YBOP COMMENTS. Coolidge- ի էֆեկտը կանգնած է ինտերնետային պոռնոյի ուժի մեջ: Coolidge- ի էֆեկտը երևույթ է, որը նկատվում է կաթնասունների տեսակների մեջ, որի արդյունքում արական սեռի ներկայացուցիչները (և ավելի քիչ կանայք) ցուցաբերում են նոր սեռական հետաքրքրություն, եթե ներկայացվում են նոր ընկալունակ սեռական գործընկերների հետ, նույնիսկ նախնական, բայց առայժմ մատչելի սեռական գործընկերներից սեռը մերժելուց հետո: Սեռական նորությունը գերակշռում է այս սովորույթը `ավելի բարձր դոպամինի հետևանքով առաջացած նոր հուզմունքով: Նորույթի անընդհատ հոսք է այն, ինչը ինտերնետային պոռնոդը այդքան է տարբերում անցյալի պոռնոններից:
- Դենիս Ֆ. Ֆիորինո,
- Արիանա Քուրի, և
- Էնթոնի Գ. Ֆիլիպսը
- Նյարդաբանության ամսագիր, 15 հունիսի 1997 թ., 17 (12): 4849-4855;
Վերացական
The Coolidge- ի ազդեցությունը նկարագրում է սեռական վարքագծի վերականգնում «սեռական տատանված» կենդանու մեջ `վեպի ընդունող կողակցին: Հաշվի առնելով մեսոլիմբիկ դոպամինի (DA) համակարգի դերը դրդապատճառ վարքի սկզբունքի եւ պահպանման գործում, microdialysis օգտագործվել է Nucleus accumbens (DA) DA փոխանցումը կուտակման ընթացքում, սեռական հակում եւ սեռական վարքի վերականգնում: Նախկին զեկույցների համաձայն, էկրանին ետեւում պատահական կնոջ ներկայացումը եւ կուտակումը կապված էին NAC DA ջրային հոսքի զգալի աճի հետ: NAC DA- ի կոնցենտրացիաների վերադարձը բազային արժեքներին համընկնում էր սեռական հաստության ժամանակահատվածի հետ, չնայած DA մետաբոլիտների, dihydroxyphenylacetic acid- ի եւ homovanillic թթվի կոնցենտրացիաները մնաց բարձր: Էկրանի ետեւում գտնվող վեպի ընդունող կանանց շնորհանդեսը հանգեցրեց NAC DA- ի փոքր աճին, որը զգալիորեն ավելացավ վեպի կանանց հետ նորացված կուտակման ժամանակ: Ներկայացված տվյալները ցույց են տալիս, որ վեպի ընդունող կանանց խթանման հատկությունները կարող են նպաստել NAC DA փոխանցումը սեռական ճանապարհով տառապած տղամարդկանց առնետում, եւ դա, իր հերթին, կարող է կապված լինել սեռական վարքի վերականգնման հետ:
ներածություն
Մի արծաթը, որը հագեցած է հագեցածության համար, կարող է հանգեցնել միմյանց նորից, եթե նախնական կինը փոխարինվի վեպի ընդունող կանանցով: Սա եկել է հայտնի որպես Քուլիչի ազդեցությունը եւ դիտվել է մի շարք հումքային տեսակների (Վիլսոն եւ այլն, 1963): Ընդհանուր գործոններ, ինչպիսիք են հոգնածության կամ շարժիչային դեպրեսիան, բավարար չեն սեռական սթրեսի ակնհայտ վիճակին բացատրելու համար, քանի որ վեպի կանանց խթանումը դեռեւս կարող է առաջացնել կուտակումը: Սեռական հագեցվածությունը կարող է նաեւ «զերծ մնալ» դեղագործորեն, նշանակալից աստիճանով, տարբեր թմրանյութերի կիրառմամբ, որոնք կարող են գործել տարբեր նյարդոտիրական համակարգերում: Այս դեղերը ներառում են yohimbine, 8-OH-DPAT (Rodriguez-Manzo եւ Fernandez-Guasti, 1994, 1995a), նալաքսոն (Pfaus եւ Gorzalka, 1987; Rodriguez-Manzo եւ Fernandez-Guasti, 1995a,b) եւ apomorphine (Mas եւ այլն, 1995c): Չնայած այս դեղերի ծայրամասային գործողությունը չի կարող բացառվել (օրինակ, ծայրահեղ ֆունկցիայի վրա ադրեներգիկ ազդեցություն), սեռավարժության հիմքում ընկած կենտրոնական մեխանիզմների ազդեցությունները առաջարկվել են ընտրովի կենտրոնական noradrenergic lesion experiments (Rodriguez-Manzo եւ Fernandez-Guasti, 1995a) եւ միկրեդիալիզի փորձեր, որոնք վերահսկում են դոպամիներգիկ նյութափոխանակությունը միջռեգիոնալ պրեոպիստիկ տարածքումMas եւ այլն, 1995a,b).
Հաշվի առնելով, որ կենտրոնական մեխանիզմները կարող են միջնորդել Coolidge- ի ազդեցությանը բնորոշ սեռական վարքագծի վերականգնմանը, հավանական թեկնածուն այն է, mesolimbic dopamine (DA) համակարգը, որը նախագծում է ventral tegmental տարածությունից մինչեւ NAC: Mesolimbic DA- ն կարծես հանդես է գալիս որպես առաջնային մոդուլատոր բարդ ինտեգրացիոն գործընթացներում, որոնք ներառում են շրջակա միջավայրի խթանման գնահատումը, ինչպես օրինակ, սեռական ընդունող կանանցից եւ նպատակային ուղղված վարքագծի կազմակերպումը, ներառյալ կուտակումը (Fibiger եւ Phillips, 1986; Բլեքբեռնը եւ այլք, 1992; Ֆիլիպսը եւ այլք, 1992; LeMoal, 1995; Salamone, 1996).
