Rimarkoj: Ĉi tiu recenzo estis produktita de la estro de NIDA, Nora Volkow, kaj ŝia teamo. Lasas malmultan dubon, ke kemiaj toksomanioj kaj kondutaj toksomanioj dividas la samajn aŭ similajn mekanismojn kaj neŭrajn cirkvitojn. Ĉi tio havas perfektan sencon, ĉar kemiaj toksomanioj kidnapas la neŭrajn cirkvitojn por ligado, sekso kaj manĝado. Ĉar sekso liberigas duoble pli da dopamino ol manĝi vian plej ŝatatan manĝaĵon, kaj uzanto de porno povas teni dopaminon levita dum horoj, estas freneze proponi, ke dependeco de porno ne povas ekzisti.
Curr Top Behav Neurosci. 2011 Okt 21.
Volkow ND, Wang GJ, Fowler JS, Tomasi D, Baler R.
fonto
Nacia Instituto pri Drogasuzo, 6001 Executive Boulevard 6001, Salono 5274, Bethesda, MD, 20892, Usono, [retpoŝte protektita].
abstrakta
Ambaŭ drogmanio kaj obezeco povas esti difinitaj kiel malordoj, en kiuj la senvalora valoro de unu speco de rekompenco (drogoj kaj manĝaĵo respektive) fariĝas nenormale plibonigita rilate al kaj koste de aliaj. Ĉi tiu modelo kongruas kun la fakto, ke kaj drogoj kaj manĝaĵoj havas potencajn plifortigajn efikojn - parte mediaciitajn de dopamina pliigoj en la limba sistemo -, kiuj, en iuj cirkonstancoj aŭ en vundeblaj individuoj, povus superforti la homeostatikajn kontrolajn mekanismojn de la cerbo. Tiaj paraleloj generis signifan intereson kompreni la dividitajn vundeblecojn kaj trajektoriojn inter toksomanio kaj obezeco. Nun, malkovroj pri cerbaj bildoj komencis malkovri oftajn trajtojn inter ĉi tiuj du kondiĉoj kaj delimigi iujn el la interkovrantaj cerbaj cirkvitoj kies disfunkcioj eble klarigos stereotipajn kaj rilatajn kondutajn deficitojn en homaj subjektoj. THese-rezultoj sugestas, ke kaj obesaj kaj drogemuloj, suferas mankojn de dopaminergiaj vojoj, kiuj reguligas neŭronajn sistemojn asociitajn ne nur kun rekompenca sentemo kaj instiga instigo, sed ankaŭ kun kondiĉado (memoro / lernado), impulsa kontrolo (kondutisma inhibicio), streĉa reaktiveco , kaj interkonceptema konscio. Ĉi tie ni kunmetas trovojn ĉefe el tomografiaj emisioj de pozitronoj, kiuj elmontras la rolon de dopamino en drogmanio kaj en obezeco, kaj proponas ĝisdatigitan laboran modelon por helpi identigi kuracajn strategiojn, kiuj povas profiti ambaŭ de ĉi tiuj kondiĉoj.
1 fono
2 La Rolo de Dopamino en Akra Rekompenco al Drogoj kaj Manĝaĵoj
3 Bildigo DA en Response to Drogoj kaj al Kondiĉigitaj Kondiĉoj en toksomanio
4 La Efiko de Malfunkcio en Inhibitora Kontrolo
5 Partopreno de Motivaj Cirkvitoj
6 Partopreno de Interoceptive Circuitry
7 La Cirkvitado de Aversio
8 Patologia Droga kaj Manĝaĵa Rekompenco: Ĝisdatigita Labora Modelo
1 fono
Dopamina (DA) estas konsiderata ŝlosilo por la rekompencaj efikoj de naturaj kaj drogaj rekompencoj. Tamen ĝia rolo en la perdo de kontrolo kaj devigaj kondutoj, kiuj estas asociitaj kun toksomanio kaj obezeco, estas multe malpli klaraj. PET-studoj ludis gravegan rolon en karakterizado de la rolo de la cerbaj DA-sistemoj en toksomanio (aldone al sia rolo en drog-rekompenco) kaj en obezeco. Efektive, drogoj de misuzo (inkluzive alkoholon) estas konsumitaj de homoj aŭ mem-administritaj de laboratoriaj bestoj, ĉar ili estas imense rekompencaj, efiko, kiu estas mediata per iliaj plibonigaj proprietoj de DA en la mezolimbia sistemo (Saĝa 2009). Hŝuldanto, en la kazo de toksomanio, bildaj studoj malkaŝis, ke la malordo efikas ne nur la DA-rekompenca cirkvito sed ankaŭ aliajn DA-vojojn implikitajn en modulado de kondiĉado / kutimoj, instigo, kaj plenumaj funkcioj (inhibicia kontrolo, saleca atribuo, kaj decido. -funkcio), kaj ke DA-deficitoj ankaŭ povas partopreni la plibonigitan streĉan reaktivecon kaj interrompon de interkomprenebla konscio asociita kun toksomanio. Klinikoj kaj klinikaj studoj ankaŭ malkaŝis aliajn neurotransmisilojn (kaj neuropeptidojn) kiuj ludas gravajn rolojn en rekompenco kaj toksomanio de drogoj. (t.e., cannabinoides, opioidoj) kaj estas intime implikitaj en la neŭroplastikaj ŝanĝoj, kiuj sekvas ripetan uzon de drogoj (t.e., glutamato, opioidoj, GABA, liberiganta faktoro de kortikotropinoj). La glutamatergia sistemo estas precipe elstara tiurilate ĉar ĝi mediacias la interrompojn tiel en longtempa potenco kaj longtempa depresio, kiuj estis observitaj en bestaj modeloj de kronika administrado de drogoj. (Thomas et al. 2008). Recenzoj pri ĉi tiuj pliaj sistemoj troveblas aliloke (Kalivas) 2009; Koob 1992).
Ĉar drogoj aktivigas la samajn rekompencajn sistemojn, kiuj subestas manĝaĵan rekompencon, estas ne tute neatendite, ke, ĝenerale, cerbaj bildaj studoj subtenis la nocion, ke mankoj en DA-modulitaj cirkvitoj ankaŭ implikiĝas en patologiaj, devigaj manĝantaj kondutoj.. Manĝaĵaj kvereloj, kiel drogaj manieroj, pliigas striitan eksterĉelan DA kaj kaŭzas la motivon okupiĝi pri la kondutoj necesaj por aĉeti kaj manĝi la manĝaĵon, donante evidentecon pri la implikiĝo de DA ne nur en manĝaĵa rekompenco sed ankaŭ en la ne-hedonaj motivaj ecoj de manĝaĵo (t.e., kaloriaj postuloj) kaj la malkresko de inhibitora kontrolo vidita en deviga manĝado (Avena et al. 2008; Volkow et al. 2008a).
Ĉi tie, ni revizias trovojn de bildaj studoj, kiuj specife fokusas sur la interkovroj en la cerbaj cirkvitoj, kiuj estas malordigitaj en obesidad kaj en drogmanio. Ĝi valoras memori, tamen, ke la regulado de manĝaĵa konsumado estas multe pli kompleksa ol la regulado de konsumado de drogoj. Ĉi-lasta estas ĉefe mediatita per la rekompencaj efikoj de drogoj, dum la unua estas modulita ne nur de ĝiaj rekompencaj efikoj (hedonaj faktoroj) sed ankaŭ de multnombraj periferiaj kaj centraj faktoroj, kiuj sentas nutrajn postulojn en la korpo necesaj por postvivado (homeostataj faktoroj). Interese, estas kreskantaj evidentaĵoj, ke homeostataj faktoroj (ekz. Insulino, leptino, ghrelin) modulas la manĝan konsumon parte pliigante aŭ malpliigante la sentivecon de cerbaj rekompencaj cirkvitoj al manĝaj stimuloj (Volkow et al. 2011a).
