 Stephanie E. Tedford
Stephanie E. Tedford- 1Departamento de Farmacología, Centro de Comportamiento Compulsivo y Adicciones, Rush University Medical Center, Chicago, IL, EE. UU.
- 2Departamento de Farmacología, Rush University Medical Center, Chicago, IL, EE. UU.
- 3Departamento de Psiquiatría, Rush University Medical Center, Chicago, IL, EE. UU.
El juego patológico es una manifestación de los trastornos del control de los impulsos. Los fundamentos biológicos de estos trastornos siguen siendo esquivos y el tratamiento dista mucho de ser ideal. Los modelos animales de trastornos del control de impulsos son una herramienta de investigación crítica para comprender esta condición y para el desarrollo de medicamentos. Modelar tales comportamientos complejos es desalentador, pero por su deconstrucción, los científicos han recapitulado en animales aspectos críticos del juego. Un aspecto del juego es la toma de decisiones de costo / beneficio en donde uno pesa los costos anticipados y los beneficios esperados de un curso de acción. La toma de decisiones basada en el riesgo / recompensa, la demora y el esfuerzo representan decisiones de costo / beneficio. Estas características se estudian en humanos y se han traducido a protocolos de animales para medir los procesos de toma de decisiones. Tradicionalmente, el reforzador positivo utilizado en estudios con animales es el alimento. Aquí, describimos cómo se puede usar la autoestimulación intracraneal para las tareas de toma de decisiones de costo / beneficio y analizamos nuestros estudios recientes que muestran cómo las terapias farmacológicas alteran estos comportamientos en ratas de laboratorio. Proponemos que estos modelos pueden tener valor en la selección de nuevos compuestos por la capacidad de promover y prevenir aspectos del comportamiento del juego.
Introducción
El juego problemático o inadaptado, incluida la condición extrema denominada juego patológico, se caracteriza por comportamientos que a menudo persisten durante períodos prolongados. El problema del juego puede tener un impacto negativo significativo en el bienestar personal, profesional y financiero. En las últimas dos décadas, las oportunidades de juego se han incrementado a través de cambios en la legislación y la introducción de nuevos lugares (por ejemplo, juegos de azar por Internet). En consecuencia, la prevalencia de los juegos de azar problemáticos ha ido en aumento. No hay tratamientos aprobados por la FDA para este trastorno y, por lo tanto, es fundamental comprender mejor estos comportamientos para desarrollar terapias eficaces.
El problema del juego es un fenómeno complejo, que incluye mayores niveles de toma de decisiones impulsiva (Alessi y Petry, 2003; Dixon et al., 2003; Holt et al., 2003; Kraplin y otros, 2014) que se derivan de evaluaciones desventajosas de costo / beneficios. Las evaluaciones clínicas de la toma de decisiones, que a menudo emplean encuestas y herramientas interactivas basadas en computadora, han sido fundamentales para determinar los perfiles de toma de decisiones subóptimos en diversas patologías, incluidos los jugadores patológicos (Ledgerwood et al., 2009; Madden et al., 2009; Michalczuk et al., 2011; Petry, 2011; Miedl et al., 2012). Con frecuencia, las evaluaciones clínicas se basan en tres aspectos diferentes de la toma de decisiones de costo / beneficio, aunque se superponen, incluyendo los siguientes: (i) la cantidad de riesgo para obtener una recompensa (toma de decisiones de riesgo / recompensa), (ii) a retraso experimentado antes de la entrega de la recompensa (toma de decisiones basada en la demora), y (iii) la cantidad de esfuerzo requerido para obtener una recompensa (toma de decisiones basada en el esfuerzo). Se han desarrollado varias tareas para medir estas características críticas de la toma de decisiones por debajo del nivel óptimo para comprender mejor los procesos que comprenden el problema del juego. En estas tareas, el sujeto elige entre una recompensa pequeña y grande, cada una asociada con contingencias de respuesta específicas. En la toma de decisiones de riesgo / recompensa (es decir, descuento de probabilidad), los sujetos eligen entre una pequeña recompensa entregada consistentemente a altas probabilidades (por ejemplo, 100% de probabilidad de recibir $ 10) y una gran recompensa entregada a diferentes probabilidades (por ejemplo, 10 – 80 % de probabilidad de recibir $ 100). En estudios clínicos y preclínicos, la ausencia de una recompensa esperada es un evento aversivo que provoca respuestas fisiológicas correspondientes (Douglas y Parry, 1994; Papini y Dudley, 1997). La preferencia por la opción más grande y "arriesgada" sobre la pequeña, cierta opción se considera que refleja la toma de decisiones de riesgo / recompensa subóptima, y se ha informado de varias patologías humanas que muestran una impulsividad mejorada (Reynolds et al., 2004; Rasmussen et al., 2010; Dai et al., 2013). En la toma de decisiones basada en el retraso (es decir, el descuento por retraso, una medida de elección impulsiva), la pequeña recompensa se entrega poco después de que se selecciona la opción, mientras que la gran recompensa se entrega después de un retraso variable (por ejemplo, $ 10 ahora o $ 100 en semanas 2). Las personas que muestran una alta impulsividad demuestran preferencia por las recompensas disponibles de inmediato (incluso si son más pequeñas), sobre las recompensas retrasadas (incluso si son más grandes), aunque la última opción puede ser más beneficiosa para la persona (Crean et al., 2000; Reynolds et al., 2004; Bickel et al., 2012). En la toma de decisiones basada en el esfuerzo, el sujeto elige entre una pequeña recompensa entregada después de una pequeña cantidad de esfuerzo, o una gran recompensa entregada después de que se haya ejercido una mayor cantidad de esfuerzo. En esta tarea, se determina la preferencia individual por la opción de gran esfuerzo / gran recompensa y el "punto" en el que el individuo cambia a la opción de bajo esfuerzo / pequeña recompensa. Aún no se han realizado estudios sobre la toma de decisiones basada en el esfuerzo en jugadores humanos, pero serían de gran interés para evaluar la función cognitiva en esta población.
