 Fabrizio Sanna1*,
Fabrizio Sanna1*,  Jessica Bratzu1,
Jessica Bratzu1,  Maria A. Piludu2,
Maria A. Piludu2,  Maria G. Corda2,
Maria G. Corda2,  Maria R. Melisová1,
Maria R. Melisová1,  Osvaldo Giorgi2 a
Osvaldo Giorgi2 a  Antonio Argiolas1,3
Antonio Argiolas1,3- 1Oddělení biomedicínských věd, Sekce neurovědy a klinické farmakologie a Centrum excelence pro neurobiologii závislostí, Univerzita v Cagliari, Cagliari, Itálie
- 2Oddělení životního a environmentálních věd, Sekce farmaceutických, farmakologických a nutričních věd, Univerzita v Cagliari, Cagliari, Itálie
- 3Ústav neurovědy, Národní rada pro výzkum, Cagliari sekce, Cittadella Universitaria, Cagliari, Itálie
Řízené krysy s vysokou (RHA) a nízkou mírou vyhýbání se (RLA), které se liší v případě rychlé vs. nedostatečné odezvy aktivní odezvy v kyvadlové schránce, vykazují rozdíly v sexuální aktivitě, receptivní samice krysy. Potkani RHA skutečně vykazují vyšší míru sexuální motivace a kopulační výkon než krysy RLA, které přetrvávají i po opakované sexuální aktivitě. Tyto rozdíly byly korelovány s vyšším tónem mezolimbického dopaminergního systému potkanů RHA proti krysám RLA, což bylo zřejmé z vyššího zvýšení dopaminu zjištěného v dialyzátu získaném z nucleus accumbens RHA než u potkanů RLA během sexuální aktivity. Tato práce ukazuje, že extracelulární dopamin a noradrenalin (NA) také zvyšují dialyzát z mediální prefrontální kůry (mPFC) samců potkanů RHA a RLA, a to v přítomnosti nepřístupných samic potkanů a výrazněji během přímé sexuální interakce. Takové zvýšení dopaminu (a jeho hlavního metabolitu 3,4-dihydroxyfenyloctová kyselina, DOPAC) a NA byly zjištěny u obou pohlavně naivních a zkušených zvířat, ale byly vyšší: (i) u RHA než u potkanů RLA; a (ii) u sexuálně zkušených potkanů RHA a RLA než v jejich naivních protějšcích. Konečně, rozdíly v dopaminu a NA v mPFC se vyskytovaly souběžně s těmi v sexuální aktivitě, protože potkani RHA vykazovali vyšší úrovně sexuální motivace a kopulační výkon než krysy RLA v sexuálně naivní a zkušené populaci. Tyto výsledky naznačují, že v mPFC se vyskytuje vyšší dopaminergní tón společně se zvýšeným noradrenergním tónem, který se může podílet na různých kopulačních vzorcích zjištěných u potkanů RHA a RLA, jak bylo navrženo pro mezolimbický dopaminergní systém.
Úvod
Řízené linie potkanů s vysokou (RHA) a nízkou mírou vyhýbání se (RLA), původně vybrané pro rychlé a špatné navázání aktivní odezvy v kyvadlové skříni (Bignami, 1965; Broadhurst a Bignami, 1965; Driscoll a Bättig, 1982; Fernández-Teruel a kol., 2002; Giorgi a kol., 2007) vykazují signifikantní rozdíly v sexuální aktivitě, pokud jsou vystaveny přítomnosti sexuálně vnímavé samice potkanů. Potkani RHA vykazují vyšší míru sexuální motivace než krysy RLA, jak je patrné z vyššího počtu erekcí bez kontaktu penisu, které tyto krysy vykazují při přítomnosti nepřístupné receptivní ženy, a lepší kopulační výkony než krysy RLA, jak se ukázalo významnými rozdíly v počtu kopulačních parametrů zjištěných hlavně v prvním kopulačním testu mezi dvěma krysími liniemi, ale také po pěti kopulačních testech, ačkoli oslabených (Sanna a kol., 2014a). Mezi nejzřetelnější rozdíly zjištěné mezi krysy RHA a RLA je procento krys, které iniciují montáž a intromitáž a které v první kopulační zkoušce ejakulují, 80% potkanů RHA proti 40% potkanů RLA v uvedeném pořadí (Sanna a kol., 2014a). Velké množství experimentálních důkazů naznačuje, že funkční odlišný dopaminergní tón se podílí na kopulačních rozdílech mezi potkany RHA a RLA. Ve skutečnosti bylo zjištěno, že různé kopulační vzory krys RHA a RLA jsou různě modifikovány apomorfinem, smíšeným agonistou dopaminového receptoru D1 / D2 podávaným v dávkách, které usnadňují sexuální chování, a haloperidolem, antagonistou receptoru dopaminového D2 podávaným na nízkých dávek, které brání sexuálnímu chování (Sanna a kol., 2014b). V souladu s tím bylo zjištěno, že krysy RLA jsou citlivější na usnadnění a inhibici sexuálního chování indukovaného apomorfinem a haloperidolem, což je prokázáno většími modifikacemi vyvolanými oběma léky, zejména při podávání v nižších dávkách v různých kopulačních parametrech RLA u potkanů RHA (Sanna a kol., 2014b). Snad důležitější pro tuto studii jsou různé kopulační vzorce krys RHA a RLA také příbuzné rozdílu v aktivitě mezolimbických dopaminergních neuronů, jejichž aktivita je dobře známa během sexuálního chování (Pfaus a kol., 1990; Pleim a kol., 1990; Pfaus a Phillips, 1991; Damsma a kol., 1992; Wenkstern a kol., 1993; Balfour a spol., 2004; Pitchers a kol., 2010, 2013; Beloate a kol., 2016). Navzdory skutečnosti, že potíže s RHA a RLA, kteří měli sexuální a sexuální zkušenost, měli podobné bazální hodnoty extracelulárního dopaminu v dialyzátu z nucleus accumbens, koncentrace extracelulárního dopaminu a kyseliny 3,4-dihydroxyfenyloctové (DOPAC, jeden z jeho hlavních metabolitů ), se zjistilo, že v dialyzátech získaných z shell shell nucleus accumbens naivních a sexuálně zkušených potkanů RHA a RLA diferencovaně vzrůstá jak v předvídatelné, tak v konzumní fázi sexuálního chování. Výše uvedené rozdíly byly výraznější mezi sexeálně naivními potkany RHA a RLA, ale přetrvávaly mezi sexuálně zkušenými potkany RHA a RLA, i když mají tendenci k poklesu u těchto potkanů, jak bylo zjištěno s rozdíly v sexuálním chování (Sanna a kol., 2015).
Funkční role zvyšování dopaminergní aktivity v nucleus accumbens během sexuální aktivity (ať už v apetitivní a konzumační fázi sexuálního chování) je stále předmětem debaty, stejně jako to, co se projevuje při krmení, které má stejně jako sexuální aktivita silný motivační valence. Takže i když dopamin v nucleus accumbens je zapojen do motivace a mezolimbické dopaminergní neurony jsou obvykle označovány jako odměňující neurony, v posledních letech 15, nedávné studie podporují hypotézu, že mezolimbický dopamin není zapojen do primárního vyjádření motivovaného nebo odměňujícího chování, ale spíše s učením a vzpomínkou na sdružení stimulů a odměn (Agmo et al., 1995; Berridge a Robinson, 1998; Ikemoto a Panksepp, 1999; Pitchers a kol., 2013, 2014; Beloate a kol., 2016; Salamone a spol., 2016). V souladu s touto hypotézou byla nedávno zjištěna blokáda dopaminových receptorů v nucleus accumbens nebo inaktivace dopaminergních neuronů ve ventrální tegmentální oblasti neschopná změnit expresi apetitivních a konzumačních aspektů kopulačního chování u samců potkanů (Pitchers a kol., 2013, 2014; Beloate a kol., 2016).
Úloha jiných oblastí mozku obsahujících dopamin ve výše uvedených rozdílech v sexuálním chování mezi krysy RHA a RLA však nelze vyloučit. Proto je dobře známo, že dopamin působí na preventivní a konzumační fáze sexuálního chování u laboratorních zvířat a také u člověka nejen v nucleus accumbens (Everitt, 1990; Pfaus a kol., 1990; Hull a kol., 1991; Pfaus a Everitt, 1995; Melis a Argiolas, 2011), ale také v jiných oblastech mozku, jako je mediální preoptická oblast, hypotalamus a jeho jádra (tj. paraventrikulární jádro (PVN)); Pfaus a Phillips, 1991; Argiolas a Melis, 1995, 2005, 2013; Hull a kol., 1995, 1999; Melis a Argiolas, 1995; Melis a kol., 2003; Succu a kol., 2007; Pfaus, 2010). Další oblast, která obsahuje dopamin a může hrát roli v sexuálním chování, je mediální prefrontální kůra (mPFC; Fernández-Guasti a kol., 1994; Agmo a Villalpando, 1995; Agmo et al., 1995; Hernández-Gonzáles a kol., 1998, 2007; Kakeyama a kol., 2003; Balfour a spol., 2006; Afonso a kol., 2007; Davis a kol., 2010; Febo, 2011). Ve skutečnosti se v této oblasti mozku objevují nervové zakončení mesokortikálních dopaminových neuronů, jejichž buněčné tělíska jsou lokalizována ve ventrální tegmentální oblasti jako mesolimbické dopaminové neurony. Pokud jde o nucleus accumbens, přesná role této oblasti mozku v sexuálním chování není zdaleka jasná. Opravdu, léze mPFC jsou obvykle zjištěny jako neschopné změnit sexuální chování samců krys s sexuálně vnímavou ženou (Fernández-Guasti a kol., 1994; Agmo a Villalpando, 1995; Agmo et al., 1995; Hernández-Gonzáles a kol., 1998, 2007; Kakeyama a kol., 2003; Balfour a spol., 2006; Afonso a kol., 2007), ani vyjádření podmíněného místa pro sexuální odměnu (Davis a kol., 2010). Nicméně léze mPFC, které nezměnily vzhled podmíněné preference místa pro sexuální odměnu, zrušily u stejných zvířat schopnost vytvářet podmíněnou averzi vůči sexuální aktivitě, když spárovány s averzními podněty (Davis a kol., 2010) a vybírání selektivních buněk při přiblížení se k chování samčích krys směrem k nepřístupným samičím ženám, které jsou sexuálně vnímavé, byly měřeny v mPFC samců potkanů (Febo, 2011). Tato zjištění vedla k názoru, že aktivace mPFC během sexuálního chování hraje roli při integraci externích a interních informací pro výkon a kontrolu cílového chování spíše než ve vyjádření vrozených reakcí na přirozené posilovače (viz Goto a Grace, 2005). V souladu s tím je mPFC společně s jádrem accumbens součástí komplexního nervového systému, který se podílí na modulaci motivovaného chování (cílové chování), což vyžaduje integraci kognitivních informací z mPFC, emoční informace z amygdaly a kontextu- souvisejících informací z hipokampu, v nucleus accumbens (Goto a Grace, 2005).
Experimentální důkazy naznačují, že uvolňování dopaminu v mPFC se podílí na adaptivní regulaci motivovaného chování a zdá se, že deregulace těchto mechanismů hraje roli v patologických nebo maladaptivních stavech, jako jsou psychiatrické poruchy, jako schizofrenie, deficity pozornosti a hyperaktivní poruchy ( ADHD), deprese (Dunlop a Nemeroff, 2007; Masana a kol., 2011) nebo zneužívání návykových látek a chování (Everitt a Robbins, 2005). Je zajímavé, že aktivita dopaminu uvolňovaného v mPFC může být ovlivněna noradrenalinem (NA), který je přítomen v mPFC na vyšších úrovních než dopamin a zejména NA transportérem (NET), který je nejen bohatší než dopaminu (DAT) v mPFC (Carboni a kol., 1990, 2006; Gresch a kol., 1995; Westernik a kol., 1998), ale také vykazuje afinitu k dopaminu ještě vyšší než u NA (Horn, 1973). Vzhledem k tomu, že dopamin a NA často spolupracují v mnoha funkcích mPFC, od pracovní paměti a tvorby pozorovacích souborů až po reverzní učení, inhibici odezvy a reakci na stres (viz Robbins a Arnsten, 2009), to vyvolává možnost, že dopamin, samotný nebo společně s NA, v mPFC může také hrát roli v rozdílech v chování mezi krysy RHA a RLA, včetně těch, které se nacházejí v sexuálním chování.
Pro testování této hypotézy byla studována aktivita mezokortikálního dopaminergního systému a noradrenergního systému v mPFC ve dvou RHA a RLA liniích potkana pomocí intracerebrální mikrodialýzy. Stručně řečeno, dopamin (a jeho hlavní metabolit DOPAC) a NA byly měřeny v dialyzátech získaných z prelimických (PrL) a infralimbických (IL) kompartmentů mPFC sexuálně nevhodných (např. Nikdy vystavených receptivní ženě) RLA potkanů (např., Kteří podstoupili pět předběžných kopulačních testů a vykazovali konstantní hladiny kopulační aktivity), když se v přítomnosti nepřístupné receptivní samice a při přímé sexuální interakci pomocí vysokotlaké kapalinové chromatografie spojené s elektrochemickou detekcí (HPLC-ECD).
Materiály a metody
Zvířata
Samci potkanů RHA a RLA s nadbytkem (N = 30 pro každý řádek, vážící ≈300 g na začátku experimentální práce) byly všechny z kolonie založené v 1998 na univerzitě v Cagliari v Itálii (Giorgi a kol., 2007). Postupy používané pro selektivní chov sardinské kolonie byly již podrobně popsány (Giorgi a kol., 2005).
Obyvatelé Envigo (San Pietro al Natisone, Itálie), kteří byli použiti ve všech experimentech, použili ovariekomizované stimulační SD samice potkanů (250-300 g na začátku experimentální práce). Zvířata byla aklimatizována čtyři na klec (38 cm × 60 cm × 20 cm) do zařízení pro bydlení oddělení biomedicínských věd Univerzity v Cagliari minimálně 10 dní před zahájením experimentů při teplotě 24 ° C, vlhkosti 60%, obrácený cyklus 12 h světlo / tma (zhasne 08: 00 h na 20: 00 h), voda a standardní laboratorní potraviny podle libosti. Zvířata byla denně manipulována s 1-2 min během habituation období, aby se omezil manipulační stres během experimentů; Navíc kontakt s personálem údržby zvířete byl omezen na jediného ošetřovatele a lůžko v domácích klecích se nikdy nezměnilo ani před nebo v den pokusů. Všechny experimenty byly provedeny mezi 10: 00 h a 18: 00 h. Tato studie byla provedena v souladu s doporučeními směrnic Evropských společenství, směrnice ze září 22, 2010 (2010 / 63 / EU) a italskými právními předpisy (DL březen 4, 2014, č. 26). Protokol byl schválen Etickou komisí pro experimentování na zvířatech na univerzitě v Cagliari (autorizační číslo 361 / 2016-PR, duben 08, 2016 na FS).
Experimentální skupiny
Byly použity sexuálně naivní a sexuálně zkušené samce potkanů RHA a RLA. Sexuálně naivní potkani byli potkani nikdy vystaveni sexuálně vnímavému ovariektomizovanému a estradiolu + progesteronu primárně-ženskému; sexuálně zkušení potkani byli potkani, kteří již podstoupili pět po sobě následujících kopulačních testů 60 min v intervalech dnů 3 s receptivní samicí (Sanna a kol., 2014a,b). Ženy se přiváděly do estra léčbou subkutánním estradiolbenzoátem (200 μg / krysa v podzemnicovém oleji) a progesteronem (0.5 mg / potkan v podzemnicovém oleji), 48 h a 6 h před kopulačními testy. Oestrus byl ověřen barvením May-Grunwald-Giemsa a mikroskopickým vyšetřením vaginálních nátěrů 1 h před experimenty. V souladu s předchozími studiemi (Sanna a kol., 2014a,b, 2015) bylo zjištěno, že pět předběžných kopulačních testů postačuje k tomu, aby samci římských krys obou linií vykazovali konstantní hladiny kopulační aktivity: např. sexuálně zkušení krysy RHA a RLA splňovaly kritérium alespoň jedné ejakulace dosažené v každém z posledních dvou testů (jedna krysa RHA a dva krysy RLA, které toto kritérium nesplnily, byly v této fázi vyřazeny). Dva dny po těchto předběžných kopulačních testech podstoupili sexuálně zkušené římské krysy stereotaktickou chirurgii pro implantaci mikrodialyzované sondy v mPFC, jak je popsáno níže (Sanna a kol., 2015).