Չնայած միջին խոռոչի DA neurons արձագանքում առաջնային պարգեւների եւ cues կանխատեսելի պարգեւատրման, վեպի կամ անկանխատեսելի բնապահպանական խթանները դրդում neuronal ակտիվացման առավել robust over կրկնվող դասընթացների (Ֆաբրեն եւ այլք, 1983; Շուլց, 1992; Mirenowicz եւ Schultz, 1994): Առկա է բազմաթիվ ապացույցներ, որոնք աջակցում են mesolimbic DA- ի կարեւոր դերակատարում դերը սեռական սեռական վարքի սկզբնավորման եւ պահպանման մեջ (Pfaus եւ Everitt, 1995), եւ մի շարք միկրոալիզի հետազոտությունների զեկույցը աճում է NAC DA անջատում արական սեռական վարքի ախորժակի եւ սպառողական փուլերումPfaus et al., 1990; Հավիմ եւ այլն, 1990; Damsma et al., 1992; Wenkstern et al., 1993; Fumero et al., 1994; Mas եւ այլն, 1995b): Այնուամենայնիվ, համեմատաբար քիչ տվյալներ կան սեռավարակների նյարդաքիմիական փոխկապակցվածության եւ սեռական վարքի վերականգնման վերաբերյալ: Դիմումը ի Վիվո միկրոդալիզը դիտելու համար mesolimbic DA- ի նյարդոտման շնորհիվ Coolidge ազդեցության ընթացքում հնարավորություն է տալիս ուսումնասիրել NAC DA- ի դերը կուտակման, սեռական հագեցման եւ կուտակման վերականգնման մեջ:
(1), թե արդյոք սեռական հագեցման սկիզբը ուղեկցվում է NAC- ում արտազատվող DA կոնցենտրացիաների վերադարձով, նախնական գնահատման արժեքների կամ ստորեւ, եւ (2), թե արդյոք վերարտադրողական վարքագծի վերականգնում է " սեռական առումով սնուցված «արական առնետը վեպի ընդունող կանանց հետ, կապված է NAC DA արտահոսքի բարձրացման հետ:
ՆՅՈՒԹԵՐ ԵՒ ՄԵԹՈԴՆԵՐ
Առարկաներ: Կանադական Sprague Dawley առնետները, որոնք ստացել են կենդանիների խնամքի կենտրոնից (բրիտանական Կոլումբիայի համալսարանում) եւ կանանց Long-Evans առնետները, որոնք ստացվել են Չարլզ գետի Կանադայից (Սեվ Կոնստանդ, Քվեբեկ, Կանադա), տեղադրվել են մետաղական ցանցի խցերում (18 × 25 × 65 սմ, հինգը մեկ վանդակում) առանձին գաղութներում: Գաղութի սենյակները պահպանվեցին ~ 20 ° C ջերմաստիճանի մեջ, հակառակ 12 ժամը թեթեւ / մութ ցիկլով: Rats- ը անսահմանափակ քանակությամբ սնունդ էր (Purina Rat Chow) եւ ջուր:
Վիրաբուժություն եւ վարքի փորձարկում, ուղեղի միկրոդալիզի առաջ:Տղամարդիկ առնետները գլխուղեղային գազային անեստեզիայում (Fluotane, Ayerst Laboratories) առնվազն 4 շաբաթ առաջ փորձարկվել էին երկակիորեն ovariectomized երկկողմանի. Կանխարգելման սեռի սեռական ընդունումը դրսեւորվել է նախքան յուրաքանչյուր փորձարկման նիստի նախապատրաստական փուլում, համապատասխանաբար, estradiol benzoate (10 μg) եւ progesterone (500 μg), 48 եւ 4 ժամ պարունակող ներարկում: Արական առնետները ցուցադրվել են սեռական վարքի համար, երկու անգամ `4 d- ից, Plexiglas պալատներում (35 × 35 × 40 սմ)` մետաղական ցանցի հատակի հետ: Միայն արական առնետները, որոնք հասել են կատարողականի չափորոշիչին, որը ներառում էր միջամտության 5- ի ներսում միջամտության 15- ի ընթացքում իգական քաղցկեղի եւ զառամինգի ներթափանցման ընթացքում, երկու ստուգման թեստերի ընթացքում ներարկվել է միկրոալիզի հետազոտման հետախուզման ուղեցույցի ուղեծրով:
Արական առնետներ (n = 5), ստերէթաքսիկ վիրահատությունից առաջ անզգայացվում էին ketamine hydrochloride (100 մգ / կգ, ip) եւ xylazine (10 մգ / կգ, ip) հետ: Microdialysis- ի հետախուզական ուղեկցող ուղեղի (19 gauge) երկաթուղային ուղեծրերը (1.7 gauge) երկաթուղային ճանապարհով զննում էին NAC- ի (կոորդինատները bregma- ից `նախորդ, + 1.1 մմ, medial, ± 1.0 մմ, ventral, 15 մմ, հարթ գանգ) եւ ապահովված է գանգի ատամնային ակրիլով եւ ոսկերիչների պտուտակներ: Երկկողմանի ուղեղային կեղեւային իմպլանտները օգտագործվել են հաջող microdialysis փորձի հնարավորության առավելագույնի հասցնելու համար: Բարեբախտաբար, ներկա փորձի մեջ յուրաքանչյուր առնետի համար անհրաժեշտ էր միայն մեկ կույտ: Արական առնետները առանձին էին տեղակայված խոշոր պլաստիկ խցերում, որոնք փորձարկված մնացորդների համար էին: Վիրահատությունից հետո մեկ շաբաթ անց առնետները փորձարկվել են սեռական վարքի համար: Դասընթացի այս մասում թեստավորման պալատը հագեցած էր լոգարիթմական Plexiglas էկրանով, որը բաժանեց պալատը խոշոր եւ փոքր բեկորներ: Տղամարդիկ առնետները մտցվել են մեծ խցիկի մեջ եւ ավելի ուշ 15 րոպե հետո, դրսից դրված է կին: 30 րոպե նախապատրաստական ժամանակաշրջանից հետո էկրանը հանվել է, եւ առնետներին թույլատրվել է քննել 4 րոպե: Երեք դասընթացներ անցկացվեցին, յուրաքանչյուրը `XNUMX դ ամսական: Բոլոր առնետները հասել են կատարողականի յուրաքանչյուր չափանիշի:
Coolidge ազդեցության փորձը: Rats- ը միակողմանիորեն ներարկվում էր միկրոալիզի հետազոտմամբ 12-18 ժամով Coolidge ազդեցության փորձի առաջ եւ տեղադրվեց թեստավորման պալատի խոշոր բաժնում `սննդի եւ ջրի ազատ հասանելիությամբ: Փորձի առավոտյան միկրոդալիզի նմուշները հավաքվեցին յուրաքանչյուր 15 րոպե: Փորձը բաղկացած էր հետեւյալ յոթ հաջորդական փուլերից `(1) բազային (առնվազն 60 րոպե); (2) կին 1 էկրանին ետեւում (15 րոպե); (3) կուտակումը 1- ի հետ, մինչեւ 30 րոպե անցնել առանց լեռան վրա: (4) կրկնօրինակը, 1- ի կրկնակի ետեւում (15 րոպե); (5) 1- ի համար 15- ի մուտքի հնարավորություն 3 րոպե ժամանակահատվածում, եթե չկա մոնտաժ (եթե տեղադրվել է, այս փուլը դիտվել է որպես փուլ 6); (2) էկրանին հետեւում կին 15- ի ներդրում (7 րոպե); 2) քնկոտություն 60- ի կանանց համար, XNUMX րոպե:
Վարքագիծը նկարահանվել է ցածր լուսավորության ներքո, օգտագործելով JVC տեսագրման համակարգ եւ դիտարկվում է տեսախցիկում, որը գտնվում է փորձարկման սենյակից դուրս: Սեռական վարքի ստանդարտ միջոցները արձանագրվել են համակարգչով եւ համապատասխան ծրագրային ապահովմամբ (Holmes et al., 1987).
Միկրոալիզի փորձարկումից հետո կենդանիներին տրվել է քլորիդային հիդրոդի գերբարձր չափաքանակ եւ ներծարծվել շաքարային եւ ձեւալինով (4%): Ուղեղը կտրտված եւ սառեցված է, եւ հետագայում, կորոնային հատվածները լցվել են cresyl violet- ի համար, որոշելու համար միկրոալիքային նմուշների տեղադրումը: Մշտական վարակների եւ նեյրո-քիմիական անալիզների համար օգտագործվել են միայն առնետները, որոնք ներգրավված են NAC- ի շրջանակներում:
Microdialysis եւ HPLC- էլեկտրաքիմիական հայտնաբերում: Միկրոդալիզային զոնդերը համակենտրոն էին դիզայնով կիսաթափանցելի խոռոչի մանրաթելային մեմբրանի հետ (2 մմ հյուսվածքի, 340 μմ արտաքին տրամագծով, 65000 մոլեկուլային քաշի կտրվածքով, Filtral 12, Hospal) հեռավոր վերջում: Սնոտները պարունակվում էին 1.0 μl / րոպեում փոփոխված Ringer- ի լուծույթով (0.01 մ նատրիումի ֆոսֆատ բուֆեր, pH 7.4, 1.3 mmCaCl2, 3.0 մմ KCl, 1.0 mmMgCl2, 147 մմ NaCl), օգտագործելով gastight ներարկիչ (Hamilton, Reno, NV) եւ ներարկիչ պոմպ (մոդել 22, Harvard Apparatus, South Natick, MA): Միկրոալիզի հետախուզման ուղեցույցի մանյակ օգտագործվել է միկրոալիզիզի հետազոտման համար ուղեցույցի ձագաթղթի մեջ: Փորձարկման պալատի վերին մասում տեղադրված հեղուկի շարժիչով (Instech 375s) կցված պողպատե կծիկ օգտագործվել է փորձարկման խողովակաշարի (Fiorino et al., 1993).