Los protocolos de toma de decisiones utilizados en las evaluaciones clínicas pueden modificarse para estudiar la toma de decisiones en ratas de laboratorio, y estos modelos son críticos para explorar los aspectos conductuales y neurofarmacológicos del juego patológico. En ratas, la toma de decisiones puede evaluarse colocando al animal en una cámara de condicionamiento operante, y permitiendo que el animal elija entre dos palancas (o dos tolvas de nariz-poke) que están disponibles al mismo tiempo. La modalidad de recompensa establecida para el reforzador positivo en estas tareas de roedores es la comida (Tapón y Floresco, 2011; Eubig et al., 2014). Discutimos aquí un método novedoso utilizado en nuestro laboratorio que emplea la estimulación eléctrica directa de las vías de recompensa cerebral (autoestimulación intracraneal; ICSS) para evaluar la toma de decisiones de costo / beneficio en ratas y la contribución de los neurotransmisores monoaminérgicos en la toma de decisiones (Rokosik y Napier, 2011, 2012; Tedford et al., 2012; Personas et al., 2013).
Autoestimulación intracraneal
Un reforzante operante es un estímulo, que cuando se hace dependiente de alguna acción, aumenta la probabilidad de que se repita esa acción. La autoestimulación intracraneal (ICSS, por sus siglas en inglés) es un comportamiento operante en el cual los animales se autoadministran la estimulación eléctrica en regiones del cerebro que se sabe están involucradas en el refuerzo positivo. ICSS se estudió por primera vez en los 1950 cuando James Olds y Peter Milner (Olds y Milner, 1954) determinaron que las ratas volverían repetidamente a una ubicación en una caja donde recibieron estimulación eléctrica para recompensar las regiones relacionadas con el cerebro. Permitieron que las ratas trabajen para esta estimulación cerebral eléctrica (EBS) respondiendo en un manipulandum operante (por ejemplo, presionando una palanca, girando una rueda) (Olds y Milner, 1954). El descubrimiento de esta técnica ha sido fundamental en el mapeo de las vías de recompensa en todo el cerebro, y si bien hay muchas regiones del cerebro que se pueden usar para apoyar el ICSS (Olds y Milner, 1954; Sabio y Bozarth, 1981; Sabio, 1996), está bien documentado que la estimulación del haz del cerebro anterior medial (MFB) promueve resultados de comportamiento profundos y confiables (Corbett y Wise, 1980; Pirch et al., 1981; McCown y otros, 1986; Tehovnik y Sommer, 1997). Los parámetros actuales de estimulación pueden manipularse para afectar el valor de refuerzo de la EBS y, por lo tanto, alterar el comportamiento de ICSS. Estos parámetros incluyen la intensidad (es decir, amperios) de la corriente eléctrica y la frecuencia actual (es decir, hercios). Las elevaciones en ambos parámetros generalmente resultan en un aumento de la excitación de las neuronas relevantes para la recompensa que se estimulan, ya sea al aumentar el número de neuronas involucradas por la estimulación (amperios) (Keesey, 1962; Wise et al., 1992) o aumentando la frecuencia con la que se dispara una población de neuronas (hercios) (Sabio y Rompre, 1989; Sabio, 2005). Las manipulaciones de intensidad de corriente alteran el número de neuronas activadas, es decir, las intensidades de corriente más grandes afectan a una población más amplia de neuronas que las corrientes más pequeñas. Por lo tanto, cuando este parámetro se mantiene constante, la población de neuronas excitadas por EBS es relativamente similar independientemente de la frecuencia actual. La variable de parámetro de estimulación de elección para estos protocolos es la frecuencia actual, ya que esta selección nos permite manipular la velocidad de disparo del mismo grupo de neuronas con efectos mínimos en el tiempo o el espacio de integración de la estimulación. Al manipular estos parámetros de EBS, hemos desarrollado modelos sofisticados de toma de decisiones de costo / beneficio que emplean ICSS (Rokosik y Napier, 2011, 2012; Tedford et al., 2012; Personas et al., 2013). Esta aplicación representa una desviación radical de los estímulos de refuerzo (es decir, alimentos) tradicionalmente utilizados en tareas que evalúan la toma de decisiones en roedores. El ICSS puede proporcionar varias ventajas experimentales sobre los métodos tradicionales de refuerzo. Para facilitar la respuesta operante de los alimentos, la ingesta diaria a menudo está restringida (Feja y Koch, 2014; Hosking et al., 2014; Mejia-Toiber et al., 2014). Esta práctica puede confundir las medidas de resultado, ya que hay una superposición sustancial en los sistemas neurobiológicos que se alteran durante la restricción crónica de alimentos y en aquellos que median la toma de decisiones impulsiva (Schuck-Paim et al., 2004; Minamimoto et al., 2009). Además, los animales reforzados con alimentos se vuelven cada vez más saciados a lo largo de una sesión, lo que disminuye el valor del refuerzo de alimentos (Bizo et al., 1998), aunque este efecto puede depender del tamaño del reforzador (Roll et al., 1995). En contraste con el refuerzo de alimentos, el valor de refuerzo de la EBS se mantiene estable durante una sesión, lo que permite evaluaciones de comportamiento más extensas y consistentes (Trowill et al., 1969). Esta característica permite que las sesiones de prueba se realicen repetidamente a lo largo del día, lo que puede ser beneficioso cuando se estudian los efectos de las terapias farmacológicas, específicamente el tratamiento crónico con medicamentos. Nuestros estudios publicados de descuentos por probabilidad (que se analizan a continuación) se realizaron varias veces al día durante los tratamientos con agonistas crónicos de dopamina (pramipexol). Proponemos que este beneficio de procedimiento es más aplicable a la condición humana y, por lo tanto, proporciona resultados traduccionales mejorados. Hasta la fecha, estudios similares que evaluaron los efectos agonistas de la dopamina en la toma de decisiones impulsiva utilizando la recompensa de alimentos solo han evaluado tratamientos agudos con fármacos (St Onge y Floresco, 2009; Zeeb et al., 2009; Madden et al., 2010; Johnson et al., 2011; Koffarnus et al., 2011) y será de gran interés comparar los resultados de comportamiento después de un tratamiento farmacológico agudo y crónico entre estos diferentes refuerzos. Si bien ICSS ofrece varias ventajas sobre el refuerzo de alimentos, ICSS también presenta varias desventajas. Por ejemplo, el ICSS requiere cirugía y recuperación del cerebro invasivas, y las etapas de la cabeza mal ajustadas pueden provocar la pérdida de sujetos a lo largo del paradigma de comportamiento. A pesar de estos inconvenientes, sostenemos que el ICSS es una alternativa viable al refuerzo de alimentos y presenta ventajas considerables al refuerzo de alimentos en estas tareas de comportamiento.
