlıqların, həddindən artıq kilolu siçovullarda ləzzətli qida istehlakının uzun müddətli davamlılığını, mənfi vəziyyəti azaltmaq üçün yüksək hedonik təsirə malik qidalara üstünlük verərək artıra bilər. mükafat.
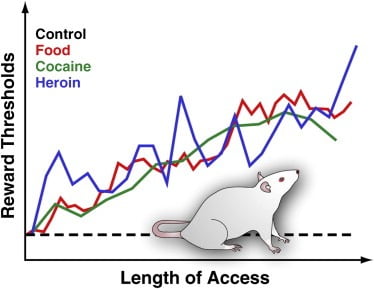
Şəkil 3. Palatable qida, kokain və ya heroin əldə etmək üçün genişləndirilmiş gündəlik çıxışı olan siçovullarda mükafat həddləri
Mükafat həddlərini ölçmək üçün, stimullaşdırıcı bir elektrod, elektrik stimullaşdırmasının güclü şəkildə təltif olunduğu və qidalanma davranışının gərgin böyüməsinə səbəb ola biləcəyi bir bölgəyə, siçovulların yan hipotalamusuna cərrahi yolla yerləşdirilir. Bərpa olunduqdan sonra heyvanlara təkər çevirərək bu bölgəni öz-özünə stimullaşdırmağa icazə verilir. Sabit özünü stimullaşdıran davranış qurulandan sonra, özünü stimullaşdıran davranışı qoruyan minimum stimullaşdırma intensivliyi təyin olunur (yəni mükafat həddi). Bu mükafat həddi mükafat sisteminin fəaliyyət səviyyəsini təmin edir. Mükafat hədləri standart laboratoriya çuğuna girən və dərman sadəlövh qalmış nəzarət siçovullarında sabit və dəyişməz olaraq qalır. Bununla birlikdə, siçovullarda dadlı qida maddələrindən (məsələn, cheesecake, pastırma, şokolad və s.) İbarət olan enerji sıx bir bəslənmə pəhrizinə gündəlik genişlənmə imkanı olan siçovullar tədricən artır. Eynilə, venadaxili kokain və ya eroin infuziyalarına gündəlik girişi genişləndirən siçovullarda mükafat hədləri tədricən yüksəlir. Yüksək mükafat həddi beyin mükafat sisteminin azalmış həssaslığını əks etdirmək üçün şərh olunur. Bu təsirlər xoşagəlməz qidaların həddindən artıq istehlakı və kilolu artımın asılılıq dərmanı həddindən artıq istehlak etməsi ilə əlaqəli beyin mükafatındakı dərin çatışmazlıqlara səbəb ola biləcəyini göstərir.
Şəkil 4. Piylənmədə Striatal Plastisite kilo fMRI ilə ölçülən ləzzətli yeməyə cavab olaraq striatal aktivliyin azalması və insanlarda striatal dopamin D2 reseptorunun (D2R) daha aşağı olması ilə əlaqələndirilir (ətraflı məlumat üçün mətnə baxın).
Piylənmədə Dopamin D2 Reseptor Siqnalı
Son bir neçə hesabat, piylənmənin inkişafı zamanı ləzzətli yeməklərin həddindən artıq istehlakına cavab olaraq mükafat çatışmazlığının yarana biləcəyi potensial mexanizmləri aşkar etmişdir. Yuxarıda qeyd edildiyi kimi, bir 6 aylıq dövrdə kilo verən qadınlar, kilo almamış qadınlarla müqayisədə bu müddət ərzində ləzzətli qidaya cavab olaraq striatal aktivliyin nəzərə çarpacaq dərəcədə azalmasını göstərdilər (Stice et al., 2010a; Şəkil 4). Sevdikləri yeməyi doyma halına gətirməyə icazə verən oruclu insanlar, striatumdakı Dopamin D2 reseptorunun (D2R) antaqonist raklopridinin (Kiçik et al. 2003) bağlanmasının daha az səviyyədə olduğunu, D2R siqnalının ləzzətli yemək istehlakına cavab olaraq azaldığını söylədi. Həqiqətən, obez fərdlər arıqlama ilə müqayisədə daha az striatal D2R mövcudluğuna malikdirlər ([Barnard et al., 2009], [Stice et al., 2008a] and [Wang et al., 2001]; Şəkil 4), arıqlamaq isə obez xəstələrdə striatal D2R sıxlığının artması ilə əlaqələndirilir (Wang et al., 2008). Striatal dopamin ötürülməsinin hedonik yeməyin tənzimlənməsində əsas rol oynadığını nəzərə alsaq, D2R siqnalındakı adaptiv azalmalar, şişman şəxslərdə striatumun dadlı qidaya reaksiyasının azalmasına kömək edə bilər. Bu fürsəti sınamaq üçün, Kiçik və iş yoldaşları, nəzarət fərdlərində və Taqia A1 allelini daşıyanlarda (Felsted et al. 2010) xoşagəlməz bir süd südü yeməsinə cavab olaraq beyin mükafatlandırma sistemlərindəki fəaliyyətini araşdırdılar. Taqia məhdudlaşdırıcı fraqment uzunluğu polimorfizmi D2R genindən (Neville et al., 2004) aşağıda və polimorfizmin A1 allelini daşıyan şəxslər, allel daşımayanlarla müqayisədə 30% - 40% daha az striatal D2R var. et al., 1999], [Ritchie and Noble, 2003] və [Stice et al., 2010b]). Bundan əlavə, A1 allele daşıyıcıları da qida hedonik reaksiyalarında iştirak edən striatal və kortikal beyin bölgələrində qlükoza mübadiləsini azaltmışdır (Jönsson et al., 1999). Taqia A1 allelini saxlayan şəxslər, obez populyasiyalarda həddindən artıq çoxdur ([Barnard et al., 2009], [Stice et al., 2008a] və [Wang et al., 2001]). Bundan əlavə, A1 alleli alkoqol, opioid və psixomotor stimullaşdırıcı asılılığa qarşı həssaslığı artırır ([Lawford et al., 2000], [Noble et al., 1993] and [Noble et al., 2000]). Striatuma dopaminerjik giriş təmin edən VTA və SN daxil olmaqla orta beyin sahələrinin, nəzarət edilən şəxslərdə xoş bir süd südü yeməsinə cavab olaraq aktivləşdirildiyi aşkar edildi (Felsted et al., 2010). Əksinə, bu beyin saytlarında fəaliyyət A1 allele daşıyıcılarında (Felsted et al. 2010) qida mükafatına cavab olaraq azaldılmışdır. A1 allel daşıyıcıları və qeyri-maneələr arasında beyin aktivləşdirilməsində oxşar tərs cavablar talamik və kortikal beyin sahələrində də aşkar edilmişdir (Felsted et al., 2010). Bu məlumatlar D2R-lərin ləzzətli qidaya mezostriatal cavab reaksiyasını tənzimləyən əsas roluna çox uyğundur. Stice və həmkarları (2008a) insan xəstələrində ləzzətli şokoladlı süd yeməyinə cavab olaraq BMI və striatumun (kaudat və putamen) aktivləşdirilməsi arasında tərs bir əlaqə tapdılar. Üstəlik, bu tərs əlaqə, Taqia A1 allelini daşıyan şəxslərdə daha aydın idi (Stice et al., 2008a). İlkin beyin görüntüləməsindən sonra 1 il sonra ölçülən bu şəxslərdə gələcək çəki artımı, ləzzətli qidaya cavab olaraq striatal aktivliyin böyüklüyünün A1 alleli olan subyektlərdə çəki artımı ilə mənfi əlaqəli olduğunu və A1 olmayan allele subyektlərində müsbət nisbətdə olduğunu göstərdi ( Stice et al., 2008a). Sonrakı araşdırmada bildirildi ki, ləzzətli yeməyin təsəvvür edilən yeməsinə cavab olaraq striatal aktivliyin həcmi, faktiki istehlakından fərqli olaraq A1 alleli ilə subyektlərdə növbəti il ərzində çəki artımı ilə tərs əlaqə oldu, lakin müsbət A1 olmayan allele mövzularında əlaqələndirilmişdir (Stice et al., 2010b). Bu tapıntılar, D2R'ların ləzzətli qidaya striatal cavab verdiyini tənzimlədiyini və çəki artımı və ya genetik amillər səbəb olduğu D2R siqnalının piylənməyə həssaslığını artıracağını göstərir.
Qüsurlu D2R Siqnal Obeziteye çatışmazlıqların mükafatlandırılmasına kömək edir
Şişman insan subyektlərindəki aşağı tənzimlənmiş striatal D2R səviyyələrinə bənzər şəkildə D2R səviyyəsi siçanlarda və siçovullarda aşağı bir pəhriz bəslənir (məsələn, [Colantuoni et al., 2001], [Geiger et al., 2009] və [Johnson and Kenny) , 2010]) və genetik olaraq piylənməyə meylli olan siçovullarda (Zucker siçovulları) (Thanos et al., 2008). Laboratoriyamız ümumiyyətlə pozulmuş striatal dopamin ötürülməsinin rolunu birbaşa araşdırdı və xüsusilə D2R siqnalında, piylənmənin inkişafı zamanı siçovullarda meydana gələn bağımlılığa bənzər mükafat çatışmazlıqlarında azaldı (bax 5 Şəkil). Xüsusilə, viral vasitəçi RNT müdaxiləsindən istifadə edərək siçovullarda striatal D2R-lərin azaldılmasının təsirlərini sınaqdan keçirdik, sonra siçovulların yalnız 18-23 saatlıq ləzzətli yüksək enerjiyə gündəlik giriş imkanı ilə BRO hədlərini qiymətləndirdik. pəhriz (yəni bir kafeterya pəhrizi) (Johnson and Kenny, 2010). DHNUMXR sıçan siçovullarda (Johnson and Kenny, 2; Şəkil 2010) LH stimullaşdırılmasına cavab vermə demək olar ki, dərhal kafeterya pəhrizinə məruz qaldıqdan sonra azalmağa başladığını gördük. Buna görə striatal D2R səviyyəsinin azalması yüksək dadlı qidaya geniş çıxışı olan siçovullarda mükafat hipofonksiyonunun meydana gəlməsini sürətləndirir, adətən ləzzətli pəhriz əldə etmək imkanı olan siçovullarda bir çox həftə davam edən bir prosesdir. Bununla birlikdə, siçovullarda striatal D2R-lərin yıxılması yalnız LH stimullaşdırılması üçün cavab vermə qabiliyyətini dəyişdirmədi, bu da azalmış striatal D2R siqnalının beyin mükafat dövranlarında digər pəhriz səbəbli uyğunlaşma reaksiyaları ilə qarşılıqlı təsir göstərməsini təklif etdi. D2R səviyyəsinin aşağı salınması ilə yanaşı, obez siçovulların beynində striatal dopaminergik ötürülmənin digər cəhətləri də dəyişdirilmişdir. Məsələn, Sprague-Dawley siçovulları yüksək enerjili bir pəhrizdə (obezliyə meylli siçovullar) sürətlə kilo vermək üçün selektivliyi artır, çəki artımına (piylənməyə davamlı siçovullara) davamlı olan siçovullara nisbətən NAc-da dopamin səviyyəsini aşağı saldılar (Geiger et al., 2008; həmçinin bax: Rada et al., 2010). Piylənməyə meylli siçovulların da dopamin biosintetik və saxlama maşınlarında azalma səviyyəsi var, bu da dopamin istehsalında və buraxılmağındakı bir arızanın obez siçovullarda striatum dopamin ötürülməsində kəsirlərin artmasına səbəb olduğunu göstərir (Geiger et al. 2008). Ləzzətli yüksək enerjili bir pəhrizin həddindən artıq istehlakı ilə piylənməni inkişaf etdirən siçovullar, NAc-da yalnız standart çow ([Davis et al. 2008] və [Geiger et al., 2009]). Əhəmiyyətli olan, standart sıyıq yeməyi, siçovulların NAc-da dopamin səviyyəsini artırmaq üçün kifayət idi, halbuki yalnız yüksək dərəcədə ləzzətli qida maddələri, ləzzətli qidaya uzun müddət girmə tarixi olan obez siçovullarda accumbal dopamin sərbəst buraxılması üçün kifayət idi. (Geiger et al., 2009). Bu tapıntılar siçovullarda piylənmənin inkişafının ən çox striatal D2R-də mesostriatal dopamin ötürülməsindəki disfunksiya ilə əlaqəli olduğunu və çatışmaz D2R siqnalının siçovullarda piylənmənin inkişafı zamanı mükafat defisitinin yaranmasına səbəb olduğunu göstərir. Bu, striatal D2R-lərin tənzimlənməsinin insanlarda çəki artımına diqqətəlayiq bir nörodaaptiv reaksiya olması ilə uyğundur ([Barnard et al., 2009], [Stice et al., 2008a] və [Wang et al., 2001]), və çatışmayan striatal D2R siqnal insan insanlarında hedonik qidaya qarşı striatal reaksiyaları kəskinləşdirə bilər və bununla da fərdin gələcək çəki artımına meyl edə bilər ([Stice et al., 2008a] və [Wang et al., 2001]).