Միկրոդալիզացված անալիզները, որոնք պարունակում են DA եւ դրա մեթաբոլիտները dihyroxyphenylacetic acid (DOPAC) եւ homovanillic թթու (HVA), առանձնացված են հակադարձ փուլային քրոմատագրությամբ (Ultrasphere սյունակ, Beckman, Fullerton, CA, ODS 5 μm, 15 սմ, 4.6 մմ, ներքին տրամագիծը ) օգտագործելով 0.083մ նատրիումի ացետատի բուֆեր, pH 3.5 (5% մեթանոլ): Analyte- ի կոնցենտրացիաները քանակականացված են էլեկտրաքիմիական (EC) հայտնաբերման միջոցով: The ապարատը բաղկացած էր Bio-Rad (Richmond, CA) պոմպից, Valco Instruments (Houston, TX) EC10W երկկողմանի իննեկտոր, ESA (Bedford, MA) Coulochem II EC- ի դետեկտոր եւ երկակի ալիքի ձայնագրիչ (Kipp եւ Զոնեն, Բոեմիա, NY): Էլեկտրաքիմիական դետեկտորների պարամետրերը հետեւյալն են `էլեկտրոդ 1, + 450 մՎ; էլեկտրոդ 2, -300 մՎ; եւ պահակային բջիջ, 450 մՎ: Տիպային զննումները վերադարձվել են vitro եւ սենյակային ջերմաստիճանում, 22% DA- ի համար, 18% DOPAC- ի համար, իսկ 18% `HVA- ի համար:
ԱՐԴՅՈՒՆՔՆԵՐ
Վարքագիծ
Քուլիջի ազդեցության փորձից վարվելակերպը ներկայացված է աղյուսակում 1. Լեռը, ներկառուցվածությունը եւ զսպելը, ինչպես նաեւ նախնական զսպումը հետո հետաձգող միջակայքը նույնն էին նախորդ ուսումնական նստաշրջանի ընթացքում (տվյալները չեն ցուցադրվում): Սա ցույց է տալիս, որ միկրոդալիզիզ կարգը չի փոխում նորմալ սեռական վարքագիծը: Սեքսուալ սթրեսի զարգացումը, որը նշվում էր չափման չափանիշներից առաջ, ejaculations- ների միջին թիվը (7.8 ± 0.5), յուրաքանչյուր ejaculation- ի նախորդող միջամտությունների առաջադեմ նվազում եւ հետվայացնող միջակայքի առաջանցիկ աճ (տվյալների ցուցադրված չէ) , նման էր նախորդ ուսումնասիրություններին (Լողափ եւ Հորդանան, 1956; Fowler եւ Whalen, 1961; Fisher, 1962; Bermant եւ այլն, 1966; Rodriguez-Manzo եւ Fernandez-Guasti, 1994; Mas եւ այլն, 1995d): Անհատական փոփոխականությունը նկատվել է իգական 1- ով ձեռք բերված ejaculations- ի քանակի, կին 1- ի հետ կնքվող ժամանակի եւ սննդի չափորոշիչին հասնելու համար պահանջվող կանանց 1- ի ներկայացումների քանակը (Աղյուսակ 1, ներքեւում): Որոշ առնետներ պահանջում էին բազմաթիվ 1 կինոնկարներ, մինչեւ 5 փուլը ավարտվերn = 3): Էկրանի ետեւում կին 1- ի տեղադրումը եւ բաժանման հեռացումը կարող էին ծառայել որպես առաջնային ախորժակ, որը հանգեցնում է կուտակման: Պետք է նաեւ նշել, որ 30 min- ի սառեցման չափանիշը առանց լեռը, չնայած նախկինում օգտագործված էր (Beach and Jordan, 1965; Mas եւ այլն, 1995b), կամայական է եւ չի երաշխավորում, որ առնետը չէր կարողանում ավելի շատ ժամանակ տրամադրել: Այնուամենայնիվ, ձգձգումները կամ հեռացումը եւ փոխարինման ընթացակարգերը հավաստիորեն չեն հանգեցրել 1- ի (օրինակ 4- ի եւ 5- ի) կանանց հետ նորացված կուտակման ժամանակ:

Behavior during the Coolidge ազդեցության փորձի
Բոլոր առնետները ցուցադրեցին Coolidge ազդեցությունը: Կին 2- ի ետեւում տեղադրելու հետ կապված գործունեությունը եւ, մասնավորապես, բաժանման հեռացումը կարող է նպաստել այս արդյունքին, սակայն, կրկին, այս իրադարձությունները ոչ թե իրենց համար էին, այլ փորձի ժամանակաշրջանները վերականգնելու համար: Օգտագործվել են կանանց 1- ի եւ կանանց 2- ի սեռական վարքի միջոցների համեմատություններ t թեստեր, Bonferroni ուղղում: Չնայած այն հանգամանքին, որ 2- ի եւ իգական սեռի ներկայացուցիչների պատասխանը, 1- ի պատասխանն էր, ի տարբերություն սեռի 2- ի առաջին կոճակակիրը, ընդհանուր առմամբ, 0.6- ով սեռական վարքագիծը ավելի քիչ էր, քանի որ նշված է զգալիորեն ավելի քիչ ejaculations (նկատի առնելով, 4.2 vs XNUMX; F = 49.86;p <0.01) և ներխուժումները (միջին = 11.2 ընդդեմ 37.0;F = 20.17; p <0.05) առաջին ժամվա ընթացքում: 1 և 2 իգական սեռի ներկայացուցիչների առաջին ժամին մոնտաժների քանակն էապես տարբեր չէր:
Կարեւոր է նշել, որ փորձարարության սառեցման մասում օգտագործված կանայք (այսինքն, 1- ի կին) դեռեւս ցուցադրվել են ուժեղ քերականություն (այսինքն `ցնցող եւ ճկուն) եւ ընկալունակ (այսինքն` լորդոզ) վարքագիծ `տղամարդկանց հետ շփման ամբողջ տեւողության համար .
Նյարդաքիմիա
DA, 3.0 ± 0.7, DA, 619.1 ± 77.7, DA- ի բազային նանոմոլային կոնցենտրացիաները եւ դրանց միկրոշրջանների մեջ գտնվող մետաբոլիտները, ներկայացված որպես միջին ± SEM, DOPAC, 234.2 ± 49.0; եւ HVA, XNUMX ± XNUMX (չբացահայտված հետաքննության համար,n = 5): Այս արժեքները ներկայացրեցին 100% ելակետային միավորներ:
Նեյրոկիմիական անալիզների համար օգտագործվել են փորձարարության յուրաքանչյուր փուլ եւ համապատասխան ընդհանուր առնետների համար պատահականորեն սահմանված տվյալների կետերը: Դրանք ներառում էին հետեւյալը. (1) յոթ նմուշներ, 1- ի (2) առաջին ներկայացումից հետո (1) չորս նմուշներ, որոնք ուղեկցում են կանանց 3- ի քոփուլինգի վարքագծի բացակայության եւ 2- ի ներկայացումից հետո (XNUMX) հինգ նմուշ: Նկար 1 նկարագրում է DA- ի կոնցենտրացիաների փոփոխությունները (գծի գրաֆիկը, միջին) եւ DA metabolites (գծային գրաֆիկ, վերեւ) զուգահեռ, պատվանդան) Քուլիչի ազդեցության փորձարկման ժամանակ:
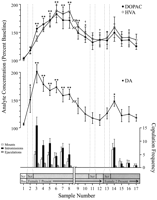
Դիտել ավելի մեծ տարբերակ:
Նկ. 1.
