Neurosci Biobehav Rev. 2011 Jan; 35 (3): 939-55. dva: 10.1016 / j.neubiorev.2010.10.014. Epub 2010 Nov 2.
- 1Bernard B. Brodie Oddělení neurologie, Univerzita v Cagliari, Cittadella Universitaria, 09042 Monserrato, CA, Itálie. [chráněno e-mailem]
ABSTRAKTNÍ
Oxytocin je silným induktorem erekce penisu při injekci do centrálního nervového systému. U samců potkanů je nejcitlivější oblastí mozku pro pro-erektilní účinek oxytocinu paraventrikulární jádro hypotalamu. Toto jádro a okolní oblasti obsahují buněčné těla všech oxytocinergních neuronů, které vyčnívají do extra-hypothalamických oblastí mozku a míchy. Tento přehled ukazuje, že oxytocin indukuje erekci penisu také při injekci v některých z těchto oblastí (např. Ventrální tegmentální oblast, ventrální subiculum hipokampu, posteromediální kortikální jádro amygdaly a thorako-bederní mícha). Mikroinjekční studie kombinované s intramolekulárními mikrodialyzačními a dvojitě imunofluorescenčními studiemi naznačují, že oxytocin v těchto oblastech přímo nebo nepřímo aktivuje mezolimbické dopaminergní neurony (hlavně prostřednictvím kyseliny glutamové). Dopamin uvolněný v nucleus accumbens naopak aktivuje nervové dráhy vedoucí k aktivaci inert-hypothalamických dopaminergních neuronů v paraventrikulárním jádru. Tím se aktivují nejen oxytocinergní neurony, které vyčnívají do míchy a zprostředkovávají erekci penisu, ale také ty, které vyčnívají do výše uvedených extra-hypothalamických oblastí, a modulují přímo nebo nepřímo (kyselinou glutamovou) aktivitu mezolimbických dopaminergních neuronů, které ovládají motivaci a odměnu. Společně tyto neurální dráhy mohou představovat komplexní hypotetický obvod, který hraje roli nejen v konzumační fázi sexuální aktivity (erektilní funkce a kopulace), ale také v motivačních a odměřujících aspektech předvídatelné fáze sexuálního chování.
1.Introduction
Erekce penisu je mužská sexuální odpověď, která hraje klíčovou roli při reprodukci savců, včetně člověka, a to lze také pozorovat v kontextech odlišných od těch, která jsou přísně spojena s reprodukcí. V závislosti na kontextu, ve kterém dochází k erekci penisu, se na jeho regulaci podílejí různé centrální a periferní nervové a / nebo humorální mechanismy (viz Meisel a Sachs, 1994, Argiolas a Melis, 1995, 2004, 2005, Sachs, 2000, 2007, McKenna, 2000, Giuliano a Rampin, 2000, 2004, Andersson, 2001, Melis a Argiolas, 1995a, 2003, Hull a kol., 2002). Mezi centrálními neurotransmitery a neuropeptidy, které kontrolují erekci penisu, jsou nejznámější dopaminu, serotoninu, excitačních aminokyselin, oxidu dusnatého, adrenokortikotropinu, oxytocinu a opioidních peptidů. Mohou usnadnit nebo inhibovat erekci penisu působením v několika oblastech mozku, tj. mediální preoptickou oblast, paraventrikulární jádro hypotalamu, ventrální tegmentální oblast, hipokampus, amygdala, jádro lůžka stria terminis, nucleus accumbens, medulla oblongata a míchu (Tabulka 1) (viz Meisel a Sachs, 1994, Witt a Insel, 1994, Stancampiano a kol., 1994, Argiolas a Melis, 1995, 2005, Argiolas, 1999, Bancila a spol., 2002, Giuliano a Rampin, 2000, McKenna, 2000, Andersson, 2001, Hull a kol., 2002, Coolen a kol., 2004).
Oxytocin, neurohypofýzový peptid dobře známý pro jeho hormonální úlohu při laktaci a porodu, je přítomen u žen a mužů nejen u neuronů s tělískymi buňkami lokalizovanými v paraventrikulárních a supraoptických jádrech hypothalamu vyčnívajícím do neurohypofýzy, ale také v neuronech vyčnívajících od paraventrikulárního jádra a okolních struktur až po extrahypothalamické oblasti mozku (tj. septum, ventrální tegmentální oblast, hippocampus, amygdala, medulla oblongata a míchu). Předpokládá se, že tyto neurony se podílejí na četných ústředních funkcích, jako je paměť, učení, affilitivní a sociálně-sexuální chování, včetně erekce penisu a kopulačního chování (viz Buijs, 1978, Sofroniew, 1983, Argiolas a Gessa, 1991; , 1992, Carter, 1992, Wagner a Clemens, 1993, Ivell a Russel, 1995, Carter a kol., 1997, Tang a další, 1998, Veronneau-Longueville a kol., 1999). Oxytocin skutečně usnadňuje erektilní funkci a mužské sexuální chování u myší, potkanů, králíků a opic (viz Argiolas a Gessa, 1991, Carter, 1992, Pedersen a kol., 1992, Argiolas a Melis, 1995, 2004, Argiolas, 1999). To se může objevit i u lidí, protože plazmatický oxytocin se zvyšuje sexuálními stimuly, zejména při ejakulaci (Carmichael et al., 1987, Murphy a kol., 1987) a manipulací prsu a genitálií, pohlavní styk (Tindall, 1974).
Jednoduchý účinek oxytocinu na mužské sexuální chování byl nejprve prokázán schopností intravenózního oxytocinu snížit latenci k první ejakulaci a zpomalit sexuální vyčerpání samců králíků spárovaných s receptivními ženami (Melin a Kihlstrom, 1963). Ovšem sexuální účinky oxytocinu byly definitivně uznány teprve v osmdesátých letech. Oxytocin podávaný centrálně v množství nanogramů byl nalezen schopný vyvolat erekci penisu (Argiolas et al., 1985, 1986) a zlepšit kopulační chování (Arletti a kol., 1985) u samců potkanů a zvýšení lordózy u samic potkanů (Arletti a Bertolini, 1985, Caldwell a kol., 1986), zjevně působením na oxytocinergní receptory děložního typu (viz Argiolas a Melis, 1995, 2004, Argiolas, 1999, Melis a Argiolas, 2003 a odkazy v ní). Oxytocin zlepšuje sexuální chování nejen u pohlavně silných samců potkanů (Arletti et al., 1985), ale také u starších samců potkanů (Arletti et al., 1990) a dominantních, ale nikoliv podřízených, opicích samců (Winslow a Insel , 1991).
Pro-erektilní účinek oxytocinu je závislý na testosteronu, protože je zrušen hypofysektomií a kastrací a obnovován doplněním testosteronem nebo jeho metabolity, estradiolem a 5-dihydro-testosteronem společně (Melis a kol., 1994a). Nejcitlivější oblastí mozku pro indukci erekce penisu pomocí oxytocinu je paraventrikulární jádro hypotalamu (Melis a kol., 1986), z nichž pocházejí všechny extra-hypotalamické oxytocinergní projekce (viz výše). Ukázalo se, že oxytocin je schopen vyvolat erekci penisu (a zívání), když je podáván v dávkách tak nízkých, jako je 3 pmol (viz níže v části 2.1). Oxytocin indukuje erekci penisu také při dvoustranném injekčním podání do oblasti CA1 hipokampu, avšak nikoliv v dorzální podušce (viz oddíl 2.3 níže), boční septa, kaudátové jádro, mediální preoptická oblast, ventromedické jádro hypotalamu a supraoptické jádro (Melis a kol., 1986). Co se týká mechanismu, kterým oxytocin působí v paraventrikulárním jádru k vyvolání této sexuální odpovědi, řada studií naznačuje, že oxytocin aktivuje své vlastní neurony. V souladu s touto hypotézou zvyšuje sexuální interakce FOS, genový produkt okamžitého časného genu c-fos v paraventrikulárních oxytocinergních neuronech vyčnívajících do míchy, které se podílejí na kontrole erekce penisu (viz Witt a Insel, 1994 a reference ) a sexuální impotence (např. neschopnost dospělého samčího potkana kopulovat s ovarektomizovanou samicí receptem vyvolanou estrogen-progesteronem) byla spojena u samců potkanů s nízkými hladinami oxytocinové mRNA v paraventrikulárním jádře hypotalamu (Arletti et al., 1997).
To, zda oxytocin ovlivňuje předběžnou fázi nebo spotřebitelskou fázi sexuálního chování, je v současné době nejasné. Vzhledem k tomu, že oxytocin indukuje erekci penisu a hlavním účinkem oxytocinu na kopulační chování je pokles post-ejakulačního intervalu u samců potkanů (Arletti et al., 1985), je rozumné předpokládat, že peptid zlepšuje sexuální výkonnost. Nicméně, jak oxytocin také zvyšuje socio-sexuální interakce (viz Pedersen et al., 1992, Carter et al., 1997, Ivell a Russel, 1995) a antagonisté oxytocinového receptoru zabraňují bezkontaktní erekci (Melis a kol., 1999a) jsou považovány za index sexuálního vzrušení (viz Sachs, 1997, 2000, 2007, Melis a kol., 1998, 1999b a odkazy v ní), nelze vyloučit možnou úlohu oxytocinu v sexuálním vzrušení a sexuální motivaci.
Tento přehled shrnuje publikované a nepublikované výsledky nedávných studií, které ukazují, že oxytocin indukuje erekci penisu nejen při vstříknutí do paraventrikulárního jádra hypotalamu, ale také v dalších extra-hypothalamických oblastech mozku, jako je ventrální tegmentální oblast (Melis a kol., 2007, 2009a, Succu a kol., 2008), ton ventral subiculum hippocampus a posterior jádro amygdala (Melis a spol., 2009b, 2010), které jsou důležitými složkami limbického systému a které jsou považovány za klíčové pro motivační a odměňování procesy. Tyto studie ukazují, že oxytocin se účastní neurálních okruhů, mezi něž patří další neurotransmitery, jako je dopamin a kyselina glutamová, a další oblasti mozku jiné než paraventrikulární jádro, např. Ventrální tegmentální oblast, nucleus accumbens, hippocampus a oblasti, identifikovány. Tyto obvody pravděpodobně zprostředkovávají interakci mezi mezolimbickým a incerto-hypothalamickým dopaminergním systémem a hrají roli nejen v konzumační fázi sexuálního chování mužů (např. Erekce penisu a kopulace), ale také v sexuální motivaci a sexuální motivaci vzrušení, čímž poskytuje nervový substrát pro vysvětlení motivačních a odměřujících vlastností sexuální aktivity.
2. Oxytocin ovlivňuje erekci penisu působením v různých oblastech mozku
2.1. Paraventrikulární jádro hypotalamu
Jak bylo uvedeno výše, paraventrikulární jádro hypotalamu bylo brzy identifikováno jako oblast mozku, která je nejcitlivější pro proreretileliální účinek oxytocinu. Při jednostranném vstřiku do tohoto jádra byl oxytocin účinný v dávkách tak nízkých, jako je 3 ng (3 pmol) (Melis a kol., 1986). Studie vztahu mezi strukturou a aktivitou odhalily, že oxytocinem indukovaná erekce penisu byla zprostředkována uterotypovými oxytocinovými receptory, spojenými s influxem Ca2 + do buněčných tělísek oxytocinergních neuronů, které vyčnívají do extra-hypothalamických oblastí mozku ak aktivaci syntázy oxidu dusnatého. Oxid dusnatý působí jako intracelulární posel s neznámým mechanismem (nezahrnuje guanylátcyklázu) vede k aktivaci oxytocinergních neuronů vyčnívajících do oblasti míchy a extra-hypothalamických oblastí mozku, což vyvolává erekci penisu (viz obr. 1) (viz. níže a Argiolas a Melis, 1995, 2004, 2005 a odkazy v ní). Schopnost oxytocinu aktivovat vlastní neurony
Obr. 1. (MISSING) Schematické znázornění oxytocinergních neuronů, které pocházejí z paraventrikulárního jádra hypotalamu a projevují se extra-hypothalamické oblasti mozku, jako je mícha, VTA, hippocampus, amygdala atd. Aktivace těchto neuronů dopaminu, excitačních aminokyselin, samotného oxytocinu, peptidů hexarelinových analogů a peptidů odvozených z VGF vede k erekci penisu, která může být redukována a / nebo zrušena stimulací receptorů CBABRN, opioidů a kanabinoidů CB1. Aktivace oxytocinergních neuronů je sekundární pro aktivaci syntázy oxidu dusnatého přítomného v těchto neuronech. Vskutku endogenní oxid dusnatý tvořený stimulací dopaminových, excitačních aminokyselin nebo oxytocinových receptorů nebo exogenního oxidu dusnatého, který pochází z donorů oxidu dusnatého přímo podávaných do paraventrikulárního jádra, aktivuje oxytocinergní neurony dosud neidentifikovaným mechanismem, zjevně nesouvisí s stimulace guanylátcyklázy. To způsobuje uvolnění oxytocinu v míchu a extra-hypothalamických oblastech mozku. Některé podrobnosti o mechanismech, kterými oxytocin indukuje erekci penisu při uvolnění v těchto oblastech, např. VTA, ventrální subiculum a amygdala, jsou popsány v příslušných sekcích oblasti mozku. Ovšem oxytocin působí na své vlastní receptory a zvyšuje produkci NO, což vede k erekci penisu, jak se nachází v PVN. Nicméně, v rozporu s PVN, v kaudální VTA NO aktivuje guanylát cyklázu. To způsobuje zvýšení koncentrace cGMP, což vede k aktivaci mezolimbických dopaminergních neuronů ak erekci penisu. Ve VS NO se aktivují glutamatergické neurony, které vyčnívají do mimo-hippocampálních oblastí, včetně VTA. Kyselina glutamová ve VTA aktivuje mezolimbické dopaminergní neurony, které byly nalezeny u oxytocinu. Mechanismy podobné těm, které jsou popsány výše, budou pravděpodobně fungovat také, když dojde k erekci penisu ve fyziologickém kontextu, a to jako když jsou samci potkani umístěni v přítomnosti nepřístupné samice receptu (např. Nekontaktní erekce) nebo během kopulace.
v paraventrikulárním jádru bylo podpořeno studiemi, které ukazují, že: (1) oxytocinové receptory jsou přítomny v tomto hypotalamickém jádru (Freund-Mercier a kol., 1987, Freund-Mercier a Stoeckel, 1995); (2) oxytocin usnadňuje jeho vlastní uvolňování in vitro a in vivo (Freund-Mercier a Richard, 1981, 1984, Moos a kol., 1984); a (3) oxytocin excituje své vlastní neurony tím, že působí v paraventrikulárním jádru (Yamashita a kol., 1987). Navíc byly v paraventrikulárním a supraoptickém jádře hypotalamu identifikovány oxytocinergní synapsy, které zasahují do buněčných tělísek magnocelulárních oxytocinergních neuronů (Theodosis, 1985). Konečně, destrukce centrálních oxytocinergních neuronů elektrolytickými nebo chemickými excitotoxickými lézemi paraventrikulárního jádra, které zcela vyčerpává obsah oxytocinu napříč centrálním nervovým systémem a míchu, ruší nejen proreretilní účinek oxytocinu, ale také zhoršuje indukované léky erekce penisu a nekontaktní erekce (viz níže a Argiolas et al., 1987a, b; Liu a kol., 1997 a odkazy v ní). Výsledky podobné těm, které se objevily s lézemi paraventrikulárního jádra, byly nalezeny u silných a selektivních antagonistů receptoru oxytocinu. Tyto sloučeniny byly skutečně injektovány do paraventrikulárního jádra v množství nanogramů, které zabraňovalo úplné erekci penisu indukované oxytocinem, zatímco při podání do laterálních komor bránily nejen erekci penisu indukované samotným oxytocinem, ale také erekci penisu indukované léčivem (viz část 3 níže (Melis et al., 1995a) a byly mimoto extrémně účinné při zhoršení kopulačního chování sexuálně silných samců krys (Argiolas a kol., 2004). Navíc sexuální interakce zvyšuje FOS, genový produkt okamžitého časného genu c-fos v paraventrikulárních oxytocinergních neuronech vyčnívajících do míchy, které se podílejí na kontrole erekce penisu (viz Witt a Insel, 2005 a tam uvedené odkazy). Konečně, sexuální impotence (např. neschopnost dospělého samčího potkana k kopulaci s estrogen-progesteronem připraveným samice) hstejně jako u samců potkanů s nízkou hladinou oxytocinové mRNA a syntázy oxidu dusnatého v paraventrikulárním jádře hypotalamu (Benelli et al., 1995, Arletti a kol., 1997) (pro rozsáhlý přehled těchto studií viz Argiolas, 1999, Argiolas a Melis, 2004, 2005).