Şəkil 5. D2 Dopamin Reseptorları, Mükafat Disfunksiyası və Piylənmədə Kompulsivlik - Siçovul striatumunda dopamin D2 reseptorlarının (D2R) yıxılması, ləzzətli yeməyə çıxışı olan siçovullarda mükafat disfunksiyasının və kompulsiv yemənin yaranmasını sürətləndirir..
Qeyri-kafi D2R Siqnal, Obezitədə kompulsif yemək yeyə bilər
Piylənmə istehlakı məhdudlaşdırmaq istəməsinə və davamlı həddindən artıq istehlakın mənfi sağlamlığı və sosial nəticələri barədə biliklərinə baxmayaraq davam edir ([Booth et al., 2008], [Delin et al., 1997] və [Puhl et əl., 2008]). Bu, bir çox obez xəstənin ağırlığını idarə etmək üçün potensial təhlükəli bariatrik (mədə bypass) əməliyyatı keçirməsi (Yurcisin et al., 2009), əməliyyatın aclığın subyektiv reytinqini azaltmasına və azaltmasına baxmayaraq tez-tez ifraz olunmasına reaksiya verir. çox miqdarda yemək istehlak etmək qabiliyyəti ([Kalarchian et al., 2002] və [Saunders, 2001]). Narkotik asılılığı, potensial dağıdıcı sağlamlıq, sosial və ya maliyyə nəticələrini bildiyinə baxmayaraq narkotik istehlakı və vərdişdə inhibe nəzarəti itkisi kimi müəyyən edilir (DSM-IV; Amerika Psixiatrik Assosiasiyası, 1994). Beləliklə, piylənmə və narkotik asılılığı kompulsif pozğunluqların əlamətlərini bölüşür, belə ki, daha az zərərli alternativ davranışların olmasına baxmayaraq istehlakda mülayim istehlak və istehlakda davamlı olmaq üçün gələcək zərərli nəticələr barədə məlumatdan istifadə edilmir.
Gəmiricilərdə kompulsif dərman qəbul etmək cəzanı proqnozlaşdıran ([Pelloux et al., 2007] və [Vanderschuren and Everitt, 2004]) cəza və ya ətraf mühit stimullaşdırılması ilə bastırmayan istehlak olaraq təyin edilmişdir. Kokain və digər sui-istifadə dərmanlarına uzun müddətli girmə dövrləri siçovullarda məcburi narkotik qəbul edən davranışların yaranmasına səbəb ola bilər ([Əhməd və Koob, 1998], [Deroche-Gamonet et al., 2004] və [Vanderschuren and Everitt, 2004] ). Həqiqətən, mənfi nəticəni proqnozlaşdıran (aversiv ayaq barmaqlığının verilməsini proqnozlaşdıran bir replika işıq) ([Belin et al., 2008]), mənfi nəticəni (proqnozlaşdıran) təsirli bir şərtləndirici stimullaşdırma (CS) tərəfindən pozulmağa davamlı olan geniş kokain istehlakı qəbulu tarixi olan siçovullar. və [Vanderschuren and Everitt, 2004]). Əksinə, eyni tərs CS, dərmana nisbətən məhdud girişi olan siçovullarda dərman axtaran cavabları dərin dərəcədə azalda bilər. Narkomaniyada məcburi narkotik istifadəsi və piylənmə halında həddindən artıq yemək arasındakı oxşarlıqları nəzərə alaraq, bir müddət əvvəl obez siçovulların ləzzətli qidaları kompulsif bəyənilən şəkildə istehlak edib-etməyəcəyini və bu müddətdə striatal D2R-lərin rol oynadığını araşdırdıq (Johnson and Kenny, 2010). Dadlı qidaya uzun müddət girmə tarixi olan piylənmiş siçovulların, mülayim bir ayaq baqajının (Johnson and Kenny, 2010) çatdırılacağını proqnozlaşdıran təhlükəli bir CS-nin (yüngül rep) olması halında da dadlı yemək yeməyə davam etdiklərini gördük. Bunun əksinə olaraq, eyni eskirici CS, enerji sıxlığı olan yeməklərə çox məhdud məruz qalmış yağsız siçovullarda ləzzətli yemək istehlakını pozdu. Buna görə çiyələkli qida istehlakı kokain istehlakının məcburi hala gələ biləcəyi qədər obez siçovullarda da məcburi hala gələ bilər. Məlumatların bu təfsirinə uyğun olaraq, əvvəllər ləzzətli yüksək yağlı pəhriz əldə edən siçanlar, pəhrizdə əvvəllər təcrübəsi olmayan siçanlara (Teegarden və Bale) nisbətən xoş bir qida əldə etmək üçün qarışıq bir mühitdə (işıqlı) daha çox vaxt sərf etdi. , 2007). Yırtıcılıq qorxusundan, açıq işıqlandırılmış açıq arenalar siçanlara qarşı çox mükəmməldir (Suarez və Gallup, 1981). Buna görə siçanlar yemləmə davranışlarının potensial mənfi nəticələrinə qarşı dayanıqlı olurlar və daha az təhlükəli yemək mövcud olsa belə ləzzətli yemək əldə etmək üçün yırtılma riskini alacaqlar.
Maraqlıdır ki, Taqia polimorfizminin A1 alleli, striatal D2R sıxlığının (Noble, 2000) azalması və ləzzətli yeməyə cavab olaraq kəskin striatal aktivləşmə (Stice et al., 2008a) ilə əlaqəli olan hərəkətlərin qarşısını almağı öyrənməkdə də çatışmazlıqlar ilə əlaqələndirilir. mənfi nəticələr (Klein et al., 2007). Şişman şəxslərdə məcburi qidalanmanın inkişafına səbəb ola biləcək həddindən artıq yeməyin mənfi nəticələri ilə bağlı məlumatdan istifadə etməməyiniz məhz bu tipdir. Dadlı qidaya çıxışı olan siçovullarda məcburi bənzər yeməyin ortaya çıxması striatal D2R yıxılmasından sonra kəskin surətdə sürətləndiyini gördük (Johnson and Kenny, 2010). Əslində, əvvəllər yalnız 2 günlük enerji sıxlığı olan qidaya geniş çıxışı olan striatal D14R yıxılması olan siçovullar mülayim CS (Johnson and Kenny, 2010; Şəkil 5) tərəfindən kəsilməyə qarşı dayanıqlı qida istehlakını nümayiş etdirdilər. Bununla birlikdə, bu xNUMX gün ərzində ləzzətli qidaya məhdud məruz qalma dövrü, kontrol siçovullarda (Johnson and Kenny, 14) kompulsif kimi yeməyə təkan vermək üçün kifayət deyildi. Bu tapıntılar, şişman siçovullarda bağımlılığa bənzər bir məcburi suqəbuledici meydana gələ biləcəyini göstərir. Bundan əlavə, ləzzətli yemək həddindən artıq istehlakına cavab olaraq mükafat həssaslığının ortaya çıxmasını sürətləndirən defisitli striatal D2010R siqnalı, kompulsiv bəslənmənin meydana gəlməsini də sürətləndirir (Şəkil 2).
D2Rs və Piylənmədə Enerji Balansının Hormonal Tənzimləyiciləri arasındakı qarşılıqlı əlaqə
VTA-ya daxil edilmiş ekzogen leptin, mesoaccumbens dopamin ötürülməsini və qidalanma davranışını ([Hommel et al., 2006] və [Krügel et al., 2003]) inhibə edir. Orta beyin dopamin sistemlərinə kəskin inhibe təsirindən əlavə, tonik leptin siqnalının mezostriatal dopamin siqnalının müvafiq səviyyəsini qorumaq üçün də lazım ola biləcəyinə dair sübutlar mövcuddur. Flier və həmkarları, ob / ob siçanlarında dopamin biosintezində əsas ferment olan orta beyin dopamin neyronlarında tirozin hidroksilazasının daha aşağı səviyyədə olduğunu (Fulton et al., 2006) tapdılar. Bundan əlavə, ob / ob siçanları NAc-a (Fulton et al., 2006) daxil olan dopamin buraxmasını azaltmış və VTA və SN-də dopamin somatodendritik vezikulyar mağazalarında azalmışdır (Roseberry et al., 2007). Ob / ob siçanlarında dopamin istehsalında və siqnal ötürücü maşınlarda olan bu çatışmazlıqlar leptin ([Fulton et al., 2006] və [Pfaffly et al., 2010]) ilə müalicə yolu ilə düzəldildi. Əslində, yalnız LH-yə yoluxmuş leptin, dopamin siqnalının müvafiq səviyyələrini qorumaq üçün LH aktındakı leptin reseptorlarını ifadə edən hüceyrələri təklif edərək, ob / ob siçanlarında (Leinninger et al. 2009) disfunksiyalı dopamin ötürülməsini düzəltmək üçün kifayət idi. Dopamin istehsalının azalması və sərbəst buraxılması ilə yanaşı, ob / ob siçanları da striatumda D2R ifadəsinin daha aşağı səviyyədə olmasına meyl göstərdilər (Pfaffly et al., 2010). Bundan əlavə, vəhşi tipli siçanlarda leptin siqnalına həssaslığın (yəni leptin müqavimətinin) inkişafı ilə nəticələnən bir rejim altında ekzogen leptinin müalicəsi (Pfaffly et al. 2). Piylənmiş siçovullar VTA-da leptin müqavimətini inkişaf etdirir ([Matheny et al., 2010] və [Scarpace et al., 2011]), həmçinin VTA-da TH-nin daha aşağı səviyyəsi, striatumda dopamin sökülməsi və striatal D2010R səviyyələrinin azalması (Geiger) et al., 2). Birlikdə götürülən bu məlumatlar leptinin midbrain dopamin sistemlərində kompleks hərəkətlərə sahib olduğunu göstərir. Bir tərəfdən, VTA-da leptin reseptorlarının kəskin aktivləşməsi mesoaccumbens dopamin ötürülməsinə inhibitor təsir göstərir və qidalanma davranışını maneə törədə bilər ([Hommel et al., 2008] və [Krügel et al., 2006]). Digər tərəfdən, orta beyindəki leptin siqnalının, müvafiq dopamin istehsalını və siqnal ötürülməsini təmin etmək lazımdır və leptin siqnalında genetik çatışmazlıqlar və ya piylənmədə leptin müqavimətinin inkişafı mesoaccumbens dopamin sistemlərini ciddi şəkildə pozur. Odur ki, piylənmənin inkişafı zamanı midbrain dopaminergik neyronlarda leptin müqavimətinin, asılılıq kimi mükafat disfonksiyonunun və kompulsif həddən artıq köklənmənin ortaya çıxmasına səbəb olan striatal D2003R siqnalının pozulmasında mərkəzi rol oynaya biləcəyi maraqlı bir ehtimaldır. (Şəkil 2) obez siçovullarda.
Leptin siqnalının mesostriatal dopaminergik ötürülməsinə mürəkkəb təsirləri ilə yanaşı, D2R-in də öz növbəsində leptin siqnalını tənzimlədiyinə dair sübutlar mövcuddur. D2R agonist bromokriptinin leptinin səviyyəsini azaldır ([Doknic et al. 2002], [Kok et al., 2006] and [Mastronardi et al. 2001]), bu da D2R-leptin səviyyəsinə inhibe təsir göstərdiyini göstərir. Bundan əlavə, D2R genində null mutasiya olan siçanlar hipotalamusda inkişaf etmiş leptin siqnalına malikdir və leptinin anorektik təsirinə daha həssasdırlar (Kim et al. 2010). Piylənmənin (hiperleptinemiya) inkişafı zamanı leptin səviyyəsinin artdığı məlumdur, lakin leptin siqnalına (yəni leptinə qarşı müqavimət) həssaslığın eyni vaxtda azalması müşahidə olunur (Hamilton et al. 1995). Beləliklə, piylənmənin inkişafı zamanı striatal D2R siqnalında azalma, leptin müqavimətinin inkişafını aradan qaldırmaq üçün dövriyyətsiz leptin səviyyəsini artıran və siqnal səmərəliliyini artıran ləzzətli qidaların həddindən artıq istehlakına və çəki artımına kompensasiyaedici cavab verə biləcəyi maraqlı bir ehtimaldır. Belə bir hərəkət leptinin ləzzətli yeməklərə ([Farooqi et al. 2007], [Fulton et al., 2006] və [Hommel et al., 2006]) reaksiyasını tənzimləyən striatal sistemlərə təsir edən təsirini artıra bilər və bununla da fəaliyyət göstərir. ləzzətli yeməyə hedonik cavab vermək. Bu tapıntını leptinin tənzimləmə rolu ilə yuxarıda təsvir olunan D2R-lərə qoyaraq, görünür ki, leptin və D2R siqnalları qidalanma davranışının homeostatik və hedonik aspektlərini tənzimləmək üçün qarşılıqlı şəkildə birləşdirilə bilər.
