PŘÍKLADY: Vynikající přehled téměř celého relevantního výzkumu androgenních receptorů, dopaminu a sexuální funkce. Fantastické kreslení vzájemných vztahů hypotalamu-odměnového obvodu.
Horm Behav. 2008 May; 53(5): 647-658.
Publikováno online 2008 únor 13. dva: 10.1016 / j.yhbeh.2008.01.010
Abstraktní
Adolescence je spojena s nárůstem chování při hledání potěšení, které se zase formují pubertální aktivací osy hypotalamo-hypofýza-gonáda. U zvířecích modelů přirozeně odměňujícího chování, jako je pohlaví, testikulární androgeny přispívají k vývoji a projevu chování u mužů. Aby se uskutečnilo zralé chování, dochází v průběhu dospívání k významnému přetvoření mozku a mnoho změn je také citlivá na androgeny, pravděpodobně působící prostřednictvím androgenních receptorů (AR). Vzhledem k jemné interakci gonadálních hormonů a vývoji mozku není překvapením, že narušení hladin hormonů během tohoto citlivého období výrazně mění chování dospívajících a dospělých. U samců křečků je pro normální projev dospělého sexuálního chování vyžadováno vystavení testosteronu během dospívání. Muži zbaveni androgenů během puberty vykazují trvalé deficity při páření. Naopak samotné androgeny nejsou dostatečné k vyvolání páření u předpubertálních samců, přestože mozková AR je přítomna před pubertou. V této souvislosti je významné znepokojení široce rozšířené užívání anabolicko-androgenních steroidů (AAS) během dospívání. Zneužití AAS má potenciál změnit časování a hladiny androgenů u dospívajících mužů. U křečků exponovaná adolescentní AAS zvyšuje agresivitu a způsobuje trvalé změny v neurotransmiterních systémech. Kromě toho AAS posilují, jak demonstruje samo-podávání testosteronu a dalších AAS. Nedávné důkazy však naznačují, že posilující účinky androgenů nemusí vyžadovat klasickou AR. Proto je pro lepší pochopení zneužití AAS nutné další vyšetření interakcí mezi androgeny a odměňujícím se chováním v adolescentním mozku.
O programu
Dospívání probouzí mozek jak pro potěšení, tak pro riziko. U lidských teenagerů se často jedná o experimentování s drogami a sexem. Ve Spojených státech je průměrný věk prvního pohlavního styku u mužů v 16.4 letech a 65% měl pohlavní styk podle třídy 12th (Kaiser Family Foundation, 2005). Podobně má tato populace nejvyšší míru užívání nelegálních drog ve Spojených státech. Podle 2004 národního šetření o užívání a zdraví uživatelů 38% věkových skupin 18-25 v uplynulém roce užívalo nelegální drogu (SAMHSA / OAS, 2005). Navíc 31% dospívajících chlapců během svého posledního sexuálního setkání užívalo drogy nebo alkohol (Kaiser Family Foundation, 2005). Kromě toho dospívání je klíčovým časem v etiologii určitých psychopatologií, jako je deprese, úzkost, poruchy stravování a porucha chování. Předpokládáme, že pubertalní sekrece gonadálních hormonů, jejich aktivace steroidních receptorů v mozku a interakce mezi hormony a zkušenosti s vývojem mozku dospívajících přispívají ke změnám v chování pozorovanými během dospívání.
Naším cílem je přezkoumat důkazy o tom, že gonadální androgeny zprostředkovávají dospívající dospívání a dospělé výkony motivovaného chování, stejně jako odměňující vlastnosti těchto chování. Také předkládáme důkazy o tom, že testosteron sám o sobě je obohacující, což pravděpodobně přispívá k maturním změnám v motivovaném chování během dospívání, kdy hladiny testosteronu stoupají. Zaměření tohoto článku je na našich studiích neurálních obvodů, které jsou základem mužského sexuálního chování, zejména u syrského křečka, se zvláštním důrazem na interakci mezi testosteronem a dopaminem (DA). Navrhujeme, aby pubertalní androgeny měly přechodné i dlouhodobé účinky na odměňování a motivované chování. Dále předpokládáme, že suplementace exogenními androgeny ve formě anabolicko-androgenních steroidů (AAS) zvyšuje normální vlivy pubertalních androgenů, což negativně ovlivňuje dospívající vývoj mozku a chování.
Adolescence jako citlivé období pro vývoj mozku
Mozek je nakonec spouštěčem i cílem působení androgenu během dospívání. U mladých chlapců (<12 let) a mladých křečků (<28 dnů věku) jsou cirkulující androgeny a gonadotropiny na bazální úrovni. Vzhledem k tomu, že sekrece luteinizačního hormonu z přední hypofýzy stoupá v reakci na hormon uvolňující hypotalamický gonadotropin, významně se zvyšuje koncentrace cirkulujícího testosteronu. K tomu dochází u Tannerovy fáze II / III (14 let) u chlapců a do 28 dnů věku u křečků. V době, kdy chlapci dosáhnou Tannerova stadia IV / V (přibližně 16 let) nebo když jsou křečkům 50–60 dnů, je endogenní testosteron v rozmezí dospělých mužů. Sekrece pubertálního hormonu se shoduje s obdobím dospívání, které u lidí probíhá přibližně od 12 do 20 let. Pubertální hormony působí nejen na periferní tkáně, aby způsobily výskyt sekundárních pohlavních znaků, které jsou zjevnými známkami puberty, ale působí také centrálně a ovlivňují jak remodelaci adolescentního mozku, tak zrání chování. Kromě toho fyziologické a neurologické změny způsobené pubertálními hormony vedou k významným změnám v prožívání jedince, které samy o sobě mohou hluboce změnit průběh vývoje mozku. Pubertální vzestup pohlavních steroidních hormonů, který je způsoben vývojově časovaným zráním reprodukční neuroendokrinní osy, tedy zase formuje vývoj chování dospívajících prostřednictvím přímých i nepřímých vlivů na nervový systém.
Lidské dospívání je nyní uznáváno jako hlavní a dynamické období nervového vývoje, během kterého jsou behaviorální obvody remodelovány a rafinovány. Ačkoli mozek dítěte starého 5 je již 90% své dospělé velikosti (Dekaban, 1978), stále přichází významná přestavba. Tento koncept byl zapálený výzkumem jak u lidí, tak u zvířat dokumentujících, že mnohé ze základních vývojových procesů, které se vyskytují během perinatálního vývoje mozku, jsou rekapitulovány během dospívání. Tyto procesy zahrnují neurogenezi (Eckenhoff a Rakic, 1988; On a posádky, 2007; Pinos, Collado, Rodriguez-Zafra, Rodriguez, Segovia a Guillamon, 2001; Rankin, Partlow, McCurdy, Giles a Fisher, 2003), programovaná smrt buněk (Nunez, Lauschke a Juraska, 2001; Nunez, Sodhi a Juraska, 2002), zpracování a prořezávání dendritických arborizací a synapsí (Andersen, Rutstein, Benzo, Hostetter a Teicher, 1997; Huttenlocher a Dabholkar, 1997; Lenroot a Giedd, 2006; Sowell, Thompson, Leonard, Vítejte, Kan a Toga, 2004), myelinizace (Benes, Turtle, Khan a Farol, 1994; Paus, Collins, Evans, Leonard, Pike a Zijdenbos, 2001; Sowell, Thompson, Tessner a Toga, 2001) a sexuální diferenciace (Chung, De Vries a Swaab, 2002; Davis, Shryne a Gorski, 1996; Nunez a kol., 2001). Vývojová trajektorie postnatálního mozku tedy není lineární, ale je charakterizována adolescentním výbuchem rychlých změn a zahrnuje jak progresivní, tak regresivní události. Jak každý vývojový biolog vědí, perioda změny signálu o rychlé vývojové změně zvýšila citlivost a zranitelnost vůči změnám závislým na zkušenostech a nežádoucím důsledkům perturbace a urážky a neexistuje žádný důvod k domněnce, že rozvoj mozku u dospívajících jedinců je výjimkou (Andersen, 2003; Spear, 2000). Takže by bylo možno předpokládat, že perturbace v načasování pubertalních hormonálních vlivů na dospívajícího mozku budou mít dlouhodobé důsledky pro chování dospělých.
Androgeny a neurální obvody pro motivované chování
Vzhledem k tomu, že dospívání je přechodná a dynamická fáze vývoje, bylo by obtížné hodnotit adolescentní mozog a chování v izolaci. Místo toho, aby ocenil jedinečný charakter adolescence, je užitečné ji kontrastovat s mozkem a chováním dospělých dospělých. Takže se zaměřením tohoto článku na mužské sexuální chování a odměnu je důležité zde představit neurální obvody pro kopulaci a sexuální motivaci u dospělých mužů, včetně úlohy gonadálních steroidních hormonů v aktivaci chování a distribuci receptorů pro androgeny (AR) a estrogeny (ER).
AR jsou přítomny v buněčných skupinách, které tvoří neurální obvody, které zprostředkovávají odměňování společenského chování, například pohlaví. MRI mozku se navíc projevují před pubertou u křečků a jsou upregulovány androgeny jak u mladistvých, tak u dospělých samců (Kashon, Hayes, Shek a Sisk, 1995; Meek, Romeo, Novak a Sisk, 1997). V mozku hlodavců dochází k podstatnému překrývání distribuce AR a ER (Wood a Newman, 1995) a aromatázy (Celotti, Negri-Cesi a Poletti, 1997), zahrnující jak α a β formy estrogenního receptoru (Shughrue, Lane a Merchenthaler, 1997). Při vazbě na ligand "klasická" AR a ER fungují jako transkripční faktory, které indukují transkripci a syntézu nových proteinů. Není překvapivé, že tyto účinky sledují relativně pomalý časový průběh se zpožděným nástupem účinku. Steroidní stimulace pohlavního chování samčího křečka (Noble a Alsum, 1975) je v souladu s akcemi prostřednictvím klasických genomických akcí. Například je třeba týdny expozice steroidů 2 obnovit páření v dlouhodobých kastrátech. Nedávné studie u potkanů také prokázaly rychlé buněčné účinky androgenů v oblastech mozku, které mají jen málo klasických receptorů (Mermelstein, Becker a Surmeier, 1996). Předpokládá se, že tyto steroidní účinky jsou zprostředkovány negenomickými receptory. Zatímco distribuce klasických AR a ER v mozku křečka je relativně omezená (Dřevo a Swann, 1999), potenciální cíle mozku pro genogenní androgenní účinky jsou mnohem širší.