2.2. Ventrální tegmentální oblast
Ventrální tegmentální oblast byla objevena teprve nedávno jako místo mozku, ve kterém oxytocin indukuje erekci penisu. Tato oblast obsahuje oxytocinergní nervové zakončení pocházející z paraventrikulárního jádra a oxytocinových receptorů (Freund-Mercier a kol., 1987, Vaccari a kol., 1998). Přesněji bylo zjištěno, že oxytocin je schopen ivyvolává erekci penisu, když je jednostranně vstřikována do kaudální, ale nikoli do ventrální tegmentální oblasti v závislosti na dávce (Melis a kol., 2007). Aktivní dávky byly vyšší než dávky potřebné při injekci do paraventrikulárního jádra a podobné těm, které indukují erekci penisu, když jsou injektovány do ventrálního subiculu hipokampu nebo do bedromediálního kortikálního jádra amygdaly (viz níže). Zdá se, že pro-erektilní účinek je zprostředkován aktivací mezolimbických dopaminergních neuronů, které vystupují do skořápky jádra accumbens, což dále aktivuje dosud neznámé neurální dráhy, které vyčnívají k inertně hypotalamickým dopaminergním neuronům, působícím na paraventrikulární oxytocinergní neurony zprostředkovávající erekci penisu (Melis a kol., 2007, 2009a).
Pokud jde o mechanismus, kterým oxytocin aktivuje dopaminergní neurotransmisi ve ventrální tegmentální oblasti, dostupné údaje naznačují, že oxytocin stimuluje oxytocinergní receptory umístěné v buněčných tělese mezolimbických dopaminergních neuronů. Tím se zvyšuje přítok Ca2 + uvnitř buněčných těl dopaminergních neuronů, čímž se aktivuje syntáza oxidu dusnatého (Succu et al., 2008). V rozporu s paraventrikulárním jádrem (viz část 3 níže) oxid dusnatý aktivuje guanylát cyklázu, čímž zvyšuje koncentraci cyklického GMP. V souladu s tímto mechanismem byl buď silný inhibitor neuronální syntázy oxidu dusnatého d) (CH2) 5Tyr (Me) 2-Orn8-vazotocin, silný antagonista oxytocinu nebo S-methylthio-1-citrulin kaudální ventrální tegmentální oblast před oxytocinem, zrušení erekce penisu a zvýšení koncentrace extracelulárního dopaminu v skořápce nucleus accumbens indukovaného oxytocinem. Navíc 8-bromocyklický GMP, cyklický analog GMP, který je účinný proti fosfodiesterázám, indukuje erekci penisu při vstřikování do kaluální ventrální tegmentální oblasti a zvyšuje koncentraci extracelulárního dopaminu v skořápce nucleus accumbens, jak bylo zjištěno u oxytocinu injikovaného do (Succu et al., 2008, Melis a kol., 2009a) (viz také obr. 2).
Vždy v souladu s tímto mechanismem haloperidol, silný antagonista dopaminového D2 receptoru, vstřikovaný do skořápky nucleus accumbens redukuje erekci penisu indukovanou oxytocinem vstřikovaným do ventrální tegmentální oblasti (Melis et al., 2007). Uvedený mechanismus je také podporován studiemi s dvojitou imunofluorescencí, která ukazují, že oxytocinová vlákna narážou na buněčné těla dopaminergních neuronů v ocasní ventrální tegmentální oblasti, které byly předtím označeny retrográdní značkou Fluorogold vstříknutou do skořápky nucleus accumbens Melis a kol., 2007, Succu a kol., 2008). Aktivace těchto dopaminergních neuronů a dopaminových receptorů v nucleus accumbens vede naopak k aktivaci neurálních cest, které ještě mají být identifikovány, což stimuluje inertní hypotalamické dopaminergní neurony k uvolňování dopaminu v paraventrikulárním jádru, čímž se aktivují oxytocinergní neurony, míchy a zprostředkování erekce penisu (viz výše a Melis a kol., 2007, Succu a kol., 2007, 2008). Ve skutečnosti oxytocin vstřikovaný do kaudální ventrální tegmentální oblasti v dávce, která vyvolala erekci penisu, zvýšila extracelulární koncentraci dopaminu v dialyzátu získaném nejen z nucleus accumbens, ale také z paraventrikulárního jádra (Succu et al., 2007).
2.3. Hippocampus
Pole CA1 hipokampu bylo další oblasti mozku bohaté na oxytocinergní vlákna a receptory identifikované v dřívějších studiích, ve kterých injekce oxytocinu vyvolala erekci penisu (viz Bujis, 1978, Sofroniew, 1983). Nicméně, v rozporu s paraventrikulárním jádrem, bylo zjištěno, že oxytocin je schopen indukovat erekci penisu pouze tehdy, když je podáván bilaterálně a ve vyšších dávkách než ty, které jsou aktivní v paraventrikulárním jádru (Melis et al., 1986, Chen et al., 1992). Injekce oxytocinu do subiculum byly v těchto dřívějších studiích neúčinné. Nedávná a pozornější studie mikroinjekce však vedou k identifikaci a oblasti ventrálního subiculu ve kterém byla injekce oxytocinu schopna indukovat erekci penisu způsobem závislým na dávce (Melis et al., 2009b). Pro-erektilní účinek oxytocinu injikovaného do této oblasti mozku byl pozorován při dávkách podobných těm, které byly zjištěny ve ventrální tegmentální oblasti po unilaterální injekci (Melis et al., 2007), jak se vyskytuje v paraventrikulárním jádru. Zřejmě oxytocin vstřikovaný do ventrální subiculum vyvolává erekci penisu aktivaci oxytocinergních receptorů v neuronech obsahujících oxid dusnatý-syntázu, což způsobuje zvýšení produkce oxidu dusnatého. Oxid dusnatý, který působí jako intercelulární posel, aktivuje neurotransmisi kyseliny glutamové, což vede k erekci penisu, případně prostřednictvím neurálních (glutamatergických) eferentních projekcí z ventrální subiculum na extra-hipokampální oblasti mozku modulující aktivitu mezolimbických dopaminergních neuronů (např. ventrální tegmentální oblast, prefrontální kůra, paraventrikulární jádro) (viz níže a Melis, 2007, 2009b, Succu a spol., 2008).
Tento mechanismus účinku je podporován intramolekulárními mikrodialyzačními experimenty, které ukazují, že oxytocin vstřikovaný do ventrální subskuliny v dávkách, které indukují erekci penisu, zvyšuje produkci oxidu dusnatého a koncentraci extracelulární kyseliny glutamové v dialyzátu z ventrálního subiculu (Melis et al. , 2010) a extracelulárního dopaminu v nucleus accumbens (Melis a kol., 2007). Tyto reakce byly antagonizovány nejen antagonistou oxytocinového receptoru d (CH2) 5Tyr (Me) 2-Orn8-vazotocinem, ale také inhibitorem neuronální oxid-syntázy oxidu dusnatého S-methylthio-lcitrulinem a hemoglobinem, do ventrální subiculum několik minut před oxytocinem (Melis et al., 2010).
Navíc, v souladu s tímto mechanismem účinku, aktivace glutamatergické neurotransmise NMDA vstřikovanou do ventrální subiculum vyvolává erekci penisu (Melis et al., 2010). Fenotyp eferentních projekcí z ventrální subiculum, které způsobují aktivaci mezolimbických dopaminergních neuronů a zvýšení extracelulárního dopaminu v nucleus accumbens, není v současné době známo. Avšak vzhledem k tomu, že erekce penisu indukovaná oxytocinem vstřikovaným do ventrálního subiculu souvisí se zvýšením extracelulární kyseliny glutamové v dialyzátu z ventrální tegmentální oblasti, ale nikoliv z nucleus accumbens a antagonizuje se (+) MK-801, (Woodruff et al., 1987), který je injektován do ventrální tegmentální oblasti, avšak nikoliv v nucleus accumbens (viz obr. 2 a Melis a kol., 2009b) , je pravděpodobné, že tyto projekce vedou k aktivaci glutamatergické neurotransmisi ve ventrální tegmentální oblasti, která pak aktivuje mezolimbické dopaminergní neurony, které vyčnívají do nucleus accumbens. Zda je zvýšená koncentrace kyseliny glutamové zjištěná ve ventrální tegmentální oblasti po injekci oxytocinu do ventrální subiculum uvolněna z neuronů pocházejících z subskulu nebo z jiných oblastí mozku (např. Prefrontální kůra) není v současnosti známa. Přesto, toto způsobuje aktivaci mezolimbických dopaminergních neuronů a zvýšené uvolňování dopaminu v nucleus accumbens. Aktivace dopaminových receptorů vede k aktivaci inert-hypothalamických dopaminergních neuronů, uvolňování dopaminu v paraventrikulárním jádru, čímž se aktivují oxytocinergní neurony, které vyčnívají do míchy a zprostředkovávají erekci penisu (viz výše a Melis a kol., 2007, 2009a, Succu a kol., 2008).
2.4. Amygdala
Amygdala je další oblast mozku bohatá na oxytocinová vlákna a receptory (viz Freund-Mercier a kol., 1987, Vaccari a kol., 1998, Uhl-Bronner a kol., 2005). Předpokládá se, že oxytocin se podílí na různých funkcích, od anxiolýzy, sociální paměti a poznávání, sociálně posilněného učení, emoční empatie, emoční zpracování obličeje a strachu u lidí k erektilní funkci a sexuálnímu chování (viz Kondo et al., 1998, Dominguez et al. al., 2001, Ebner a kol., 2005, Huber a kol., 2005, Domes a kol., 2007, Petrovic a spol., 2008, Lee et al., 2009, Donaldson and Young, 2009, Hurlemann a kol. , 2010). Nicméně, schopnost oxytocinu indukovat erekci penisu u samců potkanů, když byla indukována v posteromediálním kortikálním jádru amygdaly, byla objevena pouze nedávno (Melis et al., 2009b). Tato reakce se objevila současně se zvýšením koncentrace extracelulárního dopaminu v dialyzátu získaném z pláště jádra accumbens, jak bylo zjištěno po injekci oxytocinu do ventrální subskula (Melis et al., 2009b). Mechanismus, kterým oxytocin vstřikovaný do bedromediálního kortikálního jádra amygdaly vyvolává erekci penisu, není v tuto chvíli známo. Dostupné údaje ukazují, že jak erekce penisu, tak zvýšení koncentrace extracelulárního dopaminu v dialyzátu získaném z nucleus accumbens jsou zprostředkovány aktivací oxytocinergních receptorů, neboť oba odpovědi byly zrušeny antagonistou oxytocinového receptoru d (CH2) 5Tyr Me) 2-Orn8-vazotocin injikovaný v jádře amygdaly několik minut před oxytocinem (Melis et al., 2009b).
Ať je jakýkoli mechanismus aktivován oxytocinem v posteromediálním kortikálním jádru amygdaly, sexuální odpověď indukovaná peptidem je zrušena blokádou všech dopaminergních receptorů s cis-flupenthixolem injikovaným do skořápky nucleus accumbens a blokádou NMDA receptorů ( +) MK-801 byl injikován do ventrální tegmentální oblasti, avšak nikoliv do nucleus accumbens, jak bylo zjištěno při erekci penisu indukované oxytocinem vstřikovaným do ventrální subiculum (Melis et al., 2009b). To naznačuje, že oxytocin vstřikovaný do bedromediálního jádra amygdaly aktivuje neurotransmisi kyseliny glutamové ve ventrální tegmentální oblasti. Toto způsobuje aktivaci mezolimbických dopaminergních neuronů, což vede k erekci penisu. Vzhledem ke studiím ukazujícím neurální dráhy propojující toto jádro amygdaly s ventrálním subiculum (Canteras et al., 1995, French a Totterdell, 2003), tyto nálezy zvyšují možnost, že mezi těmito dvěma oblastmi mozku může existovat interakce, byly popsány cesty od amygdaly buď k nucleus accumbens nebo k ventral tegmental oblasti (Kelley a Domesick, 1982, Witter, 2006).
2.5. Mícha
Mícha je další oblastí centrálního nervového systému, která obsahuje oxytocinergní vlákna a receptory (Freund-Mercier a spol., 1987, Uhl-Bronner a kol., 2005), ve kterých oxytocin indukuje erekci penisu (Tang et al., 1998 Veronneau-Longueville a kol., 1999, Giuliano a Rampin, 2000, Giuliano a kol., 2001). Jak již bylo uvedeno výše, tato oxytocinergní vlákna pocházejí z paraventrikulárního jádra hypotalamu a přispívají ke sestupným cestám, které kontrolují autonomní neurony v oblasti páteře a zprostředkovávají erekci penisu. Tato vlákna skutečně vytvářejí synaptické kontakty v kolonách preganglionických sympatetických a parasympatických buněk dorsálního rohu v torakově-lumbárním a lumbosakrálním traktu s spinálními neurony, které inervují penisní kavernózní korpusy (Marson a McKenna, 1996, Giuliano a Rampin, 2000, Giuliano a kol. , 2001). Tyto synaptické kontakty byly demonstrovány značením spinálních neuronů pocházejících z penisu a dosažením míchy se specifickými retrográdními značkami injektovanými do kavernózních korpusů, kombinovaných s dvojitými imunofluorescenčními a konfokálními laserovými mikroskopickými studiemi (Tang et al., 1998, Veronneau-Longueville et al., 1999). V souladu s těmito studiemi u anestetizovaných samců potkanů intratekální injekce kumulativních dávek oxytocinu na lumbo-sacral, ale nikoliv na úrovni torako-bederní, způsobily intracavernózní zvýšení tlaku způsobem závislým na dávce. Tyto účinky byly zrušeny blokádou oxytocinergního receptoru d (CH2) 5Tyr (Me) 2-Orn8-vazotocinem a sekcí pánevních nervů (Giuliano a Rampin, 2000, Giuliano a kol., 2001). Tyto výsledky ukazují, že oxytocin, který působí na lumbo-sacral míchu, zvyšuje intracavernózní tlak a naznačuje, že oxytocin uvolněný během fyziologické aktivace paraventrikulárního jádra je silným aktivátorem páteřních pro-erektilních neuronů vyčnívajících do kavernózních korpusů. Je zajímavé, že tyto pro-erektilní spinální neurony, na kterých působí oxytocin, působí pro-erektilní účinek, také dostávají synaptické kontakty ze serotoninergních neuronů pocházejících z jádra paragiantocelulární z retikulární tvorby medulla oblongata (Marson a McKenna, 1992, Tang et al. , 1998). Zničení těchto serotoninergních neuronů u mužských potkanů usnadňuje ejakulaci a penilní reflexy (Marson a McKenna, 1992, Yells a kol., 1992). Vzhledem k tomu, léčiva, která stimulují 5HT2C receptory usnadnit erekci, pokud je podáván intracerebroventrikulárně, ale ne do paraventrikulárním jádře, a léky, které že blok 5HT2C receptorů snižují také dopamin agonistand oxytocin-indukovanou penilní erekce, zatímco antagonisté dopaminu nesnižují 5HT2C agonistou vyvolané erekci ( viz Stancampiano et al., 1994 a odkazy na ně), bylo také navrženo, aby oxytocin usnadnil působení proreretilních 5HT2C receptorů na úrovni lumboskarlové míchy (Stancampiano et al., 1994). Alternativně může oxytocin ovlivňovat aktivitu klesajících serotoninergních neuronů působením přímo v nucleus paragigantocellularis, kde pocházejí tyto neurony (viz Stancampiano et al., 1994).
3. Interakce mezi oxytocinem, dopaminem a kyselinou glutamovou v centrální nervové soustavě a erekcí penisu
Jak je připomenuto v části 1, všechny oxytocinergní neurony přítomné v centrálním nervovém systému pocházejí z paraventrikulárního jádra a okolních struktur. Aktivita těchto neuronů je pod kontrolou různých neurotransmiterů a / nebo neuropeptidů. Mezi nejvíce studované na paraventrikulární úrovni patří dopamin, kyselina glutamová, kyselina gama-aminomáselná (GABA), oxid dusnatý, endokanabinoidy, opioidní peptidy, peptidy uvolňující růstový hormon, peptidy příbuzné VGF a samotný oxytocin. Dopamin, kyselina glutamová, peptidy uvolňující růstový hormon, peptidy odvozené z VGF a oxytocin jsou stimulující, např. Tyto sloučeniny a jejich agonisté usnadňují erekci penisu při injektování do paraventrikulárního jádra, zatímco GABA, opioidní peptidy a endokanabinoidy jsou inhibiční, např. nebo jejich agonisté inhibují erekci penisu (viz Meisel a Sachs, 1994, Witt a Insel, 1994, Argiolas a Melis, 1995, 2004, 2005, Giuliano a Rampin, 2000, 2004, McKenna, 2000, Andersson, 2001, Hull a další, 2002).
Několik linek experimentálních důkazů naznačuje, že tyto oxytocinergní neurony a výše uvedené neurotransmitery a neuropeptidy se podílejí na kontrole erektilní funkce a sexuálního chování v různých fyziologických souvislostech. Navíc oxytocin uvolněný v extra-hypothalamických mozkových oblastech, jako je ventrální tegmentální oblast, hippocampus a jeho oblasti, amygdala a mícha, mohou ovlivnit aktivitu těch neuronů, na které působí oxytocinergní synapse. V současné době jsou jediné neurony důležité pro erekci penisu, na kterých narážou oxytocinergní synapsy, s jistotou identifikované buňky tělísek mezolimbických dopaminergních neuronů kaudální ventrální tegmentální oblasti vyčnívající do skořápky nucleus accumbens (Melis et al., 2007 (Succu et al., 2008) a pro-erektilní spinální neurony vyčnívající z lumbo-sacrálního traktu do kavernózních korpusů (viz Giuliano a Rampin, 2000, Giuliano a kol., 2001) (viz také oddíly 2.2 a 2.5) . Ačkoli oxytocinergní synapse a receptory byly také identifikovány ve ventrálním subiculu, amygdale a míše, oblasti, které jsou všechny důležité pro erekci penisu (viz výše), v těchto oblastech je typ neurotransmiterů přítomných v neuronech na které oxytocinergní nervové zakončení zasahují, jsou dosud neznámé.