Piylənmədə tənzimlənən beyin stres sistemləri
Orta beyində olan leptin siqnalları mesoaccumbens dopamin ötürülməsini və hedonik qidaya reaksiyasını tənzimləyir. Bununla birlikdə, VTA-da leptin reseptorlarını az ifadə edən neyronlar NAc-a az çəkir və bunun əvəzinə amigdala (CeA) mərkəzi nüvəsinə (Leshan et al., 2010) daha görkəmli proqnozlar nümayiş etdirir. Bəslənmə davranışı kontekstində CeA, zərərli ətraf mühit stimullarının qida istehlakına təsirini tənzimlədiyi bilinir (Petrovich et al., 2009). Xüsusilə, CeA'nın zədələnmələri, lakin bazolateral amigdala (BLA), ayaq balığı ilə əlaqəli şərtləndirilmiş stimullaşdırmanın qidalanma üzərində inhibe təsirini ləğv edərək, CeA'nın mənfi nəticəni proqnozlaşdıran ekoloji səbəblərə cavab olaraq qidalanma davranışına inhibe nəzarəti üçün kritik olduğunu göstərir. ( Petrovich et al., 2009). Şişman siçovullar və ya striatal D2R yıxılması və ləzzətli qidaya çıxışı olan siçovullar, mülayim bir CS-nin iştirakı ilə məcburi formada dadlı yemək istehlak etməyə davam edirlər. Bu təsirlər CeA zədələnmiş siçovullarda (Petrovich et al. 2009) aversive CS-də reaktivliyin pozulmasına heyrətamiz dərəcədə bənzəyir. Beləliklə, CeA fəaliyyətindəki dəyişikliklərin, bəlkə də striatal D2R səviyyəsinin aşağı düşməsi və ya orta beyin sahəsindəki leptin müqavimətinin inkişafı nəticəsində obez siçovullarda kompulsif bəslənmənin meydana gəlməsinə səbəb olub olmadığını müəyyən etmək vacibdir.
Zərərli ətraf mühit stimullarının qidalanma davranışına təsirlərini tənzimləməklə yanaşı, CeA, streslə əlaqəli hedonik yeməkdə də əsas rol oynaya bilər. İnsanlardakı stres, kalori ehtiyacından asılı olmayaraq enerjisi sıx dadlı qidanın seçilməsini və istehlakını güclü şəkildə motivasiya edir ([Gluck və digərləri, 2004], [Kandiah və digərləri, 2006] və [O'Connor və digərləri, 2008]). və piylənmə, streslə əlaqəli yüksək qlükokortikoid sekresiyası ilə əlaqələndirilir ([Björntorp and Rosmond, 2000] və [la Fleur, 2006]). Ətraf mühit və sosial streslər, gəmiricilərdə və meymunlarda enerjili sıx dadlı qidanın hiperfajiyasını əmələ gətirir, dadlı qida istehlakı isə stresin mənfi təsirlərini azaldır ([Dallman və digərləri, 2003], [Dallman və digərləri, 2006], [Pecoraro et al.). al., 2004] və [Wilson et al., 2008]). Bundan əlavə, stres yaradan yohimbine dərmanı, əvvəllər sönmüş dadlı qidanı siçovullarda bərpa edə bilər (qol basaraq), bu da kortikotropini sərbəst buraxan faktor-1 (CRF-1) reseptor antagonisti antalarmin (Ghitza et al., 2006) ). Stresin insanlarda və laboratoriya heyvanlarında qida istehlakına dəqiq təsirlərinin, stresin təbiətinə və böyüklüyünə, istehlak üçün istifadə olunan qidanın növünə (dadlı və mülayim), bədən çəkisinə və cinsinə (Dallman) bağlı olduğunu qeyd etmək vacibdir. , 2010).
Ləzzətli yüksək yağlı bir pəhrizdən istifadə etmək imkanı olan siçanlar CeA (Teegarden and Bale, 2007) içərisində CRF stress hormonunun ifadəsini azaldıblar. Əksinə, ləzzətli pəhrizdən "çəkilmə" edilən siçanlar CeA-da CRF ifadəsini artırdı (Teegarden and Bale, 2007). Bu son təsiri, bütün əsas sui-istifadə dərmanlarından (Koob, 2010) çəkilməyə məruz qalan siçovullarda aşkar edilən CeA-da artan CRF ifadəsinə çox oxşardır. Narkotik istifadəsi, narkotikin götürülməsinə qarşı bu mənfi neyrobioloji cavabı normallaşdıra biləcəyi üçün, CeA və digər limbik quruluşlarda hiperaktiv CRF ötürülməsinin kompulsif narkotik istifadəsinin inkişafına kömək edə biləcəyi ehtimal edildi (Koob və Zorrilla, 2010). Bu görüşə uyğun olaraq, CeA-da CRF səviyyəsini yüksəltmiş ləzzətli enerji sıxlığı olan yeməklərdən çəkilən siçanlar, əvvəllər yemək təcrübəsi olmayan siçanlara nisbətən ləzzətli (işıqlı) mühitdə daha çox vaxt sərf etdi. az qonaqpərvər (qaranlıq işıqlı) bir mühitdə mövcud idi (Teegarden and Bale, 2007). Buna görə siçanlar, CeA-da (Teegarden and Bale, 2007) CRF ötürülməsini əldə etmək üçün, daha az təhlükəli qida əldə edilsə belə, ləzzətli yemək əldə etmək üçün yedikli davranışlarının potensial mənfi nəticələrinə qarşı davamlı olurlar. Bir neçə daha çox sübut parçası məcburi yeməkdə CRF ötürülməsinin rolunu dəstəkləyir. Xüsusilə, son bir araşdırmada, CRF-1 reseptor antagonisti R121919'ın həftədə 5 gündə standart çow və gündə xNUMX gün ərzində xoş bir şəkər diyetinə girmə imkanı olan pəhrizlərində tsiklik dəyişikliyə məruz qalan siçovullarda qida istehlakına təsirləri qiymətləndirilmişdir. həftə (Cottone et al., 2). Pəhrizdəki bu tsiklik dəyişikliyin 7 həftəsindən sonra, R121919, olduqca ləzzətli pəhrizin həddindən artıq istehlakını və qarışıq çowun istehlakını artırdı (Cottone et al., 2009). Bundan əlavə, CeA-dakı CRF ifadə səviyyələri, xoşagəlməz pəhrizdən çıxarkən siklli siçovullarda artdı, lakin xoşagəlməz diyetə yenidən məruz qaldıqdan sonra ilkin səviyyələrə qayıtdı (Cottone et al., 2009). Nəhayət, CeA-da GABAergik ötürülmənin CRF tənzimlənməsi, R121919-in pozulmuş GABAergik inhibe post-sinaptik potensialının pozulduğunu əks etdirən, əvvəllər yalnız qarışıq çowa çıxışı olan nəzarət siçovulları ilə müqayisədə, dadlı pəhrizdən çəkilməyə məruz qalan siçovullarda yaxşılaşdırıldı. CeA, siçovulların idarəetmə siçovullarından dilimlərdə ötürülməsini dəyişdirməyən bir konsentrasiyada dilimlər (Cottone et al., 2009). Maraqlıdır ki, CRE-nin CeA-da GABAergic ötürülməsinə bənzər təsiri xroniki etanol ifrazatından çıxarılan siçovullarda da müşahidə edilmişdir (Roberto et al., 2010). Nəhayət, CeA, stria terminalisin yataq nüvəsi (BNST) və NAc qabığı "genişləndirilmiş amigdala" adlandırılan daha böyük bir bitki quruluşu meydana gətirir. Hedonik yeməyi stimullaşdıran yerlərdə CRF'nin NAc qabığına infuziyası ətraf mühitin motivasiya qabiliyyətini artırır Əvvəllər ləzzətli yeməyin mövcudluğu ilə cütləşdirilmiş replika (Peciña et al., 2006a). Buna görə stres, NAc qabığı neyronlarının fəaliyyətini modulyasiya edərək qida ilə əlaqəli ekoloji cəhətlərin təsirini artıra bilər. Birləşdirilən bu tapıntılar, həddindən artıq istehlaklı yemək və ya sui-istifadə dərmanlarının, beyindəki ekstrahipotalamik stres yollarında oxşar nöro-reaksiya reaksiyalarına səbəb ola biləcəyini və məcburi istehlakçı davranışlara səbəb ola biləcəyini göstərir.
Nəticələr
Son zamanlarda dadlı qidanın hedonik təsirləri ilə əlaqəli beyin sistemlərinin və dadlı qidanın həddindən artıq istehlakına və kilo almasına cavab olaraq bu dövrələrdə baş verən uyğunlaşmaların müəyyənləşdirilməsində çox irəliləyiş əldə edilmişdir. Bənzər beyin sistemlərinin və ümumi adaptiv reaksiyaların həm dadlı qida, həm də asılılıq yaradan dərmanların istehlakına cavab olaraq tetiklendiği təəccüblüdür. Xüsusilə ləzzətli qidanın və ya sui-istifadə dərmanlarının həddindən artıq istehlakı beyin mükafatlandırma sistemlərinin reaksiyasında oxşar çatışmazlıqlara səbəb olur. Dadlı qida və asılılıq yaradan dərmanlar striatal dopamin ötürülməsində və striatal D2R-lərin ekspresyonuna səbəb olur. Dadlı qida və asılılıq yaradan dərmanlar gəmiricilərdə kompulsiv bənzər istehlakçı davranışın ortaya çıxmasına səbəb olur və ekstra hipotalamik stres reaksiyalarına səbəb olur. Nəhayət, beyin mükafatlandırma sistemlərindəki ümumi genetik zəifliklər fərdləri həddindən artıq yeməyə və piylənməyə, eyni zamanda narkotik istifadəsinə və bağımlılığına meylli edə bilər. Əslində, piylənmə ilə narkotik asılılığı arasındakı bu və digər oxşarlıqlara əsaslanaraq, piylənmənin yaxınlaşan DSM-V-də diaqnostik bir kateqoriya kimi daxil edilməli olduğu iddia edilmişdir ([Devlin, 2007] və [Volkow və O'Brien, 2007] ). Bunu nəzərə alaraq, gələcək tədqiqatlar üçün kritik sahələr kompulsiv aşırı yemə və dərman istifadəsi arasındakı potensial nörobioloji örtüşmələrin daha da araşdırılmasını əhatə edəcəkdir. Məsələn, piylənmənin dorsal striatumdakı plastisiyadan doğan vərdişə bənzər istehlakçı davranışın inkişafı ilə əlaqəli olub olmadığını, narkotik asılılığının striatal yenidənqurma və vərdişə bənzər narkotik axtarmağın ortaya çıxması ilə əlaqəli olduğunu yoxlamaq vacib olacaq. davranışlar ([Everitt and Robbins, 2005], [Hollander et al., 2010] and [Kasanetz et al., 2010]). Ayrıca, icra nəzarəti və qərar qəbuletmə (yəni prefrontal korteks) və interoseptiv işləmə (insular korteks) ilə əlaqəli kortikal beyin bölgələri narkotik asılılığına çox təsir göstərmişdir ([Everitt və digərləri, 2008], [Fineberg və digərləri, 2010 ], [Koob and Volkow, 2010] və [Naqvi and Bechara, 2009]), kompulsif yemək və obezitedeki dəqiq rolları haqqında hələ çox az məlumat vardır, ([Nair və digərləri, 2011] və [Volkow et al.). , 2009]). Birlikdə, yuxarıda nəzərdən keçirilmiş məlumatlar obezliyin və narkotik asılılığının beyin mükafat dövriyyəsindəki oxşar neyro-adaptiv reaksiyalardan yarana biləcəyini və bilinən asılılıq mexanizmlərinin obezliyi anlamaq üçün heuristik bir çərçivə təmin edə biləcəyini düşünür.
Minnətdarlıq
Müəllif Narkotiklərlə Mübarizə üzrə Milli İnstitutun (NIDA) qrantları ilə dəstəklənir. Müəllif Paul Johnson və Christie Fowler'e əlyazma ilə əlaqədar dəyərli fikirləri və şərhləri üçün təşəkkür edir. Bu Scripps Araşdırma İnstitutundan 21042 əlyazma nömrəsidir.