Středová preoptická oblast (MPOA) hraje ústřední roli při kopulaci u mužů od zlaté rybky po člověka Hull, Wood a McKenna, 2006). Navíc křeček MPOA transdukuje gonadální steroidní hormony prostřednictvím bohatých AR a ER a testosteronové implantáty v MPOA jsou dostatečné k obnovení sexuální aktivity u dlouhodobých castrátůDřevo a Swann, 1999). U samců potkanů působí gonadální steroidy v MPOA k regulaci bazálního uvolňování DA (Putnam, Sato a Hull, 2003) a stimulovat páření (Hull, Du, Lorrain a Matuszewich, 1995). Zpočátku dochází k mírnému nárůstu DA, když je žena zobrazena za obrazovkou. Během kopulace se MPOA DA dále zvyšuje (+ 50% výchozí hodnoty) a tento účinek vyžaduje androgeny (Hull a kol., 1995; Putnam a kol., 2003). Není překvapením, že u kastrovaných mužů, kteří nejsou spolu, MPOA DA se nezvyšuje (Hull a kol., 1995). Je to poněkud obtížné interpretovat tento výsledek, protože nedostatek uvolňování DA je zpochybněn absencí sexuální aktivity. Avšak uvolnění DA v MPOA koreluje se ztrátou páření u krátkodobých castrátů (Hull a kol., 1995) a testosteronem vyvolanou obnovou sexuální aktivity u dlouhodobých castrátů (Du, Lorrain a Hull, 1998; Putnam, Du, Sato a Hull, 2001).
U hlodavců MPOA hrají androgenní a estrogenní metabolity testosteronu specifické role v regulaci páření (Putnam a kol., 2003; Putnam, Sato, Riolo a Hull, 2005). Lahodnost iniciovat kopulaci (mount nebo intromit) je jedním měřítkem sexuální motivace. Latentnost vůči sexuální aktivitě je citlivá na estrogeny, a to prostřednictvím udržování syntézy oxidu dusnatého MPOA, které naopak udržuje bazální DA hladiny. Castrasty léčené estrogenem vykazují vysoké bazální DA hladiny, které silně korelují se schopností iniciovat kopulaci. Nepodaří se však prokázat zvýšení produkce DA, které je důsledkem sexuální výkonnosti, vyvolané ženami a komplikacemi. V důsledku toho je jejich sexuální výkon nižší než intaktní. Na druhé straně castrati, které jsou léčeny pouze samotným aromatizačním androgenem, nevykazují zvýšené bazální DA hladiny a nezpůsobují kopulaci. Pro normální sexuální výkonnost se proto vyžadují jak estrogeny, tak androgeny. Sexuální výkonnost je obvykle vyjádřena jako frekvenční měřítka mountů, intromise a ejakulace. Pouze když jsou nahrazeny estrogeny a androgeny, vykastrovaní muži vykazují zvýšené hladiny DA (a kratší latenci) a DA zvýšené (a zvýšené frekvenční měření) indukované ženami a komplikacemi. Tímto způsobem přispívají estrogeny v MPOA k sexuální motivaci a jak estrogeny, tak androgeny k sexuálnímu výkonu.
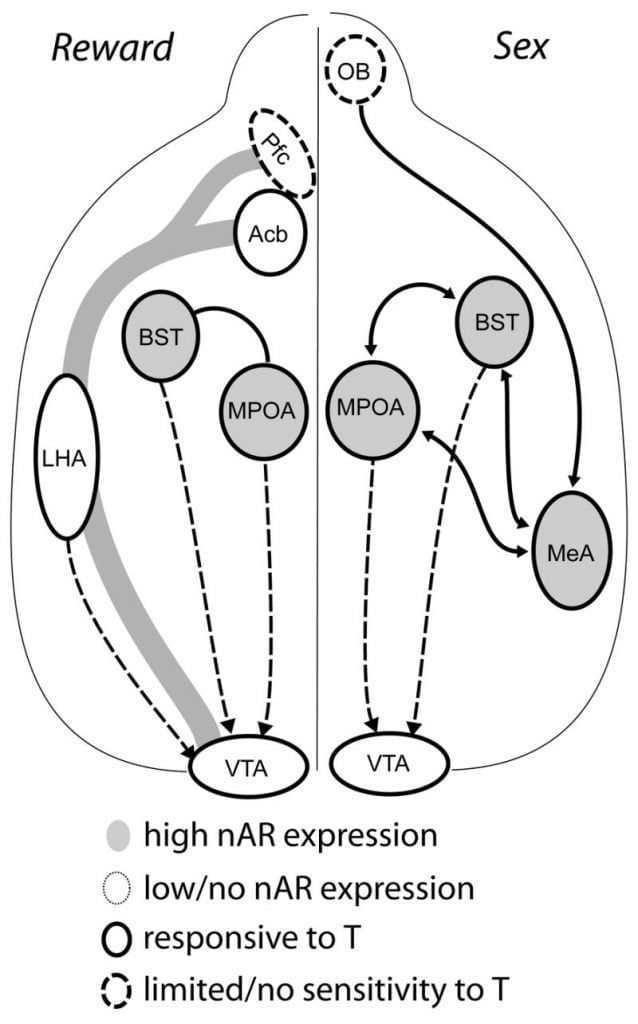
I když testosteron je nezbytný pro uvolňování MPOA DA během chování kopřivky u mužů a pro samotné páření, ani testosteron, ani samotný páření nemohou vyvolat DA v MPOA. Namísto toho se pro uvolňování DA v MPOA vyžadují chemosenzorické signály od konspecifických samic. U hlodavců jsou chemosenzorické podněty primární smyslovou modalitou k zahájení mužského sexuálního chování (Obr. 1). Chemosenzorické znamení jsou přenášeny z čichových cibulí na MPOA prostřednictvím mediálního amygdaloidního jádra a ložiskového jádra Stria terminis, struktur s bohatým AR a ER (Dřevo a Swann, 1999). K určení úlohy chemosenzorických podnětů v DA vyvolaném pářením jsme naměřili MPOA DA během páření u gonad-intaktních samců křečků s jednostrannou čichovou bulbektomií (UBx, Triemstra, Nagatani a Wood, 2005). Přestože bilaterální odstranění čichových žárovek eliminuje sexuální aktivitu a uvolnění MPOA DA, jednostranná bulbektomie neinterferuje s pářením. V této studii kopulace indukovala uvolnění MPOA DA, když byla měřena kontralaterálně k poškozené čichové baňce, ale ne v ipsilaterální polokouli (Obr. 2). Podobné výsledky byly pozorovány u samců potkanů s lézemi mediální amygdaly (Dominguez, Riolo, Xu a Hull, 2001). V související studii chemická stimulace mediální amygdaly u potkanů indukovala uvolnění MPOA DA ekvivalentní uvolnění v průběhu kopulace (Dominguez a Hull, 2001). Společně tyto údaje naznačují, že testosteron vytváří permisivní prostředí, které umožňuje vnějším smyslovým podnětům dosáhnout MPOA a vyvolat uvolňování DA během kopulace.

Obr. 2
Nakonec sexuální chování a další přírodní odměny aktivují cesty neurální odměny. Mezokortikolimbický DA obvod sestává z ventrální tegmentální oblasti (VTA), nucleus accumbens (Acb) a prefrontální kůry (Pfc). Subjekty buněk dopaminu, které se nacházejí v projektu VTA, směrem k Acb a Pfc (Koob a Nestler, 1997). U potkanů se DA uvolní do Acb během sexu (Pfaus, Damsma, Nomikos, Wenkstern, Blaha, Phillips a Fibiger, 1990). Mnohé zneužívané léky také působí v mezolimbickém systému DA ke zvýšení uvolňování DA (amfetaminy) nebo k inhibici zpětného příjmu DA (kokain, Di Chiara a Imperato, 1988), čímž posilují jejich návykové vlastnosti. Tímto způsobem má testosteron potenciál ovlivnit uvolňování DA v Acb, a to jak prostřednictvím jeho zlepšení sexuálního chování, tak prostřednictvím svých činů jako drogy zneužívání (viz níže).
Současné důkazy naznačují, že mezokortikolimbický DA systém dozrává během dospívání. Hustoty Acb DA vlákna se během dospívání dramaticky zvyšují v gerbilích, což naznačuje, že významné dozrávání dopaminergních projekcí VTA na Acb nastává v dospívajícím období (Lesting, Neddens a Teuchert-Noodt, 2005). Kromě toho je dopaminergní vstup do buněk GABA (y-aminomáselné kyseliny) v potlačení mediální prefrontální kůry obohacen a modulován serotonergními systémy během pubertalního vývoje (Benes, Taylor a Cunningham, 2000) a manipulace s androgeny u dospělých krys vede k změnám v dopaminergní hustotě axonů v prefrontální kůře (Kritzer, 2003). Pfc, Acb a VTA mají málo AR nebo ER, ačkoli ERβ je přítomen ve VTA (Shughrue a kol., 1997). Proto je pravděpodobné, že androgeny ovlivňují mezokorticolimbický DA systém prostřednictvím aferentních aterogenních aferentů nebo prostřednictvím ERp ve VTA jako v hypotalamu (Handa a kol., Toto vydání). Naše data ukazují, že buňky citlivé na androgen u mužských křečků projevují na VTA ze struktur spojených s chováním citlivým na steroidy. Například jak MPOA, tak jádro lůžka stria terminalis (BST) obsahují velké množství AR-pozitivních buněk, které vyčnívají do VTA (Sato a dřevo, 2006). Ventrální pallidum, hlavní akrobický terč (Zahm a Heimer, 1990), obsahuje také mnoho AR-pozitivních buněk, které se projevují na VTA. Tyto projekce poskytují androgenům možnost upravit aktivitu mezokortikolimbického DA systému.