Nucleus accumbens neurochemical correlates սեռական վարքի ընթացքում Coolidge ազդեցության. Առաջին ութ նմուշները ներկայացնում են ժամանակագրական տվյալների շարունակական տվյալների կետերը 1- ից մինչեւ 3 փուլեր: Sample 1- ը չորրորդ եւ վերջին precopulation բազային նմուշները (bas): 2- ի նմուշը ներկայացնում է կին 1- ի ետեւում էկրան (Scr): 15 րոպեից հետո էկրանը հանվել է, եւ առնետներին թույլատրվել է կուտակել (նմուշներ 3-8): Theկոտրել մասին x-խիսը համապատասխանում է նախնական կնոջ հետ երկար ժամանակահատվածների համար նախատեսված երեք առնետներից ստացված տվյալների բացառմանը: Վերջին ինը նմուշները նույնպես ժամանակագրական բնույթ էին կրում: 9 եւ 10 նմուշները համապատասխանում են փուլ 3- ի (այսինքն 30 րոպե առանց լեռների) հագեցման ժամանակաշրջանին: Իգական 1- ը վերազինվել է էկրանին ետեւում (օրինակ 11) եւ 15 րոպե հետո, էկրանը հեռացվել է (օրինակ 12): 15- ից հետո, կախվածությունը չհամապատասխանեցրեց, կինն 2- ն դրվեց էկրանի ետեւում (ընտրանք 13): 14-17 նմուշները համապատասխանում են 2- ի կանանց հետ: Յուրաքանչյուր 15 րոպե միկրոալիզի նմուշի հետ կապված կախվածություն, ինտրոմիա կամ սերմնահեղուկի քանակը ցույց է տրված ներքեւի գագաթը. Նյարդաքիմիական տվյալները արտահայտվում են բազային կոնցենտրացիաների տոկոսով: NAC DA- ի փոփոխությունները (փակ հրապարակներ), DOPAC (փակ շրջանակներ) եւ HVA (բաց շրջանակներ) արտահոսքը ներկայացվում է որպես գրաֆիկները. Հաջորդ համեմատությունները կատարվեցին. 1- ի բազային նմուշն ընդդեմ նմուշների 2-10; նոր բազային նմուշ 10 versus նմուշները 11 եւ 12; նոր բազային նմուշ 12 versus նմուշներ 13-17 (*p <0.05; ** p <0.01): Անկախ t թեստեր են կատարվել բազային արժեքների միջեւ (նմուշներ 1, 10 եւ 12): Առաջին բազայինի (նմուշ 1), †- ի զգալի տարբերությունների համարp <0.05:
Առանձին միակողմանի, բազմակողմանի միջոցներ ANOVA- ները կատարվեցին իգական 1- ի (նմուշներ 1-12) եւ կանանց 2- ի (նմուշներ 12-17) նյարդաքիմիական տվյալների վրա: A priori համեմատությունները կատարվել են Dunn- ի բազմակի համեմատության փորձով (Bonferroni t): 1- ի եւ 1- ի նմուշների համեմատ (2) նախնական բազային (նմուշ 10) եւ 1-2- ի (առաջին 10- ի ազդեցությունը), (11) երկրորդ բազային (օրինակ 12) նմուշները (1) , եւ (3) երրորդ բազային (նմուշ 12) versus 13-17 նմուշները (տառապում են իգական 2):
Կա կարճ ընդհանուր փոփոխություն DA efflux ի պատասխան իգական 1 [F (11,44) = 8.48; p <0.001] և կին 2 [F (5,20) = 2.83;p <0.05]: Հայտնաբերվել է DA հոսքի զգալի աճ, երբ էկրանի ետևում է կին 1-ը (+ 44%,p <0.05; նմուշ 2): Հավաքման ընթացքում DA- ի կոնցենտրացիաները հետագայում աճեցին ՝ հասնելով առավելագույն արժեքի (+ 95%;p <0.01) առաջին համապարփակ մենամարտի ընթացքում (օրինակ 3): DA- ն բարձր մակարդակի վրա մնաց բարձր և միայն վերադարձավ ելակետային կոնցենտրացիաներին 30 րոպե ժամանակահատվածում, երբ որևէ մոնտաժ չի եղել (նմուշներ 9 և 10): Ոչ էկրանի ետևում գտնվող 1-ի իգական սեռի վերականգնումը (նմուշ 11) և ոչ էլ ֆիզիկապես փոխազդելու հնարավորություն, բայց առանց տեղադրելու (նմուշ 12), բարձրացված DA- ի կոնցենտրացիաները `ելակետային երկրորդ արժեքի համեմատ (նմուշ 10): Էկրանի ետևում կին 2-ի առկայությունը (նմուշ 13) հանգեցրեց DA հոսքի փոքր աճի (12%) երրորդ բազային արժեքից (նմուշ 12), որը չի հասել վիճակագրական նշանակության: 2-րդ կանանց հետ նոր զուգակցումը հանգեցրեց զգալի (34%) աճի (p <0.05) DA հոսքում `առաջին համակցման նմուշի ընթացքում (նմուշ 14): Չնայած հաջորդ երեք նմուշների ընթացքում շարունակվել է թույլ համակարգչային վարքագիծը, DA- ի կոնցենտրացիաները իջել են ելակետային արժեքների (15–17 նմուշներ): Անկախ t «բազային» նմուշների (օրինակ `1, 10 եւ 12) միջեւ կատարված թեստերը ցույց տվեցին, որ այդ արժեքները զգալիորեն տարբեր չեն:
Երեք առնետների վերակենդանացումն այն ժամանակ վերսկսեց, երբ կին 1- ը վերաարտադրվեց, NAC DA- ի կոնցենտրացիան ավելացավ, երբ կին 1- ը ներկայացրեց էկրանին (շարք, 25-47%) եւ նմուշի նմուշի համեմատ `կուտակման ժամանակ (շարք, 13-37%): մինչեւ կանանց վերաինտեգրումը: Այդ աճը, սակայն, միայն տեղի ունեցավ, երբ սեռական վարքը ուժեղ էր եւ հանգեցրեց զզվելի:
DOPAC- ում զգալի ընդհանուր փոփոխություններ [F (11,44) = 9.57; p <0.001] և HVA [F (11,44) = 12.47; p <0] կոնցենտրացիան հայտնաբերվել է ի պատասխան կին 001-ի, բայց ոչ կնոջ 1. Մետաբոլիտի կոնցենտրացիան փոքր-ինչ աճել է (երկու դեպքում `+ 2%) էկրանի ետևում կին 15-ի ներկայացման ժամանակ (նմուշ 1), բայց դա նշանակալի չի եղել: վիճակագրորեն Այնուամենայնիվ, տեղի ունեցան զգալի աճեր DOPAC- ի և HVA- ի կոնցենտրացիաների համակցման ընթացքում (նմուշներ 2-3), հասնելով առավելագույն արժեքների (համապատասխանաբար +8 և + 80%). p <0.01) 60 րոպե հետո (նմուշ 6-ը երկու դեպքում էլ): Չնայած մետաբոլիտների կոնցենտրացիաները իջել են սեռական անգործության ժամանակահատվածում իգական սեռի 1-ի հետ շփման վերջում (9-րդ և 10-րդ նմուշներ), կոնցենտրացիաները դեռ բարձր են մնացել ելակետայինի մասովp <0.05 երկու դեպքում էլ): Էկրանի ետևում 1-ին իգական սեռի վերականգնումը (նմուշ 11), էկրանի հեռացումից հետո կին 1-ի մուտքը (նմուշ 12) և իգական սեռի 2-ի (նմուշ 13) ներմուծումը չի հանգեցրել մետաբոլիտի կոնցենտրացիայի որևէ փոփոխության: DOPAC- ի և HVA- ի կոնցենտրացիաների աննշան, բայց վիճակագրորեն աննշան աճը (երկու դեպքում + 23%) ելակետայինի համեմատ (12-րդ նմուշ) համապատասխանում էր իգական սեռի առաջին խմբաքանակի առաջին փուլին (նմուշ 2): Այս աճը, սակայն, կարճ տևեց և հրաժարվեց մնացած երեք նմուշների բազային արժեքներից (14–15): Անկախ t«բազային» նմուշների (1, 10 եւ 12) միջեւ կատարված թեստերը ցույց են տվել, որ երկրորդ եւ երրորդ բազային արժեքները (համապատասխանաբար 10 եւ 12 նմուշները), չնայած միմյանցից տարբեր չեն, զգալիորեն զգալիորեն մնացել են առաջին բազային նմուշից DOPAC- ի եւ HVA- ի համար (p <0.05 երկու դեպքում էլ):
Հիստոլոգիա
Միկրոդալիզային զոնդերը տեղակայված էին NAC- ում (նկ.2) + 1.20- ից մինչեւ + 1.70 մմ երկարություն ունեցող bregma- ից (հարթ գանգ) երկարաձգված մի շարք: Միջերկրածովյան ինքնաթիռում փոփոխություն եղավ նաեւ. Տվյալները արտացոլում են NAC- ի վահանակի եւ հիմնական ենթաբաժինների նմուշառումները:
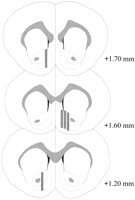
Դիտել ավելի մեծ տարբերակ:
Նկ. 2.