Las tareas de toma de decisiones de costo / beneficio requieren que se tomen decisiones entre las opciones asociadas con magnitudes de recompensa variables. En consecuencia, los reforzadores utilizados en estas tareas deben demostrar la capacidad de producir tales cambios en la magnitud de la recompensa y, posteriormente, las ratas deben poder discriminar entre la opción de reforzador pequeño (SR) y reforzador grande (LR). En los procedimientos que usan refuerzo de alimentos, esto se logra alterando el número de gránulos de alimentos obtenidos después de una respuesta. En el ICSS, el EBS se puede variar cambiando la intensidad de la corriente de estimulación o la frecuencia actual. Figura 1 ilustra la respuesta a la presión de la palanca obtenida cuando se varía la intensidad de la corriente (es decir, la frecuencia actual se mantuvo constante; Figura 1A) o cuando se varía la frecuencia actual (es decir, la intensidad de la corriente se mantuvo constante; Figura 1B). Cuando se modifica cualquiera de los parámetros, las ratas muestran una presión de palanca moderada para valores pequeños de EBS y muestran tasas de presión de palanca aumentadas para valores grandes de EBS, lo que sugiere que el valor de refuerzo de la estimulación más grande es mayor (independientemente de si se manipula la intensidad o la frecuencia de la corriente). Por lo tanto, EBS se puede adaptar para los reforzadores grandes y pequeños necesarios para los protocolos de toma de decisiones de costo / beneficio. Estos valores de refuerzo pueden determinarse en ratas individuales generando curvas de respuesta de velocidad de presión de palanca estables para cada animal (Rokosik y Napier, 2011, 2012). Alternativamente, se puede generar una curva de población a partir de un grupo de ratas a partir de las cuales se puede determinar un valor de SR y LR estandarizado (Tedford et al., 2012; Personas et al., 2013). Este último enfoque proporciona un medio más eficiente en el tiempo y, sin embargo, confiable para derivar el SR y el LR. En una segunda serie de estudios, utilizamos manipulaciones de intensidad o frecuencia actuales para establecer valores de SR / LR en una tarea de descuento de probabilidad (es decir, toma de decisiones de riesgo / recompensa). Los cambios en los valores de refuerzo de la intensidad de la corriente (es decir, la frecuencia de la corriente se mantuvo constante) y los valores de la frecuencia de la corriente (es decir, la intensidad de la corriente se mantuvo constante), ambos producen un comportamiento de descuento significativo en ratas (Figuras 1C, D). Basado en parte en la inclinación de la curva de descuento, se determinó que la frecuencia actual es el parámetro apropiado para manipular los valores de refuerzo. Una vez que se establece que las ratas pueden distinguir entre las frecuencias de corriente estandarizadas utilizadas para la RS y la LR, se pueden probar en cualquiera de nuestros paradigmas de toma de decisiones mediados por ICSS: (i) toma de decisiones de riesgo / recompensa (Rokosik y Napier, 2011, 2012), (ii) la toma de decisiones basada en el retraso (Tedford et al., 2012), o (iii) toma de decisiones basada en el esfuerzo (Personas et al., 2013).
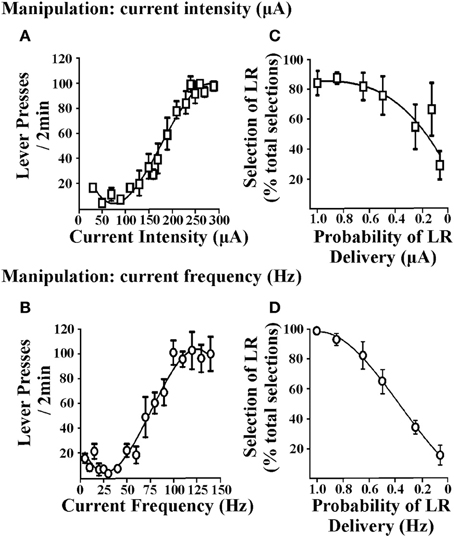
Figura 1. Efectos de los parámetros de estimulación cerebral en la respuesta de la palanca-pulsación y el descuento de probabilidad. Los dos parámetros EBS probados fueron intensidad de corriente y frecuencia de corriente. Palanca de ratas presionada para EBS (en una proporción fija-1 programa de refuerzo) en donde cada 2 min, un parámetro de EBS fue manipulado y el otro parámetro se mantuvo constante. (A) Manipulación de intensidad de corriente. Las intensidades actuales que van desde 10 a 350 μA se presentaron en orden aleatorio (n = 6); La frecuencia actual se mantuvo en 100 Hz. (B) Manipulación de la frecuencia actual. Las frecuencias actuales que van desde 5 a 140 Hz se presentaron en orden aleatorio (n = 3); La intensidad de la corriente se mantuvo constante a un nivel que fue individualizado y determinado en las sesiones de entrenamiento anteriores. La manipulación de la intensidad de la corriente o la frecuencia de la corriente produjo patrones similares de respuesta con la pulsación de palanca. Los datos se muestran como media ± sem para las últimas tres sesiones consecutivas. Las ratas se entrenaron posteriormente en la tarea de descuento de probabilidad y los valores para los reforzadores pequeños y grandes se determinaron individualmente para cada animal mediante el cálculo de las intensidades de la corriente de estimulación efectiva y las frecuencias actuales obtenidas de la curva de respuesta EBS vs. de las tasas máximas de respuesta de la palanca de prensa, respectivamente. Variando la magnitud de la intensidad de corriente (C) o frecuencia actual (D) dio como resultado el descuento del reforzador grande (LR) a medida que disminuía la probabilidad de entrega (es decir, una disminución en el porcentaje de selección de la palanca asociada con la LR sobre las selecciones totales). Los datos se muestran como media ± sem para el día uno de descuento con intensidad actual y 2 días de descuento con frecuencia actual. Figura modificada de Rokosik y Napier (2011) y reimpreso con permiso del editor.