Steroidně závislá organizace chování během dospívání
Tradiční pohled na hormonální účinek na chování dospívajících je založen na aktivačních účincích steroidních hormonů, které se týkají schopnosti steroidů usnadňovat chování v konkrétních společenských kontextech působením v cílových buňkách v neurálních obvodech, které jsou základem chování. Aktivační účinky jsou přechodné v tom smyslu, že přicházejí a odcházejí s přítomností a nepřítomností hormonu a jsou typicky spojené s výrazem chování dospělých. Naproti tomu organizační účinky se vztahují na schopnost steroidů sculptovat strukturu nervového systému během vývoje. Strukturální organizace je trvalá, přetrvává za dobu expozice hormonu a určuje nervové a behaviorální odpovědi na steroidy v dospělosti. Naše chápání vývojového vztahu mezi organizačními a aktivačními účinky steroidních hormonů se vyvinulo během posledních let 50. Phoenix a kolegové nejprve navrhli, aby behaviorální reakce dospělých (aktivace) na steroidní hormony byly programovány (organizovány) steroidními hormony během maximálně citlivého období perinatálního vývoje (Phoenix, Goy, Gerall a Young, 1959). Později Scott a kolegové položili teoretické východisko pro existenci více citlivých období pro postupné uspořádání nervového systému a poznamenali, že citlivá období se pravděpodobně vyskytují během období rychlé změny vývoje (1974). Následně Arnold a Breedlove poukázali na to, že organizmus mozku, který závisí na steroidech, může nastat mimo citlivé období vývoje (Arnold a Breedlove, 1985). Během posledních let 15 výzkum zaměřený na použití různých modelů zvířat a systémů chování jasně ukazuje, že dospívající mozky jsou citlivé jak na aktivační, tak na organizační účinky gonadálních steroidů Sisk a Zehr, 2005). A stejně jako jiné období rychlé vývojové změny představuje adolescence definované okno příležitosti pro remodelaci mozku závislého na steroidech.
Naše práce s použitím křečka jako zvířecího modelu poskytuje důkaz, že mužské společenské chování je modifikováno steroidy během dospívání (Schulz, Menard, Smith, Albers a Sisk, 2006; Schulz a Sisk, 2006). Před pubertou léčba testosteronem nemůže aktivovat sexuální chování u křečků, což naznačuje, že procesy zrání, které způsobují, že nervové okruhy, které jsou náchylné k aktivaci nebo organizaci steroidními hormony, dosud nenastaly (Meek a kol., 1997; Romeo, Richardson a Sisk, 2002a). Naopak, zatímco zjevná exprese mužského reprodukčního chování v dospělosti absolutně nevyžaduje přítomnost gonadálních steroidů během dospívání, maximální projev chování ano. Při srovnání mužského reprodukčního chování u samců kastrovaných buď prepubertálně (NoT@P) nebo postpubertálně (T@P) a poté léčených testosteronem v dospělosti, prepubertální kastráti NoT@P mají alespoň 50% deficit v mužském chování ve srovnání se samci kastrovanými po dospívání. (Obr. 3, Schulz, Richardson, Zehr, Osetek, Menard a Sisk, 2004). Navíc deficity reprodukčního chování jsou dlouhodobé a nemohou být překonány buď prodlouženou léčbou testosteronem nebo sexuálními zkušenostmi v dospělosti (Schulz a kol., 2004). Podobně po léčbě estrogenem a progesteronem vykazují muži NoT@P kratší latence lordózy a delší trvání lordózy než muži kastrovaní v dospělosti (Schulz a kol., 2004), což naznačuje, že prepuberální castrati jsou méně defeminizované než muži vystavení pubertálnímu testosteronu.

Obr. 3
Je možné, že muži NoT@P trpí sníženou sexuální motivací. Jedním ze způsobů, jak odpovědět na tuto otázku, je porovnat latence při zapojení do anogenitálního vyšetření (AGI) a nárůstu mezi muži s gonadektomií před (NoT@P) a po pubertě (T@P). Pokud je sexuální motivace závislá na expozici gonadálních hormonů během dospívání, předpovídali bychom delší latence pro zapojení se do sexuálního chování u mužů NoT@P. Při opakované expozici estrálním samičkám trvá samcům NoT@P déle, než začnou AGI a narůstají, ve srovnání s muži T@P (Obr. 4). Kromě organizování aspektů sexuální výkonnosti se tedy zdá, že pubertální hormony také organizují odměňující aspekty sexuálního chování. Na podporu této možnosti centrální podávání DA agonisty apomorfinu v dospělosti obnovuje zvyšující se chování mužů NoT@P na hladiny typické pro dospělé, což naznačuje, že testosteron během dospívání normálně organizuje dopaminergní nervové okruhy (Salas-Ramirez, Montalto a Sisk, 2006 ). Přesto zůstává mnoho zajímavých otázek. Vyvinul by si samec NoT@P barpress pro estrální samici nebo vyvinul podmíněnou preferenci místa pro páření? Budoucí výzkum bude zkoumat roli pubertálních hormonů při organizování sexuální motivace a sexuální výkonnosti.

Obr. 4
Latence a trvání anogenitálního vyšetření (AGI) projevované samci křečků gonadektomizovaných před pubertou (NoT@P) nebo po pubertě (T@P). Všichni muži byli v dospělosti 7 týdnů po gonadektomii a jeden týden před prvním testem chování léčeni testosteronem. A. T@P muži vykazovali podobné latence AGI ve třech testech s estrální samicí, zatímco NoT@P muži zvýšili AGI latence během třetího testu s estrální samicí. B. Samci T@P snížili latenci přilnutí ve třech testech chování s estrální samicí, zatímco samci noT@P nevykázali žádnou změnu latence přisednutí ve třech testech chování. Tyto údaje naznačují, že pubertální gonadální hormony mají trvalé, usnadňující účinky na motivaci dospělých mužů k sexuálnímu chování se ženou. (Nepublikovaná data od zvířat v Schulz, KM, Richardson, HN, Zehr, JL, Osetek, AJ, Menard, TA a Sisk, CL, 2004).
Prepubertální reakce na steroidy
Jedním z trvalých hádanek vývoje chování dospívajících je důvod, proč je aktivace reprodukčního chování v reakci na expozici steroidů oslabena u předkubertních samců křečků. Pokud nízké hladiny androgenů před pubertou omezují expresi mužského pohlavního chování u samic předepubertů, pak doplnění endogenních androgenů u samců předepubertů by mělo vyvolat páření. Ukázalo se, že tomu tak není (Meek a kol., 1997; Romeo, Cook-Wiens, Richardson a Sisk, 2001; Romeo, Wagner, Jansen, Diedrich a Sisk, 2002b), a to i přesto, že počet a distribuce AR a ER v porovnání s obdobným okruhem jsou podobné u hormonem léčených prepuberálních a dospělých castrátůMeek a kol., 1997; Romeo, Diedrich a Sisk, 1999; Romeo a kol., 2002a). Proto se zdá, že androgeny a AR jsou nezbytné, ale nejsou dostatečné pro vyjádření mužského sexuálního chování.
Úsilí k identifikaci faktorů, které omezují sexuální aktivitu před pubertou, bylo zatím smíšeno. Fos odpovědi na chemosenzorické podněty od estrogenních žen jsou podobné u předkubertálních a dospělých samců křečků (Romeo, Parfitt, Richardson a Sisk, 1998). Tato data ukazují, že mechanismy senzorických transdukcí jsou před pubertou zralé. Proto jsou mladiství samci schopni odhalit chemosenzorické náznaky od žen; kde se liší od dospělých, je v tom, jak reagují na tyto náznaky. Jedním z možných vysvětlení je, že prepubertální muži nejsou motivováni k sexuálnímu chování. Zjistili jsme, že předepubertální samci křečků nevykazují zvýšené dopaminergní odpovědi v MPOA jako odpověď na feromony žen, zatímco dospělé pohlavně dospělé osoby vykazují robustní MPOA dopaminergní odpovědi na stejné stimuly (Obr. 5, Schulz, Richardson, Romeo, Morris, Lookingland a Sisk, 2003). Podobně předepubertální samci nevykazují typické zvýšení počtu cirkulujících testosteronů po expozici ženským feromonům (Parfitt, Thompson, Richardson, Romeo a Sisk, 1999). Ženské feromony se tedy zdají být nepodmíněným podnětem pro neurochemické a neuroendokrinní odezvy u dospělých, ale ne předubertalních mužů, což naznačuje, že změna těchto společensky významných senzorických podnětů se mění nad vývojem puberty, možná souvisejícím se získáváním obohacujících vlastností a sexuální motivací. Navíc, ačkoliv testosteron usnadňuje AGI ženského pohlaví u předpubertálních samců, tento účinek závisí na tom, zda muž měl předchozí expozici estrosu. Snad překvapivě léčba testosteronem snižuje latenci a prodlužuje trvání AGI pouze u pohlavně naivních prepuberálních samců (Obr. 6). Dále předepubertální muži, kteří měli jednu předchozí zkušenost s ženskou, vykazovali mnohem delší latenci AGI a kratší dobu AGI než muži, kteří poprvé reagovali s receptivními ženami (Obr. 6). Tyto údaje naznačují, že interakce s estrogenní ženou jsou před pubertou spíše nepříjemné než odměňování, čímž se eliminuje jakýkoli usnadňující účinek testosteronu na AGI během následných interakcí se ženou. Bylo by zajímavé vědět, zda negativní behaviorální důsledky předčasné expozice ženám z estrosu přetrvávají v dospívání a dospělosti, zejména vzhledem k tomu, že opakovaná expozice estrosu ženám během dospívání obecně usnadňuje projev reprodukčního chování mužů (Molenda-Figueira, Salas-Ramirez, Schulz, Zehr, Montalto a Sisk, 2007).

Obr. 5
Prepubertální a dospělé dopaminové reakce na mužské mediální preoptické oblasti (MPOA) na feromony žen obsažené ve vaginálních sekretech. Dospělí muži vykazují zvýšení dopaminergní aktivity MPOA s expozicí ženských vaginálních sekrecí, zatímco preububertální samci nevykazují zvýšené dopaminergní odpovědi MPOA na feromony žen. (Překreslení z Schulz, KM, Richardson, HN, Romeo, RD, Morris, JA, Lookingland, KJ a Sisk, CL, 2003).