Mikrodialýza v mPFC během sexuálního chování
Den před mikrodialýzou byly na podkladě anestezie izofluranu (1.5% -2%, Harvard Apparatus, Holliston, MA, USA) umístěny v stereotaktickém zařízení (Stoelting Co., Wood Dale, IL, USA) a implantoval vertikální domácí mikrobiální sondu (dialyzační membrána ≈ 3 mm volného povrchu; Melis a kol., 2003) a jednostranně směřující na oddíly mPFC, PrL a IL (souřadnice: 3.0 mm přední a 0.7 mm boční až bregma a 5.5 mm ventrální k trvání; Paxinos a Watson, 2004). V den experimentu byla zvířata převedena v tmavé fázi cyklu do klece (45 cm × 30 cm × 24 cm), která byla umístěna v místnosti s ochranou proti hluku osvětlenou červeným světlem a uvnitř jiného malá plexiskla (15 cm × 15 cm × 15 cm) s otvory 25 (Ø 2 mm) ve svislých stěnách umožnila vizuální, čichovou a akustickou, ale ne přímou interakci. Po habituaci 2 h byla mikrobiální sonda připojena k mikroinfúzní pumpě CMA / 100 (Harvard Apparatus, Holliston, MA, USA) s polyethylenovou trubicí a perfundována Ringerovým roztokem (147 mM NaCl, 3 mM KCl a 1.2 mM CaCl2, pH 6.5) při průtoku 2.5 μl / min. Po ekvilibrační době 2 h perfuzního média s extracelulární kapalinou byly alikvotní podíly dialyzátu 37.5 μL shromážděny každých 15 min během experimentu v ledově ochlazených polyethylenových trubicích pro měření koncentrace dopaminu, DOPAC a NA, jak je popsáno níže. Po odebrání nejméně čtyř alikvotů dialyzátu byla do malé klece umístěné uvnitř páry klece zavedena receptivní samice potkana pro 30 min. Během těchto 30 min byly shromážděny další dva alikvotní části dialyzátu. Za těchto podmínek samci krysy nemohou přímo reagovat se samičí, ale vykazují bezkontaktní erekci (viz níže). Po uplynutí této doby byla odstraněna malá klece, kopulace byla povolena pro minuty 75 a bylo odebráno dalších pět alikvotů dialyzátu. Na konci tohoto období byla samice odstraněna z páry a byla přidána další alikvotní část dialyzátu (Pfaus a Everitt, 1995; Melis a kol., 2003; Sanna a kol., 2015). Během experimentu byly zaznamenány sexuální parametry vztahující se k předvídatelné a konzumující fázi sexuálního chování (viz níže).
Sexuální chování
Několik parametrů sexuální motivace a kopulačního výkonu souvisejících s předvídatelnou a konzumovanou fází sexuálního chování bylo zaznamenáno v průběhu celého experimentu pozorovatelem, který si nebyl vědom specifických experimentálních podmínek, např. Kdo neznal hranici a úroveň sexuální zkušenosti zvířat použitých v tomto experimentu (viz níže). Stručně řečeno, latence k první erekci bez kontaktu (NCPEL, časované od zavedení receptivní ženy ve vnitřní kleci) a jejich frekvence (NCPEF, počet nekontaktních erekcí penisu, ke kterým dochází v období, ve kterém je samice přítomné ve vnitřní kleci). Tyto feromonem zprostředkované erekce penisu, které se vyskytují u sexuálně silných samců potkanů za přítomnosti nepřístupné samice, jsou považovány za důležitý index sexuálního vzrušení (Sachs a kol., 1994; Sachs, 2000; Melis a kol., 2003). Pokud byla povolena sexuální interakce, např. Během kopulace, latence k uložení a intromitu (ML a IL, počínaje odstraněním vnitřní malé klece až do prvního úseku nebo do prvního intromise); frekvence montáže a intromitace (MF a IF, počet úchytů a intromise v první řadě kopulační aktivity a po celou dobu kopulace); latence k ejakulaci (EL, čas od prvního intromise první série až do ejakulace); byla zaznamenána frekvence ejakulace (EF, celkový počet ejakulací během kopulačního testu) a post-ejakulační interval (PEI, čas od první ejakulace až do příští intromise). Dále, kopulační účinnost (CE, počet intromise dané řady děleno součtem počtu úchytů a intromise ve stejné sérii) a interval inter-intromise (III, poměr mezi ejakulační latencí dané série a počet intromise v této sérii) byly také vypočteny pro první sérii kopulační aktivity (Sachs a Barfield, 1976; Meisel a Sachs, 1994; Melis a kol., 2003; Sanna a kol., 2014a,b, 2015).
Stanovení koncentrací dopaminu, DOPACu a noradrenalinu v dialyzátu z mPFC
Koncentrace dopaminu, DOPAC a NA byly měřeny v 20 μL stejného alikvotu dialyzátu z mPFC pomocí vysokotlaké kapalinové chromatografie (HPLC) spojené s elektrochemickou detekcí za použití dvojitého buňky 4011 (Coulochem II, ESA, Cambridge, MA, USA) jako již popsáno (Melis a kol., 2003). Detekce byla provedena v redukčním režimu při + 350 a -180 mV. HPLC byla vybavena kolonou Supelcosil C18 (7.5 cm x 3.0 mm id, velikost 3 μm, Supelco, Supelchem, Milán, Itálie), eluován 0.06 M citrát / acetát pH 4.2, obsahující methanol 20% v / v, 0.1 mM EDTA, 1 μM triethylaminu a 0.03 mM dodecylsulfátu sodného jako mobilní fáze, při průtoku 0.6 ml / min a teplotě místnosti. Citlivost testu byla 0.125 pg pro dopamin, 0.1 pg pro DOPAC a 0.2 pg pro NA.
Histologie
Na konci pokusů byly krysy usmrceny dekapitací, mozky byly okamžitě odstraněny z lebky a ponořeny do 4% vodného formaldehydu pro 12-15 dny. Po tomto období byly 40 μm koronální mozkové úseky připraveny zmrazením mikrotromu, barveny Neutrální červenou a zkontrolovány na mikroskopu s fázovým kontrastem. Pozice špičky sondy byla pak lokalizována v mPFC sledováním traktu sondy skrze řadu sekcí mozku (viz obrázek 1). Pouze krysy, u kterých bylo zjištěno, že aktivní část dialyzační membrány je správně umístěna v oddílech PrL a IL mPFC, byly vzaty v úvahu pro statistické vyhodnocení výsledků (jedna krysa z každé experimentální skupiny byla v této fázi vyřazena).
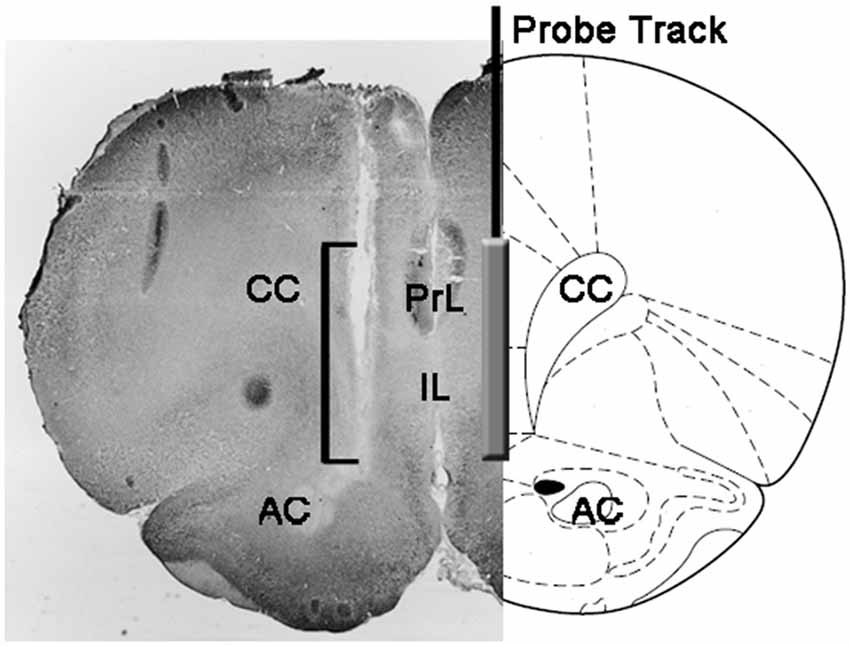 Obrázek 1. Schematické znázornění koronální sekce mozku krys vykazující stopu mikrodialyzované sondy v portech PrL a IL mediálního prefrontálního kortexu (mPFC; Paxinos a Watson, 2004). Čtvercový úhelník na mikrofotografii označuje část části Neutrální červené barvy, která zobrazuje aktivní část dialyzační membrány mikrodialyzované sondy. Zkratky: PrL, prelimbic oblast; IL, infralimbická oblast; AC, přední komise; CC, corpus callosum.
Obrázek 1. Schematické znázornění koronální sekce mozku krys vykazující stopu mikrodialyzované sondy v portech PrL a IL mediálního prefrontálního kortexu (mPFC; Paxinos a Watson, 2004). Čtvercový úhelník na mikrofotografii označuje část části Neutrální červené barvy, která zobrazuje aktivní část dialyzační membrány mikrodialyzované sondy. Zkratky: PrL, prelimbic oblast; IL, infralimbická oblast; AC, přední komise; CC, corpus callosum.
Statistika
Byly provedeny statistické analýzy biochemických (dopaminových, DOPAC a NA) a behaviorálních (NCPEL a NCPEF, ML, IL a EL, MF, IF a EF a PEI) buď zahrnující všechny experimentální subjekty (tradičně prováděné ve studiích samců potkanů kopulační chování) nebo s vyloučením těch subjektů, které během experimentu s mikrodialyzací nespolupracovaly na ejakulaci. V prvním případě, kdy byla všechna zvířata zařazena do analýzy, zvířata, která nezobrazovala NCPE v okamžiku, kdy byla samice nepřístupná nebo nebyla připojena nebo intromitována nebo ejakulována s dostupnou ženou, byly přiřazeny příslušné skóre plného rozsahu: 1800 s pokud muž nezobrazoval NCPE v minu 30, ve kterém byla žena nepřístupná; 900 s pro ML a IL, jestliže muž nezasáhl nebo nezadrčil s dostupnou ženou; 1800 s pro EL, jestliže muž nedosáhl ejakulace a 600 s pro PEI, jestliže muži neintrojovali po první ejakulaci. Ve druhém případě byla zvířata, která nebyla kopulována k ejakulaci, vyloučena z analýz. Toto bylo provedeno s cílem posoudit, zda mohou být rozdíly v sexuálním chování korelovány s rozdíly v koncentracích extracelulárního dopaminu, DOPAC a NA v mPFC římských linií potkanů během dvou fází sexuální aktivity vylučujících možnou matoucí interference v důsledku zařazení subjektů, které nevykazovaly sexuální chování tím, že jim přiřadily fixní hodnoty pro analyzované sexuální parametry.
Několik statistických analýz bylo provedeno pomocí ANOVA, aby bylo možné detekovat a lépe charakterizovat možné rozdíly mezi použitými experimentálními podmínkami. Před provedením testů ANOVA byly sady Bartlettových nebo Levineových testů v závislosti na případu prohlédnuty soubory dat každé z různých experimentálních proměnných pro homogenitu odchylek mezi čtyřmi experimentálními skupinami (prováděnými se všemi zvířaty nebo potkany kopulujícími pouze na ejakulaci). Když byly zjištěny významné rozdíly v odchylkách, datové soubory byly transformovány logaritmem (tj. Experimentální hodnoty Y byly změněny na hodnoty log Y), znovu zkontrolovány pro homogenitu odchylek a poté analyzovány pomocí ANOVA. Stručně řečeno, parametry chování získané dříve (např. S nepřístupnou samičí) a během první sérii kopulační aktivity (od prvního připojení / intromise k první mount / intromission po první ejakulaci) naivních a sexuálně zkušených samců RHA a RLA potkanů během mikrodialýzy byly analyzovány dvěma způsoby ANOVA, přičemž byla použita linie potkanů a úroveň sexuální zkušenosti mezi subjekty (průměrné hodnoty parametrů chování jsou uvedeny v tabulce 1 a F hodnoty a významové úrovně analýz ANOVA v tabulce 2).
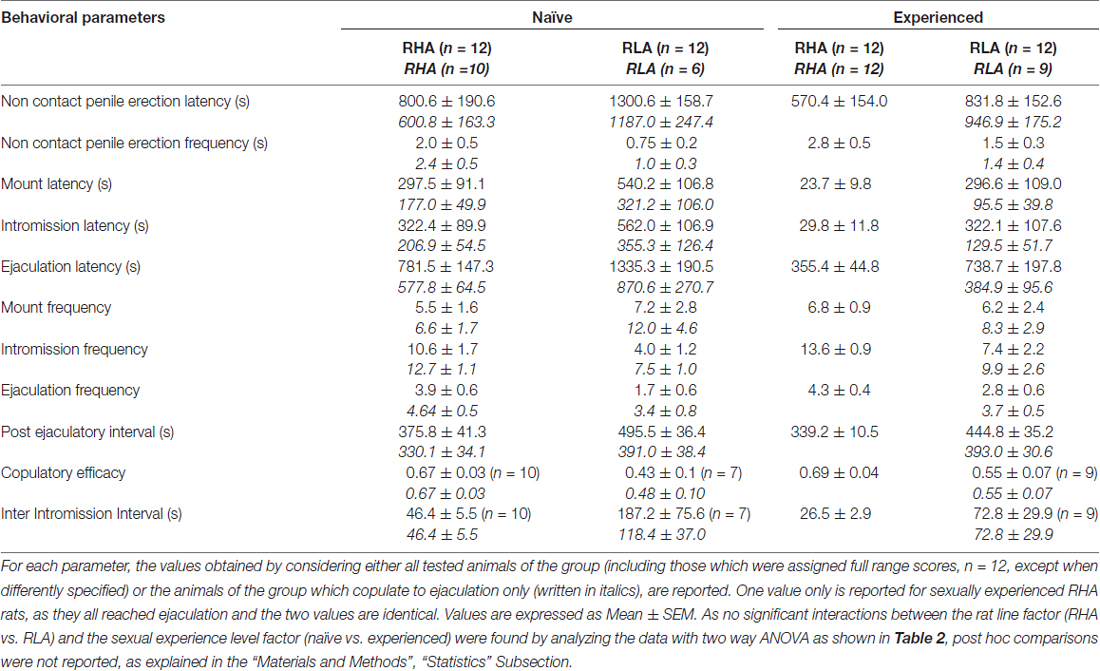 Tabulka 1. Parametry kopulace měřené v první sérii kopulační aktivity (která začíná první mount / intromission, když je samice zpřístupněna samci a končí po intervalu post-ejakulace s první mount / intrommision druhé série) a bezkontaktního penisu erekce sexuálně naivních a zkušených potkanů RHA a RLA.
Tabulka 1. Parametry kopulace měřené v první sérii kopulační aktivity (která začíná první mount / intromission, když je samice zpřístupněna samci a končí po intervalu post-ejakulace s první mount / intrommision druhé série) a bezkontaktního penisu erekce sexuálně naivních a zkušených potkanů RHA a RLA.
