 Martin Zack
Martin Zack- 1Laboratorio de Kognitiva Psikofarmakologio, Fako pri Neŭroscienco, Centro por toksomanio kaj mensa sano, Toronto, ON, Kanado
- 2Translational Neuroscience Program, Sekcio de Psikiatrio, Lernejo de Medicino, Universitato de Pensilvanio, Filadelfio, Pensilvanio, Usono
- 3Sekcio pri Biopsikologio, Fako pri Neŭroscienco, Centro por toksomanio kaj mensa sano, Toronto, ON, Kanado
La toksomanio estas konsiderata kiel cerba malsano kaŭzita de kronika ekspozicio al drogoj. Sensivigo de cerbaj dopaminaj (DA) sistemoj parte mediadas ĉi tiun efikon. Patologia hazardludo (PG) estas konsiderata kiel kondutisma toksomanio. Tial PG povas esti kaŭzita de kronika ekspozicio al vetludado. Identigi hazardlud-sensivigitan DA-sistemojn subtenus ĉi tiun eblon. Rekompenco pri hazardludo elvokas liberigon de DA. Unu epizodo de fendeta maŝina movo ŝanĝas la respondon de DA de rekompenco al apero de sonoriloj (ŝpinitaj bobenoj) al rekompenco, konforme al tempaj diferencaj lernadaj principoj. Tiel, kondiĉitaj stimuloj (CS) ludas ŝlosilan rolon en DA-respondoj al vetludado. En primatoj, DA-respondo al CS estas plej forta kiam rekompenca probablo estas 50%. Sub ĉi tiu horaro, la CS eligas esperon pri rekompenco sed donas neniujn informojn pri tio, ĉu ĝi okazos en donita provo. Dum vetludado, 50%-horaro devus provoki maksimuman DA-liberigon. Ĉi tio proksime kongruas kun rekompenca ofteco (46%) sur komerca fuzmaŝino. DA liberigo povas kontribui al sentivigo, precipe por amfetamino. Kronika ekspozicio al CS kiu antaŭdiras rekompencon 50% de la tempo povus imiti ĉi tiun efikon. Ni testis ĉi tiun hipotezon en tri studoj ĉe ratoj. Bestoj ricevis 15 × 45-min-eksponojn al CS, kiu antaŭdiris rekompencon kun probablo de 0, 25, 50, 75, aŭ 100%. La CS estis lumo; la rekompenco estis 10% sukrosa solvo. Post trejnado, ratoj ricevis sentivan reĝimon de kvin apartaj dozo (1 mg / kg) de d-amfetamino. Laste ili ricevis 0.5 aŭ 1 mg / kg amfetaminan defion antaŭ testo de 90-min-lokomotora agado. En la tri studoj, la grupo 50% montris pli grandan agadon ol la aliaj grupoj en respondo al ambaŭ defiaj dozoj. Efikaj grandecoj estis modestaj sed konsekvencaj, kiel reflektita de signifa grupo × rango-asocio (ϕ = 0.986, p = 0.025). Kronika elmontro al vetlud-simila horaro de antaŭdestinaj stimuloj povas antaŭenigi sentivigon al amfetamino tre kiel ekspozicio al amfetamino mem.
Enkonduko
La toksomanio estis karakterizita kiel cerba malsano kaŭzita de kronika ekspozicio al drogoj de misuzo (Leshner, 1997). Neŭroplastikeco pensas, ke mediacias la efikojn de tia ekspozicio (Nestler, 2001). Sensibilizado de cerbaj dopaminaj (DA) sistemoj estas formo de neuroplasticeco implikita en hiper-reaktiveco al kondiĉitaj stimuloj (CS) por drogoj, kaj deviga drogo serĉanta (Robinson kaj Berridge, 2001). Sensivigo estis operacie difinita per pliigita DA-liberigo en respondo al CS por rekompenco kaj per pliigita lokomotora respondo al farmakologia DA-defio (Robinson kaj Berridge, 1993; Pierce kaj Kalivas, 1997; Vanderschuren kaj Kalivas, 2000). Kvankam sentivigo estas nur unu el multaj cerbaj ŝanĝoj ligitaj kun toksomanio (kp. Robbins kaj Everitt, 1999; Koob kaj Le Moal, 2008), oni sugestis ke ŝanĝoj en presinaptika dopamina liberigo reprezentu oftajn neŭroadaptojn implikitajn en toksomani-serĉado de drogoj (ekz. relanĉo), en tio, ke drogoj, kiuj induktas lokomotan sentivigon opii (ekz. morfino) aŭ stimulan defion (ekz. amfetamino) , ankaŭ kaŭzas reintegriĝon de estingitaj operantaj respondoj por mem-administrado de heroino aŭ kokaino - besta modelo de reaperado (Vanderschuren et al., 1999). Evidenteco, ke instiga sentivigo (pliigita valoro de la rekompenco de drogoj) estas plej prononcita post komenca eksponiĝo al toksomaniuloj plue sugestas, ke ankaŭ la sentivigo povas esti implikita en la fruaj stadioj de toksomanioVanderschuren kaj Pierce, 2010).
Patologia hazardludo (PG) estis priskribita kiel kondutisma toksomanio kaj lastatempe reklasita al la sama kategorio kiel malsanaj dependecaj malordoj en la 5a eldono de la Diagnoza kaj Statistika Manlibro de Mensa Malordoj (Frascella et al., 2010; APA, 2013). Ĉi tio implicas, ke PG povas esti kaŭzita de kronika ekspozicio al videoludado, ke oftaj mekanismoj povas mediacii la efikojn de hazardludo kaj drogopropono (Zack kaj Poulos, 2009; Leeman kaj Potenza, 2012); kaj ke sentivigado de cerbaj DA-vojoj povas esti grava elemento de ĉi tiu procezo.
Klinikaj evidentecoj nerekte subtenas ĉi tiun eblon: Uzante tomaton de emisioj de pozitronoj (PET) Boileau kaj kolegoj trovis, ke viraj PG-subjektoj montras pli grandan striitan DA-liberigon en respondo al amfetamino (0.4 mg / kg) ol sanaj viraj kontroloj (Boileau et al., 2013). Entute grupaj diferencoj estis signifaj en la asocia kaj somatosensoria striatumo. En la limia striato, kiu inkluzivas la kernon accumbens, la grupoj ne diferencis. Tamen, en PG-subjektoj, liberigo de DA en la limia striatumo korelaciita rekte kun la severeco de PG-simptomoj. Ĉi tiuj trovoj konformas al sentivigo de cerbaj DA-vojoj en PG, sed ankaŭ sugestas iujn gravajn diferencojn kun homaj substancaj individuoj kaj kun la klasika besta modelo de amfetamina sentivigo. Male al PG-subjektoj kaj bestoj eksponitaj al malaltaj dozoj de amfetamino (kp. Robinson et al., 1982), homoj kun dependeco de substanco konstante elmontras malpliigitan liberigon de DA al stimula defio (Volkow et al., 1997; Martinez et al., 2007), kaj evidentaĵoj de bestoj sugestas, ke ĉi tio eble reflektas deficitojn en DA-funkcio dum la komencaj stadioj de sindeteno sekvante bingajn padronojn de substanco misuzo (Mateo kaj aliaj, 2005). En studoj kie stimula sentivigo estas montrita ĉe bestoj, plibonigita DA-liberigo estas kutime observata en la limia striatum anstataŭ en la dorsal (asocia, somatosensory) striatum (Vezina, 2004). Tamen, kurac-serĉado de indico en bestoj ripete elmontritaj al kokaino estis ligita kun plibonigita DA-liberigo en la dorsia striatumo, rezulto pensita indiki pli kutiman formon de instigita konduto (Ito et al., 2002). Tiel, la totala altiĝo en liberigo de DA en dorsajn regionojn en PG-subjektoj povas esti rilata al rekompenco laŭ kutimaj (neflekseblaj, rutinigitaj) rekompencoj, kiuj celas impliki "progreson de ventralo al pli dorsaj domajnoj de la striato" (Everitt kaj Robbins, 2005p. 1481), dum la severeco-dependa DA-liberigo en limbia striato en ĉi tiuj subjektoj eble respondas pli proksime al stimula sentemo tiel tipe modelita ĉe bestoj. La PET-konkludoj ne povas riveli ĉu DA hiper reactividad estis antaŭekzista trajto de ĉi tiuj PG-subjektoj, konsekvenco de hazardludo-ekspozicio, aŭ rezulto de iu alia procezo tute. Por trakti ĉi tiun demandon, necesas pruvi indukton de sentivigado per kronika hazardludo-ekspozicio en subjektoj kiuj estas normalaj antaŭ eksponi. Ĉi tio starigas demandojn pri kiaj ecoj de vetludado plej probable provokas sensibiligon.
Skinner rimarkis, ke la varia horaro de plifortigo estis fundamenta por allogi la ludadon (aŭ almenaŭ ĝian persistadon) (Skinner, NENIU). Vetada konduto en fenda maŝina ludo konformas bone al la bazaj principoj de instrumenta kondiĉado, kiel reflektas per ebla korelacio inter mona elspezo kaj veda grandeco je sinsekvaj spinoj (Tremblay et al., 2011). Tiel, reaganta varia raporto ŝajnas provizi ekstere validan modelon de fendmaŝinludo.
Lastatempaj esploroj kun bestoj disponigas fortan komencan subtenon por kaŭza efiko de videoludado sur sentivigo. Kantisto kaj kolegoj ekzamenis la efikojn de 55 1-h ĉiutagaj kunsidoj de fiksa (FR20) aŭ ŝanĝiĝema (VR20) sakarina plifortigo en operanta levil-gazetara paradigmo sur posta lokomotora respondo al malalta dozo (0.5 mg / kg) amfetamino en sana viro ( Sprague Dawley) ratoj (Kantisto et al., 2012). Ili hipotezis, ke se hazardludo kondukas al sentivigado, ratoj eksponitaj al la ŝanĝiĝema horaro, kiuj imitas hazardludon, devus montri pli grandan respondon al anfetamino ol ratoj eksponitaj al la fiksa horaro. Kiel antaŭdiris, la VR20-grupo montris respondon de 50% pli granda lokomotora ol amfetamino ol la grupo FR20. Male, la grupoj montris ekvivalentan movadon post sala injekto. Ĉi tiuj rezultoj konfirmas, ke kronika ekspozicio al varia plifortigo sufiĉas por indukti hiperreativecon al DA-defio ĉe sanaj animaloj al la hazardo laŭ la respektivaj horaroj.
Multaj demandoj ekestas el ĉi tiu rezulto: Unue, ĝis kia grado la perceptita kontingento - aŭ manko de ĝi - inter la operanta respondo kaj ĝia rezulto mediacias ĉi tiujn efikojn? Laŭ lernado, ĉi tiu efiko implikas "respondon-rezultan atendon", aŭ eble simila efiko videblas en la foresto de opona respondo, t.e., "espero-stimulo-rezulto" en pavlova paradigmo (kp. Bolles, 1972)? Due, ĉu la grado de contingencia inter la okazaĵo antecedente (respondo aŭ stimulo) kaj lia rezulto influas la gradon de sensibilización?
La dua demando koncernas la rolon de necerteco pri sentivigado. Ekzemple, ĉu ludoj kies rezulto estas vere hazardaj - tute neantaŭvideblaj - havas pli grandan potencialon indukti sensibiligon ol ludoj kie la probabloj de venko estas klare difinitaj sed ne hazardaj, eĉ se la absoluta indico de rekompenco estas malalta? La nuna esplorado traktis ĉi tiujn demandojn.
La eksperimenta dezajno estis informita de seminala studo pri rekompenco de rekompenco kaj respondo de neŭronaj DA en simioj (Fiorillo et al., 2003). La bestoj en tiu studo ricevis sukon rekompencon (US) sub 0, 25, 50, 75, aŭ 100% ŝanĝiĝemaj raporto-horaroj. La horaroj estis nomumitaj de 1 de 4 malsamaj CS (ikonoj). La 0% -programo liveris rekompencon same ofte kiel la 100-% horaro, sed preterlasis la CS. Frapado de neŭronoj de DA dum la intervalo inter komenco de CS kaj liveraĵo aŭ preterlaso de Usono estis la esenca dependa mezuro. La studo trovis ke DA-respondo pligrandiĝis kiel funkcio de la necerteco de rekompenco. Tiel, sub la 100% -ta horaro la CS elvokis malmulte da agado, sub la 25 kaj 75% horaroj, la CS elvokis moderajn kaj similajn agadajn nivelojn, kaj sub la 50% -programo la CS elvokis maksimuman agadon. En ĉiu kazo, pafo rapideco eskaladis dum la kurso de la CS-Usona intervalo, te kiam la atendado alproksimiĝis.
Ĉi tiuj rezultoj indikas, ke agado de DA ne nur varias kun tio, ĉu aŭ ne rekompenco estas certa (Fiksa Raporto) aŭ necerta (Variablo), sed ankaŭ varias laŭ inversa proporcio al la kvanto da informoj pri rekompenso transdonita de la CS. En la 100% -kondiĉo, la CS elvokas la rekompencon kaj ankaŭ perfekte antaŭdiras lian liveron. En la kondiĉoj de 25 kaj 75%, la CS elvokas la esperon kaj antaŭdiras rekompensan liveron tri el kvar fojoj. En la 50% -kondiĉo, CS elvokas la atendon, sed ne disponigas informojn pri rekompencaj transdono pli ol hazardo. Surbaze de iliaj trovoj, Fiorillo et al. konkludis: "Ĉi tiu necerteco-induktita pliigo de dopamino povus kontribui al la rekompencaj ecoj de vetludado" (p. 1901).
La efikoj de 50% varia rekompenco en ununura sesio ne devas ŝanĝiĝi dum pluraj kunsidoj, ĉar la probablo de rekompenco restas tute neantaŭvidebla en ĉiu provo. Tiel, konsiderante la kondiĉojn, kiuj maksimumigus kronikan aktivigon de DA-neŭronoj dum ripetaj epizodoj de ludado, la 50% -horaro devas generi la plej daŭran kaj la plej fortikan efikon. Ĉi tio estas rimarkinda, ĉar la longdaŭra rekompenco (rekompenco> 0) observita dum miloj da spinoj sur komerca fendo-maŝino estis 45.8% (Tremblay et al., 2011). Tiel, 50% -la rekompenco ŝajnas precize reflekti la rekompensan horaron administritan de faktaj hazardludaj aparatoj.
