COMENTARIS: Tot i que l'estudi utilitza el terme "Comportament sexual compulsiu (CSB)", els subjectes eren addictes al porno (mireu aquest comunicat de premsa). En comparació amb els controls sans, els subjectes CSB van augmentar el volum de l'amígdala esquerra i van reduir la connectivitat funcional durant el descans entre l’amígdala esquerra i l’escorça prefrontal dorsolateral bilateral DLPFC. Els autors conclouen:
Els nostres resultats actuals posen de relleu els volums elevats en una regió implicada en la rellevància motivacional i una menor connectivitat en estat de repòs de les xarxes reguladores de prefrontal de dalt a baix. La interrupció d’aquestes xarxes pot explicar els patrons de comportament aberents cap a una recompensa o una reactivitat millorable ambientalment per a indicis d’incentius destacats. Tot i que els nostres resultats volumètrics contrasten amb els dels trastorns per a l’ús de substàncies (SUD), aquestes troballes poden reflectir diferències en funció dels efectes neurotòxics de l’exposició crònica a les drogues.
Cerca de traduccions #1): "Menys connectivitat funcional entre l'amígdala i l'escorça prefrontal dorsolateral". L’amígdala té un paper clau en el processament de les emocions, inclosa la nostra resposta a l’estrès. L'amígdala està fortament implicada en molts aspectes de l'addicció, com ara els desitjos, la reactivitat i els símptomes d'abstinència. La connectivitat funcional reduïda entre l’amígdala i l’escorça prefrontal s’alinea amb les addiccions a substàncies. Es creu que una connectivitat més deficient disminueix el control de l'escorça prefrontal sobre l'impuls d'un usuari de participar en un comportament addictiu.
Cerca de traduccions #2): "Augment del volum d'amígdala" (que significa més matèria grisa). La majoria d’estudis d’addicció a les drogues informen d’una amígdala més petita en addictes (menys substància grisa). Aquest estudi suggereix que la toxicitat de les drogues pot conduir a menys matèria grisa i, per tant, reduir el volum d’amígdala en drogodependents. Sens dubte, això té un paper. Cal tenir en compte que l’amígdala és constantment activa durant la visualització de porno, especialment durant l’exposició inicial a una indicació sexual. Per exemple, si feu clic de pestanya en pestanya o cerqueu un vídeo o una imatge, s’il·luminarà l’amígdala. Potser la constant sexual la novetat i la recerca i la recerca condueixen a un efecte únic sobre l'amígdala en els usuaris de pornografia compulsiva.
Explicació alternativa per augmentar el volum d’amígdala en addictes a la pornografia: Els anys d’ús compulsiu de porno poden certament ser un factor d’estrès. A més, aquests subjectes de CSB no només eren addictes a la pornografia; també van experimentar greus conseqüències negatives com a resultat de l’ús de porno (pèrdua de feina, problemes de relació, desenvolupament d’ED induïda per pornografia). Aquí teniu un punt clau: L'estrès social crònic està relacionat amb l'augment del volum de l'amígdala:
Tot i que els mecanismes precisos de la plasticitat encara no estan plenament entesos, L’estrès moderat a sever sembla augmentar el creixement de diversos sectors de l’amígdala, mentre que els efectes en l’hipocamp i l’escorça prefrontal tendeixen a ser oposats.
Considerem la troballa anterior a la llum d’aquest fet Estudi del 2015 que va trobar que els "addictes al sexe" tenen un eix HPA hiperactiu (un sistema d’estrès hiperactiu). L’estrès crònic relacionat amb l’addicció al sexe / sexe, juntament amb factors que fan que el sexe sigui únic, podria provocar un major volum d’amígdala? Finalment, el volum d’amígdala més baix pot ser una condició preexistent als alcohòlics, igual que la descendència en famílies amb alt risc d'alcoholisme té una amígdala menor.
ENLLAÇ A L'ESTUDI COMPLET
Casper Schmidt,1,2,3 Laurel S. Morris,1,4 Timo L. Kvamme,1,2,3 Paula Hall,5 Thaddeus Birchard,5 i Valerie Voon1,4,6 *
- Publicat per primera vegada: 27 October 2016 Història completa de publicacions
- DOI: 10.1002 / hbm.23447 Veure / desar la citació
Els autors declaren que no tenen cap conflicte d'interessos per declarar.
abstracte
Fons
Els comportaments sexuals compulsius (CSB) són relativament comuns i estan associats amb una disfunció personal i social significativa. La neurobiologia subjacent encara no està ben entesa. El present estudi examina els volums cerebrals i la connectivitat funcional estatal en repòs en CSB en comparació amb voluntaris sanes coincidents (HV).
Mètodes
Les dades de RM estructurals (MPRAGE) es van recollir en subjectes de 92 (mascles 23 CSB i HV masculins coincidents 69) i analitzats mitjançant morfometria basada en voxel. Les dades de MRI funcional de l'estat de repòs utilitzant una seqüència plana de multi-ressò i un anàlisi de components independents (ME-ICA) es van recollir en subjectes 68 (subjectes 23 CSB i HV de parella 45).
Resultats
Els subjectes CSB van mostrar majors volums de matèria grisa d'amígdala esquerra (es va corregir un petit volum, s'ajustava Bonferroni P <0.01) i connectivitat funcional en estat de repòs reduït entre la llavor d’amígdala esquerra i l’escorça prefrontal dorsolateral bilateral (cervell sencer, FWE corregit per grups) P <0.05) en comparació amb l’HV.
Conclusions
El CSB s'associa amb volums elevats a les regions límbiques rellevants per a la motivació i el processament de les emocions, i la connectivitat funcional alterada entre les regions reguladores i límbiques prefrontals. Els estudis futurs haurien de tenir com a objectiu avaluar mesures longitudinals per investigar si aquests resultats són factors de risc que s’han anat abans de l’aparició dels comportaments o són conseqüències dels comportaments.
Abreviatures
- ACC del còrtex cingulat anterior
- Comportament sexual compulsiu CSB
- Líquid cefaloraquidi del LCR
- Còrtex prefrontal dorsolateral DLPFC
- Matèria grisa GM
- Model lineal general GLM
- Voluntaris saludables HV
- Magnetització MPRAGE preparada amb degradat de ressò
- Escorça orbitofrontal OFC
- Regió del ROI d'interès
- Mapatge paramètric estadístic SPM
- TR temps de repetició
- Temps de ressò de TE
- Morfometria basada en voxel de VBM
- WM white matter.
INTRODUCCIÓ
El comportament sexual compulsiu (CSB), també conegut com a trastorn hipersexual o addicció sexual, és relativament freqüent (estimat en 3% –6%) [Kraus et al., 2016] i associats a problemes significatius de trastorn i psicosocial, incloent-hi la caracterització per desitjos, impulsivitat i deteriorament social i laboral [Kraus et al., 2016]. Estudis recents s'han centrat en entendre els correlats neurobiològics subjacents [Kraus et al., 2016] encara que l’escassetat d’estudis limita la nostra comprensió dels mecanismes subjacents i com podríem conceptualitzar aquests trastorns. El CSB ha estat conceptualitzat com un trastorn de control d’impuls o com una addicció conductual [Kraus et al., 2016]. No obstant això, encara que es van proposar criteris per al trastorn hipersexual per al DSM-5 i validats en la prova de camp [Reid et al., 2012], aquest trastorn, juntament amb l’ús patològic d’Internet o de videojocs, no s’inclouen a la secció principal del DSM-5, en part a causa de les dades limitades sobre les condicions. Per tant, es necessiten més estudis sobre CSB per desenvolupar una major comprensió d’aquests trastorns. Tot i que el CSB pot tenir una sèrie de conductes, ens centrem en un grup que informa sobre les dificultats predominants amb l’ús de pornografia compulsiva. Hem utilitzat el terme CSB a partir del supòsit que "compulsiu" descriu la fenomenologia repetitiva i no pretén implicar supòsits mecanístics o etiològics.