References
1.
o Abizaid et al., 2006a
o A. Abizaid, Q. Gao, TL Horvath
o Yemək üçün düşüncələr: Beyin mexanizmləri və periferik enerji balansı
o Neuron, 51 (2006), s. 691 - 702
o
2.
o Abizaid et al., 2006b
o A. Abizaid, ZW Liu, ZB Andrews, M. Shanabrough, E. Borok, JD Elsworth, RH Roth, MW Sleeman, MR Picciotto, MH Tschöp et al.
o Ghrelin iştahı təşviq edərkən midbrain dopamin neyronlarının fəaliyyətini və sinaptik giriş təşkilini modulyasiya edir
o J. Clin. İnvest., 116 (2006), s. 3229 - 3239
o
3.
o Əhməd və Koob, 1998
o SH Əhməd, GF Koob
o Orta dərəcədən həddindən artıq dərman qəbuluna keçid: hedonik təyin olunmuş nöqtədə dəyişiklik
o Elm, 282 (1998), s. 298-300
o
4.
o Əhməd və Koob, 2005
o SH Əhməd, GF Koob
Narkomanlığa keçid: mükafat funksiyasının allostatik bir azalmasına əsaslanan mənfi bir möhkəmləndirmə modeli
o Psixofarmakologiya (Berl.), 180 (2005), s. 473-490
o
5.
o Əhməd et al., 2002
o SH Əhməd, PJ Kenny, GF Koob, A. Markou
o kokain istifadəsi ilə əlaqəli hedonik allostaz üçün neyrobioloji dəlil
o Nat. Neurosci., 5 (2002), s. 625-626
o
6.
o Allison et al., 1999
o DB Allison, KR Fontaine, JE Manson, J. Stevens, TB VanItallie
ABŞ-da piylənmə ilə əlaqəli illik ölümlər
o JAMA, 282 (1999), s. 1530 - 1538
o
7.
o Amerika Psixiatrik Assosiasiyası, 1994
o Amerika Psixiatrik Assosiasiyası
Psixi pozğunluqların diaqnostik və statistik məlumat kitabçası
o (Dördüncü Edition) Amerika Psixiatrik Assosiasiyası, Vaşinqton, DC (1994)
o
8.
o Angeles-Castellanos et al., 2007
o M. Angeles-Castellanos, J. Mendoza, C. Escobar
o Siçovullarda kortikolimbik bölgələrdə c-Fos və Per1 zülalının gündəlik ritmlərinin dəyişməsi
o Nevrologiya, 144 (2007), s. 344-355
o
9.
o Baicy et al., 2007
o K. Baicy, ED London, J. Monterosso, ML Wong, T. Delibasi, A. Sharma, J. Licinio
o Leptin əvəz edilməsi, genetik leptin çatışmazlığı olan yetkinlərdə qida istəklərinə beyin cavabını dəyişdirir
o Proc. Natl. Akad. Elm. ABŞ, 104 (2007), s. 18276 - 18279
o
10.
o Baldo et al., 2004
o BA Baldo, L. Gual-Bonilla, K. Sijapati, RA Daniel, CF Landry, AE Kelley
o, yeni bir mühitə məruz qalmamaqla, nüvənin böyüdülmə qabığının GABAA reseptoru vasitəçiliyi ilə inhibe edilməsi ilə orexin / hipokretin tərkibli hipotalamik neyronların subpopulyasiyasının aktivləşdirilməsi
o Eur. J. Neurosci., 19 (2004), s. 376-386
o
11.
o Balleine və Dickinson, 2000
o BW Balleine, A. Dickinson
o İnsulyar korteksin zədələnmələrinin instrumental kondisionerə təsiri: stimullaşdırıcı yaddaşda rolun sübutu
o J. Neurosci., 20 (2000), s. 8954-8964
o
12.
o Barnard et al., 2009
o ND Barnard, EP Noble, T. Ritchie, J. Cohen, DJ Jenkins, G. Turner-McGrievy, L. Gloede, AA Green, H. Ferdowsian
o D2 dopamin reseptoru Taq1A polimorfizmi, bədən çəkisi və 2 diabet şəklində pəhriz qəbulu
o Qidalanma, 25 (2009), s. 58-65
o
13.
o Basso və Kelley, 1999
o AM Basso, AE Kelley
o Gübrə qabığı içərisində GABA (A) reseptor stimullaşdırması ilə qidalanma: regional xəritələşdirmə və makronutrient və dad üstünlüklərinin xarakteristikası
o Behav. Neurosci., 113 (1999), s. 324-336
o
14.
o Batterham et al., 2007
o RL Batterham, DH ffytche, JM Rosenthal, FO Zelaya, GJ Barker, DJ Withers, SC Williams
o Kortikal və hipotalamik beyin bölgələrinin PYY modulyasiyası, insanlarda qidalanma davranışını proqnozlaşdırır
o Təbiət, 450 (2007), s. 106-109
o
15.
o Baxter və Murray, 2002
o MG Baxter, EA Murray
o amigdala və mükafat
o Nat. Rev. Neurosci., 3 (2002), s. 563 – 573
o
16.
o lobya et al., 2008
o MK Bean, K. Stewart, ME Olbrisch
Amerikadakı piylənmə: Klinik və sağlamlıq psixoloqları üçün təsirlər
o J. Clin. Psixol. Med. Parametrlər, 15 (2008), s. 214 - 224
o
17.
o Beaver et al., 2006
o JD Beaver, AD Lawrence, J. van Ditzhuijzen, MH Davis, A. Woods, AJ Calder
Mükafat sürücüsündəki fərdi fərqlər, yemək görünüşlərinə neyron cavablarını proqnozlaşdırır
o J. Neurosci., 26 (2006), s. 5160-5166
o
18.
o Belin et al., 2008
o D. Belin, AC Mar, JW Dalley, TW Robbins, BJ Everitt
o Yüksək dürtüsellik məcburi kokain qəbuluna keçidi proqnozlaşdırır
o Elm, 320 (2008), s. 1352-1355
o
19.
o Berridge, 1996
o KC Berridge
o Qida mükafatı: İstəyən və bəyənilən beyin substratları
o nevroloqlar. Biobehav. Rev., 20 (1996), s. 1 - 25
o
20.
o Berridge, 2009
o KC Berridge
o 'İstəyən' və 'istəyən' qida mükafatları: Beyin substratları və yemək pozğunluqlarında rol
o Physiol. Davranış., 97 (2009), s. 537 - 550
o
21.
o Björntorp və Rosmond, 2000
o P. Björntorp, R. Rosmond
Obezite və kortizol
o Qidalanma, 16 (2000), s. 924-936
o
22.
o Blundell və Herberg, 1968
o JE Blundell, LJ Herberg
qida çatışmazlığı və məhrumiyyət dövrünün yanal hipotalamusun elektrikli özünü stimullaşdırma dərəcəsinə nisbi təsiri
o Təbiət, 219 (1968), s. 627-628
o
23.
o Booth et al., 2008
o ML Booth, RL Wilkenfeld, DL Pagnini, SL Booth, LA King
o Yeniyetmələrin kilolu və piylənmə ilə bağlı təsəvvürləri: rəyin öyrənilməsinin çəkisi
o J. Paediatr. Uşaq sağlamlığı, 44 (2008), s. 248-252
o
24.
o Bragulat et al., 2010
o V. Bragulat, M. Dzemidzic, C. Bruno, CA Cox, T. Talavage, RV Considine, DA Kareken
Aclıq zamanı beyin mükafatlandırma dövrlərinin qida ilə əlaqəli qoxuları: Pilot fMRI Tədqiqatı
Obezite, Gümüş Bahar, MD (2010)
o
25.
o Cabanac və Johnson, 1983
o M. Cabanac, KG Johnson
o siçovullarda ləzzət və soyuq ifşa arasındakı qarşıdurmanın təhlili
o Physiol. Davranış., 31 (1983), s. 249 - 253
o
26.
o Campfield et al., 1995
o LA Campfield, FJ Smith, Y. Guisez, R. Devos, P. Burn
o Rekombinant siçan OB zülalı: yağlılığı və mərkəzi sinir şəbəkələrini birləşdirən periferik bir siqnal üçün dəlil
o Elm, 269 (1995), s. 546-549
o
27.
o Cannon və Palmiter, 2003
o CM Cannon, RD Palmiter
o Dopamin olmadan mükafat
o J. Neurosci., 23 (2003), s. 10827-10831
o
28.
o Carr və Simon, 1984
o KD Carr, EJ Simon
o Aclığın mükafat potensialı opioid vasitəçiliyi ilə həyata keçirilir
o Beyin Res., 297 (1984), s. 369-373
o
29.
o Xəstəliklərə Nəzarət və Qarşısının Alınması Mərkəzləri, 2009
o Xəstəliklərə Nəzarət və Qarşısının Alınması Mərkəzləri (2009). ABŞ-da piylənmə meylləri (Atlanta: Xəstəliklərə Nəzarət və Qarşısının Alınması Mərkəzləri).
o
30.
o Colantuoni et al., 2001
o C. Colantuoni, J. Schwenker, J. McCarthy, P. Rada, B. Ladenheim, JL Cadet, GJ Schwartz, TH Moran, BG Hoebel
o Həddindən artıq şəkər qəbulu, beyindəki dopamin və mu-opioid reseptorları ilə əlaqəli dəyişir
o Neuroreport, 12 (2001), s. 3549-3552
o
31.
o Cornelius et al., 2010
o JR Cornelius, M. Tippmann-Peikert, NL Slocumb, CF Frerichs, MH Silber
Narahat ayaqları sindromunda dopaminergik maddələrin istifadəsi ilə impuls idarəetmə pozğunluqları: Bir davaya nəzarət tədqiqi
o Yuxu, 33 (2010), s. 81 - 87
o
32.
o Cornier et al., 2009
o MA Cornier, AK Salzberg, DC Endly, DH Bessesen, DC Rojas, JR Tregellas
İncə və azaldılmış obez şəxslərdə vizual qida istəklərinə qarşı həddindən artıq bəslənmənin təsirləri
o PLoS ONE, 4 (2009), səh. e6310 http://dx.doi.org/10.1371/journal.pone.0006310
o
33.
o Cottone et al., 2009
o P. Cottone, V. Sabino, M. Roberto, M. Bajo, L. Pockros, JB Frihauf, EM Fekete, L. Steardo, KC Rice, DE Grigoriadis et al.
o CRF sistemi işəgötürmə məcburi yemək qaranlıq tərəfi vasitəçilik edir
o Proc. Natl. Akad. Elm. ABŞ, 106 (2009), s. 20016 - 20020
o
34.
o Dagher, 2009
o A. Dagher
o İştahın neyrobiologiyası: Aclıq asılılıq kimi
o Int. J. Obes. (Lond.), 33 (Suppl 2) (2009), s. S30 - S33
o
35.
o Dagher və Robbins, 2009
o A. Dagher, TW Robbins
o Şəxsiyyət, asılılıq, dopamin: Parkinson xəstəliyinə dair fikirlər
o Neuron, 61 (2009), s. 502 - 510
o
36.
o Dallman, 2010
o MF Dallman
o Stressə səbəb olan obezlik və emosional sinir sistemi
o tendensiyalar Endokrinol. Metab., 21 (2010), s. 159 - 165
o
37.
o Dallman et al., 2003
o MF Dallman, N. Pecoraro, SF Akana, SE La Fleur, F. Gomez, H. Houshyar, ME Bell, S. Bhatnagar, KD Laugero, S. Manalo
Xroniki stress və piylənmə: "Rahatlıq yeməyi" nin yeni bir mənzərəsi
o Proc. Natl. Akad. Elm. ABŞ, 100 (2003), s. 11696 - 11701
o
38.
o Dallman et al., 2006
o MF Dallman, NC Pecoraro, SE La Fleur, JP Warne, AB Ginsberg, SF Akana, KC Laugero, H. Houshyar, AM Strack, S. Bhatnagar, ME Bell
o Qlükokortikoidlər, xroniki stress və piylənmə
o Proq. Beyin Res., 153 (2006), s. 75 - 105
o
39.
o Davis et al., 2004
o C. Devis, S. Strachan, M. Berkson
o Mükafata həssaslıq: Aşırı kilolu olmaq və həddən artıq kilolu olmaq üçün təsir
o İştaha, 42 (2004), s. 131 - 138
o
40.
o Davis et al., 2008
o JF Davis, AL Tracy, JD Schurdak, MH Tschöp, JW Lipton, DJ Clegg, SC Benoit
o Pəhriz yağının yüksək səviyyəsinə məruz qalması siçovuldakı psixostimulyator mükafatı və mesolimbik dopamin dövriyyəsini zəiflədir.
o Behav. Neurosci., 122 (2008), s. 1257-1263
o
41.
o de Araujo et al., 2010
o IE de Araujo, X. Ren, JG Ferreira
beyin dopamin sistemlərində metabolik hissetmə
o Nəticələr Probl. Hüceyrə fərqi., 52 (2010), s. 69 - 86
o
42.
o Delin et al., 1997
o CR Delin, JM Watts, JL Saebel, PG Anderson
Morbid piylənmə üçün mədə bypass əməliyyatından sonra yemək davranışı və aclıq təcrübəsi
o obes. Surg., 7 (1997), s. 405 - 413
o
43.
o Deroche-Gamonet et al., 2004
o V. Deroche-Gamonet, D. Belin, PV Piazza
siçovuldakı asılılığa bənzər davranışa dair dəlil
o Elm, 305 (2004), s. 1014-1017
o
44.
o Devlin, 2007
o MJ Devlin
o DSM-V-də piylənmə üçün bir yer varmı?
o Int. J. Yeyin. Yalnış., 40 (Ehtiyat) (2007), s83-S88
o
45.
o Doknic et al., 2002
o M. Dokniç, S. Pekic, M. Zarkovic, M. Medic-Stojanoska, C. Dieguez, F. Casanueva, V. Popovic
o Dopaminergik ton və piylənmə: Bromokriptin ilə müalicə olunan prolaktinomalardan bir fikir
o Eur. J. Endokrinol., 147 (2002), s. 77-84
o
46.