Tato část přehledu stručně shrnuje nedávnou literaturu o mechanismech, které jsou základem pro-erektilního účinku oxytocinu injikovaného do kaudální ventrální tegmentální oblasti, ventrální subiculum hipokampu a míchy. Zvláštní pozornost je věnována interakci peptidu s dopaminem a kyselinou glutamovou v těchto oblastech a úloze, kterou tato interakce může hrát v centrální kontrole erektilní funkce. Stručný přehled účinků dopaminu a kyseliny glutamové na oxytocinergní neurony v paraventrikulárním jádru, který také hraje klíčovou roli v erektilní funkci, je poskytován jako první, aby čtenář věděl o raném stavu výzkumu v této oblasti, tyto studie již byly rozsáhle přezkoumány (viz Argiolas a Melis, 1995, 2004, 2005, Melis a Argiolas, 2003). Také v tomto případě je věnována zvláštní pozornost nejnovějším výsledkům, které naznačují důležitou roli jak pro dopamin-oxytocinové spojení, tak pro spojení glutamové kyseliny s oxytocinem nejen v sexuální výkonnosti (erekci penisu a kopulace), ale také v sexuálním vzrušení a sexuální motivace.
3.1. Interakce dopaminu a oxytocinu v paraventrikulárním jádru
Projekt schopnost agonistů dopaminu vyvolat erekci penisu aktivací centrálních oxytocinergních neuronů (Melis et al., 1987), kdy byl nalezen intracerebroventrikulární (icv) injekční antagonista oxytocinového receptoru d (CH2) 5Tyr (Me) 2-Orn8-vazotocin schopný snížit téměř úplně erekci penisu indukovanou nejen oxytocinem podávaným icv, ale také apomorfinem podávaným subkutánně (Argiolas et al., 1987b). Tyto výsledky byly následovány dalšími studiemi, které vykazovaly podobné výsledky, když byl podán icv (CH2) 5Tyr (Me) 2-Orn8-vazotocin a apomorfin byl podán přímo do paraventrikulárního jádra (Melis et al., 1989b)že agonisté dopaminu indukují erekci penisu aktivaci paraventrikulárních oxytocinergních neuronů, které vyčnívají do extra-hypothalamických oblastí mozku a zejména do míchy (viz Argiolas a Melis, 1995, 2004, 2005). V souladu s touto hypotézou bylo u anestetizovaných krys blokáda lumbo-sacrálních oxytocinergních receptorů nepeptidovým antagonistou oxytocinového receptoru nedávno zjištěna schopná zrušit zvýšení indukované apomorfinem v intrakavernním tlaku indukovaném agonistou dopaminu apomorfinem, paraventrikulo-spinální oxytocinergní dráha zapojená do erekce penisu (Baskerville a kol., 2009).
Studie zaměřené na identifikace dopaminového receptoru zodpovědného za indukci erekce penisu ukázala, že také agonisté paraventrikulárního jádra dopaminu vyvolávají erekci penisu působením na dopaminové receptory rodiny D2, jak se vyskytuje s agonisty dopaminových receptorů podávanými systémově (viz Melis a kol., 1987, Eaton a kol., 1991, Melis a Argiolas, 1995a). Apomorfin, silný smíšený agonista receptoru D1 / D2 a chinpirole, účinný selektivní agonista receptoru D2, ale nikoliv SKF 38393, selektivní agonista receptoru D1, injikovaný do tohoto hypotalamického jádra, se tedy podařilo vyvolat erekci penisu závislým na dávce , a sexuální odpověď vyvolaná těmito agonisty receptoru D2 byla zrušena antagonisty receptoru D2, jako je haloperidol a l-sulpirid, ale nikoliv SCH 23390, selektivní antagonista receptoru D1 (Melis et al., 1987). Schopnost apomorfinu indukovat erekci penisu při injekčním podání do paraventrikulárního jádra byla také potvrzena telemetrickými studiemi, které ukazují, že agonista dopaminu podaný do paraventrikulárního jádra je schopen zvýšit intrakavernózní tlak u vzhůru samců potkanů bez modifikace systémového krevního tlaku (Chen et al. , 1999, Giuliano a Allard, 2001), které byly nalezeny po systémové injekci (Bernabè et al., 1999). Tyto studie také potvrdila hlavní roli receptorů D2, neboť agonisté receptoru D1 byli obvykle zjištěni, že nebyli schopni zvýšit intracavernózní tlak po injekci do paraventrikulárního jádra (Chen a kol., 1999).
Bylo tedy k dispozici několik linek experimentálních důkazů, které naznačují, že paraventrikulární receptory D2, jejichž stimulace vyvolává erekci penisu, jsou lokalizovány na buněčných tělech oxytocinergních neuronů. Nejprve paraventrikulární jádro obsahuje dopaminergní nervové terminály, které patří k takzvaným incertohypothalamickým dopaminergním neuronům. Buněčná tělíska těchto neuronů jsou umístěna ve skupině A13 a A14 Dahlstrom a Fuxe (1964), rozsáhle působí a inervují několik hypotalamických struktur, včetně paraventrikulárních oxytocinergních neuronů, které vyčnívají do neurohypofýzy a / nebo do oblastí extra-hypothalamického mozku (Buijs et al., 1984, Lindvall a kol., 1984).
Zapojení těchto dopaminergních neuronů na paraventrikulární úrovni při kontrole erekce a kopulace penisu je podpořeno studiemi mikrodialyzy, které ukazují, že koncentrace extracelulárního dopaminu a 3,4-dihydroxyfenyloctové kyseliny (DOPAC), jeho hlavního metabolitu, jsou zvýšeny v dialyzátu získaném z paraventrikulární jádro sexuálně silných samců potkanů, které vykazují nekontaktní erekci, když jsou v přítomnosti nepřístupné ovariektomizované samice s primární reakcí na estrogen + progesteron (Melis et al., 2003).
Zvýšení koncentrací dopaminu a DOPAC bylo ještě vyšší, když byla povolena kopulace s receptivní samičkou (Melis et al., 2003), jak bylo zjištěno v mediální preoptické oblasti (Hull et al., 1995) a v nucleus accumbens (Pfaus a Everitt, 1995). Za druhé, několik studií ukazuje, že erekce penisu indukovaná stimulací paraventrikulárních receptorů D2 je zprostředkována oxytocinem uvolněným v těchto oblastech. Proto bylo zjištěno, že apomorfin podávaný v dávkách, které indukují erekci penisu, může zvýšit koncentraci oxytocinu nejen v plazmě krys a opic (Melis et al., 1989a, Cameron a kol., 1992), ale také v extra-hypothalamickém mozku oblasti, jako je hippocampus (Melis a kol., 1990). V souladu s těmito výsledky bylo nedávno prokázáno, že apomorfin vstřikovaný do paraventrikulárního jádra v dávce, která indukuje erekci penisu, je schopen zvýšit i koncentraci extracelulárního dopaminu v nucleus accumbens, což je účinek redukovaný antagonistou oxytocinového receptoru d (CH2) 5Tyr (Me) 2-Orn8-vazotocin vstřikovaný do ventrální tegmentální oblasti (Succu et al., 2007, Melis a kol., 2009a) (viz také oddíl 4). Zatřetí, bilaterální elektrolytické léze paraventrikulárního jádra, které téměř úplně eliminují oxytocin z extra-hypothalamických oblastí mozku (Hawthorn et al., 1985), ruší erekci penisu indukovanou apomorfinem (Argiolas et al., 1987a) a selektivními antagonisty receptoru oxytocinu které se podávají do laterálních komor, ale ne do paraventrikulárního jádra, redukují erekci penisu na dávce závislou apomorfinem s účinností paralelní s účinností těchto sloučenin při blokování oxytocinových receptorů (Melis et al., 1989b). Antagonisté oxytocinového receptoru jsou také extrémně účinné při snižování usnadnění sexuálního chování u mužů indukovaného nejen oxytocinem, ale také apomorfinem (Argiolas et al., 1988, 1989).
Co se týče mechanismu, kterým D2 receptory aktivované dopaminem nebo agonisty receptoru dopaminu, zvyšují aktivitu oxytocinergních neuronů, čímž uvolňují oxytocin v oblastech mozku extrahypothalamu a v míchu, četné experimentální údaježe stimulace receptorů D2 zvyšuje koncentraci intracelulárních iontů Ca2 + uvnitř těl buněk oxytocinergních neuronů, což vede k aktivaci syntázy oxidu dusnatého, enzym závislý na kalmodulinu Ca2 +, který je přítomen v těchto buněčných tělech (Vincent a Kimura, 1992, Torres a kol., 1993, Sanchez a kol., 1994, Sato-Suzuki a kol., 1998). Zvýšená produkce oxidu dusnatého způsobuje aktivaci oxytocinergních neuronů. V souladu s tím byla zabráněná erekcí penisu (1) vyvolanou apomorfinem prostřednictvím blokátorů organických kalciových kanálů a pomocí konotoxinu GVIA silný a selektivní blokátor kanálů Ca2 + závislých na napětí N-typu (McCleskey et al., 1987), který byl podán do paraventrikulární jádra (viz Argiolas a kol., 1990 a odkazy v něm); (2) apomorfinem indukovaná erekce penisu byla zabráněna inhibitory oxidu dusnatého-syntázy podané do paraventrikulárního jádra (Melis et al., 1994c); a (3) apomorfinu a dalších agonistů receptoru D2 podávaných v dávkách, které indukují erekci penisu, zvýšily produkci oxidu dusnatého v paraventrikulárním dialyzátu získaném intramolekulární mikrodialýzou, což bylo snížení, které bylo sníženo inhibitory paraventrikulární syntázy oxidu dusnatého podávaných v dávkách, D2 receptor agonista indukoval erekci penisu (Melis a kol., 1996). Mechanismus, pomocí něhož oxid dusnatý aktivuje paraventrikulární oxytocinergní neurony, je dosud neznámý, i když dostupné údaje naznačují, že oxid dusnatý působí jako intracelulární posel a že guanylátová cykláza není zapotřebí. Účinný fosfodiesterázový rezistentní analog cyklického GMP, 8-bromocyklického GMP, skutečně nebyl schopen vyvolat erekci penisu při podání do paraventrikulárního jádra (viz obrázek 2) (viz Melis a Argiolas, 1995b a odkazy v něm).
Výše uvedená interpretace byla často považována za nepřesvědčivou, hlavně proto, že stimulace receptorů dopaminu D2 je zpravidla spjata s inhibicí spíše než excitací buněčných tělísek neuronů obsahujících tyto receptory prostřednictvím různých mechanismů spojených s G proteinem (viz Sokoloff a Schwartz, 1995). Možné vysvětlení tohoto nesrovnalosti, které je v souladu s přímou stimulací paraventrikulárních oxytocinergních neuronů dopaminem, bylo nedávno navrženo objevem Dopamin D4 spojený s G proteinem (D2, D2 a D3), jejichž stimulace zvyšuje influx Ca4 + v buněčných preparátech obsahujících klonovanou verzi tohoto receptorového podtypu (Moreland et al., 2). Důležitější bylo zjištěno, že selektivní agonista receptoru D2004 (např. ABT 4) (N-methyl-724- (4-kyanofenyl) piperazinyl-2methylbenzamid maleát) je schopen indukovat erekci penisu u samců potkanů při systémových podávání (Brioni et al. 3). Tento účinek nebyl nalezen u selektivního agonistu PNU-2004E (R-2-dihydro-N, N-dimethyl-95666Himidazo [5,6-i] chinolin-4-amin) D4,5,1 receptoru (Hsieh a kol. byl také schopen zvýšit influx Ca5 + v buněčných preparátech obsahujících klonovanou verzi podtypu receptoru D2004 (Brioni et al., 2, Moreland a kol., 4). V souladu s výše uvedenými hypotézami a nálezy byl použit PD 2004 (N-methyl-2004- (168,077-kyanofenyl) piperazinyl-4methylbenzamid maleát), PIP-2EA (3- [3- (2-methoxyfenyl) piperazin-4ylmethylimidazo [2-a] pyridin) a další selektivní agonisté receptoru D1 (Heier et al., 1,2, Melis a další, 4b, Löber a kol., 1997). paraventrikulární jádro, i když méně účinně než apomorfin. Pro-erektilní účinek těchto agonistů receptoru D2006 byl zabráněn přípravkem L-2009 (4- (745,870- [chlorfenyl] piperazin-3-yl) -methyl-4H-pyrrolo [1-B] pyridin trihydrochlorid) (Patel a kol., 1, Melis a kol., 2,3, 4b, Löber a kol., 1997).
Konečně, pro-erektilní účinek výše uvedených agonistů receptoru D4 byl také snížen o inhibitory oxidu dusnatého-syntázy podané do paraventrikulárního jádra a d (CH2) 5Tyr (Me) 2-Orn8-vazotocin, selektivní antagonista receptoru oxytocinu daná ICV, ale ne v paraventrikulárním jádru. Tyto výsledky jsou v souladu s hypotézou, že agonisté receptoru D4 také stimulují oxytocinergní neurony aktivací syntázy oxidu dusnatého a uvolňováním oxytocinu v oblastech extrahypothalamického mozku, což zase usnadňuje erekci penisu, jak je ukázáno u apomorfinu a klasických agonistů D2 (Melis et al. , 2005, 2006b, Löber a kol., 2009).
Výše uvedené poznatky také podporují hypotézu, že dopamin vyvolává erekci penisu působením na D4 receptory lokalizované na buněčných tělech paraventrikulárních oxytocinergních neuronů, které způsobují zvýšený influx Ca2 + do buněčných tělísek oxytocinergních neuronů, což vede ke zvýšení produkce oxidu dusnatého. Oxid dusnatý zase aktivuje oxytocinergní neurony k uvolnění oxytocinu v oblastech mozku extra-hypothalamic a míchy, jak již bylo řečeno. V tomto ohledu, je pozoruhodné, že receptory dopaminu byly identifikovány v buněčných tělech oxytocinergních neuronů v paraventrikulárním jádru jen nedávno studiemi s dvojitou imunofluorescencí s vysokými selektivními protilátkami D2, D3 a D4 receptory as oxytocinovými protilátkami. Tyto studie ukázaly expresi všech tří podtypů D2receptoru (D2, D3 a D4), které se společně lokalizovaly v buněčných tělíštích oxytocinergních neuronů v paraventrikulárním jádru (a také v supraoptickém jádru a mediální preoptické oblasti) (Baskerville a Douglas, 2008, Baskerville a kol., 2009).
To poskytuje silnou neuroanatomickou podporu možnosti, že agonisté dopaminu a dopaminového receptoru typu D2 indukují erekci penisu aktivaci přímo oxytocinergních neuronů vyčnívajících do extrahypothalamických oblastí mozku vyvolaných výše, např. míše, ventrální tegmentální oblast, hippocampus a amygdala. Tyto nálezy však neposkytují žádnou pomoc při identifikaci subtypů receptorů D2, jejichž stimulace způsobuje erektilní odezvu. Bohužel nelze získat žádnou pomoc ani ze studií zaměřených na identifikaci oxytocinergních neuronů aktivovaných agonisty dopaminových receptorů v paraventrikulárním jádru. Navzdory různým účinkům na různé subtypy receptoru dopaminu mohou buď agonisté smíšených dopaminových receptorů (např. Apomorfin), nebo selektivní agonisté receptoru D2 (např. Quinpirole, který působí na všechny podtypy receptoru D2) nebo selektivní agonisty receptoru D4 aktivace oxytocinergních neuronů, měřená zvýšením FOS proteinu v parvocelulárních oxytocinergních neuronech paraventrikulárního jádra (Bitner et al., 2006). Toto zjištění však bylo nedávno zpochybněno, protože zvýšení FOS proteinu v paraventrikulárních oxytocinergních neuronech bylo zjištěno pouze tehdy, když byla erekce penisu indukována quinerolanem, který působí hlavně na subtypy receptorů D2 a D3, ale nikoli na PD 168077, agonistu receptoru D4, a to navzdory schopnosti obou sloučenin indukovat sexuální odpověď (Baskerville et al., 2009).
Další experimenty s selektivními agonisty ostatních podtypů D2 receptorů (zejména D2 a D3) jsou pak nutné k identifikaci přesné role každého podtypu dopaminového receptoru při kontrole erektilní funkce na paraventrikulární úrovni. V tomto ohledu, jak již bylo uvedeno výše, apomorfin, který účinně působí na všechny podtypy dopaminových receptorů (viz Brioni et al., 2004 a odkazy v něm), je mnohem účinnější než agonisté D4 receptorů při vyvolání erekce penisu při injekčním podání do paraventrikulární jádro. To může být vysvětleno vyšší afinitou apomorfinu k D4 receptorům ve srovnání s agonisty testovaných D4 receptorů nebo alternativně agonisté receptoru D4 mohou působit jako částečné agonisty receptoru D4 nebo souběžná aktivace různých subtypů receptoru dopaminu apomorfinem může produkují vyšší aktivaci oxytocinergních neuronů zprostředkujících erekci penisu než aktivace agonisty receptoru D2 pouze podtypu receptoru D4.
