Fronto. Konduto Neurosci., 23 March 2016 | http://dx.doi.org/10.3389/fnbeh.2016.00054

 Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2,
Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2, 

- 1Tradukista Esplorinstituto kaj Instituto por Sano kaj Biomedika Novigado, Kvinslanda Universitato de Teknologio, Brisbano, QLD, Aŭstralio
- 2Lernejo de Biomedicinaj Sciencoj, Universitato de Kvinslando, Brisbano, QLD, Aŭstralio
La moderna dieto fariĝis tre dolĉigita, rezultigante senprecedencajn nivelojn de sukero konsumo, precipe inter adoleskantoj. Kvankam estas konata ke kronika longtempa sukero kontribuas al la disvolviĝo de metabolaj malordoj inkluzive de obezeco kaj diabeto de tipo II, oni scias malmulte pri la rektaj konsekvencoj de longtempa binge-simila sukero-konsumo sur la cerbo. BEkaŭka sukero povas kaŭzi la liberigon de dopamino en la kerno de akcentoj (NAc) simile al drogoj pri fitraktado, ni esploris ŝanĝojn en morfologio de neŭronoj en ĉi tiu cerba regiono post mallongaj (4-semajnoj) kaj longtempaj (12-semajnoj) binge- kiel sukeroza konsumo uzante intermitan du-botelan elektan paradigmon. Ni uzis Golgi-Cox-makuladon por impregnado de mezaj spinecaj neŭronoj (MSNoj) el la NAc-kerno kaj ŝelo de mallongaj kaj longperspektivaj sukerozaj ratoj kaj komparis tiujn al aĝaj kongruaj akvokontroloj. Ni montras, ke plilongigita binge-simila sukroza konsumo signife malpliigis la totalan dendritan longon de NAc-ŝelaj MSNoj kompare kun maljunaj kontrolaj ratoj. Ni ankaŭ trovis ke la restrukturado de ĉi tiuj neŭronoj rezultis ĉefe el reduktita distala dendrita komplikeco. Al la inversa, ni observis pliigitajn spinecajn densecojn ĉe la distalaj branĉaj ordoj de NAc-ŝelaj MSNoj de longtempaj sukerozo konsumantaj ratojn. Kombinitaj, ĉi tiuj rezultoj reliefigas la neŭronajn efikojn de plilongigita binge-simila konsumado de sukroso sur NAc-ŝela morfologio.
Enkonduko
Dum la lastaj 40 jaroj, estas dokumentita pliiĝo en la konsumo de sukerkolitaj trinkaĵoj kaj manĝaĵoj enhavantaj aldonitajn sukerojn (Nielsen et al., 2002; Popkin, 2010; Ng et al., 2012), kun raportoj taksantaj, ke ĝis 75% de ĉiuj manĝaĵoj kaj trinkaĵoj enhavas altajn kvantojn da aldonitaj sukeroj (Ford kaj Dietz, 2013; Bray kaj Popkin, 2014). Dum ĉi tiu periodo, ankaŭ estis samtempa kresko en la prevalenco de obezeco kaj tipo II-diabeto, precipe ĉe adoleskantoj (Arslanian, 2002; Reinehr, 2013; Dabelea et al., 2014; Fryar et al., 2014). Lastatempaj studoj montris, ke superpezaj kaj obesaj infanoj ofte konsumas altajn kvantojn da aldonita sukero, tamen la kontribuo de alta sukero enhavanta dietojn al la pliigita efiko de trostreĉitaj kaj obesaj infanoj restas polemika (Hu, 2013; Bray kaj Popkin, 2014; Bucher Della Torre et al., 2015).
Dum kreskanta korpa indico indikas, ke la konsumo de alta sukero-dietoj povas parte kontribui al pezo-kresko inter infanoj kaj adoleskantoj (Malik et al., 2010; Te Morenga et al., 2013; Bray kaj Popkin, 2014), malpli da atento estis al adversaj ne-metabolaj konsekvencoj rezultantaj de troa konsumado de sukero. Interese, iuj oftaj kondutaj kaj psikologiaj padronoj ofte aperas inter subaro de tiuj, kiuj manĝas kaj tenas altan sukeron enhavantan dieton. Plej rimarkindaj estas la evoluo de manĝadaj malordoj inkluzive de binge-manĝado, kombinita kun la samtempa apero de psikologiaj simptomoj inkluzive de manko de instigo kaj deprimo (reviziita en Sheehan kaj Herman, 2015). Krome, ĉar binge-manĝantaj individuoj ofte montras perdon de kontrolo kaj nekapablo mem-limigi sian sukeraĵon de sukero, verŝajne ĉi tiuj kondutoj ekestas rezulte de neŭrologiaj adaptiĝoj en cerbaj regionoj, kiuj taksas hedonan valoron de tre plaĉa manĝo. (Saper et al., 2002; Lutter kaj Nestler, 2009; Kenny, 2011). Ĉi tiu racio ankaŭ estas subtenata de evidentaĵoj ĉe homoj, kiuj montras, ke sukero kaj dolĉeco povas kaŭzi avidojn similajn al tiuj induktitaj de toksomaniuloj kiel alkoholo kaj nikotino (Volkow et al., 2012).
Kvankam la toksomaniaj ecoj de sukero ankoraŭ spekulativaj, ĉi tiuj observaĵoj kombinitaj kun studoj montrantaj la kontribuon de troa konsumado de sukero al ŝanĝoj en rekompencaj cirkvitoj kaj disvolviĝo de toksomaniulaj kondutoj kaj emociaj statoj en bestaj modeloj (Avena et al., 2008; Benton, NENIU; Ventura et al., 2014), garantias la bezonon por plia enketo. Antaŭaj studoj en ronĝuloj montris, ke intermita aliro al sukerozo ŝanĝas la agadon de pluraj neurotransmisiloj ene de la mezolimbia sistemo inkluzive de dopamino, opioidoj kaj acetilcolino (Reviziita en Avena et al., 2008). Bing-simila konsumado de sukroso estas montrita por faciligi la liberigon de dopamino en la kerno de akciuloj (NAc), simile al drogoj de misuzo (Avena et al., 2008). Plue, ni montris, ke longtempa konsumo de sukroso uzanta 24 h intermitan aliron du-botela elekta paradigmo (Simms et al., 2008) modulas nikotinan acetilkolinan ricevilon (nAChR) esprimon en la NAc (Shariff et al., En gazetaro). Interese, ni ankaŭ observis, ke nAChR-komponaĵoj konataj moduli dopaminan kaj acetilcolinan agadon en la NAc, havas malsamajn efikojn al sukeroza konsumo post mallonga kaj longtempa konsumado (Shariff et al., En gazetaro).
Dum ĉi tiuj studoj pruvis similecojn en la kondutaj kaj neŭkemiaj ŝanĝoj kaŭzitaj de intermita aliro al sukero kaj drogoj de misuzo, oni ne scias, ĉu ĉi tiuj efikoj faciligas ŝanĝojn en neurona morfologio en la NAc. Tlia kontraste kun substancoj de misuzo inkluzive de kokaino, amfetamino kaj nikotino, kiuj produktas bone karakterizitajn ŝanĝojn en la morfologio de mezaj spinecaj neŭronoj (MSNoj) en la NAc, inkluzive de pliigita spina denseco kaj ŝanĝita dendrita komplekseco. (Robinson kaj Kolb, 1999, 2004; Li et al., 2003; Crombag et al., 2005). Ĉar ni antaŭe montris, ke longtempa ekspozicio (12-semajno) al alkoholo kaj sukerozo uzante la intermitan du-botelan elektan paradigmon produktas diferencan respondon al farmacoterapiaj intervenoj kompare al mallongtempa konsumado (4-semajnoj; Steensland et al., 2007; Shariff et al., En gazetaro), ni taksis la efikojn de mallonga kaj longperspektiva sukroza konsumo sur MSN-morfologio en la NAc. Ni permesis al adoleskaj ratoj konsumi sukerozon en binge-simila maniero por 4 (mallongdaŭraj) aŭ 12 (longtempaj) semajnoj kaj tiam analizis morfologion de NAc-MSNoj de mallongaj kaj longperspektivaj sukerozaj ratoj kaj komparis tion al aĝ-kongruaj kontroloj, kiuj ricevis aliron al akvo nur. Niaj rezultoj montras, ke MSN-oj de la NAc-ŝelo estas ŝanĝitaj sekvante longan sed ne mallongperspektivan konsumon de sukerozo, malpliigante dendritan longon, sed pliigis distalan dendritan spinan densecon. Plue, ni trovis la morfologion de MSNoj de la NAc-kerno restis relative sendifektaj post mallonga kaj longtempa sukeroza konsumo. Ĉi tiuj rezultoj reliefigas rektan neŭrologian konsekvencon de longtempa sukroza konsumo en binge-simila maniero. Plue, ĉi tiuj datumoj pruvas la bezonon de pliaj studoj celantaj elucidigi la molekulajn kaj neŭkemiajn ŝanĝojn, kiuj akompanas la morfologian restrukturadon de NAc-ŝelaj MSNoj induktitaj de daŭrigita binge-simila sukerosa konsumado.
Materialoj kaj metodoj
Etika Komento
Ĉiuj eksperimentaj proceduroj estis efektivigitaj konforme al la Aŭstralia Kodo por Prizorgo kaj Uzo de Bestoj por Sciencaj Celoj, 8th Edition (Nacia Konsilio pri Sanaj kaj Kuracaj Esploroj, 2013). La protokoloj estis aprobitaj de la Komitato pri Etika Besto-Komitato de la Universitato de Kvinslando kaj la Komitato pri Etika Besto de Universitato de Kvinslando.