La nuna studo uzis la samajn kondiĉojn kiel Fiorillo et al. en kronika malkovro, inter-grupaj projektado kun ratoj. Bestoj spertis ~ 3 semajnojn da ĉiutagaj kondiĉoj, kie CS (malpeza) estis parigita kun Usono (malgranda kvanto de sakarozo). Post la trejnada fazo, bestoj ripozis antaŭ takso de sentiveco indeksita de lokomotora respondo al amfetamino. Surbaze de la literaturo, oni antaŭdiris, ke ratoj elmetitaj al malsamaj rekompencaj horaroj ne diferencus laŭ sia sendrata lokomotora konduto, sed montrus signife malsamajn nivelojn de movado post amfetamino, kun la 50% -a grupo montrante pli grandan lokomotivan respondon al la drogo rilate al la aliaj grupoj laŭlonge de dozoj, ŝablono, kiun oni atendus, se la 50% da bestoj estus antaŭe elmetitaj al aldonaj dozoj de amfetamino mem (t.e. krucosentema).
Eksperimento 1
Materialoj kaj metodoj
temoj
Kvar grupoj (n = 8 / grupo) de plenkreskaj (300-350 g) masklaj ratoj Sprague-Dawley (Charles River, St. Constant, Kebekio, Kanado) estis loĝataj individue en klaraj policarbonataj skatoloj (20 × 43 × 22 cm) sub reversa 12: 12 malpeza-malhela ciklo. Ili ricevis ad libitum aliro al manĝo kaj akvo, kaj ĉiutaga uzado de eksperimentisto dum 2 semajnoj antaŭ la studo. Ĉiu grupo estis kondiĉita sub unu el kvar ŝanĝiĝemaj rekompencaj horaroj: 0, 25, 50, aŭ 100%. La grupo 75% estis preterlasita en ĉi tiu komenca studo, kiel Fiorillo et al. (2003) trovis ekvivalentan post-CS DA liberigon sub 25 kaj 75% rekompencaj horaroj, tiel ke ambaŭ kondiĉoj kondukis al pli granda DA-liberigo ol faris la 100% CS-US-kondiĉon, sed malpli ol la 50% -kondiĉo.
aparato
Aliroj al sakarozaj prezentoj kaj al la CS estis provizitaj unuope en operantkondiĉoj (33 × 31 × 29 cm). Ĉiu skatolo estis ekipita per refortista revuo, lokita sur la antaŭa muro. Lumo en la supro de la revuo servis kiel CS. Motorita, solenoid-kontrolita likva ĉapo povus esti levita al la planko de la revuo. Eventoj en la skatolo estis kontrolitaj de med-komputilaj ekipaĵoj kaj programoj, uzante propran programon skribitan en MED-PC. La lokomotora testado estis farita individue en plexiglaj kaĝoj (27 × 48 × 20 cm). Ĉiu kaĝo estis provizita per monitora sistemo konsistanta el ses fototransaj ĉeloj por detekti horizontalan movadon.
proceduro
trejnado. La studo estis farita konforme al la etikaj gvidlinioj difinitaj de la kanada Konsilio pri Bestan Zorgadon. Ratoj estis manĝ-limigitaj al 90% de ilia korpa pezo por la daŭro de la studo kaj loĝigitaj individue. Ĉiu rat ricevis 15 tagojn da sakarosa rekompenco trejnado (10% -a solvo ĉe 0.06 ml por rekompenco): 5 sinsekvajn tagojn × 3 semajnojn, kun semajnfinaj for. Bestoj estis konservitaj sur norma ŝnuro antaŭ kaj post la trejna fazo; la malkovro de sakarozo estis limigita al la 15 ~ 40-min-trejnaj sesioj. Ĉiu ĉiutaga kunsido konsistis el prezentoj de stimuloj 15 (malpeza; CS), ĉiu apartigita per intertempa intervalo de 120-a. La lumo troviĝis en la supra panelo de la revuo, kaj restis por 25-oj, kun sakarozo havebla dum la lasta 5-oj. En la kazo de grupo 0, la ciprasa sakarozo levis ĉiun 140 (por 5-oj) sed la stimula lumo ne lumis. Ĉi tio egalis la intervalon inter prezentoj de la ĉapo en grupo 0 kaj la aliaj grupoj (120 + 25 s). Ĉiu kuracista sesio daŭris ~ minimume. Mezume, grupo 40 ricevis sakarozon unufoje por ĉiu kvar CS-prezentoj; grupo 25 ricevis sakarozon unufoje por ĉiu du CS-prezentoj, kaj grupo 50 ricevis sakarozon post ĉiu CS-prezento.
provoj. Du semajnojn post la lasta sesio pri sakarozo (aŭ "kondiĉado"), la lokomotora respondo al d-amfetamino (AMPH; ip) estis taksita. Ratoj ricevis tri 2-h-sesiojn por alkutimiĝi al la lokomotivaj kestoj, sekvitaj de ses AMPH-testo-sesioj. AMPH-testotagoj okazis je intervaloj 1-wk. En testotagoj, ratoj ricevis 30-min por alkutimiĝi al kestoj kaj tiam ricevis ununuran 0.5 mg / kg-dozon de AMPH sekvita, dum apartaj ĉiusemajnaj sesioj, per kvin 1.0 mg / kg-dozo (unu dozon ĉiutage) en testaj tagoj 1 tra 5 . Post-AMPH-movado estis taksita por 90-min en ĉiu sesio.
Datuma analiza aliro
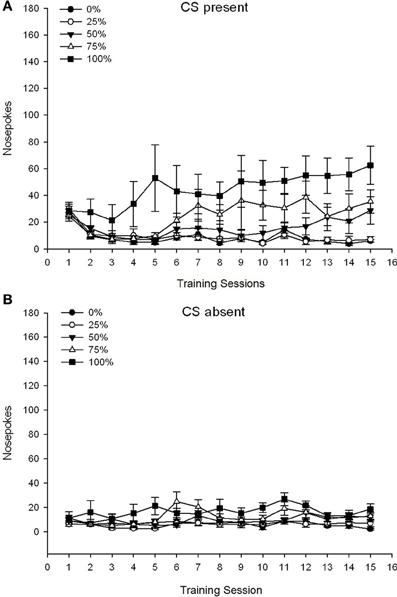
Statistikaj analizoj estis faritaj kun SPSS (v. 16 kaj v. 21; SPSS Inc., Chicago IL). Tuja kondutema respondo al la CS estis taksita laŭ la nazotruoj en la aperturon kie la sakarozo estis disdonita. La meznombro de nazotruoj dum ĉi tiu intervalo (5 s por provo) tiam estis komparita kun la meza nombro de nazotruoj por la sama daŭro (5-o) averaĝe dum la tempo kiam la CS ne ĉeestis. Grupo × Sesio ANOVA-oj kun N-ĉeestantaj ĉeestantoj kaj forestaj spuris la akiron de diskriminacia respondo al la indikoj kaj sendiskriptaj nazotrabaj respondoj sub la diversaj horaroj dum la kursaj trejnadaj sesioj 15.
Efikoj de kuracado pri lokomotoraj respondoj estis taksitaj per Grupo × Sesio-ANOVA-oj por la drog-liberaj fazoj (tri sesioj), antaŭ-sentiviga 0.5 mg / kg AMPH-defio (unu kunsido), kaj dum la kvin-sesio 1 mg / kg AMPH reĝimo pri sentivigado, kiam oni atendis, ke grupoj diferencas responde al ripetaj dozoj de AMPH. Grupo × Sesio-ANOVA-oj ankaŭ taksis respondojn lokomotorojn sen drogoj dum la antaŭ-injektado de 30-min-fazo de ĉiu AMPH-testo-sesio. Pluraj planitaj komparoj taksis la diferencon en meznombraj elfaroj de grupo 50 kontraŭ grupo 0 (sen atendokontrolo) kaj grupo 100 (neniu necertecan kontrolon), per t-testoj (Howell, 1992), uzante la terminojn de MS-eraro kaj df-eraro por la koncernanta efiko (t.e., grupo aŭ grupo × sesia interagado) de la ANOVA (Winer, 1971). Polinomaj tendencoj analizas la profilon de ŝanĝoj dum sesioj.
Por determini, ĉu aliraj reagoj en ĉeesto kaj foresto de la CS dum la trejnadaj sesioj de 15 kontribuis al variado en lokomotora respondo al AMPH, aŭ mediataj grupaj diferencoj en respondo al AMPH, oni daŭrigis analizon de kunvarianco (ANCOVA) ĉe AMPH lokomotoraj datumoj, inkluzive de totalaj nazoj (sumo por 15-sesioj) kiam la CS estis forestanta kiel la plegaria. Grava efiko de la kunvarianto indikus, ke senrespara aliro al reagoj moderigis (influis la forton de) la efikojn de grupo aŭ sesio. Malpliiĝo en la signifo de la efikoj de grupo aŭ sesio en ĉeesto de signifa kovariaro indikus ke aliraj respondoj mediaciis (respondecis) la efikojn de grupo aŭ sesio. Malpliiĝo en la signifo de grupaj aŭ seancaj efikoj se manko de signifa kunvarianta efiko simple reflektus perdon de statistika potenco pro la reasignado de df de la termino de eraro al la kunvario, kaj ne havus influon sur la interpretado de la efikoj de grupo aŭ sesio.
rezultoj
Nazo poke dum sukerokondiĉoj
CS ĉeestanta. Figuro 1A montras la mezajn nazajn okulvitrojn por grupoj 25, 50 kaj 100 dum la CS ĉeestis sur la 15 sakarozaj kondicxaj sesioj (nazoj ne koditaj por grupo 0, kiu ne ricevis CS). 3 Grupo × 15-Sesio ANOVA donis signifajn ĉefajn efikojn de Grupo, F(2, 21) = 5.63, p = 0.011, kaj Sesio, F(14, 294) = 14.00, p <0.001, kune kun signifa interaga grupo × kunsido, F(28, 294) = 2.93, p <0.001. Figuro 1A indikas, ke la ĉefa efiko de Sesio reflektis pliiĝon de nazoj tra sesioj en ĉiuj tri grupoj, kaj la ĉefa efiko de Grupo reflektis ĝenerale pli altajn ĝeneralajn poentarojn en grupo 100 kontraŭ grupo 25 kun mezaj partituroj en grupo 50. Grava Grupo × Sesio interago por la kuba tendenco, F(2, 21) = 4.42, p = 0.030, indikis rapidan pliiĝon, subenigon kaj ebenigon en nazo super sesioj en grupo 100, kontraŭ lineara pliiĝo super sesioj en grupo 50, kaj pli malprofunda lineara pliiĝo super sesioj en grupo 25.
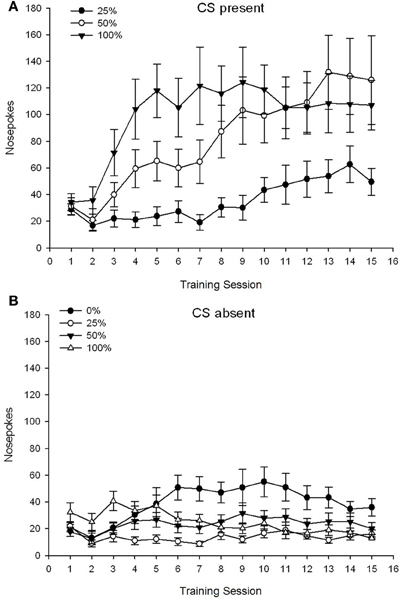
Figuro 1. Mezaj (SE) proksimaj respondoj (nazaj pokoj) sur trejnaj sesioj de 15 sukroza en grupoj de ratoj Sprague Dawley (n = 8 / grupo) eksponita al sakarosa rekompenco (10% solvo) liverata sub 0, 25, 50, aŭ 100%-ŝanĝiĝemaj horaroj. La kondiĉita stimulo estis lumo (120 s). Grupo 0 ricevis la saman nombron da rekompencoj kiel la grupo 100 en manko de kondiĉitaj stimuloj. (A) Poentaroj kiam CS ĉeestis (5 s × 15-provoj). (B) Poentaroj kiam CS forestis (mezumo por 5 × 15 s dum lumo estis malŝaltita).
CS forestas. Figuro 1B montras la mezan nazon poke por ĉiuj kvar grupoj por ekvivalenta daŭro (5 s × 15-provoj) averaĝis dum la tempo kiam la CS estis forestanta. 4 Grupo × 15-Sesio ANOVA donis signifajn ĉefajn efikojn de Grupo, F(3, 28) = 7.06, p = 0.001, kaj Sesio F(14, 392) = 2.84, p <0.001, kune kun signifa interaga grupo × kunsido, F(42, 392) = 3.93, p <0.001. Signifa interagado de Grupo × Sesio por la kvadrata tendenco, F(3, 28) = 3.91, p = 0.019, kune kun neniu interago por la kuba tendenco, F(3, 28) <0.93, p > 0.44, reflektis profilon "renversita-U" de nazpunktoj super kunsidoj en grupo 0, kontraŭ ĝenerale stabila profilo dum kunsidoj en la aliaj grupoj.
Kutimaro al lokomotoraj ĉambroj
4 Grupo × 3-Sesio ANOVA donis ĉefan efikon de Sesio, F(2, 56) = 5.67, p = 0.006, kaj neniuj aliaj signifaj efikoj, F(3, 28) <1.60, p > 0.21. Meznivelaj (SE) radiaj paŭzoj po 2 h en la lokomotoraj skatoloj estis 1681 (123) en la kunsido 1, 1525 (140) en la kunsido 2, kaj 1269 (96) en la kunsido 3. Planitaj komparoj ne trovis signifajn diferencojn inter grupo 50 kaj grupo 0 aŭ grupo 100 dum la unua aŭ fina kutimiga kunsido, t(84) <1.69, p > 0.05. Tiel, en la foresto de AMPH, ripeta ekspozicio al la provaj skatoloj estis asociita kun konsekvenca malpliiĝo de spontanea lokomotora agado en la kvar grupoj (te Sesio-efiko), kaj neniu diferenca respondo kiel funkcio de sakarosa trejnhoraro (neniu interago) .
Test-kunsidoj
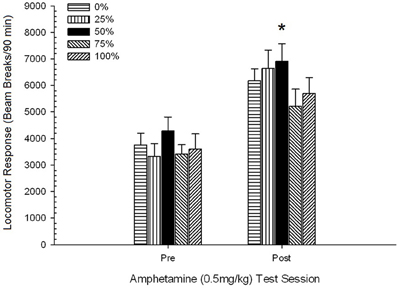
Efikoj de antaŭ-sentivigo 0.5 mg / kg AMPH-defio.
Antaŭ-injekta movado. Neŭtra maniero-ANOVA de 4-grupo de lokomotora respondo dum la antaŭ-injektado-fazo de 30-min ne donis signifajn efikojn, F(3, 28) <1.05, p > 0.38. Planitaj komparoj trovis neniun signifan diferencon inter grupo 50 kaj grupo 0 aŭ grupo 100, t(32) <0.87, p > 0.40. Sekve, bazaj diferencoj en antaŭinjekta movado ne kalkulis grupajn diferencojn en lokomotora respondo al AMPH. Duonaj (SE) trabo-paŭzoj por la specimeno estis 559 (77).