Validación del uso de ICSS para evaluar medidas de impulsividad y toma de decisiones
El desarrollo de nuevos modelos animales requiere una cuidadosa consideración con respecto a la validez. Por lo tanto, al diseñar estas tareas de toma de decisiones mediadas por el ICSS, nos hemos esforzado por verificar la validez de cara a cara y de construcción, y por determinar la probabilidad de validez predictiva.
La validez facial se refiere a la medida en que una prueba parece medir subjetivamente su fenómeno deseado. El diseño de cada tarea de toma de decisiones mediada por el ICSS se basó en los protocolos actuales empleados en humanos para el retraso y el descuento de probabilidad (Rasmussen et al., 2010; Leroi et al., 2013) y otras tareas de toma de decisiones basadas en el esfuerzo (Treadway et al., 2009; Buckholtz et al., 2010; Wardle et al., 2011). En seres humanos, las medidas de toma de decisiones de costo / beneficio se derivan de pedir a los individuos que seleccionen entre varias opciones disponibles con contingencias específicas ubicadas en cada selección (es decir, riesgo, demora o esfuerzo). Emulamos este escenario presentando ratas con dos palancas simultáneamente extendidas, en donde una selección de cualquiera de las palancas está asociada con recompensas pequeñas o más grandes que también se entregan bajo parámetros particulares de contingencia. Por lo tanto, cada una de nuestras tareas de toma de decisiones mediadas por el ICSS demuestra la validez aparente.
La validez de construcción se refiere a la capacidad del paradigma para evaluar con precisión lo que propone medir. En la toma de decisiones basada en el riesgo / recompensa y el retraso, la preferencia por la recompensa grande disminuye a medida que disminuye la probabilidad de entrega, o se incrementa la demora hacia la entrega de recompensa, respectivamente. En la toma de decisiones basada en el esfuerzo, los individuos demuestran una preferencia inicial por la opción esfuerzo alto / recompensa grande cuando el esfuerzo asociado con la recompensa grande se considera razonable. Se observa un cambio en la preferencia al bajo esfuerzo / pequeña recompensa cuando el alto esfuerzo ya no vale la pena el gasto de energía. Está bien documentado que los roedores exhiben patrones similares de riesgo / recompensa, basados en demoras y decisiones basadas en el esfuerzo en comparación con los humanos (Rachlin et al., 1991; Buelow y Suhr, 2009; Jimura et al., 2009), y hemos observado estos perfiles en cada una de nuestras tareas (Rokosik y Napier, 2011, 2012; Tedford et al., 2012; Personas et al., 2013) (por ejemplo, ver Figura 2).
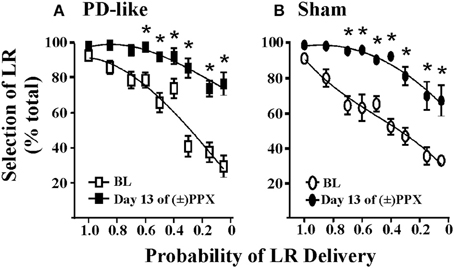
Figura 2. Efectos de pramipexol en la toma de decisiones de riesgo / recompensa utilizando una tarea de descuento de probabilidad. El crónico (±) PPX disminuye el descuento en el tipo PD (A) y control simulado (B) ratas Describiendo brevemente la tarea, como PD (n = 11) y control simulado (n = 10) las ratas fueron entrenadas en la tarea de descuento de probabilidad usando ICSS. Las probabilidades asociadas con la administración del reforzador grande (RL) se presentaron en un orden pseudoaleatorio. Una vez que se observó un comportamiento estable, las ratas se trataron crónicamente con inyecciones dos veces al día de 2 mg / kg (±) PPX durante los días de 13. Los datos mostrados se recopilaron desde el momento en que observamos el efecto máximo en el último día de tratamiento (es decir, 6 h después de la inyección) y se compararon con la línea de base de pretratamiento (BL). Se muestra el porcentaje de selección de la LR (es decir, la proporción de libre elección) frente a la probabilidad de que se haya entregado la LR. Un rmANOVA de dos vías con post hoc Newman-Keuls reveló aumentos significativos en el% de selección de lo incierto, LR después del tratamiento crónico con PPX (*p <0.05) para los grupos de ratas simuladas y similares a PD. Aunque los promedios del grupo indican un aumento inducido por PPX en la toma de decisiones subóptima de riesgo / recompensa, dos ratas en cada grupo mostraron un aumento de menos del 20% desde el valor inicial con la probabilidad más baja probada; por lo tanto, algunas ratas parecían ser insensibles a la capacidad del fármaco para modificar el descuento de probabilidad. Figura modificada de Rokosik y Napier (2012) y reimpreso con permiso del editor.