Es va realitzar una revisió de la literatura sobre addiccions de comportament mitjançant la morfometria basada en voxel (VBM) o el gruix cortical. Hem utilitzat les següents paraules de cerca a PubMed (http://www.ncbi.nlm.nih.gov/pubmed): '[("Morfometria basada en voxel" o "gruix cortical") i] ", seguit de" [joc patològic] "," [addicció a Internet] "," [trastorn a Internet] "," [addicció al joc] ". En total, es van trobar 13 estudis dins de les addiccions conductuals relacionades amb els jocs d’atzar, l’ús d’Internet o els videojocs que avaluaven el VBM o el gruix cortical. La revisió de la literatura es presenta a la taula 1 i es discuteix a continuació.
Taula 1. Revisió bibliogràfica d’estudis volumètrics i de gruix cortical sobre addiccions conductuals
títol | Addicció al comportament | Temes (P / HV) | Measure | Regions implicades |
|---|---|---|---|---|
| ||||
| [Grant et al., 2015] | Trastorn del joc | 16/17 | Espessor cortical | Disminució del gruix cortical en r-SFC, RMFC, MOFC, PCG i bl-IPC |
| [Joutsa et al., 2011] | Jocs patològics | 12/12 | Morfometria basada en voxel | No hi ha diferències volumètriques en GM o WM entre HV i pacients |
| [Koehler et al., 2013] | Jocs patològics | 20/21 | Morfometria basada en voxel | Augment del volum de GM en bl-VS i r-PFC |
| [van Holst et al., 2012] | Problemes en el joc | 40/54 | Morfometria basada en voxel | No hi ha diferències volumèriques en GM o WM entre els jugadors problemàtics i la VH |
| [Hong et al., 2013] | Addicció a Internet | 15/15 | Espessor cortical | Disminució del gruix cortical en r-LOFC |
| [Yuan et al., 2011] | Addicció a Internet | 18/18 | Morfometria basada en voxel | Disminució del volum de GM en DLPFC, SMA, OFC, CB, RACC |
| [Zhou et al., 2011] | Addicció a Internet | 15/18 | Morfometria basada en voxel | Disminució de la densitat de GM a L-ACC, PCC, IC, LING |
| [Lin et al., 2014] | Addicció als jocs d'Internet | 35/36 | Morfometria basada en voxel | Disminució de la densitat de GM a l'IFG, l-CG, IC i r-HIPP Disminució de la densitat de WM a IFG, IC, IPC, ACC |
| [Sun et al., 2014] | Addicció als jocs d'Internet | 18/21 | Morfometria basada en voxel | Augment del volum de GM al r-ITG, MTG, PHG Disminució del volum GM en l-PrCG |
| [Wang et al., 2015] | Trastorn de jocs d'Internet | 28/28 | Morfometria basada en voxel | Disminució del volum de GM en ACC, PCUN, SMA, SPC i l-DLPFC, IC, CB |
| [Cai et al., 2015] | Trastorn de jocs d'Internet | 27/30 | Volum subcortical, FreeSurfer | Augment de volums de CN i VS |
| [Weng et al., 2013] | Addicció a jocs en línia | 17/17 | Morfometria basada en voxel | Disminució del volum GM en r-OFC, SMA i bl-IC |
| [Yuan et al., 2013] | Addicció a jocs en línia | 18/18 | Espessor cortical | Augment del gruix cortical en l-PrCG, PCUN, MFC, ITG, MTG Disminució del gruix cortical en l-LOFC, IC, r-PCG, IPC |
La comprensió de les alteracions neuronals en les addiccions prové dels estudis sobre els trastorns de l’ús de substàncies (SUD). Els individus amb SUD mostren disminució del volum i del gruix cerebral cortical, especialment a les regions corticals prefrontals que sotmeten un control conductual flexible. Un recent metanàlisi d’estudis de 9 i individus dependents d’alcohol 296 van trobar volums de matèria grisa prefrontal (GM) reduïts significativament, incloent l’escorça cingulada anterior (ACC) [Xiao et al., 2015], amb el volum GM cortical frontal associat negativament a l’ús d’alcohol de la vida [Taki et al., 2006]. Els volums GM frontals es van reduir de manera similar en individus dependents de cocaïna, incloent-hi cortex orbitofrontal (OFC) [Rando et al., 2013; Tanabe et al., 2009], escorça prefrontal anterior [Rando et al., 2013] i ACC [Connolly et al., 2013], aquest últim associat amb anys d’ús de drogues [Connolly et al., 2013].
Les diferències de grup en volums i gruixos corticals han estat menys clars en les addiccions conductuals (revisades a la Taula 1). Tres petits estudis sobre trastorns del joc van mostrar troballes inconsistents o bé amb disminució del gruix cortical en múltiples regions prefrontals i parietals [Grant et al., 2015], va augmentar els volums a l'escorça prefrontal dreta [Koehler et al., 2013] o cap diferència de grup [Joutsa et al., 2011]. En un gran estudi de jugadors amb problemes menys greus, no es van observar diferències de grup en els volums del cervell [van Holst et al., 2012]. Un petit estudi sobre l'addicció a Internet va mostrar un menor gruix cortical a l'OFC [Hong et al., 2013], amb un altre informant de menor volum en l’escorça prefrontal dorsolateral (DLPFC) [Yuan et al., 2011] i dos estudis que suggereixen un volum més baix d’ACC [Yuan et al., 2011; Zhou et al., 2011]. Dos petits estudis sobre trastorns de joc a Internet van informar de la disminució de volums en els OFC [Weng et al., 2013; Yuan et al., 2013], i dos estudis més grans van reportar volums més reduïts a l'escorça cingulada [Lin et al., 2014; Wang et al., 2015] amb estudis individuals que informen de disminucions en DLPFC [Wang et al., 2015], frontal inferior [Lin et al., 2014], superior parietal [Wang et al., 2015] i parietal inferior [Yuan et al., 2013] cortices. Pel que fa a les estructures subcorticals, un petit estudi va informar de volums estriatals (VS) més elevats en el trastorn del joc [Koehler et al., 2013] sense diferències subcorticals reportades en els altres estudis. En el trastorn de jocs d'Internet, les troballes eren de manera semblant incompatibles amb un nombre més gran de parahipopocamps [Sun et al., 2014], hipocamp inferior [Lin et al., 2014] o no hi ha diferències [Wang et al., 2015; Weng et al., 2013]. Un estudi amb una mida de mostra raonable centrat en els volums subcorticals va informar de majors quantitats de caudats i SV associats amb dèficits de control cognitiu [Cai et al., 2015]. En conjunt, les troballes d’anormalitats corticals o subcorticals en el trastorn del joc són molt inconsistents. En canvi, els informes sobre anomalies corticals en l’ús d’Internet o en els jocs d'Internet de manera més consistent informen de volums reduïts amb un volum de CAC i OFC reduït que es va repetir en almenys dos estudis.
