Dev Rev. 2008; 28(1): 62-77. doi: 10.1016 / j.dr.2007.08.003
abstrakta
Adoleskeco estas disvolva periodo karakterizita de suboptimaj decidoj kaj agoj, kiuj okazigas pliigitan efikon de neintencaj vundoj kaj perforto, alkoholo kaj droguzo, neintencita gravedeco kaj sekse transdonitaj malsanoj. Tradiciaj neurobiologiaj kaj kognaj klarigoj pri adoleska konduto malsukcesis kalkuli la ne linealajn ŝanĝojn en konduto observitaj dum adoleskeco, relative al infanaĝo kaj plenaĝeco. Ĉi tiu revizio provizas biologie plaŭdeblan koncepton de la neŭralaj mekanismoj sub la koncernaj ne-liniaj ŝanĝoj en konduto, kiel pli altan respondecon al stimuloj dum impuls-kontrolo ankoraŭ relative nematura dum ĉi tiu periodo. Lastatempaj homaj bildigoj kaj bestaj studoj provizas biologian bazon por ĉi tiu vidpunkto, sugestante diferencan disvolviĝon de lombikaj rekompencaj sistemoj relative al supraj kontrolaj sistemoj dum adoleskeco relative al infanaĝo kaj plenaĝeco. Ĉi tiu evolua mastro povas pliseverigi en tiuj adoleskantoj kun predikado al risko, pliigante la riskon por malbonaj rezultoj.
Laŭ la Nacia Centro por Sanstatistiko, ĉiujare okazas pli ol 13,000-adoleskaj mortoj en Usono. Proksimume 70% de ĉi tiuj mortoj rezultas de kraŝoj de motora veturilo, neintencaj vundoj, hommortigo kaj memmortigo (Eaton et al., 2006). Rezultoj de la Nacia Enketo pri Konduto pri 2005 (YRBS) montras, ke adoleskantoj okupiĝas pri kondutoj, kiuj pliigas sian probablon de morto aŭ malsano veturante veturilon post trinki aŭ sen sigelringo, portante armilojn, uzante kontraŭleĝajn substancojn kaj okupiĝante pri neprotektita sekso. rezultigante senintencajn gravedecojn kaj MST, inkluzive de HIV-infekto (Eaton et al., 2006). Ĉi tiuj statistikoj substrekas la signifon kompreni riskajn elektojn kaj agojn en adoleskantoj.
Multaj kognaj kaj neurobiologiaj hipotezoj estis postulataj pri kial adoleskantoj okupiĝas pri suboptimaj elektaj kondutoj. En lastatempa revizio de la literaturo pri homa adoleska cerba disvolviĝo, Yurgelun-Todd (2007) sugestas, ke kognitiva disvolviĝo tra la adoleskaj jaroj asocias kun iom post iom pli granda efikeco de kognaj kontrolkapabloj. Ĉi tiu efikeco estas priskribita kiel dependa de maturiĝo de la prefrontal-kortekso kiel evidentigita per pliigita aktiveco ene de fokusaj antaŭfrontaj regionoj (Rubia et al., 2000; Tamm, Menon, kaj Reiss, 2002) kaj malpliigita agado en senrilataj cerbaj regionoj (Brown et al., 2005; Durston et al., 2006).
Ĉi tiu ĝenerala mastro, de plibonigita kognitiva kontrolo kun maturiĝo de la antaŭfronta kortekso, sugestas linean kreskon de disvolviĝo de infanaĝo ĝis plenaĝeco. Sed tamen suboptimaj elektoj kaj agoj observitaj dum adoleskeco reprezentas neliniajn ŝanĝojn en konduto distingebla de infanaĝo kaj plenkreskeco, kiel pruvis la Nacia Centro por Sanstatistiko pri adoleska konduto kaj morteco. Se kognitiva kontrolo kaj nematura antaŭfronta kortekso estis la bazo por suboptimaj elektaj kondutoj, tiam infanoj devas aspekti rimarkinde similaj aŭ eĉ pli malbonaj ol adoleskantoj, konsiderante siajn malpli evoluintajn antaŭkordajn kortekson kaj kognajn kapablojn. Tiel, nematura antaŭfronto funkcias sole, ne povas kalkuli pri adoleska konduto.
Preciza konceptado de kognaj kaj neŭobiobiologiaj ŝanĝoj dum adoleskeco devas trakti la adoleskecon kiel transiran disvolviĝan periodon (Spear, 2000), prefere ol unuopa kapto en la tempo (Casey, Tottenham, Liston, kaj Durston, 2005). Alivorte, por kompreni ĉi tiun disvolvan periodon, necesas transiroj en kaj el la adoleskeco por distingi distingajn atributojn de ĉi tiu stadio de disvolviĝo. Starigi disvolvajn trajektoriojn por kognaj kaj neŭralaj procezoj estas esenca por karakterizi ĉi tiujn transirojn kaj limigi interpretojn pri ŝanĝoj en konduto dum ĉi tiu periodo. Sur kognitiva aŭ kondutisma nivelo, adoleskantoj estas karakterizataj kiel impulsemaj (t.e. mankas kognitiva kontrolo) kaj riskantaj kun ĉi tiuj konstruoj uzataj sinonime kaj sen aprezo por apartaj disvolvaj trajektorioj de ĉiu. Sur neurobiologia nivelo, homaj bildoj kaj bestaj studoj sugestas distingajn neurobiologiajn bazojn kaj disvolvajn trajektoriojn por la neŭralaj sistemoj, kiuj subtenas ĉi tiujn apartajn konstruojn de impulsa kontrolo kaj riskaj decidoj.
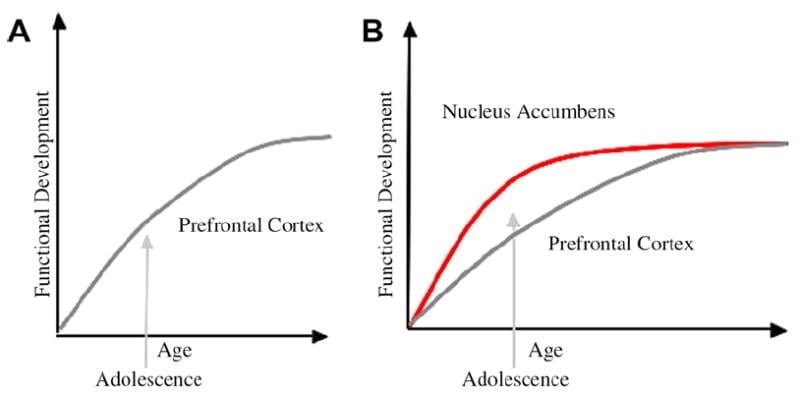
Ni disvolvis neurobiologian modelon de adoleska evoluo en ĉi tiu kadro, kiu baziĝas sur ronĝaj modeloj (Laviola, Adriani, Terranova, & Gerra, 1999; Spear, 2000) kaj freŝaj studoj pri bildigo de adoleskecoErnst et al., 2005; Galvan, Leporo, Voss, Gantfaristo, kaj Casey, 2007; Galvan et al., 2006). Figo. 1 sube montras ĉi tiun modelon. Maldekstre estas la tradicia karakterizado de la adoleskeco kiel rilata preskaŭ ekskluzive al la nematureco de la antaŭfronta kortekso. Dekstre estas nia proponita neurobiologia modelo, kiu ilustras, kiel devas konsideri kune regionajn limajn subkortikajn kaj prefrontalajn suprajn malsupren-kontrolajn regionojn. La bildstrio ilustras malsamajn disvolvajn trajektoriojn por ĉi tiuj sistemoj, kun sistemoj de limuzinoj disvolviĝantaj pli frue ol antaŭfrontaj regionoj de kontrolo. Laŭ ĉi tiu modelo, la individuo estas pli tordita de funkcie maturaj lombikaj regionoj dum adoleskeco (t.e., malekvilibro de limfika rilato al antaŭfrontalregado), kompare kun infanoj, por kiuj ĉi tiuj sistemoj (t.e., lombikaj kaj prefrontalaj) daŭre disvolviĝas; kaj kompare kun plenkreskuloj, por kiuj ĉi tiuj sistemoj plene maturiĝas. Ĉi tiu perspektivo provizas bazon por ne linearaj movoj en konduto trans disvolviĝo, pro pli frua maturiĝo de ĉi tiu limoj relative al malpli maturaj supraj malsupren antaŭfrontaj regionoj. Kun disvolviĝo kaj sperto, la funkcia konektebleco inter ĉi tiuj regionoj provizas mekanismon por suprena kontrolo de ĉi tiuj regionoj (Leporo, Voss, Gantfaristo, kaj Casey, 2007a). Plue, la modelo akordigas la kontraŭdiron de sanstatistikoj pri riska konduto dum adoleskeco, kun la ruza observado de Reyna kaj Farley (2006) ke adoleskantoj kapablas rezoni kaj kompreni riskojn de kondutoj, en kiuj ili okupiĝas. Laŭ nia modelo, en emocie elstaraj situacioj, la limbika sistemo venkos super kontrolsistemoj donita sian maturecon rilate al la antaŭfrontal-kontrolsistemo. Evidenteco de kondutismaj kaj homaj figuradaj studoj por subteni ĉi tiun modelon estas provizita en la kunteksto de agoj en rekompencaj kaj emociaj kuntekstoj (Galvan et al., 2006, 2007; Leporo, Voss, Gantfaristo, kaj Casey, 2007b; Hare et al., 2007a). Krome, ni spekulas kial la cerbo povas disvolviĝi tiamaniere kaj kial iuj adoleskantoj povas havi pli grandan riskon fari suboptimajn decidojn kondukantajn al pli malriĉaj longtempaj rezultoj (Galvan et al., 2007; Hare et al., 2007b).