Bestoj kaj Loĝado
Kvin-semajnaj (adoleskaj) viraj Wistar-ratoj (Kontrolo: 176.4 ± 4.8 g; Sukrozo: 178.3 ± 5.0 g) (ARC, WA, Aŭstralio), estis individue loĝataj en ventolita duobla nivelo Plexiglas® kaĝoj. La ratoj estis aklimigitaj al la unuopaj loĝkondiĉoj, uzado kaj inversa luma ciklo 5 tagojn antaŭ la komenco de la eksperimentoj. Ĉiuj ratoj estis loĝigitaj en klimat-kontrolita 12-hr renversita lumo / malhela ciklo (lumoj malŝaltitaj ĉe 9 am) ĉambro kun norma rato chow kaj akvo havebla ad libitum.
Intermita-Alira Du-Botela Elekto-Trinkado-Paradigmo
La intermita aliro 5% sukrosa du-botela elekto trinkanta paradigmon (Simms et al., 2008) estis adaptita el Saĝa (1973). Ĉiuj fluidoj estis prezentitaj en gradigitaj plastaj boteloj de 300 ml kun trinkakvotuboj el neoksidebla ŝtalo enmetitaj tra du ŝirmiloj en la antaŭo de la kaĝo post la komenco de la malhela ciklo de lumo. Pezoj de ĉiu botelo estis registritaj antaŭ prezento al boteloj. Du boteloj estis prezentitaj samtempe: unu botelo enhavanta akvon; la dua botelo enhavanta 5% (w / v) sukroso. La lokigo de 5% (w / v) sukerosa botelo estis alternita kun ĉiu ekspozicio por kontroli por flankaj preferoj. Boteloj estis pesitaj 24 h post la prezentado de la fluidoj, kaj mezuroj estis prenitaj al la plej proksima 0.1 g. La pezo de ĉiu rato ankaŭ estis mezurita por kalkuli la gramojn da sukroza ingesta po kilogramo de korpa pezo. En tago 1 de la trinkperiodo, ratoj (n = 6 – 9) ricevis aliron al unu botelo da 5% (p / v) sukerido kaj unu botelo da akvo. Post 24 h, la sukerosa botelo estis anstataŭigita per dua akvo-botelo, kiu estis disponebla por la sekva 24 h. Ĉi tiu mastro ripetiĝis merkredon kaj vendredon. La ratoj havis senliman aliron al akvo en ĉiuj aliaj tagoj. Binge-simila konsumado de sukroso rezultigis pliigon en la tuta sukeroza konsumado (ml) kun la tempo (Suplementa Figuro 1) kaj estis akompanata de stabilaj bazlinaj trinkniveloj surbaze de korpa pezo [20 ± 5 g / kg de la 5% (w / v)] dum mallongtempaj [~ 4-semajnoj (13-drinkadaj sesioj)] kaj longtempe [ ~ 12 semajnoj (37 trinkaj sesioj)] trinkotaj periodoj. Aparta grupo de kontrolaj ratoj (n = 6 – 9) ricevis aliron al akvo en ambaŭ boteloj (t.e., neniu sukroso) sub la samaj kondiĉoj priskribitaj supre. La meza korpa pezo de kontrolo kaj sukeroza konsumado de ratoj fine de mallongdaŭra ekspozicio estis 405.7 ± 40.8 g kaj 426.4 ± 31.2 g respektive. Ĉe la fino de longtempa ekspozicio, la meza korpa pezo por kontrolo kaj sakrosa grupoj estis 578.8 ± 53.4 g kaj 600.2 ± 45.2 g.
Golgi-Cox Staining
Post la lasta trinka sesio ratoj estis transdonitaj de la besta instalaĵo por permesi prilaboron de la cerbaj specimenoj ĉe la histologia instalaĵo ĉe la Lernejo de Biomedicinaj Sciencoj, Universitato de Kvinslando (St Lucia, Aŭstralio). Ĉiuj aprobitaj mezuroj estis prenitaj por redukti streĉon dum transporto, poste, ratoj rajtis resaniĝi tra la nokto. La sekvan tagon, ratoj estis oferitaj per natria pentobarbita superdozo (60 – 80 mg / kg, ip Vetcare, Brisbano, Aŭstralio) kaj intrakardie perfuzitaj per ~ 300 ml artefarita cerebro-spina fluido kiu enhavis, (en mM): 130 NaCl, 3 KCl, 26 NaHCO3, 1.25 NaH2PO4, 5 MgCl2, 1 CaCl2, kaj 10 D-glukozo. Ĉiu besto tiam estis decapitata kaj la cerbo forigita kaj incuba en la mallumo en Golgi-Cox-solvo, kiu enhavis 5%-kalian-dikromaton, 5%-kalianan kromaton, kaj 5%-merkurian kloridon (ĉiuj kemiaĵoj de Sigma-Aldrich), kiuj estis kreitaj freŝaj 3-tagoj antaŭ ofero kiel priskribite antaŭe (Rutledge et al., 1969). Golgi-Cox-makula inkubacio kaj post-pretigaj metodoj estis modifitaj de Ranjan kaj Mallick (2010). Cerboj de mallongtempaj sukeroza konsumado de bestoj estis inkubitaj dum 6 tagoj ĉe 37 ° C, dum cerboj el longtempaj sukeroza konsumado de bestoj estis inkubitaj dum 10 tagoj, kun unu ŝanĝo al freŝa Golgi-Cox-solvo post 4 tagoj de inkubacio.
Sekvante kovadon, koronaj sekcioj de 300 μm estis tranĉitaj uzante vibran mikrotomon Zeiss Hyrax V50 (Carl Zeiss, Germanio). Tranĉaĵoj tiam estis metitaj sinsekve en 24-putajn telerojn plenigitajn kun 30% (p / v) sukerozo en bufrita salo de 0.1 M fosfato kaj procesitaj kiel skizitaj en (Ranjan kaj Mallick, 2010). Mallonge, sekcioj estis deshidratigitaj en 50% etanolo por 5 min, tiam metitaj en 0.1 M NH4OH-solvo por 30-min, lavita dufoje kun distilita akvo por 5-min kaj metita en la film-fiksilon de Fujihunt (Fujifilm, Singapuro) por 30-min en la mallumo. La tranĉaĵoj tiam estis lavitaj dufoje en distilita akvo por 2 min ĉiu kaj deshidratigitaj en 70, 90, 95, kaj 100% etanolo dufoje por 5 min ĉiu. La sekcioj tiam estis malplenigitaj en CXA-solvo (1: 1: 1-kloroformo: xileno: alkoholo) por 10 min kaj muntitaj en DPX (Sigma-Aldrich) sur lumbildoj Superfrost Plus (Menzel-Glaser, Lomb Scientific, Aŭstralio) kaj kovriloj. (Menzel-Glaser, Germanio). La lumbildoj estis lasitaj en la mallumo por sekigi la ĉambran temperaturon dum la nokto.
Neŭrala Selektado kaj Spurado en la Nukleaj Akcioj
Koronaj tranĉaĵoj inter bregma + 2.8 kaj + 1.7 estis enketitaj por MSNoj ene de la kerno kaj ŝelo de la NAc, uzante la flankan ventrikulon kaj la antaŭan komandon kiel limŝtonojn helpe de rato-cerba atlaso (Paxinos kaj Watson, 2007) (Figuro 1). La konturfunkcio en Neurolucida 7 (MBF Bioscience, VT, Usono) estis uzata por demarkado de la NAc-kerno kaj NAc-ŝelo en ĉiu tranĉaĵo (Figuro 2). Inter 2 kaj 9-neŭronoj per regiono per besto estis spurita por dendritaj longaj parametroj uzante 63x-celon aŭ por spine-densecoj (raportitaj kiel dornoj per 100 μm) uzante 100x-celon sur Zeiss Axioskop II (Carl Zeiss, Germanio) uzante aŭtomatan xyz etapo pelita de Neŭrolucida® 7-programaro (MBF Biosciences, VT, Usono). Ĉiu spuro estis farita per blindigita maniero koncerne kuracadon. Morfologiaj parametroj de impregnitaj neŭronoj de Golgi-Cox estis analizitaj laŭ maniero simila al antaŭaj raportoj (Klenowski et al., 2015).
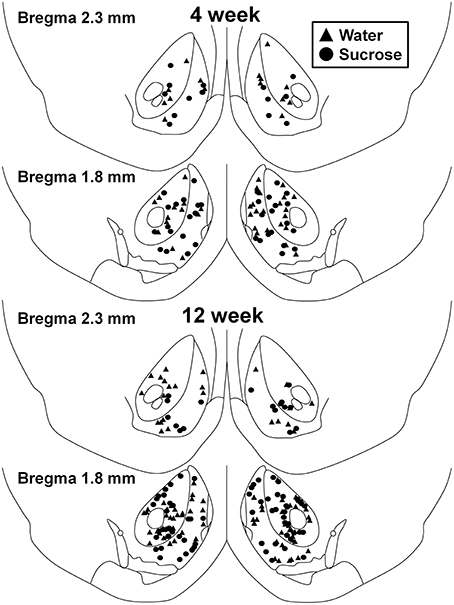
Figuro 1. Mapo montranta lokojn de mezaj spinecaj neŭronoj specimenigitaj de la kerno kaj akcentoj de 4 kaj 12-semajna sukroza konsumanta ratojn kaj aĝ-kongruajn aĝojn.. Supraj du paneloj montras lokojn de neŭronoj specimenitaj de la kerno de akcentoj kaj ŝelo de 4-semajna kontrolo (trianguloj) kaj sukrozo (rondoj) bestoj. Malsupraj du paneloj montras poziciojn de neŭronoj specimenitaj de bestoj de 12-semajna kontrolo (trianguloj) kaj sukrozo (rondoj).