Fins ara, hi ha evidències escasses de canvis neurals estructurals en individus amb CSB. Els estudis d’individus sans amb un ús pornogràfic excessiu sense el diagnòstic de CSB mostren volums menors de GM al caudat dret [Kühn i Gallinat, 2014]. Un petit estudi de RM de difusió de individus amb CSB (N = 8 per grup) va mostrar una difusivitat mitjana reduïda en tractaments de matèria blanca frontal superior (WM) en comparació amb HV [Miner et al., 2009]. Pel que fa a l’activitat funcional, l’HV masculí mostra processos d’habituació millorats amb l’activitat BOLD putaminal inferior esquerra a les imatges eròtiques estàtiques [Kühn i Gallinat, 2014] i redueix el potencial positiu tardà a imatges explícites [Prause et al., 2015]. En contrast, en un estudi fMRI basat en tasques que compara CSB amb HV, els vídeos sexuals explícits van provocar respostes més altes de VS, amígdala i dorsal ACC BOLD en CSB [Voon et al., 2014]. La connectivitat funcional entre aquestes regions es correlacionava amb un índex de desig sexual o "desitjant", però no "gust" en subjectes CSB que suggereixen el paper de la motivació d'incentius, paral·lel a l'addicció a substàncies. De la mateixa manera, en un altre estudi sobre l’addicció a la pornografia a Internet, la imatge sexual preferida estava associada a una major activitat estriatal ventral i només es correlacionava amb símptomes autoinformats de l’addicció a la pornografia a Internet i no amb altres mesures de comportament sexual o depressió [Brand et al., 2016]. Un altre estudi recent mostra també que els individus amb un comportament hipersexual problemàtic experimenten un desig sexual més freqüent i millorat durant l’exposició a estímuls sexuals i que s’observà una major activació al caudat, el lòbul parietal inferior, al giro cingulat anterior dorsal, al thalamus i al DLPFC en aquest grup [Seok i Sohn, 2015]. Els individus CSB mostren més biaix atencional primerenc a estímuls sexuals explícits [Mechelmans et al., 2014] que es correlacionava amb les preferències d’elecció per a senyals condicionades a les imatges sexuals [Banca et al., 2016]. Com a resposta a l’exposició repetida d’imatges eròtiques estàtiques, els subjectes CSB van mostrar una major habituació a l’ACC dorsal als resultats sexuals, que es correlacionaven amb les preferències d’elecció per a noves imatges sexuals [Banca et al., 2016], un efecte que es pot explicar per qualsevol de les habituacions, però també pot ser coherent amb el concepte de tolerància a les addiccions.
L’estudi actual examina el GM volumètric de CSB i revisa la literatura actual sobre estudis de gruix volumètric i cortical en trastorns d’atzar i en trastorns d’ús d’internet i de joc. També analitzem la connectivitat funcional de l'estat en repòs d’individus amb CSB i l’HV coincident amb una nova seqüència plana multi-ressò i anàlisi de components independents (ME-ICA) on els senyals BOLD s’identifiquen com a components independents amb canvi de senyal dependent del temps d'eco (TE) mentre que els senyals no BOLD s'identifiquen com a components independents del TE [Kundu et al., 2012]. Esperem que hi hagi una xarxa interrompuda de sistemes relacionats amb la salut i la recompensa subordinats a l’amígdala, al VS i al ACC dorsal.
MÈTODES
Participants
Els subjectes CSB van ser reclutats a través d’anuncis basats en Internet i de referències de terapeutes. Els HV masculins coincidents amb l'edat van ser reclutats a partir d'anuncis basats en la comunitat a la zona de East Anglia. Tots els subjectes del CSB van ser entrevistats per un psiquiatre per confirmar que complien els criteris diagnòstics del CSB (complien els criteris diagnòstics proposats tant per al trastorn hipersexual [Kafka, 2010; Reid et al., 2012] i l'addicció sexual [Carnes et al., 2007], centrant-se en l'ús compulsiu de material en línia sexualment explícit. Es va avaluar mitjançant una versió modificada de l’Arena de l’experiència sexual d'Arizona (ASES) [Mcgahuey et al., 2011], en què es van respondre preguntes a una escala de 1-8, amb puntuacions més altes que representaven un major deteriorament subjectiu. Donada la naturalesa de les indicacions, tots els subjectes de CSB i HV eren homes i heterosexuals. Tots els HV es van emparejar per edat (± 5 anys d'edat) amb assignatures de CSB. També es van examinar els subjectes per compatibilitzar-los amb l'entorn MRI tal com ho hem fet prèviament [Banca et al., 2016; Mechelmans et al., 2014; Voon et al., 2014]. Els criteris d'exclusió eren menors de 18 anys d'edat, tenint una història de SUD, sent un usuari habitual actual de substàncies il · lícites (inclòs el cannabis) i tenien un trastorn psiquiàtric greu, incloent la depressió major o moderada greu o el trastorn obsessiu-compulsiu, o història del trastorn bipolar o esquizofrènia (esgrafiat amb el Mini Inventari Internacional Neuropsiquiàtric) [Sheehan et al., 1998]. Altres adiccions compulsives o conductuals també eren exclusions. Els subjectes van ser avaluats per un psiquiatre pel que fa a l'ús problemàtic dels jocs en línia o de les xarxes socials, el joc patològic o el comportament compulsiu, el trastorn d'hiperactivitat amb dèficit d'atenció infantil o d'adults i el diagnòstic del trastorn alimentari. Els subjectes van completar l'escala de comportament impulsiu UPPS-P [Whiteside i Lynam, 2001] per avaluar la impulsivitat i l'inventari de depressió de Beck [Beck et al., 1961] per avaluar la depressió. Dues de les assignatures de 23 CSB prenien antidepressius o havien comorbitat trastorn d'ansietat generalitzada i fòbia social (N = 2) o fòbia social (N = 1) o una història infantil de TDAH (N = 1). Es va obtenir el consentiment informat per escrit i l’estudi va ser aprovat pel Comitè d’ètica de la investigació de la Universitat de Cambridge. Es pagava als subjectes per la seva participació.
Neuroimatge
Adquisició i processament de dades
Estructural.
Les imatges estructurals es van recollir incloent el gradient-eco (MPRAGE) preparat per magnetització completa mitjançant un escàner Siemens Tim Trio 3T amb una bobina de capçal de canal 32 utilitzant una seqüència MPRAGE ponderada T1 (retalls sagitals 176, temps de repetició (TR) = 9 ms; temps d'eco (TE) = 2,500 ms; temps d'inversió = 4.77 ms; matriu d'adquisició = 1,100 × 256 × 256; angle de volteig = 176 °; mida de voxel 7 × 1 × 1 mm). L’exploració es va dur a terme al Centre de creació de cervells Wolfson de la Universitat de Cambridge.