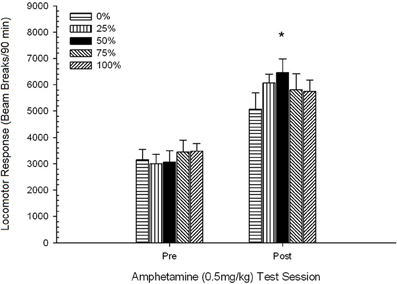
Post-injekta movado kontraŭ finaj senokupaj kutimoj. 4-Grupo × 2-Sesio ANOVA komparis la lokomotorajn respondojn de la grupoj dum la fina kutima kunsido, kaj tuj post la antaŭ-sentiviga 0.5 mg / kg AMPH-defio. Poentaroj por la kutima kunsido (120 min) estis skalitaj por korespondi kun la daŭro de la AMPH-testo (90 min) (kruda kutima poentaro × 90/120). La analizo donis signifan ĉefan efikon de Sesio, F(1, 28) = 34.16, p <0.001, kaj neniuj aliaj signifaj efikoj, F(3, 28) <2.26, p > 0.10. La Sesio-efiko reflektis pliigon de mezaj (SE) radiaj paŭzoj kiel respondo al la dozo, de 952 (72) ĝis 1859 (151). Planitaj komparoj trovis neniujn signifajn diferencojn inter grupo 50 kaj grupo 0 aŭ grupo 100 responde al la dozo, t(56) <1.72, p > 0.10. Tamen la rango-ordo de poentoj de trabo (M; SE) akordigita kun la hipotezo: grupo 50 (2205; 264)> grupo 0 (2025; 203)> grupo 100 (1909; 407)> grupo 25 (1296; 299) .
Efikoj de 1 mg / kg AMPH.
Antaŭ-injekta movado. 4 Grupo × 5-Sesio ANOVA de lokomotora respondo dum la antaŭ-injektado de 30-min-fazo pri 1 mg / kg AMPH-sesioj donis ĉefan efikon de Sesio, F(4, 112) = 43.64, p <0.0001, kaj neniuj aliaj signifaj efikoj, F(3, 28) <0.97, p > 0.42. Planitaj komparoj trovis neniun signifan diferencon inter grupo 50 kaj grupo 0 aŭ grupo 100 dum la unua aŭ fina testsesio, t(140) <0.84, p > 0.30. Sekve, bazaj diferencoj en movado ne kalkulis grupajn diferencojn en lokomotora respondo al AMPH. Meznivelaj (SE) trakaj interpunkciaj poentaroj por la antaŭ-doza kutima fazo en kunsidoj 1-5 estis: 454 (30), 809 (53), 760 (36), 505 (35), 756 (39).
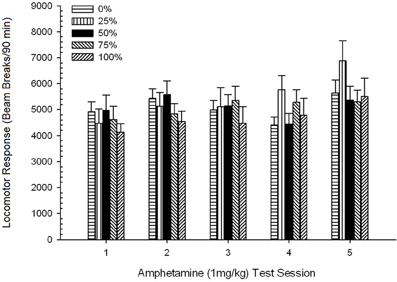
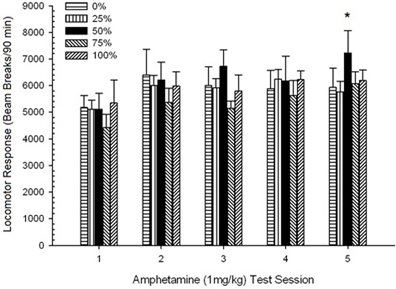
Post-injekta movado. Figuro 2 montras la efikojn de kvin injektoj de 1 mg / kg AMPH (unu por semajno) sur lokomotoraj agadopunktoj en la kvar grupoj. 4 Grupo × 5-Sesio ANOVA donis ĉefan efikon de Sesio, F(4, 112) = 8.21, p <0.001, marĝena ĉefa efiko de grupo, F(2, 45) = 3.28, p = 0.085, kaj neniu signifa interago, F(12, 122) <0.77, p > 0.68.
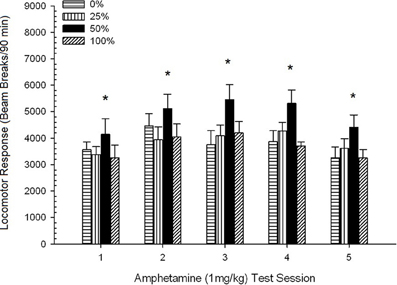
Figuro 2. Meznivela (SE) lokomotora respondo (nombro de paŭzoj de trabo en elektronika tabelo po 90 min) al 1 mg / kg d-amfetamino (ip) sur semajnaj sesioj de 5 en grupoj de ratoj Sprague Dawley (n = 8 / grupo) antaŭe eksponitaj al ĉiutagaj kunsidoj de 15 kun sakrosa rekompenco (10% solvo) transdonitaj sub 0, 25, 50, aŭ 100% ŝanĝiĝemaj horaroj. La kondiĉita stimulo estis lumo (120 s). Grupo 0 ricevis la saman nombron da rekompencoj kiel la grupo 100 en manko de kondiĉitaj stimuloj. *p <0.05 por averaĝa diferenco inter grupo 50 kaj grupo 0 same kiel grupo 100, surbaze de planitaj komparoj.
Planitaj komparoj rivelis, ke grupo 50-poentaroj diferencis signife de grupo 0, t(14) = 2.19, p = 0.037, kaj grupo 100, t(14) = 2.36, p = 0.025 [kaj diferencis marĝene de grupo 25, t(14) = 2.03, p = 0.051]. Tiel, en grupo 50, lokomotora respondo al 1 mg / kg AMPH fidinde superis tiun de la aliaj tri grupoj tra ĉiuj kvin testesesioj. Polinoma tendenco analizo detektis signifan kvadratan tendencon trans seancoj, F(1, 28) = 32.47, p <0.0001, kaj neniuj aliaj signifaj tendencoj, F(1, 28) <1.78, p > 0.19. Figuro 2 montras, ke ĉi tiu rezulto reflektis ŝablonon "inversitan U" tra sesioj.
Kontrolo por variado en nazo-poke respondanta dum trejnado de sakarozo
La sekvado ANCOVA de lokomotoraj respondoj al 1 mg / kg AMPH, kun nazotukoj (CS ĉeestantaj) kiel la plegaria, en la tri grupoj kiuj ricevis la CS, donis marĝenan ĉefan efikon de Grupo, F(2, 20) = 3.07, p = 0.069, kaj neniuj signifaj kunvariantaj rilataj efikoj, F(4, 80) <0.05, p > 0.85. Tiel, responda aliro respondanta dum trejnado ne klarigis signifan variadon en la lokomotora respondo al 1 mg / kg AMPH en grupoj 25, 50 aŭ 100.
La sekvado ANCOVA de lokomotoraj respondoj al 1 mg / kg AMPH, kun nazo-pokoj (CS-foresto) kiel kovariato, donis signifan efikon de la kovariato, F(1, 27) = 6.17, p = 0.020, signifa ĉefa efiko de Grupo, F(3, 27) = 4.13, p = 0.016, marĝena Sesio × Covariate interago, p = 0.080, kaj neniuj aliaj signifaj efikoj, F(4, 108) <1.48, p > 0.21. Tiel, nedifinita (sendistinga) aliro respondanta dum trejnado klarigis signifan variadon en lokomotora respondo al 1 mg / kg AMPH. Tamen ĉi tiu variado ne koincidis kun grupa rilata varianco, ĉar inkludo de la kunvariaĵo en la analizo pliigis anstataŭ malpliigi la signifon de la grupa efiko.
diskuto
La nazo poke datenoj dum la CS ĉeestis montras, ke grupoj akiris la asocion inter CS kaj sakarozo transdono kiel reflektita per pliigo en cued respondoj super trejnado kunsidoj. La profilo de respondado super sesioj dum la CS estis ĉeestanta sugestis ke 100 kaj 50% CS-usonaj horaroj estis same efikaj por akiri alproksimiĝon, dum la 25-%-horaro elvokis pli modestan pliiĝon en enfokusigita enfokusigas. La nazo poke datenoj dum la CS estis forestas sugestas ke grupoj kiuj ricevis iun ajn el la tri CS-sakarosa trejnado horaroj (grupo 25, 50, 100) rapide lernis redukti sian nazon pokes en foresto de la CS, dum bestoj en grupo 0 , kiu ricevis neniun CS, nur lernis malpliigi sian aliran konduton en limigita grado post vasta trejnado.
La kutimaj datumoj montras ke la grupoj ne diferencis antaŭ AMPH kaj ke ripetata ekspozicio al la testkestroj estis asociita kun malpliigita respondo de lokomotora sen drogo. Tial, inter-grupaj diferencoj kaj pliigita respondado super ripetaj dozoj de AMPH ne povas esti atribuita al antaŭekzistantaj diferencoj en lokomotora konduto.
Rezultoj de la antaŭ-sentiviga defio kun 0.5 mg / kg AMPH konfirmis, ke la medikamento pliigis lokomotoran agadon rilate al la fina nula kutimo tago. Laŭ la hipotezo, grupo 50 rangis pli alte ol grupoj 0 aŭ 100 (same kiel grupo 25) laŭ meznombraj respondoj al la dozo, kvankam la meznombraj diferencoj inter grupoj ne estis signifaj.
Por la sentemaj kunsidoj, la planitaj komparoj de inter-grupoj montris, ke antaŭa ekspozicio al 50% kondiĉita sakarozo rekompencon kaŭzis signifan kreskon de lokomotora respondo al 1.0 mg / kg dozo de amfetamino rilate al la aliaj tri horaroj. Ĉi tiu efiko estis evidenta de la unua dozo kaj ne ŝanĝis rimarkinde dum ripetaj dozoj. La tendenca analizo indikis dufazan respondon (por la plena specimeno) al ripetaj dozoj de AMPH, pliigante ĝis la tria dozo kaj malpliiĝante poste. La rezultoj de la sekvaĵo ANCOVA kun nazpunktoj (CS forestas) kiel la kunvariado konfirmis, ke diferencoj en la lokomotoraj respondoj de la kvar grupoj al 1 mg / kg AMPH ne estis peritaj de sen-responda aliro respondanta dum la sakarozaj trejnaj kunsidoj.
La grupa efiko dum la sentivigaj sesioj konformas al nia hipotezo. La dufaza sesiesta efiko ne kongruas kun la atendata kontinua eskalado de lokomotoraj respondoj kun ripetaj AMPH-dozoj. Ĉi tio povas esti rilatita al la doz-intervalo. Por trakti ĉi tiun problemon, procedo (alternaj ĉiutagaj dozoj) montrita por indukti konsekvencan grimpadon de lokomotora respondo al 1.0 mg / kg dozoj de AMPH (te konduta sentemo) devus esti uzata. La efiko de sentiviga reĝimo de AMPH en posta respondo al dua defio de 0.5 mg / kg plue subtenus la ĝeneralecon de ĉi tiu efiko. Inkludo de sala problemo antaŭ AMPH determinus la rolon de atendado aŭ injektado-rilata (ekz. Streso) efikojn sur la lokomotora respondo al AMPH. Inkluzivado de 75% kunkondiĉita sakarosa grupo helpus klarigi la rolon de rekompencaj necertecoj kontraŭ rekompenceco en la mastro de respondoj por grupoj 50 kaj grupo 25. Krome, por permesi pritaksadon (de ANCOVA) pri la kontribuo de respondo al statoj de lokomotivo sen AMI (uzante nazojn kun CS ĉeestanta kiel la plegaria), la nazo ŝvebas ankaŭ por grupo 0 dum la intervalo kiam la CS. ĉeestis en la aliaj kvar grupoj (te, tiel ke nazo poke de ĉiuj kvin grupoj - inkluzive de grupo 0 kiu ne ricevis CS - povus esti inkluzivita en la analizo de kunvarianco kun CS ĉeestanta kiel la plegaria). Ĉi tiuj rafinaĵoj estis enmetitaj en eksperimento 2.
Eksperimento 2
Materialoj kaj metodoj
La metodiko de eksperimento 2 estis simila al tiu de eksperimento 1 sed reviziita por pli bone aproksimi reĝimon trovitan fidinde indukti sensibilizadon de AMPH (Fletcher et al., 2005). Ŝanĝoj estis jenaj: (a) La 75% CS-sakarosa grupo (n = 8) estis inkluzivita; (b) Dum sukrosa trejnado, ratoj (krom por grupo 0) ricevis 20 CS (malpezajn) prezentojn (kontraste al 15 en eksperimento 1); (c) CS-prezentoj estis ĉiu apartaj per meza intertempa intervalo de 90-oj; intervalo: 30-180 s (kontraŭ 120-oj en eksperimento 1), kiu kompensas la pliiĝon en trejnadaj provoj por egaligi la daŭron de ĉiu trejnadsesio al tiu de eksperimento 1; (d) la daŭro de ĉiu el la tri kutimaj kunsidoj malpliiĝis de 120 al 90 min por korespondi kun la daŭro de la testaj sesioj; (e) salema (ip, 1 ml / kg) defio (90 min) estis aldonita (post-sakarosa trejna tago 8), por taksi la lokomotivajn efikojn de injekto en si mem (ekz. atendado, streĉo); (f) La 1-mg / kg-sentivigaj sesioj estis okazigitaj dum alternaj labortagoj (post-trejnadaj tagoj 12-21) prefere ol ĉiusemajne kiel en eksperimento 1; (g) Kune kun la antaŭ-sentiviga 0.5 mg / kg AMPH-defio (post-trejna tago 9) dua post-sentiviga 0.5 mg / kg AMPH-defio estis aldonita (post-sakarosa trejna tago 28), por testi la ĝeneralecon de la sentiviga efiko tra dozoj; (h) nazo poke dum CS estis ĉeestanta estis koditaj por ĉiuj grupoj (inkluzive de grupo 0); (i) nazo poke dum CS estis forestita estis registrita specife de la 5-s intervalo tuj antaŭ la komenco de la CS al indekso antaŭtempa alproksimiĝo respondas.
rezultoj
Nazo poke dum sukerokondiĉoj
5 Grupo × 15-Sesio × 2-Fazo (CS-ĉeestanta, CS forestanta) ANOVA de nazotruoj donis signifajn ĉefajn efikojn de Grupo, F(4, 19) = 2.89, p = 0.050, Sesio F(14, 266) = 2.28, p = 0.006, kaj Fazo, F(1, 19) = 14.72, p = 0.001, kaj ankaŭ signifa tri-vizaĝa interago, F(56, 266) = 1.38, p = 0.050. Paneloj (A, B) de Figuro 3 mapu la meznombrajn poentojn de la grupoj por la ĉeestantaj CS kaj forestantaj CS respektive. Komparo de la du paneloj malkaŝas, ke la ĉefa efiko de Fazo reflektis pli ĝeneralajn nazajn respondojn kiam la CS ĉeestis kontraŭ forestanta. Sekve, respondaj respondoj okazis signife pli ofte ol antaŭtempaj ne-responditaj respondoj. La ĉefaj efikoj de Grupo kaj Sesio ne estis facile interpretitaj pro la pli alta interago. Ĉi tiu lasta rezulto reflektis konverĝon de poentaroj por la kvin grupoj ĉe relative stabila malalta nivelo tra kunsidoj kiam la CS forestis (Figuro 3B), kune kun diverĝenco de partituroj en altajn (nivelojn 75, grupo 100), meznivelajn (grupojn 50), kaj malaltajn (grupojn 0, grupojn 25) de nazotukoj respondantaj super sesioj kiam la CS ĉeestis (Figuro) 3A). De la pli malalta orda polinoma tendencoj (lineara, kvadrata, kuba) nur la interrilata maniero por la lineara tendenco alproksimiĝis al signifo, F(4, 19) = 2.32, p = 0.094, reflektanta la ĝenerale monotonan pliiĝon de nazoj super sesioj en grupo 75 kaj relative pli rapidan stabiligon ĉe altaj, mezaj kaj malaltaj niveloj de respondo en la aliaj grupoj kiam la CS ĉeestis.