Քուլիջի ազդեցության փորձի մեջ օգտագործված արական առնետների NAC- ի մանրադիտակային զոնդերի գտնվելու վայրը: Shaded ուղղանկյուններ համապատասխանում է միկրոդիականիզացված զոնդերի բացված մեմբրանի տարածքին: Սինթետիկ կորոնալ ուղեղի հատվածները վերափոխվեցինPaxinos եւ Watson (1986).
ՔՆՆԱՐԿՈՒՄ
Նախկին զեկույցների համաձայն, ներկա արդյունքները ցույց են տալիս, որ տղամարդկանց սեռական վարքի ախորժելի եւ սպառողական բաղադրիչները կապված են բարձրացված mesolimbic DA փոխանցում,ի Վիվո մանրադիտակ (Mas եւ այլն, 1990; Pfaus et al., 1990;Հավիմ եւ այլն, 1990; Damsma et al., 1992; Wenkstern et al., 1993; Fumero et al., 1994; Mas եւ այլն, 1995a,b,d): Բացի այդ, այդ արդյունքները ապահովում են նյարդային քրոմատոլոգիական հարաբերություն սեռական սթրեսի եւ հետագայում վերարտադրության վերականգնում `ի պատասխան վեպի ընդունող կանանց (The Coolidge ազդեցությունը): Ներկայացված տվյալները ցույց են տալիս, որ վեպի ընդունող կանանց խթանման հատկությունները կարող են նպաստել NAC DA- ի փոխանցման սեռավարակ արական սեռին, որը, իր հերթին, կարող է կապված լինել սեռական վարքի վերականգնման հետ: Սա առաջին հերթին ակնհայտ է NAC DA- ի փոքր աճի վրա, որը ցուցադրվում է էկրանին ետեւում գտնվող վեպի կին կնոջ շրջանում եւ առավել համոզիչ կերպով տեղի է ունենում որպես ավելի հստակ աճ ՀՆԱՄՔՍ-ի հետ նորացված կուլտուրայի ժամանակ (նկ.1).
Էկրանի ետեւում առաջին ընկալիչ կնոջ առկայությունը հանգեցրեց NAC DA- ի հոսքի աճի (44% - ից ելակետային) ուժեղ ախորժակի աճին, որն առաջացել է նախորդ փորձարկումներում, օգտագործելով նմանատիպ դիզայն (30%, Pfaus et al., 1990Բ) 35%,Damsma et al., 1992) Այս ուսումնասիրությունների հետ համաձայնեցվեց նաև այն դիտարկումը, որ համակցման ընթացքում NAC DA- ի հոսքը հետագայում ուժեղացավ (սույն փորձի ընթացքում ելակետայինից բարձր> 95%): Չնայած մենք կարող ենք սպառողական վարքագիծը համարել կապված NAC DA- ի ուժեղացված թողարկման հետ (Wenkstern et al., 1993; Վիլսոն եւ այլն, 1995), կարեւոր է ուսումնասիրել «ախորժելի» եւ «սպառողական» տերմինները սեռական վարքի համատեքստում: Մինչդեռ այն փուլը, որի ընթացքում կինն առկա է էկրանի ետեւում, բացառապես ախորժելի կամ նախապատրաստական է, իսկ կուտակման փուլում վարքագիծը չի կարող համարվել սպառողական: Քանի որ «ախորժելի» կարող է օգտագործվել նկարագրելու բոլոր վարքագծերը, որոնք հանգեցնում են դրդապատճառ վարքի (կուտակման), հիմնական վարքագիծը, «սպառողական» փուլում ակտիվ ակտիվություն ունեցող տղամարդկանց ցուցումները, լավագույնս նկարագրվում են որպես ախորժելի, տղամարդը ծախսում է իր ժամանակի եւ ջանքերի մեծ մասը, հետեւելով կնոջը, Այս առումով մենք կարող ենք համեմատել NAC DA- ի առավելագույն փոխանցման սպառումը ինչպես նաեւ արական սեռական վարքի ինտենսիվ ախորժելի բաղադրիչները:
Երկրորդ, վեպի կանանց մատչելիությունը հանգեցրեց ամեն առարկայի նորացված կուտակմանը: Նախորդ ուսումնասիրությունները ցույց են տվել, որ առնետների մեծամասնությունը թույլ է տվել, որ հագեցած է սնուցումը, օգտագործելով նման վարքային արձանագրություն, որն օգտագործվում է ներկա փորձի մեջ, չի վերսկսել զուգավորումը, երբ փորձարկվել է 24 ժամ հետո (Լողափ եւ Հորդանան, 1956): Հավանական է, որ կանանց 2- ի վեպի խթանման հատկությունների առկայությունը, որը կարող էր ներառել խնձոր, ինչպես նաեւ տեսողական եւ լսողական դրսեւորումներ, հանգեցրեց նորացված կուտակմանը: Հետաքրքիր մի հարց, որը մնում է պատասխանել, այն է, ինչ մեխանիզմով արական սեռը տարբերվում է մի կնոջից նոր կնոջ հետ, որի հետ նա վերջին անգամ համտեսել է: Այդ մեխանիզմի համար կայքը կարող է տեղակայվել հիմնական գարեջրի համակարգում: Զեկուցվել է, որ այս համակարգի ամբողջականությունը կարեւոր է Քուլիչի ազդեցության համար համստերների (Ջոնսթոն եւ Ռասմուսեն, 1984): Այն vomeronasal- աքսեսուարային խնձորօգտագործման համակարգը, սակայն, այն ժամանակ, երբ մկների մեջ նկարագրվում էր մի ֆերոմոնալ հիշողություն:Kaba et al., 1994), նաեւ վարչապետ թեկնածու է: Այս առումով հատկանշական է, որ NAC DA- ի փոխանցման աճը օգտագործվել է օգտագործելով ի Վիվո Վոլտամետրը տղամարդկանց առնետներում ներկայացված է մահճակալներով, որոնք ենթարկվել են էստրուսի (Louillot et al., 1991; Mitchell եւ Gratton, 1992): Ավելին, Կ+ անմիջապես աքսեսուարային օֆսեթային լամպի vomeronasal նյարդային շերտի, ինչպես նաեւ օժանդակ օպտիկական լամպի ինքնին, բավական էր բարձրացնել NAC DA փոխանցումը (Mitchell եւ Gratton, 1992).