Les dades estructurals es van processar amb mapatge paramètric estadístic (SPM8; http://www.fil.ion.ucl.ac.uk/spm) (Wellcome Trust Center for Neuroimaging, Londres, Regne Unit). Les imatges anatòmiques es van reorientar manualment, situant l’origen a la comissura anterior. Les imatges es van segmentar (utilitzant el nou segment per a SPM) en líquid GM, WM i líquid cefaloraquidi (LCR) basat en mapes de probabilitat de teixit estàndard per a cada tipus de teixit. Els tres volums de classes de teixits es van resumir per produir un volum intracranial total estimat. Es va crear una plantilla personalitzada amb DARTEL [Ashburner, 2007], que defineix els paràmetres necessaris per ajustar la imatge GM nativa de cada individu a un espai comú, de manera iterativa. Aquesta plantilla DARTEL es va registrar després als mapes de probabilitat de teixits amb transformacions afines, portant imatges a l’espai MNI. Les imatges es van suavitzar espacialment amb una amplada completa a la meitat del nucli màxim de 8 mm3.
Estat de descans.
Les dades fMRI de l'estat de repòs es van adquirir per a minuts 10 amb els ulls oberts amb un escàner Siemens 3T Tim Trio amb una bobina de capçal de canal 32 al Wolfson Brain Imaging Center de la Universitat de Cambridge. Es va utilitzar una seqüència d’imatge plana de ressò multi-ressò amb la reconstrucció en línia (temps de repetició, 2.47 s; angle de xip, 78 °; mida de la matriu 64 × 64; resolució en pla, 3.75 mm; FOV, 240 mm; rodones obliques 32, altern gruix de la secció d’adquisició de llesques 3.75 mm amb un gap 10%; factor iPAT, 3; ample de banda = 1,698 Hz / píxel; temps d’eco (TE) = 12, 28, 44 i 60 ms).
Anàlisi de components independent de diversos ecos (ME-ICAv2.5 beta6; http://afni.nimh.nih.gov) s’utilitza per a l’anàlisi i el desconnexió de les dades fMRI de l’estat de repòs multi-ressò. ME-ICA descompon dades de fMRI multi-ressò en components independents amb FastICA. El canvi de senyal de percentatge de senyal BOLD depèn linealment de TE, característica de la desintegració de T2 *. Aquesta dependència de TE es mesura utilitzant el pseudo-F-estadística, kappa, amb components que s'escalen fortament amb TE amb altes puntuacions kappa [Kundu et al., 2012]. Els components que no són BOLD s'identifiquen per la independència TE mesurada pel pseudo-F-estadística, rho. Així, els components es classifiquen com BOLD o no BOLD basant-se en les seves ponderacions de valor kappa i rho, respectivament [Kundu et al., 2012]. Els components que no són BOLD s’eliminen mitjançant dades de projecció, sense soroll per a artefactes de moviment, fisiològics i d’escàner d’una manera robusta basada en principis físics. Les imatges eco-planes de les persones sense soroll es van registrar al seu MPRAGE i es van normalitzar a la plantilla de l'Institut Neurològic de Montreal (MNI). L'allisament espacial es va dur a terme amb un nucli gaussià (amplada màxima de la meitat màxima = 6 mm). El recorregut temporal de cada voxel es va filtrar temporalment per banda de banda (0.008 f <0.09 Hz). L'exploració anatòmica de cada individu es va segmentar en GM, WM i CSF. Es van eliminar els components principals significatius dels senyals de WM i CSF.
L’anàlisi de connectivitat funcional es va realitzar utilitzant un enfocament de regió d’interès (ROI) amb la caixa d’eines de connectivitat funcional CONN-fMRI [Whitfield-Gabrieli i Nieto-Castanon, 2012] per a SPM (http://www.fil.ion.ucl.ac.uk/spm/software/spm8/).
Anàlisi estadística
Les característiques dels subjectes i les puntuacions del qüestionari es van comparar entre grups amb dues cues t-test sense assumir la varianza igual. Totes les anàlisis estadístiques es van realitzar utilitzant la versió R (3.2.0) [Equip RC, 2014].
estructural
Per a les comparacions de grups, els volums GM de subjectes CSB i HV es van introduir en un model lineal general (GLM). Les dades es van corregir per al volum intracraneal total dels participants mitjançant una escala proporcional i una màscara explícita a SPM. Les comparacions de grups es van ajustar tant per puntuacions d’edat com de depressió com a covariables. Ens vam centrar en a priori hipòtesis de les regions d’interès identificades al nostre estudi anterior [Voon et al., 2014] i en meta-anàlisis d'estudis de reactivitat de la llet de drogues [Kühn i Gallinat, 2011], és a dir, VS esquerra i dreta, amígdala dreta i esquerra, i ACC dorsal amb error familiar correcte corregit (SVC) petit (FWE) corregit P <0.01 (Bonferroni es corregeix per comparacions múltiples). Per a aquestes anàlisis de SVC, hem utilitzat un ROI anatòmic VS, descrit anteriorment [Murray et al., 2008] que es va dibuixar a mà amb MRIcro basant-se en la definició de VS per Martinez et al. [2003]. El ROI d’amígdala s’ha obtingut de l’atlas d’etiquetatge anatòmic automatitzat (AAL). L’ACC dorsal es va modificar manualment mitjançant la caixa d’eines del ROI de MarsBaR [Brett et al., 2002] i es basa en el ROI cingulat de l’escorça de l’atles de l’AAL. Va ser modificat de manera que la vora anterior era la punta de la genu del cos callós [Cox et al., 2014; Desikan et al., 2006] i el posterior era l'extrem posterior de la genu del cos callós [Desikan et al., 2006]. Es van realitzar anàlisis addicionals per ajustar les puntuacions BDI.
Estat de descans
Per comparar la connectivitat entre subjectes CSB i HV, es van calcular els mapes de connectivitat del cervell sencer ROI-a-voxel per a la regió d’interès de la llavor d’amígdala esquerra basant-se en les troballes volumètriques de la diferència de grup. Els mapes de connectivitat resultants es van introduir en GLM factorials complets per comparar la connectivitat de tot el cervell entre grups que s’ajustaven per edat amb una anàlisi posterior ajustant tant per edat com per depressió. Tot el cúmul de cervells va corregir FWE P <0.05 es va considerar significatiu per a diferències de grup.
RESULTATS
Characteristics
Hi van participar vint-i-tres homes heterosexuals amb CSB (edat 26.9; SD 6.22 anys) i 69 igualats (edat 25.6; SD 6.55 anys) homes heterosexuals. 2), dels quals els subjectes de 19 CSB i 55 HV van completar els qüestionaris de comportament. Els subjectes CSB tenien BDI més elevada (P = 0.006) i UPPS-P (P <0.001) puntuacions comparades amb HV. Altres puntuacions de comportament, inclosos els patrons i la gravetat de la pornografia i l'ús d'Internet, s'han informat en altres llocs [Mechelmans et al., 2014; Voon et al., 2014].
Taula 2. Dades demogràfiques i conductuals per a subjectes de conducta sexual compulsiva i voluntaris sans
Group | edat | BDI | UPPS-P |
|---|---|---|---|
Els participants de 4 falten de 23.