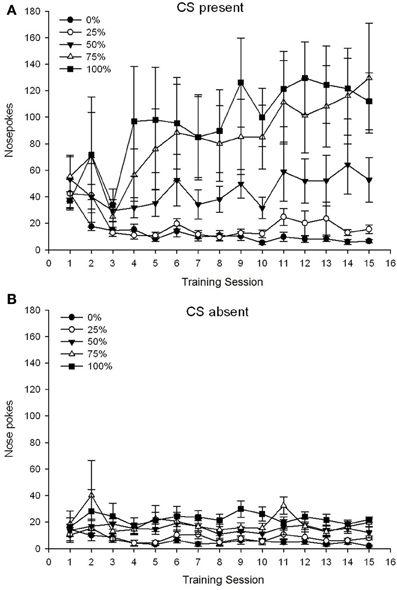
Figuro 3. Mezaj (SE) proksimaj respondoj (nazaj pokoj) sur trejnaj sesioj de 15 sukroza en grupoj de ratoj Sprague Dawley (n = 8 / grupo) eksponita al sakarosa rekompenco (10% solvo) liverita sub 0, 25, 50, 75, aŭ 100%-ŝanĝiĝemaj horaroj. La kondiĉita stimulo estis lumo (120 s). Grupo 0 ricevis la saman nombron da rekompencoj kiel la grupo 100 en manko de kondiĉitaj stimuloj. (A) Poentaroj kiam CS ĉeestis (5 s × 20-provoj). (B) Poentaroj kiam CS forestis (mezumo por 5 × 20 s dum lumo estis malŝaltita).
Kutimaro al lokomotoraj skatoloj
5 Grupo × 3-Sesio ANOVA de neakundaj lokomotoraj respondoj donis signifan ĉefan efikon de Sesio, F(2, 70) = 60.01, p <0.0001, kaj neniuj aliaj signifaj efikoj, F(4, 35) <0.70, p > 0.60. Planitaj komparoj de grupo 50 kun grupo 0 kaj kun grupo 100 en la unua kaj fina sesoj pri kutimiĝo donis neniujn gravajn efikojn, t<0.84, p > 0.40. Sekve, averaĝa sendoma lokomotora respondo en la ŝlosilaj grupoj ne diferencis antaŭ testado. Averaĝa (SE) nombro da radiaj paŭzoj po 90 min estis 2162 (118) en la unua sesio, 1 (1470) en la 116a sesio, kaj 2 (1250) en la 98a sesio.
Test-kunsidoj
Salaj. 5 Grupo × 2-Sesio ANOVA komparis respondon lokomotora sur la fina kutima sesio kaj salaj defiaj sesio. La ANOVA donis ĉefan efikon de Sesio, F(1, 35) = 62.46, p <0.0001, kaj neniuj aliaj signifaj efikoj, F(4, 35) <0.65, p > 0.64. Figuro 4 intrigoj la grupaj rimedoj kaj montras, ke la Sesia efiko reflektis ĝeneralan malkreskon de lokomotora respondo de la fina senkonsuma kutima sesio al la sala salono, kiu ne variis laŭ grupo. Tiel, la malpliiĝo de lokomotora respondo vidita dum la tri kutimaj sesioj daŭrigis la kvaran liberan ekspozicion al la testoskatoloj.
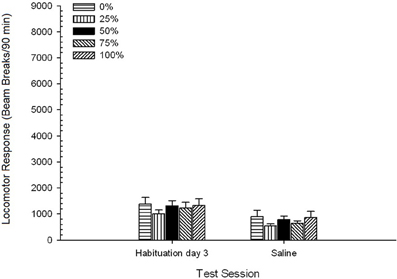
Figuro 4. Meza (SE) lokomotora respondo (nombro de trabo-paŭzoj en elektronika tabelo po 90 min) pri la lastaj el 3-senpagaj drog-sesioj kaj post posta sesio post salina injekto (ip, 1 ml / kg) en grupoj de Sprague Dawley ratoj (n = 8 / grupo) antaŭe eksponitaj al ĉiutagaj kunsidoj de 15 kun sakrosa rekompenco (solvaĵo 10%) transdonitaj sub 0, 25, 50, 75 aŭ 100% ŝanĝiĝemaj horaroj. La kondiĉita stimulo estis lumo (120 s). Grupo 0 ricevis la saman nombron da rekompencoj kiel la grupo 100 en manko de kondiĉitaj stimuloj.
Efikoj de 0.5 mg / kg AMPH.
Antaŭ-injekta movado. 5 Grupo × 2-Sesio ANOVA de antaŭ-injekta movado (30-min) dum la antaŭ-kaj post-sentiviga 0.5 mg / kg AMPH-tagaj tagoj donis signifan ĉefan efikon de Sesio, F(1, 35) = 13.39, p = 0.001, kaj neniuj aliaj signifaj efikoj, F(4, 35) <1.79, p > 0.15. Planitaj komparoj trovis neniujn signifajn diferencojn inter grupo 50 kaj grupo 0 aŭ grupo 100 dum la unua sesio, t(70) <1.00, p > 0.30. Tamen, en la dua (post-sentiviga) kunsido la grupo 50 (1203; 121) montris signife pli da antaŭinjekciaj radiaj paŭzoj (M; SE) ol la grupo 100 (756; 103), t(70) = 5.11, p <0.001, sed ne diferencis de la grupo 0 (1126; 211), t(7) <0.88, p > 0.40. Sekve, bazaj diferencoj en movado ne kalkulis grupajn diferencojn en lokomotora respondo al la unua 0.5 mg / kg dozo de AMPH sed eble kontribuis al diferencoj inter grupo 50 kaj grupo 100 en lokomotora respondo al la dua 0.5 mg / kg dozo de AMPH. . Duonaj (SE) radiaj paŭzoj por la antaŭinjekcia fazo sur la unua kaj dua 0.5 mg / kg AMPH-testaj kunsidoj estis 757 (41) kaj 974 (59).
Post-injekta movado. 5 Grupo × 2-Sesio ANOVA de lokomotora respondo al 0.5 mg / kg AMPH antaŭ kaj post la 5-doza sentiviganta reĝimo donis ĉefan efikon de Sesio, F(1, 35) = 76.05, p <0.0001, kaj neniuj aliaj signifaj efikoj, F(4, 35) <1.10, p > 0.37. Figuro 5 montras la mezajn poentarojn por ĉiu grupo kaj sesio.
Figuro 5. Meznivela (SE) lokomotora respondo (nombro de trabo-paŭzoj en elektronika tabelo per 90 min) al 0.5 mg / kg d-amfetamino en apartaj kunsidoj antaŭ kaj post 5-sesia sentema reĝimo de d-amfetamino (1.0 mg / kg; ip po sesio) en grupoj de Sprague Dawley-ratoj (n = 8 / grupo) antaŭe eksponitaj al ĉiutagaj kunsidoj de 15 kun sakrosa rekompenco (solvaĵo 10%) transdonitaj sub 0, 25, 50, 75 aŭ 100% ŝanĝiĝemaj horaroj. La kondiĉita stimulo estis lumo (120 s). Grupo 0 ricevis la saman nombron da rekompencoj kiel la grupo 100 en manko de kondiĉitaj stimuloj. *p <0.05 por averaĝa diferenco inter grupo 50 kaj grupo 0 same kiel grupo 100, surbaze de planitaj komparoj.
La figuro montras, ke la Sesia efiko implikis signifan pliiĝon de totala meznombra (SE) trabo rompiĝas per 90 min de 0.5 mg / kg dozo 1, 3674 (216) al 0.5 mg / kg dozo 2, 6123 (275). La manko de interago aŭ grupa efiko sugestis, ke sentivigxo al AMPH ne variis fidinde tra grupoj. Malgraŭ la manko de signifaj grup-rilataj efikoj en la ANOVA, inspektado de la figuro montras, ke la grupo 50 montras la plej grandan respondon al la unua kaj dua duonaj 0.5 mg / kg. Planitaj komparoj de respondo al la unua 0.5 mg / kg-dozo rivelis neniun signifan diferencon inter grupo 50 kaj grupo 0 aŭ grupo 100, testas(35) <0.48, p > 0.50. Tamen, responde al la dua (post-sentiveco) 0.5 mg / kg dozo, grupo 50 montris signife pli grandan movadon ol grupo 0, t(35) = 2.00, p <0.05, same kiel grupo 100, t(35) = 3.29, p <0.01.
En lumo de la signifa grupa diferenco en antaŭ-injekta movado ĉe la dua 0.5 mg / kg AMPH-sesio raportis pli supre, estis farita sekvaĵo 5 Grupo × 2-Sesio ANCOVA de lokomotora respondo al 0.5 mg / kg AMPH, kontrolante por antaŭ- injekta movado dum la dua sesio. Ĉi tiu analizo donis signifan efikon de la kovariato, F(1, 34) = 8.65, p = 0.006, ĉefa efiko de Sesio F(1, 34) = 10.83, p = 0.002, kaj neniuj aliaj signifaj efikoj, F(4, 34) <0.85, p > 0.50. Grave, planitaj komparoj bazitaj sur la MS-eraro kaj df-eraro de la ANCOVA konfirmis, ke averaĝa lokomotora respondo al la dua dozo de AMPH de 0.5 mg / kg restis signife pli granda en grupo 50 ol grupo 100, t(34) = 3.09, p <0.01, kaj grupo 0, t(34) = 1.88, p <0.05 (unu-vosta), kiam antaŭinjekta variado de kunsido 2 estis kontrolita. Tiel, grupo 50 montris signife pli grandan post-sentivigan lokomotoran respondon al 0.5 mg / kg AMPH ol grupon 100 aŭ grupon 0, kaj ĉi tiuj grupaj diferencoj ne estis peritaj de antaŭinjekta movado en testotagoj.
Efikoj de 1.0 mg / kg AMPH.
Antaŭ-injekta movado. 5 Grupo × 5-Sesio ANOVA de 30-min-antaŭ-injektaj poentaroj por la 1 mg / kg AMPH-sentivigaj sesioj donis ĉefan efikon de Sesio, F(4, 140) = 16.70, p <0.0001, kaj neniuj aliaj signifaj efikoj, F(4, 35) <0.94, p > 0.45. Planitaj komparoj trovis neniun signifan diferencon en antaŭinjekta movado inter grupo 50 kaj grupo 0 aŭ grupo 100 dum la unua kunsido, t(175) <1.66, p > 0.10. Tamen, dum la fina sesio, grupo 50 (1167; 140) montris signife pli da trabo-paŭzoj (M; SE) ol grupo 100 (1000; 99), t(175) = 2.35, p <0.05, sed ne diferencis de la grupo 0 (1085, 120), t(175) <1.16, p > 0.20. Sekve, diferencoj en antaŭinjekta movado kontribuis al diferencoj inter grupoj 50 kaj 100 en lokomotora respondo al la fina 1 mg / kg AMPH-dozo. Duonaj (SE) ĝeneralaj radiaj paŭzoj por la specimeno dum la antaŭinjekcia fazo por Sesioj 1 ĝis 5 estis: 810 (46), 784 (52), 760 (53), 726 (46), 1009 (51).
Post-injekta movado. 5 Grupo × 5-Sesio ANOVA de respondoj al 1 mg / kg AMPH donis signifan ĉefan efikon de Sesio, F(4, 140) = 6.72, p <0.001, marĝena interagado de Grupo × Sesio, F(16, 140) = 1.57, p = 0.085, kaj neniu ĉefa efiko de Grupo, F(4, 35) <0.44, p > 0.77. Polinomaj tendencanalizoj rivelis signifan linian tendencon, F(1, 35) = 9.19, p = 0.005, kaj kuba tendenco, F(1, 35) = 21.63, p <0.001, dum kunsidoj de 1 ĝis 5. Figuro 6 montras la mezajn lokomotorajn poentarojn por ĉiu grupo kaj sesio.
Figuro 6. Meznivela (SE) lokomotora respondo (nombro de paŭzoj de trabo en elektronika tabelo po 90 min) al 1 mg / kg d-amfetamino (ip) sur semajnaj sesioj de 5 en grupoj de ratoj Sprague Dawley (n = 8 / grupo) antaŭe eksponitaj al ĉiutagaj kunsidoj de 15 kun sakrosa rekompenco (solvaĵo 10%) transdonitaj sub 0, 25, 50, 75 aŭ 100% ŝanĝiĝemaj horaroj. La kondiĉita stimulo estis lumo (120 s). Grupo 0 ricevis la saman nombron da rekompencoj kiel la grupo 100 en manko de kondiĉitaj stimuloj.
La figuro montras, ke la Sesio-efiko reflektas signifan pliiĝon de tutaj meznombraj trairoj (SE) por la plena specimeno de sesio 1, 4624 (213) al sesio 5, 5736 (272), konfirmante la aperon de sentivigo al AMPH. La kuba tendenco signifis relativajn maksimumojn ĉe sesioj 1, 3 kaj 5, kun faltoj pri sesioj 2 kaj 4, precipe por grupoj 0 kaj 50. La figuro ankaŭ montras, ke malgraŭ la manko de signifa interago, grupo 25 montris iom post iom pli grandan lokomotivan respondon super sesioj kaj diferencis konsiderinde de la aliaj grupoj en sesioj 4 kaj 5 (9 kaj 22% pli granda ol la sekva plej alta grupo). Planitaj komparoj trovis, ke grupo 50 ne diferencis signife de grupoj 0 aŭ 100, t(175) <0.89, p > 0.40 dum la unua aŭ fina 1 mg / kg AMPH-testsesio.