Statistika Analizo
Meza kaj norma eraro de la meznombro (SEM) estis kalkulitaj por ĉiu datumaro kun la besto kiel n, uzante la mezajn morfometriajn datumojn de ĉiuj kernaj aŭ ŝelaj NAc MSNoj (n = 7 por NAc-ŝelo kaj n = 6 por NAc-kerno 4-semajno, n = 9 por 12-semajnaj grupoj). Kie indikite, senparaj duvostaj Studentoj t-provoj aŭ duflankaj ANOVAj kun Bonferroni-post-testoj estis farataj por ĉiuj analizoj implikantaj komparon de grupaj rimedoj, uzante GraphPad Prism-versio 6.02 (GraphPad Programaro, San-Diego, CA). Statistika signifo estis akceptita ĉe P <0.05. Ĉiuj datumoj en la sekcio de rezultoj estas prezentitaj kiel rimedoj ± SEM. Procentaj ŝanĝoj estas kalkulitaj relative al la kontrolvaloro.
rezultoj
Mezaj Spinecaj Neŭronoj de la Nucleus Accumbens Shell malpliiĝis Dendritic Longeco, Malpliiĝis Dendritic-Komplekseco sed Pliigita Meza Spina Denseco ĉe Distal Branch-Ordoj Sekvante Longan sed ne Baldaŭan Sukrozan Konsumon.
Post mallongdaŭraj (4-semajnoj) sukeroza konsumo, ne estis signifaj diferencoj en NAc-ŝelaj morfometriaj parametroj (Tabelo 1). Ekzistis ankaŭ neniuj signifaj diferencoj inter mallongperspektiva sukrosa konsumado kaj akvokontrolaj NAc-ŝelaj MSNoj en analizoj rilataj al centrifuga branĉa ordo. Nome, dendritaj segmentoj laŭ branĉa ordo (P = 0.4111), mezuma dendrita longo laŭ branĉa ordo (P = 0.5581) kaj mezuma spina denseco laŭ branĉa ordo (P = 0.2977, dudirektaj ANOVAoj) ne signife diferencis inter grupoj. Situa mapo montranta la proksimumajn poziciojn de la samplaj neŭronoj estas montrita en Figuro 1.
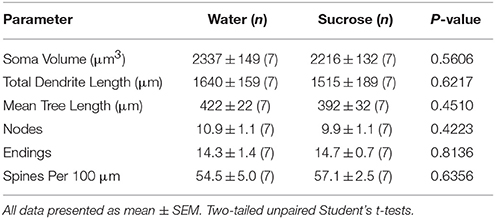
Tabelo 1. Ĝeneralaj morfologiaj parametroj de mezaj spinecaj neŭronoj de la kerno akcenta ŝelo de mallongperspektivaj sukerozaj ratoj kaj aĝaj kongruaj akvokontroloj.
Post longtempa (12 semajnoj) de sukeroza konsumo, la totala dendrita arba longo de NAc-ŝelaj MSNoj estis malpliigita de 21% kompare al akvo konsumanta kontrolojn (Akvo: 1827 ± 148 μm, n = 9; Sukroza 1449 ± 78 μm, n = 9, *P = 0.0384, duvostaj neparaj Studentoj t-testo, Figuro 2, Tablo 2). Komparo de la meznombro de dendritaj bifurcacioj (nodoj) kaj dendritaj finaĵoj inter la akvo- kaj sakrosa grupoj rivelis reduktitan (kvankam ne signifan) nivelon de dendrita komplikeco en NAc-ŝelaj MSNoj (nodoj: Akvo 12.9 ± 1.4 n = 9, Sukroza 10.1 ± 0.8 n = 9, P = 0.0879; finaĵoj: Akvo 17.9 ± 1.4 n = 9, Sukroza 14.8 ± 0.7 n = 9, P = 0.0657, duvostaj neparaj Studentoj t-testo, Tabelo 2). Ne estis ŝanĝo en soma volumo (P = 0.9400), meznombro de dendrita trunkoP = 0.1646) aŭ totala spina denseco (P = 0.3662) en NAc-ŝelaj MSNoj de longtempaj sukerozaj konsumantaj ratoj kompare al akvokontroloj. Ĉi tiuj morfometriaj parametroj estas detalaj en Tabelo 2.
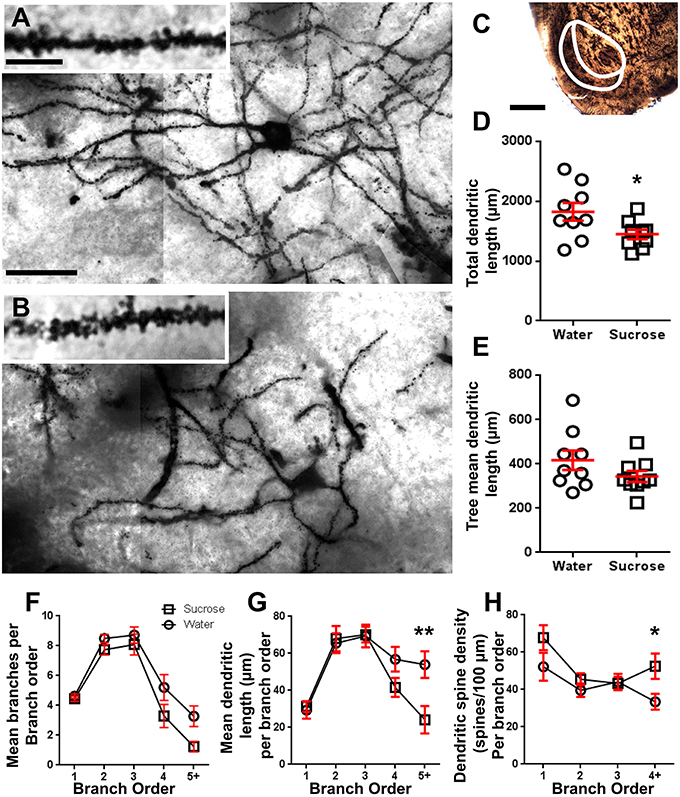 Figuro 2. Malpliiĝis dendrita arba longo kaj pliigita distala dendrita spina denseco de mezaj spinecaj neŭronoj (MSNoj) de la nukleo accumbens (NAc) ŝelo de longperspektivaj sukerozaj ratoj kompare kun kontrolaj ratoj. (A, B) montru reprezentojn de kontrolo (supro) kaj longtempa (12 semajno) sukrosa (malsupre) traktita brila kampo z-staka mozaiko de Golgi-trempitaj MSN-oj el la ŝelo de NAc (63x-pligrandiĝo). Insekto de (A, B) montras kontrolon kaj longperspektivajn brilajn bildojn de sukrozo de Golgi-trempitaj MSN-dendritoj kaj dendritaj spinoj de la ŝelo de NAc (100x-pligrandiĝo). (C) montras la anatomiajn regionojn, kiujn oni specimenigis de MSN en ĉi tiu studo. (D) montras disĵetilon de malpliiĝinta totala MSN-dendrida arbo (meznombro ± SEM) de la ŝelo de NAc en longtempaj sukosaj bestoj (kvadratoj) kompare kun kontroloj (cirkloj), senparaj studentoj t-testo, *P <0.05, n = 9; kontrolo kaj n = 9; 12-semajna sukroso. (E) montras disvastigitan intrigon de neŝanĝita meznombro de MSN-dendrita arbokolono (meznombro ± SEM) de la ŝelo de NAc dum longtempaj sukosaj bestoj (kvadratoj) kompare kun kontroloj (rondoj), neparencaj studentoj t-estest, P > 0.05, n = 9; kontrolo kaj n = 9; 12-semajna sukroso. Analizo de branĉaj ordoj (meznombro ± SEM) de dendrita segmento por ĉiu branĉa mendo (F), mezuma dendrita longo laŭ branĉa ordo (G) kaj dendrita densa spino-denseco laŭ branĉa ordo (H). Longtempa sukerozo malpliigis dendritan longon ĉe distalaj branĉaj ordoj (5 +) kaj pliigis dendritan spinan densecon ĉe distalaj branĉaj ordoj (4 +) kompare kun kontroloj (G, H), dudirektaj ANOVAJ kun post-testoj de Bonferroni, *P <0.05, **P <0.01, n = 9; kontrolo kaj n = 9; longtempa sukerozo. Skalaj Trinkejoj: (A, B) = 20 μm; enso de (A, B) = 10 μm; (C) = 1 mm.
Figuro 2. Malpliiĝis dendrita arba longo kaj pliigita distala dendrita spina denseco de mezaj spinecaj neŭronoj (MSNoj) de la nukleo accumbens (NAc) ŝelo de longperspektivaj sukerozaj ratoj kompare kun kontrolaj ratoj. (A, B) montru reprezentojn de kontrolo (supro) kaj longtempa (12 semajno) sukrosa (malsupre) traktita brila kampo z-staka mozaiko de Golgi-trempitaj MSN-oj el la ŝelo de NAc (63x-pligrandiĝo). Insekto de (A, B) montras kontrolon kaj longperspektivajn brilajn bildojn de sukrozo de Golgi-trempitaj MSN-dendritoj kaj dendritaj spinoj de la ŝelo de NAc (100x-pligrandiĝo). (C) montras la anatomiajn regionojn, kiujn oni specimenigis de MSN en ĉi tiu studo. (D) montras disĵetilon de malpliiĝinta totala MSN-dendrida arbo (meznombro ± SEM) de la ŝelo de NAc en longtempaj sukosaj bestoj (kvadratoj) kompare kun kontroloj (cirkloj), senparaj studentoj t-testo, *P <0.05, n = 9; kontrolo kaj n = 9; 12-semajna sukroso. (E) montras disvastigitan intrigon de neŝanĝita meznombro de MSN-dendrita arbokolono (meznombro ± SEM) de la ŝelo de NAc dum longtempaj sukosaj bestoj (kvadratoj) kompare kun kontroloj (rondoj), neparencaj studentoj t-estest, P > 0.05, n = 9; kontrolo kaj n = 9; 12-semajna sukroso. Analizo de branĉaj ordoj (meznombro ± SEM) de dendrita segmento por ĉiu branĉa mendo (F), mezuma dendrita longo laŭ branĉa ordo (G) kaj dendrita densa spino-denseco laŭ branĉa ordo (H). Longtempa sukerozo malpliigis dendritan longon ĉe distalaj branĉaj ordoj (5 +) kaj pliigis dendritan spinan densecon ĉe distalaj branĉaj ordoj (4 +) kompare kun kontroloj (G, H), dudirektaj ANOVAJ kun post-testoj de Bonferroni, *P <0.05, **P <0.01, n = 9; kontrolo kaj n = 9; longtempa sukerozo. Skalaj Trinkejoj: (A, B) = 20 μm; enso de (A, B) = 10 μm; (C) = 1 mm.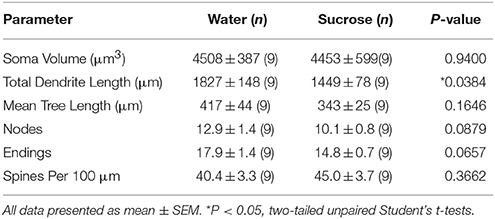
Tabelo 2. Ĝeneralaj morfologiaj parametroj de mezaj spinecaj neŭronoj de la nukleo akcenta ŝelo de longtempaj sukerozo konsumantaj ratojn kaj aĝ-kongruajn akvajn kontrolojn.