Els participants de 14 falten de 69.
| |||
| CSB (N = 23) | 26.9 (6.22) | 14.82 (11.85)a | 152.21 (16.50)a |
| HV (N = 69) | 25.6 (6.55) | 6.03 (7.20)b | 124.87 (20.73)b |
| T-valor (P-valor) | 0.88 (P = 0.380) | 3.04 (P = 0.006) | 5.81 (P <0.001) |
estructural
Els anàlisis del ROI de l’amígdala dreta i esquerra, VS esquerra i dreta i ACC dorsal van revelar que el volum de matèria grisa de l’amígdala esquerra es va incrementar en CSB en comparació amb el VH coincident (corregit per SVC FWE, P = 0.0096, Z = 3.37, xyz = −28, −4, −15) (Bonferroni corregit per SVC corregit per FWE P <0.01) (Fig. 1). Totes les altres anàlisis del ROI no eren significatives. L’ajustament de la depressió no va canviar els resultats de les diferències de grup.

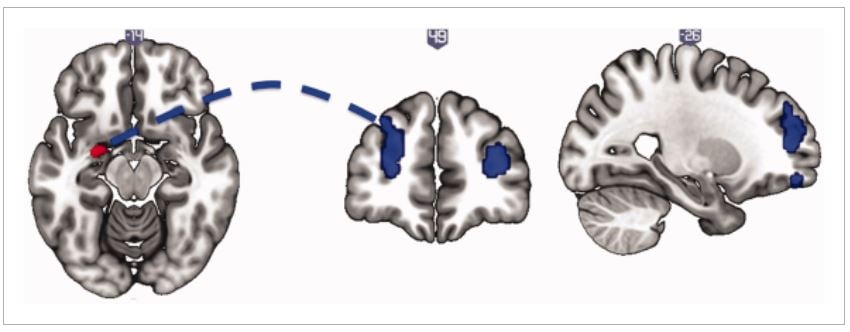
Figura 1.
Morfometria basada en Voxel en comportaments sexuals compulsius. El major volum de l'amígdala esquerra es mostra en comportaments sexuals compulsius en relació amb voluntaris sans. La imatge s’abandona a P <0.005 sense corregir per il·lustrar. [Es pot veure la figura de color a wileyonlinelibrary.com]
Estat de descans
Basant-nos en els resultats estructurals, vam examinar la connectivitat funcional de l'estat de repòs amb una llavor a l'amígdala esquerra. Hem trobat una connectivitat reduïda amb DLPFC bilateral (DLPFC dreta: P = 0.012, Z = 4.11, xyz = 31 42 16; DLPFC esquerre: P = 0.003, Z = 3.96, xyz = −27 52 23) (Fig. 2). L’ajustament de BDI no va alterar la importància de les troballes (Right DLPFC: P = 0.001, Z = 4.54, xyz = 31 61 23; DLPFC esquerre: P = 0.003, Z = 4.26, xyz = −29 49 35).

Figura 2.
Connectivitat funcional de l'estat de repòs de l’amígdala esquerra. El comportament sexual compulsiu està associat a la disminució de la connectivitat funcional de l'estat de repòs de l'amígdala esquerra (llavor, esquerra) amb l'escorça prefrontal dorsolateral bilateral (mitjana i dreta), en relació amb voluntaris sans. La imatge s’abandona a P <0.005 sense corregir per il·lustrar. [Es pot veure la figura de color a wileyonlinelibrary.com]
DISCUSSIÓ
Es van investigar les diferències neurals estructurals i funcionals en individus amb CSB en comparació amb el VH coincident. Els subjectes CSB van augmentar el volum de l'amígdala esquerra i la connectivitat funcional reduïda durant el descans entre l'amígdala esquerra i la DLPFC bilateral.
L’amígdala està implicada en el processament de la salut ambiental que dirigeix el comportament. Els nuclis de l’amígdala vinculen estímuls ambientals o interns prèviament neutres amb representacions associatives de valor afectiu, que propagen la prudència motivacional induïda per la cue [Everitt et al., 2003], així com el processament del control emocional [Cardinal et al., 2002; Gottfried et al., 2003]. El descobriment d’un augment del volum d’amígdala s’oposa amb diversos estudis sobre els trastorns d’ús d’alcohol [Makris et al 2008; Wrase et al., 2008], ja que els estudis sobre aquest tipus d’addicció reporten una disminució dels volums d’amígdala, on s’han avaluat mesures volumètriques. Una possible explicació d'aquesta discrepància és que l’ús de substàncies a llarg termini provoca canvis neuroplàstics de llarga durada i toxicitat [Kovacic, 2005; Reissner i Kalivas, 2010] que puguin contribuir a la perseverança del comportament de la recerca de drogues [Gass and Olive, 2008]. Aquesta neurotoxicitat pot contribuir sens dubte a l’atrofia generalitzada observada en les addiccions de substàncies [Bartzokis et al., 2000; Carlen et al., 1978; Mechtcheriakov et al., 2007]. Aquesta neurotoxicitat relacionada amb els fàrmacs és probablement un tema molt rellevant en el SUD, però menys en un problema en les addiccions conductuals. En un recent estudi CSB que utilitza fMRI, l’exposició a indicis sexualment explícits en CSB en comparació amb subjectes no CSB es va associar amb l’activació de l’amígdala [Voon et al., 2014]. Queda pendent de determinar si la diferència en el volum d’amígdala és un tret preexistent que predisposa els individus al CSB o relacionat amb l’exposició excessiva.
El funcionament de la DLPFC és conegut per estar associat a aspectes amplis del control cognitiu [MacDonald et al., 2000] i memòria de treball [Petrides, 2000]. El nostre descobriment de la disminució de la connectivitat funcional entre l'amígdala i la DLPFC convergeix amb la literatura existent sobre la connectivitat en aquestes regions. Aquesta connectivitat funcional és important per a la regulació de l'emoció, la qual cosa s'ha informat anteriorment de que la connectivitat reduïda entre l'amígdala i la DLPFC en persones amb trastorns de joc d'Internet està associada a nivells més alts d'impulsivitat [Ko et al., 2015]. Un altre estudi que mesura la capacitat de modular les respostes emocionals negatives mitjançant l’ús d’estratègies cognitives ha mostrat que l’activitat en àrees específiques de l’escorça frontal, inclosa la DLPFC, es covari amb l’activitat de l’amígdala i que la connectivitat funcional entre aquestes regions depenia d’aplicar estratègies cognitives regulació de l'emoció negativa [Banks et al., 2007]. La connectivitat d’amígdala i de DLPFC ha estat associada de manera similar a la depressió unipolar [Siegle et al., 2007]. El CSB s'ha associat amb símptomes depressius i d'ansietat i l'estrès pot provocar aquestes activitats; no obstant això, les nostres troballes no estaven relacionades amb les puntuacions de la depressió. La DLPFC també va estar implicada en un estudi de la VH masculina en la qual es va associar un major ús de pornografia a una menor connectivitat funcional entre el DLPFC i el striatum quan es visualitzaven imatges explícites [Kühn i Gallinat, 2014].
Advertim que aquestes troballes són preliminars tenint en compte la petita mida de la mostra de subjectes CSB, tot i que es compara aquest grup amb un gran tamany de mostra de la HV coincident. Una limitació de l’estudi és l’homogeneïtat de la població. Com no vam incloure subjectes amb altres trastorns psiquiàtrics comòrbids que podrien tenir un paper mecanicista, aquests resultats haurien d’extrapolar-se amb cautela a subjectes CSB amb altres comorbilitats. A més, les anomalies estructurals i funcionals observades entre els subjectes CSB poden estar relacionades amb trets preexistents o poden ser el resultat dels efectes de CSB i, per tant, aquest estudi no pot fer inferències causals sobre els efectes de CSB. Els estudis futurs haurien de tenir com a objectiu avaluar mesures longitudinals per determinar les diferències entre l'estat i les tendències de trets i les possibles anomalies neuronals pre-mòrdiques en mides de mostres més grans i amb sexes mixtos.