2 La Rolo de Dopamino en Akra Rekompenco al Drogoj kaj Manĝaĵoj
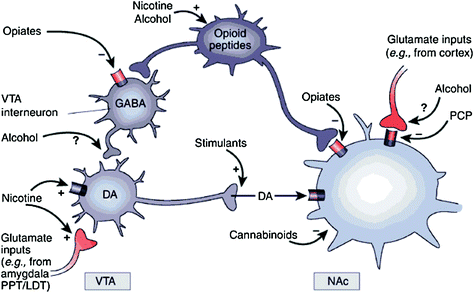
Ĉu rekte aŭ nerekte, ĉiuj toksomaniulaj drogoj montras kapablon pliigi DA en nucleus accumbens (NAc) per specifaj interagoj kun malsamaj molekulaj celoj (Nestler 2004) (Fig. 1). La mezolimbic DA-vojo [DA-ĉeloj en ventrala tegmentala areo (VTA) kiu projektas en la NAc] ŝajnas esti kerna por drog-rekompenco (Saĝa 2009). Tamen, kiel priskribite sube, aliaj DA-vojoj [mesostriatal (DA-ĉeloj en substantia nigra projekcianta en dorsan striatumon) kaj mezocortikaj (DA-ĉeloj en VTA projektantaj en frontan kortekson)] ankaŭ kontribuas al drog-rekompenco kaj toksomanio (Saĝa 2009). Entute ŝajnas, ke la rekompencaj kaj kondiĉigaj efikoj de drogoj estas ĉefe kaŭzitaj de fazo de DA-ĉela pafo, kio kondukas al grandaj kaj transiraj DA pliiĝoj. En kontrasto, la malsupreniraj ŝanĝoj en plenuma funkcio, kiuj okazas en toksomanio, estas ligitaj kun ŝanĝoj en tona DA-pafado kaj rezultigas pli malaltajn sed pli stabilajn DA-nivelojn (Grace 2000; Wanat et al. 2009). Ĉi tio siavice atentigas pri la D1-receptoroj (D1R), kiuj estas malaltaj afinecaj DA-riceviloj, kiuj stimulas ciklan AMP-signaladon, ĉar ili estas implikitaj tiel en akra drog-rekompenco kiel en kondiĉado, ĉar ĉi tiuj estas asociitaj kun la altaj DA-koncentriĝoj necesaj. stimuli D1R. En kontrasto, D2Rs, kiuj malhelpas ciklan AMP-signaladon, estas stimulitaj de kaj fazaj kaj tonikaj DA. Rimarku, ke pro la manko de specifaj radiotraceroj por la PET-bildigo de DA-riceviloj de la tipoj D1, D3, D4 kaj D5, plej multaj studoj pri la efikoj de drogoj de misuzo kaj toksomanio en la homa cerbo temigis D2Rojn.
Figo. 1 Drogoj de misuzo agas sur la rekompenco kaj akcesoraj cirkvitoj per malsamaj mekanismoj, tamen ili ĉiuj kondukas al similaj dopaminergiaj efikoj en la VTA kaj NAc. Tiel, stimulantoj akcelas senakcelan DA rekte, dum opiatoj faras tion per malaltigo de la inhibicia tono de GABAergic-interneuronoj sur DA-signalado ambaŭ en la VTA aŭ en tiam NAc. Dum la mekanismoj de aliaj drogoj misuzo estas malpli klaraj, estas evidentaĵoj sugestantaj, ke nikotino povas aktivigi VTA DA rekte tra nikotinaj acetilkolina ricevilo (nAChR) sur tiuj neŭronoj kaj nerekte per stimulado de ĝiaj riceviloj sur glutamatergaj nervaj fina stacioj, kiuj innervas la DA-ĉelojn. Alkoholo ŝajnas malhelpi GABAergic-finaĵojn en VTA, kondukante al malinhibicio de neŭronoj DA en la VTA. Kanabinoidoj agas, interalie, per la aktivigo de CB1-receptoroj sur nervaj finaĵoj glutamatergaj kaj GABAergic en la NAc, kaj sur la neŭronoj de NAc mem. Fenicididino (PCP) povas agi inhibante postsinaptajn NMDA-glutamatajn ricevilojn en la NAc. Krome, estas iuj evidentaĵoj sugestante, ke nikotino kaj alkoholo povus interagi ankaŭ kun endogenaj opioidaj kaj cannabinoidaj vojoj (ne montritaj). PPT / LDT, peduncular pontine tegmentum / latera dorsal tegmentum. Reproduktita kun permeso Nestler (2005)
En homoj, PET-studoj montris, ke pluraj drogoj [stimuliloj (Drevets et al. 2001; Volkow et al. 1999b), nikotino (Brody et al. 2009), alkoholo (Boileau et al. 2003), kaj marijuuano (Bossong et al. 2009)] pliigi DA en dorsal kaj ventra striatum (kie la NAc situas). Ĉi tiuj studoj utiligas plurajn radiotracerojn, kiel [11C] raclopride, kiu ligiĝas al D2R sed nur kiam ĉi tiuj ne estas ligantaj endogenan DA (neokupita), kiu en bazaj kondiĉoj respondas al 85-90% de la striata D2R (Abi-Dargham et al. 1998). Tiel, komparo de [11C] raclopride-ligado post placebo kaj post administrado de drogoj povas helpi nin taksi la malpliiĝojn de la havebleco de D2R induktita de la drogo (aŭ aliaj stimuloj, kiuj povas pliigi DA). Ĉi tiuj malpliiĝas en [11C] raclopride-ligado estas proporcia al la DA-pliiĝoj (Breier et al. 1997). Ĉi tiuj studoj pruvis, ke la kresko de DA-induktita de drogoj en striato estas proporcia al la intenseco de la subjektiva sperto de eŭforio aŭ "alta" [vidu recenzon (Volkow et al. 2009a)] (Fig. 2).
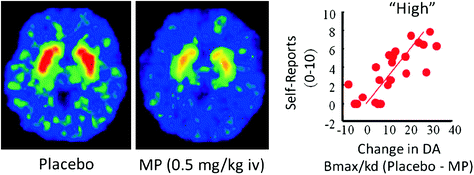
Figo. 2 Efikoj de intravena metilfenidato (MP) en raclopride-ligado kaj rilato inter striatal DA pliigoj induktitaj de MP en la striatumo kaj la mem-raportoj de "alta". Modifita de Volkow et al. (1999b)
PET-studoj ankaŭ malkaŝis klaran, rektan rilaton inter la farmacokinetika profilo de drogo (t.e., la rapideco kun kiu ĝi eniras kaj eliras la cerbon) kaj ĝiaj plifortigaj efikoj. Specife, ju pli rapida drogo atingas pintajn nivelojn en la cerbo, des pli intensa estas la "alta" (Volkow et al. 2009a). Ekzemple, por ekvivalenta nivelo de kokaino atinganta la cerbon (taksita per PET), kiam kokaino eniris la cerbon rapide (fumigita aŭ iv-administrado), ĝi estigis pli intensan "alton" ol kiam ĝi eniris pli malrapide (snorĝis) (Volkow et al. 2000). Ĉi tio konformas al preklinikaj studoj, kiuj montras similan korelacion inter la farmacokinetika profilo de drogo kaj ĝiaj plifortigaj proprecoj (Balster kaj Schuster) 1973). MiNe estas racie hipotezi, ke tiaj subitaj kaj grandaj DA pliiĝoj, kaŭzitaj de drogoj misuzoj, povas imiti la rapidajn kaj grandajn DA-pliiĝojn, kiuj rezultas de fazo de DA-pafo asociitaj en la cerbo kun la prilaborado de informoj pri rekompenco kaj saŭco. (Schultz 2010). Tiaj kreskoj de DA-induktitaj drogoj en la NAc povas esti necesaj por toksomanio, sed la fakto ke ili okazas ankaŭ ĉe ne-toksomaniuloj indikas, ke ili estas nesufiĉaj por klarigi la impulsan kaj devigan drogan uzon de toksomanio.
Nun estas evidenteco, ke kompareblaj dopaminergiaj respondoj estas ligitaj kun manĝaĵa rekompenco kaj ke ĉi tiuj mekanismoj ankaŭ povas ludi rolon en troa manĝaĵa konsumo kaj obezeco. Estas sciate, ke iuj nutraĵoj, precipe tiuj riĉaj en sukeroj kaj grasoj, potence rekompencas (Lenoir et al. 2007). Alta-kaloria manĝaĵo povas antaŭenigi tro-manĝadon (manĝado senkonektita de energiaj bezonoj) kaj deĉenigi lernitajn asociojn inter la stimulo kaj la rekompenco (kondiĉigo). En evolua termino, ĉi tiu propraĵo de plaĉaj manĝaĵoj kutimis esti avantaĝa en medioj, kie manĝaĵfontoj estis malabundaj kaj / aŭ nefidindaj, ĉar ĝi certigis, ke manĝaĵoj manĝas kiam disponeblas, ebligante konservi energion en la korpo (kiel grasa) por estonta uzo. . Bedaŭrinde, en socioj kiel la nia, kie manĝo estas abunda kaj konstante havebla, ĉi tiu adaptado fariĝis pasivo.