La validez predictiva se refiere a la capacidad de los modelos para prever relaciones futuras, y planteamos que nuestros modelos pueden usarse para predecir la capacidad de nuevos tratamientos farmacológicos para alterar la toma de decisiones de costo / beneficio. Es decir, al demostrar la prueba de concepto mediante la replicación de los efectos de los agentes farmacológicos en los comportamientos de toma de decisiones que ya se han establecido en humanos, proponemos que nuestros modelos pueden ser eficaces para predecir cómo otros fármacos pueden mediar estos comportamientos en la clínica. . Por ejemplo, un subconjunto de pacientes con enfermedad de Parkinson (EP) que son tratados con terapias con agonistas de la dopamina demuestran una mayor prevalencia del comportamiento de juego (Weintraub et al., 2010) y un mayor descuento en la toma de decisiones basada en el retraso (Housden et al., 2010; Milenkova y otros, 2011; Voon et al., 2011; Leroi et al., 2013; Szamosi et al., 2013). Por lo tanto, nuestro laboratorio se propuso modelar la EP en ratas y estudiar los efectos del pramipexol, un agonista de la dopamina comúnmente asociado con las conductas de juego (Weintraub et al., 2010), en la toma de decisiones de costo / beneficio en la rata usando la tarea de descuento de probabilidad (toma de decisiones de riesgo / recompensa) (Rokosik y Napier, 2012). Para ello, las ratas se convirtieron en "similares a PD" mediante el lesionamiento selectivo de terminales dopaminérgicos dentro del estriado dorsolateral vía infusiones bilaterales de 6-OHDA, mientras que las ratas de control recibieron infusiones del vehículo 6-OHDA (Rokosik y Napier, 2012). Las neuronas en el cuerpo estriado dorsolateral de las ratas tratadas con 6-OHDA muestran una disminución en la tirosina hidroxilasa (Rokosik y Napier, 2012), un marcador de dopamina. Las ratas similares a la PD presentan alteraciones motoras similares a las de los humanos con EP en etapa temprana, que pueden revertirse de forma dependiente de la dosis con el tratamiento con pramipexol. La dosis de pramipexol que administramos para estudiar la toma de decisiones de riesgo / recompensa alivia los déficits motores y, por lo tanto, es terapéuticamente relevante (Rokosik y Napier, 2012). Si bien no encontramos diferencias en el comportamiento "arriesgado" de referencia entre las ratas de control y las ratas similares a la EP, el tratamiento crónico con pramipexol aumenta la selección de la LR riesgosa en ambos grupos de ratas cuando las probabilidades de parto eran pequeñas (Figuras 2A, B), que indica que el pramipexol induce la toma de decisiones de riesgo / recompensa subóptimas. Estos datos concuerdan con los estudios que han evaluado los efectos del pramipexol en humanos (Spengos et al., 2006; Pizzagalli et al., 2008; Riba et al., 2008). No obstante, inferimos la validez predictiva de nuestros modelos de roedores al indicar otros agentes farmacológicos que pueden mediar en la toma de decisiones de costo / beneficio en humanos.
También hemos probado la mirtazapina, un antidepresivo atípico, en la tarea de toma de decisiones basada en el esfuerzo. Las adicciones conductuales y el abuso de sustancias comparten muchas características que se superponen, incluida la toma de decisiones subóptima, y los nuevos estudios en humanos y animales no humanos ilustran que la mirtazapina es eficaz para reducir los comportamientos motivados por el abuso de drogas (por ejemplo, opiáceos y psicoestimulantes) incluso aquellos que están asociados con recaída durante los períodos de abstinencia (para una revisión, ver Graves et al., 2012). Los datos recopilados de nuestra tarea de toma de decisiones basada en el esfuerzo mediado por ICSS indican que la mirtazapina redujo efectivamente la preferencia por un esfuerzo alto / LR, cambiando a un esfuerzo bajo / SR, lo que sugiere que la cantidad de esfuerzo requerido para el LR ya no era "valiosa". ", o que el valor de recompensa de la LR se redujo (Personas et al., 2013). Estos resultados sugieren que puede ser de interés estudiar los efectos de la mirtazapina en la toma de decisiones subóptimas en jugadores con problemas en la clínica.
Conclusión
En resumen, hemos utilizado el ICSS como un reforzador positivo en varias tareas novedosas diseñadas para medir aspectos separados, pero que se superponen, de la toma de decisiones de costo / beneficio expuesta en el problema del juego. Estas medidas se pueden utilizar para explorar más a fondo la contribución de varios sustratos neuroanatómicos y sistemas de neurotransmisores en juegos de azar problemáticos. Las tareas mediadas por ICSS proporcionan una alternativa viable al refuerzo de alimentos en estos paradigmas operantes complejos. Creemos que la validez de estas tareas indica que pueden ayudar en la selección de medicamentos por su potencial para inducir trastornos del control de impulsos, como el problema del juego, y para ayudar a identificar los medicamentos que reducen estos trastornos.
Declaracion de conflicto de interes
El Dr. Napier ha recibido apoyo de investigación de los Institutos Nacionales de la Salud, la Fundación Michael J. Fox y el Centro Nacional para el Juego Responsable. El Dr. Napier ha recibido una compensación por lo siguiente: asesoría para un centro de educación para la salud sin fines de lucro y para despachos de abogados en temas relacionados con adicciones y trastornos del control de impulsos; hablar sobre adicciones en reuniones comunitarias municipales, escuelas secundarias públicas, organizaciones sin fines de lucro basadas en la comunidad y reuniones profesionales de tribunales de drogas; proporcionar revisiones de subvenciones para los Institutos Nacionales de Salud y otras agencias; y conferencias académicas y grandes rondas. La Dra. Napier es miembro de la Alianza de Illinois sobre problemas de juego, y brinda asesoramiento especializado sobre el desarrollo de medicamentos a la Fundación Cures Within Research. El Dr. Holtz, el Dr. Persons y la Sra. Tedford declaran que la investigación se llevó a cabo en ausencia de cualquier relación comercial o financiera que pudiera interpretarse como un posible conflicto de intereses.
AGRADECIMIENTOS
Este trabajo fue apoyado por el Centro Nacional para el Juego Responsable, la Fundación Michael J. Fox, la Fundación Daniel F. y Ada L. Rice, y NS074014 de USPHSG para T. Celeste Napier y DA033121 para Stephanie E. Tedford y T. Celeste Napier .