Els nostres resultats actuals posen de relleu els volums elevats en una regió implicada en la rellevància motivacional i una menor connectivitat en estat de repòs de les xarxes reguladores de prefrontal de dalt a baix. La interrupció d’aquestes xarxes pot explicar els patrons de comportament aberents cap a una recompensa o una reactivitat millorable ambientalment per a indicis d’incentius destacats. Tot i que els nostres resultats volumètrics contrasten amb els del SUD, aquestes troballes poden reflectir diferències en funció dels efectes neurooxics de l’exposició crònica a les drogues. Les proves emergents suggereixen superposicions potencials amb un procés d’addicció que recolzen particularment les teories de la motivació d’incentius. Hem demostrat que l’activitat d’aquesta xarxa de salut és millorada després de l’exposició a indicis sexualment explícits més destacats o preferents [Brand et al., 2016; Seok i Sohn, 2015; Voon et al., 2014], juntament amb un major biaix assistencial [Mechelmans et al., 2014] i el desig específic del cua sexual, però no el desig sexual generalitzat [Brand et al., 2016; Voon et al., 2014]. L'atenció millorada a les pautes sexualment explícites es relaciona més amb la preferència per les pautes sexualment condicionades, de manera que es confirma la relació entre el condicionament sexual i el biaix atent [Banca et al., 2016]. Aquestes troballes d'una activitat millorada relacionada amb les pautes condicionades sexualment difereixen de la del resultat (o de l'estímul sense condicionament) en què l'habituació millorada, possiblement coherent amb el concepte de tolerància, augmenta la preferència per nous estímuls sexuals [Banca et al., 2016]. Junts, aquests resultats ajuden a dilucidar la neurobiologia subjacent de CSB que condueix a una major comprensió del trastorn i la identificació de possibles marcadors terapèutics.
AGRAÏMENTS
Volem donar les gràcies al personal de WBIC per la seva experiència i assistència en la recopilació de dades d'imatges i els nostres participants pel seu temps i compromís. A més, volem agrair a Thaddeus Birchard i Paula Hall la remissió de pacients a l’estudi. El Behavioral and Clinical Neuroscience Institute (BCNI) compta amb el suport del Wellcome Trust i del Medical Research Council.
REFERÈNCIES
- Ashburner J (2007): algorisme de registre d'imatges difeomorfic ràpid. Neuroimatge 38: 95 – 113.
- Banca P, Morris LS, Mitchell S, Harrison NA, Potenza MN, Voon V (2016): novetat, condicionament i tendència atencional a les recompenses sexuals. J Psychiatr Res 72: 91 – 101.
- CrossRef |
- PubMed |
- Web of Science® Times citada: 2
- CrossRef |
- PubMed |
- Web of Science® Times citada: 307
- CrossRef |
- PubMed |
- Web of Science® Times citada: 75
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times citada: 18323
- CrossRef |
- PubMed |
- Web of Science® Times citada: 1
- Bancs SJ, Eddy KT, Angstadt M, Nathan PJ, Phan KL (2007): connectivitat amígdala-frontal durant la regulació emocional. Neurosci de Soc Cogn Affect 2: 303 – 312.
- CrossRef |
- Web of Science® Times citada: 1
- CrossRef |
- PubMed |
- Web of Science® Times citada: 1087
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times citada: 342 |
- ADS
- Bartzokis G, Beckson M, Lu PH, Edwards N, Rapoport R, Wiseman E, Bridge P (2000): reducció del volum del cervell relacionada amb l'edat en addictes a anfetamina i cocaïna i controls normals: implicacions en la investigació de l'addicció. Psiquiatria Res Neuroimatge 98: 93 – 102.
- CrossRef |
- PubMed |
- Web of Science® Times citada: 16 |
- ADS
- CrossRef |
- PubMed |
- Web of Science® Times citada: 7
- CrossRef |
- PubMed |
- Web of Science® Times citada: 1782
- Wiley Online Library |
- PubMed |
- Web of Science® Times citada: 245
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times citada: 217
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times citada: 597 |
- ADS
- CrossRef |
- PubMed |
- Web of Science®
- CrossRef |
- PubMed |
- Web of Science® Times citada: 19
- CrossRef |
- PubMed |
- Web of Science® Times citada: 27
- CrossRef |
- PubMed |
- Web of Science® Times citada: 21
- CrossRef |
- PubMed |
- Web of Science® Times citada: 172
- CrossRef |
- PubMed |
- Web of Science® Times citada: 8
- CrossRef |
- PubMed |
- Web of Science® Times citada: 5
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times citada: 30
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times citada: 1
- Wiley Online Library |
- PubMed |
- Web of Science® Times citada: 76
- CrossRef |
- PubMed |
- Web of Science® Times citada: 23
- CrossRef |
- PubMed |
- Web of Science® Times citada: 63
- Beck AT, Ward C, Mendelson M (1961): inventari de depressió Beck (BDI). Arch Gen Psychiatry 4: 561 – 571.
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times citada: 1895 |
- ADS
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times citada: 134
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times citada: 313
- Marca M, Snagowski J, Laier C, Maderwald S (2016): l'activitat estriat entral a l'hora de veure imatges pornogràfiques preferides es correlaciona amb els símptomes de l'addicció a la pornografia a Internet. Neuroimatge 129: 224 – 232.
- CrossRef |
- PubMed |
- Web of Science® Times citada: 7 |
- ADS
- CrossRef |
- PubMed |
- Web of Science® Times citada: 70
- CrossRef |
- PubMed |
- Web of Science® Times citada: 28
- CrossRef |
- CAS |
- Web of Science® Times citada: 196
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times citada: 255
- CrossRef |
- PubMed |
- Web of Science® Times citada: 3
- Wiley Online Library |
- PubMed |
- Web of Science® Times citada: 20
- Brett M, Anton JL, Valabregue R, Poline JB (2002): Anàlisi de la regió d'interès mitjançant la caixa d'eines MarsBar per a SPM 99. Neuroimatge 16: S497.
- Wiley Online Library |
- PubMed |
- Web of Science® Times citada: 43
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times citada: 63
- CrossRef |
- PubMed |
- Web of Science® Times citada: 1
- PubMed |
- Web of Science® Times citada: 7675
- CrossRef |
- PubMed |
- Web of Science® Times citada: 383
- CrossRef |
- PubMed |
- Web of Science® Times citada: 4
- Wiley Online Library |
- PubMed |
- Web of Science® Times citada: 38
- CrossRef |
- PubMed |
- Web of Science® Times citada: 110
- CrossRef |
- PubMed |
- Web of Science® Times citada: 25
- CrossRef |
- PubMed |
- Web of Science® Times citada: 3
- CrossRef |
- PubMed |
- Web of Science® Times citada: 25
- CrossRef |
- Web of Science® Times citada: 1108
- CrossRef |
- PubMed
- CrossRef |
- PubMed |
- Web of Science® Times citada: 92
- CrossRef |
- PubMed |
- Web of Science® Times citada: 3
- CrossRef |
- PubMed |
- Web of Science® Times citada: 72 |
- ADS
- CrossRef |
- PubMed |
- Web of Science® Times citada: 31 |
- ADS
- CrossRef |
- PubMed |
- Web of Science® Times citada: 66
- Cai C, Yuan K, Yin J, Feng D, Bi Y, Li Y, Yu D, Jin C, Qin W, Tian J (2015): La morfometria estriada s'associa amb els dèficits de control cognitiu i la severitat dels símptomes en el trastorn del joc a Internet. Comportament de la imatge cerebral 10: 12.