Ačkoli prepuberální androgenní léčba nemůže vyvolat kopulaci, nedávná práce z naší laboratoře naznačuje, že nervový systém křečka je citlivý na organizační účinky testosteronu na reprodukční chování před adolescencemi (Schulz, Zehr, Salas-Ramirez a Sisk, 2007). Kastrace plus 19 dny vystavení testosteronu před nebo během, ale ne po adolescenci usnadnilo montážní chování, když byl testosteron v dospělosti nahrazen. Muži vystavení testosteronu předčasně také projevovali další intromise v dospělosti než muži vystavení testosteronu během nebo po pubertu (Schulz a kol., 2007). Tyto údaje naznačují, že schopnost testosteronu uspořádat behaviorální neurální obvody klesá s věkem a že dospívání označuje konec prodlouženého postnatálního citlivého období pro expozici testosteronu.
Farmakologické androgeny
Předchozí údaje naznačují, že endogenní gonadální steroidy zvyšují motivační chování během dospívání. Nyní, co se stane, když se člověk aplikuje samotné androgeny v koncentracích až do 100x normálních fyziologických koncentrací? To je problém zneužívání anabolicko-androgenních steroidů (AAS) (viz Brower, 2002; Clark a Henderson, 2003). Zde je vhodné krátké rozdělení: všechny AAS jsou deriváty testosteronu, všechny AAS mají uhlíkový skelet s kruhy 4, většinou mají uhlíky 19. AAS jsou používány hlavně pro jejich anabolické účinky (budování svalů). Nicméně, jak jejich název naznačuje, AAS mají také androgenní vlastnosti. Testosteron je logická volba ve studiích na zvířatech pro zkoumání základních mechanismů androgenní odměny. Zůstává i populární volbou pro lidské uživatele, nejčastěji ve formě dlouhodobě působících esterů testosteronu, jako je propionát testosteronu. V testu 2006 byl testosteron jednou z nejběžnějších zakázaných látek zjištěných v moči v laboratořích akreditovaných WADA (WADA, 2006). Testosteron představoval největší zlomek (34%) pozitivních testů moči na AAS na Olympijských hrách 2000 v Sydney (Van Eenoo a Delbeke, 2003). Podobně, u testů moči u uživatelů AAS, 41% testovala pozitivní testosteron (Brower, Catlin, Blow, Eliopulos, Beresford, 1991). Při vysokých dávkách produkují AAS významné změny v chování. Zejména kvůli jejich blízkému vztahu k testosteronu se zdá, že užívání AAS v mladistvých letech naruší normální steroidní prostředí rozvíjejícího se adolescentního nervového systému, včetně množství, načasování a typu expozice steroidů.
Stejně jako u jiných nelegálních drog je zneužívání lidských AAS problémem dospívání. Podle průzkumu domácích domácností 1994 o užívání drog (SAMHSA / OAS, 1996), užívají steroidy píky v pozdní dospívání ve věku 18. Kromě toho v rámci průzkumu Monitorování budoucnosti (Johnston, O'Malley a Bachman, 2003) byl celoživotní výskyt užívání steroidů mezi vysokoškolskými seniory (2.7%) srovnatelný s výskytem užívání steroidů u XCUMX% nebo heroinu (3.5%). Užívání steroidů je také stále častější u mladších věkových skupin: 1.4% studentů se stupněm 2.5th (roky 8-13) užívalo steroidy, podobně jako výskyt prasklin (14%) a heroinu (2.5%). Tento trend směřující k užívání AAS v raném věku dospívající mládeže je obzvláště znepokojující vzhledem k obavám ze společnosti 1.6, že adolescenti mohou být obzvláště zranitelní vůči zneužívání AAS a 1), že adolescentní expozice AAS na farmakologických úrovních má potenciál podstatně změnit normální zrání mozku a chování ke vzniku přehnaných morfologických a behaviorálních reakcí, akutně a chronicky.
Nevhodná agrese je reakce na chování, která je nejčastěji spojována s lidským zneužíváním AAS. Ve zveřejněných kazuistikách se užívání steroidů podílí na několika násilných vraždách (Conacher a Workman, 1989; Papež a Katz, 1990; Papež, Kouri, Powell, Campbell a Katz, 1996; Schulte, Hall a Boyer, 1993). V průzkumu současných uživatelů AAS byla zvýšená agresivita a podrážděnost nejčastějšími vedlejšími účinky při užívání AAS (Bond, Choi a Papež, 1995; Galligani, Renck a Hansen, 1996; Midgley, Heather a Davies, 2001; Parrott, Choi a Davies, 1994; Perry, Kutscher, Lund, Yates, Holman a Demers, 2003). Vzhledem k rozsahu expozice androgenu, rozmanitosti psychiatrických příznaků a potenciálu pre již existující psychiatrické dysfunkce je však obtížné určit přesnou úlohu AAS v těchto případech lidské agrese. Výsledky z prospektivních studií lidských dobrovolníků, kteří dostávali injekci AAS, byly smíšeny: Tricker a kol. (1996) a O'Connor a kol. (2004) nezaznamenaly žádné zvýšení rozhněvaného chování, zatímco jiné studie zaznamenaly zvýšenou agresi (Daly, Su, Schmidt, Pickar, Murphy a Rubinow, 2001; Hannan, Friedl, Zold, Kettler a Plymate, 1991; Kouri, Lukas, Pope a Oliva, 1995; Papež a Katz, 1994; Su, Pagliaro, Schmidt, Pickar, Wolkowitz a Rubinow, 1993). Nicméně je důležité mít na paměti, že dávky podávané dobrovolníkům jsou mnohem nižší než dávky doporučované na webových stránkách budování těla a trvání léčby je obecně krátké. Zdá se tedy, že je správné konstatovat, že AAS mají potenciál zvýšit agonistické chování, alespoň u citlivých jedinců. Pope a kol. (1994) že AAS vyvolává psychiatrické symptomy u zranitelných jedinců.
Studie na zvířatech rovněž poskytly přesvědčivý důkaz agrese indukované AAS. Mladí křečci, kteří jsou chronicky léčeni vysokými dávkami steroidů, mají kratší latenci útoku a větší počet útoků a kousnutí vůči mužskému vetřelci ve srovnání s neléčenými muži (Harrison, Connor, Nowak, Nash a Melloni, 2000; Melloni, Connor, Hang, Harrison a Ferris, 1997). Podobně mírná provokace (ocas-pinch) způsobuje přetrvávající zvýšení agrese u dospívajících potkanů samců, včetně agrese vůči ženám (Cunningham a McGinnis, 2006). Vzbuzuje ještě větší obavy, že adolescentní expozice AAS u křečků způsobuje trvalé zvýšení agonistického chování, které přetrvává po ukončení užívání steroidů (Grimes a Melloni, 2006). Tyto změny v chování jsou doprovázeny trvající remodelací neurálních obvodů v předním hypotalamu. Zejména adolescentní expozice AAS u křečků zvyšuje arginin vazopresin (AVP, Grimes a Melloni, 2006) a downreguluje serotonin a serotonergní receptory 5HT1A a 5HT1B (Ricci, Rasakham, Grimes a Melloni, 2006). Nemělo by být žádným překvapením, že AAS změní úroveň mozku AR také. Chronická expozice buď testosteronu nebo nandrolonu zvyšuje buněčnou nukleární AR u samců potkanů (Menard a Harlan, 1993; Wesson a McGinnis, 2006). Existuje tedy potenciál, aby AAS zvýšila androgen-dependentní chování jak doplňkem endogenních androgenů, tak zvýšením androgenní odezvy prostřednictvím zvýšené exprese AR.
Ve srovnání s agonistickým chováním má AAS méně výrazný účinek na chování u mužských hlodavců a reakce závisí na konkrétním použitém steroidu Clark a Henderson, 2003). U samčích křečků, kteří konzumují testosteron v perorálních roztocích, se zvýšila ejakulace způsobem závislým na dávce (Wood, 2002). Ovšem ani testosteron, ani nandrolon nezlepšily páření u dospívajících samců potkanů. Stanozolol, relativně méně účinný AAS s minimální androgenní aktivitou, skutečně inhiboval jak páření, tak agresi (Farrell a McGinnis, 2003), pravděpodobně snížením hladin endogenních androgenů.
Je zvláště důležité poznamenat, že dospívající a dospělí křečci mohou vykazovat různé reakce na chování při expozici AAS. Zatímco AAS výrazně zlepšilo agonistické chování u dospívajících mužů, stejná léčba v dospělosti způsobila jen mírné zvýšení agresivního chování a výrazně snížilo sexuální chování (Salas-Ramirez, Montaldo a Sisk, 2008). To je v souladu s pojmem adolescence jako citlivého období pro androgeny. Dále, stejně jako dospělí samci křečci získávají toleranci vůči exogennímu testosteronu (Peters a dřevo, 2005), věříme, že rozvíjející se muži získávají toleranci k testosteronu, jakmile zrají. Účinky AAS se tedy mění v rozvoji dospívajících a adolescentní expozice AAS může způsobit nadměrné agresivní a sexuální chování, které mohou přetrvávat až do dospělosti.
Zesilující účinky androgenů
Páření a boje jsou obohacující (alespoň pokud vyhrajete boj). Samci potkanů opakovaně stisknou páku, aby mohli kopulovat samici (Everitt a Stacey, 1987). Stejně tak budou samce myší a samice křečků tvořit upravenou preferenci míst (CPP) pro místa, kde dříve vyhrál boje (Martinez, Guillen-Salazar, Salvador a Simon, 1995; Meisel a Joppa, 1994). Pokud AAS může zvýšit odměňování sociálního chování nad úrovněmi normálně pozorovanými u nedospělých mužů, je logické očekávat, že samotný testosteron může být obohacující. To bylo testováno pomocí dvou osvědčených zvířecích modelů pro odměňování a posilování: CPP a samospráva. Výsledky těchto studií ukazují, že testosteron posiluje v experimentálním kontextu, kde anabolické účinky a atletické výkony jsou irelevantní. U CPP se testovaná látka opakovaně spáruje s jedinečným prostředím (například s určitou komorou v testovacím zařízení). Jakmile zvíře sdružuje vyztužující testovanou látku s tímto prostředím, bude hledat životní prostředí i bez odměny. První zprávy o androgenní odměně u laboratorních zvířat používaly systémové injekce testosteronu k indukci CPP u samců myší (Arnedo, Salvador, Martinez-Sanchis a Gonzalez-Bono, 2000; Arnedo, Salvador, Martinez-Sanchis a Pellicer, 2002) a potkanů (Alexander, Packard a Hines, 1994; de Beun, Jansen, Slangen a Van de Poll, 1992). Následně naše laboratoř použila samo-podávání testosteronu k prokázání posilování androgenu (Johnson a Wood, 2001). Zjistili jsme, že samci křečci budou dobrovolně konzumovat perorální roztoky testosteronu pomocí testů výběru lahví 2 a potravou indukovaného pití. V pozdějších studiích jsme prokázali intravenózní podávání samců potkanů a křečků (Dřevo, Johnson, Chu, Schad a Self, 2004). Intravenózní podání eliminuje potenciální zmatené účinky chuti nebo stárnutí na androgeny.