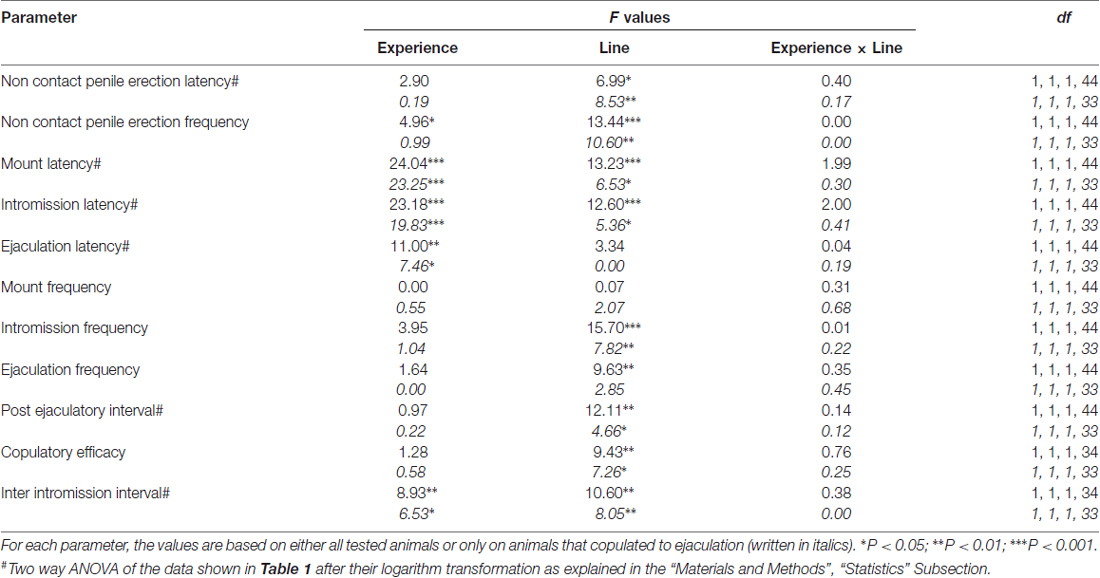 Tabulka 2. F hodnoty a úrovně významnosti dvoucestné ANOVA provedené na základě údajů uvedených v tabulce 1 (RHA vs. RLA) a úrovně sexuální zkušenosti (naivní vs. zkušené) mezi jednotlivými faktory.
Tabulka 2. F hodnoty a úrovně významnosti dvoucestné ANOVA provedené na základě údajů uvedených v tabulce 1 (RHA vs. RLA) a úrovně sexuální zkušenosti (naivní vs. zkušené) mezi jednotlivými faktory.
Kromě toho byla provedena celková analýza dat získaných od každého potkana během mikrodialýzy vypočtením nejprve hodnot AUC získaných vynesením hodnot koncentrací dopaminu, DOPAC nebo NA nebo počtu nekontaktních erekcí, mountů, intromise a ejakulací vs (začíná 1 h po zavedení samčích krys do páry do konce experimentu - 180 min rozděleno na frakce 15 min pro neurochemické hodnoty - nebo v době, kdy byla samice zavedena do páření klece pro parametry chování) a potom porovnáním vypočtených hodnot pomocí dvouvláknových ANOVA použitím linie potkanů a úrovně sexuální zkušenosti mezi faktory subjektů (průměrné hodnoty AUC neurochemických parametrů a parametrů chování jsou uvedeny v tabulce 4 a F hodnoty a významové úrovně analýz ANOVA v tabulce 5). Konečně byla provedena podrobnější analýza jednotlivých bodů (tj. Koncentrace dopaminu, DOPAC, NA a počet nekontaktních erekcí, mountů, intromise a ejakulace vs. čas) pomocí faktoriálních ANOVA pro opakovaná měření pomocí linie potkanů a úrovně sexuální zkušenosti mezi faktorem a časem (tj. Frakce dialyzátu) jako faktorů v rámci předmětu F hodnoty a úrovně významnosti těchto analýz ANOVA jsou uvedeny v tabulce 6). Podobné analýzy byly provedeny také s ohledem pouze na bazální hodnoty neurochemických parametrů (poslední čtyři alikvotní podíly dialyzátu shromážděné před zavedením do slepé klece receptivní samice), jejichž průměrné hodnoty jsou uvedeny v tabulce 3. Jako první, ale nikoli druhá, byla zjištěna interakce při provádění obecných factoriálních ANOVA pro opakovaná měření, zejména interakce Line × Time a Experience × Time pro neurochemické parametry (viz tabulka 6) byly provedeny obousměrné ANOVA s čárou nebo úrovní sexuální zkušenosti mezi subjektem a časem jako v rámci subjektivního faktoru na těchto souborech dat přímým srovnáním sexuálně naivních nebo zkušených potkanů RHA vs. RLA nebo sexuálně zkušených vs. naivních potkanů RHA nebo sexuálně zkušených vs. naivních potkanů RLA. Výsledky post hoc dvojice moudrých kontrastů provedených pomocí Tukeyho HSD testu na významných interakcích odhalených těmito obousměrnými ANOVA jsou uvedeny na obrázku 2. Ve všech ostatních případech, post hoc nebyly zaznamenány žádné srovnání, jelikož ANOVA nedokázaly odhalit žádnou významnou interakci mezi potkanovou linií, úrovní sexuální zkušenosti a časem. Statistická analýza byla provedena pomocí Graph Pad 5 (PRISM, San Diego, CA, USA) a STATISTICA 12 (Statsoft, Tulsa, OK, USA) s úrovní významu nastavenou na P <0.05.
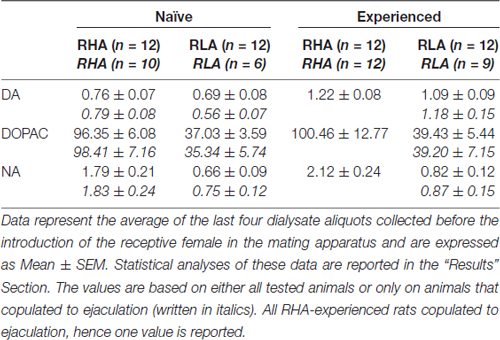 Tabulka 3. V bazálním dopaminu (DA), koncentracích 3,4-dihydroxyfenyloctové kyseliny (DOPAC) a noradrenalinu (nM) v dialyzátu z mediální prefrontální kůry (mPFC) sexuálně naivních a zkušených potkanů RHA a RLA.
Tabulka 3. V bazálním dopaminu (DA), koncentracích 3,4-dihydroxyfenyloctové kyseliny (DOPAC) a noradrenalinu (nM) v dialyzátu z mediální prefrontální kůry (mPFC) sexuálně naivních a zkušených potkanů RHA a RLA.
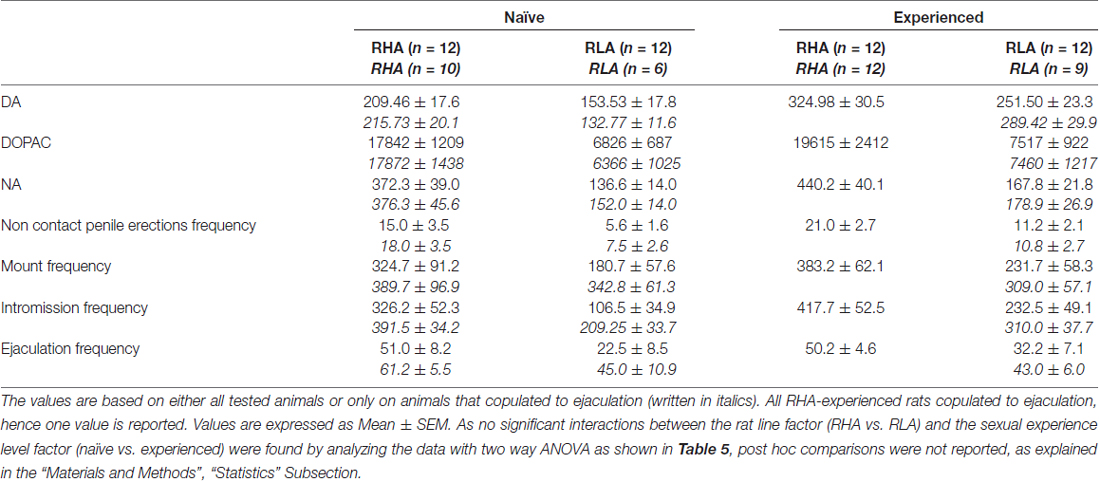 Tabulka 4. Celkové hodnocení rozdílů v koncentracích DA, DOPAC a NA a v sexuálním chování mezi pohlavně nespříznivými a zkušenými potkany RHA a RLA pomocí analýzy průměrných AUC získaných z výsledků ukázaných na obrázcích 2 a 3.
Tabulka 4. Celkové hodnocení rozdílů v koncentracích DA, DOPAC a NA a v sexuálním chování mezi pohlavně nespříznivými a zkušenými potkany RHA a RLA pomocí analýzy průměrných AUC získaných z výsledků ukázaných na obrázcích 2 a 3.
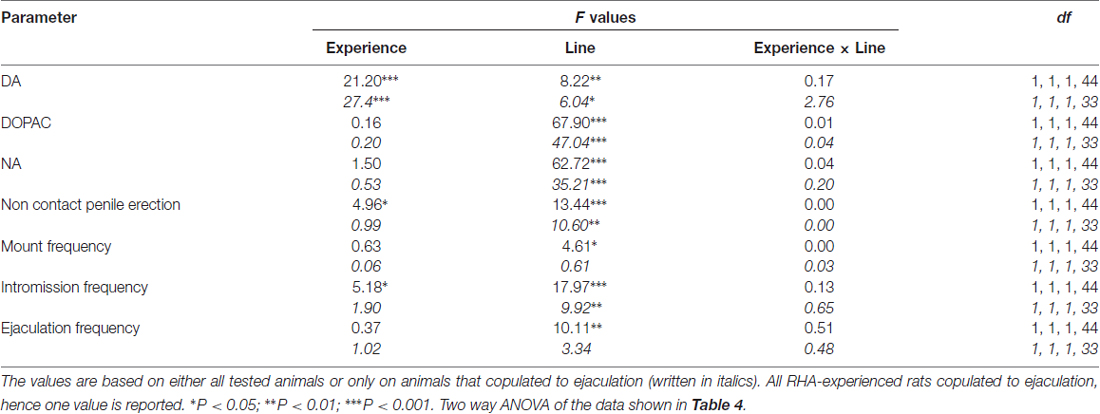 Tabulka 5. F hodnoty a úrovně významnosti dvoucestných ANOVA provedených na základě údajů uvedených v tabulce 4 (RHA vs. RLA) a úrovně sexuální zkušenosti (naivní vs. zkušené) mezi jednotlivými faktory.
Tabulka 5. F hodnoty a úrovně významnosti dvoucestných ANOVA provedených na základě údajů uvedených v tabulce 4 (RHA vs. RLA) a úrovně sexuální zkušenosti (naivní vs. zkušené) mezi jednotlivými faktory.
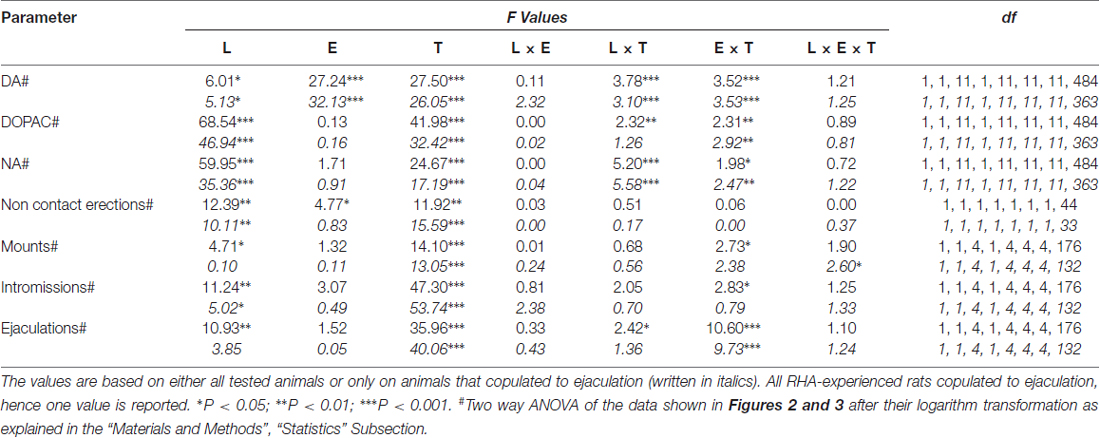 Tabulka 6. F hodnoty a úrovně významnosti obecných faktoriálních ANOVA pro opakovaná měření prováděná na výsledcích zobrazených na obrázcích 2 a 3 (L, RHA vs. RLA) a úrovně sexuální zkušenosti (E) (naivní vs. zkušené) mezi faktory subjektů a časem (T) (frakce dialyzátu) jako faktor v rámci předmětu.
Tabulka 6. F hodnoty a úrovně významnosti obecných faktoriálních ANOVA pro opakovaná měření prováděná na výsledcích zobrazených na obrázcích 2 a 3 (L, RHA vs. RLA) a úrovně sexuální zkušenosti (E) (naivní vs. zkušené) mezi faktory subjektů a časem (T) (frakce dialyzátu) jako faktor v rámci předmětu.
 Obrázek 2. Koncentrace dopaminu (DA), DOPAC a noradrenalinu (NA) v dialýzách mPFC ze sexuálně naivně (ESO) a zkušených potkanů RHA a RLA (B, D, F) během sexuální aktivity s receptivní ženou. Sexuálně naivní (tj. Nikdy předtím vystavena sexuálně vnímavé ženě) a sexuálně zkušení (kteří podstoupili pět kopulačních testů) obou linek s mikrodialyzovanou sondou stereotaxicky implantovanou do mPFC byly umístěny jednotlivě do páry klece. Mikrodialyzační sonda byla perfundována dialyzačním pufrem, jak je popsáno v části "Materiály a metody". Po odebrání čtyř alikvotů dialyzátu pro stanovení bazálních hodnot byla v malé kleci sdruženého přístroje (čas = 0, inaccess.fe) umístěna nepřístupná receptivní žena. Po 30 min byla odstraněna malá klec a kopinace byla povolena pro 75 min (kopulace), po které byla samice odstraněna ze slepé klece (fe.remov). Během experimentu byly měřeny nekontaktní erekce a parametry kopulace a alikvoty byly odebírány každý 15 min a analyzovány na dopamin, DOPAC a NA, jak je popsáno v části "Materiály a metody". Hodnoty jsou prostředky ± SEM hodnot získaných všemi krysami 12 na skupinu (RHA_all = pevné zelené čáry, RLA_all = tuhé hnědé linie) nebo ty, které se kopulují pouze na ejakulaci (RHA_cop = přerušované červené čáry; RLA_cop = přerušované modré čáry). U zkušených potkanů RHA se všechna zvířata dostala do kopulace a hodnoty jsou stejné. * *P <0.05 s ohledem na bazální hodnoty (bez samice) skupiny (zelená pro RHA_all; červená pro RHA_cop, hnědá pro RLA_all, modrá pro RLA_cop); #P <0.05 vzhledem k odpovídajícím hodnotám skupiny RLA (červená, RHA_all vs. RLA_all; zelená, RHA_cop vs. RLA_cop); §P <0.05 s ohledem na časově shodné hodnoty sexuálně naivních krys (zelená, zkušená RHA_all vs. naivní RHA_all; červená, zkušená RHA_cop vs. naivní RHA_cop; hnědá, zkušená RLA_all vs. naivní RLA_all; modrá, zkušená RLA_cop vs. naivní RLA_cop) (obousměrné ANOVA provedené na zobrazených datech po jejich logaritmické transformaci, jak je vysvětleno v pododdílech „Materiály a metody“, „Statistika“, následované testy HSD společnosti Tukey).