Interakce mezi receptory dopaminu D1 a D2 byly již popsány v kontrole sexuálního chování na úrovni mediální preoptické oblasti (viz Hull et al., 1989). V případě, že bude potvrzena neschopnost selektivních agonistů receptoru D2 indukovat erekci penisu (Hsieh et al., 2004) (viz Depoortère et al., 2009), například i po injekci těchto sloučenin do paraventrikulárního jádra , je třeba detailně analyzovat hlavní roli receptorů D3 samotných nebo společně s receptory D4 při aktivaci oxytocinergních neuronů zprostředkujících erekci penisu (viz Baskerville et al., 2009). Bohužel, selektivní agonisté receptoru D2 a D3 (např., Kteří se liší v jejich afinitě k těmto dvěma receptorovým podtypům nejméně čtyři / pět řádů in vitro), nejsou v současné době k dispozici. Z tohoto důvodu nedávný návrh, že receptory D3 zprostředkovávají erekci penisu indukovanou klasickými agonisty receptoru D2, který je založen hlavně na schopnosti předpokládaných antagonistů receptoru D3 charakterizovaných in vitro experimenty, snižuje erekci penisu indukovanou klasickými agonisty D2, jako je apomorfin, quinpirole a pramipexol, které účinně aktivují všechny podtypy receptorů dopaminového D2 (Collins a kol., 2009), musí být jistě validovány jinými experimenty. Tato validace je nutná také proto, že v této studii nebyl nalezen žádný účinek agonistů receptoru D4 na erekci penisu, což je v kontrastu s výsledky studií uvedených výše, které prokazují proreretilní účinek agonistů receptoru D4. Dokonce ani schopnost apomorfinu indukovat erekci penisu (a zívání) u myší s vyřazenými receptory D4 se stejnou účinností jako u myší s vyřazeným počtem D4 receptorů divokého typu a schopností antagonistů receptoru D3 zrušit apomorfinovou odpověď u těchto zvířat ( Collins a kol., 2009) nelze považovat za definitivní důkaz selektivní role podtypu D3 receptoru v erekci penisu indukované agonistou D2. Rozdíly v rozdílech mezi jednotlivými druhy, studie s ablací genu neurotransmiteru / neuropeptidu a / nebo neurotransmiteru / neuropeptidového receptorového receptoru (neurotransmiter / neuropeptid a / nebo neurotransmiter / neuropeptid receptor vyřazené zvěře) obvykle přidaly další zmatek a komplikace při potvrzení předpokládané sexuální role neurotransmiterů a nebo neuropeptidy a jejich receptory. Přesněji řečeno, ablace oxytocinového genu produkuje oxytocin knock out myši, které se navzájem slučují a kopulují, jako by oxytocin nebyl potřebný pro páření a kopulaci. Také homozygotní samice vyřazené oxytocinové myši vykazují normální páření a porod, i když s výrazným narušením úniku mléka (Nishimori et al., 1996, Young a kol., 1996). Ablace genu kódujícího neuronovou syntázu oxidu dusnatého také produkuje myší, které se normálně splétají a kopulují (Huang et al., 1993). Tyto nálezy však pravděpodobně znamenají důležitý rys reprodukční fyziologie, tj. Redundancí systémů zapojených do její kontroly na centrální a periferní úrovni.
Taková redundance má jistě evoluční původ, neboť zaručuje přechod genů na novou generaci pro přežití tohoto druhu. Proto skutečnost, že ablace genu receptoru D4 nemění proerektilní účinek apomorfinu, naznačuje, že receptory D4, jako je oxytocin a oxid dusnatý, jsou jen několika z mediátorů pracujících v systémech, které kontrolují erektilní funkci, spíše než což naznačuje, že tyto receptory nemají žádnou úlohu při kontrole erekce penisu a sexuálního chování. Selhání agonistů D4 k indukci erekce penisu, pokud je systémově podáváno samicím potkanům různých kmenů, byla nedávno hlášena jinou studií (Depoortère et al., 2009). Nicméně, na rozdíl od práce Collins et al. (2009), a aby obraz o roli různých podtypů receptoru D2 v kontrole erekce penisu ještě více zmátl, tato studie také ukazuje, že předpokládané selektivní D3 receptorové antagonisté podávané systémově nebyly schopné snížit apomorfinem indukovanou erekci penisu samčí krysy kmene citlivější na pro-erektilní účinek apomorfinu, zatímco sexuální odpověď (a zívání) byla antagonizována selektivní antagonistou D2 L-741,626 (3 - [[4- (4-chlorfenyl) -4-hydroxypiperidin -1-yl] methyl-1H-indol), což vede k tomu, že D2 receptory spíše než D3 a D4 receptory jsou ty, které hrají hlavní roli v erekci penisu indukované agonistou D2 (Depoortère et al., 2009). Konečně, možnost, že excitační účinek agonistů receptoru dopaminu na oxytocinergní neurony zprostředkující alespoň částečnou erekci penisu je spíše nepřímý než přímý, např. Zprostředkovaný nebo ovlivněný změnami aktivity jiných neuropeptidových neuropeptidů schopných modulovat aktivitu oxytocinergních neuronů v paraventrikulárním jádru nelze úplně vyloučit.
3.2. Interakce glutamové kyseliny s oxytocinem v paraventrikulárním jádru
Paraventrikulární jádro hypotalamu je velmi bohaté na synapse obsahující excitační aminokyselinu jako neurotransmiter (např. Kyselina glutamová a kyselina asparagová) (Van Den Pol, 1991). Excitační aminokyseliny v tomto jádru se účastní řady funkcí, včetně erekce penisu a sexuálního chování (Roeling et al., 1991, Melis a kol., 1994b, 2000, 2004b). V souladu s tím je N-methyl-d-aspartová kyselina (NMDA), selektivní agonista podtypu receptoru NMDA, ale nikoliv kyselina (±) -amino-3-hydroxy-5-methylisoxazol-4- , byl zjištěn selektivní agonista podtypu receptoru AMPA nebo selektivní agonista podtypu metabotropního receptoru (±) -trans (1) -amino-1,3-cyklopentan-dikarboxylová kyselina (ACPD), který je schopen indukovat erekci penisu po injekci do paraventrikulární jádro volně se pohybujících krys (Melis et al., 1994b). Pro-erektilní účinek přípravku 948 MR Melis, A. Argiolas / Neuroscience a Biobehavioral Review 35 (2011) 939-955 NMDA byl zabráněn (+) MK-801, nekompetičním antagonistou NMDA receptoru (Woodruff a kol., 1987 ), injikované do paraventrikulárního jádra (Melis a kol., 1994b). V souladu s těmito výsledky bylo v telemetrických studiích zaměřených na sledování intracavernózního tlaku zjištěno, že NMDA je mnohem aktivnější než agonisté jiných subtypů receptorových excitačních aminokyselin, když jsou injikovány do paraventrikulárního jádra při zvyšujícím se intracavernosálním tlaku u bledých nebo anestetizovaných samců potkanů (Zahran et al. al., 2000, Chen a Chang, 2003).
Jak bylo naznačeno výše pro oxytocin a dopamin, je pravděpodobné, že NMDA receptory zprostředkující erekci penisu jsou lokalizovány v buněčných tělech oxytocinergních neuronů, protože excitační aminokyselinové nervové zakončení zasahují do oxytocinergních buněčných tělísek v paraventrikulárním jádru (Van Den Pol, 1991). Analogicky k tomu, co se objevuje u agonistů dopaminových receptorů, je proreretilní účinek NMDA zřejmě zprostředkován aktivací oxytocinergní neurotransmise, která je zrušena oxytocinovým antagonistou d (CH2) 5Tyr (Me) 2-Orn8-vazotocinem daným icv, ale ne do paraventrikulárního jádra (viz Argiolas a Melis, 1995, 2004, 2005 a odkazy v nich). Stejně tak je aktivace oxytocinergní neurotransmience indukovaná NMDA také sekundární pro aktivaci syntázy oxidu dusnatého, protože NMDA-indukovanou erekci penisu brání inhibitory oxidu dusnatého syntázy (N-nitro-N-methyl-arginin methylester a N- methyl-thio-l-citrulin) podávaného do paraventrikulárního jádra a NMDA injekčně do paraventrikulárního jádra v dávkách, které indukují erekci penisu, zvyšuje produkci oxidu dusnatého v hypotalamickém jádru (viz Argiolas a Melis, 1995, 2004, 2005 a tam uvedené odkazy ). Pokud jde o agonisty dopaminových receptorů, může být NMDA indukovaná aktivace oxidu-syntázy dusičnanů také sekundární vzhledem ke zvýšení influxu Ca2 + v oxytocinergních buněčných tělech prostřednictvím NMDA receptorů sdružených s Ca2 + kanálem, jak je ukázáno v několika nervových přípravcích (viz Snyder, 1992; Southam a Garthwaite, 1993, Schuman a Madison, 1994 a odkazy v nich). Oxid dusnatý zase aktivuje oxytocinergní neurotransmisi (viz výše). Vznik glutamatergických projekcí, které aktivují paraventrikulární oxytocinergní neurony zprostředkující erekci penisu, není znám, ačkoli některé neuroanatomické a elektrofyziologické důkazy naznačují, že mohou pocházet alespoň částečně v hippocampu (Saphier a Feldman, 1987, Chen a kol., 1992) . Ačkoli je zapotřebí další práce pro lepší charakterizaci vzniku glutamatergických projekcí paraventrikulárního jádra, zapojení kyseliny glutamové do paraventrikulárního jádra při kontrole erekce penisu a sexuálního chování je jasně podporováno studiemi mikrodialyzy. Proto byly extracelulární koncentrace kyseliny glutamové a kyseliny asparagové zvýšeny v dialyzátu získaném z paraventrikulárního jádra samců potkanů vykazujících bezkontaktní erekce, když byly v přítomnosti nepřístupných samic potkanů připravených estrogenem a progesteronem primárně (Melis et al. 2004b), erekce penisu, které jsou také zprostředkovány aktivací centrálního oxytocinergního přenosu (Melis et al., 1999a, b). Takové zvýšení bylo zjištěno ještě vyšší, když byla povolena kopulace s receptivní ženou (Melis et al., 2004a). V souladu s hypotézou, že se zvýšená aktivita excitačních aminokyselin vyskytuje v paraventrikulárním jádru během erekce a kopulace penisu, dochází k redukci blokády NMDA receptorů v paraventrikulárním jádru jak u nekontaktní erekce, tak u kopulačního chování (během kterého dochází při erekci penisu kopule) , a toto snížení je následováno snížením zvýšení produkce oxidu dusnatého, ke kterému dochází v tomto hypothalamickém jádru v těchto fyziologických souvislostech (Melis et al., 2000). Zvýšení extracelulární koncentrace kyseliny glutamové sekundární ke snížení uvolňování GABA z konců GABAergních nervů ovlivňujících excitační aminokyselinové synapse vedle těla oxytocinergních buněk se také objevilo v paraventrikulárním jádru po blokaci kanabinoidních receptorů CB1 antagonistou CB1 SR 141761A, (viz Succu et al., 2006, Castelli et al., 2007), které byly podávány do laterálních komor nebo přímo do paraventrikulárního jádra v dávkách, které indukují erekci penisu. Takové zvýšení vedlo k aktivaci syntázy oxidu dusnatého v buněčných tělíštích oxytocinergních neuronů, což vedlo ke zvýšení produkce oxidu dusnatého. Oxid dusnatý zase aktivuje oxytocinergní neurony zprostředkovávající erekci penisu, jak je popsáno výše. V souladu s takovým mechanismem byla indukce erekce penisu SR 141761A snížena blokádou receptorů NMDA a inhibitorů syntázy oxidu dusnatého, ale nikoliv blokádou dopaminových nebo oxytocinových receptorů v paraventrikulárním jádru, zatímco to bylo zabráněno blokádou centrální oxytocinové receptory antagonisty oxytocinového receptoru, kterým je podáván icv
3.3. Oxytocin-dopaminová interakce ve ventrální tegmentální oblasti
Oxytocin indukuje erekci penisu, když je injektován do kaudální části ventrální tegmentální oblasti způsobem závislým na dávce (Melis a kol., 2007). Nejnižší aktivní dávka, která byla injekčně podána jednostranně, byla 20 ng, zatímco nejvyšší testovaná dávka byla 100 ng. Oxytocinový účinek je zprostředkován aktivací oxytocinergních receptorů, protože sexuální odpověď je zrušena předchozí injekcí oxytocinového antagonisty d (CH2) 5Tyr (Me) 2-Orn8-vazotocinu v kaudální ventrální tegmentální oblasti. Tyto receptory jsou lokalizovány v buněčných tělesech dopaminergních neuronů, které se projevují hlavně na skořápce nucleus accumbens. Studie s dvojitou imunofluorescencí (1) ukazují, že v ocasní ventrální tegmentální oblasti jsou oxytocinergní vlákna v těsném kontaktu s buněčnými tělísky dopaminergních neuronů, z nichž většina byla pozitivně značena pro tyrosin-hydroxylázu a obsahovala retrográdní značku Fluorogold (Melis et al., 2007) a (2) ventrální tektonální oblast vyvolaná oxytocinem vyvolaná erekce penisu dochází současně se zvýšením koncentrace extracelulárního dopaminu v dialyzátu získaném z pláště jádro akumbens (Melis a kol., 2007). Oxytocinem indukovaná erekce penisu se vyskytuje současně se zvýšením produkce oxidu dusnatého ve ventrální tegmentální oblasti, přičemž obě reakce jsou antagonizovány nejen d (CH2) 5Tyr (Me) 2-Orn8-vazotocinem a inhibitorem syntázy oxidu dusnatého S- methyl-thio-1-citrulinu, ale také _-konotoxinem, napěťově závislým blokátorem kanálků Ca2 + a ODQ (1H- [1,2,4] oxadiazol [4,3-a] chinoxalin-1- cyklázy, které jsou uvedeny v kaudální ventrální tegmentální oblasti před oxytocinem (Succu et al., 2008). Jelikož mnoho z dopaminergních buněk označených Fluorogoldem, které jsou v kontaktu s oxytocinergními vlákny, které byly pozitivní pro tyrosinhydroxylázu v kaluální ventrální tegmentální oblasti, byly také pozitivně označeny pro syntázu oxidu dusnatého a guanylátcyklázu (Succu et al., 2008), oxytocin vyvolaná erekce penisu může být zprostředkována následujícím mechanismem. Aktivace oxytocinergních receptorů v dopaminergních buňkách těmito peptidy zvyšuje influx Ca2 + uvnitř buněčných tělísek dopaminergních neuronů. Tím se aktivuje syntáza oxidu dusnatého, enzym závislý na kalmodulinu Ca2 +, čímž se zvyšuje tvorba oxidu dusnatého. Oxid dusnatý aktivuje guanylátcyklázu, což vede ke zvýšené koncentraci cyklických GMP. Cyklická GMP aktivuje dopaminergní neurony, které vyčnívají do nucleus accumbens. Úloha cyklického GMP v erekci penisu indukované oxytocinem vstřikovaným do kaudální ventrální tegmentální oblasti je také podporována schopností 8-bromocyklického GMP, aktivního analogu cyklického GMP rezistentního na fosfodiesterázu, vyvolat erekci penisu po injekci do (Cauca et al., 2008, Melis a další, 2009a). To je v rozporu s mechanismem, kterým oxid dusnatý aktivuje oxytocinergní neurony v paraventrikulárním jádru, neboť 8-bromocyklický GMP není schopen vyvolat erekci penisu při injekci do tohoto jádra (Melis a Argiolas, 1995b) (obr. 2). Pokud jde o nervové dráhy aktivované dopaminem v nucleus accumbens vedoucím k erekci penisu, jsou tyto dosud neznámé. Dostupná data však naznačují, že tyto cesty aktivují dopaminovou neurotransmisi v paraventrikulárním jádře hypotalamu. Vzhledem k tomu, erekce penicilie indukovaná oxytocinem probíhá souběžně se zvýšením extracelulárního dopaminu nejen v nucleus accumbens, ale také v paraventrikulárním jádru a je antagonizován antagonistou dopaminového receptoru haloperidolem vstřikovaným do paraventrikulárního jádra (Melis et al., 2007). ATyto výsledky společně podporují myšlenku, že oxytocinergní neurony pocházející z paraventrikulárního jádra a vystupující do kaudální ventrální tegmentální oblasti při aktivovaném uvolňování oxytocinu v této oblasti, čímž aktivují NO-cyklický GMP signalizační systém, který naopak aktivuje mezolimbické dopaminergní neurony (Melis a kol., 2007, 2009a, Succu a kol., 2008). Dopamin uvolněný v nucleus accumbens naopak aktivuje neurální dráhy vedoucí k aktivaci inert-hypothalamických dopaminergních neuronů, které stimulují paraventrikulární oxytocinergní neurony, které vyčnívají do míchy zprostředkované erekci penisu. Současně dopamin uvolněný v paraventrikulárním jádru může také aktivovat oxytocinergní neurony, které vyčnívají do extra-hypothalamických oblastí mozku, jako je ventrální tegmentální oblast, hipokampus, amygdala a možná další oblasti mozku.
Jak bylo připomenuto výše, v souladu s touto hypotézou apomorfin vstřikovaný do paraventrikulárního jádra v dávce, která indukuje erekci penisu, také zvyšuje koncentraci extracelulárního dopaminu v nucleus accumbens, což je účinek redukovaný antagonistou oxytocinového receptoru d (CH2) 5Tyr (Me ) 2-Orn8-vazotocin vstřikovaný do ventrální tegmentální oblasti (Succu a kol., 2007, Melis a kol., 2009a). Společně by výše uvedené neurální dráhy mohly představovat hypotetický neuronový obvod zahrnující dopamin, oxytocin a jiné neurotransmitery (např. Kyselinu glutamovou, viz níže), které ovlivňují nejen sexuální výkon, ale i sexuální motivaci a sexuální odměnu (viz část 4).