La tradicia klarigo pri adoleska konduto sugestis esti pro ŝuldata evoluo de la antaŭfrontaleca kortekso (A). Nia modelo enkalkulas la disvolviĝon de la antaŭfronta kortekso kune kun subkortikaj limuzaj regionoj (ekz., Nucleus accumbens), kiuj estis implikitaj en riskaj elektoj kaj agoj (B).
Disvolviĝo de cel-direktita konduto
Ŝtono de kognitiva evoluo estas la kapablo subpremi netaŭgajn pensojn kaj agojn favore al cel-direktitaj, precipe en ĉeesto de devigaj instigoj (Casey, Galvan, & Hare, 2005; Casey et al., 2000b; Casey, Tomaso, David-filo, Kunz, kaj Franzen, 2002a; Casey, Tottenham, kaj Fossella, 2002b). Multaj klasikaj evoluaj studoj montris, ke ĉi tiu kapablo disvolviĝas dum la infanaĝo kaj adoleskeco (Kazo, NENIU; Flavell, Feach, kaj Chinsky, 1966; Keating & Bobbitt, 1978; Pascual-Leone, NENIU). Pluraj teoriuloj argumentis, ke kognitiva disvolviĝo estas pro kreskoj de prilabora rapideco kaj efikeco kaj ne pro pliigo de mensa kapablo (ekz. Bjorklando, 1985; Bjorklando, 1987; Kazo, NENIU). Aliaj teoriuloj inkluzivis la konstruon de procezoj “inhibitoriaj” en sia konto de kognitiva disvolviĝo (Harnishfeger & Bjorkland, 1993). Laŭ ĉi tiu konto, nematura kogno estas karakterizata de susceptibilidad al interfero de konkurantaj fontoj, kiuj devas esti subpremitaj (ekz. Brainerd & Reyna, 1993; Casey, Thomas, Davidson, Kunz, kaj Franzen, 2002a; Dempster, 1993; Diamanto, 1985; Munakata & Yerys, 2001). Tiel cel-direktita konduto postulas regadon de impulsoj aŭ prokrasto de gratifiko por optimumigo de rezultoj kaj tiu kapablo ŝajnas maturiĝi tra infanaĝo kaj adoleskeco.
Adoleska konduto estis priskribita kiel impulsema kaj riska, preskaŭ sinonime, tamen ĉi tiuj konstruaĵoj dependas de malsamaj kognaj kaj neŭralaj procezoj, kiuj sugestas distingajn konstruojn kun malsamaj disvolvaj trajektorioj. Specife, revizio de la literaturo sugestas, ke impulsiveco malpliiĝas kun la aĝo tra infanaĝo kaj adoleskeco (Casey et al., 2002a; Casey, Galvan et al., 2005; Galvan et al., 2007) kaj estas asociita kun plilongigita evoluo de la antaŭfronta kortekso (Casey, Galvan et al., 2005), kvankam estas diferencoj en la grado, en kiu difinita individuo estas impulsema aŭ ne, sendepende de aĝo.
En kontrasto, al impulsa / kognitiva kontrolo, risko-kreskado ŝajnas pliiĝi dum adoleskeco rilate infanaĝon kaj plenkreskecon kaj estas asociita kun subkortikaj sistemoj, konataj okupiĝante pri taksado de rekompencoj. Studoj pri homaj bildoj, kiuj estos reviziitaj, sugestas kreskon de subkortika aktivado (ekz. Akciuloj) dum farado de riskaj elektoj (Kuhnen & Knutson, 2005; Matthews kaj aliaj, 2004; Montague & Berns, 2002) troigita en adoleskantoj, rilate al infanoj kaj plenkreskuloj (Ernst et al., 2005; Galvan et al., 2006). Ĉi tiuj trovoj sugestas malsamajn trajektoriojn por rekompenco aŭ instigo-bazita konduto, kun pli frua disvolviĝo de ĉi tiuj sistemoj relative al kontrolaj sistemoj, kiuj montras longan kaj linean disvolvan kurson, se temas pri superregado de netaŭgaj elektoj kaj agoj favore al cel-direktitaj.
Evidenteco de neŭroimagaj studoj pri homa cerba disvolviĝo
Lastatempaj esploroj pri adoleska cerba disvolviĝo baziĝis sur progresoj en neŭroimagaj metodaroj facile uzeblaj kun evoluantaj homaj loĝantaroj. Ĉi tiuj metodoj dependas de magneta resonanca bildigo (MRI), metodoj (vidu Figo. 2) kaj inkluzivas: struktura MRI, uzata por mezuri la grandecon kaj formon de strukturoj; funkcia MRI kiu estas uzata por mezuri padronojn de cerba aktiveco; kaj difusa tensia bildigo (DTI), kiu estas uzata por indeksigi konekteblecon de blankaj materiaj fibroj. Evidenteco por nia evolua modelo de konkurenco inter kortikaj kaj subkortikaj regionoj estas subtenata de nematura struktura kaj funkcia konektebleco respektive mezurita de DTI kaj fMRI.
MRI-studoj pri homa cerba disvolviĝo
Pluraj studoj uzis strukturan MRI por mapi la anatomian kurson de normala cerba disvolviĝo (vidu recenzon de Durston et al., 2001). Kvankam la totala cerba grandeco estas proksimume 90% de sia plenkreska grandeco en aĝo de ses jaroj, la grizaj kaj blankaj materiaj subkomponentoj de la cerbo daŭre spertas dinamikajn ŝanĝojn dum la adoleskeco. Datumoj de lastatempaj longformaj MRI-studoj indikas, ke volumeno de griza materio havas inversigitan U-formon, kun pli granda regiona variaĵo ol blanka materio (Giedd, 2004; Gogtay et al., 2004; Sowell et al, 2003; Sowell, Thompson, & Toga, 2004). Enerale regionoj submetataj al primaraj funkcioj, ekzemple motoraj kaj sensaj sistemoj, maturiĝas plej frue; Areoj de pli alta ordo, kiuj integras ĉi tiujn primarajn funkciojn, maturiĝas posteGogtay et al., 2004; Sowell, Thompson, & Toga, 2004). Ekzemple, studoj uzantaj MRI-bazitajn mezurojn montras, ke perdo de kortikaj grizaj substancoj okazas plej frue en la ĉefaj sensiempaj areoj kaj plej malfrue en la dorsolateraj antaŭ-orientaj kaj flankaj tempaj korturoj (Gogtay et al., 2004). Ĉi tiu ŝablono kongruas kun nehomaj primatoj kaj homaj postmortemaj studoj montrante, ke la antaŭfronta kortekso estas unu el la lastaj cerbaj regionoj maturiĝiBurĝo, Goldman-Rakic kaj Rakic, 1994; Huttenlocher, 1979). Kontraste al griza materio, blanka materio volumeno pliiĝas laŭ proksimume lineara ŝablono, pliiĝante dum tuta evoluo bone en plenaĝecon (Gogtay et al., 2004). Ĉi tiuj ŝanĝoj supozeble reflektas daŭran mielinigon de axonoj per oligodendrocitoj plibonigante neuronal kondukadon kaj komunikadon.