Obrázek 2. Koncentrace dopaminu (DA), DOPAC a noradrenalinu (NA) v dialýzách mPFC ze sexuálně naivně (ESO) a zkušených potkanů RHA a RLA (B, D, F) během sexuální aktivity s receptivní ženou. Sexuálně naivní (tj. Nikdy předtím vystavena sexuálně vnímavé ženě) a sexuálně zkušení (kteří podstoupili pět kopulačních testů) obou linek s mikrodialyzovanou sondou stereotaxicky implantovanou do mPFC byly umístěny jednotlivě do páry klece. Mikrodialyzační sonda byla perfundována dialyzačním pufrem, jak je popsáno v části "Materiály a metody". Po odebrání čtyř alikvotů dialyzátu pro stanovení bazálních hodnot byla v malé kleci sdruženého přístroje (čas = 0, inaccess.fe) umístěna nepřístupná receptivní žena. Po 30 min byla odstraněna malá klec a kopinace byla povolena pro 75 min (kopulace), po které byla samice odstraněna ze slepé klece (fe.remov). Během experimentu byly měřeny nekontaktní erekce a parametry kopulace a alikvoty byly odebírány každý 15 min a analyzovány na dopamin, DOPAC a NA, jak je popsáno v části "Materiály a metody". Hodnoty jsou prostředky ± SEM hodnot získaných všemi krysami 12 na skupinu (RHA_all = pevné zelené čáry, RLA_all = tuhé hnědé linie) nebo ty, které se kopulují pouze na ejakulaci (RHA_cop = přerušované červené čáry; RLA_cop = přerušované modré čáry). U zkušených potkanů RHA se všechna zvířata dostala do kopulace a hodnoty jsou stejné. * *P <0.05 s ohledem na bazální hodnoty (bez samice) skupiny (zelená pro RHA_all; červená pro RHA_cop, hnědá pro RLA_all, modrá pro RLA_cop); #P <0.05 vzhledem k odpovídajícím hodnotám skupiny RLA (červená, RHA_all vs. RLA_all; zelená, RHA_cop vs. RLA_cop); §P <0.05 s ohledem na časově shodné hodnoty sexuálně naivních krys (zelená, zkušená RHA_all vs. naivní RHA_all; červená, zkušená RHA_cop vs. naivní RHA_cop; hnědá, zkušená RLA_all vs. naivní RLA_all; modrá, zkušená RLA_cop vs. naivní RLA_cop) (obousměrné ANOVA provedené na zobrazených datech po jejich logaritmické transformaci, jak je vysvětleno v pododdílech „Materiály a metody“, „Statistika“, následované testy HSD společnosti Tukey).
výsledky
Krysy RHA a RLA vykazují rozdílný počet nekontaktních erekcí a různé vzory kopulačního chování
V souladu s dřívějšími studiemi (Sanna a kol., 2014a,b, 2015) se jiný počet pohlavně naivních samců potkanů RHA a RLA začal zabývat sexuální aktivitou, když byli spolu s receptivní ženou během sběru alikvotů dialyzátu z mPFC pomocí intracerebrální mikrodialýzy. Stručně, v této studii 10 z 12 z pohlavně naivní samčích RHA potkanů (83%) kopulovali do ejakulace v prvním kopulačním testu proti 6 pouze u krys RLA na počátku 12 (50%) během experimentu s mikrodialyzací. Tento rozdíl byl také nalezen u sexuálně zkušených samců potkanů RHA a RLA (po pěti kopulačních testech), i když byl oslaben, přičemž všechny krysy 12 RHA dosáhly ejakulace proti 9 z krys 12 RLA během mikrodialýzy. Vždy v souladu s předchozími studiemi i v této studii dvě linie římských potkanů vykazovaly různé latence a frekvence erekcí penisu bez kontaktů, když byly vystaveny nepřístupné receptivní ženě, stejně jako různé vzorce kopulačního chování během sexuální interakce. Tyto rozdíly byly zjištěny buď při uvážení údajů všech experimentálních zvířat dané skupiny, bez ohledu na to, zda se kopulovaly na ejakulaci nebo ne (např. Přiřazením plného skóre těm zvířatům, které neukázaly chování, tj. Potkanům, ejakulace) nebo při uvážení údajů o zvířatech, která ukázala pouze chování dané skupiny (např. zvířata, která se kopulovala do ejakulace; 1). Statistická analýza hodnot nekontaktní erekce (NCPE) a kopulačních parametrů naměřených v první sérii kopulační aktivity dvoucestnými ANOVA odhalila významné rozdíly mezi dvěma krysami, a to buď při zvažování všech experimentálních potkanů nebo kopulačních potkanů ve čtyřech experimentálních skupinách (viz tabulka 2). Ve skutečnosti, když byly vzaty v úvahu údaje získané od všech experimentálních zvířat, počet nekontaktních erekcí byl vyšší a NCPEL, ML, IL a PEI byly signifikantně kratší u potkanů RHA ve srovnání s krysami RLA. Dále EF a CE byly vyšší, zatímco III byl významně kratší u potkanů RHA ve srovnání s potkanmi RLA. Některé z výše uvedených rozdílů mezi dvěma římskými krysími liniemi se při opakovaných kopulačních testech zmenšovaly nebo zmizely. Některé z těchto rozdílů však přetrvávají i po stabilizaci sexuálního chování opakovaným sexuálním zážitkem. Podobné výsledky byly získány, když se zvažovaly pouze hodnoty zvířat ze čtyř experimentálních skupin, které se kopulovaly do ejakulace, s výjimkou EF (viz tabulky 1, 2).
Bazální koncentrace extracelulárního dopaminu, DOPAC a noradrenalinu v dialyzátech mPFC od sexuálně nespěšných a zkušených RHA a RLA krys
Při současných experimentálních podmínkách byly dávky dopaminu, DOPAC a NA v dialyzátech získaných z mPFC všech (bez ohledu na to, zda se kopulovaly na ejakulaci nebo ne), sexuálně nevhodnými potkany RHA a RLA, 2.32 pg a 2.11 pg pro dopamin, 322.56 pg a 124.42 pg pro DOPAC a 6.05 pg a 2.24 pg pro NA v 20 μl dialyzátu. Podobné hodnoty byly měřeny v dialyzátech získaných z mPFC u potkanů s RHA a RLA, kteří byli sexuálně testovaní (dopamin: 3.73 pg a 3.33 pg, DOPAC: 353.62 pg a 133.32 pg, NA: 7.17 pg a 2.78 pg u potkanů RHA a RLA) . Tyto hodnoty naznačují koncentraci ≅0.8-1.2 nM a ≅95-100 nM pro extracelulární dopamin a DOPAC a ≅1.8-2.2 nM pro extracelulární NA, v mPFC potkanů RHA a koncentraci ≅0.8-1.2 nM a ≅35-40 nM pro extracelulární dopamin a DOPAC a ≅0.6-0.9 nM pro extracelulární NA, v mPFC krys RLA (tabulka 3). Výše uvedené hodnoty byly získány po ekvilibrační době 2 h dialyzačního pufru s mimobuněčnou tekutinou mPFC. Vzhledem k tomu, že se odhaduje, že obnovení autentického dopaminu se DOPAC a NA dialyzačních sond blíží hodnotě 20, lze odhadnout, že koncentrace extracelulárního dopaminu, DOPAC a NA jsou blízké hodnotám xNUMX-4 nM v obou liniích pro dopamin, 5 a ≅ 500 nM pro DOPAC a ≅200 a ≅10 nM pro NA v mPFC u potkanů RHA a RLA. Faktory ANOVA pro opakovaná měření provedená na výše uvedených hodnotách po transformaci logaritmu odhalily významné rozdíly v bazální hladině dopaminu mezi potkany, kteří dosud nebyli dosud nakaženi a měli sexuální zkušenost (F(1,44,132) = 26.05, P <0.001) a v bazálních hladinách DOPAC (F(1,44,132) = 63.36, P <0.001) a NA (F(1,44,132) = 42.14, P <0.001) mezi krysami RHA a RLA (poslední čtyři vzorky odebrané před zavedením samice do malé klece). Podobné hodnoty byly nalezeny při zohlednění pouze potkanů, které kopulovaly k ejakulaci během mikrodialyzačního experimentu (tabulka 3). Také v tomto případě faktoriální ANOVA pro opakovaná měření provedená na hodnotách po transformaci logaritmu odhalily významné rozdíly v bazálních hladinách dopaminu mezi potkany, kteří dosud nebyli dosud nakaženi a měli sexuální zkušenost (F(1,33,99) = 26.97, P <0.001) a v bazálních hladinách DOPAC (F(1,33,99) = 42.95, P <0.001) a NA (F(1,33,99) = 27.63, P <0.001) mezi krysami RHA a RLA.
Koncentrace extracelulárního dopaminu, DOPAC a noradrenalinu v dialyzátech mPFC od sexuálně nespavých a zkušených RHA a RLA potkanů se mění diferencovaně během sexuální aktivity
Přítomnost nepřístupné ženy v malé vnitřní kleci a následná přímá sexuální interakce zvýšila koncentrace extracelulárního dopaminu, DOPAC a NA v dialyzátorech mPFC získaným z obou pohlavně dosud neléčených a zkušených samců RHA a RLA potkanů, i když s významnými rozdíly mezi dvě linie potkanů a úroveň zkušeností, a to buď při zvažování všech experimentálních zvířat nebo pouze těch, které se kopulovaly na ejakulaci (obrázek 2). Ve skutečnosti obousměrné ANOVA hodnot AUC koncentrací dopaminu, DOPAC a NA ukázaly, že celkový obsah dopaminu, DOPAC a NA celého testu byl významně vyšší u potkanů RHA ve srovnání s krysami RLA a v sexuálně kteří byli ve srovnání s naivními potkany (viz tabulka 4 pro hodnoty AUC a tabulku 5 for F hodnoty a významová úroveň). Tyto rozdíly byly dále potvrzeny bodovými faktory ANOVA pro opakovaná měření dávky dopaminu, DOPAC a NA u obou sexuálně naivních a zkušených potkaních linií v průběhu experimentu, které odhalily významné hlavní účinky Line, Experience, Time a signifikantní Line × Time a Zkušenosti × Doba interakcí prvního řádu (viz tabulka 6 for F hodnoty a významová úroveň).
Koncentrace extracelulárního dopaminu, DOPAC a noradrenalinu v dialyzátech mPFC získaných z sexuálně nevhodných RHA a RLA potkanů se mění diferencovaně během sexuální aktivity
U sexeálně naivních potkanů RHA a RLA byl extracelulární dopamin, ale ne DOPAC, nejprve zvýšil, když byl podáván v přítomnosti nepřístupné samice, zatímco NA se zvýšila pouze u potkanů RHA. U naivních potkanů RHA se extracelulární dopamin zvyšuje v prvním 15 minu sexuální interakce, jak bylo zjištěno u DOPAC a NA. Přírůstky extracelulárního dopaminu, DOPAC a NA trvaly po celou dobu kopulace, přičemž dopamin, DOPAC a NA dosáhly nejvyšší hodnoty u 60 min, 75 min a 75 min, potom extracelulárního dopaminu a v menší míře DOPAC a NA , po odebrání samičky se snížily na hodnoty podobné základním.
Na druhé straně první významný nárůst a vrcholová hodnota v extracelulárním dopaminu, DOPAC a NA se vyskytla u naivních RLA krys během prvního 15 min kopulace. Avšak v rozporu s potkany RHA u krys RLA extracelulárního dopaminu se DOPAC a NA snažily vrátit k hodnotám podobným základním hodnotám v prvním 30 min přímou interakcí se samičí (viz obrázky 2A, C, E).
Konečně, během kopulace byly přírůstky extracelulárního dopaminu větší a perzistentnější u pohlavně naivních RHA než potkanů RLA (viz obr. 2A), zatímco v nepřítomnosti samice byla jejich koncentrace podobná, i když ne totožná, napříč dvěma liniemi krys (viz výše a tabulka 3). Je zajímavé, že ačkoli nebyl zjištěn žádný rozdíl mezi dvěma římskými liniemi v procentuálním nárůstu DOPAC a v menší míře v koncentracích NA během přítomnosti vnímavé ženy, je rozdíl více než dvojnásobný v absolutních hodnotách koncentrací DOPAC a NA byla pozorována mezi dvěma potkaními liniemi jak v bazálních podmínkách (viz tabulka 3) a během testu s receptivní ženou (viz obr 2C, E).
Koncentrace extracelulárního dopaminu, DOPAC a noradrenalinu v dialýzách mPFC získaných z pohlavně vyzkoušených RHA a RLA potkanů se mění diferencovaně během sexuální aktivity
Jak bylo zjištěno u potkanů RHA a RLA, kteří dosud nebyli sexuálně naivní, přítomnost nepřístupné ženy a následná přímá sexuální interakce vedla ke zvýšení extracelulárního dopaminu a DOPAC v dialyzačním mPFC získaném z pohlavně zkušených samců potkanů RHA a RLA. Avšak také v tomto případě byly zjištěny rozdíly související s potkaním linií buď při zvažování všech pokusných zvířat, nebo v menší míře pouze u těch, které se kopulovaly na ejakulaci (obrázek 2). U sexuálně zkušených potkanů RHA a RLA došlo k prvnímu zvýšení extracelulárního dopaminu u nepřístupné samice. Poté se koncentrace dopaminu během kopulace zvýšily v obou potkacích liniích. Koncentrace dopaminu dosáhly vrcholových hodnot po 45-60 min kopulace a pomalu klesaly na bazální hodnoty na konci kopulačního testu (viz obr. 2B). Podobně jako u pohlavně naivních potkanů byly koncentrace extracelulárního dopaminu vyšší u potkanů s RHA sexuálně testovanými krysami ve srovnání s krysami RLA, zejména v alikvotních podílech dialyzátu odebraných během centrální části kopulační fáze (po 30-45 min kopulace). Koncentrace DOPAC vzrostly během přítomnosti nepřístupné samice pouze u potkanů RHA, zatímco v kopulaci se zvýšily v obou případech u krysích linií (vrcholové hodnoty při 75-90 min) a měly tendenci vrátit se k bazálním hodnotám na konci kopulační test (viz obr 2D). Koncentrace NA také vzrostly u obou sexuálně zkušených potkaních linií po zavedení žen dosahujících vrcholových hodnot při 90 min u potkanů RHA a při 45 min u potkanů RLA (viz obr. 2F). V rozporu s dopaminem (viz výše) byl podobný rozdíl zjištěn u vzrůstu koncentrací DOPAC a NA u pohlavně zkušených krys z obou linií, když se uvážily všechna zvířata vs. zvířata, která dosáhli pouze ejakulace. Nicméně, jak bylo zjištěno u pohlavně naivních potkanů, ačkoli procentní zvýšení koncentrací DOPAC a NA bylo podobné, významné rozdíly v absolutních hodnotách DOPAC a NA byly zjištěny mezi pohlavně testovanými potkany RHA a RLA během testu (tj. Před, během a po přítomnost receptivní samice), přičemž potkani RHA vykazují vyšší hodnoty než krysy RLA (viz tabulka 3 pro základní hodnoty a obrázky 2D, F pro hodnoty podél celého testu).
Sexuální zkušenost ovlivňuje změny v koncentracích dopaminu a noradrenalinu v dialýzách mPFC získaných z potkanů RHA a RLA během sexuální aktivity
Srovnání koncentrace extracelulárního dopaminu a NA u pohlavně nakažených a zkušených potkanů RHA a RLA ukázalo, že sexuální zkušenost změnila koncentraci extracelulárního dopaminu a v menší míře NA u obou krysích linií buď při zvažování všech pokusných zvířat nebo ty, které se kopulovaly pouze na ejakulaci (obr 2A, B, E, F). Proto došlo k významným rozdílům v bazálních hodnotách dopaminu mezi pohlavně naivně a zkušenými potkany obou linií (viz tabulka 3) a hodnoty dopaminu během kopulační aktivity byly obecně vyšší u pohlavně testovaných potkanů RHA a RLA ve srovnání s jejich naivními protějšky (viz obrázek 2B). Navíc u sexuálně zkušených potkanů RHA, ačkoliv zvýšení dopaminu zjištěné během přítomnosti nepřístupné samice sledovalo stejný časový vzorec pozorovaný u pohlavně naivních potkanů, byly během kopulace zjištěny rozdíly ve časovém vzoru. Ve skutečnosti se v tomto případě v rozporu s tím, co bylo pozorováno u pohlavně naivních potkanů RHA (jeden hlavní pík u 60 min), byly zjištěny dvě hlavní hodnoty píku, první po 15 min a druhé po 45 min kopulace. Po tomto posledním zvýšení hodnoty dopaminu měly tendenci se vrátit k bazálním hodnotám (viz obr 2B).
Podobný obrázek byl nalezen u potkanů RLA, kteří byli naivní a předtím experimentovaní. Ve skutečnosti, podobně jako u potkanů RHA, také u zkušených krys RLA byl nalezen časový vzorec charakterizovaný třemi hlavními nárůstem extracelulárního dopaminu: první s nepřístupnou samičí a další dvě během kopulace, tj. Po 15 min a 60 min kopulace (viz obr 2A, B). Obecně se zdá, že sexuální zkušenost způsobuje více dlouhotrvajících změn v časovém schématu uvolňování dopaminu v obou potkaních liniích při přechodu od sexuálně naivního do zkušeného stavu.