V souvislosti se zneužíváním AAS je důležité rozlišit centrální a periferní účinky androgenů. Vzhledem k tomu, že testosteron má rozsáhlé účinky v celém těle, lze tvrdit, že odměna a posilování systémovými injekcemi testosteronu je sekundární pro systémové anabolické a androgenní účinky testosteronu. Jinými slovy, možná testosteron snižuje únavu svalů a zlepšuje funkci kloubů tak, aby zvířata jen tak cítit lepší. Toto vysvětlení bylo skutečně použito v klinické literatuře (i když bez experimentálních důkazů) k argumentaci proti potenciálu závislosti a závislosti na AAS (DiPasquale, 1998). Nicméně Packard a kol. (Packard, Cornell a Alexander, 1997) ukázaly, že injekce testosteronu přímo do mozku krys mohou indukovat CPP. Podobně naše laboratoř prokázala intracerebroventrikulární (icv) testosteronové samo-podávání u samců křečků (Wood a kol., 2004). Intracerebrální CPP a samo-podávání icv s testosteronem argumentují pro centrální cíle, které zprostředkovávají posilování androgenu.
Je pozoruhodné, že posilování testosteronu nemusí nutně sledovat stejné mechanismy, které byly dříve stanoveny pro účinky steroidů na sexuální chování. Jak bylo dříve diskutováno, MPOA je klíčovým místem pro organizaci sexuálního chování pohlaví u mužů (Hull, Meisel a Sachs, 2002). U křečků má MPOA bohaté steroidní receptory a testosteronové implantáty v MPOA obnovují sexuální aktivitu v dlouhodobých kastrátech (Dřevo a Swann, 1999). Časový průběh těchto účinků steroidů je pomalý: montážní chování přetrvává po týdnech po orchidektomii a je zapotřebí prodlužovat expozici steroidů, aby se obnovilo párování dlouhodobých kastrátůNoble a Alsum, 1975). Avšak injekce testosteronu do MPOA samců potkanů neprodukují CPP (King, Packard a Alexander, 1999). To naznačuje, že další oblasti mozku jsou důležité pro posilování androgenu.
Naproti tomu samci potkanů vytvoří injekci testosteronu CPP v Acb (Packard a kol., 1997). Stejně jako u jiných léků zneužívání, DA pravděpodobně bude klíčovým neurotransmiterem pro posilování testosteronu: CPP indukovaná systémovou injekcí testosteronu je blokována antagonisty D1u a D2 dopaminového receptoru (Schroeder a Packard, 2000). Nicméně, na rozdíl od jiných léků zneužívání, naše studie u křečků naznačují, že testosteron neindukuje uvolňování Acb DA (Triemstra, Sato a Wood, v tisku). Podobně, studie samců potkanů ukazují, že androgeny nemají žádný účinek na bazální DA hladiny nebo amfetaminem stimulované uvolňování DA (Birgner, Kindlundh-Hogberg, Nyberg a Bergstrom, 2006; ale také vidět Clark, Lindenfeld a Gibbons, 1996). Testosteron navíc vykazuje relativně malý vliv na hladiny tkání Acb DA (Thiblin, Finn, Ross a Stenfors, 1999). Společně tyto údaje naznačují, že ačkoliv může posilování testosteronu v ACb v konečném důsledku změnit aktivitu DA, mechanismy mohou být odlišné od mechanismů kokainu nebo jiných stimulantů. Z tohoto pohledu nedávné údaje naznačují, že chronická expozice AAS může změnit citlivost na DA změnou metabolismu DA (Kurling, Kankaanpaa, Ellermaa, Karila a Seppala, 2005), hladiny DA receptorů (Kindlundh, Lindblom, Bergstrom, Wikberg a Nyberg, 2001; Kindlundh, Lindblom a Nyberg, 2003) nebo přepravce DA (Kindlundh, Bergstrom, Monazzam, Hallberg, Blomqvist, Langstrom a Nyberg, 2002).
V současné době nejsou známé specifické steroidní signály, receptory a místa mozku pro testosteronovou výztuž. Na základě nedávné studie křečků z naší laboratoře se zdá, že posilující účinky testosteronu jsou zprostředkovány jak androgeny, tak estrogeny (DiMeo a Wood, 2006). Obvykle zneužívaná AAS zahrnuje jak aromatizovatelné, tak aromatizovatelné androgeny (Gallaway, 1997; WADA, 2006). To znamená, že jak AR, tak ER mohou přenést steroidní stimuly na odměnu. Existuje další možnost, že zesílení testosteronu může být zprostředkováno kombinací klasických a negenomických receptorů.
Několik linií důkazů ukazuje na působení negenomových receptorů při zesilujících účincích AAS. Kromě řídké distribuce AR v Acb a VTA je časový průběh posilování androgenu rychlý (<30 minut) a zpracování signálu přes klasický AR nemusí být pro vyztužení dostatečně rychlé. V souladu s tím jsme k testování role negenomové AR při posilování AAS použili dvě doplňkové techniky (Obr. 7). V jednom pokusu (Sato, Johansen, Jordán a Wood, 2006), dovolili jsme potkanům s mutací testikulární feminizace (Tfm, viz toto téma) samo-podat dihydrotestosteron (DHT), aromatizovatelný androgen. Tfm mutace výrazně snižuje vazbu ligandu u AR. Nicméně potkani Tfm a jejich divoký typ samčích sourozenců podali zhruba stejné množství DHT. Toto tvrdí pro negenomické účinky DHT. V následné studii jsme stanovili, že samci křečci by samy podávali DHT konjugovaný s bovinním sérovým albuminem (BSA, Obr. 8, Sato a dřevo, 2007). Konjugáty DHT-BSA jsou membránově nepropustné; takže jejich účinky jsou omezeny na povrch buněk. Škrtící samci podávaný DHT, jak bylo dříve prokázáno (DiMeo a Wood, 2006). Ukázali podobnou preferenci pro konjugáty DHT-BSA, ale nedokázaly samy podávat samotný BSA.


Tato data ukazují na centrální roli ARs buněčných povrchů v androgenní výztuži. V současné době není přesná povaha takových receptorů známa. Bylo navrženo, že androgeny mohou působit na buněčný povrch prostřednictvím vazby na vyhrazenou membránu AR (Thomas, Dressing, Pang, Berg, Tubbs, Benninghoff a Doughty, 2006, viz také tento problém). Toto může být ve formě extrajaderné klasické AR, jak je uvedeno v hipokampu (Sarkey a kol., V tomto čísle). Alternativně mohou předchozí studie také popisovat místa vazby steroidů na jiných neurotransmiterních systémech. Konkrétně, řada steroidních hormonů, včetně AAS, může alostericky modulovat receptor GABA-A (Henderson, 2007; Lambert, Belelli, Peden, Vardy a Peters, 2003). Podobně sulfátované neurosteroidy mohou modifikovat aktivitu N-methyl-D-aspartátových receptorových subtypů (Malayev, Gibbs a Farb, 2002) receptory. To je důležitá oblast pro budoucí výzkum.
Proč by měla být membrána AR? Jak bylo uvedeno výše, existuje úzký vztah mezi vylučováním androgenu a odměňováním sociálního chování. Můžeme spekulovat, že zvýšení sekrece testosteronu, které následuje po páření nebo boje, slouží k posílení chování. Pokud tomu tak je, je třeba mít rychlé spojení stimulace (chování) a odměny (testosteron). To lze nejlépe dosáhnout vazbou na membránu AR. V tomto ohledu by bylo zajímavé určit, zda upnutí sekrece androgenu během páření snižuje odměňující účinky sexuálního chování.
Shrnutí
Zde přezkoumáme důkazy, že androgeny jsou silnými mediátory dospělého motivovaného chování a dále, že načasování expozice androgenu během vývojových programů arogen-dependentní chování v dospělosti. Anabolické steroidy se rychle stávají zvýhodněnými drogami zneužívanými adolescenty v USA. Zatímco AAS nemusí mít návykovou schopnost kokainu nebo heroinu, začínáme chápat potenciál pro posilování a závislost na androgenu. Zejména proto, že sportovní aktivity mladých lidí se stávají konkurenceschopnějšími, stoupá tlak na rozvoj sportovců, kteří používají steroidy, a to od mladšího věku. Tento trend je znepokojivý vzhledem k novým důkazům o sterilizačním nervovém zrání u adolescentů.
Navzdory zvýšenému povědomí veřejné i vědecké komunity o hlubokých nervových změnách doprovázejících dospívání bylo experimentální studium vývojové neurobiologie puberty omezené. Zvířecí modely vývoje dospívajících jsou potřebné k prozkoumání toho, jak načasování expozice hormonů během vývoje zvyšuje individuální riziko psychopatologie a užívání drog a jaké typy zkušeností zmírňují nebo zesilují behaviorální účinky odchylek v pubertálním načasování. Například sociální faktory, jako například vliv vrstevníků, zhoršují účinky pubertalního časování pro užívání látky a alkoholu (Biehl, Natsuaki a Ge, 2007; Patton, Novy, Lee a Hickok, 2004; Simons-Morton a Haynie, 2003; Wichstrom a Pedersen, 2001). Zvířecí modely načasování pubertu také budou informovat lidské výzkumné úsilí a potenciálně povedou k účinnějším terapeutickým intervencím během dospívání.