Kvankam malpli da atento estis donita al subkortikaj regionoj kiam oni ekzamenas strukturajn ŝanĝojn, kelkaj el la plej grandaj ŝanĝoj en la cerbo vidiĝas en ĉi tiuj regionoj, precipe en la basaj ganglioj (Sowell et al., 1999, vidu Figo. 3) kaj precipe ĉe maskloj (Giedd et al., 1996). Evoluaj ŝanĝoj en struktura volumo ene de bazaj ganglioj kaj antaŭfrontaj regionoj estas interesaj sekve de konataj disvolvaj procezoj (ekz. Dendrita arborigo, ĉela morto, sinaptika pritondado, mielinigo) okazantaj dum infanaĝo kaj adoleskeco. Ĉi tiuj procezoj permesas bonegan agordadon kaj plifortigon de ligoj inter antaŭfrontaj kaj subkortikaj regionoj kun disvolviĝo kaj lernado, kiuj eble koincidas kun pli granda kognitiva kontrolo. Kiel ĉi tiuj strukturaj ŝanĝoj rilatas al kognaj ŝanĝoj? Multaj studoj rilatigis frontan lobon strukturan maturiĝon kaj kognan funkcion uzante neuropsikologiajn kaj kognajn mezurojn (ekz. Sowell et al., 2003). Specife, raportoj estis raportitaj inter MRI-bazitaj antaŭfrontalaj kortikaj kaj bazaj regionaj ganglioj kaj mezuroj de kognitiva kontrolo (t.e. kapablo superregi maltaŭgan respondon favore al alia aŭ subpremi atenton al senrilata stimula atributo favore al rilata stimula atributo (Casey, Trainor et al., 1997). Ĉi tiuj trovoj sugestas, ke kognaj ŝanĝoj speguliĝas en strukturaj cerbaj ŝanĝoj kaj substrekas la gravecon de subkortika (basaj ganglioj) same kiel kortikan (ekz. Prefrontalan kortekson) disvolviĝon.

DTI-studoj pri homa cerba disvolviĝo
La MRI-bazitaj morfometry-studoj reviziitaj sugestas, ke kortikaj ligoj perfekte agordas kun la elimino de troa abundeco de sinapsoj kaj plifortigo de koncernaj rilatoj kun disvolviĝo kaj sperto. Lastatempaj progresoj en MRI-teknologio, kiel DTI, provizas eblan ilon por ekzameni la rolon de specifaj blankaj materioj por la disvolviĝo de la cerbo kaj konduto kun pli da detalo. Gravaj al ĉi tiu papero estas la neŭroimaj studoj, kiuj ligis la disvolviĝon de fibraj traktoj kun plibonigoj de kognitiva kapablo. Specife, asocioj inter DTI-bazitaj mezuroj pri antaŭfrontal blanka materio kaj kognitiva kontrolo en infanoj estis montritaj. En unu studo, disvolviĝo de ĉi tiu kapablo pozitive korelaciis kun antaŭfrontal-parietalaj fibraj traktoj (Nagy, Westerberg, kaj Klingberg, 2004) konforma al funkciaj neŭroimagaj studoj montrantaj diferencan varbadon de ĉi tiuj regionoj en infanoj relative al plenkreskuloj.
Uzante similan aliron, Liston et al. (2005) montris, ke blankaj materioj inter antaŭfrontal-bazaj ganglioj kaj -postaj fibraj traktoj daŭre disvolviĝas tra infanaĝo ĝis plenkreskeco, sed nur tiuj spuroj inter la prefrontal-kortekso kaj basaj ganglioj estas korelaciitaj kun impulsa kontrolo, kiel mezurite per agado dum irado / nogo tasko. La prefrontalaj fibraj traktoj estis difinitaj de regionoj de interesoj identigitaj en fMRI-studo uzante la saman taskon. Inter ambaŭ disvolvaj DTI-studoj, fibraj traktaj mezuroj estis korelaciitaj kun evoluo, sed specifeco de apartaj fibraj traktoj kun kognitiva agado estis montrita disociante la apartan traktadon (Liston et al., 2005) aŭ kogna kapablo (Nagy et al., 2004). Ĉi tiuj trovoj substrekas la gravecon ekzameni ne nur regionajn, sed cirkvitajn rilatajn ŝanĝojn kiam faras asertojn pri aĝoj dependantaj de aĝo en neŭraj substratoj de kognitiva disvolviĝo.
Funkciaj MRI-studoj pri kondutisma kaj cerba disvolviĝo
Kvankam strukturaj ŝanĝoj mezuritaj per MRI kaj DTI estis asociitaj kun kondutaj ŝanĝoj dum disvolviĝo, pli rekta alproksimiĝo por ekzamenado de strukturo-funkcia asocio estas mezuri ŝanĝojn en la cerbo kaj konduto samtempe, kiel kun fMRI. La kapablo mezuri funkciajn ŝanĝojn en la evolua cerbo kun MRI havas signifan potencialon por la kampo de disvolva scienco. Kadre de la aktuala papero, fMRI disponigas rimedon por limigi interpretojn de adoleska konduto. Kiel dirite antaŭe, oni opinias, ke la disvolviĝo de la antaŭfronta kortekso ludas gravan rolon en la maturiĝo de pli altaj kognaj kapabloj kiel ekzemple decidiĝo kaj kognitiva kontrolo (Casey, Tottenham, kaj Fossella 2002b; Casey, Trainor et al., 1997). Multaj paradigmoj estis uzataj kune kun fMRI por taksi la neurobiologiajn bazojn de ĉi tiuj kapablecoj, inkluzive taskojn de flanker, Stroop kaj go / nogo (Casey, Castellanos et al., 1997; Casey, Giedd, kaj Thomas, 2000a; Durston et al., 2003). Kolektive, ĉi tiuj studoj montras, ke infanoj rekrutas malsamajn sed ofte pli grandajn, pli disvastiĝantajn antaŭfrontajn regionojn kiam ili plenumas ĉi tiujn taskojn ol plenkreskuloj. La mastro de agado ene de cerbaj regionoj centra por taska agado (t.e., kiu rilatas al kognitiva agado) fariĝas pli fokusa aŭ pli agordita kun aĝo, dum regionoj ne korelaciaj kun taska agado malpliiĝas en aktiveco kun aĝo. Ĉi tiu ŝablono estis observita en ambaŭ transversaj sekcioj (Brown et al., 2005) kaj longitudaj studoj (Durston et al., 2006) kaj trans diversaj paradigmoj. Kvankam neuroimagaj studoj ne povas definitive karakterizi la me mechanismanismon de tiaj evoluaj ŝanĝoj (ekz. Dendrita arborigo, sinaptika rikolto) la trovoj reflektas disvolviĝon ene de kaj rafinado de projekcioj de kaj de, aktivigitaj cerbaj regionoj kun maturiĝo kaj sugestas, ke ĉi tiuj ŝanĝoj okazu super longa tempodaŭro (Brown et al., 2005; Bunge, Dudukovic, Thomason, Vaidya, kaj Gabrieli, 2002; Casey, Trainor et al., 1997; Casey et al., 2002a; Crone, Donohue, Honomichl, Wendelken, & Bunge, 2006; Luna et al., 2001; Moses et al., 2002; Schlaggar et al., 2002; Tamm et al., 2002; Thomas et al., 2004; Turkeltaub, Gareau, Floroj, Zeffiro, kaj Edeno, 2003).