Referencias
Alessi, SM y Petry, NM (2003). La gravedad patológica del juego está asociada con la impulsividad en un procedimiento de descuento por demora. Behav. Procesos 64, 345–354. doi: 10.1016/S0376-6357(03)00150-5
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Bickel, WK, Jarmolowicz, DP, Mueller, ET, Koffarnus, MN y Gatchalian, KM (2012). Descuento excesivo de refuerzos retrasados como un proceso de trans-enfermedad que contribuye a la adicción y otras vulnerabilidades relacionadas con la enfermedad: evidencia emergente. Pharmacol Ther. 134, 287 – 297. doi: 10.1016 / j.pharmthera.2012.02.004
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Bizo, LA, Bogdanov, SV, y Killeen, PR (1998). La saciedad causa una disminución dentro de la sesión en la respuesta instrumental. J. Exp. Psychol. Anim Behav. Proceso. 24, 439 – 452. doi: 10.1037 / 0097-7403.24.4.439
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Buckholtz, JW, Treadway, MT, Cowan, RL, Woodward, ND, Li, R., Ansari, MS, y otros. (2010). Diferencias de la red dopaminérgica en la impulsividad humana. Ciencia: 329, 532. doi: 10.1126 / science.1185778
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Buelow, MT y Suhr, JA (2009). Construir la validez de la tarea de juego iowa. Neuropsicol. Rdo. 19, 102 – 114. doi: 10.1007 / s11065-009-9083-4
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Corbett, D., y Wise, RA (1980). Autoestimulación intracraneal en relación con los sistemas dopaminérgicos ascendentes del mesencéfalo: un estudio de mapeo de electrodos móviles. Brain Res.. 185, 1–15. doi: 10.1016/0006-8993(80)90666-6
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Crean, JP, de, WH y Richards, JB (2000). Recompensar el descuento como medida del comportamiento impulsivo en una población psiquiátrica ambulatoria. Exp. Clinica Psicofarmacol. 8, 155 – 162. doi: 10.1037 / 1064-1297.8.2.155
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Dai, Z., Harrow, SE, Song, X., Rucklidge, J. y Grace, R. (2013). Apuestas, retrasos y descuentos de probabilidad en adultos con y sin TDAH. J. Atten. Desorden. doi: 10.1177 / 1087054713496461. [Epub antes de imprimir].
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Dixon, MR, Marley, J., y Jacobs, EA (2003). Retraso en el descuento por apostadores patológicos. J. Appl. Behav. Anal. 36, 449 – 458. doi: 10.1901 / jaba.2003.36-449
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Douglas, VI, y Parry, PA (1994). Efectos de recompensa y no recompensa sobre la frustración y la atención en el trastorno por déficit de atención. J. Abnorm. Psicologia infantil. 22, 281 – 302. doi: 10.1007 / BF02168075
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Eubig, PA, Noe, TE, Floresco, SB, Sable, JJ y Schantz, SL (2014). Diferencias de sexo en respuesta a la anfetamina en ratas adultas de larga evolución que realizan una tarea de descuento por demora. Pharmacol. Biochem. Behav. 118, 1 – 9. doi: 10.1016 / j.pbb.2013.12.021
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Feja, M., y Koch, M. (2014). La inactivación de la corteza prefrontal medial ventral altera el control de los impulsos pero no afecta el retraso en el descuento en ratas. Behav. Brain Res. 264, 230 – 239. doi: 10.1016 / j.bbr.2014.02.013
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Graves, SM, Rafeyan, R., Watts, J. y Napier, TC (2012). La mirtazapina y los compuestos similares a la mirtazapina como posible farmacoterapia para los trastornos por abuso de sustancias: evidencia del banco y de la cama. Pharmacol. El r. 136, 343 – 353. doi: 10.1016 / j.pharmthera.2012.08.013
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Holt, DD, Green, L. y Myerson, J. (2003). ¿El descuento es impulsivo? Evidencia de descuentos temporales y probabilísticos en estudiantes universitarios de juego y no apostadores. Behav. Procesos 64, 355–367. doi: 10.1016/S0376-6357(03)00141-4
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Hosking, JG, Cocker, PJ y Winstanley, CA (2014). Contribuciones disociables de la corteza cingulada anterior y la amígdala basolateral en una tarea de toma de decisiones de costo / beneficio de roedores del esfuerzo cognitivo. Neuropsicofarmacología 39, 1558 – 1567. doi: 10.1038 / npp.2014.27
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Housden, CR, O'Sullivan, SS, Joyce, EM, Lees, AJ y Roiser, JP (2010). Aprendizaje de recompensa intacto pero descuento por retraso elevado en pacientes con enfermedad de Parkinson con conductas del espectro impulsivo-compulsivo. Neuropsicofarmacología 35, 2155 – 2164. doi: 10.1038 / npp.2010.84
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Jimura, K., Myerson, J., Hilgard, J., Braver, TS y Green, L. (2009). ¿Las personas son realmente más pacientes que otros animales? Evidencia del descuento humano de recompensas de líquidos reales. Psychon. Toro. Rdo. 16, 1071 – 1075. doi: 10.3758 / PBR.16.6.1071
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Johnson, PS, Madden, GJ, Brewer, AT, Pinkston, JW y Fowler, SC (2011). Efectos del pramipexol agudo en la preferencia por los horarios de refuerzo similares a los juegos de azar en ratas. Psicofarmacología (Berl) 231, 11–18. doi: 10.1007/s00213-010-2006-5
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Keesey, RE (1962). La relación entre la frecuencia, la intensidad y la duración del pulso y la tasa de respuesta para la estimulación intracraneal. J. comp. Fisiol. Psicol. 55, 671 – 678. doi: 10.1037 / h0049222
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Koffarnus, MN, Newman, AH, Grundt, P., Rice, KC, y Woods, JH (2011). Efectos de los compuestos dopaminérgicos selectivos en una tarea de descuento por demora. Behav. Pharmacol. 22, 300 – 311. doi: 10.1097 / FBP.0b013e3283473bcb
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Kraplin, A., Dshemuchadse, M., Behrendt, S., Scherbaum, S., Goschke, T. y Buhringer, G. (2014). Toma de decisiones disfuncional en el juego patológico: especificidad de patrón y el papel de la impulsividad. Psiquiatría Res.. 215, 675 – 682. doi: 10.1016 / j.psychres.2013.12.041