Jak je znázorněno na obrázcích 2C-F, byly pozorovány pouze malé rozdíly v koncentraci DOPAC a NA mezi naivními a zkušenými krysy obou linií. Nicméně významný trend extracelulárních NA k vyšším hodnotám u zkušených ve srovnání s naivními potkany byl pozorován u bazálních (před zavedením receptivní samice viz Tabulka 3) a v celkových množstvích NK, jak jsou zjištěny hodnotami AUC (viz tabulka 4) počítáno na extracelulární koncentrace získané z celého testu (před a během přítomnosti receptivní samice). Nakonec časový vzorec koncentrací NA byl velmi podobný, když porovnávaly zkušené potkany obou krysích linií s jejich naivními protějšky, což naznačovalo, že sexuální zkušenost může vést k obecnému zvýšení extracelulárního NA, a nikoliv ke změnám vzoru jeho nárůstu (viz obr. 2E, F).
Změny koncentrací extracelulárního dopaminu a noradrenalinu v dialýzách mPFC získaných z sexuálně naivních a zkušených RHA a RLA potkanů se vyskytují souběžně s změnami v sexuálním chování
Rozdíly v koncentracích extracelulárního dopaminu, DOPAC a NA v dialyzátorech získaných z mPFC nalezených během testu u obou sexuálně naivních a zkušených krys RHA a RLA (obrázek 2) se objevily společně s modifikacemi v různých sexuálních parametrech měřených během dvou hlavních fází pokusů (tj. ženy nepřístupné a dostupné pro muže), mezi něž patří mimo jiné nekontaktní erekce penisu (obr. 3A, B), úchyty (obr 3C, D), intromise (obr 3E, F) a ejakulací (obr 3G, H). Tyto rozdíly byly zjištěny buď při zvažování všech pokusných zvířat nebo těch, které se kopulovaly pouze na ejakulaci. Ve skutečnosti je předběžná analýza hodnot AUC těchto parametrů obousměrnými hodnotami ANOVA (viz tabulka 4 pro hodnoty AUC a tabulku 5 for F hodnoty a významová hladina), následovaná hodnotovou analýzou hodnot těchto parametrů v experimentu pomocí faktoriální analýzy ANOVA pro opakovaná měření odhalila významné hlavní účinky line, zkušeností, času, významných interakcí prvního řádu × čas a zkušenosti × čas, a významná interakce druhého řádu pro montáž (viz tabulka 6 for F hodnoty a významová úroveň). V souladu s předchozími studiemi (Sanna a kol., 2015): (i) sexuálně naivní RHA potkany vykazovaly více erekcí bez kontaktu penisu, mountů, intromise a ejakulací než jejich RLA protějšky; (ii) sexuálně zkušené krysy RHA vykazovaly vyšší počet erekcí, intromise a ejakulací bez kontaktu penisu než jejich RLA protějšky; a (iii) sexuálně zkušení potkani obou linií a zvláště potkaní RLA vykazovali více erekcí, intromise a ejakulací bez kontaktu penisu než jejich sexuálně naivní protějšky. Ve skutečnosti, zatímco u potkanů RHA se většina změn způsobených sexuálními zkušenostmi vyskytla spíše v časovém průběhu než ve frekvenci parametrů chování (např. Zkušení krysy kopulované hlavně v první polovině testu, zatímco naivní potkani ukázali obdobný kopulační trend v průběhu celého testu), u potkanů RLA bylo dosaženo výraznějšího obecného zlepšení sexuálního chování, zejména v sexuální výkonnosti, sexuální zkušeností, což bylo zjištěno změnami hodnot parametrů kopulace u zkušených potkanů RLA ve srovnání s jejich naivní protějšky.
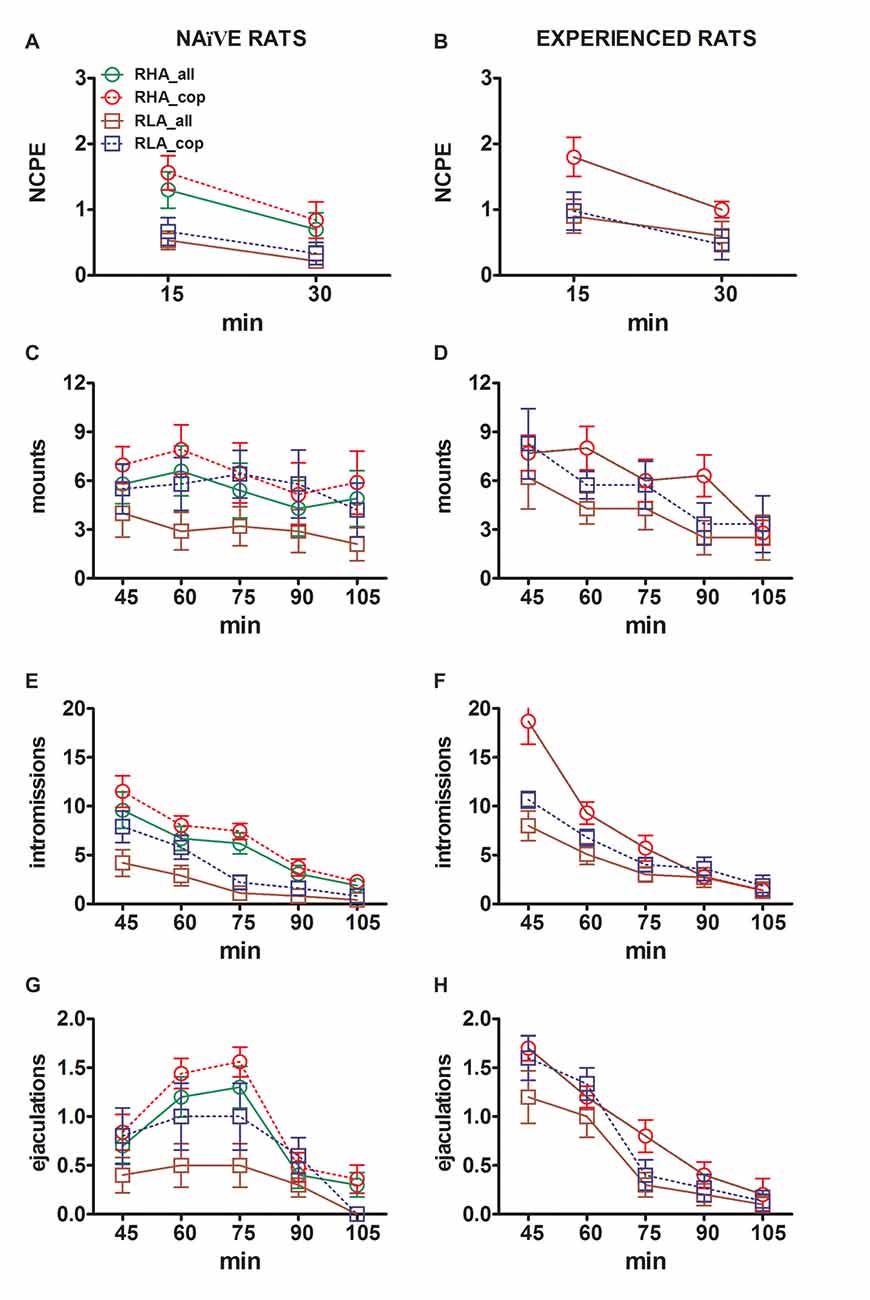 Obrázek 3. Změny v počtu nekontaktních erekcí penisu (NCPE), mountů, intromise a ejakulací zaznamenaných z sexuálně naivní (A, C, E, G) a zkušených potkanů RHA a RLA (B, D, F, H) během experimentů s mikrodialyzací uvedenými na obrázku 2. Všechny experimentální podmínky byly totožné s podmínkami popsanými v legendě na obr 2. Hodnoty jsou prostředky ± SEM hodnot získaných u všech potkanů 12 v každé skupině (včetně těch, které byly přiřazeny skóre na plný úvazek, RHA = tuhé zelené linie, RLA = tuhé hnědé linie) nebo ty, které se kopulovaly pouze na ejakulaci (RHA = ; RLA = přerušované modré čáry). U zkušených potkanů RHA se všechna zvířata dostala do kopulace a hodnoty jsou stejné. Vzhledem k neexistenci významných interakcí mezi faktorem potkaní linky (RHA vs. RLA) byl zjištěn faktor úrovně sexuálního zážitku (naivní vs. zkušený) a čas analýzou dat s dvoucestnými ANOVA, jak je uvedeno v tabulce 6, post hoc srovnání nebyly hlášeny, jak je vysvětleno v podkapitole "Materiály a metody", "Statistika".
Obrázek 3. Změny v počtu nekontaktních erekcí penisu (NCPE), mountů, intromise a ejakulací zaznamenaných z sexuálně naivní (A, C, E, G) a zkušených potkanů RHA a RLA (B, D, F, H) během experimentů s mikrodialyzací uvedenými na obrázku 2. Všechny experimentální podmínky byly totožné s podmínkami popsanými v legendě na obr 2. Hodnoty jsou prostředky ± SEM hodnot získaných u všech potkanů 12 v každé skupině (včetně těch, které byly přiřazeny skóre na plný úvazek, RHA = tuhé zelené linie, RLA = tuhé hnědé linie) nebo ty, které se kopulovaly pouze na ejakulaci (RHA = ; RLA = přerušované modré čáry). U zkušených potkanů RHA se všechna zvířata dostala do kopulace a hodnoty jsou stejné. Vzhledem k neexistenci významných interakcí mezi faktorem potkaní linky (RHA vs. RLA) byl zjištěn faktor úrovně sexuálního zážitku (naivní vs. zkušený) a čas analýzou dat s dvoucestnými ANOVA, jak je uvedeno v tabulce 6, post hoc srovnání nebyly hlášeny, jak je vysvětleno v podkapitole "Materiály a metody", "Statistika".
Diskuse
Tato studie potvrzuje a rozšiřuje poznatky z předchozích studií, které ukazují, že krysy RHA a RLA, které vykazují zřetelně odlišné styly v reakci na averzivní stavy (Driscoll a Bättig, 1982; Giorgi a kol., 2003a; Steimer a Driscoll, 2003) vykazují odlišné vzorce chování v přítomnosti nepřístupné ženy a během klasických kopulačních testů (Sanna a kol., 2014a,b, 2015). V souladu s tím také tato studie ukazuje, že sexuálně dosud neléčené krysy RHA (které nikdy nebyly vystaveny receptivní ženě dříve) vykazovaly vyšší sexuální motivaci a lepší sexuální výkonnost ve srovnání s sexuálně naivními potkany RLA, např. Potkani RHA vykazují vyšší počet erekcí penisu bez kontaktu (feromon- vyvolané erekce penisu považovány za index sexuálního vzrušení; Sachs a kol., 1994; Sachs, 2000) a různé pro-sexuální změny v několika kopulačních parametrech, které byly zjištěny v předchozích studiích (Sanna a kol., 2014a,b, 2015). Tyto rozdíly přetrvávaly v obou římských liniích potkanů po získání sexuálních zkušeností, tedy po stabilizaci sexuálního chování s pěti expozicemi a kopulačními testy s receptivní ženou. Tato studie také poprvé ukazuje, že u obou potkaních linek RHA a RLA, buď v naivním nebo zkušeném stavu, nedošlo k erekci penisu a sexuální interakci souběžně s nárůstem koncentrace extracelulárního dopaminu (a jeho hlavního metabolitu DOPAC ) a NA v dialyzátech získaných intracerebrální mikrodialýzou z mPFC obou římských potkaních linií. Je zajímavé, že toto zvýšení extracelulárního dopaminu a NA v mPFC se zdá být spojeno s rozdíly buď v potkaní fenotypu nebo v úrovni sexuální zkušenosti. V souladu s tím, jak se ukázalo v sekci Výsledky, oba sexuálně naivní a zkušení RHA potkani vykazovali vyšší a dlouhodobé zvýšení extracelulárního dopaminu a NA v porovnání s jejich RLA protějšky jak v apetitivní, tak konzumační fázi sexuálního chování. Navíc sexuálně zkušení krysy obou římských linií vykazovaly vyšší a dlouhodobé zvýšení extracelulárního dopaminu a NA v mPFC v obou fázích sexuálního chování v porovnání s jejich naivními protějšky. Konečně všechny výše uvedené rozdíly v extracelulárním dopaminu a NA v mPFC byly zřejmě souvisely s rozdíly v behaviorálních parametrech sexuálního chování, např. Sexuálně zkušení krysy obou římských linií vykazovaly vyšší sexuální motivaci a lepší kopulační výkony ve srovnání s jejich naivními protějšky a sexuálně naivní a zkušení potkani RHA vykazovali vyšší sexuální motivaci a lepší kopulační výkony ve srovnání s jejich sexuálně naivní a zkušenými RLA protějšky.
Pokud jde o nárůst extracelulárního dopaminu, který byl nalezen v dialyzačním zařízení mPFC během sexuální aktivity, došlo k prvnímu zvýšení v prvním 15 min po zavedení sexuálně vnímavé ženy do páření, kdy byla samice pro muže nedostupná, což obvykle ukazuje v této podmínce nekontaktní erekce penisu. Druhé a větší zvýšení extracelulárního dopaminu se vyskytlo během prvního 15 min poté, co byla samice zpřístupněna samci pro kopulaci. Je zajímavé, že zatímco v naivním stavu vzrostl extracelulární dopamin, hlavně v první části testu s dostupnou samičkou s tendencí vrátit se k bazálním hodnotám v druhé části, v sexuálně zkušebném stavu byl dopamin vyšší v průběhu testu a dosáhl špičkových hodnot po 45-60 min kopulace v obou římských potkaních liniích.