Kiel ĉi tiu metodaro povas informi nin pri tio, ĉu adoleskantoj ja mankas sufiĉan kognan regadon (impulseman) aŭ riskas siajn elektojn kaj agojn? Impulso-kontrolo kiel mezurita per kognaj kontrolaj taskoj kiel la tasko de iro / nogo montras linean padronon de disvolviĝo tra infanaĝo kaj adoleskeco kiel priskribite supre. Tamen, lastatempaj neŭroimagaj studoj komencis ekzameni rekompencon-prilaborado specifa al risko-prenado en adoleskantoj (Bjork et al., 2004; Ernst et al., 2005; Majo et al., 2004). Ĉi tiuj studoj koncentriĝis ĉefe sur la regiono de la akciuloj, parto de la bazaj ganglioj implikitaj en antaŭdiro de rekompenco, anstataŭ karakterizado de la disvolviĝo de ĉi tiu regiono lige kun supraj kontrolaj regionoj (prefrontal-kortekso). Kvankam lastatempa raporto pri malpli ventra prefrontal-agado en adoleskantoj relative al plenkreskuloj dum mona decidiĝa tasko pri riska konduto estis montrita (Eshel, Nelson, Blair, Pine, & Ernst, 2007).
Entute, malmultaj studoj ekzamenis kiel la evoluo de rekompenccirkvitoj en subkortikaj regionoj (ekz., Akciuloj) ŝanĝiĝas lige kun disvolviĝo de kortikaj antaŭfrontaj regionoj. Plie, kiel ĉi tiuj neŭraj ŝanĝoj koincidas kun rekompenco, impulsemo kaj risko-konduto restas relative nekonataj. Nia neurobiologia modelo proponas, ke la kombinaĵo de pli alta respondeco al rekompenco kaj nematureco en kondutregulaj regionoj eble aĉos adoleskantojn serĉi tujan, anstataŭ longtempajn gajnojn, eble klarigante sian pliiĝon de riska decido kaj impulsaj kondutoj. Spuri subkortikan (ekz. Akciindan) kaj kortikan (ekz. Antaŭfrontalan) disvolviĝon de decidiĝo tra infanaĝo tra plenaĝeco, provizas pliajn limigojn pri tio, ĉu ŝanĝoj raportitaj en adoleskeco estas specifaj al ĉi tiu periodo de disvolviĝo, aŭ reflektas maturiĝon konstante okazantan en iom linia aranĝo de infanaĝo ĝis plenaĝeco.
Empiria evidenteco de freŝa fMRI-studo helpas subteni nian neurobiologian modelon kaj prenas transiran aliron al kompreno de adoleskeco per ekzamenado de ŝanĝoj antaŭ kaj sekvantaj adoleskecon. En ĉi tiu studo (Galvan et al., 2006), ni ekzamenis kondutajn kaj neŭralajn respondojn por rekompenci manipuladojn trans disvolviĝo, enfokusigante cerbajn regionojn implicitajn en rekompenc-lernado kaj konduto en bestoj (Hikosaka & Watanabe, 2000; Pecina, Cagniard, Berridge, Aldridge, kaj Zhuang, 2003; Schultz, 2006) kaj plenkreskaj bildaj studoj (ekz. Knutson, Adams, Fong, & Hommer, 2001; O, Doherty, Kringelbach, Rolls, Hornak, Andrews, 2001; Zald et al., 2004) kaj en studoj pri toksomanio (Hyman & Malenka, 2001; Volkow & Li, 2004). Surbaze de rodentaj modeloj (Laviola et al., 1999; Spear, 2000) kaj antaŭa bilda laboro (Ernst et al., 2005), ni hipotezis, ke rilate al infanoj kaj plenkreskuloj, adoleskantoj montrus troigitan aktivadon de la akuzantoj, koncerte kun malpli matura rekrutado de supraj malsupren antaŭfrontaj regionoj. Lastatempa laboro montranta malfruan funkcian konekteblecon inter ĉi tiuj antaŭfrontalaj kaj limuzikaj subkortikaj regionoj en adoleskeco rilate al plenkreskuloj, provizas mekanismon por la manko de suprena kontrolo de ĉi tiuj regionoj (Hare et al., 2007a).
Niaj trovoj konformis al modeloj de ronĝuloj (Laviola, Macri, Morley-Fletcher, & Adriani, 2003) kaj antaŭaj bildigaj studoj (Ernst et al., 2005) sugestante plibonigitajn akciigajn agadojn rekompencitajn dum adoleskeco. Efektive, relative al infanoj kaj plenkreskuloj, adoleskantoj montris troigitan akcizan respondon antaŭ atendo de rekompenco. Tamen, infanoj kaj adoleskantoj montris malpli matura respondo en regionoj de antaŭfronto kontrolo ol plenkreskuloj. Ĉi tiuj trovoj sugestas, ke malsamaj disvolvaj trajektorioj por ĉi tiuj regionoj povas substreki la plibonigon de akciaj aktivecoj, relative al infanoj aŭ plenkreskuloj, kiuj eble rilatas al la pliigitaj impulsaj kaj riskaj kondutoj observitaj dum ĉi tiu periodo de disvolviĝo (vidu Figo. 4).

Malsamaj varboj de antaŭfrontaj kaj subkortikaj regionoj estis raportitaj tra kelkaj disvolvaj fMRI-studoj (Casey et al., 2002b; Monk et al., 2003; Thomas et al., 2004). Tipe ĉi tiuj trovoj estis interpretitaj laŭ nematuraj prefrontaj regionoj prefere ol malekvilibro inter prefrontala kaj subkorta seksa regiona disvolviĝo. Pruvoj de antaŭfrontaj regionoj por gvidi taŭgajn agojn en malsamaj kuntekstojMiller kaj Cohen, 2001) nematura prefrontala agado povus malhelpi taŭgan korinklinon de estontaj rezultoj kaj takso de riskaj elektoj, kaj eble tiel malpli influi sur kompensvaloro ol la akcumbenoj. Ĉi tiu ŝablono kongruas kun antaŭa esplorado montranta altan subkortikan, relativan al kortika agado kiam decidoj estas influitaj tuje per longperspektivaj gajnoj (McClure, Laibson, Loewenstein, & Cohen, 2004). Plue, komuna agado estis montrita per fMRI por pozitive korelacii kun postaj riskoj-prenantaj kondutoj (Kuhnen & Knutson, 2005). Dum adoleskeco, relative al infanaĝo aŭ plenkreskeco, nematura ventra antaŭfronta kortekso eble ne provizas sufiĉan supran malsupren kontrolon de fortike aktivigitaj rekompencaj regionoj (ekz., Akciuloj), rezultigante malpli influon de antaŭfrostaj sistemoj (orbitofronta kortekso) relative al la akciuloj rekompencaj. taksado
Kial la cerbo estus programita tiel disvolvi?
Adoleskeco estas la transira periodo inter infanaĝo kaj plenaĝeco ofte okazanta kun pubereco. Pubereco markas la komencojn de seksa maturiĝo (Graber & Brooks-Gunn, 1998) kaj povas esti difinita per biologiaj markiloj. La adoleskeco povas esti priskribita kiel progresema transiro al plenkreskeco kun nebula ontogenetika tempo-kurso.Spear, 2000). Evoluke parolante, adoleskeco estas la periodo, kiam oni akiras sendependecajn kapablojn por pliigi sukceson post disiĝo de la protekto de la familio, kvankam pliigas eblecojn por malutilaj cirkonstancoj (ekz. Vundo, depresio, angoro, konsumado de drogoj kaj toksomanio (Kelley, Schochet, & Landry, 2004). Sendependec-serĉantaj kondutoj estas ĝeneralaj en specioj, kiel ekzemple pliigoj en sociaj direktitaj sociaj interagoj kaj intensigoj en novec-serĉantaj kaj riskantaj kondutoj. Psikosiaj faktoroj influas adoleskan inklinon al riska konduto. Tamen, riska konduto estas la produkto de biologie movita malekvilibro inter pliigita noveco kaj serĉado de sentoj kune kun nematura "memregula kompetenteco" (Steinberg, 2004). Niaj neurobiologiaj datumoj sugestas, ke ĉi tio okazas per diferenca disvolviĝo de ĉi tiuj du sistemoj (limbic kaj kontrolo).
Spekulado sugestas, ke ĉi tiu evolua ŝablono estas evolua trajto. Vi devas okupiĝi pri riska konduto por forlasi vian familion kaj vilaĝon por trovi kunulon kaj riskon, samtempe, kiam hormonoj pelas adoleskantojn serĉi seksajn partnerojn. En la hodiaŭa socio, kiam la adoleskeco povas etendiĝi senfine, kun infanoj vivantaj kun gepatroj kaj financaj dependecoj kaj elektantaj samideanoj poste en la vivo, ĉi tiu evoluo povas esti konsiderata nekonvena.