o Everitt və Robbins, 2005
o BJ Everitt, TW Robbins
Narkomaniya üçün sinir gücləndirmə sistemləri: hərəkətlərdən vərdişlərə qədər
o Nat. Neurosci., 8 (2005), s. 1481-1489
o
47.
o Everitt et al., 2008
o BJ Everitt, D. Belin, D. Economidou, Y. Pelloux, JW Dalley, TW Robbins
o nəzərdən keçirin. Məcburi dərman axtaran vərdiş və bağımlılığı inkişaf etdirmək üçün həssaslığın əsasını təşkil edən sinir mexanizmləri
o Filos. Trans. R. Soc. Lond. B Biol. Elm., 363 (2008), s. 3125 - 3135
o
48.
o Farooqi et al., 2007
o IS Farooqi, E. Bullmore, J. Keogh, J. Gillard, S. O'Rahilly, PC Fletcher
o Leptin, striatal bölgələri və insanın yemək davranışını tənzimləyir
o Elm, 317 (2007), səh. 1355
o
49.
o Felsted et al., 2010
o JA Felsted, X. Ren, F. Chouinard-Decorte, DM Kiçik
Birincili qida mükafatına beyin reaksiyasında genetik olaraq müəyyən edilmiş fərqlər
o J. Neurosci., 30 (2010), s. 2428-2432
o
50.
o Figlewicz et al., 2001
o DP Figlewicz, MS Higgins, SB Ng-Evans, PJ Havel
o Leptin, qida məhdudlaşdıran siçovullarda saxaroza şəraitli yer seçimini geri qaytarır
o Physiol. Davranış., 73 (2001), s. 229 - 234
o
51.
o Figlewicz et al., 2003
o DP Figlewicz, SB Evans, J. Murphy, M. Hoen, DG Baskin
o siçovulun ventral tegramal bölgəsində / substantia nigra (VTA / SN) insulin və leptin üçün reseptorların ifadəsi
o Beyin Res., 964 (2003), s. 107-115
o
52.
o Fineberg et al., 2010
o NA Fineberg, MN Potenza, SR Chamberlain, HA Berlin, L. Menzies, A. Bechara, BJ Sahakian, TW Robbins, ET Bullmore, E. Hollander
Heyvan modellərindən endopenotiplərə qədər olan məcburi və dürtüsel davranışları: Povest rəy
o Neyropsikofarmakologiya, 35 (2010), s. 591-604
o
53.
o Finkelstein et al., 2005
o EA Finkelstein, CJ Ruhm, KM Kosa
piylənmənin iqtisadi səbəbləri və nəticələri
o Annu. Rev. Xalq Sağlamlığı, 26 (2005), s. 239-257
o
54.
o Flegal et al., 2010
o KM Flegal, MD Carroll, CL Ogden, LR Curtin
o ABŞ-da böyüklər arasında piylənmənin yayılması və meylləri, 1999-2008
o JAMA, 303 (2010), s. 235 - 241
o
55.
o Foo və Mason, 2005
o H. Foo, P. Mason
o Qidalanma zamanı həssas təzyiq
o Proc. Natl. Akad. Elm. ABŞ, 102 (2005), s. 16865 - 16869
o
56.
o Franken və Muris, 2005
o IH Franken, P. Muris
o Mükafat həssaslığındakı fərdi fərqlər, sağlam qadınlarda qida istəyi və nisbi bədən çəkisi ilə əlaqədardır
o İştaha, 45 (2005), s. 198 - 201
o
57.
o Friedman et al., 2011
o A. Fridman, E. Lax, Y. Dikştein, L. İbrahim, Y. Flaumenhaft, E. Sudai, M. Ben-Tzion, G. Yadid
Yanal habenula'nın elektrikli stimullaşdırılması saxaroza özünü idarə etməsinə maneə törədir
o Neuropharmakology, 60 (2011), s. 381-387
o
58.
o Fulton et al., 2000
o S. Fulton, B. Woodside, P. Shizgal
leptin ilə beyin mükafat dövrə modulyasiya
o Elm, 287 (2000), s. 125-128
o
59.
o Fulton et al., 2006
o S. Fulton, P. Pissios, RP Manchon, L. Stiles, L. Frank, EN Pothos, E. Maratos-Flier, JS Flier
o mezoaccumbens dopamin yolunun leptinin tənzimlənməsi
o Neuron, 51 (2006), s. 811 - 822
o
60.
o Gao və Horvath, 2007
o Q. Gao, TL Horvath
o Qidalanma və enerji xərcləməsinin neyrobiologiyası
o Annu. Rev. Neurosci., 30 (2007), s. 367 – 398
o
61.
o Gautier et al., 2000
o JF Gautier, K. Chen, AD Salbe, D. Bandy, RE Pratley, M. Heiman, E. Ravussin, EM Reiman, PA Tataranni
Şişman və arıq kişilərdə doyma üçün fərqli beyin cavabları
o Diabet, 49 (2000), s. 838 - 846
o
62.
o Geiger et al., 2008
o BM Geiger, GG Behr, LE Frank, AD Caldera-Siu, MC Beinfeld, EG Kokkotou, EN Pothos
Obezite meylli siçovullarda qüsurlu mesolimbik dopamin ekzotsitozuna dair dəlil
o FASEB J., 22 (2008), s. 2740-2746
o
63.
o Geiger et al., 2009
o BM Geiger, M. Haburcak, NM Avena, MC Moyer, BG Hoebel, EN Pothos
o siçovul piylənməsində mesolimbik dopamin neyrotransmissiyasının çatışmazlıqları
o Nevrologiya, 159 (2009), s. 1193-1199
o
64.
o Ghitza et al., 2006
o UE Ghitza, SM Grey, DH Epstein, KC Rice, Y. Shaham
o Neyropsikofarmakologiya
o Anksiogen dərmanı yohimbine, siçovul residivi modelində qidalı qidaları bərpa edir: CRF (1) reseptorlarının rolu, 33 (2006), s. 2188-2196
o
65.
o Gluck et al., 2004
o ME Gluck, A. Geliebter, J. Hung, E. Yahav
o Kortizol, aclıq və binge yemək pozğunluğu olan obez qadınlarda soyuq bir stress testindən sonra yemək istəmək
o Psixosom. Med., 66 (2004), s. 876 - 881
o
66.
o Goldstone et al., 2009
o AP Goldstone, CG Prechtl de Hernandez, JD Beaver, K. Muhammed, C. Croese, G. Bell, G. Durighel, E. Hughes, AD Waldman, G. Frost, JD Bell
o Oruc tutmaq beyin mükafat sistemlərini yüksək kalorili qidalara istiqamətləndirir
o Eur. J. Neurosci., 30 (2009), s. 1625-1635
o
67.
o Halaas et al., 1995
o JL Halaas, KS Gajiwala, M. Maffei, SL Cohen, BT Chait, D. Rabinowitz, RL Lallone, SK Burley, JM Friedman
obez gen tərəfindən kodlanmış plazma zülalının çəki azaldan təsirləri
o Elm, 269 (1995), s. 543-546
o
68.
o Hamilton et al., 1995
o BS Hamilton, D. Paglia, AY Kwan, M. Deitel
o Massively obez insanlarda olan omental yağ hüceyrələrində artan obez mRNA ifadəsi
o Nat. Med., 1 (1995), s. 953 - 956
o
69.
o Hernandez və Hoebel, 1988
o L. Hernandez, BG Hoebel
o Qida mükafatı və kokain mikrodializ ilə ölçülən nüvədəki hüceyrədaxili dopamin artırır
o Həyat Elmi., 42 (1988), s. 1705-1712
o
70.
o Hill et al., 2003
o JO Hill, HR Wyatt, GW Reed, JC Peters
Obezite və ətraf mühit: Buradan hara gedirik?
o Elm, 299 (2003), s. 853-855
o
71.
o Hoebel, 1969
o BG Hoebel
o Qidalanma və özünü stimullaşdırmaq
o Ann. NY Acad. Elm., 157 (1969), s. 758 - 778
o
72.
o Hoebel və Balagura, 1967
o BG Hoebel, S. Balagura
o insulin və qlükaqon tərəfindən dəyişdirilmiş yanal hipotalamusun özünü stimullaşdırması
o Physiol. Davranış., 2 (1967), s. 337 - 340
o
73.
o Hoebel və Teitelbaum, 1962
o BG Hoebel, P. Teitelbaum
o qidalanma və özünü stimullaşdırmaq hipotalamik nəzarət
o Elm, 135 (1962), s. 375-377
o
74.
o Hoebel və Tompson, 1969
o BG Hoebel, RD Thompson
o İntraqastrik qidalanma və ya piylənmə nəticəsində yaranan yanal hipotalamik stimullaşdırmaya laqeydlik
o J. Comp. Fiziol. Psixol., 68 (1969), s. 536 - 543
o
75.
o Hofmann et al., 2010
o W. Hofmann, GM van Koningsbruggen, W. Stroebe, S. Ramanathan, H. Aarts
Zövq inkişaf etdikcə: Cazibədar qidaya Hedonik cavablar
o Psixol. Elm., 21 (2010), s. 1863 - 1870
o
76.
o Hollandiya və Gallagher, 2004
o PC Holland, M. Gallagher
o Amygdala-frontal qarşılıqlı təsir və mükafat gözləməsi
o Curr. Rəy. Neurobiol., 14 (2004), s. 148-155
o
77.
o Hollander et al., 2010
o JA Hollander, HI Im, AL Amelio, J. Kocerha, P. Bali, Q. Lu, D. Willoughby, C. Wahlestedt, MD Conkright, PJ Kenny
o Striatal microRNA, CREB siqnalının köməyi ilə kokain qəbulunu nəzarət edir
o Təbiət, 466 (2010), s. 197-202
o
78.
o Hommel et al., 2006
o JD Hommel, R. Trinko, RM Sears, D. Georgescu, ZW Liu, XB Gao, JJ Thurmon, M. Marinelli, RJ DiLeone
o midbrain dopamin neyronlarında siqnal verən Leptin reseptoru qidalanmanı tənzimləyir
o Neuron, 51 (2006), s. 801 - 810
o
79.
o Imaizumi et al., 2001
o M. İmaizumi, M. Takeda, A. Suzuki, S. Sawano, T. Fushiki
o Siçanlarda yüksək yağlı qidaya üstünlük: qaynadılmış kartofla müqayisədə qızardılmış kartof
o İştaha, 36 (2001), s. 237 - 238
o
80.
o Jerlhag et al., 2006
o E. Jerlhag, E. Egecioglu, SL Dickson, M. Andersson, L. Svensson, JA Engel
o Ghrelin, siçanlarda mərkəzi xolinergik sistemlər vasitəsi ilə lokomotor fəaliyyətini və dupamin daşmasını artırır: beyin mükafatına cəlb edilməsinin təsirləri
o Addict. Biol., 11 (2006), s. 45 - 54
o
81.
o Jerlhag et al., 2007
o E. Jerlhag, E. Egecioglu, SL Dickson, A. Douhan, L. Svensson, JA Engel
o Ghrelin tegramal bölgələrə daxil olması, lokomotor fəaliyyətini stimullaşdırır və nüvədəki hüceyrələrdə hüceyrədaxili konsentrasiyanı artırır
o Addict. Biol., 12 (2007), s. 6 - 16
o
82.
o Jhou et al., 2009
o TC Jhou, HL Alanlar, MG Baxter, CB Saper, PC Holland
o Orta beyin dopamin neyronlarına bir GABAergik afferent olan rostromedial tegramal nüvə (RMTg), əks stimulları kodlayır və motor cavablarını maneə törədir
o Neuron, 61 (2009), s. 786 - 800
o
83.
o Johnson və Kenny, 2010
o PM Johnson, PJ Kenny
Asılılıq kimi mükafat disfunksiyasında Dopamin D2 reseptorları və obez siçovullarda məcburi yemək
o Nat. Neurosci., 13 (2010), s. 635-641
o
84.
o Johnson et al., 1996
o PI Johnson, MA Parente, JR Stellar
o NMDA-nın nüvəli birləşmələri və ya ventral pallidumun zərərli siçovullara qidanın təsirli təsirini artırır
o Beyin Res., 722 (1996), s. 109-117
o
85.
o Jönsson et al., 1999
o EG Jönsson, MM Nöthen, F. Grünhage, L. Farde, Y. Nakashima, P. Propping, GC Sedvall
o Dopamin D2 reseptor genindəki polimorfizmlər və sağlam könüllülərin striatal dopamin reseptorlarının sıxlığı ilə əlaqələri
o Mol. Psixiatriya, 4 (1999), s. 290 - 296
o
86.
o Kalarchian et al., 2002
o MA Kalarchian, MD Marcus, GT Wilson, EW Labouvie, RE Brolin, LB LaMarca
Uzunmüddətli təqib zamanı mədə bypass xəstələri arasında yemək
o obes. Surg., 12 (2002), s. 270 - 275
o
87.
o Kandiah et al., 2006
o J. Kandiah, M. Yake, J. Jones, M. Meyer
o Stress kollec qadınlarında iştaha və rahat yemək seçimlərinə təsir göstərir
o Nutr. Res., 26 (2006), s. 118 - 123
o
88.