3.4. Interakce kyseliny oxytocin-glutamové ve ventrálním subiculu hipokampu
Oxytocin indukuje erekci penisu při injektování do ventrálního, ale nikoliv do zadního subiculu, v závislosti na dávce (Melis et al., 2009b). Sexuální odpověď je zprostředkována stimulací oxytocinových receptorů a je zrušena předchozí injekcí d (CH2) 5Tyr (Me) 2-Orn8-vazotocinu podávaného do stejného místa oxytocinu, které se nachází v jiných oblastech mozku (viz výše) . Pokud jde o lokalizaci těchto receptorů, dostupné údaje naznačují, že jsou lokalizovány v buněčných tělíštích neuronů bohatých na syntézu oxidu dusnatého.
Mikrodialyzační studie tedy ukazují, že erekce penisu indukovaná oxytocinem probíhá souběžně se zvýšením produkce oxidu dusnatého ve ventrální subcélu a toto zvýšení je zrušeno nejen předchozí injekcí inhibitoru oxysyntázy dusičnanu sodného S-methylthio-1-citrulinu a hemoglobinem zachytávajícím oxid dusnatý, ale také dávkou, která antagonizuje erekci penisu (Melis et al., 2), ale také d (CH5) 2Tyr (Me) 8-Orn2010-vazotocin. Důležitější je, že erekce penisu indukovaná oxytocinem se vyskytuje současně s nárůstem koncentrace extracelulární kyseliny glutamové ve ventrálním sububulu, který je pouze částečně antagonizován nekompetičním antagonistou NMDA receptoru (+) MK-801 podaným do ventrální subiculum (Melis a kol., 2010). Společně tyto výsledky naznačují, že nově vytvořený oxid dusnatý, působící jako intercelulární posel, aktivuje neurotransmisi kyseliny glutamové, která vede k erekci penisu, případně prostřednictvím nervových eferentních projekcí z ventrální subiculum do extra-hipokampálních oblastí mozku. V souladu s touto hypotézou, NMDA, vstřikovaná do ventrální subskulky, indukuje erekci penisu závislým na dávce a tento účinek je úplně antagonizován předcházející injekcí do (+) MK-801, ale ne S-methylthio-1-citrulinem , hemoglobin nebo d (CH2) 5Tyr (Me) 2-Orn8-vazotocin (Melis a kol., 2010). Pokud jde o neuronové eferentní dráhy, které vyčnívají do extrahipokampálních oblastí mozku aktivovaných excitačními aminokyselinami (tj. Kyselinou glutamovou) ve ventrální subiculu, je pravděpodobné, že tyto jsou glutamatergické, stejně jako většina hippocampálních eferentních projekcí. V současné době by mohlo být pouze navrženo, aby tyto projekce modulovaly aktivitu mezolimbických dopaminergních neuronů, které následně modulují aktivitu inert-hypothalamických dopaminergních neuronů v paraventrikulárním jádře, což vede k aktivaci oxytocinergních neuronů zprostředkování erekce penisu, jak již bylo řečeno (viz výše).
V důsledku toho dochází k erekci penisu indukované oxytocinem ventrální subiculum souběžně se zvýšením koncentrace extracelulárního dopaminu v skořápce nucleus accumbens a toto zvýšení, jako je erekce penisu, je zrušeno d (CH2) 5Tyr (Me) 2- Orn8-vazotocin podávaný do ventrálního subiculu před oxytocinem (Melis et al., 2009b). Navíc, jelikož indukovaná ventrikulární subiculum indukovaná oxytocinem je erekce penisu také snížena (+) MK-801 vstřikovaným do ventrální tegmentální oblasti, ale nikoliv do nucleus accumbens (Melis et al., 2009b) a dochází souběžně ke zvýšení extracelulární glutamové kyseliny ve ventrální tegmentální oblasti, ale nikoliv v nucleus accumbens, přičemž oba odpovědi byly zrušeny pomocí d (CH2) 5Tyr (Me) -Orn8-vazotocinu, injikovány do ventrální subiculum před oxytocinem (viz obr. 3) je pravděpodobné, že aktivace mezolimbických dopaminergních neuronů je sekundární vzhledem ke zvýšené glutamatergické neurotransmisi ve ventrální tegmentální oblasti. To naznačuje, že interakce glutamové kyseliny s dopaminem ovlivňující erekci penisu existuje ve ventrální tegmentální oblasti. Další studie jsou potřebné k ověření, zda proreretilelní eferentní glutametgické cesty od subpikula do ventrální tegmentální oblasti jsou přímé nebo nepřímé, tj. Přes prefrontální kůru nebo jiné oblasti mozku (viz Melis et al., 2009b a odkazy v ní). Protože paraventrikulární jádro také přijímá glutamatergické projekce od hipokampu (viz výše a Saphier a Feldman, 1987), a kyselina glutamová aktivuje paraventrikulární oxytocinergní neurony včetně těch, které vyčnívají do ventrální tegmentální oblasti (viz Argiolas a Melis, 2005 a tam uvedené odkazy) a oxytocin ve ventrální tegmentální oblasti vyvolává erekci penisu a zvyšuje aktivitu mezolimbického dopaminergního neuronu(viz výše), je lákavé spekulovat, že paraventrikulární oxytocinergní neurony se mohou také alespoň částečně podílet na aktivaci mezolimbických dopaminergních neuronů oxytocinem vstříknutým do ventrální subskula (viz část 4).
4. Závěrečné poznámky
Zkoumané studie potvrzují a rozšiřují počáteční nálezy, které ukazují, že u samců potkanů hraje oxytocin klíčovou úlohu v centrální kontrole erekce penisu na úrovni paraventrikulárního jádra hypotalamu a míchy. Zejména nejčastějiže oxytocin ovlivňuje erekci penisu také tím, že působí v jiných oblastech mozku, tj. ventrální tegmentální oblast, ventrální subiculum a posteromediální kortikální jádro amygdaly.
Na úrovni paraventrikulárním, nejdůležitější nové zjištění, je možná objev exprese všech dopaminových receptorů z rodiny D2 (D2, D3 a D4) v buněčných těl oxytocinergic neuronů v paraventrikulárním jádře (i v supraoptic a mediální preoptická oblast) (Baskerville a Douglas, 2008, Baskerville a kol., 2009). To poskytuje silnou neuroanatomii podpora hypotézy, že agonisté dopaminu a dopaminového receptoru mohou aktivovat přímo oxytocinergní neurony spojené s erektilní funkcí a projevující se nejenom na míchu, ale také na extra-hypothalamické oblasti mozku. V tomto ohledu je také důležité zjistit, že zvýšení blokády oxytocinergních receptorů v lumbo-sacralové míchě (Baskerville et al., 2009) je sníženo zvýšením intrakavernózního tlaku indukovaného agonistou dopaminového receptoru. Ačkoliv tyto důkazy byly získány u anestetizovaných samců potkanů, zjištění potvrzuje aktivaci paraventrikulo-spinální oxytocinergní klesající dráhy, která se účastní erekce penisu vyvolané agonisty dopaminového receptoru. Hje však třeba ještě zjistit, zda je erekce penisu indukovaná stimulací dopaminových receptorů přítomných v oxytocinergních buněčných tělískách sekundární pro aktivaci specifického subtypu receptoru dopaminu rodiny D2 (D2, D3 nebo D4) nebo pokud tyto podtypy receptorů spolupracují při modulaci erektilní odezvy, případně různými způsoby v závislosti na kontextu, ve kterém dochází k erekci penisu (viz Moreland et al., 2004, Enguehard-Gueiffier a další, 2006, Melis a kol. 2006a, b; Löber a kol., 2009, Collins a kol., 2009, Depoortère a kol., 2009, Baskerville a kol., 2009).
Dalším důležitým novým zjištěním je, že oxytocin indukuje erekci penisu, když je injekčně aplikován nejen do paraventrikulárního jádra nebo do oblasti CA1 hipokampu, ale také do ventrální tegmentální oblasti, ventrální subiculum a bedromediálního kortikálního jádra amygdaly. Tyto oblasti mozku nebyly testovány v dřívějších studiích, které ukazují, že oxytocin zvyšuje spontánní epizody erekce penisu u samců potkanů, i když dostávají jako lumbo-sacral mícha oxytocinergní projekce z paraventrikulárního jádra. Oxytocin byl skutečně nalezen schopný zvýšit spontánní epizody erekce penisu, které se vyskytují u dospělých samců potkanů v nepřítomnosti jakýchkoli sexuálních podnětů, jako jsou ty, které pocházejí z přítomnosti přístupného nebo nepřístupného samice potkana s ovarektomií ovariektomatizovaným (estrogen-progesteronem primárně) nebo manipulace genitálií, když je vstřikována do paraventrikulárního jádra a CA1 pole hipokampu, ale nikoliv v dorzálním subiculu, boční přepážce, kaudátové jádro, mediální preoptická oblast, ventromedické jádro a superoptické jádro (Melis et al. , 1986). Ve všech těchto studiích byla erekce penisu zpravidla počítána, když se penis objevil z penisního pláště pozorovatelem, který nevěděl o dané léčbě přímo během experimentu nebo později sledováním experimentu zaznamenaného na videokazetě s videokamerou. Každá epizoda erekce penisu trvá 0.5-1 min a obvykle je doprovázena pérováním penisu a / nebo flexi kyčle. V těchto krysách se obvykle neprovádí žádný pokus, aby se zjistil účinek sexuální zkušenosti, věku nebo jestliže tyto krysy mohou být rozděleny na nízké nebo vysoké odpovědi na pro-erektilní účinek oxytocinu injikovaného do různých oblastí mozku. To se týká také většiny studií pro-erektilního účinku jiných neuropeptidů a léků, které zvyšují spontánní epizody erekce penisu, včetně dopaminových agonistů, excitačních aminokyselin, ACTH-MSH, hexarelinových a VGF peptidů. Pro-erektilní účinek těchto sloučenin byl opakovaně potvrzen telemetrickými metodami, které určují výskyt erekce penisu zvýšením intracavernózního tlaku, ke kterému dochází spontánně nebo po podání těchto sloučenin různými cestami, tj. Systémově, intracerebroventrikulárně nebo intracerebroventrikulárně přímo do specifických mozkových jader, po implantaci tlakového mikrotransduktoru přímo do kavernózních korpusů (viz Bernabè et al., 1999). Ve ventrální tegmentální oblasti, ventrální subiculum a posteromediální jádro amygdaly také oxytocin indukuje erekci penisu působením na oxytocinergní receptory. To vede k aktivaci mezolimbických dopaminergních neuronů pocházejících z ventrální tegmentální oblasti a vystupující do skořápky nucleus accumbens, měřeno zvýšením koncentrace extracelulárního dopaminu v dialyzátu získaném z pláště jádra accumbens a snížením erektilní odpovědi indukované peptidem injikovaným do těchto extra-hypothalamických oblastí, které byly nalezeny po blokádě dopaminergních receptorů v nucleus accumbens (viz níže). Pokud jde o mechanismy aktivované stimulací oxytocinergních receptorů v těchto oblastech mozku, které vedou k aktivaci mezolimbických dopaminergních neuronů a k erekci penisu, nejlépe vyjasněné jsou ty, které se vyskytují v kaudální ventrální tegmentální oblasti. Výsledky farmakologické a imuno-fluorescence ukazují, že oxytocinové nervové zakončení zasahují do buněčných tělísek dopaminergních neuronů, které vyčnívají do skořápky nucleus accumbens (Melis et al., 2007, 2009a, Succu et al., 2008). Mnohé z těchto neuronů jsou bohaté na syntézu oxidu dusnatého a na guanylátcyklázu. Stimulace oxytocinergních receptorů v buněčných tělesech těchto dopaminergních neuronů způsobuje aktivaci syntázy oxidu dusnatého vedoucí ke zvýšení produkce oxidu dusnatého. Oxid dusnatý aktivuje guanylátcyklázu, čímž zvyšuje koncentraci cyklických GMP, což vede k aktivaci mezolimbických dopaminergních neuronů ak uvolňování dopaminu v nucleus accumbens, měřeno zvýšením extracelulárního dopaminu v dialyzátu z nucleus accumbens získaného intramolekulární mikrodialýzou (Succu et al., 2008 ). Dopamin uvolněný v nucleus accumbens zase aktivuje nervové dráhy vedoucí k erekci penisu. To je podpořeno schopností antagonistů dopaminových receptorů haloperidolu a / nebo cis-flupentixolu vstřikovaných do nucleus accumbens, aby se snížila erekce penilu vyvolaná oxytocinem v oblasti tektonové oblasti (Succu et al., 2008). Jedna z pro-erektilních cest se zdá, že aktivuje incerto-hypothalamické dopaminergní neurony, zejména ty, které projevují buněčné tělíska paraventrikulárních oxytocinergních neuronů. Ve skutečnosti oxytocin vstřikovaný do kaudální ventrální tegmentální oblasti zvyšuje extracelulární dopamin nejen v nucleus accumbens, ale také v paraventrikulárním jádru a blokáda dopaminových receptorů v paraventrikulárním jádru významně snižuje erekci penilu vyvolanou oxytocinem v oblasti tektonické oblasti (Succu et al. , 2007, 2008, Melis a kol., 2007, 2009a). Existence těchto nucleus accumbens dopamin-paraventrikulární dopamin-paraventrikulární oxytocin-ventrální tegmentální oblast oxytocin-dopaminové vazby je také navržena schopností pro-erektilní dávky apomorfinu a agonisty receptoru D4 PD 168077 injikovaného do paraventrikulárního jádra pro zvýšení extracelulárního dopaminu v skořápce nucleus accumbens (Succu et al., 2007), reakce který je zrušen pomocí d (CH2) 5Tyr (Me) -Orn8-vazotocinu podaného do ventrální tegmentální oblasti (Melis et al., 2009a, viz také níže). Nicméně další práce je nutná k identifikaci neurálních cest, které spojují nucleus accumbens s inertním hypotalamovým dopaminergním systémem.
Mechanismus, kterým oxytocin indukuje erekci penisu a aktivuje mezolimbické dopaminergní neurony, když je injekčně podán do ventrální subiculum nebo do posteromediálního jádra amygády, je v současné době jen částečně chápán. V těchto oblastech také oxytocin aktivuje své vlastní receptory, které vedou k aktivaci syntázy oxidu dusnatého, čímž se zvyšuje produkce oxidu dusnatého. Oxid dusnatý zase aktivuje neznámé eferentní projekce, které zřejmě zvyšují glutamatergickou neurotransmisi ve ventrální tegmentální oblasti. Kyselina glutamová pak stimuluje mezolimbické dopaminergní neurony vedoucí k erekci penisu. Tato hypotéza je podpořena hlavně schopností oxytocinu vstřikovaného do ventrálního subiculu pro zvýšení extracelulární kyseliny glutamové ve ventrální tegmentální oblasti (obr. 3) a nekompetitivního NMDA receptorového antagonisty (+) MK-801 vstřikovaného do ventrální tegmentální oblasti, avšak nikoliv v nucleus accumbens, ke snížení erekce penisu indukované oxytocinem vstřikovaným buď do ventrální podkožky, amygdaly (Melis a kol., 2009b). V současné době jsou k dispozici další podrobnosti týkající se erekce penisu vyvolané oxytocinem v ventrálním subiculu. Zde se zdá, že oxytocinem indukovaná erekce penisu je sekundární pro aktivaci oxytocinergních receptorů umístěných v buněčných tělíštích neuronů obsahujících oxid dusnatý-syntázu. To způsobuje zvýšení tvorby oxidu dusnatého, který aktivuje glutamatergickou neurotransmisi tím, že působí jako mezibuněčný posel s mechanismem podobným tomu, který je popsán pro dlouhodobou potenciaci (viz Snyder, 1992, Southam a Garthwaite, 1993, Schuman a Madison, 2004) . V souladu s touto hypotézou vzniká oxytocinem indukovaná erekce penisu souběžně se zvýšením extracelulární kyseliny glutamové v dialyzátu z ventrální subiculum a stimulace excitačních aminokyselinových receptorů ve ventrálním subiculu pomocí NMDA vyvolává erekci penisu. Kyselina glutamová zase aktivuje neurální eferentní projekce, které vedou k aktivaci mezolimbických dopaminergních neuronů ve ventrální tegmentální oblasti, jak bylo uvedeno výše. Pokud jsou tyto mechanismy funkční i v posteromediálním jádře amygdaly, není v současnosti známo. Kromě toho jsou nezbytné další studie, které dokládají, že (1) oxytocinergní nervové zakončení a receptory ve ventrálním subiculu a v posteromediálním jádře amygdaly jsou lokalizovány v buněčných tělesech neuronů obsahujících oxid dusnatý-syntázu (2), jsou-li tyto neurony nakonec pod kontrolou excitačních aminokyselin (glutamatergických) synapsí a (3) k identifikaci signalizačního systému aktivovaného oxidem dusnatým. V tomto ohledu je třeba poznamenat, že schopnost oxytocinu zvyšovat aktivitu oxidu dusnatého syntázy v buňkách těla dopaminergních neuronů v ocasní ventrální tegmentální oblasti a dosud neznámých neuronů v ventrálním sub-jekule a v posteromediálním jádru amygdaly, vyvolává schopnost oxytocinu aktivovat synthózu oxidu dusnatého v buněčných tělech oxytocinergních neuronů v paraventrikulárním jádru (Melis et al., 1997). Nicméně zatímco ve ventrální tegmentální oblasti vede zvýšená produkce oxidu dusnatého v buněčných tělesech dopaminergních neuronů k aktivaci těchto neuronů aktivací guanylátcyklázy a zvyšováním cyklické GMP, nedochází k tomu v paraventrikulárním jádru. Proto 8-bromokylická GMP vstřikovaná do paraventrikulárního jádra neindukuje erekci penisu, a to při injektování do ventrální tegmentální oblasti. Další signální dráhy liší od dusičné systému oxidecyclic GMP je pak zapojen na úrovni paraventrikulárního při aktivaci oxytocinergic neuronů zprostředkujících erekci endogenní a / nebo exogenního oxidu dusnatého (Melis a Argiolas, 1995b;. Melis et al, 1997) (obr . 1). Na druhé straně se zdá, že cyklická GMP ve ventrální tegmentální oblasti hraje také klíčovou roli při aktivaci mezolimbických dopaminergních neuronů a ve zvýšení extracelulárního dopaminu vyskytujícího se v dialyzátu získaném z pláště jádra accumbens samců potkanů vybrané pro ukázání nebo neexistenci erekcí penisu bez kontaktu, když se v přítomnosti nedosažitelné ovariektomizované receptivní (ošetřené estrogenem a progesteronem) potkanem samice. Při těchto experimentálních podmínkách u samců potkanů vykazujících bezkontaktní erekci penisu bylo zjištěno zvýšení očekávané koncentrace extracelulárního dopaminu a toto zvýšení je dále zvýšeno, i když jen skromně, inhibitory fosfodiesterázy podávanými do kaluální ventrální tegmentální oblasti (Sanna et al., 2009).