Estas evidenteco ĉe specioj pri pli alta serĉado de novecoj kaj risko dum la adoleskaj jaroj. Serĉi samseksajn samulojn kaj kvereli kun gepatroj, kiuj ĉiuj helpas forigi la adoleskanton for de la hejma teritorio por pariĝo en aliaj specioj, inkluzive de ronĝuloj, nehomaj primatoj kaj iuj birdoj (Spear, 2000). Rilate al plenkreskuloj, periadoleskaj ratoj montras pliigajn nov-serĉantajn kondutojn en libera elekto-noviga paradigmo (Laviola et al., 1999). Neŭkemia indico indikas, ke la ekvilibro en la adoleska cerbo inter kortikaj kaj subkortikaj dopaminaj sistemoj, komencas ŝanĝi pli grandajn kortikajn dopaminajn nivelojn dum adoleskeco (Spear, 2000). Simila longedaŭra dopaminergia konservado tra adoleskeco en plenaĝecon estis montrita ankaŭ en la nehoma homa primata kortekso (Rosenberg & Lewis, 1995). Tiel ĉi tiu alta ŝajna risko ŝajnas esti inter specioj kaj havas gravajn adaptajn celojn.
Biologiaj antaŭdiroj, disvolviĝo kaj risko
Individuaj diferencoj en impulsa kontrolo kaj prenado de riskoj estas rekonataj en psikologio antaŭ iom da tempo (Benthin, Slovic kaj Severson, 1993). Eble unu el la klasikaj ekzemploj de unuopaj diferencoj raportitaj pri ĉi tiuj kapabloj en la socia, kognitiva kaj disvolva psikologio-literaturo estas prokrasto de gratifiko (Mischel, Shoda, kaj Rodriguez, 1989). Prokrasto de dankemo estas tipe taksita en 3- al 4-jaraĝaj infanetoj. La infanito demandas, ĉu ili preferas malgrandan rekompencon (unu kuketon) aŭ grandan rekompencon (du kuketojn). La infano tiam estas dirita, ke la eksperimentisto forlasos la ĉambron por prepari sin por venontaj agadoj kaj klarigas al la infano, ke se ŝi restos sur sia sidloko kaj ne manĝas kuketon, ŝi ricevos la grandan rekompencon. Se la infano faras aŭ ne povas atendi, ŝi sonu sonorilon por alvoki la eksperimentanton kaj tiel ricevi la pli malgrandan rekompencon. Post kiam estas certe, la infano komprenas la taskon, ŝi sidas ĉe la tablo kun la du rekompencoj kaj la sonorilo. Distancoj en la ĉambro estas minimumigitaj, sen ludiloj, libroj aŭ bildoj. La eksperimentanto revenas post 15 min aŭ post kiam la infano sonorigis la sonorilon, manĝis la rekompencojn aŭ montris ajnajn signojn de ĝeno. Mischel montris, ke infanoj tipe kondutas laŭ unu el du manieroj: (1) ĉu ili sonas la sonorilon preskaŭ tuj por havi la kuketon, kio signifas, ke ili nur ricevas unu; aŭ (2) ili atendas kaj optimumigas siajn gajnojn kaj ricevas ambaŭ kuketojn. Ĉi tiu observado sugestas, ke iuj individuoj estas pli bonaj ol aliaj en sia kapablo kontroli impulsojn fronte al tre elstaraj instigoj kaj ĉi tiu parodio povas esti detektita en frua infanaĝo (Mischel et al., 1989) Kaj ili ŝajnas resti dum la adoleskeco kaj juna plenaĝeco (Eigsti et al., 2006).
Kio povus klarigi individuajn diferencojn en optimuma decidado kaj konduto? Iuj teoriuloj postulis, ke dopaminergiaj mesolimbaj cirkvitoj, implicitaj en prilaboro, subigas riska konduto. Individuaj diferencoj en ĉi tiu cirkvito, kiel alelaj variantoj en dopamin-rilataj genoj, rezultantaj en tro malmulte aŭ tro da dopamino en subkortikaj regionoj, povus rilati al la propensieco okupi riskan konduton (O'Doherty, 2004). La kerno accumbens pruviĝis pliiĝi en aktiveco tuj antaŭ ol fari riske elektojn pri mon-riskaj paradigmoj (Kuhnen & Knutson, 2005; Matthews et al., 2004; Montague & Berns, 2002) kaj kiel priskribite antaŭe, adoleskantoj montras troigajn aĉajn agadojn al rekompencaj rezultoj rilate infanojn aŭ plenkreskulojn (Ernst et al., 2005; Galvan et al., 2006). Kolektive, ĉi tiuj datumoj sugestas, ke adoleskantoj povas esti pli inklinaj al riskaj elektoj kiel grupo (Ĝardenisto & Steinberg, 2005), sed iuj adoleskantoj estos pli inklinaj ol aliaj al riska konduto, metante ilin en eble pli grandan riskon por negativaj rezultoj. Tial gravas konsideri individuan variecon kiam oni ekzamenas kompleksajn cerbajn kondutajn rilatojn rilatigitaj kun risko-preno kaj rekompenco-prilaborado en evoluaj populacioj.
Esplori individuajn diferencojn en risko-konduto, Galvan et al. (2007) lastatempe ekzamenis la asocion inter agado en rekompencaj neŭralaj cirkvitoj antaŭvide al granda mona rekompenco kun personecaj trajtoj de risko kaj impulsemo en adoleskeco. Funkcia magneta resonanca bildigo kaj anonima mem-raporta skalo de riska konduto, riska percepto kaj impulsiveco estis akiritaj en individuoj inter 7 kaj 29-jaroj. Ekzistis pozitiva asocio inter kutimaj agadoj kaj la probableco okupiĝi pri riska konduto trans disvolviĝo. Ĉi tiu aktiveco variis kiel funkcio de taksoj de individuoj pri antaŭviditaj pozitivaj aŭ negativaj konsekvencoj de tia konduto. Tiuj individuoj, kiuj perceptis riskajn kondutojn kiel malfacilajn konsekvencojn, aktivigis la akuzantojn malpli rekompenci. Ĉi tiu asocio estis antaŭenigita plejparte de la infanoj, kaj la plenkreskuloj taksis la konsekvencojn de tia konduto kiel eble. Impulsivecaj rangoj ne estis asociitaj kun akciza agado, sed prefere kun aĝo. Ĉi tiuj trovoj sugestas, ke dum adoleskeco, iuj individuoj povas esti pli inklinaj al riskaj kondutoj pro evoluaj ŝanĝoj koncerte kun varieco en predikto de donita individuo por okupiĝi pri riska konduto, anstataŭ simplaj ŝanĝoj en impulsemo (vidu Figo. 5).

Adoleska konduto plurfoje karakteriziĝis kiel impulsema kaj riska (Steinberg, 2004, 2007), tamen ĉi tiu recenzo de la bilda literaturo sugestas malsamajn neurobiologiajn substratojn kaj malsamajn disvolvajn trajektoriojn por ĉi tiuj kondutoj. Specife, impulsiveco estas asociita kun nematura ventroforma disvolviĝo kaj iom post iom malpliiĝas de infanaĝo ĝis plenaĝeco (Casey, Galvan et al., 2005). La negativa korelacio inter impulsivecoj kaj aĝo en la studo Galvan et al. (2007) plue subtenas ĉi tiun nocion. En kontrasto, risko-asocio estas asociita kun pliigo de akciza agado (Kuhnen & Knutson, 2005; Matthews et al., 2004; Montague & Berns, 2002), tio troigas en adoleskantoj, rilate al infanoj kaj plenkreskuloj (Ernst et al., 2005; Galvan et al., 2006). Tiel adoleskaj elektoj kaj konduto ne povas esti klarigitaj per impulsiveco aŭ longedaŭra disvolviĝo de la antaŭfrontaleca kortekso sole, ĉar infanoj tiam estus antaŭdiritaj esti pli grandaj riskantoj. La trovoj provizas neŭralan bazon de kial iuj adoleskantoj havas pli grandan riskon ol aliaj, sed plue provizas bazon pri kiel adoleska konduto diferencas de infanoj kaj plenkreskuloj en risko-prenado.