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Ledgerwood, DM, Alessi, SM, Phoenix, N. y Petry, NM (2009). Evaluación conductual de la impulsividad en jugadores patológicos con y sin historial de trastornos por uso de sustancias versus controles sanos. Drogas alcohol dependiente. 105, 89 – 96. doi: 10.1016 / j.drugalcdep.2009.06.011
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Leroi, I., Barraclough, M., McKie, S., Hinvest, N., Evans, J., Elliott, R. y McDonald, K. (2013). Influencias dopaminérgicas sobre la función ejecutiva y el comportamiento impulsivo en los trastornos del control de impulsos en la enfermedad de Parkinson. J. Neuropsychol. 7, 306 – 325. doi: 10.1111 / jnp.12026
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Madden, GJ, Johnson, PS, Brewer, AT, Pinkston, JW y Fowler, SC (2010). Efectos del pramipexol en la elección impulsiva en ratas wistar macho. Exp. Clinica Psicofarmacol. 18, 267 – 276. doi: 10.1037 / a0019244
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Madden, GJ, Petry, NM y Johnson, PS (2009). Los jugadores patológicos descuentan las recompensas probabilísticas menos abruptamente que los controles emparejados. Exp. Clinica Psicofarmacol. 17, 283 – 290. doi: 10.1037 / a0016806
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
McCown, TJ, Napier, TC y Breese, GR (1986). Efectos de la implantación crónica de electrodos en las neuronas dopaminérgicas. in vivo. Pharmacol. Biochem. Behav 25, 63–69. doi: 10.1016/0091-3057(86)90231-5
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Mejia-Toiber, J., Boutros, N., Markou, A. y Semenova, S. (2014). Elección impulsiva y comportamiento similar a la ansiedad en ratas adultas expuestas al etanol intermitente crónico durante la adolescencia y la edad adulta. Behav. Brain Res. 266, 19 – 28. doi: 10.1016 / j.bbr.2014.02.019
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Michalczuk, R., Bowden-Jones, H., Verdejo-Garcia, A. y Clark, L. (2011). Impulsividad y distorsiones cognitivas en jugadores patológicos que asisten a la Clínica Nacional de Apuestas al Problema del Reino Unido: un informe preliminar. Psychol. Medicina. 41, 2625 – 2635. doi: 10.1017 / S003329171100095X
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Miedl, SF, Peters, J. y Buchel, C. (2012). Representaciones de recompensas neuronales alteradas en jugadores patológicos reveladas por demoras y descuentos de probabilidad. Arco. Gen. Psiquiatría 69, 177 – 186. doi: 10.1001 / archgenpsychiatry.2011.1552
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Milenkova, M., Mohammadi, B., Kollewe, K., Schrader, C., Fellbrich, A., Wittfoth, M. y col. (2011). Elección intertemporal en la enfermedad de Parkinson. Mov. Desorden. 26, 2004 – 2010. doi: 10.1002 / mds.23756
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Minamimoto, T., La Camera, G., y Richmond, BJ (2009). Midiendo y modelando la interacción entre el tamaño de la recompensa, el retraso en la recompensa y el nivel de saciedad en la motivación de los monos. J. neurofisiol. 101, 437 – 447. doi: 10.1152 / jn.90959.2008
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Olds, J., y Milner, P. (1954). Refuerzo positivo producido por la estimulación eléctrica del área septal y otras regiones del cerebro de rata. J. comp. Fisiol. Psicol. 47, 419 – 427. doi: 10.1037 / h0058775
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Papini, MR, y Dudley, RT (1997). Consecuencias de las sorprendentes omisiones de recompensa. Rev. Gen. Psychol. 1, 175 – 197. doi: 10.1037 / 1089-2680.1.2.175
Personas, AL, Tedford, SE y Napier, TC (2013). “Evaluaciones preclínicas de compuestos serotoninérgicos de función mixta para el tratamiento de trastornos del juego. programa no. 635.12, ”en Planificador de reuniones de neurociencia (San Diego, CA: Society for Neuroscience).
Petry, NM (2011). El descuento de las recompensas probabilísticas se asocia con la abstinencia del juego en jugadores patológicos que buscan tratamiento. J. Abnorm. Psicol. 121, 151 – 159. doi: 10.1037 / a0024782
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Pirch, JH, Napier, TC y Corbus, MJ (1981). La estimulación cerebral como un indicio de potenciales relacionados con eventos en la corteza de rata: efectos de anfetamina. En t. J. Neurosci. 15, 217 – 222.
Pizzagalli, DA, Evins, AE, Schetter, EC, Frank, MJ, Pajtas, PE, Santesso, DL, et al. (2008). La dosis única de un agonista de la dopamina altera el aprendizaje de refuerzo en humanos: evidencia de comportamiento de una medida basada en laboratorio de respuesta de recompensa. Psicofarmacología (Berl) 196, 221–232. doi: 10.1007/s00213-007-0957-y
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Rachlin, H., Raineri, A. y Cross, D. (1991). Probabilidad subjetiva y retraso. J. Exp. Anal. Behav. 55, 233 – 244. doi: 10.1901 / jeab.1991.55-233
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Rasmussen, EB, Lawyer, SR, y Reilly, W. (2010). El porcentaje de grasa corporal está relacionado con el retraso y el descuento de probabilidad para los alimentos en humanos. Behav. Procesos 83, 23 – 30. doi: 10.1016 / j.beproc.2009.09.001
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Reynolds, B., Richards, JB, Horn, K. y Karraker, K. (2004). Retrasar el descuento y el descuento de probabilidad en relación con el estado de fumar cigarrillos en adultos. Behav. Procesos 65, 35–42. doi: 10.1016/S0376-6357(03)00109-8
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Riba, J., Kramer, UM, Heldmann, M., Richter, S. y Munte, TF (2008). El agonista de la dopamina aumenta el riesgo pero embota la actividad cerebral relacionada con la recompensa PLASOS UNO 3: e2479. doi: 10.1371 / journal.pone.0002479
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Rokosik, SL, y Napier, TC (2011). La autoestimulación intracraneal como refuerzo positivo para estudiar la impulsividad en un paradigma de descuento de probabilidad. J. Neurosci. Métodos 198, 260 – 269. doi: 10.1016 / j.jneumeth.2011.04.025