Schopnost oxytocinu vstřikovaného do ventrální tegmentální oblasti, ve ventrální subiculum av posteromediálním kortikálním jádru amygdaly spolu s agonisty dopaminu injikovanými do paraventrikulárního jádra vyvolat erekci penisu a aktivovat mezolimbické dopaminergní neurony si zaslouží nějaký komentář . Za prvé, mechanismy podobné těm, které jsou uvedeny výše, mohou být účinné, jestliže dochází k erekci penisu ve fyziologickém kontextu, například během kopulace (při výskytu erekce penisu kopula) nebo během erekcí penisu bez kontaktu. Tyto erekce jsou feromonové zprostředkované penilní erekce nerozeznatelné od těch, vyvolané léčivy nebo oxytocinu, které se vyskytují, když jsou sexuálně silnými samci krys do přítomnosti nepřístupném receptivních (vaječníky estrogen + progesteron ošetřený) samice krysy a jsou považovány jako index sexuální vzrušení (Sachs, 1997, 2007). Ačkoli tyto výsledky neprokazují, že oxytocin v těchto oblastech hraje roli při erekci penisu ve fyziologickém kontextu nebo po podání léku, dodávají další sílu k raným nálezům, což naznačuje, že tyto oblasti mozku patří k těm, kde se oxytocin, nejen spontánní epizody erekce penisu pozorované po léčbě pro-erektilních léků, ale také zlepšení sexuálního chování mužů (a žen) (viz Argiolas a Melis, 2004 a odkazy v ní). V souladu s tím se koncentrace oxytocinu zvyšuje u hippocampu samců potkanů léčených pro-erektilní dávkou apomorfinu, klasického agonisty dopaminu (Melis a kol., 1990) a d (CH2) 5Tyr (Me) -Orn8-vazotocinu, který blokuje oxytocin (Argiolas a kol., 1987a), ale také usnadňující účinek apomorfinu na chování mužského kopulace (Argiolas a kol., 1987b) u pohlavně silných samců potkanů, během kterých při erekci penisu kopule nastat. d (CH2) 5Tyr (Me) -Orn8-vasotocin je také velmi účinný při snižování bezkontaktní erekce u sexuálně silné samců krys, při podávání v nanogram množství do postranních komor, ale ne do PVN (Melis a kol., 1999a).
Za druhé, mezolimbické dopaminergní neurony hrají klíčovou roli v motivačních a obohacujících vlastnostech přírodních posilovacích podnětů, jako jsou potraviny, voda a sexuální aktivity (Fibiger a Phillips, 1988, Wise a Rompre, 1989, Everitt, 1990). Konkrétně se předpokládá, že dopamina uvolněná z těchto neuronů zprostředkovává transpozici motivačních aspektů přirozených podnětů do cíleně orientovaného chování, například v případě sexuální aktivity, hledání sexuálního partnera a sexuálního styku k dosažení odměny a spokojenosti (Goto a Grace, 2005). V souladu s tím se koncentrace extracelulárního dopaminu zvyšuje v dialyzátu z nucleus accumbens u sexuálně silných samců potkanů během expozice nepřístupnému ovariectomizovanému samici potkana s estrogenem a progesteronem primovaným a tento vzestup byl ještě vyšší, když samčí potkan byl ponechán kopulovat s receptivní samicí (Pfaus a Everitt, 1995).
Zatřetí, současné výsledky podporují hypotézu, že neurální okruh spojuje paraventrikulární jádro s ventrální tegmentální oblastí přímo nebo nepřímo (přes ventrální subiculum a / nebo posteromediální kortikální jádro amygdaly) a nucleus accumbens a odtud přes neznámé cesty zpět do paraventrikulárního jádra pro kontrolu aktivity oxytocinergních neuronů vyčnívajících do míchy zprostředkujících erekci penisu a oxytocinergních neuronů vyčnívajících do ventrální tegmentální oblasti, ventrální subiculum a bedromediální kortikální jádro amygdaly, čímž se moduluje aktivity mezolimbických dopaminergních neuronů (obr. 4). Tento komplexní neurální okruh může hrát roli při integraci nervových aktivit, které se podílejí na kontrole spotřebitelských (erektilní-ejakulační) a předvídatelných (motivačních a odměňování) aspektů mužského sexuálního chování ve fyziologických souvislostech. Ve skutečnosti se extracelulární dopamin zvyšuje v nucleus accumbens (Pfaus a Everitt, 1995) a v paraventrikulárním jádru sexuálně silných samců potkanů během expozice nepřístupné receptivní samici potkana, když se vyskytují nekontaktní erekce a ještě více, když je povolena kopulace, např. když se v penisu objevují erekce penisu (Melis et al., 2003). Takže i když jsou nezbytné další studie k objasnění úlohy endogenního oxytocinu ve ventrální tegmentální oblasti, ventrální subiculum a amygdala během erekcí bez kontaktu a sexuálního chování, pak lze rozumně předpokládat, že tento hypotetický neurální okruh, a přitom přispívá k (Everitt, 1990, Pfaus a Everitt, 1995), které se mohou projevit jako příznivé pro pohlavní chování, mohou současně aktivovat mezolimbický dopaminergní systém poskytující nervový substrát. V tomto ohledu je třeba poznamenat, že mezolimbický dopaminergní systém aktivovaný oxytocinem vstřikovaným do ventrální tegmentální oblasti je stejný, aktivovaný zneužíváním léků, jako jsou opiáty, kanabinoidy, amfetamin, kokain a alkohol (Tanda a kol., 1997) a že oxytocin byl nalezen schopný snížit toleranci a závislost na kokainu, morfinu, alkoholu a kanabinoidech (Kovacs et al., 1998, Cui a kol., 2001). Na závěr se zdá, že oxytocin uvolněný nejen ve ventrální tegmentální oblasti, ale také ve ventrálním subiculu a posteromediálním kortikálním jádru amygdaly, může aktivovat mezolimbické dopaminergní neurony, které se mohou podílet na apetistických a odměřujících účincích sexuální aktivity . Aktivace mezolimbických dopaminergních neuronů může být přímá přes oxytocinergní receptory v buněčných tělech mezolimbických dopaminergních neuronů nebo nepřímá aktivací neurotransmisí kyseliny glutamové ve ventrální tegmentální oblasti.
Dopamin vydána v nucleus accumbens skořepinové moduluje zase aktivitu incerto-hypotalamické dopaminergických neuronů v paraventrikulárním jádře způsobí buď erekci (aktivací oxytocinergic neuronů vyčnívajících do míchy), nebo sexuální motivaci a odměnu (aktivací oxytocinergic neurony vyčnívající do ventrální tegmentální oblasti, ventrální subiculum nebo posteromediální kortikální jádro amygdaly). Vzhledem k tomu, že dopamin je také uvolňován v jádře jádra a v paraventrikulárním jádru, když dojde k erekci penisu ve fyziologických kontextech (např. Nekontaktní erekce a kopulace) (Succu et al., 2007, Melis et al., 2003, 2007). je pravděpodobné, že centrální oxytocinergní neurony se účastní nervových obvodů zprostředkujících interakci mezi mezolimbickým a incerto-hypothalamovým dopaminergním systémem. Tyto neurální okruhy mohou hrát roli nejen v konzumační fázi sexuálního chování (např. Erekci penisu a kopulace), ale také v sexuální motivaci, sexuálním vzrušení a sexuální odměně.
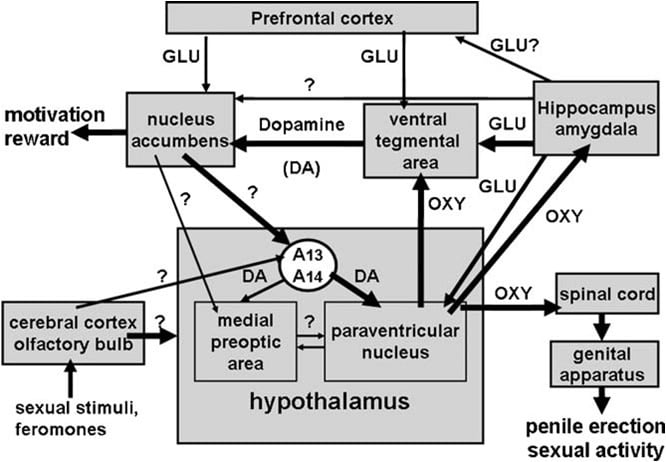
Schematické znázornění hypotetického neuronového obvodu zahrnujícího oxytocin ovlivňující sexuální motivaci, odměňování a sexuální výkonnost, jak to naznačují výsledky této kapitoly a předchozí zprávy. Oxytocinergic neurony pocházející paraventrikulárním jádře a vyčnívající do míchy, když je aktivována například dopaminu a kyselina glutamová (ale také dalších neurotransmiterů a / nebo neuropeptidů), usnadňuje erekci a sexuální výkonnost aktivací oxytocinergic neuronů vyčnívajících do míchy. Dopamin a kyselina glutamová (ale také neurotransmitery a neuropeptidy) v paraventrikulárním jádru také aktivují oxytocinergní neurony, které vyčnívají do ventrální tegmentální oblasti, čímž aktivují mezolimbické dopaminergní neurony, které vyčnívají do nucleus accumbens, modulují sexuální motivaci a odměnu. Dopamin uvolněný v nucleus accumbens (NAs) aktivuje stále ještě neznámé neurální dráhy, které zvyšují aktivitu incerto-hypothalamických dopaminergních neuronů (pocházejících z A13-A14 skupin Dahlstrom a Fuxe), které působí na oxytocinergní neurony, včetně těch, míchy, což vede k erekci penisu. Tento obvod může být také aktivován oxytocinem vstřikovaným nejen do kaudální ventrální tegmentální oblasti, ale také do ventrální subiculum a amygdaly, které také dostávají oxytocinergní inervaci z paraventrikulárního jádra, případně přímým nebo nepřímým glutamatergickým eferentem na ventrální které vedou k modulaci sexuální motivace i erekce penisu. Konečně výše uvedené obvody mohou být také aktivovány sexuálními stimuly a feromony, neboť extracelulární dopamin a kyselina glutamová se zvyšují v paraventrikulárním jádru (a v mediální preoptické oblasti) během feromonem zprostředkované bezkontaktní erekce a kopulace (pro příslušné odkazy viz Seznam odkazů).
Poděkování Tato práce byla částečně podpořena grantem italského ministerstva univerzity a výzkumu AA a MRM
Reference
Andersson, KE, 2001. Farmakologie erekce penisu. Pharmacol. Rev 53, 417-450. Argiolas, A., 1994. Oxid dusnatý je centrálním mediátorem erekce penisu. Neurofarmakologie 33, 1339-1344. Argiolas, A., 1999. Neuropeptidy a sexuální chování. Neurosci. Biobehav. Rev 23, 1127-1142. Argiolas, A., Gessa, GL, 1991. Centrální funkce oxytocinu. Neurosci. Biobehav. Rev 15, 217-231. Argiolas, A., Melis, MR, 1995. Neuromodulace erekce penisu: přehled role neurotransmiterů a neuropeptidů. Prog. Neurobiol. 47, 235-255. Argiolas, A., Melis, MR, 2004. Úloha oxytocinu a paraventrikulárního jádra v sexuálním chování savců. Physiol. Behav. 83, 309-317. Argiolas, A., Melis, MR, 2005. Centrální kontrola erekce penisu: úloha paraventrikulárního jádra hypotalamu. Prog. Neurobiol. 76, 1-21. Argiolas, A., Collu, M., Gessa, GL, Melis, MR, Serra, G., 1988. Oxytocinový antagonista d (CH2) 5Tyr (Me) -Orn8-vazotocin antagonizuje chování samčího kopulace u potkanů. E. J. Pharmacol. 149, 389-392. Argiolas, A., Collu, M., D'Aquila, P., Gessa, GL, Melis, MR, Serra, G., 1989. Apomorfinová stimulace chování kardiovaskulárního onemocnění u potkanů brání antagonista oxytocinu d (CH2) 5Tyr (Me) -Orn8-vazotocin. Pharmacol. Biochem. Behav. 33, 81-83. Argiolas, A., Melis, MR, Gessa, GL, 1985. Intraventrikulární oxytocin vyvolává zívání a erekci penisu u potkanů. E. J. Pharmacol. 117, 395-396. Argiolas, A., Melis, MR, Gessa, GL, 1986. Oxytocin: extrémně silný induktor erekce penisu a zívání u samců potkanů. E. J. Pharmacol. 130, 265-272. Argiolas, A., Melis, MR, Mauri, A., Gessa, GL, 1987a. Paraventrikulární léze jádra brání zivění a erekci penisu indukované apomorfinem a oxytocinem, ale nikoliv ACTH u potkanů. Brain Res. 421, 349-352. Argiolas, A., Melis, MR, Vargiu, L., Mauri, A., Gessa, GL, 1987b. d (CH2) 5Tyr (Me) -Orn8-vazotocin, silný antagonista oxytocinu, antagonizuje erekci penisu a zívání indukovanou oxytocinem a apomorfinem, ale nikoli ACTH 1-24. E. J. Pharmacol. 134, 221-224. Argiolas, A., Melis, MR, Stancampiano, R., Gessa, GL, 1990. Konotoxin zabraňuje vzniku apomorfinu a oxytocinu vyvolané erekci penisu a zívání u samců potkanů. Pharmacol. Biochem. Behav. 37, 253-257. Arletti, R., Bertolini, A., 1985. Oxytocin stimuluje chování lordózy u samic potkanů. Neuropeptidy 6, 247-255. Arletti, R., Bazzani, C., Castelli, M., Bertolini, A., 1985. Oxytocin zvyšuje výkon krysících mužů u potkanů. Horm. Behav. 19, 14-20. Arletti, R., Benelli, A., Bertolini, A., 1990. Sexuální chování starých samců potkanů je stimulováno oxytocinem. E. J. Pharmacol. 179, 377-382. Arletti, R., Calzà, L., Giardino, L., Benelli, A., Cavazzutti, E., Bertolini, A., 1997. Sexuální impotence je spojena se sníženou produkcí oxytocinu a zvýšenou produkcí opioidních peptidů v paraventrikulárním jádře hypotalamu. Neurosci. Lett. 233, 65-68. Bancila, M., Giuliano, F., Rampin, O., Mailly, P., Brisorgueil, MJ, Clas, A., Verge, D., 2002. Důkazy pro přímý projev paraventrikulárního jádra hypotalamu na předpokládané serotoninergní neurony jader paragigantocelulární, které se podílejí na kontrole erekce u potkanů. E. J. Neurosci. 16, 1240-1249. Burnett, AL, Lowenstein, CJ, Bredt, DS, Chang, TSK, Snyder, SH, 1992. Oxid dusnatý: fyziologický mediátor erekce penisu. Věda 257, 401-403. Baskerville, TA, Douglas, AJ, 2008. Interakce mezi dopaminem a oxytocinem při kontrole sexuálního chování. Prog. Brain Res. 170, 277-289. Baskerville, TA, Allard, J., Wayman, C., Douglas, AJ, 2009. Interakce mezi dopaminem a oxytocinem v erekci penisu. E. J. Neurosci. 30, 2151-2164. Benelli, A., Bertolini, A., Poggioli, R., Cavazzutti, E., Calzà, L., Giardino, L., Arletti, R., 1995. Oxid dusnatý se podílí na mužském sexuálním chování potkanů. E. J. Pharmacol. 294, 505-510. Bernabè, J., Rampin, O., Sachs, BD, Giuliano, F., 1999. Intrakavernózní tlak během erekce u potkanů: integrační přístup založený na telemetrickém záznamu. Am. J. Physiol. 