Pluraj neurotransmisiloj, inkluzive de DA, kanabinoidoj, opioidoj, kaj serotonino, same kiel hormonoj kaj neuropeptidoj implikitaj en homeostatika regulado de manĝaĵa konsumado, kiel insulino, oreksino, leptino kaj ghrenlin, estis implikitaj en la rekompencaj efikoj de manĝaĵo (Atkinson 2008; Cason et al. 2010; Cota et al. 2006). De ĉi tiuj, DA estis la plej detale esplorita kaj estas la plej karakterizita. Eksperimentoj en ronĝuloj montris, ke post unua eksponiĝo al manĝaĵa rekompenco, la pafo de DA-neŭronoj en la VTA pliiĝas kun rezulta kresko de liberigo de DA en NAc (Norgren et al. 2006). Simile, ĉe sanaj normal-pezaj homaj subjektoj, la ingesta gusto de manĝaĵo pruviĝis liberigi DA en la dorsa striatumo proporcie al la taksoj de agrabla manĝo (Malgranda et al. 2003) (Fig. 3). Tamen, kaj kiel videblas en studoj kun drogaj fitraktantoj, la kresko de la striatala DA-induktita de manĝaĵo sole ne povas klarigi la diferencon inter normala konsumado de manĝaĵoj kaj troa deviga manĝaĵa konsumo, ĉar ĉi tiuj okazas ankaŭ ĉe sanaj individuoj, kiuj ne manĝas troe. Tiel, kiel okazas kun toksomanio, malsuprenirantaj adaptoj probable implikas la perdon de kontrolo pri manĝaĵa konsumado.
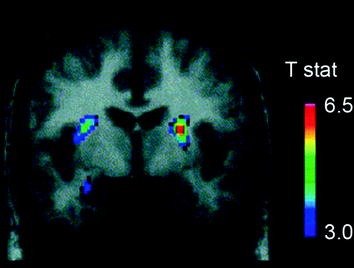
Figo. 3 Dopamina liberigo induktita de nutrado. Korona sekcio el T-mapo de statistike signifaj reduktoj de [11C] liganta potencialo (BP) de raclopride post nutrado. La kolora stango reprezentas la t statistikajn valorojn. (Reproduktita kun permeso Malgranda et al. 2003)
3 Bildigo DA en Response to Drogoj kaj al Kondiĉigitaj Kondiĉoj en toksomanio
La rolo de DA en plifortigado estas pli kompleksa ol nur kodigo por rekompenco (heda plezuro); ekzemple, stimuloj, kiuj induktas rapidajn kaj grandajn DA-pligrandigojn, ankaŭ deĉenigas kondiĉitajn respondojn kaj provokas instigan instigon por akiri ilin (Owesson-White et al. 2009). Ĉi tio gravas ĉar per procezo de kondiĉado, neŭtralaj stimuloj ligitaj al la plifortigilo (ĉu natura aŭ droga plifortigilo) akiras la kapablon mem pliigi DA en striatumo (inkluzive NAc) antaŭ atendo de la rekompenco, tiel engendante. forta instigo serĉi la drogon (Owesson-White et al. 2009). Tamen, senkompata rekompenco kaj kondiĉado de mekanismoj en la procezo de toksomanio estas pli malfacila ol por konsumado de manĝaĵoj ĉar drogoj de misuzo, per siaj farmacologiaj efikoj, rekte aktivigas DA-neŭronojn (t.e., nikotino) aŭ pliigas DA-liberigon (t.e., amfetaminon).
Cerbaj bildigaj studoj, kiuj komparis la kreskojn de DA induktitaj de la stimula drog-metilfenidato (MP) aŭ amfetamino (AMPH) inter subjektoj kun kokaino kontraŭ kontroloj, montris markitan atenuon de kreskoj de MP aŭ AMPH-induktita de DA en striatum (50% pli malalta en detoxifitaj fitraktantoj) kaj 80% en aktivaj fitraktantoj) kaj pli malaltaj mem-raportoj pri la rekompencaj efikoj de la drogo relative al ne-drogaj misuzoj (Martinez et al. 2007; Volkow et al. 1997) (Fig. 4). Ĉi tio surpriziĝis pro tio ke MP kaj AMPH estas farmacologie similaj al kokaino kaj metamfetamino respektive, kaj drogaj fitraktantoj ne povas distingi inter ili kiam ili estas administritaj intravenan. Ĉar oni markis la markitajn reduktojn de la drog-induktitaj DA-kreskoj, ĉu la kokainaj fitraktantoj estis aŭ ne detoxifitaj, tio indikas, ke la stato de retiriĝo ne estas konfuzanta faktoro (Volkow et al. 2011b). Ĉi tiuj kaj rilataj rezultoj (Volkow et al. 2009a) estas konformaj al la hipotezo, ke la hedonika respondo fariĝas manka ĉe drogemuloj, kaj plue fortigas la nocion, ke la akraj farmakologiaj plibonigaj efikoj de la drogo en NAc ne povas klarigi per si mem la pliigitan motivon konsumi ilin.
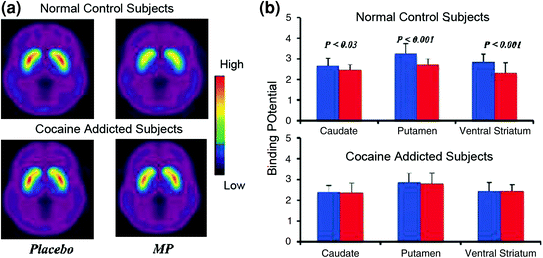
Figo. 4 DA ŝanĝoj induktitaj de iv MP en kontroloj kaj en aktivaj toksomaniaj subjektoj. Duona bildoj ne aranĝeblaj (BPND) de [11C] racloprido en aktivaj toksomaniulaj subjektoj (n = 19) kaj en kontroloj (n = 24) testitaj post placebo kaj post iv MP. b D2R-havebleco (BPND) en kaŭdato, putameno kaj ventrala striatumo post placebo (blua) kaj post MP (ruĝa) en kontroloj kaj en kokain-toksomaniuloj. MP reduktis D2R en kontroloj sed ne en kokain-toksomaniuloj. Atentu, ke kokainaj misuzantoj montras ambaŭ malpliiĝojn de baz-striatala havebleco de D2R (placebo-mezuro) kaj malkreskojn de DA-liberigo kiam donita iv MP (mezurita kiel malkreskoj en D2R-havebleco de baslinio). Kvankam oni povus pridubi la mezuron, en kiu la malalta striatala D2R-havebleco en kokain-toksomania subjekto limigas la kapablon detekti pliajn malpliiĝojn de MP, la fakto, ke koka-toksomaniuloj montras reduktojn en la havebleco de D2R kiam eksponitaj al kokainaj indikoj indikas, ke la atenuitaj efikoj. de MP sur [11C] raclopride-liganta reflekto malpliigis DA-liberigon. Reproduktita kun permeso (Volkow et al. 1997; Wang et al. 2010)
La respondo de VTA DA neŭronoj al rekompencaj stimuloj ŝanĝiĝas kun ripetita ekspozicio.
Dum DA-ĉeloj ekbruliĝas al la unua ekspozicio al nova rekompenco, ripetita ekspozicio al DA kaŭzas, ke la neŭronoj ĉesu pafi ĉe konsumo kaj ekbruligas anstataŭe kiam ili estas elmontritaj al stimuloj antaŭdiraj al la rekompenco. (Schultz et al. 1997). Ĉi tio probable subfosas la rolon de DA en lernado kaj kondiĉado. Efektive, drog-indika fazo DA signalado povas eventuale eligi neŭroadaptojn en akciaj cirkvitoj, kiuj rilatas al kutima formado kaj kondutisma kondiĉo. These-ŝanĝoj estas ĉefe induktitaj de D1R-signalado kaj sinaptaj ŝanĝoj en glutamataj modulitaj NMDA kaj AMPA-riceviloj (Luscher kaj Malenka 2011; Zweifel et al. 2009). La varbado de ĉi tiuj cirkvitoj estas grava en la progresado de malsano ĉar la sekvaj kondiĉitaj respondoj helpas klarigi la intensan deziron de la drogo (avido) kaj la devigan uzon, kiu okazas kiam toksomaniuloj submetiĝas al drogoj. Ĉi tiu hipotezo konformas al sendependaj observoj (Volkow et al. 2006b; Wong et al. 2006) kiu montras la potencon de koka-asociaj kuraj eksponoj por levi DA-nivelojn en la dorsa striatumo kaj deĉenigi samtempan kreskon de la subjektiva sperto de avido ĉe detoxigita kokainuzo (Fig. 5). Ĉar la dorsstria strato ludas rolon en lernado de kutimoj (Belin et al. 2009; Yin et al. 2004), la asocio verŝajne reflektas la plifortigon de kutimoj dum progresas kronikeco de toksomanio. Ĉi tio sugestas, ke baza interrompo en toksomanio povus rilati al la kaŭzitaj respondoj de la kaŭzita de DA, kiuj rezultas en kutimoj kondukantaj al intensa avido kaj deviga konsumado de drogoj. Interese, dum aktive uzado de kokain-toksomaniaj subjektoj, la DA-pliiĝoj deĉenigitaj per kondiĉitaj resendoj ŝajnas esti eĉ pli grandaj ol tiuj produktitaj de la stimulanta drogo mem kiel taksite en du apartaj grupoj de subjektoj (Volkow et al. 2011b, 2006b), sugestante, ke kondiĉitaj respondoj povas instigi la DA-signaladon, kiu konservas la motivon preni la drogon eĉ kiam ĝiaj farmakologiaj efikoj ŝajnas esti reduktitaj. Tiel, kvankam drogoj povas komence indiki sentojn de tuja rekompenco per liberigo de DA en la ventra striato, kun ripeta uzo, kaj dum kutimo disvolviĝas, ŝajnas esti transiro de la drogo al la kondiĉita stimulo. Laŭ studoj en laboratorio-bestoj, glutamatergaj projekcioj de prefrontal-kortekso kaj de amigdala en VTA / SN kaj NAc mediacias ĉi tiujn kondiĉitajn respondojn (Kalivas 2009). Tiel, la nura antaŭdiro de rekompenco eventuale fariĝos la rekompenco, kiu motivas la konduton necesan por konsumo de drogoj (aŭ manĝaĵoj).