Poděkování
Děkujeme Eleni Antzoulatosové, Cortney Ballardové, Lucy Chu, Kelly Petersové, Jennifer Triemstra, Jane Venierové, Lisy Rogersové a Pamele Montaltové za pomoc při těchto studiích. Tato práce je podporována granty od NIH (DA12843 k RIW, MH68764 k CLS a MH070125 k KMS).
Poznámky pod čarou
Zřeknutí se odpovědnosti vydavatele: Jedná se o soubor PDF s neupraveným rukopisem, který byl přijat k publikaci. Jako službu pro naše zákazníky poskytujeme tuto ranní verzi rukopisu. Rukopis podstoupí kopírování, sázení a přezkoumání výsledného důkazu před jeho zveřejněním ve své konečné podobě. Vezměte prosím na vědomí, že během výrobního procesu mohou být objeveny chyby, které by mohly ovlivnit obsah, a veškeré právní odmítnutí týkající se časopisu.
Reference
- Alexander GM, Packard MG, Hines M. Testosteron odměňuje afektivní vlastnosti u samců potkanů: důsledky pro biologický základ sexuální motivace. Behaviorální neurovědy. 1994;108: 424-8. [PubMed]
- Andersen SL. Trajektorie vývoje mozku: bod zranitelnosti nebo okno příležitosti? Neurovědy a biobehaviorální recenze. 2003;27: 3-18. [PubMed]
- Andersen SL, Rutstein M, Benzo JM, Hostetter JC, Teicher MH. Pohlavní rozdíly v nadprodukci a eliminaci dopaminového receptoru. Neuroreport. 1997;8: 1495-8. [PubMed]
- Arnedo MT, Salvador A, Martinez-Sanchis S, Gonzalez-Bono E. Odměňování vlastností testosteronu u intaktních samců myší: pilotní studie. Farmakologie, biochemie a chování. 2000;65: 327-32.
- Arnedo MT, Salvador A, Martinez-Sanchis S, Pellicer O. Podobné účinky na testosteron u myší, které jsou hodnoceny jako osoby s krátkou a dlouhou latencí útoku. Závislost biologie. 2002;7: 373-9. [PubMed]
- Arnold AP, Breedlove SM. Organizační a aktivační účinky pohlavních steroidů na mozek a chování: reanalýza. Hormony a chování. 1985;19: 469-98. [PubMed]
- Benes FM, Taylor JB, Cunningham MC. Konvergence a plasticita monoaminergních systémů v mediální prefrontální kůře v postnatálním období: důsledky pro vývoj psychopatologie. Mozková kůra. 2000;10: 1014-27. [PubMed]
- Benes FM, Turtle M, Chán Y, Farol P. Myelinizace klíčové reléové zóny v hipokampální formaci se vyskytuje v lidském mozku v dětství, dospívání a dospělosti. Archivy obecné psychiatrie. 1994;51: 477-84. [PubMed]
- Biehl MC, Natsuaki MN, Ge XJ. Vliv pubertalního časování na užívání alkoholu a těžké pití trajektorií. Časopis mládeže a dospívání. 2007;36: 153-167.
- Birgner C, Kindlundh-Hogberg AM, Nyberg F, Bergstrom L. Neuroscience Letters. 2006. Změna extracelulárních hladin DOPAC a HVA v jádře krys jádra accumbens v reakci na subchronické podávání nandrolonu a následnou provokaci amfetaminem.
- Bond AJ, Choi PY, Pope HG., Jr Posouzení pozornosti a nálady pozornosti u uživatelů a neužívajících anabolicko-androgenní steroidy. Závislost na drogách a alkoholu. 1995;37: 241-5. [PubMed]
- Brower KJ. Anabolické steroidní abusy a závislost. Aktuální zprávy o psychiatrii. 2002;4: 377-87. [PubMed]
- Brower KJ, Catlin DH, Blow FC, Eliopulos GA, Beresford TP. Klinické vyšetření a vyšetření moči na anabolické a androgenní steroidní abusy a závislost. American Journal of Drug & Alcohol Abuse. 1991;17: 161-171. [PubMed]
- Celotti F, Negri-Cesi P, Poletti A. Steroidní metabolismus v mozku savců: redukce a aromatizace 5alpha. Brain Research Bulletin. 1997;44: 365-75. [PubMed]
- Chung WC, De Vries GJ, Swaab DF. Sexuální diferenciace ložiskového jádra Stria terminalis u lidí může být rozšířena do dospělosti. Journal of Neuroscience. 2002;22: 1027-33. [PubMed]
- Clark AS, Henderson LP. Behaviorální a fyziologické reakce na anabolické a androgenní steroidy. Neurovědy a biobehaviorální recenze. 2003;27: 413-36. [PubMed]
- Clark AS, Lindenfeld RC, Gibbons CH. Anabolické androgenní steroidy a odměna za mozku. Farmakologie, biochemie a chování. 1996;53: 741-5.
- Conacher GN, Workman DG. Násilné trestné činy, které mohou být spojeny s užíváním anabolických steroidů. American Journal of Psychiatry. 1989;146: 679. [PubMed]
- Cunningham RL, McGinnis MY. Fyzická provokace pubertálních anabolických onemocněných samčích potkanů s anabolickými androgenními steroidy vyvolává agresi vůči ženám. Hormony a chování. 2006;50: 410-6. [PubMed]
- Daly RC, Su TP, Schmidt PJ, Pickar D, Murphy DL, Rubinow DR. Cerebrospinální tekutina a změny chování po podání methyltestosteronu: předběžné nálezy. Archivy obecné psychiatrie. 2001;58: 172-7. [PubMed]
- Davis EC, Shryne JE, Gorski RA. Strukturální sexuální dimorfismy v anteroventrálním periventrikulárním jádru potkaního hypothalamu jsou perinatálně citlivé na gonadální steroidy, ale se rozvíjejí peripubertálně. Neuroendokrinologie. 1996;63: 142-8. [PubMed]
- de Beun R, Jansen E, Slangen JL, Van de Poll NE. Testosteron jako chutný a diskriminační podnět u potkanů: účinky závislé na pohlaví a dávce. Fyziologie a chování. 1992;52: 629-34. [PubMed]
- Dekaban AS. Změny hmotnosti mozku v průběhu lidského života: vztah mozkových závaží k výškám těla a tělům. Annals of Neurology. 1978;4: 345-56. [PubMed]
- Di Chiara G, Imperato A. Léky užívané lidmi přednostně zvyšují koncentrace synaptických dopaminů v mezolimbickém systému volně se pohybujících krys. Sborník z Národní akademie věd Spojených států amerických. 1988;85: 5274-8. [PMC bezplatný článek] [PubMed]
- DiMeo AN, Wood RI. Samostatné podávání estrogenu a dihydrotestosteronu u samců křečků. Hormony a chování. 2006;49: 519-26. [PubMed]
- DiPasquale M. Anabolické steroidy. In: Tarter RE, Ammerman RT, Ott PJ, redaktoři. Příručka zneužívání látek. Plenum Press; NY: 1998. str. 547-565.
- Dominguez J, Riolo JV, Xu Z, Hull EM. Regulace mediální amygádou kopulace a mediálního předoptického uvolňování dopaminu. Journal of Neuroscience. 2001;21: 349-355. [PubMed]
- Dominguez JM, Hull EM. Stimulace mediální amygdaly zvyšuje uvolnění mediálního preoptického dopaminu: důsledky sexuálního chování u potkanů samců. Výzkum mozku. 2001;917: 225-229. [PubMed]
- Du J, Lorrain DS, Hull EM. Kastrace snižuje extracelulární, ale zvyšuje intracelulární dopamin v mediální preoptické oblasti samců potkanů. Výzkum mozku. 1998;782: 11-17. [PubMed]
- Eckenhoff MF, Rakic P. Příroda a osud proliferativních buněk v hipokampálním dentátovém gyru během životnosti opice rhesus. Journal of Neuroscience. 1988;8: 2729-47. [PubMed]
- Everitt BJ, Stacey P. Studie instrumentálního chování se sexuální výztuží u samců potkanů (Rattus norvegicus): II. Účinky lézí preoptické oblasti, kastrace a testosteronu. Journal of Comparative Psychology. 1987;101: 407-19. [PubMed]
- Gallaway S. Steroidní bible. Belle International Press; Sacramento, CA: 1997.
- Galligani N, Renck A, Hansen S. Profil osobnosti mužů užívajících anabolické androgenní steroidy. Hormony a chování. 1996;30: 170-5. [PubMed]
- Grimes JM, Melloni RH., Jr Prodloužené změny v serotoninovém nervovém systému po ukončení expozice adolescentních anabolických a androgenních steroidů u křečků (Mesocricetus auratus) Behaviorální neurovědy. 2006;120: 1242-51. [PubMed]
- Hannan CJ, Jr, Friedl KE, Zold A, Kettler TM, Plymate SR. Změny psychologické a sérové homovanilové kyseliny u mužů podávaných androgenní steroidy. Psychoneuroendokrinologie. 1991;16: 335-43. [PubMed]
- Harrison RJ, Connor DF, Nowak C, Nash K, Melloni RH., Jr Chronická anabolická a androgenní steroidní léčba během dospívání zvyšuje přední hypotalamický vazopresin a agresi u intaktních křečků. Psychoneuroendokrinologie. 2000;25: 317-38. [PubMed]
- J J, posádky FT. Neurogenesa klesá během dospívání mozku od dospívání k dospělosti. Farmakologie, biochemie a chování. 2007;86: 327-33.
- Henderson LP. Steroidní modulace přenosu zprostředkovaného GABAA receptory v hypotalamu: účinky na reprodukční funkci. Neurofarmakologie. 2007;52: 1439-53. [PMC bezplatný článek] [PubMed]
- Hull EM, Du J, Lorrain DS, Matuszewich L. Extracelulární dopamin v oblasti mediální preoptiky: důsledky sexuální motivace a hormonální kontroly kopulace. Journal of Neuroscience. 1995;15: 7465-7471. [PubMed]
- Hull EM, Meisel RL, Sachs BD. Mužské sexuální chování. V: Pfaff DW, Arnold AP, Etgen AM, Fahrbach SE, Rubin RT, editory. Hormony, mozky a chování. Academic Press; New York: 2002. str. 3-137.
- Hull EM, Wood RI, McKenna KE. Neurobiologie mužského pohlavního chování. V: Neill JD, redaktor. Fyziologie reprodukce. Vol. 1. Elsevier Press; New York: 2006. str. 1729-1824.