Sekve de la karakterizado de la ĝenerala dendrita morfologio de longtempaj sukerozo konsumanta NAc-ŝelajn MSNojn, ni analizis dendritajn arborizojn kaj spinajn densecojn koncerne iliajn branĉajn ordojn. Nia ampleksa takso de la dendritaj arboj kvantigis la nombron da dendritaj segmentoj laŭ branĉa ordo, la mezuma longo de dendritaj segmentoj laŭ branĉa ordo kaj mezuma spina denseco laŭ branĉa ordo de NAc-ŝelaj MSNoj de akvokontrolo kaj longtempa sukrozo konsumanta ratojn. Resumo pri la datumoj kaj analizo de branĉaj ordoj estas prezentita en Tabelo 3.
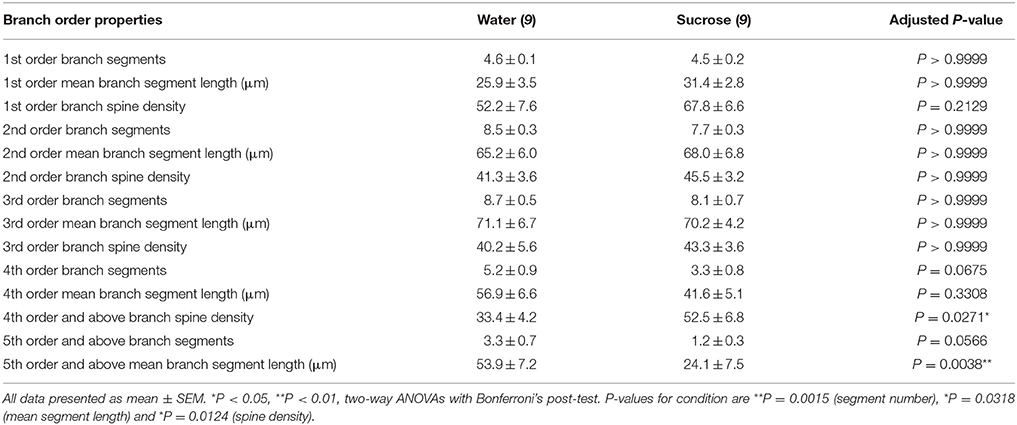 Tabelo 3. Branĉaj ordoj de mezaj spinaj neŭronoj el longtempaj sukerozo kaj akvo trinkantaj ratoj.
Tabelo 3. Branĉaj ordoj de mezaj spinaj neŭronoj el longtempaj sukerozo kaj akvo trinkantaj ratoj.La meznombro de dendritaj branĉaj nombroj laŭ branĉa ordo de NAc-ŝelaj MSNoj estis signife reduktita en longtempaj sukerozaj konsumantaj ratoj kompare al akvaj kontroloj (**)P = 0.0015, dudirekta ANOVA). Post-testoj de Bonferroni malkaŝis tendencon al reduktita nombro de branĉaj segmentoj ĉe 4th (Akvo: 5.2 ± 0.9, n = 9; Sukroza 3.3 ± 0.8, n = 9, P = 0.0675, Figuro 2F, Tablo 3), kaj 5a ordo kaj supre branĉaj ordoj (Akvo: 3.3 ± 0.7, n = 9; Sukroza 1.2 ± 0.3, n = 9, P = 0.0566, Figuro 2F, Tablo 3). La meza dendrita dendro-longo laŭ branĉa ordo de NAc-ŝelaj MSNoj ankaŭ estis signife reduktita en longtempaj sukeroza konsumado de ratoj kompare al akvokontroloj (*P = 0.0444, dudirekta ANOVA). Post-testoj de Bonferroni montris redukton de 55% ĉe filioj de 5a ordo kaj preter (Akvo: 53.9 ± 7.2 μm, n = 9; Sukroza 24.1 ± 7.5 μm, n = 9, **P = 0.0038, Figuro 2G, Tablo 3).
Analizo de branĉaj ordoj montris signifan kreskon de la dendrita spina denseco de NAc-ŝelaj MSNoj de longtempa sukeroza konsumado de ratoj kompare kun kontroloj (*P = 0.0124, dudirekta ANOVA). Post-testoj de Bonferroni montris spinan densecan kreskon de 57% ĉe distalaj 4-ordaj branĉoj kaj plu (Akvo: 33.4 ± 4.2, n = 9; Sukroza 52.5 ± 6.8, n = 9, P = 0.0271 *, enmeto de Figuroj 2A, B, H, Tablo 3). Reprezentaj bildoj de entuta MSN-arkitekturo kaj distala spina denseco (enigita) estas bildigitaj en Figuroj 2A, B.
Kunigitaj, ĉi tiuj rezultoj indikas, ke mallongperspektiva sukerozo havas malmultan efikon al morfologiaj parametroj de MSNoj ene de la ŝelo de NAc. Tamen, post plilongigita konsumo, estas signifa malkresko en la neuronal arba longo kaj komplikeco, precipe en distalaj dendritaj branĉoj. Samtempaj distalaj dornaj kreskoj ankaŭ estas videblaj en NAc-ŝelaj MSNoj de longtempa sukeroza konsumado de ratoj.
Mezaj Spinecaj Neŭronoj de la Kerno de Nukleaj Akcentoj reduktis Branĉan Kompleksecon post Longa sed ne Baldaŭa Sukrosa Konsumo
Sekvante mallongperspektivan sukerozan konsumon, ne estis signifaj diferencoj en NAc-kernaj MSN-morfometriaj parametroj (Tabelo) 4). Ankaŭ ne estis signifaj diferencoj inter 4-semajna sukerosa konsumo kaj akvo-kontrolaj MSN-kernoj en analizoj rilataj al centrifuga branĉa ordo. Nome, dendritaj segmentoj laŭ branĉa ordo (P = 0.7717), mezuma dendrita longo laŭ branĉa ordo (P = 0.2096), kaj mezuma spina denseco laŭ branĉa ordo (P = 0.3521, dudirektaj ANOVAJ) ne diferencis inter grupoj.
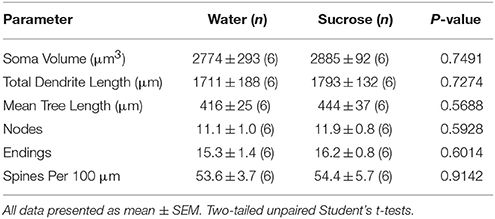 Tabelo 4. Ĝeneralaj morfologiaj parametroj de mezaj spinecaj neŭronoj de la kerno akcenta kerno de mallongperspektivaj sukerozaj ratoj kaj aĝaj kongruaj akvokontroloj..
Tabelo 4. Ĝeneralaj morfologiaj parametroj de mezaj spinecaj neŭronoj de la kerno akcenta kerno de mallongperspektivaj sukerozaj ratoj kaj aĝaj kongruaj akvokontroloj..Prolongigita sukeroza konsumo ankaŭ havis neniun signifan sur NAc-kernaj MSN-morfometraj parametroj 5). La meznombro de dendritaj branĉaj nombroj laŭ branĉa ordo de NAc-kernaj MSNoj reduktiĝis signife en longtempaj sukerozaj konsumantaj ratoj kompare al akvaj kontroloj (*P = 0.0416, dudirekta ANOVA), tamen ne estis signifaj diferencoj en meznoma dendrita longo laŭ branĉa ordo (P = 0.0995) kaj mezuma spina denseco laŭ branĉa ordo (P = 0.4888, dudirektaj ANOVAJ) inter MSNoj en la kerno de NAc de longtempaj sukeroza konsumado de ratoj kompare kun akvokontroloj. Prenitaj kune, niaj datumoj montras, ke la NAc-kerno ne tiom respondas al longtempa konsumo de sukerozo kompare al MSN-oj de la regiono de la ŝeloj de NAc.
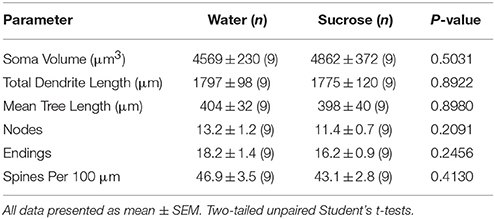 Tabelo 5. Ĝeneralaj morfologiaj parametroj de mezaj spinecaj neŭronoj de la kerno akcenta kerno de longtempaj sukerozo konsumantaj ratojn kaj aĝ-kongruajn akvajn kontrolojn.