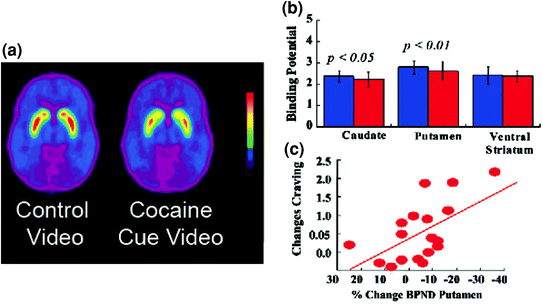
Figo. 5 DA ŝanĝoj induktitaj de kondiĉitaj resumoj en aktivaj toksomaniulaj subjektoj. Duona neplenumebla bindebla potencialo (BPND) bildoj de [11C] raclopride en kokain-toksomaniaj subjektoj (n = 17) testis dum spektado de neŭtrala video (naturscenoj) kaj dum spektado de kokaino-vidbendoj (subjektoj administrantaj kokainon). b D2R-havebleco (BPND) en kaŭdato, putameno kaj ventrala striato por la neŭtrala video (blua) kaj la kokain-cues-video (ruĝa). La kokainaj limoj malpliiĝis D2R en kaŭdato kaj putameno. c Korelacioj inter ŝanĝoj en D2R (reflektaj kreskoj de DA) kaj mem raportoj pri kokainaj avidoj induktitaj de la video kun kokaino. Modifita de ref. (Volkow et al. 2006b)
INerestime, ĉi tiu speco de funkcia "ŝaltilo" ankaŭ estis raportita pri naturaj plifortigiloj, kiuj probable indu ekvivalentan kaj laŭgradan ŝanĝon en pliigoj de DA, de ventralo ĝis pli dorsaj regionoj de la striatumo dum la transiro de nova stimulo nekonate. rekompencante tiun de la rilataj asertoj, kiuj antaŭdiras ĝin. Ĉi tiu transiro estas farita per DA-signalado, kiu ŝajnas kodi "rekompencon-antaŭdiro-eraro" (Schultz 2010). La vastaj glutamatergaj aferentoj al DA-neŭronoj el regionoj implikitaj en prilaborado de sensoraj (insulaj aŭ primaraj gustaj kortekso), homeostatikaj (hipotalamo), rekompenco (NAc), emociaj (amigdala kaj hipokampo), kaj multimodaj (orbitofrontala kortekso por atingo de saleco) , modulas sian aktivecon en respondo al rekompencoj kaj al kondiĉitaj signoj (Geisler kaj Wise 2008). Pli specife, projekcioj de la amigdala kaj la orbitofrontala kortekso (OFC) al DA-neŭronoj kaj al NAc estas engaĝitaj pri kondiĉitaj respondoj al manĝaĵo (Petrovich 2010). Efektive, bildstudaj studoj montris, ke kiam oni ne petis viran subjekton malhelpi sian avidon pri manĝaĵo - dum ili estis eksponitaj al manĝaĵoj, ili montris malpliiĝan metabolan agadon en amigdala kaj OFC (same kiel en hipokampo), insula kaj striatum, kaj ke la malpliiĝoj de OFC estis asociitaj kun reduktoj en avido de manĝaĵoj (Wang et al. 2009). Simila inhibicio de la metabola agado en la OFC (kaj ankaŭ en NAc) estis observita en la konsumantoj de kokaino kiam oni petis ilin inhibici avidojn de drogoj post ekspozicio al kokainaj manieroj (Volkow et al. 2009b).
Ankoraŭ, la apero de tiaj potencaj malhelpaj avidoj, kiuj ankaŭ por manĝaĵo okazas en sanaj individuoj, kiuj ne manĝas tro, ne estus tiel devastaj, se ili ne kuplus kun kreskantaj deficitoj en la kapablo de cerbo malhelpi malbonkondutajn kondutojn.
4 La Efiko de Malfunkcio en Inhibitora Kontrolo
La kapablo malhelpi antaŭtempajn respondojn estas ligita por kontribui al la kapablo de individuo eviti okupiĝi pri netaŭgaj kondutoj, kiel preni drogojn aŭ manĝi preter la punkto de saĝeco, kaj tiel pliigi lian vundeblecon al toksomanio (aŭ obezeco) (Volkow kaj Fowler) 2000; Volkow et al. 2008a).
PET-studoj malkovris signifajn reduktojn en la disponebleco de D2R en la striato de toksomaniuloj, kiuj persistas dum monatoj post longa malstreĉiĝo [reviziita en (Volkow et al. 2009a)]. Simile, preklinikaj studoj en ronĝuloj kaj ne-homaj primatoj montris, ke ripetaj ekspozicioj al drogoj estas asociitaj kun reduktoj de striaj D2R-niveloj (Nader et al. 2006; Thanos et al. 2007; Volkow et al. 2001). En la striato, D2Rs mediacias signaladon en la striata nerekta vojo, kiu modulas antaŭfrontajn regionojn; kaj ĝia malreguligo pruviĝis plibonigi sentivigon al la efikoj de drogoj en bestaj modeloj (Ferguson et al. 2011). En homoj toksomaniitaj al drogoj, la redukto de striataj D2R estas asociita kun malpliigita agado de antaŭfrontaj regionoj kiel evidentigita per malpliiĝoj de la basko-glukoza metabolo (markilo de cerba funkcio) en OFC, cingulata antaŭa giro (ACC), kaj dorsolatera prefrontal-kortekso (DLPFC) ) (Volkow et al. 2001, 1993, 2007) (Fig. 6). Koncerne OFC, ACC, kaj DLPFC estas implikitaj kun saleca atribuo, inhibicia kontrolo / emocio-regulado kaj decidado respektive, estis postulite ke ilia malĝusta regulado per D2R-mediata DA-signalado en toksomaniuloj povis subjekti la plibonigitan motivan valoron de drogoj en sia konduto kaj la perdo de kontrolo pri konsumado de drogoj (Volkow kaj Fowler 2000). Krome, ĉar mankoj en OFC kaj ACC estas asociitaj kun deviga konduto kaj impulsemo (Fineberg et al. 2009), Malpliboniga modulado de ĉi tiuj regionoj de DA verŝajne kontribuos al la konsumado de komputa kaj impulsema drogo vidita en toksomanio (Goldstein kaj Volkow 2002). Efektive, en misuzantoj de metamfetamino, malalta striatala D2R estis asociita kun impulsemo (Lee et al. 2009), kaj ĝi ankaŭ antaŭdiris devigan kokainan administradon en ronĝuloj (Everitt et al. 2008). Reversa scenaro, en kiu komenca vundebleco por uzado de drogoj antaŭekzistas en antaŭfrostaj regionoj, kaj per kiu ripetaj uzoj de drogoj plue malpliiĝas en stria D2R, ankaŭ eblas. Efektive, studo farita en subjektoj, kiuj malgraŭ alta risko de alkoholismo (pozitiva familia historio de alkoholismo) ne estis alkoholuloj, malkaŝis pli altan ol normala havebla D2R-havebleco, kiu estis asociita kun normala metabolo en OFC, ACC, kaj DLPFC (Volkow et al. 2006a). Ĉi tio sugestas, ke en ĉi tiuj subjektoj kun risko por alkoholismo, la normala antaŭfronto estis ligita al plibonigita striatala D2R-signalado, kiu siavice eble protektis ilin kontraŭ alkoholuzo.