- Cardinal RN, JA Parkinson, Hall J, Everitt BJ (2002): Emoció i motivació: el paper de l’amígdala, l’estriat ventral i l’escorça prefrontal. Neurosci Biobehav Rev 26: 321 – 352.
- Carlen PL, Wortzman G, Holgate RC, Wilkinson DA, Rankin JC (1978): Atròfia cerebral reversible en alcohòlics crònics recentment abstinguts mesurats per tomografies computades. Ciència 200: 1076 – 1078.
- Carnes P, Delmonico DL, Griffin E (2007): A les ombres de la xarxa: lliure de comportament sexual compulsiu en línia, 2nd ed. Centre City, MN: Publishing Hazelden.
- Connolly CG, Bell RP, Foxe JJ, Garavan H (2013): canvis de matèria grisa dissociats amb addicció prolongada i abstenció en usuaris de cocaïna. PLoS One 8: e59645.
- Cox SR, Ferguson KJ, Royle NA, Shenkin SD, MacPherson SE, MacLullich AMJ, Deary IJ, Wardlaw JM (2014): Una revisió sistemàtica de tècniques de parcel·lació del lòbul frontal cerebral en ressonància magnètica. Brain Struct Funct 219: 1 – 22.
- Desikan RS, Ségonne F, Fischl B, Quinn BT, Dickerson BC, Blacker D, Buckner RL, Dale AM, Maguire RP, Hyman BT (2006): Un sistema d’etiquetatge automatitzat per subdividir l’escorça cerebral humana a les exploracions de ressonància magnètica a regions amb base gyral. d'interès. Neuroimatge 31: 968 – 980.
- Everitt BJ, Cardinal RN, JA Parkinson, TW Robbins (2003): Comportament apetitiu: impacte dels mecanismes d'aprenentatge emocional dependents de l'amígdala. Ann NY Acad Sci 985: 233-250.
- Gass JT, Olive MF (2008): substrats glutamatèrgics de drogodependència i alcoholisme. Biochem Pharmacol 75: 218 – 265.
- Gottfried JA, O'Doherty J, Dolan RJ (2003): Codificació del valor predictiu de la recompensa en l'amígdala humana i l'escorça orbitofrontal. Ciència 301: 1104-1107.
- Grant JE, Odlaug BL, Chamberlain SR (2015): Reducció del gruix cortical en el trastorn del joc: un estudi de RM morfomètric. Eur Arch Psychiatry Clin Neurosci 265: 655 – 661.
- Van Holst RJ, de Ruiter MB, van den Brink W, Veltman, Goudriaan AE (2012): un estudi de morfometria basat en voxel que compara jugadors problemàtics, consumidors d'alcohol i controls saludables. Depèn de l’alcohol de drogues 124: 142 – 148.
- Hong SB, Kim JW, Choi EJ, Kim HH, Suh JE, Kim CD, Klauser P, Whittle S, Yűcel M, Pantelis C, Yi SH (2013): reducció del gruix de la cortical orbitofrontal en adolescents masculins amb addicció a Internet. Behav Brain Funct 9: 11.
- Joutsa J, Saunavaara J, Parkkola R, Niemelä S, Kaasinen V (2011): Anomalia extensa de la integritat de la matèria blanca del cervell en el joc patològic. Psychiatry Res - Neuroimaging 194: 340-346.
- Kafka MP (2010): Trastorn hipersexual: un diagnòstic proposat per DSM-V. Arch Sex Behav 39: 377 – 400.
- Ko CH, Hsieh TJ, Wang PW, Lin WC, Yen CF, Chen CS, Yen JY (2015): alteració de la densitat de matèria grisa i interrupció de la connectivitat funcional de l'amígdala en adults amb trastorns de joc a Internet. Prog Neuropsychopharmacol Biol Psychiatry 57: 185 – 192.
- Koehler S, Hasselmann E, Wüstenberg T, Heinz A, Romanczuk-Seiferth N (2013): Volum més elevat de l'estri ventral i l'escorça prefrontal dreta en jocs d'atzar patològics. Brain Struct Funct 220: 469 – 477.
- Kovacic P (2005): mecanisme unificador per a l'addicció i la toxicitat de drogues maltractades amb aplicació a mediadors de dopamina i glutamat: transferència d'electrons i espècies reactives d'oxigen. Hypotheses Med 65: 90 – 96.
- Kraus SW, Voon V, Potenza MN (2016): Neurobiologia del comportament sexual compulsiu: ciències emergents. Neuropsicofarmacologia 41: 385 – 386.
- Kühn S, Gallinat J (2011): biologia comuna del desig a través de drogues legals i il·legals: una meta-anàlisi quantitativa de la resposta del cervell a la reactivitat. Eur J Neurosci 33: 1318 – 1326.
- Kühn S, Gallinat J (2014): Estructura del cervell i connectivitat funcional associada al consum de pornografia: el cervell sobre la pornografia. JAMA Psychiatry 71: 827 – 834.
- Kundu P, Inati SJ, Evans JW, Luh WM, Bandettini PA (2012): Diferenciació dels senyals BOLD i no BOLD en sèries temporals fMRI mitjançant EPI multi-eco. Neuroimatge 60: 1759 – 1770.
- Lin X, Dong G, Wang Q, Du X (2014): volum anormal de matèria grisa i de matèria blanca en "addictes als jocs d'Internet". Addict Behav 40C: 137 – 143.
- MacDonald AW, JD Cohen, VA Stenger, Carter CS (2000): Dissociant el paper de l'escorça prefrontal dorsolateral i cingulada anterior en control cognitiu. Ciència 288: 1835 – 1838.
- Makris N, Oscar-Berman M, Jaffin SK, Hodge SM, Kennedy DN, Caviness VS, Marinkovic K, Breiter HC, Gasic GP, Harris GJ (2008): Disminució del volum del sistema de recompensa del cervell en l'alcoholisme. Biol Psychiatry 64: 192 – 202.
- Martinez D, Slifstein M, Broft A, Mawlawi O, Chatterjee R, Hwang DR, Huang Y, Cooper T, Kegeles L, Zarahn E, Abi-Dargham A, Haber SN, Laruelle M (2003): Imatge de transmissió de dopamina mesolímbica humana amb tomografia per emissió de positrons. Part II: Alliberament de dopamina induïda per l'anfetamina a les subdivisions funcionals de l'estriat. J Cereb Blood Flow Metab 23: 285 – 300.
- Mcgahuey CA, Gelenberg AJ, Cindi A, Moreno FA, Delgado PL, Mcknight KM, Manber R (2011): Journal of sex & marital the arizona sexual experience scale (asex): fiabilitat i validesa. J Sex Marital Ther 26: 37-41.