Vedle výše uvedených rozdílů v extracelulárním dopaminu během různých fází sexuální aktivity mezi potkany RHA a RLA v sexuálně naivní a zkušené populaci tato studie rovněž ukazuje neočekávané důležité a velmi významné rozdíly v extracelulární koncentraci DOPAC mezi dvěma římskými potkanými liniemi dříve , bazální hodnoty) a po zavedení vnímavé samičky do spojovacího zařízení. V souladu s tím byly bazální hodnoty extracelulárního DOPAC v mPFC dialyzátu nalezeny u potkanů RHA, které mají přibližně 2.6-krát vyšší než hodnoty zjištěné u potkanů RLA. Tento rozdíl mezi krysami RHA a RLA se objevil a přetrvával v sexuálně naivních a zkušených stavech navzdory velmi skromným rozdílům v bazálních hodnotách extracelulárního dopaminu v dialyzátorech mPFC získaným z těchto dvou římských potkaních linií v těchto experimentálních podmínkách. Celkově mohou tyto rozdíly signalizovat odlišný obrat dopaminu na úrovni mPFC mezi dvěma římskými liniemi krys. V souladu s tím koncentrace DOPAC obvykle odrážejí množství uvolněného dopaminu a následně znovu zachycují dopaminergní nervové terminály a převedou na DOPAC monoaminooxydázou (MAO; Carlsson, 1975). Proto vyšší bazální koncentrace DOPAC zjištěné u potkanů RHA naznačují, že vyšší dávky dopaminu se uvolňují a opětovně zachycují dopaminergními nervovými zakončeními v mPFC potkanů RHA, např. Vyšší bazální aktivita mezokortikálního dopaminergního systému potkanů RHA ve srovnání s RLA potkanů. Vyšší mesokortický dopaminergní tón u potkanů RHA než u potkanů RLA je také podpořen vyššími koncentracemi extracelulárního dopaminu zjištěnými během sexuální aktivity v dialyzačním roztoku mPFC u obou sexuálně naivních a zkušených potkanů RHA s ohledem na jejich RLA protějšky. V tomto ohledu je třeba připomenout, že vyšší dopaminergní tón je považován za zodpovědný, přinejmenším částečně, za mnoho z různých a dokonce i opačných behaviorálních znaků přítomných u potkanů RHA a RLA. V souladu s tím jsou krysy RHA aktivní kopery, vysoce impulzivní, novinky a hledající senzace a jsou náchylné k přijímání a zneužívání několika tříd drog závislosti, zatímco RLA krysy jsou reaktivní kopery, hyperemocionální a jsou náchylné k rozvoji depresivních symptomůZeier a kol., 1978; Giorgi a kol., 1994, 2003b, 2007; Corda a kol., 1997, 2014; Escorihuela a kol., 1999; Steimer a Driscoll, 2003; Lecca a kol., 2004; Giménez-Llort a kol., 2005; Carrasco a kol., 2008; Fattore a kol., 2009; Moreno a kol., 2010; Coppens a kol., 2012; Díaz-Morán a kol., 2012; Sabariego a kol., 2013; Manzo a kol., 2014a,b; Oliveras a kol., 2015), včetně vyšší sexuální motivace a lepší kopulační výkon sexuálně naivních a zkušených potkanů RHA proti krysám RHA (Sanna a kol., 2015). Vyšší nárůst koncentrací extracelulárního dopaminu zjištěného v dialyzátu z mPFC u sexuálně naivních a zkušených krys RHA se podobá vyšším koncentracím extracelulárního dopaminu zjištěnému v dialyzátu z nucleus accumbens u sexuálně naivních a zkušených potkanů RHA s ohledem na jejich RLA protějšky (Sanna a kol., 2015). Nicméně, v rozporu s mPFC, bazální hladiny extracelulárního dopaminu a DOPAC v dialyzátu z nucleus accumbens obou sexuálně naivních a zkušených krys RHA a RLA se významně nelišily (Sanna a kol., 2015). Proto vyšší mezolimbický dopaminergní tón v nucleus accumbens u sexuálně naivních a zkušených potkanů RHA může být odvozen pouze vyššími extracelulárními koncentracemi dopaminu zjištěnými v dialyzátech z nucleus accumbens potkanů RHA ve srovnání s protějšky RLA v přítomnosti vnímavých samice a nikoliv jinou bazální dopaminergní aktivitou odhalenou vyššími a nižšími bazálními hladinami koncentrace DOPAC u potkanů RHA a krys RLA, jak bylo zjištěno v mPFC v této studii. To je také v souladu s výsledky předchozích zpráv, které ukazují, že extracelulární hladiny dopaminu v dialyzátu z mPFC potkanů RHA byly vyšší než u potkanů RLA, zatímco podobné hladiny dopaminu byly zjištěny v dialyzátu ze skořápky nucleus accumbens obou RHA a RLA potkanů za různých experimentálních podmínek (D'Angio a kol., 1988; Scatton a kol., 1988; Willig a kol., 1991; Giorgi a kol., 2003a, 2007). Přes výše uvedené rozdíly mezi mPFC a jádrem accumbens se zdá, že sexuální zkušenost produkuje v mPFC dlouhodobější vzestup časového schématu uvolňování dopaminu v obou římských liniích potkanů při přechodu od sexuálně naivného do zkušeného stavu, i když takové zvýšení byly u potkanů RHA obvykle vyšší než krysy RLA. To se poněkud liší od toho, co se vyskytuje v nucleus accumbens, kde sexuální zážitky vedly k posunu extracelulárního dopaminu ke první části kopulačního testu u potkanů RHA, zatímco se zvýšily v celém kopulačním testu u potkanů RLASanna a kol., 2015).
Podle našeho názoru tato studie také poprvé ukazuje, že nejen extracelulární dopamin, ale také extracelulární NA se během sexuální aktivity dialyzátu zvyšuje z mPFC u potkanů RHA i RLA, jak se vyskytuje u dopaminu, ale také v tomto případě rozdíly související s římskou linií potkanů a se sexuální zkušeností. Konkrétně, v rozporu s bazálními hladinami dopaminu, které byly velmi podobné u dialyzátu z mPFC obou krys RHA a RLA, byly bazální hladiny extracelulární NA v dialyzátu z mPFC krys RHA významně vyšší (asi 2,5-fold ) než u potkanů RLA. Důvody pro tyto rozdíly jsou neznámé, ale jak bylo diskutováno výše pro dopamin a DOPAC, tyto výsledky mohou naznačovat, že v mozkové kůře potkanů RHA existuje nejvyšší bazální noradrenergní tón v porovnání s krysami RLA. Takový vyšší noradrenergní tón se může také podílet na různých a často opačných behaviorálních vlastnostech přítomných u potkanů RHA a RLA, jak bylo popsáno výše pro dopamin. Další podpora pro vyšší noradrenergní tón u potkanů RHA než u potkanů RLA v mPFC pochází z výsledků, které ukazují, že rozdíly v bazálních extracelulárních hladinách NA přetrvávaly také během sexuální aktivity u obou sexuálně naivní a zkušených podmínek, např. dialyzát mPFC u potkanů RHA při expozici nepřístupné samičce a během všech období kopulace mnohem víc než u potkanů RLA, u kterých se hladiny NA zvýšily pouze v prvních 15 min období kopulace, po které se hodnoty NA vrátily zpět k bazálním hodnotám . Tyto změny v extracelulárních hladinách NA byly pozorovány jak v pohlavně naivním, tak v zkušebním stavu, avšak zvýšení hladin NA bylo vyšší u pohlavně postižených potkanů RHA a RLA ve srovnání s jejich sexuálně prospěšnými protějšky. Takže sexuální zkušenost zřejmě vyvolává další potenciaci noradrenergní aktivity u mRFC potkanů RHA a v menší míře u potkanů RLA, jak již bylo diskutováno u dopaminu. Souběžně vyšší noradrenergní a dopaminergní tóny u mRFC potkanů RHA ve srovnání s krysami RLA si zaslouží nějaký komentář. Ve skutečnosti bylo hlášeno, že noradrenergní aktivita v mPFC může ovlivňovat uvolňování dopaminu různými mechanismy (viz např. Carboni a kol., 1990, 2006; Gresch a kol., 1995; Westernik a kol., 1998 a odkazy v ní). Jeden z nich souvisí s aktivitou NET, u které se prokázalo, že váže dopamin s afinitou ještě vyšší než afinita k NA v synaptozomech získaných z homogenátů PFC (Horn, 1973). Pokud k tomu dojde v mPFC in vivo, dopamin uvolněný v mPFC by mohl být znovu zachycen nejen dopaminergními, ale i noradrenergními nervovými terminály při rychlosti dokonce vyšší než rychlost NA. To by mohlo způsobit zvýšení uvolňování dopaminu z dopaminergních terminálů (např. Zvýšený dopaminergní tón), aby se kompenzovalo množství dopaminu odstraněného z synaptické štěrbiny; takže vyšší noradrenergní tón nalezený u potkanů RHA by mohl alespoň částečně přispět k vyššímu dopaminergnímu tónu zjištěnému u mRFC potkanů RHA ve srovnání s krysami RLA. Další studie jsou nutné k ověření této možnosti. Pozoruhodně rozdíl v noradrenergní aktivitě zjištěný v této studii v mPFC mezi potkany RHA a RLA může také hrát roli v jejich různé odpovědi na antidepresivní léčbu. Ve skutečnosti jsou krysy RHA považovány za model odolnosti a RLA potkanů modelem zranitelnosti vůči depresi. V souladu s tím akutní nebo chronická léčba antidepresivy, jako je desipramin, fluoxetin a chlorimipramin, významně zlepšily reakce na chování v testu nuceného plavání u potkanů RLA bez ovlivnění odpovědí potkanů RHA (Piras a kol., 2010, 2014).
Tato studie konečně potvrzuje a rozšiřuje předchozí poznatky, které ukazují, že mPFC se podílí na kontrole sexuálního chování, ačkoli přesná role této oblasti mozku v sexuálním chování není zdaleka jasná (Fernández-Guasti a kol., 1994; Agmo a Villalpando, 1995; Agmo et al., 1995; Hernández-Gonzáles a kol., 1998, 2007; Kakeyama a kol., 2003; Balfour a spol., 2006; Afonso a kol., 2007; Davis a kol., 2010; Febo, 2011). Nedávno se ukázalo, že selektivní spalování buněk v mPFC při přiblížení se chování samčích krys směrem k nepřístupné sexuálně vnímatelné ženě bylo provedeno pomocí paradigmatu záznamu jedné buňky (Febo, 2011). Je zajímavé, že neurony, které nereagovaly během první expozice muže na nepřístupnou receptivní ženu, se aktivovaly během druhé expozice, jako kdyby předchozí zkušenost byla schopna indukovat plastické změny, které vedly ke změnám reakce na chování. Nicméně, léze s kyselinou ibotenovou z mPFC nebyly schopné změnit sexuální chování samců potkanů s sexuálně vnímavou ženou, ani vyjádření podmíněných místních preferencí pro sexuální odměnu. Nicméně schopnost vytvářet podmíněnou averzi vůči sexuální aktivitě, když byla spárována s averzními podněty, byla v těchto zraněných zvířatech zcela odstraněna (Davis a kol., 2010). To naznačuje, že aktivace mPFC během sexuálního chování hraje roli v integraci externích a interních informací pro výkon a kontrolu cílového chování spíše než ve vyjádření vrozených reakcí na přirozené posilovače (viz Goto a Grace, 2005). Důležitější pro tuto práci bylo několik studií, které ukázaly, že oblasti mozku, které se podílejí na sexuální motivaci a sexuálním chování, jako jsou nucleus accumbens, ventrální tegmentální oblast, mediální preoptická oblast, jádro lůžka stria terminalis, bazolaterální amygdala a parvocelulární subfraascicular thalamic nucleus z mPFC během sexuální aktivity (viz Balfour a spol., 2006). Mnohé z těchto oblastí mozku se účastní komplexního neurálního obvodu zapojeného do kontroly sexuálního chování, od sexuální motivace a odměny k sexuálnímu výkonu. Tento obvod zahrnuje oxytocinergní neurony, které pocházejí z PVN hypotalamu (PVN) a projevují se na ventrální tegmentální oblast, nucleus accumbens, hippocampus, amygdala, PFC, jádro lůžka stria terminalis, medulla oblongata a mícha, mezolimbic / mesocortical dopaminergic neurony vyčnívající z ventrální tegmentální oblasti do nucleus accumbens a PFC, incertohypothalamic dopaminergní neurony a glutamatergické neurony, které se účastní lokální a systémové úrovně v několika oblastech obvodu (viz Melis a Argiolas, 1995, 2011; Melis a kol., 2003, 2007, 2009, 2010; Succu a kol., 2007, 2008, 2011). Zvýšení extracelulárního dopaminu (ale také NA) u dialyzátů z mPFC pozorovaného během sexuální aktivity může naznačovat, že mPFC je aktivní částí tohoto obvodu, to znamená, že mezokortické dopaminergní neurony se podílejí na kontrole sexuální aktivity společně s noradrenergními, mezolimbickými dopaminergními, oxytocinergními a glutamatergickými neurony, ačkoli se zdá, že není nezbytně nutné pro vyjádření sexuálního chování (Davis a kol., 2010). V souladu s touto hypotézou tato studie ukazuje, že rozdíly v uvolňování dopaminu a NA v mPFC během sexuální aktivity se objevily souběžně s rozdíly v počtu erekcí bez kontaktu penisu a změnami v kopulačních parametrech s vyšším uvolňováním dopaminu a NA během apetitívní a konzumační fáze spojené s vyšší sexuální motivací a lepšími kopulačními výkony v římských liniích potkanů i v sexuálně naivních a zkušených podmínkách.
Diskuse uvedená výše je založena hlavně na známém zvýšení extracelulárního dopaminu, ke kterému dochází v nucleus accumbens samců potkanů během sexuální aktivity (Pfaus a kol., 1990; Pleim a kol., 1990; Pfaus a Phillips, 1991; Damsma a kol., 1992; Wenkstern a kol., 1993). Avšak dopamin a jiné neurotransmitery by mohly být uvolňovány v mPFC (a v nucleus accumbens) v jiných experimentálních podmínkách, jako je například během stresu nebo s ním (Thierry a kol., 1976; Fadda a kol., 1978), které nejsou přísně spjaty se sexuálním chováním. Tudíž lze tvrdit, že zvýšení dopaminu zjištěné v extracelulárním dopaminu v mPFC (tato studie) nebo v nucleus accumbens (Sanna a kol., 2015), je spíše spojena se stresem sekundárním nebo s vyrovnáním se s přítomností a / nebo interakcí s sexuálně vnímavou ženou spíše než se sexuální aktivitou. To může mít určitý význam u krys RHA a RLA, které vykazují zřetelně odlišné styly a různé aktivace osy hypothalamus-hypofýza-nadledvina v reakci na averzivní stavy (krysy RLA vykazují aktivaci této osy mnohem vyšší než u potkanů RHA; Carrasco a kol., 2008; Díaz-Morán a kol., 2012). To také zvyšuje možnost, že stresové hormony (tj. Kortikosteroidy) mohou ovlivňovat dopaminergní aktivitu v mPFC a dalších oblastech mozku. Krysy RLA jsou reaktivní kopery a vykazují hyperemocionální chování charakterizované hypomotilitou a mrazem, zatímco potkani RHA vykazují proaktivní chování při manipulaci se stresem (Driscoll a Bättig, 1982; Willig a kol., 1991; Escorihuela a kol., 1999; Steimer a Driscoll, 2003; Giorgi a kol., 2007). I když je nemožné zcela vyloučit, že k tomu může dojít u sexuálně naivních krys (které nikdy předtím neinteragovaly s vnímavou ženou) a zvláště u sexuálně naivních krys RLA (viz výše), je to nepravděpodobné. V našich experimentálních podmínkách se dělá vše, co je možné, aby se zabránilo všem druhům stresu (viz část „Materiály a metody“) a experimenty jsou organizovány tak, aby muži neinteragovali s ženou okamžitě, ale až po 30 minutách expozice jí , během kterého je naměřený nárůst dopaminu mnohem nižší než nárůst, ke kterému dochází během sexuální interakce. Pokud zvýšení dopaminu zjištěné v mPFC (a v nucleus accumbens) římských potkanů (sexuálně naivní RHA a RLA krysy mají bazální hladiny extracelulárního dopaminu v dialyzátu z mPFC a nucleus accumbens, velmi podobné) byly skutečně sekundární vůči stres způsobený nebo zvládáním přítomnosti / interakce se ženou: (i) zvýšení dopaminu by bylo zjištěno mnohem vyšší v prvním období, kdy jsou muži vystaveni přítomnosti ženy, a ne později, jak bylo zjištěno během páření; a ii) lze očekávat, že zvýšení dopaminu zmizí nebo se výrazně sníží po získání stabilní sexuální aktivity, tj. po dokončení procesů učení (včetně zvládání novosti), které budou hrát při provádění sexuálních aktivit jen malou roli na rozdíl od toho se i nadále opakuje, když se sexuální aktivita opakuje. Jinými slovy, extracelulární dopamin v mPFC (a v nucleus accumbens) se během kopulace vždy zvyšuje, a to i poté, co se naučíte sexuální chování. K objasnění funkční role tohoto zvýšení aktivity dopaminu v mPFC (a v nucleus accumbens; viz část „Úvod a odkazy“) jsou zapotřebí další studie.
Na závěr tato studie poprvé ukazuje, že přítomnost a ještě více interakce s sexuálně vnímavou ženou vede k nárůstu extracelulárního dopaminu a NA v dialyzátu z mPFC sexuálně naivního (nikdy vystaveného sexuálním podnětům ) a sexuálně zkušenými (kteří podstoupili pět předběžných kopulačních testů a ukázali stabilní sexuální výkony) krysy RHA a RLA, které vykazují zřetelně odlišné styly při odezvě na averzivní stavy a různé vzorce kopulačního chování u sexuálně vnímavé ženy. V souladu s tím byly koncentrace dopaminu a NA zjištěny v dialyzátu vyšší než u sexuálně naivních a zkušených potkanů RHA ve srovnání s jejich RLA protějšky během předběžné a konzumační fáze sexuální aktivity. Tyto výsledky mohou být částečně způsobeny dopaminergním a noradrenergním tónem u mRFC potkanů RHA vyšší, než u potkanů RLA, jak bylo uvedeno pro dopamin v nucleus accumbens (Sanna a kol., 2015). Kromě toho byl extracelulární mPFC dopamin a NA zjištěn vyšší u sexuálně zkušených RHA a v menší míře u potkanů RLA ve srovnání s jejich sexuálně prospěšnými protějšky v bazálních podmínkách, stejně jako během apetitivní a konzumační fáze sexuálního chování. To naznačuje, že sexuální zkušenost indukuje plastické procesy, které dále zvyšují dopamin a neurotransmisi NA v mPFC, jak bylo zjištěno pro dopamin v nucleus accumbens obou římských linií krys (Sanna a kol., 2015). Výše uvedené rozdíly v dopaminergní a noradrenergní neurotransmisi v mPFC dvou římských linií mohou hrát roli nejen v různých vzorcích sexuálního chování, ale také v různých a často opačných charakteristikách chování, které charakterizují tyto dvě linie potkanů.