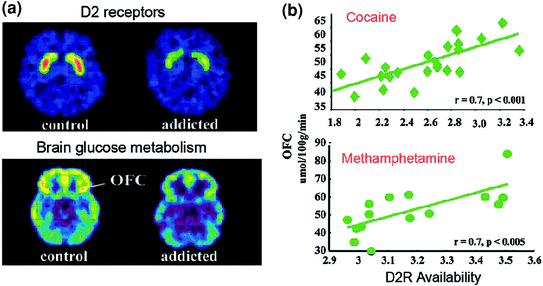
Figo. 6 Korelacioj inter striata D2R-havebleco kaj metabolo en prefrontalaj cerbaj regionoj. Axial-cerbaj bildoj por kontrolo kaj por kokain-toksomaniita temo por bazaj bildoj de havebleco de D2R en striatumo (akirita kun [11C] raclopride) kaj de cerba glukoza metabolo en OFC (akirita kun [18FDG). b Korelacioj inter striaj D2R kaj metabolo en OFC en koka-toksomaniuloj kaj metamfetaminecaj subjektoj. Reproduktita de Volkow et al. (2009a) Kopirajto (2009), kun permeso de Elsevier
Antaŭvideble, evidentecoj de malregulaĵo en kontrolaj cirkvitoj ankaŭ estis trovitaj inter obesaj individuoj. Ambaŭ preklinikaj kaj klinikaj studoj havigis evidentojn pri malpliigita striatala D2R-signalado, kiu, kiel menciite supre, estas ligita kun rekompenco (NAc) sed ankaŭ kun la starigo de kutimoj kaj rutinoj (dorsal-striatum) en obesito.y (Geiger et al. 2009; Wang et al. 2001). Grave, malpliigita striatala havebleco de D2R estis ligita al komputa manĝaĵa konsumado en obesaj ronĝuloj (Johnson kaj Kenny 2010) kaj kun malpliigita metabola agado en OFC kaj ACC en homoj obesaj (Volkow et al. 2008b) (Fig. 7a – c). Konsiderante, ke disfunkcio en OFC kaj ACC rezultigas kompulsivon [vidu recenzon (Fineberg et al. 2009)], tio povus esti parto de la mekanismo per kiu malalta signalado D2R faciligas hiperfagion (Davis et al. 2009). Krome, ĉar malpliiĝanta D2R-rilata signalado ankaŭ probable reduktas la sentivecon al aliaj naturaj rekompencoj, ĉi tiu deficito ĉe obesaj homoj ankaŭ povus kontribui al kompensita manĝado. (Geiger et al. 2008).
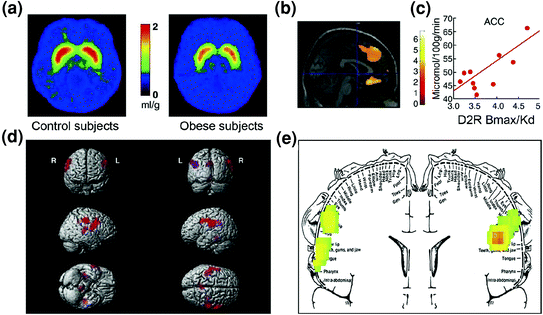
Figo. 7 Hiperfagio povus rezulti el veturado por kompensi malfortigitan rekompencan cirkviton (prilaboritan per dopamin-reguligitaj kortikostriaj cirkvitoj) kombinitan kun pli alta sentiveco al palatabileco (hedonaj ecoj de manĝaĵoj procesitaj parte tra la somatosensoria kortekso). a Mezumitaj bildoj por disponebleco de ricevilo DA D2 (D2R) en kontroloj (n = 10) kaj en malsanaj grasuloj (n = 10). b Rezultoj de (Statistika Parametrika Mapado) SPM identiganta la areojn en la cerbo, kie D2R estis asociita kun glukoza metabolo, ĉi tiuj inkluzivis la median OFC, ACC kaj la dorsolateral PFC (regiono ne montrita). c Regresa deklivo inter striata D2R kaj metabola agado en ACC en obesaj temoj. d Tridimensie bildigitaj SPM-bildoj montrantaj la areojn kun pli alta metabolo ĉe grasuloj ol ĉe maldikaj temoj (P <0.003, nekorektita). e Kolor-kodigitaj SPM-rezultoj montritaj en korona ebeno kun supermetita diagramo de la somatosensa homunkulo. La rezultoj (z-valoro) estas prezentitaj per la ĉielarka skalo kie ruĝa> flava> verda. Kompare kun maldikaj subjektoj, grasaj subjektoj havis pli altan bazan metabolon en la somatosensaj lokoj, kie la buŝo, lipoj kaj lango estas reprezentataj kaj kiuj estas implikitaj kun prilaborado de manĝaĵoj. Modifita, kun permeso, de Volkow k.a. (2008a) (a – c) kaj Wang et al. (2002) (d, e)
Ĉi tiu hipotezo konformas al preklinikaj evidentaĵoj, kiuj montras, ke malpliigita DA-agado en VTA rezultigas draman kreskon de konsumado de altaj grasaj manĝaĵoj (Stoeckel et al. 2008). Simile, kompare kun normal-pezaj individuoj, obesaj homoj, kiuj estis prezentitaj kun bildoj de alta kaloria manĝaĵo (stimuloj, al kiuj ili estas kondiĉitaj), montris pliigitan neŭralan aktivadon en regionoj, kiuj estas parto de rekompenco kaj instigaj cirkvitoj (NAc, dorsal striatum, OFC , ACC, amigdala, hipokampo kaj insula) (Killgore kaj Yurgelun-Todd 2005). Kontraŭe, en normal-pezaj kontroloj, la aktivigo de ACC kaj OFC (regionoj implikitaj en saleca atribuo, kiu projektas en la NAc) dum prezento de alt-kaloria manĝaĵo estis trovita esti negative korelaciita kun ilia korpa mas-indekso (IMC) ( Stice et al. 2008b). Ĉi tio sugestas dinamikan interagadon inter la kvanto de manĝaĵo manĝita (reflektita parte en la BMI) kaj la reaktiveco de rekompencaj regionoj al alt-kaloria manĝaĵo (reflektita en la aktivigo de OFC kaj ACC) ĉe normal-pezaj individuoj, kio estas perdita en obezeco.
Surprize, obesaj homoj elmontris malpli da aktivigo de rekompencaj cirkvitoj de efektiva manĝaĵa konsumo (nomata konsuma manĝaĵa rekompenco) ol maldikaj individuoj, dum ili montris pli grandan aktivigon de somatosensoraj kortikaj regionoj, kiuj procesas palatecon, kiam ili antaŭvidis konsumon (Stice et al. 2008b). Ĉi-lasta observado korespondis al regionoj kie antaŭa studo montris plibonigitan agadon ĉe obesaj subjektoj testitaj ĉe la komenco (ne stimulado) (Wang et al. 2002) (Fig. 7d, e). Pliigita agado de regionoj, kiuj procesas plaĉecon, povas fari obesajn subjektojn favori manĝon super aliaj naturaj plifortigiloj, dum malpliigita aktivigo de dopaminergiaj celoj per la reala manĝaĵa konsumo povus konduki al troa konsumado kiel rimedo por kompensi malfortan signaladon de D2R-meditado (Stice et al. 2008a). Ĉi tiu reduktita respondo de la rekompenca cirkvito al manĝaĵa konsumo en obesaj subjektoj estas rememoriga pri la malpliigitaj DA kreskoj deĉenigitaj de drogokonsumado en toksomaniuloj kiam kompare kun ne-toksomaniuloj.