Kontrolo por variado en nazo-poke respondanta dum trejnado de sakarozo
Du 5 Grupo × 2-Sesio ANCOVA-oj de lokomotora respondo al 0.5 mg / kg AMPH antaŭ kaj post la sentiviga reĝimo, inkluzive de totala nazotukoj dum sakarozo-trejnado kun CS ĉeestanta kaj kun CS-foresto kiel apartaj kovariataj, trovis neniujn signifajn efikojn por ĉu kovariato. F(1, 18) <1.03, p > 0.31. Sekve, aliro respondanta dum trejnado ne mediaciis grupajn diferencojn en respondo al 0.5 mg / kg AMPH.
Du 5 Grupo × 5-Sesio ANCOVA-oj de lokomotora respondo al 1 mg / kg dum la sentivigaj kunsidoj kun tutaj nazfrapoj (ĉeestantaj CS, CS-mankantaj) kiel apartaj kunvariantoj ne donis signifajn efikojn de la kunvarianto dum la CS ĉeestis, F(4, 104) <1.04, p > 0.38, kaj marĝena ĉefa efiko de la kunvariado dum la CS forestis, F(1, 18) = 3.32, p = 0.085.
diskuto
La rezultoj de ĉi tiu studo ne subtenis konstante la hipotezon, ke grupo 50 montros pli altan lokomotoran respondon super sesioj kompare kun la aliaj grupoj. La XFMXG mg / kg-datumoj de AMPH konfirmis la aperon de sentivigxo kun la alternaj tagaj dozon reĝimo. La mastro inter grupoj indikis tendencon por pli granda sentivigado dum la lastaj sesioj en grupo 1, sen tia pruvo por grupo 25. En kontrasto, la 50-mg / kg-dozo rezultoj indikis tendencon por pli granda sentivigo en grupo 0.5, dum en la sama tempo konfirmas signifan totalan pliiĝon de lokomotora respondo trans grupoj al la dua kontraŭ la unua 50 mg / kg AMPH-dozo. La nula efiko de sala injekto konfirmis, ke espero aŭ injekto-rilato ne kontribuis al la AMPH-efikoj.
La datumoj pri naso-poke denove malkaŝis totalan kreskon de alproksimiĝo dum la trejnaj kunsidoj kiam la CS ĉeestis, kun neniu responda kresko kiam la CS forestis. Tial la bestoj ekhavis la asocion inter la CS kaj la perspektivo de sakrosa rekompenco. Grupaj diferencoj en la frekvenco de nazaj pokoj kiam la CS ĉeestis konformis proksimume al la ofteco de rekompenco sub la respektivaj horaroj, kaj grupoj 75 kaj 100 montras la plej multajn nazajn pokojn, grupo 50 montras interajn nombrojn de nazaj pokoj, kaj grupojn 0 kaj 25 Montrante la plej malmultajn nazajn pufojn. Ĉi tiuj rezultoj sugestas, ke la CS venis al kontrolprocento respondanta en maniero konforma al la ĝenerala probablo de rekompenco. Kvankam spekulema, unu ebla klarigo pri la pli malaltaj nazaj poke-ritmoj kun CS ĉeestantaj en grupo 50 en eksperimento 2 vs eksperimento 1 eble estas la mallongigo de la interprocezo, ĉar pli longaj inter-provaj intertempoj (eksperimento 1) ŝajnas instigi impulsemajn tendencoj kaj ĉi tio estas asociita kun pliigita rotacio de DA en kortikaj antaŭaj, prelimbaj kaj infralimbaj kortikoj (Dalley et al., 2002). Tial, la 30% redukto en intertempa intervalo en eksperimento 2 (kaj 3) eble havas ŝanĝitajn kortikajn DA-nivelojn kaj antaŭenigis pli da elekta (te gvidata de la relativa frekvenco de rekompenco) kontraŭ impulsiĝema (ne gvidata de rekompensa frekvenco) aliro respondante en grupo 50 dum trejnadaj provoj en eksperimento 2 kompare kun eksperimento 1.
La manko de signifaj covariate-rilataj efikoj por nazotranĉoj en la CS-nuna kondiĉo en la ANCOVA indikas ke aliro respondanta dum sakarozo-trejnado ne mediis la efikojn de la malsamaj CS-sakarozaj horaroj pri respondoj al AMPH. La marĝene signifa efiko de la kunvarianto por la malĉefa kondiĉo en la ANCOVA de lokomotoraj respondoj al 1 mg / kg AMPH sugestas, ke la tendenco al trofrua liber-respondo klarigis kelkajn el la variemo en lokomotoraj efikoj de AMPH dum la sentivigaj sesioj.
Kune, la indikaĵoj sugestas, ke la efikoj de kondiĉa historio povas esti pli distingeblaj kun 0.5-AMPH ol kun 1 mg / kg AMPH, kaj ke protokolo kiu generas sentivigon en manko de iu ajn alia manipulado povas malhelpi aŭ redundigi la efikojn de supozita sensitiga-antaŭeniga konduta manipulado (te kronika variablo rekompenco).
Konduta sentivigo al AMPH estas fortika efiko en la laboratorio. Tamen ekster la laboratorio nur minoritato de homoj, kiuj ludas kronike, grimpas al patologiaj niveloj. Kvankam risko por sentivigo rilatas al risko por toksomanio (aŭ serĉado de drogoj), precipe por psikostimulantoj (Vezina, 2004; Flagel et al., 2008), multaj faktoroj krom sensibiliza risko eble predikos unu al toksomanio (ekz., Verdejo-Garcia et al., 2008; Conversano et al., 2012; Volkow et al., 2012). Tamen, ecoj de trajtoj kiuj donas al sentiveco povas interagi kun kondiĉita historio por akcenti la efikojn de neantaŭvidebla rekompenco (te 50% CS-usona programo) pri DA-reagemo. Por esplori ĉi tiun eblon, eksperimento 3 uzis la saman procedon kiel eksperimento 2 sed uzis Lewis-streĉadon anstataŭ Sprague Dawley-ratoj.
Sprague Dawley-ratoj montras mezajn nivelojn de DA-transportiloj, kun pli malaltaj niveloj ol Wistar-streĉaj ratoj (Zamudio et al., 2005), sed pli altaj niveloj ol ratoj Wistar-Kioto ("deprima" simila speco) en la kerno accumbens, amigdala, ventra tegmentala areo kaj substantia nigra (Jiao et al., 2003). Ĉi tiu profilo povas fari ke Sprague Dawley-ratoj nur modere sentas al mediaj aŭ farmacologiaj manipuladoj de DA-funkcio. En kontrasto, Lewis ratoj montras malaltajn nivelojn de DA-transportiloj same kiel D2 kaj D3 DA-ricevilojn en la kerno akumbens kaj dorsal striatum kompare kun aliaj streĉoj (ekz. F344) (Flores et al., 1998). Ĉi tiuj morfologiaj diferencoj povas kontribui al la diferenca respondo de Lewis-ratoj al DA-manipuladoj. Lewis-ratoj ankaŭ montras gamon da akcentitaj respondoj al eksperimentaj drogmanipuladoj kompare kun aliaj trostreĉoj (ekz. F344). Plej grave, Lewis-ratoj montras pli grandan sentemon al metamfetamino, karakterizita per malalta respondo al komencaj dozoj sed pli alta respondo al postaj dozoj (Camp et al., 1994). Lewis ratoj ankaŭ montras pli grandan lokomotivan sentiviĝon al gamo da dozoj de kokaino (Kosten et al., 1994; Haile et al., 2001). Surbaze de ĉi tiu padrono de efikoj, ni supozis, ke Lewis-ratoj ebligus nin esplori, ĉu susceptibilidad al sentivigado pligrandigas la efikojn de kondiĉa kalendaro je posta respondo al AMPH.
Eksperimento 3
Materialoj kaj metodoj
La metodaro estis la sama kiel en eksperimento 2, krom la uzo de Lewis-ratoj (200-225 g al alveno, Charles River, Quebec, Canada).
rezultoj
Nazo poke dum sukerokondiĉoj
5 Grupo × 15-Sesio × 2-Fazo (CS-ĉeestanta, CS forestanta) ANOVA de nazotruoj donis signifajn ĉefajn efikojn de Grupo, F(4, 34) = 6.12, p = 0.001, Sesio, F(14, 476) = 3.42, p <0.001, kaj Fazo, F(1, 34) = 20.83, p <0.001, same kiel signifa tridirekta interago, F(56, 476) = 1.56, p = 0.008. Paneloj (A, B) de Figuro 7 mapu la meznombrajn poentojn de la grupoj por la ĉeestantaj CS kaj forestantaj CS respektive. Komparo de la du paneloj malkaŝas, ke la ĉefa efiko de Fazo reflektis pli ĝeneralajn nazajn respondojn kiam la CS ĉeestis kontraŭ forestanta. Sekve, respondaj respondoj okazis signife pli ofte ol antaŭmaturaj respondoj. La ĉefaj efikoj de Grupo kaj Sesio ne estis facile interpretitaj pro la pli alta interago. La tridirekta interago reflektis konverĝon de poentaroj por la kvin grupoj je relative stabila malalta nivelo tra kunsidoj kiam la CS forestis [Panelo (B)], kune kun diverĝo de dudekopo kiam la CS ĉeestis en relative diskretaj profiloj por ĉiu grupo kiu egalas sian rangordon de rekompencofrekvenco: de plej alta (grupo 100) ĝis plej malalta (grupo 25) [Panelo (A)]. Nur la lineara tendenco por la interago estis signifa, F(4, 34) = 4.03, p = 0.009, reflektante la ĝenerale konsekvencan pliiĝon de nazoj dum sesioj en grupo 100 kiam la CS ĉeestis kiel kontraŭ la relative malkonsekvenca profilo de pliiĝo de nazoj tra sesioj en la aliaj grupoj dum ĉi tiu fazo.
Figuro 7. Mezaj (SE) aliroj respondoj (nazaj pokoj) sur 15-sakrosaj trejnaj kunsidoj en grupoj de Lewis ratoj (n = 8 / grupo) eksponita al sakarosa rekompenco (10% solvo) liverita sub 0, 25, 50, 75, aŭ 100%-ŝanĝiĝemaj horaroj. La kondiĉita stimulo estis lumo (120 s). Grupo 0 ricevis la saman nombron da rekompencoj kiel la grupo 100 en manko de kondiĉitaj stimuloj. (A) Poentaroj kiam CS ĉeestis (5 s × 20-provoj). (B) Poentaroj kiam CS forestis (mezumo por 5 × 20 s dum lumo estis malŝaltita).
Kutimaro al lokomotoraj skatoloj
5 Grupo × 3-Sesio ANOVA donis ĉefan efikon de Sesio, F(2, 70) = 23.07, p <0.0001, kaj neniuj aliaj signifaj efikoj, F(8, 70) <1.47, p > 0.18. Kurba ŝablono de meznombraj lokomotoraj poentaroj aperis de kunsido 1, 1076 (74), tra kunsido 2, 644 (48), ĝis kunsido 3, 762 (59). Planitaj komparoj de grupo 50 kun grupo 0 kaj kun grupo 100 en la unua kaj fina kutimigaj kunsidoj malkaŝis signife malpli da faskoj en grupo 50 (M = 911; SE = 109) kontraŭ grupo 0 (M = 1103; SE = 176) dum kutima sesio 1, t(105) = 2.02, p <0.05, sed neniu diferenco inter grupo 50 kaj grupo 100 (M = 1066; SE = 150), t(105) <1.20, p > 0.20, en ĉi tiu kunsido. Grupo 50 ne diferencis signife de aŭ grupo 0 aŭ grupo 100 en la fina kunsido, t(105) <0.93, p > 0.30. Sekve, averaĝa sendomaka lokomotora respondo en la ŝlosilaj grupoj ne diferencis konstante antaŭ testado.
Test-kunsidoj
Salaj. 5 Grupo × 2-Sesio ANOVA de lokomotoraj respondoj sur la fina kutima sesio kaj la sala testo-sesio donis signifan ĉefan efikon de Sesio, F(1, 35) = 50.12, p <0.0001, kaj neniuj aliaj signifaj efikoj, F(4, 35) <0.57, p > 0.68. Figuro 8 montras la grupon meznivelajn poentarojn por la du sesioj kaj indikas, ke la Sesia efiko reflektas signifan malpliiĝon de kutimo al sala testo. Tiel, ricevo de la injekto en si mem (ekz. atendado, streso) ne plibonigis lokomotoron respondi.
Figuro 8. Meza (SE) lokomotora respondo (nombro da trabo-paŭzoj en elektronika tabelo po 90 min) pri la lastaj el 3-sen-drogaj sesioj de drogoj kaj en posta sesio post salina injekto (ip, 1 ml / kg) en grupoj de Lewis-ratoj. (n = 8 / grupo) antaŭe eksponitaj al ĉiutagaj kunsidoj de 15 kun sakrosa rekompenco (solvaĵo 10%) transdonitaj sub 0, 25, 50, 75 aŭ 100% ŝanĝiĝemaj horaroj. La kondiĉita stimulo estis lumo (120 s). Grupo 0 ricevis la saman nombron da rekompencoj kiel la grupo 100 en manko de kondiĉitaj stimuloj.
Efikoj de 0.5 mg / kg AMPH.
Antaŭ-injekta movado. 5 Grupo × 2-Sesio ANOVA de antaŭ-injekta movado donis signifan ĉefan efikon de Sesio, F(1, 35) = 15.04, p <0.001, kaj neniuj aliaj signifaj efikoj, F(4, 35) <1.19, p > 0.33. Planitaj komparoj trovis neniun signifan diferencon inter grupo 50 kaj grupo 0 aŭ grupo 100 en ambaŭ testaj sesioj, t(70) <0.99, p > 0.30. Sekve, bazaj diferencoj en antaŭinjekta movado ne kalkulis grupajn diferencojn en lokomotora respondo al 0.5 mg / kg AMPH. Duonaj (SE) radiaj paŭzoj por la antaŭinjekcia fazo por la unua kaj dua (post-sentiveco) 0.5 mg / kg sesioj estis 325 (25) kaj 473 (36).