Kolektive, ĉi tiuj datumoj sugestas, ke kvankam adoleskantoj kiel grupo estas konsiderataj riskantoj (Ĝardenisto & Steinberg, 2005), iuj adoleskantoj estos pli inklinaj ol aliaj al riska konduto, metante ilin en plej grandan riskon por negativaj rezultoj. Ĉi tiuj trovoj substrekas la gravecon konsideri individuan variablon kiam oni ekzamenas kompleksajn cerbajn-kondutajn rilatojn rilatigitaj kun risko-preno kaj rekompenco-prilaborado en evoluaj populacioj. Plue, ĉi tiuj individuaj kaj evoluaj diferencoj povus helpi klarigi vundeblecon ĉe iuj individuoj al risko-asocio kun uzo de substancoj kaj finfine al toksomanio.
konkludoj
Studoj pri homaj bildoj montras strukturajn kaj funkciajn ŝanĝojn en frontostriaj regionoj (Giedd et al., 1996, 1999; Jernigan et al., 1991; Sowell et al., 1999; por revizio, Casey, Galvan et al., 2005) kiuj ŝajnas paralelaj kreskoj de kognitiva kontrolo kaj memregulado (Casey, Trainor et al., 1997; Luna & Sweeney, 2004; Luna et al., 2001; Rubia et al., 2000; Steinberg, 2004; Vidu ankaŭ Steinberg, 2008, ĉi tiu afero). Ĉi tiuj ŝanĝoj montras movadon en aktivigo de antaŭfrontaj regionoj de disvastiga al pli fokusa varbado tra la tempo (Brown et al., 2005; Bunge et al., 2002; Casey, Trainor et al., 1997; Durston et al., 2006; Moses et al., 2002) kaj levita varbado de subkortikaj regionoj dum adoleskeco (Casey et al., 2002a; Durston et al., 2006; Luna et al., 2001). Kvankam studoj pri neŭroimage ne povas definitive karakterizi la mekanismon de tiaj evoluaj ŝanĝoj, ĉi tiuj ŝanĝoj en volumo kaj strukturo povas reflekti disvolviĝon ene de rafinado de projekcioj al kaj el ĉi tiuj cerbaj regionoj dum maturiĝo sugestas pri agordado de la sistemo kun disvolviĝo.
Kunigitaj, la trovoj ĉi tie sintezitaj indikas, ke pliigita riska konduto en adoleskeco asocias kun malsamaj disvolvaj trajektorioj de subkortika plezuro kaj kortikala regado. Ĉi tiuj evoluaj ŝanĝoj povas pligravigi per individuaj diferencoj en aktiveco de rekompencaj sistemoj. Kvankam adoleskeco estis distingita kiel periodo karakterizata de kondutoj serĉantaj rekompencon kaj riskon (Ĝardenisto & Steinberg, 2005; Spear, 2000) Individuaj diferencoj en neŭralaj respondoj rekompenci, predisponi iujn adoleskantojn al pli multaj riskoj ol aliaj, metante ilin en pli granda risko por negativaj rezultoj. Ĉi tiuj trovoj donas kernan bazon sintezante la diversajn trovojn ligitajn al risko-konduto en adoleskeco kaj komprenante individuajn diferencojn kaj disvolvajn markilojn por inklinoj okupi negativan konduton.
Dankojn
Ĉi tiu laboro estis subtenita parte per subvencioj de la Nacia Instituto pri Drogaj Misuzoj R01 DA18879 kaj la Nacia Instituto pri Mensa Sano 1P50 MH62196.
Referencoj
- Bentin A, Slovic P, Severson H. Psikometria studo de percepto pri risko de adoleskantoj. Ĵurnalo de Adoleskado. 1993;16: 153-168. [PubMed]
- Bjork JM, Knutson B, Fong GW, Caggiano DM, Bennett SM, Hommer DW. Incentive-provokita cerba aktivado en adoleskantoj: Similecoj kaj diferencoj de junaj plenkreskuloj. Ĵurnalo de Neŭrikeco. 2004;24: 1793-1802. [PubMed]
- Bjorklanda DF. La rolo de konceptaj scioj en disvolviĝo de organizo en infana memoro. En: Brainerd CJ, Pressley M, redaktistoj. Bazaj procezoj en memora evoluo: Progreso en esplorado de kognaj disvolviĝoj. Nov-Jorko: Springer-Verlag; 1985 pp 103 – 142.
- Bjorklanda DF. Kiel aĝo-ŝanĝoj en scia bazo kontribuas al disvolviĝo de infana memoro: Interpreta recenzo. Disvolviga Revizio. 1987;7: 93-130.
- Burĝa JP, Goldman-Rakic PS, Rakic P. Synaptogenesis en la antaŭfronta kortiko de rhesusaj simioj. Cerba Kortiko. 1994;4: 78-96. [PubMed]
- Brainerd CJ, Reyna VF. Memora sendependeco kaj memora enmiksiĝo en kogna evoluo. Psikologia Revizio. 1993;100: 42-67. [PubMed]
- Bruna TT, Lugar HM, Coalson RS, Miezin FM, Petersen SE, Schlaggar BL. Evoluaj ŝanĝoj en homa cerba funkcia organizo por vort-generado. Cerba Kortiko. 2005;15: 275-290. [PubMed]
- Bunge SA, Dudukovic NM, Thomason ME, Vaidya CJ, Gabrieli JD. Nematuriga frontala lobo kontribuoj al kognitiva kontrolo en infanoj: Evidentoj de fMRI. Neŭrono. 2002;33: 301-311. [PubMed]
- Kazo R. Validigo de nov-Piagetiana kapabla konstruo. Ofurnalo pri Eksperimenta Infan Psikologio. 1972;14: 287-302.
- Kazo R. Intelekta disvolviĝo: Naskiĝo ĝis plenaĝeco. New York: Akademia Gazetaro; 1985
- Casey BJ, Castellanos FX, Giedd JN, Marsh WL, Hamburger SD, Schubert AB. et al. Impliki dekstran frontostriajn cirkvitojn en respondo-inhibicio kaj atento-deficita / hiperaktiveca malordo. Urnalo de la Usona Akademio de Psikiatrio pri Infanoj kaj Adoleskantoj. 1997;36: 374-383. [PubMed]
- Casey BJ, Galvan A, Hare TA. Ŝanĝoj en cerba funkcia organizo dum kognitiva evoluo. Nuna Opinio en Neurobiologio. 2005;15: 239-244. [PubMed]
- Casey BJ, Giedd JN, Thomas KM. Struktura kaj funkcia cerba evoluo kaj ĝia rilato al kogna evoluo. Biologia Psikologio. 2000a;54: 241-257. [PubMed]
- Casey BJ, Thomas KM, Davidson MC, Kunz K, Franzen PL. Disiigada stria kaj hipokampa funkcio evolue kun stimula-respondo-kongrua tasko. Ĵurnalo de Neŭrikeco. 2002a;22: 8647-8652. [PubMed]
- Casey BJ, Thomas KM, kimra TF, Badgaiyan-RD, Eccard-CH, Jennings JR, kaj aliaj. Disigado de respondo-konflikto, atentelekto, kaj atendado kun funkcia magneta resono. Procedoj de la Nacia Akademio de Sciencoj. 2000b;97: 8728-8733.
- Casey BJ, Tottenham N, Fossella J. Klinikaj, bildigaj, lezaj kaj genetikaj aliroj al modelo de kognitiva kontrolo. Disvolva Psikobiologio. 2002b;40: 237-254. [PubMed]
- Casey BJ, Tottenham N, Liston C, Durston S. Bildigo de la evolua cerbo: Kion ni lernis pri kognitiva disvolviĝo? Tendencoj en Scienca Scienco. 2005;9: 104-110.
- Casey BJ, Trainor RJ, Orendi JL, Schubert AB, Nystrom LE, Giedd JN, et al. Studo pri evolua funkcia MRI de prefrontala aktivado dum plenumado de nerevokebla tasko. Ofurnalo de Cognitive Neuroscience. 1997;9: 835-847.