La prefrontal-kortekso (PFC) ludas gravegan rolon en plenuma funkcio, inkluzive de inhibicia kontrolo (Miller kaj Cohen 2001). Ĉi tiuj procezoj estas modulitaj de D1R kaj D2R (supozeble ankaŭ D4R) kaj tiel, la malpliigita aktiveco en PFC, tiel en toksomanio kiel en obesidad, verŝajne kontribuos al malbona kontrolo kaj alta kompulsio. La pli malalta ol normala havebleco de D2R en la striato de obesaj individuoj, kiu estis asociita kun reduktita aktiveco en PFC kaj ACC (Volkow et al. 2008b) tial verŝajne kontribuas al ilia manka kontrolo pri konsumado de manĝaĵoj. Efektive, la negativa korelacio inter IMC kaj striata D2R raportita en obesoj (Wang et al. 2001) kaj en superpeso (Haltia et al. 2007a) individuoj subtenas ĉi tion. Pli bona kompreno de la mekanismoj, kiuj kondukas al difektita PFC-funkcio en obezeco (aŭ toksomanio), povus faciligi la disvolviĝon de strategioj por plibonigi, aŭ eble eĉ reverti specifajn difektojn en kernaj kognaj domenoj. Ekzemple, malfrua rabatado, kiu estas la tendenco devalori rekompencon kiel funkcio de la tempo de prokrasto de ĝia liverado, estas unu el la plej amplekse esploritaj kognaj operacioj rilate al malordoj asociitaj kun impulsiveco kaj kompulsiveco. Malfrua rabatado estis plej ĝisfunde esplorita en drogaj misuzantoj, kiuj montras troigitan preferon por malgrandaj sed tujaj super grandaj sed malfruaj rekompencoj (Bickel et al. 2007). Tamen, la malmultaj studoj faritaj kun obesaj homoj ankaŭ malkovris evidentaĵojn pri prefero por altaj, tujaj rekompencoj, malgraŭ pliigita ebleco suferi pli altajn estontajn perdojn (Brogan et al. 2010; Weller et al. 2008). Kaj pli lastatempe, alia studo trovis pozitivan korelacion inter IMC kaj hiperbola rabatado, per kio estontaj negativaj rekompencoj estas malpliigitaj ol estontaj pozitivaj rekompencoj (Ikeda et al. 2010). Interese, malfrua rabatado ŝajnas dependi de la funkcio de ventrala striatumo (Gregorios-Pippas et al. 2009) kaj de la PFC, inkluzive de flanka OFC (Bjork et al. 2009), kaj estas sentema al DA-manipuladoj (Pine et al. 2010). Specife, plibonigi DA-signaladon (kun L-DOPA-traktado) pliigis impulsivecon kaj tempan rabatadon.
5 Partopreno de Motivaj Cirkvitoj
Dopaminergia signalado ankaŭ modulas instigon. Kondutismaj trajtoj kiel vigleco, persistemo kaj investado de daŭra penado por atingi celon estas ĉiuj submetataj al modulado de DA aganta tra pluraj celaj regionoj, inkluzive de NAc, ACC, OFC, DLPFC, amigdala, dorsal striatum, kaj ventral pallidum (Salamone et al. 2007). Malreguligita DA-signalado estas asociita kun plibonigita instigo por eltiri drogojn, karakteriza toksomanio, tial la drog-toksomaniuloj ofte okupiĝas pri ekstremaj kondutoj por akiri drogojn, eĉ kiam ili kunportas konatajn severajn kaj adversajn konsekvencojn (Volkow kaj Li 2005). Ĉar konsumado de drogoj fariĝas la ĉefa instiga enmiksiĝo en drogmanio (Volkow et al. 2003), toksomaniuloj estas instigitaj kaj motivitaj de la procezo por akiri la drogon, sed inklinas esti retiritaj kaj apatiaj kiam eksponitaj al ne-drogaj agadoj. Ĉi tiu ŝanĝo estis studita komparante la cerbajn aktivadajn ŝablonojn okazantajn kun ekspozicio al kondiĉitaj signoj kun tiuj okazantaj en la foresto de tiaj indikoj. Kontraste al la malkreskoj en la antaŭfronta agado raportita en detoxifitaj kokainaj fitraktantoj kiam ne stimulitaj kun drogaj aŭ drogaj sondoj [vidu recenzon (Volkow et al. 2009a)], ĉi tiuj prefrontalaj regionoj fariĝas aktivigitaj kiam kokainaj fitraktantoj estas elmontritaj al avidaj induktaj stimuloj (ĉu drogoj aŭ kuracaj rimedoj) (Grant et al. 1996; Volkow et al. 1999a; Wang et al. 1999). Ĉi tiu rezulto rememorigas la observadon, ke maltrankviligantoj de kokaino, studitaj baldaŭ post epizodo de kokaina bingado, montris kreskon de metabola agado en OFC kaj ACC (ankaŭ dorsal striatum), kiu estis asociita kun avido (Volkow et al. 1991).
Plie, kiam la respondoj al iv MP estas komparataj inter kokainaj toksomaniuloj kaj ne-toksomaniuloj, la unua respondis kun pliigita metabolo en ventra ACC kaj media OFC (efiko asociita kun avido), dum la dua montris la kontraŭan respondon, nome malpliiĝis. metabolo en ĉi tiuj regionoj (Volkow et al. 2005). Ĉi tio sugestas, ke la aktivigo de ĉi tiuj antaŭfrontaj regionoj kun ekspozicio al drogoj povas esti specifa al toksomanio kaj asociita kun la plibonigita deziro por la drogo. Krome, studo kiu instigis kokain-toksomaniulojn al intence malhelpado de avido kiam eksponitaj al drogaj indikoj montris, ke tiuj subjektoj sukcesantaj malhelpi avidon montris malpliiĝon de metabolo en meda OFC (kiu prilaboras motivan valoron de plifortigilo) kaj NAc (kiu antaŭdiras rekompenco) (Volkow et al. 2009b). Ĉi tiuj trovoj plue konfirmas la implikiĝon de OFC, ACC, kaj striatum en la plibonigita instigo por akiri la drogon viditan en toksomanio.
Antaŭvideble, la OFC ankaŭ estis implicita en atribui salecan valoron al manĝaĵo (Grabenhorst et al. 2008; Rolls kaj McCabe 2007), helpante taksi ĝiajn atendatajn agrablaĵojn kaj plaĝecon kiel funkcio de ĝia kunteksto. PET-studoj kun FDG por mezuri cerban glukozon-metabolon en normalaj pezaj individuoj raportis, ke eksponiĝo al manĝaĵ-kresko pliigis metabolan aktivecon en OFC, kio estis efiko asociita kun la percepto de malsato kaj deziro de manĝaĵo (Wang et al. 2004). La plibonigita OFC-aktivigo per la nutraĵa stimulo probable reflektas maldensajn dopaminergiajn efikojn kaj partoprenos la partoprenadon de DA en la veturado por manĝaĵ-konsumo. La OFC ludas rolon en lernado de stimulo-plifortigaj asocioj kaj kondiĉado (Cox et al. 2005; Gallagher et al. 1999), subtenas kondiĉitan nutraĵon de nutrado (Weingarten) 1983), kaj probable kontribuas al troo senkonsidere de malsatkatastaloj (Ogden kaj Wardle) 1990). Efektive, misfunkcio de la OFC estis ligita al troo (Machado kaj Bachevalier) 2007).
Malgraŭ iuj malkonsekvencoj inter studoj, cerbaj bildigaj datumoj ankaŭ subtenas la nocion, ke strukturaj kaj funkciaj ŝanĝoj en cerbaj regionoj implikitaj en plenuma funkcio (inkluzive de inhibitora kontrolo) povus esti asociitaj kun alta IMC ĉe alie sanaj individuoj. Ekzemple, MRI-studo farita ĉe maljunaj virinoj, uzante voxel-bazitan morfometrion, trovis negativan korelacion inter IMC kaj grizaj materiaj volumoj (inkluzive de frontaj regionoj), kiu, en la OFC, estis asociita kun malplibonigita plenuma funkcio (Walther et al. 2010). Uzante PET por mezuri cerbon-glukozan metabolon en sanaj kontroloj, ni raportis negativan korelacion inter IMC kaj metabola aktiveco en DLPFC, OFC, kaj ACC. En ĉi tiu studo, la metabola aktiveco en antaŭfrontaj regionoj antaŭdiris la agadon de la subjektoj en provoj de plenuma funkcio (Volkow et al. 2009c). Simile, nuklea magneta resono (NMR) spektroskopa studo en sana mezaĝo kaj maljunaj kontroloj montris, ke IMC estis negative asociita kun la niveloj de N-acetil-aspartato (markilo de neŭrona integreco) en frontala kortekso kaj ACC (Gazdzinski et al. 2008; Volkow et al. 2009c).
Studoj de cerbaj bildoj komparantaj obeajn kaj maldikajn individuojn ankaŭ raportis pli malaltan densecon de griza materio en frontaj regionoj (frontala operculum kaj meza frontala giro) kaj en post-centra giro kaj putamen (Pannacciulli et al. 2006). Alia studo trovis neniujn diferencojn en volumoj de griza materio inter obesaj kaj maldikaj subjektoj, tamen, ĝi registris pozitivan korelacion inter blanka materio-volumo en bazaj cerbaj strukturoj kaj talio al kokso-proporcioj, tendenco kiu estis parte inversigita per dietado (Haltia et al. 2007b). Interese, ke kortikaj areoj, kiel la DPFC kaj OFC, kiuj estas implikitaj en inhibitora kontrolo, ankaŭ estis trovitaj esti aktivigitaj en sukcesaj dieteroj kiel respondo al manĝa konsumado (DelParigi et al. 2007), sugestante eblan celon por kondutisma formado en traktado de obesidad (kaj ankaŭ en toksomanio).