Tabelo 5. Ĝeneralaj morfologiaj parametroj de mezaj spinecaj neŭronoj de la kerno akcenta kerno de longtempaj sukerozo konsumantaj ratojn kaj aĝ-kongruajn akvajn kontrolojn.diskuto
La pliigita havebleco de tre dolĉigita manĝaĵo en la okcidenta dieto ne nur kontribuis al la pliigita prevalenco kaj ekonomia ŝarĝo de obezeco kaj tipo II-diabeto, sed ankaŭ kaŭzis aperon de manĝaj malordoj kiel binge manĝado (Swanson et al., 2011; Kessler et al., 2013; Davis, 2015). Kvankam la toksomaniaj ecoj de sukeroj inkluzive de fruktozo kaj sukerozo restas spekulativaj, ekzistas frapa simileco en la kondutaj kaj neŭralaj korelacioj kiuj manifestiĝas kiel rezulto de tro manĝado kaj plilongigita drogo-uzo. (Avena et al., 2008, 2011). Krome sukero aktivigas la rekompencajn cirkvitojn de la cerbo en maniero simila al misuzo de drogoj (Volkow et al., 2012), kaj rezultoj de homaj studoj sugestas, ke sukero kaj dolĉeco povas estigi avidojn kompareblajn kun grando al tiuj induktitaj de toksomaniuloj kiel alkoholo kaj nikotino (Volkow et al., 2012). Ni tial uzis modelon de binge-sukarosa konsumo en ratoj por determini la efikojn de mallonga (4-semajnoj) kaj longperspektivaj (12-semajnoj) sukrosa konsumo sur neŭtrala morfologio de MSNoj en la NAc, ŝlosila komponento de la interkovra rekompenco. tio estas modulata de sukero kaj toksomaniaj drogoj. Ni montras, ke MSN-oj de la NAc-ŝelo de kronikaj longperspektivaj konsumantaj ratoj signife malpliigis dendritan longon kaj kompleksecon, sed pliigis distalan dendritan spinan densecon. Longtempa sukerozo havis neniun efikon sur la morfologio de MSNoj de la NAc-kerno, dum mallongdaŭra sukerozo ankaŭ havis neniun signifan efikon sur MSN-morfologio de la NAc-kerno aŭ ŝelo. Ĉi tiuj rezultoj ne nur montras rektan efikon de plilongigita binge-simila sukroza konsumado sur neuronal morfologio de NAc-ŝelaj MSNoj, sed ankaŭ emfazas la eble damaĝajn konsekvencojn de la plilongigita konsumado de alta sukero enhavanta dietojn.
La NAc, kiu formas parton de la ventrala striatumo, konsistas ĉefe el MSN-oj, kiuj estas morfologie karakterizitaj kiel mezgrandaj neŭronoj kun ampleksaj dendritaj arborizoj kaj alta spina denseco (Kemp kaj Powell, 1971; Graveland kaj DiFiglia, 1985; Rafols et al., 1989; Kawaguchi et al., 1990). Glutamatergaj kaj dopaminergiaj neŭronoj estas la du primaraj aferentaj enigaĵoj al la NAc, ĉefe kontakti la dendritajn ŝafojn kaj spinojn de MSNoj. (Groves, 1980; Kaiya kaj Namba, 1981; Groves et al., 1994). Specife, la NAc-ŝelo kaj kerno ricevas glutamatergajn enigaĵojn de funkcie distingaj kortikaj areoj (Brog et al., 1993). La NAc-ŝelo estas ankaŭ avidita de ekscitataj aferentoj de subkortikaj regionoj kiel la hipokampo, talamo kaj bazolateral amigdalo (Brog et al., 1993; Wright kaj Groenewegen, 1995). Antaŭaj studoj pruvis, ke ĉi tiuj glutamatergaj enigaĵoj ludas pivotan rolon en instigo kaj cel-direktitaj kondutoj kiel manĝaĵo kaj rekompenco (Maldonado-Irizarry et al., 1995; Kelley kaj Swanson, 1997; Reynolds kaj Berridge, 2003; Rikardo kaj Berridge, 2011). La alia superreganta enigo de nafaj MSNoj devenas de dopaminergiaj aferentoj kiuj projektas de la ventra tegmenta areo (Lindvall kaj Björklund, 1978; Veening et al., 1980; Kalivas kaj Miller, 1984). Interese, antaŭaj studoj uzantaj similajn modelojn de intermita sukero-aliro montris, ke la rezulta konsumo de binge-rezulto pliiĝas en eksterĉela dopamino en la NAc simile (kvankam en plej malgranda mezuro) al drogoj de misuzo. (Rada et al., 2005; Avena et al., 2006), kaj povas moduli dopamin-ricevan esprimon (Colantuoni et al., 2001, 2002) en la NAc-kerno kaj ŝelo. Interese, ke binge-simila konsumado de sukerozo kaŭzas eskalon de konsumado kun la tempo simile al mem-administrado de drogoj de misuzo kiel kokaino kaj heroojn (Ahmed kaj Koob, 1998; Ahmed et al., 2000, 2003) kiu estas asociita kun la evoluo de "toksomaniulo".
Nia analizo de branĉa ordo-morfometrio montras, ke la totala redukto de dendrita longo de NAc-ŝelaj MSN-oj kaŭzitaj de longtempa sukera konsumado, rezultas ĉefe el reduktoj de la komplekseco de distalaj branĉaj ordoj. Ni observis reduktitajn distalajn branĉojn (4th kaj 5th-ordo kaj super branĉaj ordoj) kaj signife reduktitan mezuman longon je 5th-ordo kaj super dendritoj, kombinitaj kun pliigitaj dornaj densecoj ĉe ĉi tiuj branĉaj ordoj. Ofta faktoro probable influanta ĉi tiun specon de dendrita restrukturado inkluzivas ŝanĝojn en sinaptika konektebleco kaj / aŭ funkcio (Russo et al., 2010). Antaŭaj studoj montris, ke glutamatergaj sinapsoj sur MSNoj formiĝas ĉefe sur dornoj, precipe ĉe dendraj distaloj (Groenewegen et al., 1999). Aldone, kun-lokalizo de dopamina kaj glutamatergiaj enigaĵoj de la prefrontal-kortekso (Sesack kaj Pickel, 1992), hipocampo (Totterdell kaj Smith, 1989; Sesack kaj Pickel, 1990), kaj amigdala (Johnson et al., 1994) estis observitaj sur dendritaj spinoj de MSNoj. Ĉi tiuj observoj kombinitaj kun la pliigita spina denseco post longtempa sukeroza konsumo vidita en nia studo, subtenas la formadon de pliigitaj ekscitaj enigoj. Sekve, la ebleco ekestas, kie konstantaj efikoj kaŭzitaj de plilongigita binge-simila sukerosa konsumado povus faciligi pliigitan ekscitigan sinaptan aktivecon ĉe la distalaj dendritoj de MSNoj en la NAc-ŝelo. Konsekvence, redukto kaj / aŭ retiriĝo de distalaj dendritoj povas rezulti per sinaptika homeostata mekanismo (Reissner kaj Kalivas, 2010), tamen tio restas determinita.
Estas interese rimarki, ke Crombag kaj kolegoj montris, ke ne estis spina denseco pliigita en la ŝelo de NAc sekvante 4-semajnan sukerozan konsumon per la naz-poke-memadministra paradigmo malgraŭ pli fortika akiro kaj pli alta responda indico al sukerozo kompare. kun amfetamino (Crombag et al., 2005). Ilia observado de foresto de ŝanĝo en spina denseco ĉe 4-semajnoj spegulas niajn trovojn. Kontraŭe, tamen, nia studo pruvas, ke post longa daŭro (12-semajno) ekspozicio al kronika sukeroza konsumo, estas signifa kresko de distala spina denseco sur la MSNoj de la sukerozo-spertaj ratoj. Plue, nia laboratorio antaŭe montris, ke longtempa (12 semajno) sukeroza konsumo faciligas diferencan farmakologian respondon al farmakoterapeŭtoj, kiuj estis montritaj moduli dopaminajn kaj acetilcolinajn respondojn ĉe la nivelo de la NAc (Shariff et al., En gazetaro). Kune, ĉi tio sugestas, ke longdaŭraj (12-semajnoj kaj pli postaj) sukroza ekspozicio, pli ĝuste reflekta de real-mondaj scenoj, rezultigas morfologiajn adaptiĝojn ĉe la nivelo de NAc.
Koncerne al drogoj misuzo, ripeta eksponiĝo al diversaj drogoj produktas daŭrajn ŝanĝojn en la strukturo de dendritoj kaj dendritaj spinoj. Ekzemple, amfetaminoj kaj kokaino ambaŭ pliigas spinan densecon en la NAc kaj en ŝelo kaj kerno (Robinson kaj Kolb, 2004). Ekspozicio al nikotino ankaŭ pruviĝis pliigi spinan densecon en la ŝelo de NAc. Al la inversa, morfina ekspozicio kondukas al malpliigo de la spina denseco kaj dendrita branĉokomplekseco (Robinson kaj Kolb, 2004). Rilate al longtempa konsumo de sacarose, ni observis kreskon de spina denseco simila al amfetamino, kokaino kaj nikotino kaj kontraŭe al la efiko de morfino. Tamen, male al amfetamino kaj kokaino, sed simila al nikotino, la kresko de spina denseco dum longtempa ekspozicio al sukerozo estas limigita al la ŝelo de NAc. Ankaŭ estas interese, ke ŝanĝoj en ambaŭ dendrita branĉado (Robinson kaj Kolb, 1999) kaj spina denseco (Li et al., 2003) produktita de amfetamino aŭ kokaino estas limigita al distalaj dendritoj de MSNoj en la NAc, kio reflektas la trovojn en nia studo. Plie, kaj korodivebla al la ŝanĝoj priskribitaj supre, ankaŭ konsumo de sukerozo antaŭe estis montrita pliigante ekscitigan sinaptan forton sur akumulaj dopaminaj neŭronoj (Stuber et al., 2008b) same kiel aliajn komponentojn de la mezolimbia rekompenca vojo (Stuber et al., 2008a; Chen et al., 2010). Kunigitaj, ĉi tio pozas sukerozon kiel potencan modulatoron de neŭrma morfologio sekve plilongigitan pezan uzadon, kiu similas al la efikoj observataj de drogoj de misuzo.