- Crone E, Donohue S, Honomichl R, Wendelken C, Bunge S. Cerbaj regionoj mediaciante pri fleksebla regulo-uzo dum disvolviĝo. Ĵurnalo de Neŭrikeco. 2006;26: 11239-11247. [PubMed]
- Dempster FN. Rezisto al enmiksiĝo: Evoluaj ŝanĝoj en baza prilaborado. En: Howe ML, Pasnak R, redaktistoj. Aperantaj temoj en kognitiva disvolviĝo Volumo 1: Fundamentoj. Nov-Jorko: Springer; 1993 pp 3 – 27.
- Diamanto A. Disvolviĝo de la kapablo uzi revokon por gvidi agon, kiel indikite de la agado de beboj en AB. Infanevoluo. 1985;56: 868-883. [PubMed]
- Durston S, Davidson MC, Thomas KM, Worden MS, Tottenham N, Martinez A, kaj aliaj. Parametra manipulado de konflikto kaj reago-konkurenco per rapida memkontraŭa okazaĵo rilata fMRI. Neuroimage. 2003;20: 2135-2141. [PubMed]
- Durston S, Davidson MC, Tottenham N, Galvan A, Spicer J, Fossella J, et al. Ŝanĝo de disvastiga al fokusa kortika agado kun disvolviĝo. Evolua Scienco. 2006;1: 18-20. [PubMed]
- Durston S, Hulshoff Pol HE, Casey BJ, Giedd JN, Buitelaar JK, van Engeland H. Anatomia MRI de la evolua homa cerbo: Kion ni lernis? Revuo por Usona Akademio de Infana Adoleska Psikiatrio. 2001;40: 1012-1020.
- Eaton LK, Kinchen S, Ross J, Hawkins J, Harris WA, Lowry R, et al. Gvatado pri junulara riska konduto - Usono, 2005, resumoj pri gvatado. Morbemo kaj Morteco Semajna Raporto. 2006;55: 1-108. [PubMed]
- Eigstio-IM, Zayas V, Mischel W, Shoda Y, Ayduk O, Dadlani MB, et al. Antaŭdirante kognan kontrolon de antaŭlernejo al malfrua adoleskeco kaj juna plenaĝa. Psikologia Scienco. 2006;17: 478-484. [PubMed]
- Ernst M, Nelson EE, Jazbec S, McClure EB, Monk CS, Leibenluft E, et al. Amigdalo kaj kerno accumbens en respondoj al ricevo kaj preterlaso de gajnoj en plenkreskuloj kaj adoleskantoj. Neuroimage. 2005;25: 1279-1291. [PubMed]
- Eshel N, Nelson EE, Blair RJ, Pine DS, Ernst M. Neŭralaj substratoj elekteblaj en plenkreskuloj kaj adoleskantoj: Disvolviĝo de la ventrolateraj antaŭfrontalaj kaj antaŭaj cingulaj kortikoj. Neuropsychologia. 2007;45: 1270-1279. [PMC libera artikolo] [PubMed]
- Flavell JH, Feach DR, Chinsky JM. Spontana verba ripetado en memora tasko kiel funkcio de aĝo. Infanevoluo. 1966;37: 283-299. [PubMed]
- Galvan A, Hare TA, Parra CE, Penn J, Voss H, Glover G, et al. Pli frua evoluo de la akcumbenoj rilate al orbitofronta kortekso povus subteni konduton de risko en adoleskantoj. Ĵurnalo de Neŭrikeco. 2006;26: 6885-6892. [PubMed]
- Galvan A, Hare T, Voss H, Glover G, Casey BJ. Kapti risko kaj adoleska cerbo: Kiu riskas? Evolua Scienco. 2007;10: F8-F14. [PubMed]
- Ĝardenisto M, Steinberg L. Peer-influo sur risko, risko prefero, kaj riska decidiĝo en adoleskeco kaj plenaĝeco: eksperimenta studo. Evolua Psikologio. 2005;41: 625-635. [PubMed]
- Giedd JN. Struktura magneta resono de la adoleska cerbo. Anales de la Akademio de Sciencoj de Novjorko. 2004;1021: 77-85. [PubMed]
- Giedd JN, Blumenthal J, Jeffries NE, Castellanos FX, Liu H, Zijdenbos A, et al. Cerba disvolviĝo dum infanaĝo kaj adoleskeco: longforma MRI-studo. Naturo-Neurokienco. 1999;2: 861-863.
- Giedd JN, Snell JW, Lange N, Rajapakse JC, Casey BJ, Kozuch PL, et al. Kvanta magneta resonanca bildigo de homa cerba disvolviĝo: Aĝoj 4 – 18. Cerba Kortiko. 1996;6: 551-560. [PubMed]
- Gogtay N, Giedd JN, Lusk L, Hayashi KM, Greenstein D, Vaituzis AC, et al. Dinamika mapado de homa kortika evoluo dum infanaĝo tra frua adulto. Proceedings de la Nacia Akademio de Sciencoj de Usono. 2004;101: 8174-8179. [PMC libera artikolo] [PubMed]
- JA Graber, Brooks-Gunn J. Puberteco. En: Blechman EA, Brownell KD, redaktistoj. Kondutisma medicino kaj virinoj ampleksa manlibro. New York, NY: Guilford Press; 1998 pp 51 – 58.
- Leporo TA, Voss HU, Glover GH, Casey BJ. La adoleska cerbo kaj ebla risko por angoro kaj depresio. 2007a Submetite por publikigado.
- Leporo TA, Voss HU, Glover GH, Casey BJ. Konkurenco inter antaŭfrontalaj kaj subkortikaj limuzaj sistemoj bazas emocian reaktivecon dum adoleskeco. 2007b Submetite por publikigado.
- Harnishfeger KK, Bjorkland F. La ontogenio de malhelpaj mekanismoj: Renovigita aliro al kognitiva disvolviĝo. En: Howe ML, Pasnek R, redaktistoj. Emerĝaj temoj pri kogna evoluo. Vol. 1 Nov-Jorko: Springer-Verlag; 1993 pp 28 – 49.
- Hikosaka K, Watanabe M. Prokrasti agadon de orbitaj kaj flankaj prefrontaj neŭronoj de la simio variantaj kun malsamaj rekompencoj. Cerba Kortiko. 2000;10: 263-271. [PubMed]
- Huttenlocher PR. Sinaptika denseco en homa fronta kortekso - Evoluaj ŝanĝoj kaj efikoj de maljuniĝo. Brain-Esploro. 1979;163: 195-205. [PubMed]
- Hyman SE, Malenka RC. Toksomanio kaj cerbo: la neurobiologio de devigado kaj ĝia persisto. Naturaj Revizioj Neŭrokienco. 2001;2: 695-703.
- Jernigan TL, Zisook S, Heaton RK, Moranville JT, Hesselink JR, Braff DL. Anomalioj de magnetaj resonancaj bildigoj en lenticularaj kernoj kaj cerba kortekso en skizofrenio. Arkivoj de enerala Psikiatrio. 1991;48: 811-823.
- Keating DP, Bobbitt BL. Individuaj kaj evoluaj diferencoj en kognaj procezaj komponentoj de mensa kapableco. Infanevoluo. 1978;49: 155-167.