o Karhunen et al., 1997
o LJ Karhunen, RI Lappalainen, EJ Vanninen, JT Kuikka, MI Uusitupa
Obez və normal çəkili qadınlarda qida qəbulu zamanı regional beyin qanının axması
o Beyin, 120 (1997), s. 1675 - 1684
o
89.
o Kasanetz et al., 2010
o F. Kasanetz, V. Deroche-Gamonet, N. Berson, E. Balado, M. Lafourcade, O. Manzoni, PV Piazza
Bağımlılığa keçid, sinaptik plastiklikdə davamlı bir dəyərsizləşmə ilə əlaqələndirilir
o Elm, 328 (2010), s. 1709-1712
o
90.
o Kelley et al., 1996
o AE Kelley, EP Bless, CJ Swanson
o siçovullarda bəslənmə və saxaroza içməyə bəslənən tiryək antaqonistlərinin təsirini araşdırmaq
o J. Farmakol. Exp. Ther., 278 (1996), s. 1499 - 1507
o
91.
o Kelley et al., 2005
o AE Kelley, BA Baldo, WE Pratt, MJ Will
o Kortikostriatal-hipotalamik dövrə və qida motivasiyası: enerji, fəaliyyət və mükafat inteqrasiyası
o Physiol. Davranış., 86 (2005), s. 773 - 795
o
92.
o Kenny et al., 2006
o PJ Kenny, SA Chen, O. Kitamura, A. Markou, GF Koob
Şərti çəkilmə eroin istehlakını sürətləndirir və mükafat həssaslığını azaldır
o J. Neurosci., 26 (2006), s. 5894-5900
o
93.
o Kim et al., 2010
o KS Kim, YR Yoon, HJ Lee, S. Yoon, SY Kim, SW Shin, JJ An, MS Kim, SY Choi, W. Sun, JH Baik
o Dopamin D2 reseptoru olmayan siçanlarda inkişaf etmiş hipotalamik leptin siqnalları
o J. Biol. Chem., 285 (2010), s. 8905 - 8917
o
94.
o Klein et al., 2007
o TA Klein, J. Neumann, M. Reuter, J. Hennig, DY von Cramon, M. Ullsperger
səhvlərdən öyrənməkdə genetik olaraq müəyyən edilmiş fərqlər
o Elm, 318 (2007), s. 1642-1645
o
95.
o Kojima et al., 1999
o M. Kojima, H. Hosoda, Y. Tarix, M. Nakazato, H. Matsuo, K. Kangawa
o Ghrelin, böyümə hormonunu ifraz edən mədədən asitləşdirilmiş peptiddir
o Təbiət, 402 (1999), s. 656-660
o
96.
o Kok et al., 2006
o P. Kok, F. Roelfsema, M. Frölich, J. van Pelt, AE Meinders, H. Pijl
o Dopamin D2 reseptorlarının aktivləşdirilməsi obez qadınlarda sirkadiyalı leptinin konsentrasiyasını azaldır
o J. Clin. Endokrinol. Metab., 91 (2006), s. 3236 - 3240
o
97.
o Koob, 2010
o GF Koob
Bağımlılığın qaranlıq tərəfində CRF və CRF ilə əlaqəli peptidlərin rolu
o Beyin Res., 1314 (2010), s. 3-14
o
98.
o Koob və Le Moal, 2008
o GF Koob, M. Le Moal
Bağımlılık və beyin antireward sistemi
o Annu. Rev. Psixol., 59 (2008), s. 29-53
o
99.
o Koob və Volkow, 2010
o GF Koob, ND Volkow
Bağımlılığın neyrokirkulyasiyası
o Neyropsikofarmakologiya, 35 (2010), s. 217-238
o
100.
o Koob və Zorrilla, 2010
o GF Koob, EP Zorrilla
Bağımlılığın neyrobioloji mexanizmləri: Kortikotropini sərbəst buraxan amilə diqqət
o Curr. Rəy. Tədqiq et. Dərmanlar, 11 (2010), s. 63 - 71
o
101.
o Kringelbach et al., 2003
o ML Kringelbach, J. O'Doherty, ET Rolls, C. Andrews
o İnsanın orbitofontral korteksinin maye qida stimuluna qoşulması onun subyektiv xoşluğu ilə əlaqələndirilir
o Cereb. Korteks, 13 (2003), s. 1064 - 1071
o
102.
o Krügel et al., 2003
o U. Krügel, T. Schraft, H. Kittner, W. Kiess, P. Illes
siçovul nüvəsi böyüdücü bazal və bəslənən dopamin sərbəst buraxılması leptin tərəfindən depressiyaya uğrayır
o Eur. J. Farmakol., 482 (2003), s. 185-187
o
103.
o la Fleur, 2006
o SE la Fleur
o Siçovullarda qidalanma davranışına qlükokortikoidlərin təsiri
o Physiol. Davranış., 89 (2006), s. 110 - 114
o
104.
o LaBar et al., 2001
o KS LaBar, DR Gitelman, TB Parrish, YH Kim, AC Nobre, MM Mesulam
O aclıq insanlarda kortikolimbik aktivləşdirməni qida stimullarına seçici olaraq modulyasiya edir
o Behav. Neurosci., 115 (2001), s. 493-500
o
105.
o Latagliata et al., 2010
o EC Latagliata, E. Patrono, S. Puglisi-Allegra, R. Ventura
Zərərli nəticələrə baxmayaraq qidalanma prefrontal kortikal noradrenergik nəzarət altındadır
o BMC Neurosci., 11 (2010), səh. 15
o
106.
o Lawford et al., 2000
o BR Lawford, RM Young, EP Noble, J. Sargent, J. Rowell, S. Shadforth, X. Zhang, T. Ritchie
o D (2) dopamin reseptoru A (1) allele və opioid asılılığı: heroin istifadəsi və metadon müalicəsinə reaksiya
o Am. J. Med. Genet., 96 (2000), s. 592 - 598
o
107.
o Leinninger et al., 2009
o GM Leinninger, YH Jo, RL Leshan, GW Louis, H. Yang, JG Barrera, H. Wilson, DM Opland, MA Faouzi, Y. Gong et al.
o Leptin, mesolimbik dopamin sistemini tənzimləmək və bəslənməni yatırmaq üçün leptin reseptorunu ifadə edən yan hipotalamik neyronlarla hərəkət edir.
o Hüceyrə Metab., 10 (2009), s. 89-98
o
108.
o Lenoir et al., 2007
o M. Lenoir, F. Serre, L. Cantin, SH Ahmed
o Sıx şirinlik kokain mükafatından üstündür
o PLoS ONE, 2 (2007), səh. e698 http://dx.doi.org/10.1371/journal.pone.0000698
o
109.
o Leshan et al., 2010
o RL Leshan, DM Opland, GW Louis, GM Leinninger, CM Patterson, CJ Rodos, H. Münzberg, MG Myers Jr.
o Ventral tegramal bölgədə leptin reseptor neyronları genişləndirilmiş mərkəzi amigdalanın kokain və amfetamin tənzimlənmiş transkript neyronlarını xüsusi olaraq layihələndirir və tənzimləyir.
o J. Neurosci., 30 (2010), s. 5713-5723
o
110.
o Louis et al., 2010
o GW Louis, GM Leinninger, CJ Rodos, MG Myers Jr.
lateral hipotalamik LepRb neyronları tərəfindən orexin neyronlarının birbaşa innervasiyası və modulyasiyası
o J. Neurosci., 30 (2010), s. 11278-11287
o
111.
o Luppino et al., 2010
o FS Luppino, LM de Wit, PF Bouvy, T. Stijnen, P. Cuijpers, BW Penninx, FG Zitman
o Kilolu, piylənmə və depressiya: sistematik bir baxış və uzunlamasına tədqiqatların meta analizi
o Arch. Genetik psixiatriya, 67 (2010), s. 220-229
o
112.
o Lutter və Nestler, 2009
o M. Lutter, EJ Nestler
o Homeostatik və hedonik siqnallar qida qəbulunu tənzimləyir
o J. Nutr., 139 (2009), s. 629-632
o
113.
o Macht və Mueller, 2007
o M. Macht, J. Mueller
o Şokoladın eksperimental olaraq yaranan əhval vəziyyətlərinə dərhal təsiri
o İştaha, 49 (2007), s. 667 - 674
o
114.
o Maldonado-Irizarry və Kelley, 1995
o CS Maldonado-Irizarry, AE Kelley
o Nüvənin əsas və qabıq subregionlarının eksitotoksik lezyonları bədənin ağırlığının tənzimlənməsini və siçovuldakı motor fəaliyyətini fərqli şəkildə pozur
o Beyin Res. Bull., 38 (1995), s. 551 - 559
o
115.
o Maldonado-Irizarry et al., 1995
o CS Maldonado-Irizarry, CJ Swanson, AE Kelley
o Nüvədəki glutamat reseptorları yanal hipotalamusun üzərindəki qidalanma davranışını tənzimləyir
o J. Neurosci., 15 (1995), s. 6779-6788
o
116.
o Malik et al., 2008
o S. Malik, F. McGlone, D. Bedrossian, A. Dagher
o Ghrelin, iştahsız davranışa nəzarət edən bölgələrdə beyin fəaliyyətini modulyasiya edir
o Hüceyrə Metab., 7 (2008), s. 400-409
o
117.
o Man et al., 2009
o MS Man, HF Clarke, AC Roberts
o Orbitof Frontal korteksin və medial striatumun qida mükafatlarına üstünlük verilən cavabların tənzimlənməsində rolu
o Cereb. Korteks, 19 (2009), s. 899 - 906
o
118.
o Margules və Olds, 1962
o DL Margules, J. Olds
Siçovulların yan hipotalamusundakı "qidalanma" və "mükafatlandıran" sistemlər
o Elm, 135 (1962), s. 374-375
o
119.
o Markou və Frank, 1987
o A. Markou, RA Frank
o Operant və elektrod yerləşdirilməsinin özünü stimullaşdıran qatar müddəti cavab funksiyalarına təsiri
o Physiol. Davranış., 41 (1987), s. 303 - 308
o
120.
o Markou və Koob, 1991
o A. Markou, GF Koob
o Postokokain anhedoniyası. Kokainin çəkilməsinin heyvan modeli
o Neyropsikofarmakologiya, 4 (1991), s. 17-26
o
121.
o Mastronardi et al., 2001
o CA Mastronardi, WH Yu, VK Srivastava, WL Dees, SM McCann
o Lipopolisakkaridlə əlaqəli leptinin sərbəst buraxılması neyron tərəfindən idarə olunur
o Proc. Natl. Akad. Elm. ABŞ, 98 (2001), s. 14720 - 14725
o
122.
o Matheny et al., 2011
o M. Matheny, A. Shapiro, N. Tumer, PJ Scarpace
o Bölgəyə xas pəhriz və leptin təsirli hüceyrə leptinə qarşı müqavimət siçovullarda ventral tegramal bölgəni əhatə edir
o Neuropharmakology (2011) http://dx.doi.org/10.1016/j.neuropharm.2010.11.002 mətbuatda. Onlayn Noyabr 5, 2010
o
123.
o Matsumoto və Hikosaka, 2007
o M. Matsumoto, O. Hikosaka
o Yanal habenula, dopamin neyronlarında mənfi mükafat siqnalları mənbəyi kimi
o Təbiət, 447 (2007), s. 1111-1115
o
124.
o Morton et al., 2006
o GJ Morton, DE Cummings, DG Baskin, GS Barsh, MW Schwartz
o Mərkəzi sinir sistemi qida qəbulunu və bədən çəkisini nəzarət edir
o Təbiət, 443 (2006), s. 289-295
o
125.
o Dağ və Hoebel, 1967
o G. Dağı, BG Hoebel
o yan hipotalamik özünü stimullaşdırma: qida qəbulu ilə müəyyən edilmiş hədd artmışdır
o Psixon. Elm., 9 (1967), s. 265 - 266
o
126.
o Nair et al., 2011
o SG Nair, BM Navarre, C. Cifani, CL Pickens, JM Bossert, Y. Şaham
o Anksiogen dərmanı yohimbine səbəb olan yüksək yağlı qidaya reaksiya verən dorsal medial prefrontal korteks dopamin D1 ailəli reseptorların rolu
o Neyropsikofarmakologiya, 36 (2011), s. 497-510
o
127.
o Nakazato et al., 2001
o M. Nakazato, N. Murakami, Y. Date, M. Kojima, H. Matsuo, K. Kangawa, S. Matsukura
o Bəslənmənin mərkəzi tənzimlənməsində grrelin rolu
o Təbiət, 409 (2001), s. 194-198
o
128.
o Naqvi və Bechara, 2009
o NH Naqvi, A. Bechara
Gizli asılılıq adası: İnsula
o tendensiyalar Neurosci., 32 (2009), s. 56-67
o
129.