Autorské příspěvky
FS, AA, MRM, OG a MGC navrhli projekt. FS a JB navrhly, provedly a analyzovaly údaje ze sexuálního chování a experimentů s mikrodialyzací. MAP, OG a MGC vybrané a chované říční krysy. FS, AA, MRM, OG a MGC sledovaly studii. FS, AA, MRM, OG a MGC napsaly rukopis. Všichni autoři diskutovali o výsledcích a komentovali rukopis.
Financování
Tato práce byla částečně podpořena granty italského ministerstva univerzity a výzkumu (MIUR) (Vědecká nezávislost mladých vědců, SIR 2014, kód č. RBSI14IUX7) do FS, z autonomní oblasti Sardinie (ARS) (LR 7 / 2007 , "Podpora vědeckého výzkumu a technologických inovací na Sardinii", Evropský sociální fond, 2007-2013, Projektový kód č. CRP-59842) na OG a od Univerzity v Cagliari po AA, MRM, FS, OG a MGC.
Prohlášení o konfliktu zájmů
Autoři prohlašují, že výzkum byl proveden bez obchodních či finančních vztahů, které by mohly být považovány za potenciální střet zájmů.
Poděkování
Dr. Barbara Tuveri, univerzita v Cagliari, pro bydlení a péči o zvířata.
Reference
Afonso, VM, Sison, M., Lovic, V. a Fleming, AS (2007). Mediální prefrontální kortikální léze u potkaních samic ovlivňují sexuální a mateřské chování a jejich následnou organizaci. Behav. Neurosci. 121, 515-526. dva: 10.1037 / 0735-7044.121.3.515
Agmo, A. a Villalpando, A. (1995). Centrální nervové stimulátory usnadňují sexuální chování u samců potkanů s mediálními prefrontalovými lézemi. Brain Res. 696, 187–193. doi: 10.1016/0006-8993(95)00853-i
Agmo, A., Villalpando, A., Picker, Z., a Fernández, H. (1995). Léze mediální prefrontální kůry a sexuální chování u samců potkanů. Brain Res. 696, 177–186. doi: 10.1016/0006-8993(95)00852-h
Argiolas, A. a Melis, MR (1995). Neuromodulace erekce penisu: přehled role neurotransmiterů a neuropeptidů. Prog. Neurobiol. 47, 235–255. doi: 10.1016/0301-0082(95)00020-8
Argiolas, A. a Melis, MR (2005). Centrální kontrola erekce penisu: úloha paraventrikulárního jádra hypotalamu. Prog. Neurobiol. 76, 1-21. dva: 10.1016 / j.pneurobio.2005.06.002
Argiolas, A. a Melis, MR (2013). Neuropeptidy a centrální kontrola sexuálního chování z minulosti do současnosti: přehled. Prog. Neurobiol. 108, 80-107. dva: 10.1016 / j.pneurobio.2013.06.006
Balfour, ME, Brown, JL, Yu, L. a Coolen, LM (2006). Potenciální příspěvky eferentů z mediální prefrontální kůry k nervové aktivaci po sexuálním chování u samců potkanů. Neurovědy 137, 1259-1276. dva: 10.1016 / j.neuroscience.2005.11.013
Balfour, ME, Yu, L. a Coolen, LM (2004). Sexuální chování a sexuální doprovodné příznaky aktivují mezolimbický systém u samců potkanů. Neuropsychopharmacology 29, 718-730. dva: 10.1038 / sj.npp.1300350
Beloate, LN, Omrani, A., Adan, RA, Webb, IC a Coolen, LM (2016). Aktivace dopamínových buněk v oblasti tepenné oblasti v průběhu sexuálního chování mužského pohlaví reguluje neuroplasticitu a křížovou senzibilizaci d-amfetaminu po abstinenci pohlaví. J. Neurosci. 36, 9949-9961. dva: 10.1523 / JNEUROSCI.0937-16.2016
Berridge, K. a Robinson, T. (1998). Jaká je role dopaminu v odměně: hedonický dopad, odměňování nebo pobídka? Brain Res. Rev. 28, 309–369. doi: 10.1016/s0165-0173(98)00019-8
Bignami, G. (1965). Výběr vysokých dávek a nízké míry kondicionování potíží u potkanů. Anim. Behav. 13, 221–227. doi: 10.1016/0003-3472(65)90038-2
Broadhurst, PL a Bignami, G. (1965). Korelační účinky psychogenetické selekce: studie romských vysokých a nízkých kmenů vyhýbání krys. Behav. Res. Ther. 3, 273–280. doi: 10.1016/0005-7967(64)90033-6
Carboni, E., Silvagni, A., Vacca, C. a Di Chiara, G. (2006). Kumulativní účinek blokády norepinefrinu a dopaminového nosiče na zvýšení extracelulárního dopaminu ve skořápce nucleus accumbens, ložiskovém jádru stria terminalis a prefrontální kůře. J. Neurochem. 96, 473-481. dva: 10.1111 / j.1471-4159.2005.03556.x
Carboni, E., Tanda, GL, Frau, R. a Di Chiara, G. (1990). Blokáda noradrenalinového nosiče zvyšuje koncentrace extracelulárního dopaminu v prefrontální kůře: důkaz, že dopamina je přijímána in vivo noradrenergními terminály. J. Neurochem. 55, 1067–1070. doi: 10.1111/j.1471-4159.1990.tb04599.x
Carlsson, A. (1975). "Receptorem zprostředkovaná kontrola metabolismu dopaminu", v Pre- a postsynaptické receptory, vyd. E. Usdin a WE Bunney (New York, NY: Marcel Deker), 49-65.
Carrasco, J., Márquez, C., Nadal, R., Tobeña, A., Fernández-Teruel, A. a Armario, A. (2008). Charakterizace centrálních a periferních složek osy hypothalamus-hypofýza-nadledviny u inbrisovaných kmenů římských krys. Psychoneuroendokrinologie 33, 437-445. dva: 10.1016 / j.psyneuen.2008.01.001
Coppens, CM, de Boer, SF, Steimer, T. a Koolhaas, JM (2012). Impulsivita a agresivní chování u potkanů s nízkým a nízkým rizikem v Římě: základní rozdíly a změny v sociálním stresu dospívající osoby. Physiol. Behav. 105, 1156-1160. dva: 10.1016 / j.physbeh.2011.12.013
Corda, MG, Lecca, D., Piras, G., Di Chiara, G., a Giorgi, O. (1997). Biochemické parametry dopaminergní a GABAergní neurotransmise u CNS římských potkanů s nízkým vyhýbáním a římského nízkého vyhýbání se. Behav. Genet. 27, 527-536. dva: 10.1023 / A: 1021452814574
Corda, MG, Piras, G., Piludu, MA, a Giorgi, O. (2014). Diferenciální účinky dobrovolné spotřeby etanolu na produkci dopaminu v skořápce nucleus accumbens u romských krys s vysokým a nízkým vyloučením: studie chování a mozkové mikrodialýzy. Svět J. Neurosci. 4, 279-292. dva: 10.4236 / wjns.2014.43031
Dahlstrom, A. a Fuxe, K. (1964). Důkazy o existenci neuronů obsahujících monoaminy v centrální nervové soustavě. I. Demonstrace monoaminů v tělech buněk neuronů mozku. Acta Physiol. Scand. Suppl. 62, 1-54.
Damsma, G., Pfaus, J., Wenkstern, D., Phillips, A., a Fibiger, H. (1992). Sexuální chování zvyšuje přenos dopaminu v nucleus accumbens a striatum samců potkanů: srovnání s novinkou a pohybem. Behav. Neurosci. 106, 181-191. dva: 10.1037 / 0735-7044.106.1.181
D'Angio, M., Serrano, A., Driscoll, P. a Scatton, B. (1988). Stresující enviromentální podněty zvyšují extracelulární hladiny DOPAC v prefrontálním kortexu hypoemocionálních (římské vysoké vyhýbání se), ale nikoli hyperemocionálních (římské nízké vyhýbání se) potkanům. an in vivo voltametrická studie. Brain Res. 451, 237–247. doi: 10.1016/0006-8993(88)90768-8
Davis, JF, Loos, M., Di Sebastiano, AR, Brown, JL, Lehman, MN a Coolen, LM (2010). Léze mediální prefrontální kůry způsobují maladaptivní sexuální chování u samců potkanů. Biol. Psychiatrie 67, 1199-1204. dva: 10.1016 / j.biopsych.2009.12.029
Díaz-Morán, S., Palència, M., Mont-Cardona, C., Cañete, T., Blázquez, G., Martínez-Membrives, E. a kol. (2012). Stylový styl a reakce na stresový hormon u geneticky heterogenních krys: srovnání s kmeny římských potkanů. Behav. Brain Res. 228, 203-210. dva: 10.1016 / j.bbr.2011.12.002
Driscoll, P. a Bättig, K. (1982). "Behaviorální, emoční a neurochemické profily potkanů vybraných pro extrémní rozdíly v aktivním, oboustranném vyhýbání se výkonu" Genetika mozku, ed. I. Lieblich (Amsterdam: Elsevier), 95-123.
Dunlop, BW a Nemeroff, CB (2007). Role dopaminu v patofyziologii deprese. Oblouk. Gen. Psychiatrie 64, 327-337. dva: 10.1001 / archpsyc.64.3.327
Escorihuela, RM, Fernández-Teruel, A., Gil, L., Aguilar, R., Tobeña, A. a Driscoll, P. (1999). Inbrední římští potíže s vysokou a nízkou vyhýbací schopností: rozdíly v úzkosti, chování při hledání novinek a v režimu shuttleboxu. Physiol. Behav. 67, 19–26. doi: 10.1016/S0031-9384(99)00064-5
Everitt, BJ (1990). Sexuální motivace: neurální a behaviorální analýza mechanismů, které jsou základem reakce apelace a kopulace samců potkanů. Neurosci. Biobehav. Rev. 14, 217–232. doi: 10.1016/s0149-7634(05)80222-2
Everitt, BJ a Robbins, TW (2005). Neuronové systémy posílení pro drogovou závislost: od akcí po zvyky k nátlaku. Nat. Neurosci. 8, 1481-1489. dva: 10.1038 / nn1579
Fadda, F., Argiolas, A., Melis, MR, Tissari, AH, Onali, PL a Gessa, GL (1978). Stresem indukované zvýšení hladin kyseliny 3,4-dihydroxyfenyloctové (DOPAC) v mozkové kůře a n. Akumulaci: zvrácení diazepamem. Life Sci. 23, 2219–2224. doi: 10.1016/b978-0-08-023768-8.51934-4
Fattore, L., Piras, G., Corda, MG a Giorgi, O. (2009). Římské linky potkanů s vysokým a nízkým výskytem potíží se liší při pořizování, udržování, vyhynutí a obnovení intravenózního kokainového samosprávy. Neuropsychopharmacology 34, 1091-1101. dva: 10.1038 / npp.2008.43
Febo, M. (2011). Prefrontální buněčné palby u samců potkanů při přiblížení se k sexuálně vnímavým ženám: interakce s kokainem. Synapse 65, 271-277. dva: 10.1002 / syn.20843
Fernández-Guasti, A., Omaña-Zapata, I., Luján, M. a Condés-Lara, M. (1994). Opatření ligatury sedacího nervu na sexuální chování sexuálně zkušených a nezkušených samců potkanů: účinky deformace čelního pólu. Physiol. Behav. 55, 577–581. doi: 10.1016/0031-9384(94)90119-8
Fernández-Teruel, A., Driscoll, P., Gil, L., Aguilar, R., Tobeña, A. a Escorihuela, RM (2002). Trvalé účinky obohacování životního prostředí na hledání novinek, sacharin a příjem ethanolu ve dvou liniích potkanů (RHA / Verh a RLA / Verh) se liší od chování motivujícího k pobídce. Pharmacol. Biochem. Behav. 73, 225–231. doi: 10.1016/s0091-3057(02)00784-0
Giménez-Llort, L., Cañete, T., Guitart-Masip, M., Fernández-Teruel, A. a Tobeña, A. (2005). Dva odlišné fenotypy indukované apomorfinem u krys římského vysokého a nízkého vyloučení. Physiol. Behav. 86, 458-466. dva: 10.1016 / j.physbeh.2005.07.021
Giorgi, O., Lecca, D., Piras, G., a Corda, MG (2005). Diferenciální aktivace uvolňování dopaminu v nucleus accumbens jádře a skořápce po akutních nebo opakovaných injekcích amfetaminu: srovnávací studie v římských liniích potkanů s vysokým a nízkým vyloučením. Neurovědy 135, 987-998. dva: 10.1016 / j.neuroscience.2005.06.075
Giorgi, O., Lecca, D., Piras, G., Driscoll, P. a Corda, MG (2003a). Disociace mezi mezokortikálním uvolňováním dopaminu a chováním souvisejícím se strachem ve dvou psychogeneticky vybraných liniích potkanů, které se liší ve strategiích zvládnutí a na odporné podmínky. Eur. J. Neurosci. 17, 2716-2726. dva: 10.1046 / j.1460-9568.2003.02689.x
Giorgi, O., Piras, G., Lecca, D., Hansson, S., Driscoll, P. a Corda, MG (2003b). Diferenciální neurochemické vlastnosti centrální serotonergní translace u potkanů s nízkým a nízkým vyloučením. J. Neurochem. 86, 422-431. dva: 10.1046 / j.1471-4159.2003.01845.x
Giorgi, O., Orlandi, M., Escorihuela, RM, Driscoll, P., Lecca, D. a Corda, MG (1994). GABAergní a dopaminergní přenos v mozku římských vysoce vyhýbavých a římských potkanů s nízkou vyhýbací schopností. Brain Res. 638, 133–138. doi: 10.1016/0006-8993(94)90642-4
Giorgi, O., Piras, G. a Corda, MG (2007). Psychogeneticky vybrané římské linie s vysokým a nízkým vyloučením krys: model, který zkoumá individuální zranitelnost vůči drogové závislosti. Neurosci. Biobehav. Rev. 31, 148-163. dva: 10.1016 / j.neubiorev.2006.07.008
Jdi, Y. a Grace, AA (2005). Dopaminergní modulace limbického a kortikálního posunu nucleus accumbens cíleného chování. Nat. Neurosci. 8, 805-812. dva: 10.1038 / nn1471
Gresch, PJ, Sved, AF, Zigmond, MJ a Finlay, JM (1995). Lokální vliv endogenního norepinefrinu na extracelulární dopamin v mediální prefrontální kůře potkana. J. Neurochem. 65, 111-116. dva: 10.1046 / j.1471-4159.1995.65010111.x
Hernández-Gonzáles, M., Guevara, MA, Cervantes, M., Morali, G., a Corsi-Cabrera, M. (1998). Charakteristické kmitočtové pásmo kortikofronálního EEG během sexuální interakce samčích krys v důsledku faktoriální analýzy. J. Physiology 92, 43–50. doi: 10.1016/s0928-4257(98)80022-3
Hernández-Gonzáles, M., Prieto-Beracoechea, CA, Arteaga-Silva, M. a Guevara, MA (2007). Různá funkčnost mediální a orbitální prefrontální kůry během sexuálně motivovaného úkolu u potkanů. Physiol. Behav. 90, 450-458. dva: 10.1016 / j.physbeh.2006.10.006
Horn, AS (1973). Vztah mezi strukturou a aktivitou pro inhibici vychytávání katecholaminů do synaptozomů z noradrenergních a dopaminergních neuronů v mozkových homogenatech krys. Br. J. Pharmacol. 47, 332–338. doi: 10.1111/j.1476-5381.1973.tb08331.x
Hull, EM, Du, J., Lorrain, DS a Matuszewich, L. (1995). Extracelulární dopamin v oblasti mediální preoptiky: důsledky pro sexuální motivaci a hormonální kontrolu kopulace. J. Neurosci. 15, 7465-7471.