- Kelley AE, Schochet T, Landry C. Anales de la Akademio de Sciencoj de Novjorko. 2004;1021: 27-32. [PubMed]
- Knutson B, Adams CM, Fong GW, Hommer D. Anticipado pri kreskanta mona rekompenco selekte rekrutas kernon accumbens. Ĵurnalo de Neŭrikeco. 2001;21: RC159. [PubMed]
- Kuhnen CM, Knutson B. La neŭra bazo de financa risko. Neŭrono. 2005;47: 763-770. [PubMed]
- Laviola G, Adriani W, Terranova ML, Gerra G. Faktoroj psikobiologiaj de risko por vundebleco al psikosimulantoj en homaj adoleskantoj kaj bestaj modeloj. Revizioj pri Neurokienco kaj Bioportaj. 1999;23: 993-1010. [PubMed]
- Laviola G, Macri S, Morley-Fletcher S, Adriani W. Abstraktaj risko-kondutoj en adoleskaj musoj: Psikobiologiaj determinantoj kaj frua epigenetika influo. Revizioj pri Neurokienco kaj Bioportaj. 2003;27: 19-31. [PubMed]
- Listono C, Watts R, Tottenham N, Davidson MC, Niogi S, Ulug AM, et al. Frontostriatal-mikrostrukturo modulas efikan kontrakton de kognan kontrolon. Cerba Kortiko. 2005;16: 553-560. [PubMed]
- Luna B, Sweeney JA. La apero de kunlabora cerba funkcio: FMRI-studoj pri la evoluo de respondo-inhibicio. Anales de la Akademio de Sciencoj de Novjorko. 2004;1021: 296-309. [PubMed]
- Luno B, Thulborn KR, Munoz DP, Merriam EP, Garver KE, Minshew NJ, et al. Maturiĝo de vaste distribuita cerba funkcio servas kognan disvolviĝon. Neuroimage. 2001;13: 786-793. [PubMed]
- Matthews SC, et al. Selektema aktivigo de la kerno akcenta dum prenado de riskoj. Neuroreporto. 2004;15: 2123-2127. [PubMed]
- JC de majo, Delgado MR, Dahl RE, Stenger VA, Ryan ND, Fiez JA, et al. Event-rilata funkcia magneta resono al rekompensaj cerbaj cirkvitoj en infanoj kaj adoleskantoj. Biologia Psikiatrio 2004;55: 359-366. [PubMed]
- McClure SM, Laibson DI, Loewenstein G, Cohen JD. Apartaj neŭralaj sistemoj taksas tujajn monajn rekompencojn. Scienco. 2004;306: 503-507. [PubMed]
- Miller EK, Cohen JD. Teorio integra de la funkcio de la krozo prefrontal. Jara Revizio de Neŭrikeco. 2001;24: 167-202.
- Mischel W, Shoda Y, Rodriguez MI. Prokrasto de kontentigo ĉe infanoj. Scienco. 1989;244: 933-938. [PubMed]
- Monk CS, McClure EB, Nelson EE, Zarahn E, Bilder RM, Leibenluft E, et al. Adoleska nematureco en atentema cerba engaĝiĝo al emociaj vizaĝaj esprimoj. Neuroimage. 2003;20: 420-428. [PubMed]
- Montague PR, Berns GS. Neŭra ekonomio kaj la biologiaj substratoj de taksado. Neŭrono. 2002;36: 265-284. [PubMed]
- Moses P, Roe K, Buxton RB, Wong EC, Frank LR, Stiles J. Funkcia MRI de monda kaj loka pretigo ĉe infanoj. Neuroimage. 2002;16: 415-424. [PubMed]
- Munakata Y, Yerys BE. Ĉio kune nun: Kiam la disiĝoj inter scio kaj ago malaperas. Psikologia scienco. 2001;12: 335-337.
- Nagy Z, Westerberg H, Klingberg T. Maturiĝo de blanka materio estas asociita kun la disvolviĝo de kognaj funkcioj dum infanaĝo. Ofurnalo de Cognitive Neuroscience. 2004;16: 1227-1233. [PubMed]
- O'Doherty J, Kringelbach ML, Rolls ET, Hornak J, Andrews C. Abstraktaj rekompencoj kaj punaj reprezentadoj en la homa orbitofrontala kortekso. Naturo Neŭrosko. 2001;4: 95-102. [PubMed]
- O'Doherty JP. Rekompencaj reprezentadoj kaj rekompenco-lernado en la homa cerbo: Enrigardoj de neŭromaĝado. Aktualaj Opinioj en Neurobiologio. 2004;14: 769-776.
- Pascual-Leone JA. Matematika modelo por transiro en la evoluaj stadioj de Piaget. Acta Psychologica. 1970;32: 301-345.
- Pecina S, Cagniard B, Berridge KC, Aldridge JW, Zhuang X. Hiperdopaminergiaj mutaciaj musoj havas pli altan "deziron" sed ne "plaĉantan" por dolĉaj rekompencoj. Ĵurnalo de Neŭrikeco. 2003;23: 9395-9402. [PubMed]
- Reyna VF, Farley F. Risko kaj racieco en adoleska decidiĝo: Implikaĵoj por teorio, praktiko, kaj publika politiko. Psikologia Scienco en Publika Intereso. 2006;7: 1-44.
- Rosenberg DR, Lewis DA. Postnata maturiĝo de la dopaminerin-inervo de simio-antaŭfrontaj kaj motoraj kurboj: tirozina hidroxilasa imuno-histokemia analizo. La urnalo de Kompara Neŭrologio. 1995;358: 383-400. [PubMed]
- Rubia K, Overmeyer S, Taylor E, Brammer M, Williams SC, Simmons A, et al. Funkcia frontaligo kun aĝo: Mapado de neŭroevoluaj trajektorioj kun fMRI. Revizioj pri Neurokienco kaj Bioportaj. 2000;24: 13-19. [PubMed]
- Schlaggar BL, Bruna TT, Lugar HM, Visscher KM, Miezin FM, Petersen SE. Funkciaj neuroanatomiaj diferencoj inter plenkreskuloj kaj lernejaj infanoj en la prilaborado de ununuraj vortoj. Scienco. 2002;296: 1476-1479. [PubMed]
- Schultz W. Kondutismaj teorioj kaj neŭrofiziologio de rekompenco. Jara Revizioj de Psikologio. 2006;57: 87-115.
- Sowell ER, Peterson BS, Thompson-ĉefministro, Welcome SE, Henkenius AL, Toga AW. Mapante kortikan ŝanĝon tra la homa vivdaŭro. Naturo-Neurokienco. 2003;6: 309-315.
- Sowell ER, Thompson-ĉefministro, Holmes CJ, Jernigan TL, Toga AW. Indico en vivo por post-adoleska cerba maturiĝo en frontaj kaj striataj regionoj. Naturo-Neurokienco. 1999;2: 859-861.
- Sowell ER, Thompson-gazetaro, Toga AW. Mapado de ŝanĝoj en la homa kortekso tra la tuta tempo. Neurocientisto. 2004;10: 372-392. [PubMed]
- Lanco LP. La adoleskaj cerbaj kaj aĝaj kondutaj manifestacioj. Revizioj pri Neurokienco kaj Bioportaj. 2000;24: 417-463. [PubMed]
- Steinberg L. Risko-prenado en adoleskeco: Kio ŝanĝiĝas, kaj kial? Anales de la Akademio de Sciencoj de Novjorko. 2004;1021: 51-58. [PubMed]
- Steinberg L. Risko-prenado en adoleskeco: Novaj perspektivoj de cerba kaj kondutisma scienco. Aktualaj Direktoj en Psikologia Scienco. 2007;16: 55-59.
- Steinberg L. Perspektivo pri socia neŭoscienco pri adoleska risko-preno. Disvolviga Revizio. 2008;28: 78-106. [PMC libera artikolo] [PubMed]
- Tamm L, Menon V, Reiss AL. Maturiĝo de cerba funkcio asociita kun reaga inhibo. Urnalo de la Usona Akademio de Psikiatrio pri Infanoj kaj Adoleskantoj. 2002;41: 1231-1238. [PubMed]
- Thomas KM, Hunt RH, Vizueta N, Sommer T, Durston S, Yang Y, et al. Evidenteco de evoluaj diferencoj en implicita sekvenca lernado: FMRI-studo de infanoj kaj plenkreskuloj. Ofurnalo de Cognitive Neuroscience. 2004;16: 1339-1351. [PubMed]
- Turkeltaub PE, Gareau L, Floroj DL, Zeffiro TA, Eden GF. Disvolviĝo de nervaj mekanismoj por legado. Naturo-Neurokienco. 2003;6: 767-773.
- Volkow ND, Li TK. Drogodependeco: La neŭrobiologio de konduto malaperis. Naturaj Revizioj Neŭrokienco. 2004;5: 963-970.
- Yurgelun-Todd D. Emociaj kaj kognaj ŝanĝoj dum adoleskeco. Nuna Opinio en Neurobiologio. 2007;17: 251-257. [PubMed]
- Zald DH, Boileau I, El-Dearedy W, Gunn R, McGlone F, Dichter GS, et al. Transdono de dopamina en la homa striato dum taskoj de monaj rekompencoj. Ĵurnalo de Neŭrikeco. 2004;24: 4105-4112. [PubMed]