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Rokosik, SL y Napier, TC (2012). Incremento del descuento probabilístico inducido por pramipexol: comparación entre un modelo de roedor de la enfermedad de Parkinson y los controles. Neuropsicofarmacología 37, 1397 – 1408. doi: 10.1038 / npp.2011.325
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Roll, JM, McSweeney, FK, Johnson, KS y Weatherly, JN (1995). La saciedad contribuye poco a las disminuciones dentro de la sesión en la respuesta. Aprender. Motiv. 26, 323–341. doi: 10.1016/0023-9690(95)90018-7
Schuck-Paim, C., Pompilio, L., y Kacelnik, A. (2004). Las decisiones dependientes del estado causan aparentes violaciones de la racionalidad en la elección de los animales. PLASOS Biol. 2: e402. doi: 10.1371 / journal.pbio.0020402
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Spengos, K., Grips, E., Karachalios, G., Tsivgoulis, G., y Papadimitriou, G. (2006). Juego patológico reversible en tratamiento con pramipexol. Nervenarzt 77, 958–960. doi: 10.1007/s00115-006-2128-5
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
St Onge, JR y Floresco, SB (2009). Modulación dopaminérgica de la toma de decisiones basada en el riesgo. Neuropsicofarmacología 34, 681 – 697. doi: 10.1038 / npp.2008.121
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Tapón, CM, y Floresco, SB (2011). Contribuciones del núcleo accumbens y sus subregiones a diferentes aspectos de la toma de decisiones basada en el riesgo. Cogn. Afectar. Behav. Neurosci. 11, 97 – 112. doi: 10.3758 / s13415-010-0015-9
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Szamosi, A., Nagy, H. y Keri, S. (2013). Retrasar el descuento de la recompensa y el volumen del núcleo caudado en individuos con duplicación del gen alfa-sinucleína antes y después del desarrollo de la enfermedad de Parkinson. Neurodegenerador. Dis. 11, 72 – 78. doi: 10.1159 / 000341997
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Tedford, SE, Persons, AL, Chen, L., Rokosik, SL y Napier, TC (2012). “La autoestimulación intracraneal como refuerzo positivo para estudiar la impulsividad en un paradigma de descuento retrasado en ratas. programa No. 926.03, ”en Planificador de reuniones de neurociencia (Nueva Orleans, LA: Society for Neuroscience).
Tehovnik, EJ y Sommer, MA (1997). Diseminación efectiva y curso de tiempo de la inactivación neural causada por la inyección de lidocaína en la corteza cerebral de los monos. J. Neurosci. Métodos 74, 17–26. doi: 10.1016/S0165-0270(97)02229-2
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Treadway, MT, Buckholtz, JW, Schwartzman, AN, Lambert, WE y Zald, DH (2009). ¿Vale la pena el 'EEfRT'? El gasto de esfuerzo para recompensar la tarea como medida objetiva de motivación y anhedonia. PLASOS UNO 4: e6598. doi: 10.1371 / journal.pone.0006598
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Trowill, JA, Panksepp, J., y Gandelman, R. (1969). Un modelo de incentivo para recompensar la estimulación cerebral. Psychol. Rdo. 76, 264 – 281. doi: 10.1037 / h0027295
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Voon, V., Sohr, M., Lang, AE, Potenza, MN, Siderowf, AD, Whetteckey, J., et al. (2011). Trastornos del control de impulsos en la enfermedad de Parkinson: un estudio multicéntrico de casos y controles. Ana. Neurol. 69, 986 – 996. doi: 10.1002 / ana.22356
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Wardle, MC, Treadway, MT, Mayo, LM, Zald, DH y de, WH (2011). Amplificación del esfuerzo: efectos de la d-anfetamina en la toma de decisiones basada en el esfuerzo humano. J. Neurosci. 31, 16597 – 16602. doi: 10.1523 / JNEUROSCI.4387-11.2011
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Weintraub, D., Koester, J., Potenza, MN, Siderowf, AD, Stacy, M., Voon, V., et al. (2010). Trastornos de control de impulso en la enfermedad de Parkinson: un estudio transversal de pacientes con 3090. Arco. Neurol. 67, 589 – 595. doi: 10.1001 / archneurol.2010.65
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Sabio, RA (1996). Las drogas adictivas y la estimulación cerebral recompensa. Annu. Rev. Neurosci. 19, 319 – 340. doi: 10.1146 / annurev.neuro.19.1.319
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Sabio, RA (2005). Sustratos Forebrain de recompensa y motivación. J. comp. Neurol. 493, 115 – 121. doi: 10.1002 / cne.20689
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Wise, RA, Bauco, P., Carlezon, WA Jr., y Trojniar, W. (1992). Mecanismos de autoestimulación y recompensa de drogas. Ana. NY Acad. Ciencia. 654, 192 – 198. doi: 10.1111 / j.1749-6632.1992.tb25967.x
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Wise, RA y Bozarth, MA (1981). Sustratos cerebrales para refuerzo y autoadministración de fármacos. Prog. Neuropsicofarmacol. 5, 467 – 474. doi: 10.1016 / 0364-7722 (81) 90028-X
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Wise, RA, y Rompre, PP (1989). Dopamina cerebral y recompensa. Annu. Rev. Psychol. 40, 191 – 225. doi: 10.1146 / annurev.psych.40.1.191
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Zeeb, FD, Robbins, TW y Winstanley, CA (2009). La modulación serotoninérgica y dopaminérgica de la conducta de juego se evalúa utilizando una tarea de juego de ratas novedosa. Neuropsicofarmacología 34, 2329 – 2343. doi: 10.1038 / npp.2009.62
Resumen Pubmed | PubMed texto completo | Texto completo de CrossRef
Palabras clave: toma de decisiones de costo / beneficio, descuentos, toma de decisiones basada en el esfuerzo, juego, autoestimulación intracraneal
Cita: Tedford SE, Holtz NA, Persons AL y Napier TC (2014) Un nuevo enfoque para evaluar el comportamiento similar al juego en ratas de laboratorio: el uso de la autoestimulación intracraneal como refuerzo positivo. Frente. Behav. Neurosci. 8: 215. doi: 10.3389 / fnbeh.2014.00215
Recibido: 06 marzo 2014; Aceptado: 27 May 2014;
Publicado en línea: 11 junio 2014.
Editado por:
Patrick Anselme, Universidad de Lieja, Bélgica
Revisado por:
Christelle Baunez, Centro Nacional de la Investigación Científica, Francia
Yueqiang Xue, Centro de Ciencias de la Salud de la Universidad de Tennessee, EE. UU.