o Neville et al., 2004
o MJ Neville, EC Johnstone, RT Walton
o ANKK1-ın eyniləşdirilməsi və xarakteristikası: 2q11 xromosoma qrupunda DRD23.1 ilə sıx əlaqəli yeni bir kinaz geni
o Hum. Mutat., 23 (2004), s. 540 - 545
o
130.
o Nirenberg və Waters, 2006
o MJ Nirenberg, C. Waters
o Dopamin agonist istifadəsi ilə əlaqəli kompulsif yemək və çəki artımı
o Mov. İtirmə., 21 (2006), s. 524 - 529
o
131.
o Soylu, 2000
o EP Soylu
o D2 dopamin reseptor geninin polimorfizmi vasitəsilə bağımlılıq və onun mükafatlandırma prosesi: Rəy
o Eur. Psixiatriya, 15 (2000), s. 79 - 89
o
132.
o Noble et al., 1993
o EP Noble, K. Blum, ME Khalsa, T. Ritchie, A. Montgomery, RC Wood, RJ Fitch, T. Ozkaragoz, PJ Sheridan, MD Anglin et al.
o kokaindən asılılığı olan D2 dopamin reseptor geninin allel birləşməsi
Narkotik alkoqoldan asılılıq., 33 (1993), s. 271-285
o
133.
o Noble et al., 2000
o EP Noble, X. Zhang, TL Ritchie, RS Sparkes
o DRD2 lokusundakı Haplotiplər və ağır alkoqolizm
o Am. J. Med. Genet., 96 (2000), s. 622 - 631
o
134.
o O'Connor et al., 2008
o DB O'Connor, F. Jones, M. Conner, B. McMillan, E. Ferguson
Gündəlik əngəllərin və yemək tərzinin yemək davranışına təsiri
o Sağlamlıq Psixolu, 27 (1, Ehtiyat) (2008), səh. S20 - S31
o
135.
o O'Doherty et al., 2002
o JP O'Doherty, R. Deichmann, HD Critchley, RJ Dolan
ilkin zövq mükafatını gözləyərkən sinir cavabları
o Neuron, 33 (2002), s. 815 - 826
o
136.
o O'Rahilly, 2009
o S. O'Rahilly
o İnsan genetikası metabolik xəstəliyin yollarını işıqlandırır
o Təbiət, 462 (2009), s. 307-314
o
137.
o Osswald et al., 2010
o KD Oswald, DL Murdaugh, VL King, MM Boggiano
binge yeməyin heyvan modelindəki nəticələrə baxmayaraq ləzzətli yemək üçün motivasiya
o Int. J. Yeyin. Razılıq. (2010) http://dx.doi.org/10.1002/eat.20808 mətbuatda. Fevral 22, 2010, onlayn nəşr olundu
o
138.
o Park və Carr, 1998
o TH Park, KD Carr
o salinli və naltreksonla müalicə olunan siçovullarda xoş ovqat və yeməklə əlaqəli mühitin yaratdığı fos tipli immunoreaktivliyin neyroanatomik nümunələri
o Beyin Res., 805 (1998), s. 169-180
o
139.
o Peciña və Berridge, 2005
o S. Peciña, KC Berridge
o Nüvədəki Hedonik isti nöqtə accumbens shell: mu-opioidlər şirinliyin artan hedonik təsirinə səbəb olur?
o J. Neurosci., 25 (2005), s. 11777-11786
o
140.
o Peciña et al., 2006a
o S. Peciña, J. Schulkin, KC Berridge
o Nucleus kortikotropini sərbəst buraxan amil, saxaroza mükafatı üçün replika stimullaşdırmasını artırır: Stresdə paradoksal müsbət həvəsləndirici təsir göstərirmi?
o BMC Biol., 4 (2006), səh. 8
o
141.
o Peciña et al., 2006b
o S. Peciña, KS Smith, KC Berridge
beyindəki hedonik isti nöqtələr
o Neuroscientist, 12 (2006), s. 500-511
o
142.
o Pecoraro et al., 2004
o N. Pecoraro, F. Reyes, F. Gomez, A. Bhargava, MF Dallman
Xroniki stress, stres əlamətlərini azaldan xoşagəlməz qidalanmanı təşviq edir: Xroniki stressin irəli və geribildirim təsirləri
o Endokrinologiya, 145 (2004), s. 3754-3762
o
143.
o Pelchat et al., 2004
o ML Pelchat, A. Johnson, R. Chan, J. Valdez, JD Ragland
o İstək şəkilləri: FMRI zamanı yemək istəyi aktivləşdirilməsi
o Neuroimage, 23 (2004), s. 1486-1493
o
144.
o Pelleymounter et al., 1995
o MA Pelleymounter, MJ Cullen, MB Baker, R. Hecht, D. Winters, T. Boone, F. Collins
O ob ob siçanlarında bədən çəkisinin tənzimlənməsinə obez gen məhsulunun təsiri
o Elm, 269 (1995), s. 540-543
o
145.
o Pelloux et al., 2007
o Y. Pelloux, BJ Everitt, A. Dickinson
Cəza altında siçovulların axtardığı kompulsif dərmanlar: Dərman qəbul etmə tarixinin təsiri
o Psixofarmakologiya (Berl.), 194 (2007), s. 127-137
o
146.
o Perello et al., 2010
o M. Perello, I. Sakata, S. Birnbaum, JC Chuang, S. Osborne-Lawrence, SA Rovinsky, J. Woloszyn, M. Yanagisawa, M. Lutter, JM Zigman
o Ghrelin, yüksək yağlı pəhrizin tələffüz dəyərini bir oreksinə bağlı bir şəkildə artırır
o Biol. Psixiatriya, 67 (2010), s. 880 - 886
o
147.
o Petrovich et al., 2009
o GD Petrovich, CA Ross, P. Mody, PC Holland, M. Gallagher
o Mərkəzi, lakin bazolateral deyil amigdala, mülayim öyrənilən istəklərlə qidalanma nəzarəti üçün vacibdir
o J. Neurosci., 29 (2009), s. 15205-15212
o
148.
o Pfaffly et al., 2010
o J. Pfaffly, M. Michaelides, GJ Wang, JE Pessin, ND Volkow, PK Thanos
o Leptin, leptin çatışmazlığı olan obez (ob / ob) siçanlarda bağlanmış striatal dopamin D2 reseptorunu artırır.
o Sinaps, 64 (2010), s. 503 - 510
o
149.
o Puhl et al., 2008
o RM Puhl, CA Moss-Racusin, MB Schwartz, KD Brownell
o Çəki stiqmatizasiyası və qərəz azaldılması: Kilolu və piylənmiş böyüklərin perspektivləri
o Sağlamlıq Təhsili. Res., 23 (2008), s. 347 - 358
o
150.
o Rada et al., 2010
o P. Rada, ME Bocarsly, JR Barson, BG Hoebel, SF Leibowitz
o yağla zəngin bir pəhriz yeməyə meylli olan Sprague-Dawley siçovullarında azaldılmış dopamin dumbamin
o Physiol. Davranış., 101 (2010), s. 394 - 400
o
151.
o Ritchie və Noble, 2003
o T. Ritchie, EP Soylu
beyin reseptoru bağlayıcı xüsusiyyətləri olan D2 dopamin reseptor geninin yeddi polimorfizmi assosiasiyası
o neyrokim. Res., 28 (2003), s. 73 - 82
o
152.
o Roberto et al., 2010
o M. Roberto, MT Cruz, NW Gilpin, V. Sabino, P. Schweitzer, M. Bajo, P. Cottone, SG Madamba, DG Stouffer, EP Zorrilla et al.
o Kortikotropin sərbəst buraxan amigdala gamma-aminobutyric turşusunun sərbəst buraxılması spirt asılılığında əsas rol oynayır.
o Biol. Psixiatriya, 67 (2010), s. 831 - 839
o
153.
o Roitman et al., 2004
o MF Roitman, GD Stuber, PE Phillips, RM Wightman, RM Carelli
o Dopamin qidalanma təklif edən bir modekond kimi fəaliyyət göstərir
o J. Neurosci., 24 (2004), s. 1265-1271
o
154.
o Roitman et al., 2008
o MF Roitman, RA Wheeler, RM Wightman, RM Carelli
o nüvədəki real vaxt kimyəvi reaksiyalar mükafatlandırıcı və aversiv stimulları fərqləndirir
o Nat. Neurosci., 11 (2008), s. 1376-1377
o
155.
o Rolls, 2008
o ET Rolls
o Dad, olfaksiya, iştah və duyğu baxımından orbitofrontal və pregenual cingulate korteksin funksiyaları
o Acta Physiol. Hung., 95 (2008), s. 131 - 164
o
156.
o Rolls, 2010
o ET Rolls
o Dad, ətirli və qida toxuması beyində və piylənmədə mükafat emalı
o Int. J. Obes. (Lond.) (2010) http://dx.doi.org/10.1038/ijo.2010.155 mətbuatda. Onlayn Avqust 3, 2010
o
157.
o Rolls et al., 1983
o ET Rolls, BJ Rolls, EA Rowe
o İnsanda qida və suyun görünməsi və dadı üçün həssas və motivasiyaya xas olan doyma
o Physiol. Davranış., 30 (1983), s. 185 - 192
o
158.
o Roseberry et al., 2007
o AG Roseberry, T. Painter, GP Mark, JT Williams
leptin çatışmazlığı olan siçanlarda vezikulyar somatodendritik dopamin anbarlarının azalması
o J. Neurosci., 27 (2007), s. 7021-7027
o
159.
o Rothemund et al., 2007
o Y. Rothemund, C. Preuschhof, G. Bohner, HC Bauknecht, R. Klingebiel, H. Flor, BF Klapp
o obez şəxslərdə yüksək kalorili vizual qida stimulları ilə dorsal striatumun differensial aktivləşdirilməsi
o Neuroimage, 37 (2007), s. 410-421
o
160.
o Salas et al., 2010
o R. Salas, P. Baldwin, M. de Biasi, PR Montague
o insan habenulasında mənfi mükafat proqnozu səhvlərinə BOLD cavablar
o Ön. Xum. Neurosci., 4 (2010), səh. 36
o
161.
o Saper et al., 2002
o CB Saper, TC Chou, JK Elmquist
o qidalanma ehtiyacı: yeməyə homeostatik və hedonik nəzarət
o Neuron, 36 (2002), s. 199 - 211
o
162.
o Saunders, 2001
o R. Saunders
o Məcburi yemək və mədə damar əməliyyatı: Aclığın bunun nə ilə əlaqəsi var?
o obes. Surg., 11 (2001), s. 757 - 761
o
163.
o Scarpace et al., 2010
o PJ Scarpace, M. Matheny, Y. Zhang
o Təkər çalışması yüksək yağlı üstünlükləri aradan qaldırır və ventral tegramal bölgədə leptin siqnalını artırır
o Physiol. Davranış., 100 (2010), s. 173 - 179
o
164.
o Schiltz et al., 2007
o CA Schiltz, QZ Bremer, CF Landry, AE Kelley
Qida ilə əlaqəli istəklər, dərhal erkən gen və proenkefalin ifadəsi ilə qiymətləndirildiyi kimi beyin beyin funksional bağlantısını dəyişdirir.
o BMC Biol., 5 (2007), səh. 16
o
165.
o Schur et al., 2009
o EA Schur, NM Kleinhans, J. Goldberg, D. Buchwald, MW Schwartz, K. Maravilla
Beyin enerjisinin tənzimlənməsi və mükafat mərkəzlərində qida sapları ilə aktivləşdirmə vizual stimulun seçimi ilə dəyişir
o Int. J. Obes. (Lond.), 33 (2009), s. 653 - 661
o
166.
o Sclafani et al., 1998
o A. Sclafani, RJ Bodnar, AR Delamater
qida şərti üstünlüklərinin farmakologiyası
o İştaha, 31 (1998), səh. 406
o
167.
o Sescousse et al., 2010
o G. Sescousse, J. Redouté, JC Dreher
o İnsan orbitofortal korteksində mükafat dəyərinin kodlaşdırılması memarlığı
o J. Neurosci., 30 (2010), s. 13095-13104
o
168.
o Shomaker et al., 2010
o LB Shomaker, M. Tanofsky-Kraff, JM Zocca, A. Courville, M. Kozlosky, KM Columbo, LE Wolkoff, SM Brady, MK Crocker, AH Ali et al.
o Yetkinlərdə aclıq olmadıqda yemək: Normal bir yeməkdən sonra ilə müqayisədə böyük bir sıra yeməkdən sonra qəbul edin.
o Am. J. Clin. Nutr., 92 (2010), s. 697 - 703
o
169.
o Simmons et al., 2005
o WK Simmons, A. Martin, LW Barsalou
o İştahaaçan qidaların şəkilləri zövq və mükafat üçün gustatory korteksləri aktivləşdirir
o Cereb. Korteks, 15 (2005), s. 1602 - 1608
o
170.
o Kiçik, 2010
o DM Kiçik
o İnsanın insulindəki dadın təmsil olunması
o Beyin quruluşu. Funksiya, 214 (2010), s. 551 - 561
o
171.