- Huttenlocher PR, Dabholkar AS. Regionální rozdíly v synaptogenezi v lidské mozkové kůře. Journal of Comparative Neurology. 1997;387: 167-78. [PubMed]
- Johnson LR, dřevo RI. Orální samo-podávání testosteronu u samců křečků. Neuroendokrinologie. 2001;73: 285-92. [PubMed]
- Johnston LD, O'Malley PM, Bachman JG. Střední studenti (publikace NIH č. 03-5375) I. Bethesda, MD: Národní ústav pro zneužívání drog; 2003. Monitorování výsledků budoucích národních průzkumů o užívání drog, 1975-2002.
- Kaiser Family Foundation. Americká dospívající sexuální aktivita. 2005. str. # 3040-02.
- Kashon ML, Hayes MJ, Shek PP, Sisk CL. Regulace imunoreaktivity mozkových androgenních receptorů androgenem u předpubertálních samců fretek. Biologie reprodukce. 1995;52: 1198-205. [PubMed]
- Klíčová slova: dopaminergní účinky po chronické léčbě nandrolonem vizualizovaným v mozku u potkanů pozitronovou emisní tomografií. MUDr. Pokrok v neuro-psychofarmakologii a biologické psychiatrii. 2002;26: 1303-8.
- Kindlundh AM, Lindblom J, Bergstrom L, Wikberg JE, Nyberg F. Anabolic-androgenní steroidní nandrolon dekanoát ovlivňuje hustotu dopaminových receptorů u mužského potkaního mozku. European Journal of Neuroscience. 2001;13: 291-6. [PubMed]
- Kindlundh AM, Lindblom J, Nyberg F. Chronické podávání s nandrolon dekanoátem indukuje změny v obsahu genového transkriptu dopaminových D (1) a D (2) -receptorů v mozku krys. Výzkum mozku. 2003;979: 37-42. [PubMed]
- King BE, Packard MG, Alexander GM. Affective properties of intra-medial preoptic area injekce testosteronu u samců potkanů. Neuroscience Letters. 1999;269: 149-52. [PubMed]
- Koob GF, Nestler EJ. Neurobiologie drogové závislosti. Journal of Neuropsychiatry & Clinical Neurosciences. 1997;9: 482-97. [PubMed]
- Kouri EM, Lukas SE, papež HG, Jr, Oliva PS. Zvýšená agresivní odpověď u mužských dobrovolníků po podání postupně rostoucích dávek testosteron-cypionátu. Závislost na drogách a alkoholu. 1995;40: 73-9. [PubMed]
- Kritzer MF. Dlouhodobá gonadektomie ovlivňuje hustotu tyrosinhydroxylázových, ale nikoliv dopamin-beta-hydroxylázových, cholin acetyltransferázových nebo serotoninových imunoreaktivních axonů v mediálních prefrontálních kortexích dospělých samců potkanů. Mozková kůra. 2003;13: 282-296. [PubMed]
- Kurling S, Kankaanpaa A, Ellermaa S, Karila T, Seppala T. Účinek subchronické léčby nandrolondemanoátem na dopaminergních a serotonergních neuronových systémech v mozku potkanů. Výzkum mozku. 2005;1044: 67-75. [PubMed]
- Lambert JJ, Belelli D, Peden DR, Vardy AW, Peters JA. Neurosteroidní modulace receptorů GABAA. Pokrok v neurobiologii. 2003;71: 67-80. [PubMed]
- Lenroot RK, Giedd JN. Vývoj mozku u dětí a dospívajících: pohledy z anatomické magnetické rezonance. Neurovědy a biologické chování. 2006;30: 718-29.
- Lesting J, Neddens J, Teuchert-Noodt G. Ontogeny dopaminové inervace v nucleus accumbens gerbilů. Výzkum mozku. 2005;1066: 16-23. [PubMed]
- Malayev A, Gibbs TT, Farb DH. Inhibice odpovědi NMDA pomocí pregnenolon-sulfátu odhaluje subtyp selektivní modulaci receptorů NMDA pomocí sulfátových steroidů. British Journal of Pharmacology. 2002;135: 901-9. [PMC bezplatný článek] [PubMed]
- Martinez M, Guillen-Salazar F, Salvador A, Simon VM. Úspěšná intermální agrese a podmíněná preference místa u myší. Fyziologie a chování. 1995;58: 323-8. [PubMed]
- Meek LR, Romeo RD, Novak CM, Sisk CL. Aktivity testosteronu u křečků předepubertálních a postpubertálních samčích křečků: disociace účinků na reprodukční chování a imunoreaktivita androgenního receptoru mozku. Hormony a chování. 1997;31: 75-88.
- Meisel RL, Joppa MA. Podmínka preferovaných míst pro křečci po agresivních nebo sexuálních setkáních. Fyziologie a chování. 1994;56: 1115-8. [PubMed]
- Melloni RH, Jr, Connor DF, Hang PT, Harrison RJ, Ferris CF. Anabolicko-androgenní expozice steroidů během dospívání a agresivní chování u zlatých křečků. Fyziologie a chování. 1997;61: 359-64. [PubMed]
- Menard C, Harlan R. Upravení imunoreaktivity androgenového receptoru v mozku krys androgen-anabolickými steroidy. Výzkum mozku. 1993;622: 226-236. [PubMed]
- Mermelstein PG, Becker JB, Surmeier DJ. Estradiol snižuje proud vápníku v neostriálních neuronech krys přes membránový receptor. Journal of Neuroscience. 1996;16: 595-604. [PubMed]
- Midgley SJ, Heather N, Davies JB. Úrovně agresivity u skupiny uživatelů anabolických a androgenních steroidů. Medicína, věda a právo. 2001;41: 309-14.
- Molenda-Figueira HN, Salas-Ramirez KY, Schulz KM, Zehr JL, Montalto PR, Sisk CL. Sociální zkušenost dospívajících obnovuje dospělé ejakulační chování u mužských syrských křečků, kteří postrádají pubertální testosteron. Společnost pro behaviorální neuroendokrinologii; Pacific Grove, CA: 2007.
- Noble RG, Alsum PB. Hormonálně závislý sexuální dimorfismus u zlatého křečka (Mesocricetus auratus) Fyziologie a chování. 1975;14: 567-74. [PubMed]
- Nunez JL, Lauschke DM, Juraská JM. Smrt buněk ve vývoji zadní kůry u samců a samic potkanů. Journal of Comparative Neurology. 2001;436: 32-41. [PubMed]
- Nunez JL, Sodhi J, Juraská JM. Ovariální hormony po postnatálním dni 20 snižují počet neuronů v primární vizuální kůře potkanů. Journal of Neurobiology. 2002;52: 312-21. [PubMed]
- O'Connor DB, Archer J, Wu FC. Účinky testosteronu na náladu, agresi a sexuální chování u mladých mužů: dvojitě zaslepená placebem kontrolovaná studie. Journal of Clinical Endocrinology & Metabolism. 2004;89: 2837-45. [PubMed]
- Packard MG, Cornell AH, Alexander GM. Odměňování afektivních vlastností intra-nucleus accumbens injekcí testosteronu. Behaviorální neurovědy. 1997;111: 219-24. [PubMed]
- Parfitt DB, Thompson RC, Richardson HN, Romeo RD, Sisk CL. GnRH mRNA se zvyšuje s pubertem u mužského syrského křeččího mozku. Journal of Neuroendocrinology. 1999;11: 621-7. [PubMed]
- Parrott AC, Choi PY, Davies M. Použití anabolických steroidů amatérskými sportovci: účinky na psychologické náladové stavy. Journal of Sports Medicine & Physical Fitness. 1994;34: 292-8. [PubMed]
- Patton PE, Nový MJ, Lee DM, Hickok LR. Diagnóza a reprodukční výsledek po chirurgickém ošetření úplné dělohy, duplikovaného děložního čípku a vaginálního septa. American Journal of Obstetrics & Gynecology. 2004;190: 1669-75. 1675-8. [PubMed]
- Paus T, Collins DL, Evans AC, Leonard G., Pike B, Zijdenbos A. Maturace bílé hmoty v lidském mozku: přehled studií magnetické rezonance. Brain Research Bulletin. 2001;54: 255-66. [PubMed]
- Perry PJ, Kutscher EC, Lund BC, Yates WR, Holman TL, Demers L. Míra agrese a změny nálady u mužských vzpomínek s použitím androgenních anabolických steroidů a bez nich. Věstník forenzních věd. 2003;48: 646-51. [PubMed]
- Peters KD, dřevo RI. Androgenní závislost u křečků: předávkování, tolerance a potenciální opioidní mechanismy. Neurovědy. 2005;130: 971-81. [PubMed]
- Pfaus JG, Damsma G, Nomikos GG, Wenkstern DG, Blaha CD, Phillips AG, Fibiger HC. Sexuální chování zvyšuje centrální přenos dopaminu u samců potkanů. Výzkum mozku. 1990;530: 345-348. [PubMed]
- Phoenix CH, Goy RW, Gerall AA, Mladé WC. Organizace účinku prenatálně podávaného testosteron propionátu na tkáně zprostředkování páření chování u ženského morčete. Endokrinologie. 1959;65: 369-82. [PubMed]
- Pinos H, Collado P, Rodriguez-Zafra M, Rodriguez C, Segovia S, Guillamon A. Rozvoj pohlavních rozdílů v lokusu coeruleus krysy. Brain Research Bulletin. 2001;56: 73-8. [PubMed]
- Papež HG, Jr, Katz DL. Vražda a vražda u uživatelů anabolických steroidů [viz komentář] Journal of Clinical Psychiatry. 1990;51: 28-31. [PubMed]
- Papež HG, Jr, Katz DL. Psychiatrické a lékařské účinky užívání anabolicko-androgenních steroidů. Kontrolovaná studie sportovců 160. Archivy obecné psychiatrie. 1994;51: 375-82. [PubMed]
- Papež HG, Jr, Kouri EM, Powell KF, Campbell C, Katz DL. Používání anabolicko-androgenních steroidů u vězňů 133. Komplexní psychiatrie. 1996;37: 322-7. [PubMed]
- Putnam SK, Du J, Sato S, Hull EM. Testosteronové obnovení kopulačního chování koreluje s uvolňováním mediálních preoptických dopaminů u kastrovaných samců potkanů. Hormony a chování. 2001;39: 216-224. [PubMed]
- Putnam SK, Sato S, Hull EM. Účinky metabolitů testosteronu na kopulaci a mediální předoptické uvolňování dopaminu u kastrovaných samců potkanů. Hormony a chování. 2003;44: 419-26. [PubMed]
- Putnam SK, Sato S, Riolo JV, Hull EM. Účinky metabolitů testosteronu na kopulaci, mediální preoptický dopamin a NOS-imunoreaktivita u kastrovaných samců potkanů. Hormony a chování. 2005;47: 513-522. [PubMed]
- Rankin SL, Partlow GD, McCurdy RD, Giles ED, Fisher KR. Postnatální neurogeneze v vasopresinu a jádru obsahujícím oxytocin v prasečím hypotalamu. Výzkum mozku. 2003;971: 189-96. [PubMed]
- Ricci LA, Rasakham K, Grimes JM, Melloni RH. Jr Serotonin-1A receptorová aktivita a exprese modulují adolescentní anabolickou / androgenní steroidem indukovanou agresi u křečků. Farmakologie, biochemie a chování. 2006;85: 1-11.