15- ով առաջին 2 րոպե հափշտակումը կապված էր NAC DA- ի զգալի աճի հետ: Ի տարբերություն կանանց 1- ի, 2- ի հետ փոխազդեցությունը չի առաջացրել NAC DA- ում նույն ուժգնության աճը կամ ախորժելի (12%) կամ սպառողական (34%) փուլերում: Այս փոքր աճը NAC DA- ում, սակայն, լավ է համեմատվում կանանց 2- ի հետ 1- ով համեմատած սեռական վարքի կրճատված մակարդակի հետ: Մեթաբոլիտի կոնցենտրացիաները մնացին բարձր մակարդակով սահունության փուլում, ինչը հանգեցրեց նոր բազային կոնցենտրացիաների (նմուշներ 10 եւ 12), որոնք զգալիորեն բարձրացվել են նախնական սկզբնական արժեքից (օրինակ 1):
DOPAC- ի եւ HVA- ի կոնցենտրացիաների աճի ժամանակային ուշացումը համադրման ընթացքում համահունչ է դրանց կազմմանը `որպես ծնող բաղադրիչի մետաբոլիտներ, DA: Առաջարկվել է, որ միկրոդիսիզի մեթաբոլիտի կոնցենտրացիաները, գոնե ֆարմակոլոգիական առաջացնող բնական վարքագծի ժամանակ, ապահովում են նյարդային ակտիվության օգտակար ցուցանիշ (Damsma et al., 1992; Fumero et al., 1994): Այն փաստը, որ մետաբոլիտի կոնցենտրացիաները մնացել են նույնիսկ սեռական անգործության ժամանակահատվածներում այս փորձարկումում, երբ DA կոնցենտրացիաները վերադարձել են նախնական բազային արժեքներին, կասկածի տակ են դնում այս առաջարկը:
Այս փորձառության մեջ հայտնաբերված DA մետաբոլիտի կոնցենտրացիաների համառ բարձրացումը, առնետներում դիտվող ԴԱ մետաբոլիտների միջին մեխանիկական նախաբեղմային տարածքի (mPOA) պրոֆիլը, առաջին օրվանից հետո,Mas եւ այլն, 1995a,b): NAC- ի կամ mPOA- ի DOPAC- ի եւ HVA- ի կոնցենտրացիաների կայուն բարձրացումները միշտ չէ, որ նկատվում են, երբ ճաշացանկը կայուն տեւողությունն է, որը շատ ավելի կարճ է, քան պահանջվող ժամանակը բավարարելու համար: Օրինակ, բազմաթիվ հետազոտություններ ցույց են տվել, որ DOPAC- ի կոնցենտրացիաները աճել են եւ մնացել բարձրացված `կուտակման ժամանակ, սակայն շուտով հրաժարվել են նախնական արժեքներից:Pfaus et al., 1990; Հավիմ եւ այլն, 1990; Damsma et al., 1992;Hull et al., 1993; Wenkstern et al., 1993; Hull et al., 1995): Հետազոտության կողմից Mas եւ դ. (1995b), mPOA- ի DOPAC- ի եւ HVA- ի բուսական արտանետումներից բաղադրիչները շարունակում էին բարձրացվել 4 հաջորդական օրերի ընթացքում, որոնք համապատասխանում էին սեռական անգործության ժամանակահատվածին: Չորրորդ օրն անմիջապես կենդանիների վերարտադրումը սկսեց, երբ metabolites- ի բազային կոնցենտրացիաները մոտ էին մոտավոր արժեքների: Հեղինակները համեմատեցին նյարդային քիմիական փոփոխությունների օրինակները, որոնք հայտնաբերվել են DA receptor blockers- ի (Zetterström et al., 1984; Imperato եւ DiChiara, 1985) եւ առաջարկել են, որ սեռական անգործության վիճակը կարող է միջամտել պրոլակտինային արձակման միջոցով, որը կարող է հանդես գալ որպես «էնդոկրին նեյրոլտտիկ» (Mas եւ այլն, 1995a,b,d): Ակնհայտ է, որ նյարդային համակարգերի վարումը ուղեկցվում է արտազատվող մետաբոլիտի կոնցենտրացիաների ավելացմամբ եւ DA efflux- ով (Zetterström et al., 1984; Imperato եւ DiChiara, 1985): Ցավոք, Mas եւ դ. (1995a,b) չկարողացան հայտնաբերել mPOA DA- ի կոնցենտրացիաները: Սույն ուսումնասիրության մեջ NAC- ի DA կոնցենտրացիաները վերադարձվել են precopulation արժեքներին, մինչդեռ DOPAC- ի եւ HVA- ի կոնցենտրացիաները մնացել են բարձր: Այս նմուշը հակասում է NAC- ում գործող էնդոգեն նեյրոէլպտիկի դերի հետ, որը սեռական հակում է առաջացնում:
Հաշվի առնելով mesolimbic DA neurons- ի ներգրավվածությունը մոտիվացված վարքով (Fibiger եւ Phillips, 1986; Բլեքբեռնը եւ այլք, 1992; Kalivas եւ այլոք, 1993; LeMoal, 1995) եւ նրանց զգայունությունը նոր վտանգավոր շրջակա միջավայրի խթանների նկատմամբ (Ֆաբրեն եւ այլք, 1983; Շուլց, 1992; Mirenowicz եւ Schultz, 1994), NAC DA- ի արտերկրյա խտությունների դիտարկված աճը, ի պատասխան վեպի կանանց, համապատասխանում է այն ենթադրությանը, որ այս DA համակարգում գործունեությունը կարեւոր է սեռական վարքի վերականգնման համար: Բացի դրանից, DA հաղորդման մեջ ախորժակի եւ սպառողական մեծությունների մասին հաշվետվություններըHull et al., 1993, 1995;Mas եւ այլն, 1995b; Sato et al., 1995) եւ նեյրոնային ակտիվություն (Shimura et al., 1994) սեռական վարքի ընթացքում տղամարդկանց առնետների mPOA- ում նշում են, որ այս կառուցվածքը կարող է նպաստել նաեւ Coolidge- ի ազդեցությանը բնորոշ նորացված կուտակմանը:
Ընդհանուր դերակատարություն ունենալով mesolimbic DA համակարգի մոտիվացված վարքագծում, լավ է հաստատված, որ DA- ի արտերկրյա կոնցենտրացիաները նույնպես բարձրանում են նախքան, ընթացքում եւ անմիջապես հետո կերակուրը սպառելուց հետո, նախնական արժեքների վերադարձը ~ 30 րոպե հետո (Վիլսոն եւ այլն, 1995): Հայտնի է, որ սննդակարգով սննդակարգը ազդում է իր զգայական հատկությունների վրա: Մարդիկ եւ կենդանիները մերժում են այն կերակուրը, որով նրանք կերակրում էին սնունդը եւ սնվելու այլ կերակուրներ, որոնք չեն կերել (Rolls, 1986): Սա բարձրացնում է այն հարցը, թե արդյոք NAC- ում արտանետվող DA արտանետումները կբարձրացնեն ընտրված սննդի նոր տեսակի ներկայացման միջոցով, այլ ոչ թե սննդով վերջերս սպառված սննդով, այնպես, ինչպես սույն ուսումնասիրության մեջ ներկայացված է նմանատիպ ուսումնասիրության համատեքստում: սեռական մոտիվացիա: Եթե հաստատված լինեն, բնական հարգանքների, հագեցվածության եւ մազալիմիկի DA փոխանցման զգայական հատկությունների միջեւ այս ընդհանուր կապը նշանակալի դեր կխաղա այս նյարդային համակարգի համար, մոտիվացիոն գործընթացների կարգավորման մեջ, որի խաթարումը կարող է հանգեցնել սննդի եւ սննդի լուրջ խանգարումների: .