Hull, EM, Lorrain, DS, Du, J., Matuszewich, L., Lumley, LA, Putnam, SK a kol. (1999). Interakce hormonů a neurotransmiterů při kontrole sexuálního chování. Behav. Brain Res. 105, 105–116. doi: 10.1016/s0166-4328(99)00086-8
Hull, EM, Weber, MS, Eaton, RC, Dua, R., Markowski, VP, Lumley, L. a kol. (1991). Dopaminové receptory ve ventrální tegmentální oblasti ovlivňují motorové, ale ne motivační nebo reflexní komponenty kopulace u samců potkanů. Brain Res. 554, 72–76. doi: 10.1016/0006-8993(91)90173-S
Ikemoto, S. a Panksepp, J. (1999). Úloha jádra accumbens dopaminu v motivovaném chování: sjednocující interpretace se zvláštním zřetelem k odměňování. Brain Res. Rev. 31, 6–41. doi: 10.1016/S0165-0173(99)00023-5
Kakeyama, M., Sone, H., Miyabara, Y. a Tohyama, C. (2003). Perinatální expozice 2,3,7,8-tetrachlordibenzo-p-dioxinu mění aktivitu závislou expresi BDNF mRNA u neokortexu a pohlavního chování u samců potkanů v dospělosti. Neurotoxikologie 24, 207–217. doi: 10.1016/S0161-813x(02)00214-0
Lecca, D., Piras, G., Driscoll, P., Giorgi, O., a Corda, MG (2004). Diferenciální aktivace výstupu dopaminu v skořápce a jádře nucleus accumbens je spojena s motorickými reakcemi na návykové drogy: studie na dialyzaci mozku u laboratorních potkanů s vysokým a nízkým vyloučením. Neurofarmakologie 46, 688-699. dva: 10.1016 / j.neuropharm.2003.11.011
Manzo, L., Gómez, MJ, Callejas-Aguilera, JE, Donaire, R., Sabariego, M., Fernández-Teruel, A., et al. (2014a). Vztah mezi preferencemi etanolu a hledáním pojmů / hledáním novinek. Physiol. Behav. 133, 53-60. dva: 10.1016 / j.physbeh.2014.05.003
Manzo, L., Gómez, MJ, Callejas-Aguilera, JE, Fernández-Teruel, A., Papini, MR a Torres, C. (2014b). Anti-úzkostná samolepka vyvolaná ztrátou pobídky u potkanů. Physiol. Behav. 123, 86-92. dva: 10.1016 / j.physbeh.2013.10.002
Masana, M., Bortolozzi, A., a Artigas, F. (2011). Selektivní zlepšení mezokortikálního dopaminergního přenosu noradrenergními léky: terapeutické příležitosti u schizofrenie. Int. J. Neuropsychopharmacol. 14, 53-68. dva: 10.1017 / s1461145710000908
Meisel, RL a Sachs, BD (1994). "Fyziologie mužského sexuálního chování", Fyziologie reprodukce, Vol. 2, 2nd Edn., Vyd. E. Knobil a J. Neil (New York, NY: Raven Press), 3-96.
Melis, MR a Argiolas, A. (1995). Dopamin a sexuální chování. Neurosci. Biobehav. Rev. 19, 19–38. doi: 10.1016/0149-7634(94)00020-2
Melis, MR a Argiolas, A. (2011). Centrální kontrola erekce penisu: opětovné navození role oxytocinu a jeho interakce s dopaminem a kyselinou glutamovou u samců potkanů. Neurosci. Biobehav. Rev. 35, 939-955. dva: 10.1016 / j.neubiorev.2010.10.014
Melis, MR, Melis, T., Cocco, C., Succu, S., Sanna, F., Pillolla, G. a kol. (2007). Oxytocin vstřikovaný do ventrální tegmentální oblasti vyvolává erekci penisu a zvyšuje extracelulární dopamin v nucleus accumbens a paraventrikular jádře hypotalamu samců potkanů. Eur. J. Neurosci. 26, 1026-1035. dva: 10.1111 / j.1460-9568.2007.05721.x
Melis, MR, Succu, S., Cocco, C., Caboni, E., Sanna, F., Boi, A., et al. (2010). Oxytocin indukuje erekci penisu, když je injekčně podán do ventrální subiculum: úloha oxidu dusnatého a kyseliny glutamové. Neurofarmakologie 58, 1153-1160. dva: 10.1016 / j.neuropharm.2010.02.008
Melis, MR, Succu, S., Mascia, MS, Cortis, L., a Argiolas, A. (2003). Extracelulární dopamin zvyšuje v paraventrikulárním jádru samců potkanů během sexuální aktivity. Eur. J. Neurosci. 17, 1266-1272. dva: 10.1046 / j.1460-9568.2003.02558.x
Melis, MR, Succu, S., Sanna, F., Boi, A., a Argiolas, A. (2009). Oxytocin, vstřikovaný do ventrální subskultury nebo do bedromediálního kortikálního jádra amygdaly, vyvolává erekci penisu a zvyšuje hladinu extracelulárního dopaminu v nucleus accumbens samců potkanů. Eur. J. Neurosci. 30, 1349-1357. dva: 10.1111 / j.1460-9568.2009.06912.x
Moreno, M., Cardona, D., Gómez, MJ, Sánchez-Santed, F., Tobeña, A., Fernández-Teruel, A., et al. (2010). Charakterizace impulsivity v římských kmenech potkanů s vysokým a nízkým vyloučením: behaviorální a neurochemické rozdíly. Neuropsychopharmacology 35, 1198-1208. dva: 10.1038 / npp.2009.224
Oliveras, I., Río-Álamos, C., Cañete, T., Blázquez, G., Martínez-Membrives, E., Giorgi, O., et al. (2015). Inhibice předpulsů předpovídá výkon prostorové pracovní paměti u inbredních potkanů s vysokým a nízkým vyloučením a u geneticky heterogenních krys NIH-HS: význam pro studium pre-pozorných a kognitivních anomálií při schizofrenii. Přední. Behav. Neurosci. 9: 213. dva: 10.3389 / fnbeh.2015.00213
Paxinos, G. a Watson, C. (2004). Krysa mozku ve stereotaxických souřadnicích, 4th Edn, New York: Academic Press.
Pfaus, JG (2010). Dopamina: pomáhá mužům kopulovat alespoň za 200 miliony let: teoretická poznámka o Kleitz-Nelson et al. (2010). Behav. Neurosci. 124, 877-880. dva: 10.1037 / a0021823
Pfaus, JG, Damsma, G., Nomikos, GG, Wenkstern, DG, Blaha, CD, Phillips, AG a kol. (1990). Sexuální chování zvyšuje centrální přenos dopaminu u samců potkanů. Brain Res. 530, 345–348. doi: 10.1016/0006-8993(90)91309-5
Pfaus, JG a Everitt, BJ (1995). "Psychofarmakologie sexuálního chování", Psychopharmacology: čtvrtá generace pokroku, ed. FE Bloom a DJ Kupfer (New York, NY: Raven Press), 742-758.
Pfaus, JG a Phillips AG (1991). Role dopaminu v předvídatelných a konzumačních aspektech sexuálního chování u samců potkanů. Behav. Neurosci. 105, 727-747. dva: 10.1037 / 0735-7044.105.5.727
Piras, G., Giorgi, O. a Corda, MG (2010). Účinky antidepresiv na výkon v testu nuceného plavání u dvou psychogeneticky vybraných linií krys, které se liší ve strategiích zvládnutí a na odporné podmínky. Psychofarmakologie 211, 403–414. doi: 10.1007/s00213-010-1904-x
Piras, G., Piludu, MA, Giorgi, O., a Corda, MG (2014). Účinky chronické antidepresivní léčby u předpokládaného genetického modelu zranitelnosti (krysy s nízkou obezitou) a rezistence (římské krysy s vysokou vyhýbací schopností) na stresem vyvolanou depresi. Psychofarmakologie 231, 43–53. doi: 10.1007/s00213-013-3205-7
Pitchers, KK, Frohmader, KS, Vialou, V., Mouzon, K., Nestler, EJ, Lehman, MN a kol. (2010). ΔFosB v nucleus accumbens je rozhodující pro posílení účinků sexuální odměny. Genes Brain Behav. 9, 831–840. doi: 10.1111/j.1601-183X.2010.00621.x
Pitchers, KK, Coppens, CM, Beloate, LN, Fuller, J., Van, S., Frohmader, KS a kol. (2014). Endogenní opioidně indukovaná neuroplasticita dopaminergních neuronů ve ventrální tegmentální oblasti ovlivňuje přírodní a opiátovou odměnu. J. Neurosci. 34, 8825-8836. dva: 10.1523 / JNEUROSCI.0133-14.2014
Pitchers, KK, Vialou, V., Nestler, EJ, Laviolette, SR, Lehman, MN a Coolen, LM (2013). Přírodní a drogové odměny působí na společné mechanismy neurální plasticity s DFosB jako klíčovým prostředníkem. J. Neurosci. 33, 3434-3442. dva: 10.1523 / JNEUROSCI.4881-12.2013
Pleim, E., Matochik, J., Barfield, R., a Auerbach, S. (1990). Korelace uvolňování dopaminu v nucleus accumbens s mužským sexuálním chováním u potkanů. Brain Res. 524, 160–163. doi: 10.1016/0006-8993(90)90507-8
Robbins, TW a Arnsten, AFT (2009). Neuropsychofarmakologie fronto-výkonné funkce: monoaminergní modulace. Ann. Rev. Neurosci. 32, 267-287. dva: 10.1146 / annurev.neuro.051508.135535
Sabariego, M., Morón, I., Gómez, MJ, Donaire, R., Tobeña, A., Fernández-Teruel, A., et al. (2013). Stimulační ztráta a exprese hipokampálního genu u inbredních krys s nízkou (RLA-I) nízkým (RLA-I) potlačením s vysokým (RHA-I) románem. Behav. Brain Res. 257, 62-70. dva: 10.1016 / j.bbr.2013.09.025
Sachs, BD (2000). Kontextuální přístupy k fyziologii a klasifikaci erektilní funkce, erektilní dysfunkce a sexuální vzrušení. Neurosci. Biobehav. Rev. 24, 541–560. doi: 10.1016/S0149-7634(00)00022-1
Sachs, BD, Akasofu, K., Citro, JK, Daniels, SB, a Natoli, JH (1994). Bezkontaktní stimulace od estrusových žen vyvolává erekci penisu u potkanů. Physiol. Behav. 55, 1073–1079. doi: 10.1016/0031-9384(94)90390-5
Sachs, BD a Barfield, RJ (1976). Funkční analýza chování maskulinního kopulace u potkanů. Adv. Stud. Behav. 7, 91–154. doi: 10.1016/s0065-3454(08)60166-7
Salamone, JD, Pardo, M., Yohn, SE, López-Cruz, L., San Miguel, N. a Correa, M. (2016). Mezolimbický dopamin a regulace motivovaného chování. Curr. Horní. Behav. Neurosci. 27, 231–257. doi: 10.1007/7854_2015_383
Sanna, F., Corda, MG, Melis, MR, Piludu, MA, Giorgi, O., a Argiolas, A. (2014a). Samci římských vysokých a nízkých vyhýbacích krys ukazují různé vzorce kopulačního chování: srovnání s krysami sprague dawley. Physiol. Behav. 127, 27-36. dva: 10.1016 / j.physbeh.2014.01.002
Sanna, F., Piludu, MA, Corda, MG, Argiolas, A., Giorgi, O., a Melis, MR (2014b). Dopamín se podílí na různých vzorcích kopulačního chování u potkanů s vysokým a nízkým vyloučením v Římě: studie s apomorfinem a haloperidolem. Pharmacol. Biochem. Behav. 124, 211-219. dva: 10.1016 / j.pbb.2014.06.012
Sanna, F., Piludu, MA, Corda, MG, Melis, MR, Giorgi, O., a Argiolas, A. (2015). Zapojení dopaminu v rozdílech v sexuálním chování u potkanů s vysokým a nízkým rizikem potratu: intracerebrální mikrodialýza. Behav. Brain Res. 281, 177-186. dva: 10.1016 / j.bbr.2014.12.009
Scatton, B., D'Angio, M., Driscoll, P. a Serrano, A. (1988). An in vivo voltammetrická studie odezvy mezokortických a mesoakcumbens dopaminergních neuronů na environmentální stimuly u kmenů potkanů s rozdílnou úrovní emocionality. Ann. NY Acad. Sci. 537, 124–137. doi: 10.1111/j.1749-6632.1988.tb42101.x
Steimer, T. a Driscoll, P. (2003). Odchylná odezva na stres a styly v psychogeneticky vybraných římských krysích s vysokou (RHA) a nízkou (RLA) vyhýbáním: behaviorální, neuroendokrinní a vývojové aspekty. Stres 6, 87-100. dva: 10.1080 / 1025389031000111320
Succu, S., Sanna, F., Argiolas, A. a Melis, MR (2011). Oxytocin injikovaný do hipokampální ventrální subiculum vyvolává erekci penisu u samců potkanů zvýšením glutamatergické neurotransmisi ve ventrální tegmentální oblasti. Neurofarmakologie 61, 181-188. dva: 10.1016 / j.neuropharm.2011.03.026
Succu, S., Sanna, F., Cocco, C., Melis, T., Boi, A., Ferri, GL a kol. (2008). Oxytocin indukuje erekci penisu po injekci do ventrální tegmentální oblasti samců potkanů: úloha oxidu dusnatého a cyklického GMP. Eur. J. Neurosci. 28, 813-821. dva: 10.1111 / j.1460-9568.2008.06385.x
Succu, S., Sanna, F., Melis, T., Boi, A., Argiolas, A. a Melis, MR (2007). Stimulace dopaminových receptorů v paraventrikulárním jádru hypotalamu samců krys vyvolává erekci penisu a zvyšuje extracelulární dopamin v nucleus accumbens: zapojení centrálního oxytocinu. Neurofarmakologie 52, 1034-1043. dva: 10.1016 / j.neuropharm.2006.10.019
Thierry, AM, Tassin, JP, Blanc, G. a Glowinski, J. (1976). Selektivní aktivace mezokortikálního DA systému stresem. Příroda 263, 242-244. dva: 10.1038 / 263242a0
Wenkstern, D., Pfaus, J., a Fibiger, H. (1993). Přenos dopaminu se zvyšuje v nucleus accumbens samců potkanů během prvního vystavení sexuálně citlivým samicím potkanům. Brain Res. 618, 41–46. doi: 10.1016/0006-8993(93)90426-N
Westernik, BHC, De Boer, P., De Vries, JB a Long, S. (1998). Antipsychotické léky vyvolávají podobné účinky na uvolňování dopaminu a noradrenalinu v mediálním prefrontálním kortexu mozku krys. Eur. J. Pharmacol. 361, 27–33. doi: 10.1016/s0014-2999(98)00711-0
Willig, F., M'Harzi, M., Bardelay, C., Viet, D. a Delacour, J. (1991). Řízené kmeny jako psychogenetický model pro studium pracovní paměti: chování a biochemické údaje. Pharmacol. Biochem. Behav. 40, 7–16. doi: 10.1016/0091-3057(91)90313-q
Klíčová slova: sexuální chování, dopamin, noradrenalin, mPFC, mikrodialýza, potkaní RHA a RLA
Citace: Sanna F, Bratzu J, Piludu MA, Corda MG, Melis MR, Giorgi O a Argiolas A (2017) Dopamin, Noradrenalin a rozdíly v sexuálním chování mezi romskými vysokými a nízkopravskými samci potkanů: Mikrodialyzační studie v mediální prefrontální kortex . Přední. Behav. Neurosci. 11: 108. dva: 10.3389 / fnbeh.2017.00108
Přijato: 22 prosinec 2016; Přijato: 19 May 2017;
Publikováno: 07 Červenec 2017.