Post-injekta movado. 5 Grupo × 2-Sesio ANOVA de lokomotora respondo al 0.5 mg / kg dozo liveritaj antaŭ kaj post kronika 1 mg / kg AMPH donis ĉefan efikon de Sesio, F(1, 34) = 87.44, p <0.0001, kaj neniuj aliaj signifaj efikoj, F(4, 34) <0.94, p > 0.45. Figuro 9 intrigoj la mezaj lokomotoraj poentaroj por ĉiu grupo kaj sesio kaj montras ke la Sesiofekto reflektis pliigitan totalan respondon al la dua 0.5 mg / kg dozo, kongrua kun sentivigado. La figuro ankaŭ montras, ke la grupoj agis tre simile dum sesio 1, sed tiu grupo 50 montras pli da lokomotora agado ol la aliaj grupoj dum sesio 2. Pluraj planitaj komparoj responde al la unua 0.5 mg / kg-dozo rivelis neniujn signifajn diferencojn inter grupo 50 kaj grupo 0 aŭ grupo 100, t(35) <1.28, p > 0.20. Tamen, la grupo 50 montris signife pli grandan lokomotoran respondon al la dua dozo de 0.5 mg / kg ol la grupo 0, t(35) = 4.32, p <0.001, aŭ grupo 100, t(35) = 2.24, p <0.05.
Figuro 9. Meznivela (SE) lokomotora respondo (nombro de trabo-paŭzoj en elektronika tabelo per 90 min) al 0.5 mg / kg d-amfetamino en apartaj kunsidoj antaŭ kaj post 5-sesia sentema reĝimo de d-amfetamino (1.0 mg / kg; ip po sesio) en grupoj de ratoj Lewis (n = 8 / grupo) antaŭe eksponitaj al ĉiutagaj kunsidoj de 15 kun sakrosa rekompenco (solvaĵo 10%) transdonitaj sub 0, 25, 50, 75 aŭ 100% ŝanĝiĝemaj horaroj. La kondiĉita stimulo estis lumo (120 s). Grupo 0 ricevis la saman nombron da rekompencoj kiel la grupo 100 en manko de kondiĉitaj stimuloj. *p <0.05 por averaĝa diferenco inter grupo 50 kaj grupo 0 same kiel grupo 100, surbaze de planitaj komparoj.
Efikoj de 1 mg / kg AMPH.
Antaŭ-injekta movado. 5 Grupo × 5-Sesio ANOVA de antaŭ-injektaj poentaroj 30-min por la sentivigaj sesioj donis ĉefan efikon de Sesio, F(4, 140) = 4.10, p = 0.004, kaj neniuj aliaj signifaj efikoj, F(4, 35) = 1.25, p > 0.31. Laŭplanaj komparoj trovis, ke faskoj dum la antaŭinjekcia fazo (M; SE) estis signife pli malaltaj en grupo 50 (395; 62) ol en grupo 100 (508; 62), t(175) = 2.58, p <0.01, sed ne grupo 0, t(175) <1.83, p > 0.10, dum 1 mg / kg AMPH-sesio 1. Dum la fina 1 mg / kg AMPH-sesio, planitaj komparoj ankaŭ trovis, ke antaŭinjekta movado en grupo 50 (378; 60) estis signife pli malalta ol en grupo 100 (650; 75 ), t(175) = 6.17, p <0.001, sed ne en grupo 0, t(175) <1.84, p > 0.10. Ĉar la direkto de ĉi tiuj grupaj diferencoj (kontrolgrupo = grupo 50) estis kontraŭa al la hipotezita ŝablono, grupaj diferencoj en postinjekta movado, kiuj kongruas kun la hipotezo, ne povas esti atribuitaj al antaŭinjektaj bazaj diferencoj. Mezaj (SE) ĝeneralaj radiaj paŭzoj dum la antaŭinjekcia fazo por Sesioj 1 ĝis 5 estis: 442 (34), 452 (32), 542 (40), 411 (26), 504 (37).
Post-injekta movado. 5 Grupo × 5 Sesioj ANOVA de respondoj al la 1 mg / kg-dozo produktis signifan ĉefan efikon de Sesio, F(4, 140) = 6.15, p <0.001, kaj neniuj aliaj signifaj efikoj, F(4, 35) <0.57, p > 0.68. Polinomaj tendencanalizoj rivelis signifan linian tendencon, F(1, 35) = 9.34, p = 0.004, kaj kuba tendenco, F(1, 35) = 5.08, p = 0.031, la lasta rezulto indikas relativajn maksimumojn ĉe sesioj 3 kaj 5. Figuro 10 argumentas tiujn poentojn kaj montras, ke malgraŭ la manko de signifa interago en la ANOVA, grupo 50 elmontris multe pli grandan lokomotivon ol la aliaj kvar grupoj en respondo al la fina 1 mg / kg dozo. Sekve, planitaj komparoj rivelis signife pli grandan mezan poentarojn pri sesio 5 en grupo 50 ol en ĉiuj aliaj grupoj, t(35) > 3.68, p <0.001.
Figuro 10. Meznivela (SE) lokomotora respondo (nombro de trabo-paŭzoj en elektronika tabelo per 90 min) al 1 mg / kg d-amfetamino (ip) sur 5-semajnaj sesioj en grupoj de Lewis-ratoj (n = 8 / grupo) antaŭe eksponitaj al ĉiutagaj kunsidoj de 15 kun sakrosa rekompenco (solvaĵo 10%) transdonitaj sub 0, 25, 50, 75 aŭ 100% ŝanĝiĝemaj horaroj. La kondiĉita stimulo estis lumo (120 s). Grupo 0 ricevis la saman nombron da rekompencoj kiel la grupo 100 en manko de kondiĉitaj stimuloj. *p <0.05 por averaĝa diferenco inter grupo 50 kaj grupo 0 same kiel grupo 100, surbaze de planitaj komparoj.
Kontrolo por variado en nazo-poke respondanta dum trejnado de sakarozo
Du 5 Grupo × 2-Sesio ANCOVA-oj de lokomotora respondo al 0.5 mg / kg AMPH antaŭ kaj post la sentiviga reĝimo, inkluzive de totala nazotukoj dum sakarozo-trejnado kun CS ĉeestanta kaj kun CS-foresto kiel apartaj kovariataj, trovis neniujn signifajn efikojn por ĉu kovariato. F(1, 32) <0.44 p > 0.51. Du 5-grupaj × 5-sesiaj ANCOVAs de lokomotora respondo al 1 mg / kg AMPH dum la sentivaj kunsidoj kun totalaj nazaj pikoj (CS ĉeestas, CS forestas) kiel apartaj kunvariaĵoj donis neniujn gravajn efikojn de la kunvariaĵo dum la CS ĉeestis aŭ forestis, F(1, 33) <0.14, p > 0.71. Sekve, sendroga aliro respondanta ne respondecis pri grupaj diferencoj en lokomotoraj respondoj al ambaŭ dozoj de AMPH.
diskuto
Sentiĝo disvolviĝis al la efikoj de ripetita 1.0 mg / kg da amfetamino. La kutimaj kaj salaj datumoj konfirmas, ke ĉi tiu efiko ne ŝuldiĝis al antaŭekzistaj diferencoj, atendoj aŭ streĉaj respondoj al la injekto. La ANCOVA-oj kun nazpunktoj konfirmas, ke ĉi tiuj efikoj ne ŝuldiĝis al konduto pri sendrogiĝo. La datumoj pri nazpunktado mem indikis, ke la grupoj akiris la asocion inter la CS kaj eblo de sakarosa rekompenco. La ranga nivelo de la grupoj de respondo al la fino de trejnado egalis la ĝeneralan oftecon de rekompenco sub la malsamaj horaroj de plej alta (grupo 100) ĝis plej malalta (grupo 0), kiel ĝi faris en eksperimento 2. La relative pli malalta entuta mezumo nazaj niveloj en ĉi tiu eksperimento kompare kun eksperimentoj 1 kaj 2 povas reflekti pli selekteman aliron respondantan al signaloj pri rekompenco ĉe Lewis-ratoj (Kosten et al., 2007).
La 0.5-mg / kg-donacaj datumoj montris, ke la komenca lokomotora respondo al AMPH en Lewis-ratoj (figuro) 9) estis iomete subpremita kompare kun Sprague Dawley-ratoj (eksperimento 2; figuro.) 5), sed la ena grupo pliiĝis laŭ la dua dozo en Lewis-ratoj estis konsiderinda (preskaŭ duobla respondo al la unua 0.5 mg / kg-dozo) post la 5-sesa AMPH-reĝimo Plej precipe grupo 50 montras pli grandan lokomotoran respondon. ol ĉiuj grupoj krom grupo 25 al la dua (te post-sentiviga) 0.5 mg / kg AMPH-dozo kaj pli granda lokomotora respondo ol ĉiuj aliaj grupoj, inkluzive de grupo 25, al la fina 1 mg / kg AMPH-dozo (fina sentiviga sesio) .
Resuma analizo de grupaj rangotabeloj laŭ eksperimentoj
Por determini la fidindecon de grupaj diferencoj en sentivigo, ne-parametrika analizo taksis la kontantecon inter grupo kaj rango de mezuma lokomotora respondo al la dua (post-kronika AMPH) 0.5 mg / kg dozo kaj la fina 1.0 mg / kg dozo de AMPH. el la 3-eksperimentoj. La analizo donis signifan efikon, φ = 0.986, p = 0.025, reflektante la fakton, ke grupo 50 rangis unua en ĉiuj krom unu el la komparoj. La supera rango de grupo 50 kompare kun ĉiuj aliaj grupoj responde al la dua (post-kronika AMPH) 0.5 mg / kg dozo estas prezentita en figuro 5 (eksperimento 2) kaj Figuro 9 (eksperimento 3). La supera rango de grupo 50 relative al aliaj grupoj responde al la fina 1.0 mg / kg-dozo estas prezentita en figuro 2 (eksperimento 1) kaj Figuro 10 (eksperimento 3). La sola escepto al ĉi tiu modelo estis la respondo al la fina 1.0 mg / kg dozo en Sprague-Dawley-ratoj en eksperimento 2.
Ĝenerala diskuto
La nuna serio de eksperimentoj testis la hipotezon, ke kronika ekspozicio al vetlud-simila horaro de rekompenco povas sensigi cerbajn DA-vojojn simile al kronika ekspozicio al drogoj de misuzo. La evidenteco por tia efiko sugestus, ke neuroplastikeco, samkiel pensita kontribui al drogmanio, povas esti induktita per kronika ekspozicio al impredecieblaj rekompencaj horaroj. Konforme al la literaturo pri drogodependeco, lokomotora respondo al 0.5 kaj 1.0 mg / kg dozoj de AMPH-indeksita DA-sistema reaktiveco, kun pli granda lokomotivo en respondo al postaj dozoj operacie difinantaj sentivigojn (kp. Robinson kaj Berridge, 1993; Pierce kaj Kalivas, 1997; Vanderschuren kaj Kalivas, 2000).
Entute, la rezultoj kongruas kun nia hipotezo. Tamen, ili ankaŭ indikas konsiderindan variablon en eksperimentaj efikoj pro proceduraj faktoroj. La efikoj de kondiĉa horaro estis modestaj sed koheraj, kun grupo 50 montranta pli grandan respondon ol la aliaj kvar grupoj al ambaŭ dozo post la kvin dozon-reĝimo. Kvankam entute F-Valoroj por grup-rilataj efikoj en la variancia analizo ofte ne-signifaj, esencaj grupaj diferencoj estis konfirmitaj kun duope planitaj komparoj. Tiurilate, oni notu, ke "La aktuala pensado tamen estas tiu entuta signifo [por F en la ANOVA] ne necesas. Unue, la hipotezoj testataj de la totala testo kaj multobla komparo estas tute malsamaj, kun sufiĉe malsamaj niveloj de potenco. Ekzemple, la ĝenerala F efektive distribuas diferencojn inter grupoj laŭ la nombro da gradoj de libereco por grupoj. Ĉi tio efikas dilui la totalon F en la situacio kie pluraj grupaj rimedoj egalas unu la alian sed malsimilas al ia alia meznombro "(Howell, 1992p. 338). Ĉi tio estas ĝuste la situacio aplikita en la nunaj eksperimentoj, kie atendita grupo 50 diferenciĝos de grupoj 0 kaj grupaj 100-kontroloj sed neniu diferenco inter ĉi tiuj regaj grupoj estis antaŭdirita por grupo 25 aŭ grupo 75.
La nazo poke datumoj konfirmis ke, en ĉiu eksperimento, la bestoj akiris la asocion inter la CS kaj la perspektivo de sakarozo rekompenco. La korespondado inter nazo-poke-frekvenco por la malsamaj grupoj kaj totala frekvenco de rekompenco sub iliaj respektivaj trejnadaj horaroj sugestas, ke la meza indico de sakarozo rekompencas senpagan aliron al medikamentoj. Tamen, la manko de mediacianta efiko de nazotufoj sur grup-rilataj lokomotoraj respondoj al AMPH en la ANCOVA indikis ke apartaj procezoj bazas la du kondutojn.
En iuj kazoj, la efiko de kondiĉa horaro estis evidenta kiel respondo al la unua AMPH-dozo; en aliaj kazoj ĝi aperis nur post ripetaj dozo. Grupaj diferencoj en lokomotora respondo al la unua AMPH-dozo sugestas, ke ekspono al hazardludaj rekompensaj horaroj sufiĉas memstare por indukti sensibiligon. Grupaj diferencoj en lokomocio sekvanta multojn AMPH-dozojn indikas pli subtilan efikon kiu povus esti karakterizita kiel "susceptibilidad", kiu nur manifestas kiam kombinita kun daŭra ekspozicio al la primara sentiviga agento (te amfetamino).
Diferencoj en la ŝablono de respondo trans eksperimentoj sugestas, ke pli longa intervalo inter trejnado kaj komenca AMPH-defio povas maksimumigi la okazon detekti la enan sensivigan efikon de la traktado. Ĉi tio siavice sugestas, ke efikoj de kondiĉita rekompenca ekspozicio eble kovriĝos kun la tempo, fenomeno ankaŭ vidita kun stimula sentivigo (Grimm et al., 2006). La modelo de respondo al la du dozoj de anfetamino sugestas, ke la dozo 0.5 mg / kg povas esti pli efika por malkaŝi la efikojn de kondiĉita historio. Ĉi tio siavice sugestas, ke kondiĉaj efikoj sub la nuna trejnada protokolo estas iomete subtilaj kaj povas esti kamuflitaj per plafonaj efikoj sub dozo de AMPH kaj kondiĉoj kiuj generas de novo sentiveco.
En eksperimento 3, la bifásica ŝablono de respondo al 0.5 mg / kg dozon kaj progresiva ekapero de supereco en grupo 50 konformas al la atendita profilo por ratoj Lewis en respondo al metamfetamino (Camp et al., 1994). Ĉi tio subtenas la validecon de la nunaj trovoj kaj sugestas koincidon inter la faktoroj kiuj moderigas vundeblecon al psikosimulanta sentivigado kaj al vetludaj horaroj de rekompenco.