- Mechelmans DJ, Irvine M, Banca P, Porter L, Mitchell S, Mole TB, Lapa TR, Harrison NA, Potenza MN, Voon V (2014): S'ha millorat el biaix atenció cap a senyals sexualment explícites en individus amb i sense comportaments sexuals compulsius. PLoS One 9: e105476.
- Mechtcheriakov S, Brenneis C, Egger K, Koppelstaetter F, Schocke M, Marksteiner J (2007): Un patró diferent de l'atròfia cerebral estès en pacients amb addicció a l'alcohol revelat per morfometria basada en voxel. J Neurol Neurosurg Psychiatry 78: 610 – 614.
- Miner MH, Raymond N, Mueller B. a, Lloyd M, Lim KO (2009): investigació preliminar de les característiques impulsives i neuroanatòmiques del comportament sexual compulsiu. Psychiatry Res - Neuroimaging 174: 146-151.
- Murray GK, Corlett PR, Clark L, Pessiglione M, Blackwell AD, Honey G, Jones PB, Bullmore ET, Robbins TW, Fletcher PC (2008): Substància nigra / ventral tegmental recompensa d'error de predicció de predicció de psicosi. Mol Psiquiatria 13: 267-276.
- Petrides M (2000): el paper de l'escorça prefrontal dorsolateral mitjana en la memòria de treball. Res Brain Res 133: 44 – 54.
- Prause N, Steele VR, Staley C, Sabatinelli D, Proudfit GH (2015): Modulació de potencials positius tardans per imatges sexuals en usuaris de problemes i controls inconsistents amb la "addicció del porno". Biol Psychol 109: 192 – 199.
- Rando K, Tuit K, Hannestad J, Guarnaccia J, Sinha R (2013): Les diferències sexuals en la disminució del volum líquid i cortical de la matèria grossa a la dependència de la cocaïna: un estudi morfomètric basat en voxel. Addicte Biol 18: 147 – 160.
- Equip de RC (2014): R: un idioma i un entorn per a la computació estadística. Viena, Àustria: Fundació R per a la Informàtica Estadística. ISBN 3-900051-07-0.
- Reid RC, Carpenter BN, Hook JN, Garos S, Manning JC, Gilliland R, Cooper EB, McKittrick H, Davtian M, Fong T (2012): Informe de resultats en un assaig de camp DSM-5 per al trastorn hipersexual. J Sex Med 9: 2868 – 2877.
- Reissner KJ, Kalivas PW (2010): utilitzar l'homeòstasi del glutamat com a objectiu per al tractament de trastorns addictius. Behav Pharmacol 21: 514.
- Seok JW, Sohn JH (2015): substrats neuronals del desig sexual en individus amb comportament hipersexual problemàtic. Front Behav Neurosci 9: 1 – 11.
- Sheehan DV, Lecrubier Y, Sheehan KH, Amorim P, Janavs J, Weiller E, Hergueta T, Baker R, Dunbar GC (1998): La mini-entrevista neuropsiquiàtrica internacional (mini): el desenvolupament i la validació d'una entrevista psiquiàtrica de diagnòstic estructurat per DSM-IV i ICD-10. J Clin Psychiatry 59: 22 – 33.
- Siegle GJ, Thompson W, Carter CS, Steinhauer SR, Tase ME (2007): Augment de l'amígdala i disminució de les respostes dorsolaterals prefrontals BOLD en la depressió unipolar: característiques relacionades i independents. Biol Psychiatry 61: 198 – 209.
- Sun Y, Sun J, Zhou Y, Ding W, Chen X, Zhuang Z, Xu J, Du Y (2014): Valoració de les alteracions de la microestructura in vivo en matèria grisa utilitzant DKI a la addicció als jocs d'Internet. Behav Brain Funct 10: 37.
- Taki Y, Kinomura S, Sato K, Goto R, Inoue K, Okada K, Ono S, Kawashima R, Fukuda H (2006): tant el volum global de matèria grisa com el volum regional de matèria grisa es correlacionen negativament amb la ingesta d'alcohol a la vida no alcohòlica -home japonès independent: una anàlisi volumètrica i una morfometria basada en voxel. Alcohol Clin Exp Res 30: 1045 – 1050.
- Tanabe J, Tregellas JR, Dalwani M, Thompson L, Owens E, Crowley T, Banich M (2009): La matèria grisa de l'escorça orbitofrontal medial es redueix en individus que depenen de la substància abstinents. Biol Psychiatry 65: 160 – 164.
- Voon V, Mole TB, Banca P, Porter L, Morris L, Mitchell S, Lapa TR, Karr J, Harrison NA, Potenza MN, Irvine M (2014): Correlats neuronals de reactivitat sexual en individus amb i sense comportaments sexuals compulsius . PLoS One 9: e102419.
- Wang H, Jin C, Yuan K, Shakir TM, Mao C, Niu X, Niu C, Guo L, Zhang M (2015): L'alteració del volum de matèria grisa i el control cognitiu en adolescents amb trastorns de joc a Internet. Front Behav Neurosci 9: 1 – 7.
- Weng CB, Qian RB, Fu XM, Lin B, Han XP, Niu CS, Wang YH (2013): anomalies de matèria grisa i de matèria blanca en l'addicció als jocs en línia. Eur J Radiol 82: 1308 – 1312.
- Whiteside SP, Lynam DR (2001): el model de cinc factors i la impulsivitat: utilitzar un model estructural de personalitat per entendre la impulsivitat. Pers Individus Diff 30: 669 – 689.
- Whitfield-Gabrieli S, Nieto-Castanon A (2012): caixa d'eines de connectivitat funcional per a xarxes cerebrals correlacionades i anticorrelades. Connexió cerebral 2: 125 – 141.
- Wreas J, Makris N, Braus DF, Mann K, Smolka MN, Kennedy DN, Caviness VS, Hodge SM, Tang L, Albaugh M, Ziegler D. a, Davis OC, Kissling C, Schumann G, Breiter HC, Heinz A ( 2008): volum de l'amígdala associat a la recaiguda i al desig d'abús d'alcohol. Am J Psychiatry 165: 1179 – 1184.
- Xiao P, Dai Z, Zhong J, Zhu Y, Shi H, Pan P (2015): dèficits regionals de matèria grisa en dependència de l’alcohol: metaanàlisi d’estudis de morfometria basada en voxel. Depèn de l’alcohol de drogues 153: 22 – 28.
- Yuan K, Qin W, Wang G, Zeng F, Zhao L, Yang X, Liu P, Liu J, Sun J, KM von Deneen, Gong Q, Liu Y, Tian J (2011): anomalies de la microestructura en adolescents amb addicció a Internet trastorn. PLoS One 6: e20708.
- Yuan K, Cheng P, Dong T, Bi Y, Xing L, Yu D, Zhao L, Dong M, KM von Deneen, Liu Y, Qin W, Tian J (2013): anormalitats del gruix cortical en l'adolescència final amb addicció al joc en línia . PLoS One 8: e53055.
- Zhou Y, Lin FC, Du YS, Qin LD, Zhao ZM, Xu JR, Lei H (2011): anomalies de matèria grisa en addicció a Internet: un estudi de morfometria basat en voxel. Eur J Radiol 79: 92 – 95.

{kind=link}
{kind=link}