Kvankam pliaj esploroj estas bezonataj por malkovri la ĉelajn kaj sinaptikajn mekanismojn kontribuantajn al la morfologiaj ŝanĝoj viditaj en ĉi tiu studo, niaj rezultoj pruvas signifajn neuronajn efikojn kaŭzitajn de longtempa sukeroza konsumo. En aparta, konsidero ne ekzamenita en nia studo estas ĉu la observitaj morfologiaj efikoj de sukarido povas ankaŭ esti eliritaj per ne-kaloriaj edulkorantoj kiel sakarino. Tiurilate gravas, ke Lenoir kaj kolegoj montris, ke intensa dolĉeco superas kokainan rekompencon, ĉu ĝi estas generita de sakarino aŭ sukerozo (Lenoir et al., 2007). Plue, lastatempa studo publikigita de nia laboratorio (Shariff et al., En gazetaro) pruvas, ke varenicline, nikotinika acetilkolina parta agonisto reduktis ambaŭ sukerozan kaj sakarinan konsumon en ronĝuloj sekvante la saman longtempan intermitan aliron-reĝimon uzatan en la nuna studo. Interese, antaŭaj studoj montris similecojn en la akraj efikoj de ne-kaloriaj edulkorantoj kiel sakarino kaj sukerozo ĉe la nivelo de NAc (Scheggi et al., 2013; Tukey et al., 2013; Carelli kaj Okcidenta, 2014). Tamen, pliaj studoj bezonas determini, ĉu ne-kaloriaj dolĉigiloj povas indukti longdaŭrajn efikojn similajn al ŝanĝoj en la morfologio de la ŝeloj de NAc-ŝelo kaŭzitaj de longtempa sukerozo raportita ĉi tie.
La manko de efiko al NAc MSN-morfologio sekvanta mallongperspektivan sukerozon, elstarigas la gravecon de efektivigado de longtempaj studoj por taksi la efikon de plilongigita fitraktado de drogoj aŭ naturaj rekompencoj kiel sukrozo. Koncerne dependecon, ne nur ripetaj cikloj de ingesta konsumado kaj sindeteno estas ŝlosilaj komponentoj de la toksomania ciklo, kreskanta atestaĵo malkaŝis, ke la transiro al dependeco estas progresiva procezo, kiu ofte okazas dum plilongigita tempodaŭro. Kvankam la toksomaniaj ecoj de sukeroj restas necertaj, la plaŭdemo de toksomanio al aliaj ne-drogaj rekompencoj kiel sekso, vetludo kaj manĝaĵo estas ĉiam pli esploritaj. La rezultoj de ĉi tiu studo aldonas meriton al la hipotezo, ke sukeroj kiel ekzemple sukerozo eble havas toksajn propraĵojn sekvante longtempan, similan konsumon. Niaj rezultoj ankaŭ havas implicojn por la kreskanta nombro de infanoj kaj adoleskantoj, kiuj konservas malsanajn manĝajn kutimojn (alta sukero kaj binge manĝado) en plenaĝecon. Lige al pliigita risko de evoluigi metabolajn efikojn, eblas ankaŭ ke neurologiaj kaj psikiatriaj konsekvencoj influantaj humoro kaj instigo povus rezulti ankaŭ de ĉi tiuj kondutoj.
Aŭtoro Kontribuoj
Partoprenis esploradon: PK, SB. Efektivigitaj eksperimentoj: PK, MS, AB, MF, EM. Analizo de datumoj: PK, MF, MS. Interpretis la datumojn kaj kontribuis al verkado de la manuskripto: PK, MS, MF, EM, MB, SB. Ĉiuj aŭtoroj legis kaj aprobis la finan manuskripton por sendado.
Konflikto pri Interesa Rakonto
La aŭtoroj deklaras, ke la esplorado estis farita sen manko de komercaj aŭ financaj rilatoj, kiujn oni povus konsideri kiel ebla konflikto de intereso.
La revizoroj SC, SA kaj pritraktanta Redaktoro deklaris sian apartan aliĝon, kaj la Redaktoro pri pritraktado deklaras, ke la procezo tamen plenumas la normojn de justa kaj objektiva revizio.
Dankojn
Ĉi tiu laboro estis subtenita de financado de subvencioj de la Aŭstralia Esplora Konsilio (FT1110884) al SB kaj la Nacia Sano kaj Medicina Esplora Konsilio (1061979) al SB kaj MB.
Suplementa Materialo
La Suplementa Materialo por ĉi tiu artikolo povas esti trovita enrete ĉe: http://journal.frontiersin.org/article/10.3389/fnbeh.2016.00054
Suplementa Figuro 1. Sukero kaj konsumado de prefero de 4 kaj 12 semajnaj sukerozaj konsumantaj ratoj. (A, B) montru grimpadon en totala sukeroza konsumado (ml) dum 4 kaj 12 semajnoj da ekspozicio. (C, D) montru altan preferon por sukerozo super akvo dum periodoj de sukeroza prezento.
Referencoj
Ahmed, SH, kaj Koob, GF (1998). Transiro de modera al troa konsumado de drogoj: ŝanĝo en hedonan fikspunkton. scienco 282, 298-300. doi: 10.1126 / science.282.5387.298
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Ahmed, SH, Lin, D., Koob, GF, kaj Parsons, LH (2003). Eskalado de kokain-memadministrado ne dependas de ŝanĝita induktita kun kokaino-kerno kaŭzas dopaminajn nivelojn. J. Neurochem. 86, 102-113. doi: 10.1046 / j.1471-4159.2003.01833.x
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Ahmed, SH, Walker, JR, kaj Koob, GF (2000). Persista pliiĝo en la instigo preni heroinon en ratoj kun historio de drogakcelado. Neuropsychofarmacology 22, 413–421. doi: 10.1016/S0893-133X(99)00133-5
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Arslanian, S. (2002). Tipo 2-diabeto en infanoj: klinikaj aspektoj kaj riskaj faktoroj. Horma Res 57 (Suppl. 1), 19 – 28. doi: 10.1159 / 000053308
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Avena, NM, Bocarsly, ME, Hoebel, BG, kaj Gold, MS (2011). Superkovroj en la nosologio de misuzo kaj troigo: la tradukaj implicoj de "manĝa toksomanio". Curr. Drug Abuse Rev. 4, 133-139. doi: 10.2174 / 1874473711104030133
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Avena, NM, Rada, P., kaj Hoebel, BG (2008). Evidenteco por sukero toksomanio: kondutaj kaj neŭkemiaj efikoj de intermita, troa konsumado de sukero. Neurosci. Biobehav. Rev. 32, 20-39. doi: 10.1016 / j.neubiorev.2007.04.019
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Avena, NM, Rada, P., Moise, N., kaj Hoebel, BG (2006). Sukcena ŝafa nutrado laŭ bengala horaro liberigas akombensan dopaminon ree kaj forigas la acetilkolinan satigan respondon. Neurokienco 139, 813 – 820. doi: 10.1016 / j.neuroscience.2005.12.037
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Benton, D. (2010). La plaŭdemo de sukero toksomanio kaj ĝia rolo en obezeco kaj manĝadaj malordoj. Kliniko. Nutr. 29, 288 – 303. doi: 10.1016 / j.clnu.2009.12.001
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Bray, GA, kaj Popkin, BM (2014). Dieta sukero kaj korpa pezo: ĉu ni atingis krizon en la epidemio de obezeco kaj diabeto ?: sano estas damnita! Verŝu sur la sukeron. Diabeta Prizorgo 37, 950 – 956. doi: 10.2337 / dc13-2085
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Brog, JS, Salyapongse, A., Deutch, AY, kaj Zahm, DS (1993). La padronoj de afera senĝenigo de la kerno kaj ŝelo en la parto "akciuloj" de la ventra striatumo: imunohistokemia detekto de retrograde transportita fluoro-oro. J. Comp. Neŭrolo. 338, 255 – 278. doi: 10.1002 / cne.903380209
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Bucher Della Torre, S., Keller, A., Laure Depeyre, J., kaj Kruseman, M. (2015). Sukero-dolĉigitaj trinkaĵoj kaj obesidad risko en infanoj kaj adoleskantoj: sistema analizo pri kiel metodika kvalito povas influi konkludojn. J. Acad. Nutr. Dieto. [Epub antaŭ presaĵo]. doi: 10.1016 / j.jand.2015.05.020
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Carelli, RM, kaj West, EA (2014). Kiam bona gusto fariĝas malbona: neŭralaj mekanismoj sub la apero de negativa afekto kaj asociita natura rekompenca malvalorigo de kokaino. Neuropharmacology 76 (Pt B), 360 – 369. doi: 10.1016 / j.neuropharm.2013.04.025
Chen, BT, Hopf, FW, kaj Bonci, A. (2010). Sinaptika plastikeco en la mezolimbia sistemo: terapiaj implikaĵoj por fitraktado. Ann. NY Acad. Sci. 1187, 129-139. doi: 10.1111 / j.1749-6632.2009.05154.x
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Colantuoni, C., Rada, P., McCarthy, J., Patten, C., Avena, NM, Chadeayne, A., et al. (2002) Evidentas, ke intermita kaj troa sukero konsumas endogenan opioidan dependecon. Obes. Res. 10, 478 – 488. doi: 10.1038 / oby.2002.66
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Colantuoni, C., Schwenker, J., McCarthy, J., Rada, P., Ladenheim, B., Cadet, JL, et al. (2001) Troa konsumado de sukero ŝanĝas ligon al dopamino kaj mu-opioidaj riceviloj en la cerbo. Neuroreporto 12, 3549–3552. doi: 10.1097/00001756-200111160-00035
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Crombag, HS, Gorny, G., Li, Y., Kolb, B., kaj Robinson, TE (2005). Kontraŭaj efikoj de amfetamina mem-administrada sperto sur dendritaj spinoj en la medial kaj orbita prefrontal-kortekso. Cereb. Cortex 15, 341 – 348. doi: 10.1093 / cercor / bhh136
Dabelea, D., Mayer-Davis, EJ, Saydah, S., Imperatore, G., Linder, B., Divers, J., et al. (2014) Antaŭvaloro de tipo 1 kaj tipo 2-diabeto inter infanoj kaj adoleskantoj de 2001 ĝis 2009. JAMO 311, 1778 – 1786. doi: 10.1001 / jama.2014.3201
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Davis, C. (2015). La epidemiologio kaj genetiko de malforta manĝa malordo (BED). CNS-spektanto. 20, 522-529. doi: 10.1017 / s1092852915000462
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Ford, ES, kaj Dietz, WH (2013). Tendencoj pri konsumado de energio inter plenkreskuloj en Usono: trovoj de NHANES. Estas. J. Clin. Nutr. 97, 848 – 853. doi: 10.3945 / ajcn.112.052662
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Fryar, CD, Carroll, MD, kaj Ogden, CL (2014). Prevaloro de Troa Pezeco, Obezeco, Kaj Ekstrema Obeo Inter Plenkreskuloj: Usono, 1960 – 1962 tra 2011 – 2012. Atlanta, GA: CDC.