o Kiçik et al., 2003
o DM Kiçik, M. Jones-Gotman, A. Dagher
o Dorsal striatumda qidalanma nəticəsində yaranan dopamin azadlığı, insan sağlam könüllülərindəki yemək ləzzəti ilə əlaqələndirilir
o Neuroimage, 19 (2003), s. 1709-1715
o
172.
o Smith and Berridge, 2007
o KS Smith, KC Berridge
Mükafat üçün opioid limbik dövrə: nüvələrin hedonic isti nöqtələri ilə ventral pallidum arasında qarşılıqlı əlaqə
o J. Neurosci., 27 (2007), s. 1594-1605
o
173.
o Söderpalm və Berridge, 2000
o AH Söderpalm, KC Berridge
o Diazepam, morfin və ya muskololdan sonra qida qəbulu: Nüvədəki mikroinjeksiyonlar
o Farmakol. Biokim. Davranış., 66 (2000), s. 429 - 434
o
174.
o Stice et al., 2008a
o E. Stice, S. Spoor, C. Bohon, DM Kiçik
Oezlik və qida üçün kəskin striatal reaksiya arasındakı münasibət Taqia A1 alleli tərəfindən idarə olunur
o Elm, 322 (2008), s. 449-452
o
175.
o Stice et al., 2008b
o E. Stice, S. Spoor, C. Bohon, MG Veldhuizen, DM Kiçik
Qidalanma və gözlənilən qida qəbulundan piylənməyə olan mükafatın əlaqəsi: Funksional maqnetik rezonans görüntüləmə işi
o J. Abnorm. Psixol., 117 (2008), s. 924 - 935
o
176.
o Stice et al., 2010a
o E. Stice, S. Yokum, K. Blum, C. Bohon
o Çəki artımı, keyfiyyətsiz qidaya olan striatal reaksiyanın azalması ilə əlaqələndirilir
o J. Neurosci., 30 (2010), s. 13105-13109
o
177.
o Stice et al., 2010b
o E. Stice, S. Yokum, C. Bohon, N. Marti, A. Smolen
o Qidaya mükafat vermə dövriyyəsi, bədən kütləsinin gələcəkdə artacağını proqnozlaşdırır: DRD2 və DRD4-ın moderator təsirləri
o Neuroimage, 50 (2010), s. 1618-1625
o
178.
o Stoeckel, 2010
o Stoeckel, LE (2010). Piylənmənin Qoldilok prinsipi. Elmi Amerika. İyun 8, 2010. http://www.scientificamerican.com/article.cfm?id=the-goldilocks-principle-obesity.
o
179.
o Stratford və Kelley, 1997
o TR Stratford, AE Kelley
o Nüvədəki GABA accumbens qabığında qidalanma davranışının mərkəzi tənzimlənməsində iştirak edir
o J. Neurosci., 17 (1997), s. 4434-4440
o
180.
o Stratford və Kelley, 1999
o TR Stratford, AE Kelley
o, qidalanma davranışına nəzarət edən nüvənin böyüməsi qabığı və yanal hipotalamus arasındakı funksional əlaqənin sübutu
o J. Neurosci., 19 (1999), s. 11040-11048
o
181.
o Suarez və Gallup, 1981
o SD Suarez, GGJ Gallup
o siçovullarda və siçanlarda açıq sahə davranışının etoloji təhlili
o öyrənin. Motiv., 12 (1981), s. 342 - 363
o
182.
o Bazar və s., 1983
o SR Bazar günü, SA Sanders, G. Collier
o Doyma və yemək nümunələri
o Physiol. Davranış., 30 (1983), s. 915 - 918
o
183.
o Swinburn et al., 2009
o B. Swinburn, G. Sacks, E. Ravussin
ABŞ-da piylənmə epidemiyasını izah etmək üçün artan qida enerjisi təchizatı kifayətdir
o Am. J. Clin. Nutr., 90 (2009), s. 1453 - 1456
o
184.
o Taha və Sahələr, 2005
o SA Taha, HL sahələri
o Nüvə accumbensindəki fərqli neyron populyasiyalar tərəfindən rahatlıq və iştahaaçan davranışların kodlaşdırılması
o J. Neurosci., 25 (2005), s. 1193-1202
o
185.
o Taha et al., 2009
o SA Taha, Y. Katsuura, D. Noorvash, A. Seroussi, HL Sahələri
o Seriya deyil, striatal və pallidal dövranlar, opioid təsirli qida qəbulunu tənzimləyir
o Nevrologiya, 161 (2009), s. 718-733
o
186.
o Teegarden və Bale, 2007
o SL Teegarden, TL Bale
o Pəhriz seçimində azalma, artan emosionallıq və pəhriz residivi üçün risk yaradır
o Biol. Psixiatriya, 61 (2007), s. 1021 - 1029
o
187.
o Teegarden et al., 2009
o SL Teegarden, AN Scott, TL Bale
erkən yaşda yüksək yağlı bir diyetə məruz qalma, pəhriz seçimlərində və mərkəzi mükafat siqnalında uzun müddətli dəyişikliklərə kömək edir
o Nevrologiya, 162 (2009), s. 924-932
o
188.
o Thanos et al., 2008
o PK Thanos, M. Michaelides, YK Piyis, GJ Wang, ND Volkow
o Qida məhdudiyyəti in-vivo muPET görüntüləmə ([2C] raklopride) və in-vitro ([2H] spiperone) avtoreqrafiyasında qiymətləndirildiyi kimi, piylənmənin siçovul modelində dopamin D11 reseptorunu (D3R) artırır.
o Sinaps, 62 (2008), s. 50 - 61
o
189.
o Thompson və Swanson, 2010
o RH Thompson, LW Swanson
o Hipotezaya əsaslanan struktur bağlantısı təhlili beyin memarlığının iyerarxik modeli üzərində şəbəkəni dəstəkləyir
o Proc. Natl. Akad. Elm. ABŞ, 107 (2010), s. 15235 - 15239
o
190.
o Vanderschuren və Everitt, 2004
o LJ Vanderschuren, BJ Everitt
Dərman istəməsi kokainin özünü idarə etməsindən sonra məcbur olur
o Elm, 305 (2004), s. 1017-1019
o
191.
o Vendruscolo et al., 2010a
o LF Vendruscolo, AB Gueye, M. Darnaudéry, SH Ahmed, M. Cador
Yetkinlik dövründə şəkərin həddindən artıq olması böyüklərdəki siçovullarda motivasiya və mükafat funksiyasını seçici olaraq dəyişdirir
o PLoS ONE, 5 (2010), səh. e9296 http://dx.doi.org/10.1371/journal.pone.0009296
o
192.
o Vendruscolo et al., 2010b
o LF Vendruscolo, AB Gueye, JC Vendruscolo, KJ Clemens, P. Mormède, M. Darnaudéry, M. Cador
o yetkinlik dövründə saxaroza məruz qalan yetkin siçovullarda spirtli içkilərin azaldılması
o Neuropharmakology, 59 (2010), s. 388-394
o
193.
o Volkow və O'Brien, 2007
o ND Volkow, CP O'Brien
o DSM-V üçün problemlər: Piylənmə beyin pozğunluğu kimi daxil edilməlidirmi?
o Am. J. Psixiatriya, 164 (2007), s. 708-710
o
194.
o Volkow və Wise, 2005
o ND Volkow, RA Müdrik
o Narkomanlıq piylənməni anlamağımıza necə kömək edə bilər?
o Nat. Neurosci., 8 (2005), s. 555-560
o
195.
o Volkow et al., 2009
o ND Volkow, GJ Wang, F. Telang, JS Fowler, RZ Goldstein, N. Alia-Klein, J. Logan, C. Wong, PK Thanos, Y. Ma, K. Pradhan
o BMI və sağlam yetkinlərdə prefrontal metabolik fəaliyyət arasında tərs birləşmə
Obezite (Gümüş Bahar), 17 (2009), s. 60-65
o
196.
o Vucetic və Reyes, 2010
o Z. Vucetic, TM Reyes
qida qəbulunu və mükafatı idarə edən mərkəzi dopaminergik dövrə: piylənmənin tənzimlənməsi üçün təsirlər
o Wiley Interdiscip. Rev. Syst Biol. Med., 2 (2010), s. 577 - 593
o
197.
o Wang et al., 2001
o GJ Wang, ND Volkow, J. Logan, NR Pappas, CT Wong, W Zhu, N. Netusil, JS Fowler
o Beyin dopamin və piylənmə
o Lancet, 357 (2001), s. 354 - 357
o
198.
o Wang et al., 2002
o GJ Wang, ND Volkow, JS Fowler
o Dopamin insanlarda qida motivasiyasında rolu: Piylənmə üçün təsirlər
o Ekspert rəyi. Ther. Hədəflər, 6 (2002), s. 601 - 609
o
199.
o Wang et al., 2004a
o GJ Wang, ND Volkow, F. Telang, M. Jayne, J. Ma, M. Rao, W. Zhu, CT Wong, NR Pappas, A. Geliebter, JS Fowler
o İştahsız qida stimullarına məruz qalmaq insan beynini aktivləşdirir
o Neuroimage, 21 (2004), s. 1790-1797
o
200.
o Wang et al., 2004b
o GJ Wang, ND Volkow, PK Thanos, JS Fowler
o, neyrofunksional görüntüləmə ilə qiymətləndirilən piylənmə və narkomaniya arasındakı oxşarlıq: bir konsepsiya icmalı
o J. Addict. Dis., 23 (2004), s. 39 - 53
o
201.
o Wang et al., 2008
o GJ Wang, D. Tomasi, W. Backus, R. Wang, F. Telang, A. Geliebter, J. Korner, A. Bauman, JS Fowler, PK Thanos, ND Volkow
o Mədə pozğunluğu insan beynində doyma dövranını aktivləşdirir
o Neuroimage, 39 (2008), s. 1824-1831
o
202.
o Wassum et al., 2009
o KM Wassum, SB Ostlund, NT Maidment, BW Balleine
Fərqli opioid sxemləri qonaqlıq verici hadisələrin rahatlığını və arzuolunanlığını təyin edir
o Proc. Natl. Akad. Elm. ABŞ, 106 (2009), s. 12512 - 12517
o
203.
o Wassum et al., 2011
o KM Wassum, IC Cely, BW Balleine, NT Qızdırma
Bazolateral amigdala'dakı o io-Opioid reseptorunun aktivləşdirilməsi artımların öyrənilməsinə vasitəçilik edir, lakin qida mükafatının stimullaşdırıcı dəyərində azalmır.
o J. Neurosci., 31 (2011), s. 1591-1599
o
204.
o Wilkinson və Peele, 1962
o HA Wilkinson, TL Peele
o Aclıq doyma ilə intrakranial özünü stimullaşdırmanın modifikasiyası
o Am. J. Physiol., 203 (1962), s. 537 - 540
o
205.
o Will et al., 2003
o MJ Will, EB Franzblau, AE Kelley
o Nucleus accumbens mu-opioidlər paylanmış beyin şəbəkəsinin aktivləşdirilməsi yolu ilə yüksək yağlı pəhriz qəbulunu tənzimləyir.
o J. Neurosci., 23 (2003), s. 2882-2888
o
206.
o Wilson et al., 2008
o ME Wilson, J. Fisher, A. Fischer, V. Lee, RB Harris, TJ Bartness
Sosial vəziyyətdə olan meymunlarda qida qəbulunun miqdarı: sosial vəziyyətin kalorili istehlaka təsiri
o Physiol. Davranış., 94 (2008), s. 586 - 594
o Maddə | PDF (553 K) |
Scopus-da qeydiyyatdan keçin
| Scopus-a istinadən (22)
207.
o Yurcisin et al., 2009
o BM Yurcisin, MM Gaddor, EJ DeMaria
piylənmə və bariatrik əməliyyat
o Klinika. Sinə Med., 30 (2009), s. 539-553, ix
o
208.
o Zhang et al., 1998
o M. Zhang, BA Gosnell, AE Kelley
o Yüksək yağlı qidaların qəbulu, nüvənin birləşmələrində mu opioid reseptorun stimullaşdırılması ilə seçilmiş şəkildə artırılır
o J. Farmakol. Exp. Ther., 285 (1998), s. 908 - 914
Scopus-da qeydlərə baxın
| Scopus-a istinadən (148)
209.
o Zheng et al., 2007
o H. Zheng, LM Patterson, HR Berthoud
ventral tegramal bölgədəki Orexin siqnalının, nüvələrin böyüdülməsini opioid stimullaşdırması ilə meydana gələn yüksək yağlı iştaha tələb olunur.
o J. Neurosci., 27 (2007), s. 11075-11082
Scopus-da qeydlərə baxın
| Scopus-a istinadən (64)
210.
o Zheng et al., 2009
o H. Zheng, NR Lenard, AC Shin, HR Berthoud
Müasir dünyada iştaha nəzarəti və enerji balansının tənzimlənməsi: Mükafatla işləyən beyin əvəzetmə siqnallarını ləğv edir
o Int. J. Obes. (Lond.), 33 (Suppl 2) (2009), s. S8 - S13
Scopus-da qeydlərə baxın
| Scopus-a istinadən (34)