Հիշատակում
- ↵
- Beach FA,
- Հորդանանի Լ
(1956) Սեռական առնետի սեռական սպառումը եւ վերականգնումը: QJ Exp Psychol 8- ը `121-133:
- ↵
- Բերմանտ Գ,
- Lott DF,
- Անդերսոն Լ
(1966) Քուլիդի ազդեցության ժամանակային բնութագրերը տղամարդկանց առնետների քերծվածքային վարքագծում: J Comp Physiol Հոգեբուժություն 65- ը `447-452:
- ↵
- Բլեքբեռն JR,
- Pfaus JG,
- Phillips AG- ն
(1992) Dopamine գործառույթները ախորժելի եւ պաշտպանողական վարքագծի մեջ: Prog Neurobiol- ը 39- ը `247-279:
- ↵
- Դամմա Գ,
- Pfaus JG,
- Wenkstern D,
- Phillips AG,
- Fibiger HC- ը
(1992) Սեռական վարքագիծը դոպամինի փոխանցումն ավելացնում է արական առնետների ճառագայթների ակտեմենտի եւ ստրիատումում `համեմատելի նորության եւ շարժունակության հետ: Ահավ Նեյրոսցի 106- ը `181-191:
- ↵
- Ֆաբրե Մ,
- Rolls ET,
- Էշթոն JP,
- Ուիլյամս Գ
(1983) Նեյրոնների ակտիվությունը, որը զբաղեցնում է կապիկի երկնային շրջանում: Այո 9- ը `213-235:
- ↵
- Fibiger HC,
- Phillips AG- ն
(1986) Պարգեւատրումը, մոտիվացիան, ճանաչումը, mesotelencephalic dopamine համակարգերի հոգեբանաբանությունը: Ֆիզիոլոգիայի ձեռնարկը. նյարդային համակարգի IV, eds Bloom FE, Geiger SD (American Physiology Society, Bethesda, MD), pp 647-675:
- ↵
- Ֆիորինո DF,
- Coury AG,
- Fibiger HC,
- Phillips AG- ն
(1993) Վրացական ներգաղթային տարածքում պարգեւավճարների կայանների էլեկտրական խթանումը մեծացնում է դիպլոմային փոխանցումը առնետի ակմեմենտներում: Այո 55- ը `131-141:
- ↵
- Ֆիշեր Ա
(1962) Սեռական շնչառության վրա խթանման տատանումների ազդեցությունը արական առնետում: J Comp Physiol Հոգեբուժություն 55- ը `614-620:
- ↵
- Fowler H,
- Ուլեն Ռ
(1961) Մարդկային առնետում խթանող խթանման եւ սեռական վարքի փոփոխումը: J Comp Physiol Հոգեբուժություն 54- ը `68-71:
- ↵
- Ֆումերո Բ,
- Fernendez-Vera JR,
- Gonzalez-Mora JL,
- Mas M
(1994) Մարմնի շրջանառության փոփոխություններ, որոնք առնչվում են տղամարդկային սեռական վարքագծի հետ `միկրոալիզի հետազոտություն: Brain Res 662- ը `233-239:
- ↵
- Հոլմս GM,
- Holmes DG,
- Sachs BD- ն
(1987) IBM-PC- ի վրա հիմնված տվյալների հավաքագրման համակարգ, կրծող սեռական վարքի եւ ընդհանուր իրադարձության ձայնագրության համար: Physiol Behav- ը 44- ը `825-828:
- ↵
- Հալլ EM,
- Eaton RC,
- Մովսես Ջ,
- Լորայր Դ
(1993) Կոպուլյացիան ավելացնում է դոպամինի ակտիվությունը արական առնետների միջերկրածովային շրջանում: Կյանքի գիտություն 52- ը `935-940:
- ↵
- Հալլ EM,
- Ջիանֆանգ Դ,
- Լորենայն DS,
- Մատուսեեւիչ Լ
(1995) Արտազեկումային դոպամին `միջին միջամտության շրջանում. Սեռական մոտիվացիայի եւ գլխուղեղի հորմոնալ վերահսկողության հետեւանք: J Neurosci 15- ը `7465-7471:
- ↵
- Imperato A,
- Դիխիարա Գ
(1985) Dopamine- ի թողարկում եւ նյութափոխանակություն արթնացնող աթոռներում, համակարգային նեյրոէլպտիկներից հետո, երբ ուսումնասիրվում է տրանստրիատիվ դիալիզը: J Neurosci 5- ը `297-306:
- ↵
- Johnston RE,
- Ռասմուսեն Կ
(1984) Կանացի խոզապուխտների անհատական ճանաչում տղամարդկանց կողմից. Քիմիական զուգահեռների եւ քաղցկեղի եւ վոմերոնալ համակարգերի դերը: Physiol Behav- ը 33- ը `95-104:
- ↵
- Կաբա Հ,
- Հայաշի Յ,
- Higuchi T,
- Nakanishi S
(1994) Խառնուրդի հիշողությունը ներդիրավորումը `metabotropic glutamate reseptor- ի ակտիվացման միջոցով: գիտություն 265- ը `262-264:
- ↵
- Kalivas PW,
- Sorg BA,
- Կեռիկներ MS
(1993) Դեղագիտության եւ հոգեսթիմուլյացիաների նկատմամբ զգայունացման նեյրոնային սխեման: Behav ֆարմակոլ 4- ը `315-334:
- ↵
- LeMoal M
(1995) Mesocorticolimbic dopaminergic neurons: Ֆունկցիոնալ եւ կարգավորող դերեր: Psychopharmacology- ում. առաջընթացի չորրորդ սերունդ, EDS Bloom FE, Kupfer DJ (Raven, Նյու Յորք), pp 283-294:
- ↵
- Լուիլոտ Ա,
- Gonzalez-Mora JL,
- Գուադալուպե Տ,
- Mas M
(1991) Սեքսի հետ կապված հյուսվածքի խթանները տղամարդկանց առնետների ճառագայթային ակամենսներում դոպամինի ազատման ընտրովի աճի առաջացում են առաջացնում: Brain Res 553- ը `313-317:
- ↵
- Mas M,
- Gonzalez-Mora JL,
- Լուիլոտ Ա,
- Sole C,
- Գվադալուպե Թ
(1990) Ավելի բարձր դոպամինային արձակում է արյան ողնաշարի ճարպակալման ճառագայթների ակունգենսներում, որոնք վկայում են վիվո վոլտամետրերի մասին: Neurosci լատիշ 110- ը `303-308:
- ↵
- Mas M,
- Ֆումերո Բ,
- Fernandez-Vera JR,
- Gonzalez-Mora JL
(1995a) Սեռական սպեցիֆիկացիայի եւ վերականգնման նեյրոքիմիական փոխկապակցվածություն, որը գնահատվել է in vivo microdialysis- ով: Brain Res 675- ը `13-19:
- ↵
- Mas M,
- Ֆումերո Բ,
- Gonzalez-Mora JL
(1995b) Վոլտամետրիկ եւ միկրոդիլիզի դիտում ուղեղի մոնոամինային նյարդային հաղորդիչի արձակում `սոցիոլոգիական փոխազդեցությունների ընթացքում: Այո 71- ը `69-79:
- ↵
- Mas M,
- Ֆումերո Բ,
- Պերեզ-Ռոդրիգես I
(1995c) Սեռական առումով առնետների մեջ apomorphine- ի զուգակցված վարքի ներդաշնակեցում: Eur J Pharmacol 280- ը `331-334:
- ↵
- Mas M,
- Ֆումերո Բ,
- Պերեզ-Ռոդրիգես I,
- Gonzalez-Mora JL
(1995d) Սեռական շնչառության նյարդաքիմիա: Անխախտ ցանկության փորձարարական մոդելը: սեռական ֆունկցիայի եւ դիսֆունկցիայի դեղագործության մեջ, Ed Bancroft J- ը (Raven, Նյու Յորք), pp 115-126:
- ↵
- Mirenowicz J,
- Շուլց Վ
(1994) Պրոտատե դոպամիներգիկ նեյրոնների վարձատրության պատասխանների կարեւորությունը: J Neurophysiol- ը 72- ը `1024-1027:
- ↵
- Mitchell JB,
- Գաթթոն Ա
(1992) Mesolimbic dopamine- ի արձագանքը, որն առաջացել է օժանդակ սարքերի ակտիվացման միջոցով. Բարձր արագությամբ քրոնոամերոմետրիկ ուսումնասիրություն: Neurosci լատիշ 140- ը `81-84:
- ↵
- Paxinos G,
- Վաթսոն Ջ
(1986) Սաթե ուղեղը ստերեոտաքսիկ կոորդինատներում (2nd ed): (Ակադեմիական, Սան Դիեգո):
- ↵
- Pfaus JG,
- Դամմա Գ,
- Nomikos GG,
- Wenkstern D,
- Blaha CD,
- Phillips AG,
- Fibiger HC- ը
(1990) Սեռական վարքագիծը մեծացնում է կենտրոնական դոպամինի փոխանցումը արական առնետի մեջ: Brain Res 530- ը `345-348:
- ↵
- Pfaus JG,