Tra eksperimentoj, la post-sensibiliza lokomotora respondo de la grupo 50 ĝenerale superis tiun de la aliaj grupoj sub malsamaj dozoj de amfetamino kaj en malsamaj streĉoj de bestoj. Tamen, la alta ene-grupa ŝanĝiĝemo kaj modesta inter-grupaj efikgrandecoj indikas rolon por aliaj faktoroj en la DA-reaktiveco al amfetamino post eksponado al diversaj horaroj de rekompencita sukroza rekompenco. Kvankam respondoj de DA-neŭronoj por rekompenci signalojn povas provizi krudan modelon de vetludado (Fiorillo et al., 2003), kiel ĉiuj modeloj, estas perdo de informo por parsimonio - te, por montri ŝlosilan procezon. Rezulte, la modelo de efikoj tra CS-US kondiĉoj en la origina Fiorillo et al. studo ne plene ĝeneraligas al lokomotora respondo al anfetamino. Pliaj plibonigoj de la modelo estas postulataj por plene kapti la aspektojn de vetludado, kiuj efikas sur la DA-funkcia sistemo.
Kunigitaj, la rezultoj de ĉi tiu serio de eksperimentoj provizas provizorajn subtenojn por la hipotezo, ke kronika ekspozicio al vetlud-similaj horaroj de rekompenco plibonigas la reakcion de la cerba DA-sistemo al psikostimulanta defio. Kiel tia, ili etendas la trovojn de Singer et al. (2012) kiu pruvis tion, rilate al antaŭfiksita horaro, antaŭa eksponiĝo al ŝanĝiĝema plifortiga horaro en operanta paradigmo plibonigas postan lokomotivan respondon al amfetamino. Pli specife, la nunaj trovoj atentigas pri necerteco de rekompenco-liverado kiel la kritika faktoro sub la efikoj de ŝanĝiĝema rekompenco. La grando de efikoj en la operanta paradigmo estis substance pli granda ol la trovitaj efikoj en la nunaj eksperimentoj. Ĉi tio eble reflektas pli grandan kronikan ekspozicion al la videoludado kiel ludado (55 vs. 15-tagoj); ĝi eble reflektas la efikojn de postuli operan respondon por eligi la rekompencon (t.e., rolon por agentejo) anstataŭ pasiva ekspozicio, kiel en la nuna studo. Pliigi la daŭron de trejnado en la nuna paradigmo helpus solvi ĉi tiujn demandojn.
La valideco de ŝanĝiĝema rekompenco kaj plifortigaj horaroj kiel modeloj de vetludado ne povas esti elprenita de ĉi tiuj eksperimentoj. Estonta esplorado, kiu ekzamenas la efikon de kondiĉa historio sur risko-konduto en ronĝulaj videoludaj taskoj, povus pritrakti ĉi tiun temon. Simile, la korespondado inter la kondutisma sentivigo trovita ĉi tie kaj la levita striatala DA respondo al amfetamino ĵus trovita en patologiaj ludantoj devas atendi plian esploron (Boileau et al., 2013). Mikro-dializo povus trakti ĉi tiun demandon, kaj la prognozo bazita sur la homaj datumoj estus ke pli granda DAa liberigo en la grupo 50 "hazardludfenotipo" estus plej klare observata en la dorsaj (sensiĝimotoraj) striaj anstataŭ la ventra (limba) striato . Validigo de 50% -ta variablo CS + rekompenco en ĉi tiuj aliaj paradigmoj subtenus ĝian utilecon kiel bona fide eksperimenta modelo de PG.
Dum iuj ludmanieroj klare kaŭzas instrumentan respondon (ekz. Ludaŭtomatoj), en aliaj ludmanieroj (ekz. Loterio) la ligo inter la ago (aĉeti la bileton, t.e., meti veton), la indikoj por rekompenco (t.e. , loteriaj nombroj) kaj la rekompenco mem (la alloga nombro kaj mona rekompenco) estas multe pli difuza. Tamen, aktivado de DA dum la CS-Usono-intervalo povas okazi. Ĉi tio povas klarigi kial, kiam la "venka numero" estas anoncita, atento estas fiksita kiam ĉiu individua loteria pilko falas sinsekve por komponi la specifan sinsekvon de ciferoj en la gajnanta numero. Kvankam la probablo de specifa cifero aperi estas matematike difinita, la rezulto por ĉiu individua loteria pilko estas binara - trafita (kongruas kun la numero de la ludanto) aŭ maltrafas (ne kongruas kun la numero de la ludanto) - kaj la rezulto pri iu ajn donita provo estas nekonata. Tia scenaro povas pli bone karakterizi la sperton de grupo 50 en la nunaj eksperimentoj, kie rekompenco estis donita nekontingente sed ankaŭ neantaŭvideble kaj la CS nur indikis la eblon de rekompenco sen malkaŝi ĉu ĝi okazus dum donita provo. Slot-maŝinoj pli forte ligas kun PG ol loterioj (Cox et al., 2000; Bakken et al., 2009), indikante gravan rolon por instrumentaj faktoroj (kaj tugeco) en la rekompencaj aspektoj de hazardludo por ĉi tiu loĝantaro (Loba et al., 2001). Tamen, la Pavlovia procezo modelita en la nunaj eksperimentoj (CS + necerta rekompenco) ŝajnas esti necesa se ne sufiĉa elemento de la hazardluda sperto.
Kune kun la manko de klara instrumenta postulo, kelkaj aliaj dezajnaj ecoj eble kontribuis al la relative modesta kaj varia ŝablono de eksperimentaj efikoj. La grupoj malsamis en totala sakarozo malkovro same kiel la contingencia inter CS kaj sakarozo rekompenco. Kvankam ĉi tio eble kontribuis al inter-grupa variebleco, ĝi ne povas facile klarigi kial bestoj kun plej granda sakarosa ekspozicio (grupo 100) montras malpli da sentivigxo ol grupo 50. Krome, grupo 0 ricevis neniun stimulon antaŭ la malkovro de sakarozo ĉe ĉiu testo. Kvankam ĉi tio malebligis atendeblan atendon de rekompenco, ĝi ne kontrolis la ĉeeston de stimulo antaŭ rekompenco, kiu ekzistis en ĉiuj aliaj grupoj. Por trakti ĉi tiun problemon, estontaj esploroj devus inkluzivi kondiĉon kie bestoj ricevas rekompencon en ĉiu testo post hazarda ekspozicio al neŭtrala stimulo (te kies ĉeesto ne indikas potencialon de rekompenco).
Alia dezajno-limigo estas la ebla apero de aldona konduto, kiu povus influi la efikojn de trejnada horaro. Fronte al necerteco, bestoj povas disvolvi superstiĉajn kondutojn desegnitajn por plibonigi perceptitan kontrolon kaj malpliigi DA-aktivigon de induktaj necertecoj (kp. Harris et al., 2013). Estas do eble, ke neregataj aspektoj de la eksperimenta dezajno ebligis al la bestoj kompensi la efikojn de kondiĉa horaro. Tia efiko povus kontribui al la relative modesta kaj varia respondo al anfetamina en grupo 50 post CS + sakarosa trejnado. Estontaj esploroj registros spontanean konduton, krom nazotruoj, dum trejnadaj sesioj por testi ĉi tiun eblecon, kaj kontroli ĝin statistike se ĝi aperos. Ĉar tia konduto estus atendita kontraŭstari aŭ malfortigi la efikojn de hor-induktita necerteco, lokomotora respondo al anfetamino en grupo 50 devus esti plibonigita kiam ĝi estas kontrolita (procedure aŭ statistike). Tial, la nuna (nekontrolata) dezajno provizas konservativan teston de la efikoj de 50% CS + rekompenco al anfetamina sentivigo.
Koncerne al ekstera valideco, la uzo de viraj ratoj ankaŭ limigas ĝeneraligeblajn rezultojn. La manko de klara "puno" diferencas ankaŭ de hazardludo, kie oftaj monaj perdoj oftas kaj praktikas gravajn motivajn efikojn (Nieuwenhuis et al., 2005; Singh kaj Khan, 2012). La kapablo akumuli rekompencon ankaŭ forestas de la aktuala paradigmo kaj akumulaj gajnoj en fendo-maŝina ludo estis trovitaj interagi kun DA-manipuladoj en homoj (Tremblay et al., 2011; Smart kaj aliaj, 2013). Simile, la ŝanco por premtipo estas grava diferenco inter la nuna modelo kaj reala vetludado.
Malgraŭ ĉi tiuj limoj, la nunaj rezultoj sugestas, ke 50% ŝanĝiĝema CS + rekompenco povas engaĝi DA-vojojn implikitajn en plifortigaj efikoj de hazardludo (Fiorillo et al., 2003; Anselme, 2013). Interkruciĝrezulto de respondo al AMPH sekvante ĉi tiun hazardlud-similan horaron kongruas kun pivota rolo por DA en hazardludo kaj psikostimulaj drogaj efikoj (Zack kaj Poulos, 2009), kaj etendas pli fruajn studojn pri interkruciĝo de motivado ludi de AMPH en patologiaj ludantoj (Zack kaj Poulos, 2004). La aktualaj rezultoj ankaŭ nerekte sugestas, ke modestaj dozoj de AMPH, kiuj ne kaŭzas suprafiziologian liberigon de DA, eble pli bonas modeligi cerban aktivecon en respondo al intermitaj rekompencaj signaloj (t.e., dum vetludado) ol ekspozicio al alta (t.e., binge-simila) dozon da stimulaj drogoj (kp. Vanderschuren kaj Pierce, 2010). Rekta subteno por ĉi tiu korespondado povus esti derivita per taksado de liberigo de DA en respondo al la 50% variablo CS-usona kalendaro kaj malsamaj dozoj de AMPH uzanta mikrodializon.
De eksperimenta starpunkto, la nuna Pavlova modelo kaj la antaŭa operanta modelo de varia plifortigo ŝajnas generi fenotipon similan al la homa patologia vetludanto. Kiel tia, ili provizas valoran komplementon al taskoj pri vetludado de roduloj, kiuj modeligas hazardludan konduton (kiel dependa mezuro) sed ĝis nun nur dungis sanajn bestojn, la ekvivalento de homaj sociaj ludantoj. Surbaze de la literaturo, la bestoj kronike ekspoziciitaj al varia rekompenco eble diferencas en ĉi tiuj taskoj, precipe kiel respondo al DA-ergaj medikamentoj. Kombinado de la reta vetludotipa fenotipo kun vetludado povas permesi sisteman evoluon de medikamentoj por la kuracado de PG, kiuj eble ne estos plene plenumitaj kun sanaj bestoj sole. Pliaj plibonigoj en la eksperimenta desegno kaj trejna reĝimo, kiel priskribita supre, devus plibonigi la korespondadon inter bestoj trejnitaj en ĉi tiu paradigmo kaj realaj patologiaj ludantoj.
De la klinika-sociologia vidpunkto, la trovo de eksponiĝo al 50% variablo CS + rekompenco, kiu tre kongruas kun la rekompenca horaro sur komerca fendmaŝino (Tremblay et al., 2011), ŝanĝas la cerbon DA-sistemon per fidindaj kaj daŭraj manieroj sugestas, ke en iuj kazoj hazardluda agado, kiel drogoj de misuzo, povas esti "patogeno" kapabla kaŭzi toksomanion. Tamen, la modesta efikogrando kaj alta variado en respondo al rekompenco de 50% CS + sugestas, ke, kiel medikamentoj de misuzo, la tendenco de vetas rekompensajn horarojn por antaŭenigi dependecon dependas forte de la antaŭekzista riska profilo de la vetludanto. Tamen, por ŝpari alt-riskajn individuojn eksponajn al eblaj malfavoraj hazardludaj efikoj, ŝajnas akcepteble, ke politikoj aplikitaj por malinstigi uzon kaj minimumigi damaĝojn de drogaj drogoj ankaŭ povus esti etenditaj al vetludado ankaŭ.
Konflikto pri Interesa Rakonto
La aŭtoroj deklaras, ke la esplorado estis farita sen manko de komercaj aŭ financaj rilatoj, kiujn oni povus konsideri kiel ebla konflikto de intereso.
Dankojn
Ĉi tiu esplorado estis financita per subvencioj de La Natura Scienco kaj Inĝenierio-Esplora Konsilio de Kanado al Paul J. Fletcher. Ni sincere dankas al s-ino Djurdja Djordjevic pro la preparado de la ciferoj.
Referencoj
Anselme, P. (2013). Dopamino, motivado, kaj la evolua graveco de ludado-simila konduto. Behav. Brain Res. 256, 1-4. doi: 10.1016 / j.bbr.2013.07.039
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
APA (2013). Diagnostika kaj Statistika Manlibro pri Mentraj Malsanoj, 5-a Edno. Arlington, VA: Usona Psikiatra Eldonejo.
Bakken, IJ, Gotestam, KG, Grawe, RW, Wenzel, HG, kaj Oren, A. (2009). Problemoj pri hazardludo kaj hazardludo en Norvegio 2007. Skandalo. J. Psychol. 50, 333-339. doi: 10.1111 / j.1467-9450.2009.00713.x
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Boileau, I., Pagisto, D., Chugani, B., Lobo, DS, Houle, S., Wilson, AA, et al. (2013) En vivo evidenteco por pli granda dopamina liberigo de anfetaminoj en patologia hazardludo: studo pri tomografia emisio de pozitronoj kun [C] - (+) - PHNO. Mol. Psikiatrio doi: 10.1038 / mp.2013.163. [Epub antaŭ presaĵo].
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Bolles, RC (1972). Plifortigo, atendado, lernado. Psikolo. Rev. 79, 394 – 409. doi: 10.1037 / h0033120
Camp, DM, Browman, KE, kaj Robinson, TE (1994). La efikoj de metamfetamino kaj kokaino sur motora konduto kaj eksterĉela dopamino en la ventrala striatumo de Lewis kontraŭ Fischer 344-ratoj. Brain Res. 668, 180–193. doi: 10.1016/0006-8993(94)90523-1
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Conversano, C., Marazziti, D., Carmassi, C., Baldini, S., Barnabei, G., kaj Dell'Osso, L. (2012). Patologia hazardludo: sistema revizio de biokemiaj, neŭrobildaj kaj neŭropsikologiaj trovoj. Harv. Rev-Psikiatrio 20, 130-148. doi: 10.3109 / 10673229.2012.694318
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Cox, BJ, Kwong, J., Michaud, V., kaj Enns, MW (2000). Problemo kaj probabla patologia videoludado: konsideroj de komunuma enketo. Povas. J. Psikiatrio 45, 548-553.