6 Partopreno de Interoceptive Circuitry
Neuroimagaj studoj malkaŝis, ke la meza insulo ludas kritikan rolon en avidoj pri manĝaĵoj, kokainoj kaj cigaredoj. (Bonson et al. 2002; Pelchat et al. 2004; Wang et al. 2007). La graveco de la insulo estis elstarigita per studo, kiu raportis, ke fumantoj kun damaĝo en ĉi tiu regiono (sed ne kontrolis fumantojn, kiuj suferis ekster-insulajn lezojn) kapablis ĉesi fumi facile kaj sen sperti nek avidojn nek relokiĝon (Naqvi et al . 2007). La insulo, precipe ties pli antaŭaj regionoj, estas reciproke konektita al pluraj regionoj de la limuzino (ekz. Ventromedaj antaŭfrontalaj kortekso, amigdala kaj ventrala striatumo) kaj ŝajnas havi interkonceptan funkcion, integrante la aŭtonomajn kaj viscerajn informojn kun emocio kaj instigo, tiel provizante konscia konscio pri ĉi tiuj urĝoj (Naqvi kaj Bechara 2009). Efektive, cerbaj lezaj studoj sugestas, ke la ventromedaj PFC kaj insuloj estas necesaj komponentoj de la distribuitaj cirkvitoj, kiuj subtenas emocian decidadon (Clark et al. 2008). Konforme al ĉi tiu hipotezo, figuradaj studoj konstante montras diferencigan aktivadon de la insula dum avido (Brody et al. 2009; Goudriaan et al. 2010; Naqvi kaj Bechara 2009; Wang et al. 1999). Laŭ ĝi sugestis, ke la reaktiveco de ĉi tiu cerba regiono funkcias kiel biomarkilo por helpi antaŭdiri reaperadon (Janes et al. 2010).
La insuleto estas ankaŭ primara gustuma areo, kiu partoprenas multajn aspektojn de manĝadaj kondutoj, kiel gusto. Krome, la vizaĝa insulaĵo (ligita al primara gusto-kortekso) liveras informojn al la OFC, kiu influas ĝian multimodan reprezentadon de la plaĉo aŭ rekompenca valoro de venonta manĝaĵo (Rolls 2008). Pro la implikiĝo de la insuleto en la interkaptan sencon de la korpo, en emocia konscio (Craig 2003) kaj en instigo kaj emocio (Ruloj 2008), kontribuo de insula difekto en obezeco povus esti atendita. Efektive, gastrika distingo rezultigas aktivadon de la posta insulo, kiu probable reflektas ĝian rolon en la konscio pri korpaj statoj (ĉi-kaze de pleneco) (Wang et al. 2008). Plie, en malgrasaj, sed ne en obesaj subjektoj, gastra distingo rezultigis aktivadon de la amigdala kaj senaktivigon de la antaŭa insulo (Tomasi et al. 2009). La manko de amigdala respondo ĉe obesaj subjektoj povus reflekti senkonscian interkonceptan konscion pri korpaj ŝtatoj ligitaj kun saĝeco (plena stomako). Kvankam la modulado de insula agado fare de DA estis nebone esplorita, oni agnoskas, ke DA okupiĝas pri la respondoj al gustumo de plaĉaj manĝaĵoj, kiuj estas mediaciitaj per la insuleto (Hajnal kaj Norgren). 2005). Studoj pri homaj bildoj montris, ke gustumado de plaĉaj manĝaĵoj aktivigis la insulan kaj mezkranan areon (DelParigi et al. 2005; Frank et al. 2008). Tamen, la DA-signalado povus esti necesa ankaŭ por senti la kalorian enhavon de manĝaĵo. Ekzemple, kiam virinoj kun normala pezo gustigis dolĉigilon kun kalorioj (sukerozo), la insulaj kaj dopaminergiaj mezkruraj areoj aktivis, dum gustumo de kalorioj sen kalorioj (sukralose) nur aktivigis la insulaĵon (Frank et al. 2008). Obesaj subjektoj montras pli grandan insulan aktivadon ol normalaj kontroloj dum gustumo de likva manĝo, kiu konsistas el sukero kaj graso (DelParigi et al. 2005). Kontraŭe, subjektoj, kiuj resaniĝis de anoreksio nervosa, montras malpli da aktivigo en la insulo dum gustumado de sukerozo kaj neniun asocion de sentoj de plaĉo kun insuleca aktivado kiel observite en la normalaj kontroloj (Wagner et al. 2008). Se kombinitaj, ĉi tiuj rezultoj faras probable, ke malregulaĵo de la insula respondo al gustaj stimuloj povus esti implikita en la neplenaĝa kontrolo de diversaj apetitaj kondutoj.
7 La Cirkvitado de Aversio
Kiel menciite antaŭe, trejnado (kondensado) sur kvazo, kiu antaŭdiras rekompencon, kondukas al dopaminergiaj ĉeloj pafantaj en respondo al la antaŭdiro de rekompenco, kaj ne al rekompenco mem. Aliflanke, kaj konforme al ĉi tiu logiko, oni observis, ke dopaminergiaj ĉeloj ekbruliĝos malpli ol kutime, se la atendata rekompenco malsukcesos konkuriĝi (Schultz et al. 1997). Akumula indico (Christoph et al. 1986; Lisoprawski et al. 1980; Matsumoto kaj Hikosaka 2007; Nishikawa et al. 1986) notas la habenula kiel unu el la regionoj, kiuj kontrolas la malpliiĝojn de pafo de dopaminergiaj ĉeloj en VTA, kiuj eble sekvos la malsukceson ricevi atendatan rekompencon (Kimura et al. 2007). Tiel, pliigita sentiveco de la habenula, kiel rezulto de kronikaj ekspozicioj al drogoj, povus subteni pli grandan reaktivecon al drogoj. Efektive, aktivigo de la habenula, en kokain-toksomaniulaj subjektoj, estis asociita kun kondutema relanĉo al drogoj post ekspona indico (Brown et al. 2011; Zhang et al. 2005). En la kazo de nikotino, la nikotinaj riceviloj α5 en la habenula ŝajnas moduli la aversajn respondojn al grandaj dozoj de nikotino (Fowler et al. 2011); kaj la α5 kaj α2-riceviloj en la habenula estas implicitaj en nicotina retiriĝo (Salas et al. 2009). Pro la kontraŭa respondo de la habenula al tiu de DA-neŭronoj por rekompenci (malaktiviĝo) kaj ĝia aktivado post eksponiĝo al aversaj stimuloj, ni raportas ĉi tie al la signalado de la habenula kiel unu elportanta "kontraŭreala" enigaĵo.
La habenula ŝajnas ludi similan rolon rilate al manĝaĵa rekompenco. Tre palpebla manĝa dieto povas indukti obezecon ĉe ratoj, kaj la pezaj kreskoj korelaciante kun pliigoj de μ-opioidaj peptidaj ligoj en la bazolateral kaj bazomedia amigdalo. Interese, la meza habenula montris signife pli altan μ-opioidan peptidan ligon (ĉirkaŭ 40%) post eksponiĝo al la plaĉa manĝaĵo ĉe la ratoj, kiuj akiris pezon (tiuj, kiuj konsumis pli da manĝaĵo) sed ne en tiuj, kiuj ne faris (Smith et al. 2002). Ĉi tio sugestas, ke la habenula eble okupiĝos pri tro manĝado sub kondiĉo de havebla manĝebla manĝo. Plie, neŭronoj en la rostromedia tegmenta kerno, kiuj ricevas gravan enigon de la flanka habenula, projektas al VTA DA neŭronoj kaj estas aktivigitaj post manĝa senigado (Jhou et al. 2009). Ĉi tiuj trovoj estas konformaj al la rolo por la habenula mediado de respondoj al avversaj stimuloj aŭ ŝtatoj kiel tiuj, kiuj okazas dum dietado aŭ retiriĝo de drogoj.
La implikiĝo de la habenula kiel kontraŭreala kerno ene de emociaj retoj konformas al antaŭaj teoriaj modeloj de toksomanio, kiuj postulis sentivigitajn kontraŭ-rekompencajn respondojn (mediaciitaj per pliigita sentiveco de la amigdala kaj pliigita signalado kvankam la kortikopropona liberiga faktoro) kiel veturado de konsumado de drogoj en toksomanio. (Koob kaj Le Moal 2008). Similaj antirrespondaj respondoj ankaŭ povas kontribui al troa manĝaĵa konsumo en obezeco.