Graveland, GA, kaj DiFiglia, M. (1985). La ofteco kaj distribuo de mezgrandaj neŭronoj kun indentaj kernoj en la primato kaj ronĝa neostriatumo. Brain Res. 327, 307–311. doi: 10.1016/0006-8993(85)91524-0
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Groenewegen, HJ, Wright, CI, Beijer, AV, kaj Voorn, P. (1999). Konverĝo kaj apartigo de ventraj striaj enigoj kaj rezultoj. Ann. NY Acad. Sci. 877, 49–63. doi: 10.1111/j.1749-6632.1999.tb09260.x
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Groves, PM (1980). Sinaptaj finaĵoj kaj iliaj postsinaptaj celoj en neostriatum: sinaptaj specialiĝoj malkaŝitaj de analizo de seriaj sekcioj. Prok. Natl. Acad. Sci. Usono 77, 6926-6929. doi: 10.1073 / pnas.77.11.6926
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Groves, PM, Linder, JC, kaj Young, SJ (1994). Dopaminergiaj akonoj markitaj kun hidroksidopamina: tridimensiaj rekonstruoj de akonoj, sinapsoj kaj postsinaptaj celoj en rato-neostriato. Neurokienco 58, 593–604. doi: 10.1016/0306-4522(94)90084-1
Hu, FB (2013). Solvita: estas sufiĉa scienca evidenteco, ke malpliiĝanta sukero-dolĉigita trinkaĵkonsumo malpliigos la prevalencon de obezeco kaj rilataj obesaj malsanoj. Obes. Rev. 14, 606 – 619. doi: 10.1111 / obr.12040
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Johnson, LR, Aylward, RL, Hussain, Z., kaj Totterdell, S. (1994). Eniro de la amigdala al la rato-kerno acumbens: ĝia rilato kun imunoreaktiveco de tirozina hidroksilasa kaj neŭronoj identigitaj. Neurokienco 61, 851–865. doi: 10.1016/0306-4522(94)90408-1
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Kaiya, H., kaj Namba, M. (1981). Du specoj de dopaminergiaj nervaj finaĵoj en la rato neostriatum. Ultraestruktura studo. Neurosci. Lett. 25, 251–256. doi: 10.1016/0304-3940(81)90400-6
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Kalivas, PW, kaj Miller, JS (1984). Neŭrotensinaj neŭronoj en la ventrala tegmenta areo projektas al la media kerno accumbens. Brain Res. 300, 157–160. doi: 10.1016/0006-8993(84)91351-9
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Kawaguchi, Y., Wilson, CJ, kaj Emson, PC (1990). Projekciaj subtipoj de rataj neostriaj matricĉeloj rivelitaj per intracelula injekto de biocitino. J. Neurosci. 10, 3421-3438.
Kelley, AE, kaj Swanson, CJ (1997). Nutrado induktita de blokado de AMPA kaj kainataj riceviloj ene de la ventrala striatumo: studo pri mikrofinfuzio. Konduto. Brain Res. 89, 107–113. doi: 10.1016/S0166-4328(97)00054-5
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Kemp, JM, kaj Powell, TP (1971). La sinaptia organizo de la kaŭda kerno. Filozofio. Trans. R. Soc. Lond. B Biol. Sci. 262, 403-412. doi: 10.1098 / rstb.1971.0103
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Kenny, PJ (2011). Rekomendaj mekanismoj en obezeco: novaj komprenoj kaj estontaj direktoj. Neŭrono 69, 664-679. doi: 10.1016 / j.neuron.2011.02.016
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Kessler, RC, Berglund, PA, Chiu, WT, Deitz, AC, Hudson, JI, Shahly, V., et al. (2013) La prevalenco kaj korelacioj de binge manĝa malordo en la Monda Mensa Sano-Enketoj de Monda Organizo. Biol. Psikiatrio 73, 904-914. doi: 10.1016 / j.biopsych.2012.11.020
Klenowski, PM, Fogarty, MJ, Belmer, A., Noakes, PG, Bellingham, MC, kaj Bartlett, SE (2015). Struktura kaj funkcia karakterizado de dendritaj arboj kaj GABAergic-sinaptaj enigaĵoj sur interneŭronoj kaj ĉefaj ĉeloj en la rato basolateral amigdalo. J. Neurophysiol. 114, 942-957. doi: 10.1152 / jn.00824.2014
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Lenoir, M., Serre, F., Cantin, L., kaj Ahmed, SH (2007). Intensa dolĉeco superas kokainan rekompencon. PLOJ UN 2: e698. doi: 10.1371 / journal.pone.0000698
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Li, Y., Kolb, B., kaj Robinson, TE (2003). La situo de konstantaj anfetamin-induktitaj ŝanĝoj en la denseco de dendritaj spinoj sur mezaj spinecaj neŭronoj en la kerno accumbens kaj caudate-putamen. Neuropsychofarmacology 28, 1082-1085. doi: 10.1038 / sj.npp.1300115
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Lindvall, O., kaj Björklund, A. (1978). Anatomio de la dopaminergiaj neŭronaj sistemoj en la cerbo de rato. Adv. Biochem. Psikofarmakolo. 19, 1-23.
Lutter, M., kaj Nestler, EJ (2009). Hejmaj kaj hedonaj signaloj interagas en la regulado de konsumado de manĝaĵoj. J. Nutr. 139, 629-632. doi: 10.3945 / jn.108.097618
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Maldonado-Irizarry, CS, Swanson, CJ, kaj Kelley, AE (1995). La receptoroj de glutamato en la kerno akcentas nutradan konduton per la flanka hipotalamo. J. Neurosci. 15, 6779-6788.
Malik, VS, Popkin, BM, Bray, GA, Poste, JP, kaj Hu, FB (2010). Suker-dolĉigitaj trinkaĵoj, obezeco, tipo 2 diabeto mellitus, kaj kardiovaskula malsano. Trafiko 121, 1356 – 1364. doi: 10.1161 / CIRCULATIONAHA.109.876185
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Ng, SW, Slining, MM, kaj Popkin, BM (2012). Uzo de kaloriaj kaj nealoriaj dolĉigiloj en pakitaj manĝaĵoj de usonaj konsumantoj, 2005-2009. J. Acad. Nutr. Dieto. 112, 1828 – 1834 e1821 – e1826. doi: 10.1016 / j.jand.2012.07.009
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Nielsen, SJ, Siega-Riz, AM, kaj Popkin, BM (2002). Tendencoj en energia konsumado en Usono inter 1977 kaj 1996: similaj deĵoroj viditaj inter aĝaj grupoj. Obes. Res. 10, 370 – 378. doi: 10.1038 / oby.2002.51
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Paxinos, G., kaj Watson, C. (2007). La Cerbo de Rato en Stereotoksaj Koordinatoj. Amsterdamo; Boston, MA: Akademia Gazetaro / Elsevier.