- Everitt BJ- ն
(1995) Սեքսուալ վարքի հոգեֆարմարաբանությունը: Psychopharmacology- ում. առաջընթացի չորրորդ սերունդ, EDS Bloom FE, Kupfer DJ (Raven, Նյու Յորք), pp 743-758:
- ↵
- Pfaus JG,
- Գորզալկա Բ
(1987) Օփիոիդներ եւ սեռական վարք: Neurosci Biobehav Rev 11- ը `1-34:
- ↵
- Phillips AG,
- Blaha CD,
- Pfaus JG,
- Բլեքբեռն JR
(1992) Դրական էմոցիոնալ վիճակների նեյրոբիոլոգիական փոխկապակցվածությունը. Դոպամին, ակնկալիք եւ պարգեւ: Զգացմունքների ուսումնասիրության միջազգային ուսումնասիրության մեջ, Ed Strongman (Wiley, Նյու Յորք), pp 31-50:
- ↵
- Pleim ET,
- Matochik JA,
- Բարֆելը RJ,
- Auerbach SB- ն
(1990) առնետների մեջ դոպամինի ազատման հարաբերակցությունը առնետների առնական սեռական վարքի հետ: Brain Res 524- ը `160-163:
- ↵
- Rodriguez-Manzo G,
- Ֆերնանդեզ-Գուանտին Ա
(1994) Սեռական սպառվածության վերացում serotonergic եւ noradrenergic գործակալների կողմից: Այո 62- ը `127-134:
- ↵
- Rodriguez-Manzo G,
- Ֆերնանդեզ-Գուանտին Ա
(1995a) Կենտրոնական noradrenergic համակարգի մասնակցությունը սեռական սպառված առնետների քորոցային վարքագծի վերականգնմանը yohimbine, naloxone եւ 8-OH-DPAT- ի կողմից: Brain Res Bull 38- ը `399-404:
- ↵
- Rodriguez-Manzo G,
- Ֆերնանդեզ-Գուանտին Ա
(1995b) Opioid antagonists եւ սեռական սնուցման երեւույթը: Psychopharmacol 122- ը `131-136:
- ↵
- Rolls BJ- ն
(1986) Sensory-specific հագեցվածություն: Nutr Rev 44- ը `93-101:
- ↵
- Salamone JD- ն
(1996) Մոտիվացիայի վարքային նյարդահիմություն. Մեթոդաբանական եւ կոնցեպտուալ հարցեր միջուկի ականջի դոպամինի դինամիկ ակտիվության ուսումնասիրության մեջ: J Neurosci մեթոդները 64- ը `137-149:
- ↵
- Սաթո Յ,
- Wada H,
- Հորիտա Հ,
- Suzuki N,
- Շիբույա Ա,
- Ադայի Հ,
- Կաթո Ռ,
- Tsukamoto T,
- Կումամոտո Յ
(1995) Առնետների ճարպակալման ժամանակ դոպամինի բացթողումը միջերկրական առաջխաղացման շրջանում: Brain Res 692- ը `66-70:
- ↵
- Շուլց Վ
(1992) Դոպամինային նեյրոնների ակտիվությունը վարակիչի մեջ: Սեմին Նեուրոսցի 4- ը `129-138:
- ↵
- Շիմուրա Տ,
- Յամամոտո Տ,
- Շիմոկոչի Մ
(1994) Առողջական առաջխաղացման տարածքը ներգրավված է արական սեռական հարաբերության մեջ եւ սեռական արատների եւ կատարման մեջ. Ազատ շարժվող կենդանիների նեյրոնային գործունեության վերագնահատում: Brain Res 640- ը `215-222:
- ↵
- Wenkstern D,
- Pfaus JG,
- Fibiger HC- ը
(1993) Դոպամինի փոխանցումը մեծացնում է արական սեռի ատամնաբույժների վրա սեռական ընդունող կանանց առնետների առաջին ազդեցության ժամանակ: Brain Res 618- ը `41-46:
- ↵
- Wilson C,
- Nomikos GG,
- Collu M,
- Fibiger HC- ը
(1995) Մոտիվացված վարքի դոպամիներգիկ փոխկապակցվածությունը `քշելու կարեւորությունը: J Neurosci 15- ը `5169-5178:
- ↵
- Wilson JR,
- Kahn RE,
- Beach FA- ն
(1963) Մոդիֆիկացիա տղամարդկանց առնետների սեռական վարքագծում `արտադրվող խթանիչ կնոջ փոխելով: J Comp Physiol Հոգեբուժություն 56- ը `636-644:
- ↵
- Zetterström T,
- Sharp T,
- Ungerstedt U
(1984) Նեյրոլեպտիկ դեղերի ազդեցությունը ստիատալալ դոպամինի ազատման եւ նյութափոխանակության վրա, ներխուժական շառավիղում, ներգրավված դիալիզի միջոցով: Eur J Pharmacol 106- ը `27-37:
Հոդվածներ, հղում կատարելով այս հոդվածին
- Ventral Tegmental տարածքում դոպամիներգիկ նեյրոնների էնդոկենտի օփիոիդային հորմոնալ նյարդապոլիտուցիկությունը ազդում է բնական եւ օպիատիվ պարգեւի վրա Նեյրոնագիտության ամսագիր, 25 Հունիս 2014, 34 (26), 8825-8836
- Անպաշտպանությունը պաթոլոգիական խաղերի տարբեր տեսակի պարգեւների զգայունության մեջ Ուղեղ, 1 օգոստոս 2013, 136 (8), 2527-2538
- Բնական եւ դեղորայքային պարգեւատրումների մասին ակտը {Delta} FosB- ի հետ որպես սովորական միջնորդի ընդհանուր նյարդային պլաստիկության մեխանիզմների վերաբերյալ Նեվոլոգիայի ամսագիր, 20 փետրվարի 2013, 33 (8), 3434-3442
- Neuronal արձագանքներ են միջուկը Accumbens Shell ընթացքում սեռական վարքագծի տղամարդիկ աթոռներ Նեվոլոգիայի ամսագիր, 1 փետրվարի 2012, 32 (5), 1672-1686
- Ռոմանտիկ սերը. Կատվի ուղեղի ուղեղի ընտրության համար Թագավորական հասարակության փիլիսոփայական գործարքները B: Կենսաբանական գիտություններ, 29 դեկտեմբեր 2006, 361 (1476): 2173-2186
- Հեդոնային տաք կետերը ուղեղում The Neuroscientist, 1 Դեկտեմբեր 2006, 12 (6): 500-511
- Մաթեմատիկայի կենտրոնական եւ բազոլարային Amygdalar միջուկները եւ Dopaminergic փոխկապակցվածները սնուցում է սեռական Nucleus Accumbens եւ Medial Prefrontal Cortex Նեվոլոգիայի ամսագիր, 15 Դեկտեմբեր 2002, 22 (24), 10958-10965
- Դիֆամինի համակենտրոնացումների հաճախականությունը տղամարդկանց առնետների դորալային եւ վրացական շերտերում տատանումների աճն է Conspecifics- ի ներդրման ընթացքում Նեվոլոգիայի ամսագիր, 1 Դեկտեմբեր 2002, 22 (23), 10477-10486
- Book Review: Արտանետումների Դոպամինի դինամիկան կոկաինի սուր եւ քրոնիկ գործողություններում The Neuroscientist, 1 Օգոստոս 2002, 8 (4): 315-322
- Պավլովյան մոտեցման վարքի ձեռքբերման եւ կատարման միջուկում գտնվող NMDA, AMPA / Kainate եւ դոպամին ընկալիչների դիֆերենցիալ ներգրավում Նեվոլոգիայի ամսագիր, 1 Դեկտեմբեր 2001, 21 (23), 9471-9477
- Ատամների հետ կապված թմրանյութերի հետ կապված խանգարումների միջոցով կոկաինի փնտրման վարքագծի վերահսկում. Ամիգդալայում եւ միջուկային հյուսվածքներում մարվող օպերատիվ-արձագանքող եւ արտազատվող դոպամինի մակարդակի վերականգնման հետեւանքները PNAS, 11 Ապրիլ 2000, 97 (8): 4321-4326
- Սեռական վարքագծի դինամիկացում եւ դոպամինային ուժեղացում, D-Amphetamine-induced վարքագծային սենսիտիզացիան հետո արական ողորկների միջուկում Նեվոլոգիայի ամսագիր, 1 հունվարի 1999, 19 (1), 456-463
- Նյարդաբանություն ՝ ուղեղի ուշադրության կենտրոնում Գիտություն, 3 հոկտեմբերի 1997, 278 (5335), 35-37
- Դիֆամիներգիկ փոխազդեցությունը զգայուն հատուկ ճառագայթման է Medial Prefrontal ծղոտե ներքնակ եւ միջուկային ակումբենս է Rat Նեվոլոգիայի ամսագիր, 1 հոկտեմբերի 1999, 19 (19), RC29