8 Patologia Droga kaj Manĝaĵa Rekompenco: Ĝisdatigita Labora Modelo
TLi kapablas rezisti la bezonon uzi drogon aŭ manĝi preter punpunkto postulas taŭgan funkciadon de neŭronaj cirkvitoj implikitaj en suprena kontrolo por kontraŭi la kondiĉitajn respondojn, kiuj antaŭdiras rekompencon de la ingesta manĝo / drogo kaj la deziro ingesti la manĝaĵo / drogo. Ĉi tie, ni reliefigis ses el ĉi tiuj cirkvitoj: rekompenco / malkuraĝigo, kutimado / kutimoj, inhibicia kontrolo / plenuma funkcio, instigo / veturado, interkompreniĝo kaj aversia evitado / streĉa reaktiveco (Fig. 8). Surbaze de la bildaj datumoj prezentitaj ĉi tie, ni proklamas, ke temas pri la diskreteco inter la atendo pri la drogaj / manĝaj efikoj (kondiĉitaj respondoj) kaj la malpacaj neŭrofiziologiaj efikoj, kiuj konservas la prenon de drogoj aŭ la konsumon de nutraĵoj en provo atingi la atendita rekompenco. Ankaŭ, ĉu testate dum fruaj aŭ longaj periodoj de sindeteno / dietado, toksomaniuloj / obesaj subjektoj montras pli malaltan D2R en striato (inkluzive NAc), kiuj estas asociitaj kun malpliiĝoj de baza agado en frontaj cerbaj regionoj implicitaj en saleca atribuo (orbitofrontala kortekso) kaj inhibitoro. kontrolo (ACC kaj DLPFC), kies interrompo rezultigas kompulsivecon kaj impulsemon. Finale, evidentaĵoj ankaŭ aperis sur la rolo de interkaptaj kaj avaraj cirkvitoj en la sistemaj malekvilibroj, kiuj rezultigas devigan konsumon de aŭ drogoj aŭ nutraĵoj.
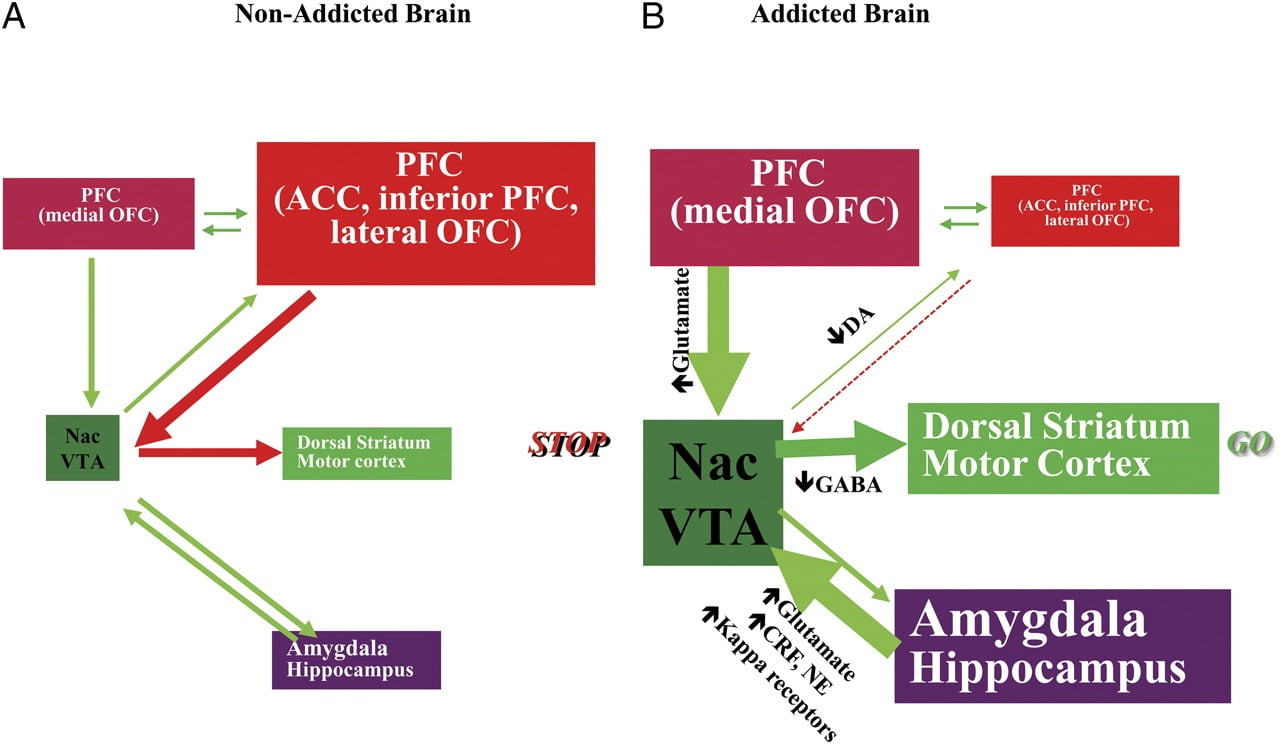
Figo. 8 Modelo proponanta reton de interagaj cirkvitoj, interrompoj, kiuj kontribuas al la kompleksa aro de stereotipaj kondutoj sub la drogmanio kaj kronika supertraktado: rekompenco (nucleus accumbens, VTA, kaj ventral pallidum), kondiĉado / memoro (amigdala, meza OFC por atribuo de saleco, hipokampo kaj dorsal striatumo por kutimoj), plenuma kontrolo (DLPFC, ACC, malplena fronta kortekso, kaj laterala OFC), instigo / veturado (meza OFC por atribuo de saleco, ventrala ACC, VTA, SN, dorsal-striato, kaj motora kortekso). Nac, nucleus accumbens, interkaptado (Insula kaj ACC), kaj aversio / evitado (Habenula). a Kiam ĉi tiuj cirkvitoj ekvilibrigas, tio rezultigas taŭgan inhibician regadon kaj decidadon. b Dum toksomanio, kiam la plibonigita atenda valoro de la drogo en la rekompenco, instigo kaj memoraj cirkvitoj venkas la kontrolan cirkviton, favoras pozitivan retroscivolon iniciatitan de la konsumo de la drogo kaj eternigita per plibonigita aktivigo de la instigo / veturado kaj memoraj cirkvitoj. Ĉi tiuj cirkvitoj ankaŭ interagas kun cirkvitoj implikitaj en humora regulado, inkluzive de streĉa reaktiveco (kiu implikas la amigdala, hipotalamo, habenula) kaj interparoladon (kiu implikas la insulon kaj ACC kaj kontribuas al konscio pri avido). Pluraj neurotransmisiloj estas implikitaj en ĉi tiuj neŭroadaptoj, inkluzive de glutamato, GABA, norepinefrino, liberiganta faktoro de kortikotropino kaj opioidaj riceviloj. CRF, liberiganta faktoro de kortikotropino; NE, norepinefrino. Modifita kun permeso de Volkow et al. (2011b)
Kiel sekvo de la sinsekva interrompo en ĉi tiuj cirkvitoj, individuoj povas sperti 1) plibonigitan motivan valoron de la drogo / manĝaĵo (sekundara al lernitaj asocioj per kondiĉado kaj kutimoj) koste de aliaj plifortigiloj (malĉefe al malpliigita sentiveco de la rekompenca cirkvito). ), 2) malhelpa kapablo malhelpi la intencajn (cel-direktitajn) agojn ekigitajn de la forta deziro preni la drogon / manĝaĵon (malĉefe al malvalora plenuma funkcio), kiuj rezultas en deviga drogo / manĝa preno, kaj 3) plibonigita streĉa reaktiveco kaj perversa evitado, kiu rezultigas impulsan drogon, por eskapi de la avara stato.
Ĉi tiu modelo sugestas ĝeneraligitan terapian alproksimiĝon al toksomanio desegnita por malpliigi la plifortigajn propraĵojn de drogo / manĝaĵo, restarigi / plibonigi la rekompencajn propraĵojn de naturaj plifortigiloj, malhelpi kondiĉitajn lernitajn asociojn, plibonigi instigon por ne-drogaj / manĝaĵoj, malpliigi streĉan reaktivecon , plibonigu humoron kaj plifortigas ĝeneraluzan inhibician regadon.
Dankojn
La aŭtoroj ŝatus danki la subtenon de la NIAAA-intramural programo de la Naciaj Institutoj de Sano.
Referencoj