276, R441-R449. Bitner, RS, Nikkel, AL, Otte, S., Martino, B., Barlow, EH, Bhatia, P., Stewart, AO, Brioni, JD, Decker, MWN, Moreland, RB, 2006. Signalizace dopaminového D4 receptoru u paraventrikulárního hypotalamického jádra potkanů: důkaz o přirozené vazbě zahrnující okamžitou časnou indukci genu a fosforylaci proteinkinázy aktivované mitogenem. Neurofarmakologie 50, 521-531. , Brian, JD, Moreland, RB, Cowart, M., Hsieh, GC, Stewart, AO, Hedlund, P., Donnelly-Roberts, DL, Nakane M., Lynch 3rd., J. Kolasa, T. Polakowski , JS, Osinski, MA, Marsh, K., Andersson, KE, Sullivan, JP, 2004. Aktivace receptorů dopaminu D4 pomocí ABT-724 vyvolává erekci penisu u potkanů. Proc. Natl. Acad. Sci. Spojené státy americké 101, 6758-6763. Buijs, RM, 1978. Intra- a extra-hypotalamické vazopresinové a oxytocinové cesty u potkanů. Cell Tissue Res. 192, 423-435. Buijs, RM, Geffard, M., Pool, CW, Hoorneman, EMD, 1984. Dopaminergní inervace superoptického a paraventrikulárního jádra. Studium lehké a elektronové mikroskopie. Brain Res. 323, 65-72. Caldwell, JD, Prange, AJ, Pedersen, CA, 1986. Oxytocin usnadňuje sexuální receptivitu samic potkanů ošetřených estrogenem. Neuropeptidy 7, 175-189. Cameron, JL, Pomerantz, SM, Layden, LM, Amico, JA, 1992. Dopaminergní stimulace koncentrací oxytocinu v plazmě samic a samic opic apoporfinem a agonistou receptoru D2. J. Clin. Endocrinol. Metab. 75, 855-860. Canteras, NS, Simerly, RB, Swanson, LW, 1995. Organizace projekcí z mediálního jádra amygdaly: studie PHAL u potkanů. J. Comp. Neurol. 360, 213-245. Carmichael, MS, Humbert, R., Dixen, J., Palmisano, G., Greeleaf, W., Davidson, JM, 1987. Plazmatický oxytocin zvyšuje lidskou sexuální odpověď. J. Clin. Endocrinol. Metab. 64, 27-31. Carter, CS, 1992. Oxytocin a sexuální chování. Neurosci. Biobehav. Rev 16, 131-144. Carter, CS, Lederhendler, II., Kirkpatrick, B., 1997. Interaktivní neurobiologie příbuzenstva, Annals of the New York Academy of Sciences, sv. 807. New York Academy of Sciences, New York. Perez, AP, Melis, T., Succu, S., Sanna, F., Melis, MR, Collu, S., Ennas, MG, Diaz, G., Mackie, K., Argiolas, A. , 2007. Kanabinoidní receptory CB1 v paraventrikulárním jádru a centrální kontrola erekce penisu: imunocytochemie, autoradiografie a behaviorální studie. Neurovědy 147, 197-206. Chen, KK, Chang, LS, 2003. Účinek agonistů receptoru excitační aminokyseliny na erekci penisu po podání do paraventrikulárního jádra hypotalamu u potkanů. J. Urol. 62, 575-580. Chen, KK, Chan, JYH, Chang, LS, Chen, MT, Chang, SHH, 1992. Vyvolání erekce penisu po aktivaci tvorby hippocampu u potkanů. Neurosci. Lett. 141, 218-222. Chen, KK, Chan, JYH, Chang, LS, 1999. Dopaminergní neurotransmise v paraventrikulárním jádře hypotalamu v centrální regulaci erekce penisu u potkanů. J. Urol. 162, 237-242. Collins, GT, Truccone, A., Haji-Abdi, F., Newman, AH, Grundt, P., Rice, KC, Husbands, SM, Greedy, BM, Enguehard-Gueiffer, , J., Wang, S., Katz, JL, Grandy, DK, Sunahara, RK, Woods, JH, 2009. Pro-erektilní účinky dopamin D2 podobných agonistů jsou zprostředkovány receptorem D3 u potkanů a myší. J. Pharmacol. Exp. Ther. 329, 210-217. Coolen, LM, Allard, J., Truitt, WA, McKenna, KE, 2004. Centrální regulace ejakulace. Physiol. Behav. 83, 203-215. Cui, SS, Bowen, RC, Gu, GB, Hannesson, BK, Yu, PH, Zhang, X., 2001. Prevence abstinenčního syndromu kanabinoidů lithiem: zapojení oxytocinergní neuronální aktivace. J. Neurosci. 21, 9867-9876. Dahlstrom, A., Fuxe, K., 1964. Důkazy o existenci neuronů obsahujících monoaminy v centrální nervové soustavě. I. Demonstrace monoaminů v tělech buněk neuronů mozku. Acta Physiol. Scand. 62 (Suppl. 232), 1-54. Depoortère, R., Bardin, L., Rodrigues, M., Abrial, E., Aliaga, M., Newman-Tancredi, A., 2009. Penis erekce a zívání indukované agonisty dopaminových receptorů podobných D2u u potkanů: vliv kmene a přínos dopaminových D2, ale nikoliv D3 a D4 receptorů. Behav. Pharmacol. 20, 303-311. Domes, G., Heinrichs, M., Buchel, C., Braus, DF, Herpertz, SC, 2007. Oxytocin zeslabuje reakce amygdaly na emocionální tváře bez ohledu na valence. Biol. Psychiatrie 62, 11871190. Dominguez, J., Riolo, JV, Xu, Z., Hull, ME, 2001. Regulace mediální amygádou kopulace a mediálního předoptického uvolňování dopaminu. J. Neurosci. 21, 349-355. Donaldson, ZR, Young, LJ, 2009. Oxytocin, vazopresin a neurogenetika společenského života. Věda 322, 900-904. Eaton, RG, Markowski, VF, Lumley, LA, Thompson, JT, Moses, J., Hull, EM, 1991. D2 receptory v paraventrikulárním jádru regulují genitální odpovědi a kopulaci u samců potkanů. Pharmacol. Biochem. Behav. 39, 177-181. Ebner, K., Bosch, OJ, Krömer, SA, Singewald, N., Neumann, ID, 2005. Uvolňování oxytocinu v centrální amygdii potkana moduluje chování při zvládání stresu a uvolňování excitačních aminokyselin. Neuropsychofarmakologie 30, 223-230. Enguehard-Gueiffier, C., Hübner, H., El Hakmaoui, A., Allouchi, H., Gmeiner, P., Argiolas, A., Melis, MR, Gueiffier, A., 2006. 2 - [(4-Fenylpiperazin-XN-MX-yl) methyl] imidazo (di) aziny jako selektivní D1-ligandy. Indukce erekce penisu přípravkem 2- [4- (2-methoxyfenyl) piperazin-1-ylmethyl] imidazo [1,2-a] pyridin (PIP3EA), silný a selektivní agonista D4. J. Med. Chem. 49, 3938-3947. Everitt, BJ, 1990. Sexuální motivace: neurální a behaviorální analýza mechanismů, které jsou základem potíží apetitivních a kopulátorských potkanů. Neurosci. Biobehav. Rev 14, 217-232. Fibiger, HC, Phillips, AG, 1988. Mezokortikolimbický dopaminový systém a odměna. Ann. N. Y. Acad. Sci. 5, 206-215. Francouzština, SJ, Totterdell, S., 2003. Jednotlivé nucleus accumbens-projection neurons dostávají bazolaterální amygdaly a ventrální subjunkční aferenty u potkanů. Neurovědy 119, 19-31. 954 MR Melis, A. Argiolas / Neuroscience a biobehaviorální recenze 35 (2011) 939-955 Freund-Mercier, MJ, Richard, P., 1981. Excitační účinky intraventrikulárních injekcí oxytocinu na reflexe ejekce mléka u potkanů. Neurosci. Lett. 23, 193-198. Freund-Mercier, MJ, Richard, P., 1984. Elektrofyziologické důkazy pro usnadnění kontroly oxytocinových neuronů oxytocinem během kojení u potkanů. J. Physiol. (Lond.) 352, 447-466. Freund-Mercier, MJ, Stoeckel, ME, 1995. Somatodendritické autoreceptory na oxytocinových neuronech. In: Ivell, R., Russel, JA (Eds.), Oxytocin, buněčné a molekulární přístupy v medicíně a výzkumu. Adv. Exp. Med. Biol., 365. Plenum Press, New York a Londýn, str. 185-194. Freund-Mercier, MJ, Stoeckel, ME, Palacios, JM, Pazos, JM, Richard, PH, Porte, A., 1987. Farmakologické charakteristiky a anatomická distribuce vazebných míst 3H oxytocinu v mozku potkana Wistar studovaným autorádiografií. Neurovědy 20, 599-614. Giuliano, F., Rampin, O., 2000. Centrální kontrola erekce penisu. Neurosci. Biobehav. Rev 24, 517-533. Giuliano, F., Allard, J., 2001. Dopamin a sexuální funkce. Int. J. Impotence Res. 13 (Suppl. 3), 18-28. Giuliano, F., Rampin, O., 2004. Neurální kontrola erekce. Physiol. Behav. 83, 189-201. Giuliano, F., Bernabé, J., McKenna, KE, Longueville, F., Rampin, O., 2001. Spinální procretilní účinek oxytocinu u anestetizovaných krys. Am J. Physiol. Regul. Integ. Comp. Physiol. 280, R1870-R1877. Jdi, Y., Grace, AA, 2005. Dopaminergní modulace limbického a kortikálního posunu nucleus accumbens v cílovém chování. Nat. Neurosci. 8, 805-812. Hawthorn, J., Ang, VT, Jenkins, JS, 1985. Účinky lézí v hypotalamických paraventrikulárních, supraoptických a suprachiasmatických jádrech na vasopresin a oxytocin v mozku potkana a míchy. Brain Res. 346, 51-57. Heier, RF, Dolak, LA, Duncan, JN, Hyslop, DK, Lipton, MF, Martin, LJ, Mauragis, MA, Piercey, MF, Nichols, NF, Schreur, MW, 1997. Syntéza a biologické aktivity (R) -5,6-dihydro-N, N-dimethyl-4H-imidazo [4,5,1-ij] chinolin-5-aminu) a jeho metabolitů. J. Med. Chem. 40, 639-646. Hsieh, GC, Hollingsworth, PR, Martino, B., Chang, R., Terranova, MA, O'Neill, AB, Lynch, JJ, Moreland, RB, Donnelly-Roberts, DL, Kolasa, T., Mikusa, JP McVey, JM, Marsh, KC, Sullivan, JP, Brioni, JD, 2004. Centrální mechanismy regulující erekci penisu u potkanů s vědomím: dopaminergní systémy související s procretilním účinkem apomorfinu. J. Pharmacol. Exp. Ther. 308, 330-338. Huang, PL, Dawson, TM, Bredt, DS, Snyder, SH, Fishman, MC, 1993. Cílené rozrušení genu syntézy oxidu dusnatého. Buňka 75, 1273-1286. Huber, D., Veinante, P., Stoop, R., 2005. Vasopresin a oxytocin vyvolávají zřetelné neuronové populace v centrální amygdale. Věda 308, 245-248. Hull, EM, Warner, RK, Bazzett, TJ, Eaton, RC, Thompson, JT, 1989. Poměr D2 / D1 v mediální preoptické oblasti ovlivňuje kopulaci samců potkanů. J. Pharmacol. Exp. Ther. 251, 422-427. Hull, EM, Du, J., Lorrain, DS, Matuszewich, L., 1995. Extracelulární dopamin v oblasti mediální preoptiky: důsledky sexuální motivace a hormonální kontroly kopulace. J. Neurosci. 15, 7465-7471. Hull, EM, Meisel, RL, Sachs, BD, 2002. Mužské sexuální chování. V: Pfaff, DW, Arnold, AP, Etgen, AM, Fahrbach, SE, Rubin, RT (Eds.), Hormony, mozky a chování. Academic Press, New York, str. 3-137. Hurlemann, R., Patin, A., Onur, OA, Cohen, MX, Baumgartner, T., Metzler S., Dziobek I., Gallinat J., Wagner M., Maier W. Kendrick, KM, 2010. Oxytocin zvyšuje u člověka závislost na amygláli, sociálně posilované učení a emoční empatii. J. Neurosci. 30, 4999-5007. Ivell, R., Russel, JA, 1995. Oxytocin: buněčné a molekulární přístupy v medicíně a výzkumu. Pokroky v experimentální medicíně a biologii, sv. 395. Plenum Press, New York. Kelley, AE, Domesick, VB, 1982. Distribuce projekce od hippocampal formace k nucleus accumbens u krysy: studie anterograde a retrograde-křenová peroxidasa. Neurovědy 7, 2321-2335. Kimura, Y., Naitou, Y., Wanibuchi, F., Yamaguchi, T., 2008. Aktivace receptoru 5-HT (2C) je běžným mechanismem pro procretilní účinky apomorfinu, oxytocinu a melanotanu II u potkanů. E. J. Pharmacol. 589, 157-162. Kondo, Y., Sachs, BD, Sakuma, Y., 1998. Význam mediální amygdaly u erekce penisu u potkanů vyvolaných vzdálenými podněty od estrogenních žen. Behav. Brain Res. 91, 215-222. Kovacs, GL, Sarnyai, Z., Szabo, G., 1998. Oxytocin a závislost: recenze. Psychoneuroendokrinologie 23, 945-962. Lee, HJ, Macbeth, AH, Pagani, JH, Scott Young 3rd, W., 2009. Oxytocin: velký prostředník života. Prog. Neurobiol. 88, 127-151. Lindvall, O., Bjorklund, A., Skagerberg, G., 1984. Selektivní ischochemická demonstrace terminálních systémů dopaminu u di- a telencefalonu potkana: nový důkaz dopaminergní inervace hypotalamických neurosecretorových jader. Brain Res. 306, 19-30. Liu, YC, Salamone, JD, Sachs, BD, 1997. Zhoršená sexuální odpověď po lézích paraventrikulárního jádra hypotalamu u samců potkanů. Behav. Neurosci. 111, 1361-1367. Löber, S., Tschammer, N., Hübner, H., Melis, MR, Argiolas, A., Gmeiner, P., 2009. Azulenová kostra jako nový bioizoster: návrh silných dopaminových D4 receptorových ligandů vyvolávajících erekci penisu. Chem. Med. Chem. 4, 325-328. McCleskey, EW, Fox, AP, Feldman, DH, Cruz, LJ, Olivera, BM, Tsien, RW, Yoshikami, D., 1987. Konotoxin: přímá a trvalá blokáda specifických typů kalciových kanálů v neuronech, nikoliv však ve svalech. Prot. Nat. Acad. Sci. Spojené státy americké 84, 4327-4331. McKenna, KE, 2000. Některé návrhy týkající se organizace kontroly centrální nervové soustavy při erekci penisu. Neurosci. Biobehav. Rev 24, 535-540. Marson, L., McKenna, KE, 1992. Role 5-hydroxytryptaminu při sestupné inhibici spinálních sexuálních reflexů. Exp. Brain Res. 88, 313-318. Marson, L., McKenna, KE, 1996. Skupiny buněk CNS, které se podílejí na kontrole svalů ischiocavernosus a bulbospongiosus: transneuronální sledovací studie s použitím viru pseudorabies. J. Comp. Neurol. 374, 161-179. Meisel, RL, Sachs, BD, 1994. Fyziologie mužského sexuálního chování. In: Knobil, E., Neil, J. (Eds.), Fyziologie reprodukce, sv. 2, druhé vydání. Raven Press, New York, str. 3-96. Melin, P., Kihlstrom, JE, 1963. Vliv oxytocinu na sexuální chování u samců králíků. Endokrinologie 73, 433-435. Melis, MR, Argiolas, A., 1995a. Dopamin a sexuální chování. Neurosci. Biobehav. Rev 19, 19-38. Melis, MR, Argiolas, A., 1995b. Dárci oxidu dusnatého indukují erekci penisu a zívání při injekci do centrálního nervového systému samců potkanů. E. J. Pharmacol. 294, 1-9. Melis, MR, Argiolas, A., 2003. Centrální oxytocinergní neurotransmise: léčebný cíl pro terapii psykogenní erektilní dysfunkce. Curr. Cíle na léky 4, 55-66. Melis, MR, Argiolas, A., Gessa, GL, 1986. Oxytocinem indukované zívání a erekce penisu: místo působení v mozku. Brain Res. 398, 259-265. Melis, MR, Argiolas, A., Gessa, GL, 1987. Apomorfinem indukované zívání a erekce penisu: místo působení v mozku. Brain Res. 415, 98-104. Melis, MR, Argiolas, A., Gessa, GL, 1989a. Apomorfin zvyšuje hladiny oxytocinu v plazmě u potkanů. Neurosci. Lett. 98, 351-355. Melis, MR, Argiolas, A., Gessa, GL, 1989b. Důkaz, že apomorfin indukuje erekci penisu a zívání uvolňováním oxytocinu v centrální nervové soustavě. E. J. Phamacol. 164, 565-570. Melis, MR, Argiolas, A., Stancampiano, R., Gessa, GL, 1990. Účinek apomorfinu na koncentrace oxytocinu v různých oblastech mozku a plazmě samců potkanů. E. J. Pharmacol. 182, 101-107. Melis, MR, Mauri, A., Argiolas, A., 1994a. Apomorfinová a oxytocinem indukovaná erekce penisu a zívání u intaktních a kastrovaných samců potkanů: účinek sexuálních steroidů. Neuroendokrinologie 59, 349-354. Melis, MR, Stancampiano, R., Argiolas, A., 1994b. Penis erekce a zívání vyvolané paraventrikulárními injekcemi NMDA jsou zprostředkovány oxytocinem. Pharmacol. Biochem. Behav. 