Dalley, JW, Theobald, DE, Eagle, DM, Passetti, F., kaj Robbins, TW (2002). Difektoj en impulsa kontrolo asociitaj kun ton-levita serotonergia funkcio en pre-fronta kortekso de rato. Neuropsychofarmacology 26, 716–728. doi: 10.1016/S0893-133X(01)00412-2
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Everitt, BJ, kaj Robbins, TW (2005). Neŭralaj sistemoj de plifortigo por drogmanio: de agoj al kutimoj al devigo. Nat. Neurosci. 8, 1481-1489. doi: 10.1038 / nn1579
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Fiorillo, KD, Tobler, PN, kaj Schultz, W. (2003). Diskreta kodigo de rekompenco kaj necerteco de dopamina-neŭronoj. scienco 299, 1898-1902. doi: 10.1126 / science.1077349
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Flagel, SB, Watson, SJ, Akil, H., kaj Robinson, TE (2008). Individuaj diferencoj en la atribuo de incita saĝeco al rekompenco-rilato: influo sur kokain-sentivigo. Behav. Brain Res. 186, 48-56. doi: 10.1016 / j.bbr.2007.07.022
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Fletcher, PJ, Tenn, CC, Rizos, Z., Lovic, V., kaj Kapur, S. (2005). Sentiveco al amfetamino, sed ne PCP, malhelpas atenteman fiksitan aron: renversiĝo de agonisto de D1-ricevilo injektita en la median prefrontalan kortekson. Psikofarmacologio (Berl.) 183, 190–200. doi: 10.1007/s00213-005-0157-6
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Flores, G., Wood, GK, Barbeau, D., Quirion, R., kaj Srivastava, LK (1998). Lewis kaj Fischer ratoj: komparo de dopamina transportilo kaj ricevilniveloj. Brain Res. 814, 34–40. doi: 10.1016/S0006-8993(98)01011-7
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Frascella, J., Potenza, MN, Brown, LL, kaj Childress, AR (2010). Dividitaj cerbaj vundeblecoj malfermas la vojon al nesubstancaj toksomanioj: ĉizado de toksomanio ĉe nova artiko? Ann. NY Acad. Sci. 1187, 294-315. doi: 10.1111 / j.1749-6632.2009.05420.x
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Grimm, JW, Buse, C., Manaois, M., Osincup, D., Fyall, A., kaj Wells, B. (2006). Tempa dispartigo de kokainaj dozon-respondaj efikoj sur sukroza avido kaj lokomocio. Behav. Pharmacol. 17, 143 – 149. doi: 10.1097 / 01.fbp.0000190686.23103.f8
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Haile, CN, Hiroi, N., Nestler, EJ, kaj Kosten, TA (2001). Malsamaj kondutismaj respondoj al kokaino estas asociitaj kun dinamiko de mezolimbaj dopaminaj proteinoj en ratoj Lewis kaj Fischer 344. Synapse 41, 179 – 190. doi: 10.1002 / syn.1073
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Harris, JA, Andrew, BJ, kaj Kwok, DW (2013). Revua aliro dum signalo por manĝaĵo dependas de Pavloviano, ne instrumenta, kondiĉado. J. Exp. Psikolo. Anim. Konduto Procezo. 39, 107 – 116. doi: 10.1037 / a0031315
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Howell, DC (1992). Statistikaj Metodoj por Psikologio. Boston, MA: Duxbury.
Ito, R., Dalley, JW, Robbins, TW, kaj Everitt, BJ (2002). Dopamina liberigo en la dorsa striatumo dum serĉado de kokaino sub la kontrolo de drog-aso. J. Neurosci. 22, 6247-6253.
Jiao, X., Pare, WP, kaj Tejani-Butt, S. (2003). Streĉaj diferencoj en la distribuado de dopaminaj transportiloj en rato-cerbo. Prog. Neuropsychopharmacolo. Biol. Psikiatrio 27, 913–919. doi: 10.1016/S0278-5846(03)00150-7
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Koob, GF, kaj Le Moal, M. (2008). Recenzo. Neurobiologiaj mekanismoj por kontraŭaj instigaj procezoj en toksomanio. Filozofoj. Trans. R. Soc. Lond. B Biol. Sci. 363, 3113-3123. doi: 10.1098 / rstb.2008.0094
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Kosten, TA, Miserendino, MJ, Chi, S., kaj Nestler, EJ (1994). La streĉoj de Fischer kaj Lewis montras diferencajn kokainajn efikojn en kondiĉita loko preferita kaj kondutisma sento, sed ne en lokomotora agado aŭ kondiĉita gusto aversio. J. Pharmacol. Eks. Nu. 269, 137-144.
Kosten, TA, Zhang, XY, kaj Haile, CN (2007). Streĉu diferencojn en konservado de kokain-memadministrado kaj ilia rilato al novedzinaj respondoj. Behav. Neŭrosko. 121, 380 – 388. doi: 10.1037 / 0735-7044.121.2.380
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Leeman, RF, kaj Potenza, MN (2012). Similecoj kaj diferencoj inter patologiaj vetludoj kaj malsanoj uzantaj substancojn: fokuso al impulsemo kaj kompulsiveco. Psikofarmacologio (Berl.) 219, 469–490. doi: 10.1007/s00213-011-2550-7
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Leshner, AI (1997). La toksomanio estas cerba malsano, kaj gravas. scienco 278, 45-47. doi: 10.1126 / science.278.5335.45
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Loba, P., Stewart, SH, Klein, RM, kaj Blackburn, JR (2001). Manipuladoj de la ecoj de normaj videoludaj terminaloj (VLT) ludoj: efikoj en patologiaj kaj ne patologiaj ludantoj. J. Gambl. Stud. 17, 297 – 320. doi: 10.1023 / A: 1013639729908
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Martinez, D., Narendran, R., Foltin, RW, Slifstein, M., Hwang, DR, Broft, A., et al. (2007) Dopeta liberigo de amfetamino: markite malakra en dependeco de kokaino kaj prognozo de la elekto mem-administri kokainon. Estas. J. Psikiatrio 164, 622-629. doi: 10.1176 / appi.ajp.164.4.622
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Mateo, Y., Lack, CM, Morgan, D., Roberts, DC, kaj Jones, SR (2005). Malpligrandigita dopamina fina funkcio kaj nesentemo al kokaino sekve de kokina memregado kaj deprivado. Neuropsychofarmacology 30, 1455-1463. doi: 10.1038 / sj.npp.1300687
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Nestler, EJ (2001). Molekula bazo de longtempa plastikeco sub la toksomanio. Nat. Rev. Neurosci. 2, 119-128. doi: 10.1038 / 35053570
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Nieuwenhuis, S., Heslenfeld, DJ, von Geusau, NJ, Marso, RB, Holroyd, CB, kaj Yeung, N. (2005). Aktiveco en homaj rekompencaj cerbaj regionoj dependas forte de kunteksto. Neuroimage 25, 1302-1309. doi: 10.1016 / j.neuroimage.2004.12.043
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Pierce, RC, kaj Kalivas, PW (1997). Cirkvitmodelo de la esprimo de kondutisma sentivigo al amfetamin-similaj psikostimuliloj. Cerbo Res. Cerbo Res. Rev. 25, 192. doi: 10.1016 / S0165-0173 (97) 00021-0
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Robbins, TW, kaj Everitt, BJ (1999). Drogodependeco: malbonaj kutimoj aldoniĝas. naturo 398, 567-570. doi: 10.1038 / 19208
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Robinson, TE, Becker, JB, kaj Presty, SK (1982). Longtempa plifaciligo de amfetamin-induktita rotacia konduto kaj striatala dopamina liberigo produktita de sola eksponiĝo al amfetamino: seksaj diferencoj. Brain Res. 253, 231–241. doi: 10.1016/0006-8993(82)90690-4
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Robinson, TE, kaj Berridge, KC (1993). La neŭra bazo de drogodirekto: stimulo-sentemiga teorio de toksomanio. Cerbo Res. Cerbo Res. Rev. 18, 247 – 291. doi: 10.1016 / 0165-0173 (93) 90013-P
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Robinson, TE, kaj Berridge, KC (2001). Incentivo-sentivigo kaj toksomanio. toksomanio 96, 103-114. doi: 10.1046 / j.1360-0443.2001.9611038.x
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Kantisto, BF, Scott-Railton, J., kaj Vezina, P. (2012). Neantaŭvidebla plifaciligo de sakarino plibonigas lokomotoron respondantan al amfetamino. Behav. Brain Res. 226, 340-344. doi: 10.1016 / j.bbr.2011.09.003
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Singh, V., kaj Khan, A. (2012). Decido en la variantoj de rekompenco kaj puno de la iowa hazardluda tasko: evidenteco de "antaŭvido" aŭ "enkadrigo"? Fronto. Neŭrosko. 6: 107. doi: 10.3389 / fnins.2012.00107
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Skinner, BF (1953). Scienco kaj Homa Konduto. New York, NY: Senpaga Gazetaro.
Inteligenta, K., Desmond, RC, Poulos, CX, kaj Zack, M. (2013). Modafinil pliigas rekompencan salajron en fendo-maŝinludo en patologiaj ludantoj de malalta kaj alta impulsiveco. Neuropharmacology 73, 66-74. doi: 10.1016 / j.neuropharm.2013.05.015
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Tremblay, AM, Desmond, RC, Poulos, CX, kaj Zack, M. (2011). Haloperidol modifas instrumentajn aspektojn de videoludado en patologiaj ludantoj kaj sanaj kontroloj. Toksomaniuloj. Biol. 16, 467-484. doi: 10.1111 / j.1369-1600.2010.00208.x
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Vanderschuren, LJ, kaj Kalivas, PW (2000). Altecoj en dopaminergiaj kaj glutamatergaj transdono en la indukto kaj esprimo de kondutisma sentivigo: kritika revizio de preklinikaj studoj. Psikofarmacologio (Berl.) 151, 99-120. doi: 10.1007 / s002130000493
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Vanderschuren, LJ, kaj Pierce, RC (2010). Sensivigaj procezoj en drogmanio. Curr. Supro Konduto Neŭroscio. 3, 179 – 195. doi: 10.1007 / 7854_2009_21
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Vanderschuren, LJ, Schoffelmeer, AN, Mulder, AH, kaj De Vries, TJ (1999). Dopaminergiaj mekanismoj mediaciaj la longdaŭra esprimo de lokomotora sensivigo post antaŭ-eksponiĝo al morfino aŭ amfetamino. Psikofarmacologio (Berl.) 143, 244-253. doi: 10.1007 / s002130050943
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Verdejo-Garcia, A., Lawrence, AJ, kaj Clark, L. (2008). Malfortikeco kiel vundeblecmarko por substanco-uzaj malordoj: revizio de trovoj de alta riska esplorado, problemaj ludantoj kaj genetikaj asociitaj studoj. Neŭrosko. Biobehav. Rev. 32, 777-810. doi: 10.1016 / j.neubiorev.2007.11.003
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Vezina, P. (2004). Sentiĝo de reakcia dopamina neurona reagemo kaj memadministrado de psikomotraj stimulaj drogoj. Neŭrosko. Biobehav. Rev. 27, 827-839. doi: 10.1016 / j.neubiorev.2003.11.001
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Volkow, ND, Wang, GJ, Fowler, JS, Logan, J., Gatley, SJ, Hitzemann, R., et al. (1997) Malpliiĝis striatala dopaminergia respondo en detoxigitaj kokainaj dependaj subjektoj. naturo 386, 830 – 833. doi: 10.1038 / 386830a0
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Volkow, ND, Wang, GJ, Fowler, JS, kaj Tomasi, D. (2012). Cirkvitoj de toksomanio en la homa cerbo. Annu. Rev-ino Farmacolo. Toksikolo. 52, 321 – 336. doi: 10.1146 / annurev-pharmtox-010611-134625
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Winer, B. (red.). (1971) Statistikaj Principoj en Eksperimenta Dezajno. New York, NY: McGraw-Hill.
Zack, M., kaj Poulos, CX (2004). Amfetaminoj primokas motivon al ludoj kaj hazardludaj rilataj semantikaj retoj en problemaj ludantoj. Neuropsychofarmacology 29, 195-207. doi: 10.1038 / sj.npp.1300333
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Zack, M., kaj Poulos, CX (2009). Paralelaj roloj por dopamino en patologiaj vetludoj kaj psikosimulantoj. Curr. Droga misuzo Rev. 2, 11-25. doi: 10.2174 / 1874473710902010011
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Zamudio, S., Fregoso, T., Miranda, A., De La Cruz, F., kaj Flores, G. (2005). Streĉas diferencojn de dopaminaj receptoroj kaj dopamin-rilataj kondutoj ĉe ratoj. Cerbo Res. Virbovo. 65, 339 – 347. doi: 10.1016 / j.brainresbull.2005.01.009
Pubeda Abstraktaĵo | Plena Teksto Pubita | Plena Teksto de CrossRef
Ŝlosilvortoj: patologia hazardludo, sentivigo, amfetamino, dopamino, necerteco
Citado: Zack M, Featherstone RE, Mathewson S kaj Fletcher PJ (2014) Kronika ekspozicio al vetlud-simila horaro de rekompencaj antaŭdiroj stimuloj povas antaŭenigi sentivigon al amfetamino ĉe ratoj. Fronto. Konduto Neurosci. 8: 36. doi: 10.3389 / fnbeh.2014.00036
Ricevita: 01 Novembro 2013; Akceptita: 23 januaro 2014;
Eldonita interrete: 11 Februaro 2014.
Eldonita de:
Bryan F. Kantisto, Universitato de Miĉigano, Usono
Reviziita de:
Louk Vanderschuren, Universitato de Utrecht, Nederlando
Ruud Van Den Bos, Radboud Universitato Nijmegen, Nederlando
Patrick Anselme, Universitato de Liège, Belgio
Kopirajto © 2014 Zack, Featherstone, Mathewson kaj Fletcher. Ĉi tio estas malferma-alira artikolo distribuita sub la kondiĉoj de la Krea Komunaĵo Atribuka Permesilo (CC BY). La uzo, dissendo aŭ reproduktaĵo en aliaj forumoj estas permesita, se la originala aŭtoro aŭ licencanto estas akredititaj kaj ke la originala publikigado en ĉi tiu ĵurnalo estas citita, laŭ akceptebla akademia praktiko. Neniu uzo, dissendo aŭ reprodukto estas permesata, kiu ne plenumas ĉi tiujn terminojn.
* Korespondado: Martin Zack, Kognitiva Psikofarmakologia Laboratorio, Fako pri Neŭroscienco, Centro por toksomanio kaj Mensa Sano, 33 Russell Street, Toronto, ON M5S 2S1, Kanada retpoŝto: [retpoŝte protektita]