Popkin, BM (2010). Kio misas kun la usona aliro al obezeco? Virtuala Mentoro 12, 316–320. doi: 10.1001/virtualmentor.2010.12.4.pfor2-1004
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Rada, P., Avena, NM, kaj Hoebel, BG (2005). Ĉiutage fuŝado de sukero ree liberigas dopaminon en la akciza ŝelo. Neurokienco 134, 737 – 744. doi: 10.1016 / j.neuroscience.2005.04.043
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Rafols, JA, Cheng, HW, kaj McNeill, TH (1989). Studo de Golgi pri la musa striato: dendritaj ŝanĝoj en aĝo de malsamaj neŭronaj populacioj. J. Comp. Neŭrolo. 279, 212 – 227. doi: 10.1002 / cne.902790205
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Ranjan, A., kaj Mallick, BN (2010). Modifita metodo por konsekvenca kaj fidinda Golgi-cox-makulado en signife reduktita tempo. Fronto. Neurol. 1: 157. doi: 10.3389 / fneur.2010.00157
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Reinehr, T. (2013). Tipo 2 diabeto mellitus en infanoj kaj adoleskantoj. Mondo J. Diabeto 4, 270 – 281. doi: 10.4239 / wjd.v4.i6.270
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Reissner, KJ, kaj Kalivas, PW (2010). Uzi glutamatan homeostazon kiel celon por kuracado de toksomanioj. Behav. Pharmacol. 21, 514–522. doi: 10.1097/FBP.0b013e32833d41b2
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Reynolds, SM, kaj Berridge, KC (2003). Glutamataj motivaj ensembloj en nukleaj akcentoj: rostrocaudaj ŝelaj gradientoj de timo kaj nutrado. Eŭro. J. Neurosci. 17, 2187-2200. doi: 10.1046 / j.1460-9568.2003.02642.x
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Rikardo, JM, kaj Berridge, KC (2011). Nucleus accumbens dopamina / glutamate-interagado ŝanĝas modojn por generi deziron kontraŭ timado: D (1) sole por apetita manĝo sed D (1) kaj D (2) kune pro timo. J. Neurosci. 31, 12866-12879. doi: 10.1523 / JNEUROSCI.1339-11.2011
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Robinson, TE, kaj Kolb, B. (1999). Transformoj en la morfologio de dendritoj kaj dendritaj spinoj en la kerno acumbens kaj prefrontal-kortekso post ripetita traktado kun amfetamino aŭ kokaino. Eŭro. J. Neurosci. 11, 1598-1604. doi: 10.1046 / j.1460-9568.1999.00576.x
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Robinson, TE, kaj Kolb, B. (2004). Struktura plastikeco asociita kun eksponiĝo al drogoj de misuzo. Neuropharmacology 47 (Suppl. 1), 33 – 46. doi: 10.1016 / j.neuropharm.2004.06.025
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Russo, SJ, Dietz, DM, Dumitriu, D., Morrison, JH, Malenka, RC, kaj Nestler, EJ (2010). La dependigita sinapso: mekanismoj de sinapta kaj struktura plasticeco en kerno accumbens. Tendencoj Neurosci. 33, 267 – 276. doi: 10.1016 / j.tins.2010.02.002
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Rutledge, LT, Duncan, J., kaj Beatty, N. (1969). Studo pri piramidaj ĉelaj aksonaj koleratoj en nerompita kaj parte izolita plenkreska cerba kortekso. Brain Res. 16, 15–22. doi: 10.1016/0006-8993(69)90082-1
Saper, CB, Chou, TC, kaj Elmquist, JK (2002). La bezono nutriĝi: homeostatika kaj hedonika kontrolo de manĝado. Neŭrono 36, 199–211. doi: 10.1016/S0896-6273(02)00969-8
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Scheggi, S., Secci, ME, Marchese, G., De Montis, MG, kaj Gambarana, C. (2013). Influo de palabileco sur instigo funkcii por kaloria kaj ne-kaloria manĝaĵo en ne-senhavaj kaj manĝeblaj ratoj. Neurokienco 236, 320 – 331. doi: 10.1016 / j.neuroscience.2013.01.027
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Sesack, SR, kaj Pickel, VM (1990). En la rato mezaj kernoj, hipokampaj kaj katecolaminergiaj finaĵoj konverĝas al spaj neŭronoj kaj konsentas unu kun la alia. Brain Res. 527, 266–279. doi: 10.1016/0006-8993(90)91146-8
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Sesack, SR, kaj Pickel, VM (1992). Prefrontalaj kortikaj efikoj en la rato sinapsis sur nebeligitaj neuronaj celoj de katecolaminaj finaĵoj en la kerno accumbens septi kaj sur dopamina neŭronoj en la ventra tegmentala areo. J. Comp. Neŭrolo. 320, 145 – 160. doi: 10.1002 / cne.903200202
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Ŝerifo, M., Quik, M., Holgate, JY, Morgan, M., Patkar, OL, Tam, V., et al. (en gazetaro). Neŭronaj nikotinaj acetilkolinaj modulatoroj reduktas sukeron. PLOJ UN.
Sheehan, DV, kaj Herman, BK (2015). La psikologiaj kaj medicinaj faktoroj asociitaj kun ne traktita binge manĝa malordo. Prim. Care Companion CNS Malordo. 17 doi: 10.4088 / PCC.14r01732
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Simms, JA, Steensland, P., Medina, B., Abernathy, KE, Chandler, LJ, Wise, R., et al. (2008) Intermita aliro al 20% etanolo induktas altan konsumon de etanolo en ratoj Long-Evans kaj Wistar. Alkoholo. Kliniko. Ekspliko Res. 32, 1816-1823. doi: 10.1111 / j.1530-0277.2008.00753.x
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Steensland, P., Simms, JA, Holgate, J., Richards, JK, kaj Bartlett, SE (2007). Varenicline, alfa4beta2 parta agonisto de niketina acetilcolina, selektive malpliigas kaj serĉas etanolon. Prok. Natl. Acad. Sci. Usono 104, 12518-12523. doi: 10.1073 / pnas.0705368104
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Stuber, GD, Hopf, FW, Hahn, J., Cho, SL, Guillory, A., kaj Bonci, A. (2008a). Volonta konsumado de etanolo plibonigas ekscitigan sinaptan forton en la ventrala tegmenta areo. Alkoholo. Kliniko. Ekspliko Res. 32, 1714-1720. doi: 10.1111 / j.1530-0277.2008.00749.x
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Stuber, GD, Klanker, M., de Ridder, B., Bowers, MS, Joosten, RN, Feenstra, MG, et al. (2008b). Rekomendaj prognozaj indicoj plibonigas ekscitigan sinaptan forton sur dubonaj dopaminaj neŭronoj. scienco 321, 1690-1692. doi: 10.1126 / science.1160873
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Swanson, SA, Korvo, SJ, Le Grange, D., Swendsen, J., kaj Merikangas, KR (2011). Antaŭvaloro kaj korelacioj de manĝaj malordoj en adoleskantoj. Rezultoj de la nacia komorbideca enketo pri adoleska suplemento. Arko. Genia psikiatrio 68, 714-723. doi: 10.1001 / archgenpsychiatry.2011.22
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Te Morenga, L., Mallard, S., kaj Mann, J. (2013). Dietaj sukeroj kaj korpa pezo: sistema revizio kaj metaanalizoj de randomigitaj kontrolitaj provoj kaj kohortaj studoj. BMJ 346: e7492. doi: 10.1136 / bmj.e7492
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Totterdell, S., kaj Smith, AD (1989). Konverĝo de hipokampo kaj dopaminergia enigo al identigitaj neŭronoj en la kerno de la rato. J. Chem. Neuroanato. 2, 285-298.
Tukey, DS, Ferreira, JM, Antoine, SO, D'Amour, JA, Ninan, I., Cabeza de Vaca, S., kaj aliaj. (2013). Sakarosa ingestaĵo kaŭzas rapidan trafikon de ricevilo AMPA. J. Neurosci. 33, 6123-6132. doi: 10.1523 / JNEUROSCI.4806-12.2013
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Veening, JG, Cornelissen, FM, kaj Lieven, PA (1980). La tópica organizado de la aferentoj al la kaŭdatoputamenoj de la rato. Studo sur peroksidaj peroksidoj. Neurokienco 5, 1253–1268. doi: 10.1016/0306-4522(80)90198-0
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Ventura, T., Santander, J., Torres, R., kaj Contreras, AM (2014). Neurobiologia bazo de avido de karbonhidratoj. nutrado 30, 252 – 256. doi: 10.1016 / j.nut.2013.06.010
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Volkow, ND, Wang, GJ, Fowler, JS, Tomasi, D., kaj Baler, R. (2012). Manĝaĵo kaj drogokompenso: interkovrantaj cirkvitoj en homa obezeco kaj toksomanio. Curr. Supro. Konduto Neurosci. 11, 1–24. doi: 10.1007/7854_2011_169
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Saĝa, RA (1973). Volontula konsumado de etanolo en ratoj post eksponiĝo al etanolo laŭ diversaj horaroj. Psikopharmacologia 29, 203-210. doi: 10.1007 / BF00414034
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Wright, CI, kaj Groenewegen, HJ (1995). Skemoj de konverĝo kaj apartigo en la mediana kerno de la rato: rilatoj de prefrontalaj kortikaj, mezliniaj talamoj kaj basalaj amigdaloidaj aferentoj. J. Comp. Neŭrolo. 361, 383 – 403. doi: 10.1002 / cne.903610304
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
Ŝlosilvortoj: binge-simila konsumo, longdaŭra, meza spina neŭro, nukleo accumbens, sukerozo
Citado: Klenowski PM, Shariff MR, Belmer A, Fogarty MJ, Mu EWH, Bellingham MC kaj Bartlett SE (2016) Prolonged Consumption of Sucarose in Binge-Like Manner, Alters the Morphology of Medium Spine Neurons in the Nucleus Accumbens Shell. Fronto. Konduto Neurosci. 10: 54. doi: 10.3389 / fnbeh.2016.00054
Ricevita: 03 decembro 2015; Akceptita: 07 March 2016;
Eldonita: 23 March 2016.
Eldonita de:
Nora Abrous, Institut des Neurosciences de Bordeaux, Francio
Reviziita de:
Serĝo H. Ahmed, Centre National de la Recherche Scientifique, Francio
Stéphanie Caille, Centre National de la Recherche Scientifique, Francio
Kopirajto © 2016 Klenowski, Shariff, Belmer, Fogarty, Mu, Bellingham kaj Bartlett. Ĉi tio estas malferma-alira artikolo distribuita sub la kondiĉoj de la Krea Komunaĵo Atribuka Permesilo (CC BY). La uzo, dissendo aŭ reproduktaĵo en aliaj forumoj estas permesita, se la originala aŭtoro aŭ licencanto estas akredititaj kaj ke la originala publikigado en ĉi tiu ĵurnalo estas citita, laŭ akceptebla akademia praktiko. Neniu uzo, dissendo aŭ reprodukto estas permesata, kiu ne plenumas ĉi tiujn terminojn.
* Korespondado: Selena E. Bartlett, [retpoŝte protektita]