- Romeo RD, Cook-Wiens E, Richardson HN, Sisk CL. Dihydrotestosteron aktivuje sexuální chování u dospělých samců křečků, ale ne u mladistvých. Fyziologie a chování. 2001;73: 579-84. [PubMed]
- Romeo RD, Diedrich SL, Sisk CL. Imunoreaktivita estrogenového receptoru u předpubertálních a dospělých mužských syrských křečků. Neuroscience Letters. 1999;265: 167-70. [PubMed]
- Romeo RD, Parfitt DB, Richardson HN, Sisk CL. Feromony vyvolávají ekvivalentní hladiny Fos-imunoreaktivity u předpubertálních a dospělých samčích sýrských křečků. Hormony a chování. 1998;34: 48-55. [PubMed]
- Romeo RD, Richardson HN, Sisk CL. Puberty a zrání mužského mozku a sexuálního chování: přepracování behaviorálního potenciálu. Neurovědy a biobehaviorální recenze. 2002a;26: 381-91. [PubMed]
- Romeo RD, Wagner CK, Jansen HT, Diedrich SL, Sisk CL. Estradiol indukuje hypotalamické progesteronové receptory, ale neaktivuje chování pohlavních orgánů u samců křečků (Mesocricetus auratus) před pubertou. Behaviorální neurovědy. 2002b;116: 198-205. [PubMed]
- Salas-Ramirez KY, Montalto PR, Sisk CL. Anabolické androgenní steroidy (AAS) mají rozdílný vliv na sociální chování dospívajících a dospělých mužských syrských křečků. Hormony a chování. 2008 v tisku.
- SAMHSA / OAS. DHHS publikace č. (SMA) 1996. 1994 Národní průzkum domácností o zneužívání drog, hlavní zjištění 1994; str. 96-3085.
- SAMHSA / OAS. Výsledky z Národního průzkumu 2004 o užívání drog a zdraví: Národní zjištění. 2005. Řada NSDUH H-28, publikace DHHS č. SMA 05-4062.
- Sato SM, Johansen J, Jordánsko CL, Wood RI. Androgenní samoadministrace u potkanů Tfm. 10th výroční zasedání společnosti pro behaviorální neuroendokrinologii.2006.
- Sato SM, Wood RI. Samostatné podávání anabolicko-androgenních steroidů (aas) nepropustných pro membránu u syrských křečků. 11th Výroční zasedání společnosti Fof Behavioral Neuroendocrinology.2007.
- Schroeder JP, Packard MG. Role podtypů dopaminových receptorů při získávání kondicionovaných podmínek testosteronu u potkanů. Neuroscience Letters. 2000;282: 17-20. [PubMed]
- Schulte HM, Hall MJ, Boyer M. Domácí násilí spojené s užíváním anabolických steroidů. American Journal of Psychiatry. 1993;150: 348. [PubMed]
- Schulz KM, Menard TA, Smith DA, Albers HE, Sisk CL. Expozice testikulárních hormonů během dospívání organizuje chování bokového značení a vázání vazopresinového receptoru v bočním přepážce. Hormony a chování. 2006;50: 477-83. [PubMed]
- Schulz KM, Richardson HN, Romeo RD, Morris JA, Lookingland KJ, Sisk CL. Mediální preoptická oblast dopaminergních odpovědí na feromony žen se během puberty vyvine u samčího křeččího křečka. Výzkum mozku. 2003;988: 139-45. [PubMed]
- Schulz KM, Richardson HN, Zehr JL, Osetek AJ, Menard TA, Sisk CL. Gonadální hormony maskulinizují a defeminizují reprodukční chování během puberty u mužského syrského křečka. Hormony a chování. 2004;45: 242-9. [PubMed]
- Schulz KM, Sisk CL. Pubertalní hormony, dospívající mozek a zrání společenského chování: poučení ze syrského křečka. Molekulární a buněčná endokrinologie. 2006:254–255. 120–6.
- Schulz KM, Zehr JL, Salas-Ramirez KY, Sisk CL. Plánovač setkání s neurovědy. Sandiego, CA: Společnost pro neurovědy; 2007. Je dospívání druhým citlivým obdobím pro organizační účinky testosteronu na dospělé reprodukční chování mužů? 2007 Online.
- Scott JP, Stewart JM, De Ghett VJ. Kritická období v organizaci systémů. Vývojová psychobiologie. 1974;7: 489-513. [PubMed]
- Shughrue PJ, Lane MV, Merchenthaler I. Srovnávací distribuce estrogenového receptoru alfa a beta-mRNA v centrální nervové soustavě potkana. Journal of Comparative Neurology. 1997;388: 507-25. [PubMed]
- Simons-Morton BG, Haynie DL. Psychosociální prediktory zvýšené fáze kouření u šestého srovnávače. American Journal of Health Chování. 2003;27: 592-602. [PubMed]
- Sisk CL, Zehr JL. Pubertalní hormony organizují dospívající mozky a chování. Hranice v neuroendokrinologii. 2005;26: 163-74. [PubMed]
- Sowell ER, Thompson PM, Leonard CM, Vítejte SE, Kan E, Toga AW. Podélné mapování tloušťky kortiků a růstu mozku u normálních dětí. Journal of Neuroscience. 2004;24: 8223-31. [PubMed]
- Sowell ER, Thompson PM, Tessner KD, Toga AW. Mapování pokračujícího růstu mozku a snižování hustoty šedé hmoty v dorzální frontální kůře: Inverzní vztahy během dozadolescentního zrání mozku. Journal of Neuroscience. 2001;21: 8819-29. [PubMed]
- Spear LP. Adolescentní mozkové a věkové behaviorální projevy. Neurovědy a biologické chování. 2000;24: 417-63.
- Su TP, Pagliaro M, Schmidt PJ, Pickar D, Wolkowitz O, Rubinow DR. Neuropsychiatrické účinky anabolických steroidů u mužských normálních dobrovolníků. JAMA. 1993;269: 2760-4. [PubMed]
- Thiblin I, Finn A, Ross SB, Stenfors C. Zvýšení dopaminergních a 5-hydroxytryptaminergních aktivit u mužského potkaního mozku po dlouhodobé léčbě anabolickými androgenními steroidy. British Journal of Pharmacology. 1999;126: 1301-6. [PMC bezplatný článek] [PubMed]
- Thomas P, Dressing G, Pang Y, Berg H, Tubbs C, Benninghoff A, Doughty K. Progestin, estrogenní a androgenní G-proteinové receptory v gonádách ryb. Steroidy. 2006;71: 310-6. [PubMed]
- Tricker R, Casaburi R, Storer TW, Clevenger B, Berman N, Shirazi A, Bhasin S. Účinky suprafyziologických dávek testosteronu na rozzlobené chování u zdravých eugonadálních mužů - studie klinického výzkumného centra. Journal of Clinical Endocrinology & Metabolism. 1996;81: 3754-8. [PubMed]
- Triemstra JL, Nagatani S, dřevo RI. Chemozenzorické náznaky jsou nezbytné pro uvolňování dopaminu vyvolaného pářením v MPOA samčích syrských křečků. Neuropsychopharmacology. 2005;30: 1436-42. [PubMed]
- Triemstra JL, Sato SM, dřevo RI. Testosteron a nucleus accumbens dopamina u mužského syrského křečka. Psychoneuroendokrinologie (v tisku)
- Van Eenoo P, Delbeke FT. Výskyt dopingu ve Flandrech ve srovnání s prevalencí dopingu v mezinárodních sportech. Mezinárodní žurnál sportovní medicíny. 2003;24: 565-570. [PubMed]
- WADA. Nepříznivé analytické nálezy hlášené akreditovanými laboratořemi. 2006. http://www.wada-ama.org.
- Wesson DW, McGinnis MY. Stohování anabolických androgenních steroidů (AAS) během puberty u potkanů: neuroendokrinní a behaviorální hodnocení. Farmakologie, biochemie a chování. 2006;83: 410-9.
- Wichstrom L, Pedersen W. Použití anabolicko-androgenních steroidů v dospívání: vítězství, dobré vyhlídky nebo špatné? Věstník studií o alkoholu. 2001;62: 5-13. [PubMed]
- Wood RI, Johnson LR, Chu L, Schad C, Self DW. Zesilování testosteronu: intravenózní a intracerebroventrikulární samoadministrace u potkanů samců a křečků. Psychopharmacology. 2004;171: 298-305. [PubMed]
- Wood RI, Newman SW. Androgenní a estrogenní receptory koexistují s jednotlivými neurony v mozku syrského křečka. Neuroendokrinologie. 1995;62: 487-97. [PubMed]
- Dřevo RI, Swann JM. Neuronální integrace chemosenzorických a hormonálních signálů, které kontrolují mužské sexuální chování. V: Wallen K, Schneider JS, redaktoři. Reprodukce v kontextu. MIT Press; Cambridge: 1999. str. 423-444.
- Zahm DS, Heimer L. Dvě transpelidální dráhy pocházející z krys jádra accumbens. Journal of Comparative Neurology. 1990;302: 437-46. [PubMed]