48, 203-207. Melis, MR, Stancampiano, R., Argiolas, A., 1994c. Prevence metylesteru NG-nitro-l-argininu z erekce penisu a zívání vyvolaného apomorfinem a oxytocinem: místo působení v mozku. Pharmacol. Biochem. Behav. 48, 799-804. Melis, MR, Succu, S., Argiolas, A., 1996. Agonisté dopaminu zvyšují tvorbu oxidu dusnatého v paraventrikulárním jádru hypotalamu: korelace s erekcí penisu a zívání. E. J. Neurosci. 8, 2056-2063. Melis, MR, Succu, S., Iannucci, U., Argiolas, A., 1997. Oxytocin zvyšuje produkci oxidu dusnatého v paraventrikulárním jádru hypotalamu: korelace s erekcí penisu a zívání. Reg. Peptidy 69, 105-112. Melis, MR, Succu, S., Mauri, A., Argiolas, A., 1998. Produkce oxidu dusnatého se zvyšuje v paraventrikulárním jádře hypotalamu samců potkanů během bezkontaktní erekce penisu a kopulace. E. J. Neurosci. 10, 1968-1974. Melis, MR, Spano, MS, Succu, S., Argiolas, A., 1999a. Oxytocinový antagonista d (CH2) 5Tyr (Me) 2-Orn8-vazotocin redukuje bezkontaktní erekci penisu u samců potkanů. Neurosci. Lett. 265, 171-174. Melis, MR, Succu, S., Spano, MS, Argiolas, A., 1999b. Morfin, vstřikovaný do paraventrikulárního jádra hypotalamu, zabraňuje bezkontaktní erekci a narušuje kopulaci: zapojení oxidu dusnatého. E. J. Neurosci. 11, 1857-1864. Melis, MR, Spano, MS, Succu, S., Argiolas, A., 2000. Účinek antagonistů excitační aminokyseliny, dopaminu a oxytocinu na nekontaktní erekci penisu a produkci paraventrikulárního oxidu dusnatého u samců potkanů. Behav. Neurosci. 114, 849-857. Melis, MR, Succu, S., Mascia, MS, Cortis, L., Argiolas, A., 2003. Extracelulární dopamin zvyšuje v paraventrikulárním jádru samců potkanů během sexuální aktivity. E. J. Neurosci. 17, 1266-1272. Melis, MR, Succu, S., Mascia, MS, Argiolas, A., 2004a. Antagonismus kanabinoidních receptorů CB1 v paraventrikulárním jádru samců potkanů vyvolává erekci penisu. Neurosci. Lett. 359, 17-20. Melis, MR, Succu, S., Mascia, MS, Cortis, L., Argiolas, A., 2004b. Extracelulární excitační aminokyseliny se zvyšují v paraventrikulárním jádru samců potkanů během sexuální aktivity: hlavní úloha NMDA receptorů v erektilní funkci. E. J. Neurosci. 19, 2569-2575. Melis, MR, Succu, S., Mascia, MS, Argiolas, A., 2005. PD-168,077, selektivní agonista receptoru dopaminu D4, vyvolává erekci penisu po injekci do paraventrikulárního jádra samců potkanů. Neurosci. Lett. 379, 59-62. Melis, MR, Succu, S., Mascia, MS, Sanna, F., Melis, T., Succu, S., Castelli, MP, Argiolas, A., 2006a. SR 141716A-indukovaná erekce penisu u samců potkanů: postižení paraventrikulární kyseliny glutamové a oxidu dusnatého. Neurofarmakologie 50, 219-228. Melis, MR, Succu, S., Sanna, F., Mascia, MS, Melis, T., Enguehard-Gueiffier, C., Hubner H., Gmenier P., Gueiffier A., Argiolas A., 2006b. PIP3EA a PD168077, dva selektivní agonisté receptoru dopaminu D4, vyvolávají erekci penisu u samců potkanů: místo a mechanismus účinku v mozku. E. J. Neurosci. 24, 2021-2030. Melis, MR, Melis, T., Cocco, C., Succu, S., Sanna, F., Pillolla, G., Boi, A., Ferri, GL, Argiolas, A., 2007. Oxytocin vstřikovaný do ventrální tegmentální oblasti vyvolává erekci penisu a zvyšuje extracelulární dopamin v nucleus accumbens MR Melis, A. Argiolas / Neuroscience a Biobehaviorální recenze 35 (2011) 939-955 955 a paraventrikulární jádro hypotalamu samců potkanů. E. J. Neurosci. 26, 1026-1035. Melis, MR, Sanna, F., Succu, S., Zarone, P., Boi, A., Argiolas, A., 2009a. Úloha oxytocinu v předvídatelné a konzumační fázi pohlavního chování samců potkanů. In: Jastrow, H., Feuerbach, D. (Eds.), Příručka výzkumu oxytocinu: Syntéza, skladování a uvolňování, akční a lékové formy. Nova Publishers Inc, New York, USA, str. 109-125. Melis, MR, Succu, S., Sanna, F., Boi, A., Argiolas, A., 2009b. Oxytocin vstřikovaný do ventrální podkožní vrstvy nebo do bedromediálního kortikálního jádra amygdaly indukuje erekci penisu a zvyšuje extracelulární dopamin v nucleus accumbens samců potkanů. E. J. Neurosci. 30, 1349-1357. Melis, MR, Succu, S., Cocco, C., Caboni, E., Sanna, F., Boi, A., Ferri, GL, Argiolas, A., 2010. Oxytocin indukuje erekci penisu, když je injekčně podán do ventrální subiculum: úloha oxidu dusnatého a kyseliny glutamové. Neurofarmakologie 58, 1153-1160. Molec, RB, Nakane, M., Donnelly-Roberts, DL, Miller, LN, Chang, R., Uchic, ME, Terranova, MA, Gubbins, Masters, JN, Brioni, JD, 2004. Srovnávací farmakologie buněčných linií stabilních receptorů lidského dopaminu D (2) spojených s průtokem vápníku přes Galpha (qo5). Biochem. Pharmacol. 68, 761-772. Moos, F., Freund-Mercier, MJ, Guerne, Y., Guerne, JM, Stoeckel, ME, Richard, P., 1984. Uvolňování oxytocinu a vazopresinu magnetickými jádry in vitro: specifický učinný účinek oxytocinu na jeho vlastní uvolnění. J. Endocrinol. 102, 63-72. Murphy, MR, Seckl, JR, Burton, S., Checkley, SA, Lightman, SL, 1987. Změny sekrece oxytocinu a vazopresinu během sexuální aktivity u mužů. J. Clin. Endocrinol. Metab. 65, 738-741. Nishimori, K., Young, LJ, Guo, Q., Wang, Z., Insel, TR, Matzuk, MM, 1996. Oxytocin je vyžadován pro ošetřování, ale není nezbytný pro porod nebo reprodukční chování. Proc. Natl. Acad. Sci. Spojené státy americké 93, 11699-11704. Patel, S., Freedman, S., Chapman, KL, Emms, Fletcher, AE, Knowles, M., Marwood, R., McCallister, G., Myers, J., Curtis, J., Kulagowski, JJ, Leeson, PD, Ridgill, M., Graham, M., Matheson, S., Rathbone, D., Watt, AP, Bristow, LJ, Rupniak, NM, Baskin, E., Lynch, JJ, Ragan, CI , 1997. Biologický profil L745,870, selektivní antagonista s vysokou afinitou k receptoru dopaminu D4. J. Pharmacol. Exp. Ther. 283, 636-647. Pedersen, CA, Caldwell, JD, Jirikowski, GF, Insel, TR, 1992. Oxytocin v mateřském, sexuálním a sociálním chování, Annals of the New York Academy of Sciences, sv. 652. New York Academy of Sciences, New York. Petrovic, P., Kalisch, R., Singer, T., Dolan, RJ, 2008. Oxytocin zmírňuje afektivní hodnocení podmíněných tváří a amygdální aktivity. J. Neurosci. 28, 6607-6615. Pfaus, JG, Everitt, BJ, 1995. Psychofarmakologie sexuálního chování. V: Knobil, FE, Kupfer, DJ (Eds.), Psychopharmacology: čtvrtá generace pokroku. Raven Press, New York, str. 742-758. Rajfer, J., Aronson, WJ, Bush, PA, Dorey, FJ, Ignarro, LJ, 1992. Oxid dusnatý jako prostředník relaxace corpus cavernosum v reakci na neadrenergní, noncholinergní neurotransmisi. N. Engl. J. Med. 326, 90-94. Roeling, TAP, Van Erp, AMM, Meelis, W., Kruk, MR, Veening, JG, 1991. Behaviorální účinky NMDA injekčně podané do hypotalamického paraventrikulárního jádra krysy. Brain Res. 550, 220-224. Sachs, BD, 1997. Erekce vyvolaná u samců krys vzdušnou vůní z estrousů. Physiol. Behav. 62, 921-924. Sachs, BD, 2000. Kontextuální přístupy k fyziologii a klasifikaci erektilní funkce, erektilní dysfunkce a sexuální vzrušení. Neurosci. Biobehav. Rev 24, 541-560. Sachs, BD, 2007. Kontextová definice mužského sexuálního vzrušení. Horm. Behav. 51, 569-578. Sanchez, F., Alonso, JR, Arevalo, R., Blanco, E., Aijon, J., Vazquez, R., 1994. Koexistence NADPH-diaforázy s vazopresinem a oxytocinem v hypotalamických magnocelulárních neurosecretorových jádrech krys. Cell Tissue Res. 276, 31-34. Sanna, F., Succu, S., Boi, A., Melis, MR, Argiolas, A., 2009. Inhibitory fosfodiesterázy 5 usnadňují nekontaktní erekci samců potkanů: místo působení v mozku a mechanismus účinku. J. Sex. Med. 6, 2680-2689. Saphier, D., Feldman, S., 1987. Účinky septálních a hipokampálních podnětů na neurony paraventrikulárního jádra. Neurovědy 20, 749-755. Sato-Suzuki, I., Kita, I., Oguri, M., Arita, H., 1998. Stereotypní reakce zívání vyvolané elektrickou a chemickou stimulací paraventrikulárního jádra potkana. J. Neurophysiol. 80, 2765-2775. Schuman, EM, Madison, DV, 1994. Oxid dusnatý a synaptická funkce. Ann. Rev Neurosci. 17, 153-183. Snyder, SH, 1992. Oxid dusnatý: nejprve v nové třídě neurotransmiterů? Věda 254, 494-496. Sofroniew, MV, 1983. Vasopresin a oxytocin v savčím mozku a míchu. Trendy Neurosci. 6, 467-472. Sokoloff, P., Schwartz, JC, 1995. Nové dopaminové receptory o půl deset let později. Trends Pharmacol. Sci. 16, 270-275. Southam, E., Garthwaite, J., 1993. Signální dráha oxidů dusíku - cyklická GMP v mozku krys. Neurofarmakologie 32, 1267-1277. Stancampiano, R., Melis, MR, Argiolas, A., 1994. Penis erekce a zívání indukované agonisty 5-HT1c u samců potkanů: vztah k dopaminergnímu a oxytocinergnímu přenosu. E. J. Pharmacol. 261, 149-155. Succu, S., Mascia, MS, Sanna, F., Melis, T., Argiolas, A., Melis, MR, 2006. Kanabinoidní antagonista receptoru CB1 SR 141716A vyvolává erekci penisu zvýšením extracelulární kyseliny glutamové v paraventrikulárním jádru samců potkanů. Behav. Brain Res. 169, 274-281. Succu, S., Sanna, F., Melis, T., Boi, A., Argiolas, A., Melis, MR, 2007. Stimulace dopaminových receptorů v paraventrikulárním jádru hypotalamu samců krys vyvolává erekci penisu a zvyšuje extracelulární dopamin v nucleus accumbens: zapojení centrálního oxytocinu. Neurofarmakologie 52, 1034-1043. Succu, S., Sanna, F., Cocco, C., Melis, T., Boi, A., Ferri, GL, Argiolas, A., Melis, MR, 2008. Oxytocin indukuje erekci penisu po injekci do ventrální tegmentální oblasti samců potkanů: úloha oxidu dusnatého a cyklického GMP. E. J. Neurosci. 28, 813-821. Tanda, G., Pontieri, FE, Di Chiara, G., 1997. Kanabinoidy a heroinová aktivace přenosu mezolimbického dopaminu běžným mechanismem mu1 opioidního receptoru. Věda 276, 2048-2050. Tang, Y., Rampin, O., Calas, A., Facchinetti, P., Giuliano, F., 1998. Oxytocinergní a serotoninergní inervace identifikovaných lumbosakrálních jader kontrolujících erekci penisu u samců potkanů. Neurovědy 82, 241-254. Theodosis, DT, 1985. Oxytocin-imunoreaktivní terminály synapse na oxytocinových neuronech v superoptických jádrech. Příroda (Londýn) 313, 682-684. Tindall, JS, 1974. Stimulace, které způsobují uvolňování oxytocinu. In: Geiger, SR, Knobil, E., Sawyer, WH, Greef, R., Astwood, EB (Eds.), Příručka fyziologie. Sekta. 7, Endocrinology, sv. IV. Americká fyziologická společnost, Washington DC, str. 257-267. Torres, G., Lee, S., Rivier, C., 1993. Ontogeny krysí hypothalamické syntázy oxidu dusnatého a kolokalizace neuropeptidů. Mol. Buňka. Neurosci. 4, 155-163. Uhl-Bronner, S., Waltisperger, E., Martinez-Lorenzana, G., Condes, LM, Freund-Mercier, MJ, 2005. Sexuálně dimorfní exprese vazebných míst oxytocinu v předním mozku a míchu potkana. Neurovědy 135, 147-154. Vaccari, C., Lolait, SJ, Ostrowski, NL, 1998. Srovnávací distribuce vazopresinu V1b a ribonukleových kyselin receptorů oxytocinu v mozku. Endokrinologie 139, 5015-5033. Van Den Pol, A., 1991. Glutamátová a aspartátová imunoreaktivita v hypotalamických presynaptických axonech. J. Neurosci. 11, 2087-2101. Veronneau-Longueville, F., Rampin, O., Freund-Mercier, MJ, Tang, Y., Calas, A., Marson, L., McKenna, KE, Stoeckel, ME, Benoit, G. , 1999. Oxytocinergní inervace autonomních jader kontrolujících erekci penisu u potkanů. Neurovědy 93, 1437-1447. Vincent, SR, Kimura, H., 1992. Histochemické mapování syntázy oxidu dusnatého v mozku krys. Neurovědy 46, 755-784. Wagner, CK, Clemens, LG, 1993. Cesta obsahující neurofyzin z paraventrikulárního jádra hypotalamu do pohlavně dimorfního motorického jádra v bederní páteři. J. Comp. Neurol. 336, 106-116. Winslow, JT, Insel, TR, 1991. Sociální status ve dvojicích opic vejce mužského pohlaví určuje reakci na chování centrálního podávání oxytocinu. J. Neurosci. 11, 2032-2038. Moudrý, RA, Rompre, P.-P., 1989. Mozkový dopamin a odměna. Ann. Rev Psychol. 40, 191-225. Witt, DM, Insel, TR, 1994. Mužské sexuální chování aktivuje protein c-fos v oxytocinových neuronech v paraventrikulárním jádře hypotalamu. J. Neuroendocrinol. 6, 13-18. Witter, MP, 2006. Spojení subikulu krysy: topografie ve vztahu k sloupcové a laminární organizaci. Behav Brain Res. 174, 251-264. Woodruff, GN, Foster, AC, Gill, R., Kemp, JA, Wong, EH, Iversen, LL, 1987. Interakce mezi MK-801 a receptory pro N-methyl-d-aspartát: funkční důsledky. Neurofarmakologie 26, 903-909. Yamashita, H., Shigeru, O., Inenaga, K., Kasai, M., Uesugi, S., Kannan, H., Kaneko, T., 1987. Oxytocin převážně exituuje předpokládané oxytocinové neurony v superoptickém jádře potkana in vitro. Brain Res. 416, 364-368. Yells, DP, Hendricks, SE, Prendergast, MA, 1992. Léze jádra paragigantocelulární: účinky na chování kojení u samců potkanů. Brain Res. 596, 73-79. Young, WS, Shepard, E., Amico, J., Hennighausen, L., LaMarca, ME, McKinney, C., Ginns, EI, 1996. Nedostatek oxytocinu u myší zabraňuje vylučování mléka, ale ne plodnost nebo porod. J. Neuroendocrinol. 8, 847-854. Zahran, AR, Vachon, P., Courtois, F., Carrier, S., 2000. Zvýšení intrakavernózního penisního tlaku po injekci agonistů receptoru excitačních aminokyselin v hypotalamickém paraventrikulárním jádře anestetizovaných krys. J. Urol.