YBOPコメント:Coolidge効果は、インターネットポルノの力の背後にあります。 クーリッジ効果は哺乳類種で見られる現象であり、以前のまだ利用可能な性的パートナーからのセックスを拒否した後でも、新しい受容性的パートナーに導入された場合、男性(およびそれほどではないが女性)は新たな性的興味を示します。 性的な目新しさは、この慣れを、より高いドーパミンによって引き起こされる新たな興奮で上書きします。 絶え間ない斬新さの流れが、インターネットポルノを過去のポルノと大きく異なるものにしています。
- デニス・F・フィオリーノ、
- アリアン・クーリー、そして
- アンソニーG.フィリップス
-
神経科学ジャーナル、 15年1997月17日、12(4849):4855-XNUMX;
抽象
クーリッジ効果は、新たな受容性仲間に反応した、「性的に満腹した」動物における性的行動の再開を説明しています。 動機付けられた行動の開始と維持における中辺縁系ドーパミン(DA)システムの役割を考えると、交尾中の側坐核(NAC)DA伝達、性的満腹感、および性行動の再開を監視するために微小透析が使用された。 以前の報告と一致して、スクリーンと交尾の背後にある女性の発情の提示は、NAC DA流出の有意な増加と関連していた。 NAC DA濃度のベースライン値への復帰は、性的満腹期間と一致したが、DA代謝産物、ジヒドロキシフェニル酢酸およびホモバニリン酸の濃度は上昇したままであった。 スクリーンの後ろに新規の受容的な女性を提示すると、NAC DAがわずかに増加したが、これは新規の女性との交尾交尾中に有意に増強された。 本データは、新規の受容性女性の刺激特性が、性的に満足している雄性ラットにおけるNAC DA伝達を増加させるのに役立ち得ることを示唆し、そしてこれは、今度は性的行動の再開に関連し得る。
イントロダクション
満腹まで交尾した雄ラットは、最初の雌が新規の受容雌と交換された場合に再び交尾するように誘導することができる。 これはクーリッジ効果として知られるようになり、多くの哺乳動物種で観察されています(Wilsonら、1963) 新しい女性からの刺激は依然として交尾を誘発する可能性があるため、疲労や運動性うつ病などの一般的な要因では、性的満足感の見かけの状態を説明するのに十分ではありません。 性的満腹感はまた、異なる神経伝達物質系に作用することができる様々な薬物の投与によって、薬理学的に、かなりの程度まで「逆転」させることができる。 これらの薬はヨヒンビン、8-OH-DPAT(ロドリゲス - マンゾとフェルナンデス - グースティ、1994, 1995a)、ナラキソン(Pfaus and Gorzalka、1987; ロドリゲス - マンゾとフェルナンデス - グースティ、1995a,b)およびアポモルヒネ(Masら、1995c) これらの薬物の末梢作用(例えば勃起機能に対するアドレナリン作用)を除外することはできないが、選択的中枢性ノルアドレナリン作動性病変実験に基づいて性的満腹の根底にある中枢機構に対する作用が提案されている(ロドリゲス - マンゾとフェルナンデス - グースティ、1995a内側視索前野のドーパミン作動性代謝をモニターした)マイクロダイアリシス実験Masら、1995a,b).
中枢機構がCoolidge効果に特徴的な性行動の再開を仲介する可能性があることを考えると、有望な候補は腹側被蓋野からNACまで予測する中辺縁系ドーパミン(DA)システムである。 中辺縁系DAは、性的に受容的な女性からの手がかりなどの環境的刺激の評価、および交尾を含む目標指向行動の組織化を含む複雑な統合的プロセスにおいて主要なモジュレーターとして作用するように思われる(Fibiger and Phillips、1986; Blackburnら、1992; フィリップス他、1992; レモール、1995; サラモン、1996).
中脳DAニューロンは一次報酬に反応し、報酬を予測する手がかりを与えるが、新規または予測不可能な環境刺激は、反復訓練セッションにわたって最も強くニューロン活性化を誘導する(Fabreら、1983; シュルツ、1992; MirenowiczとSchultz、1994) ラットの性行動の開始と維持における中辺縁系DAの重要な促進的役割を支持する多くの証拠がある(Pfaus and Everitt、1995また、多くのマイクロダイアリシス研究では、男性の性行動の食欲および完了期におけるNAC DA流出の増加を報告している(Pfausら、1990; Pleimら、1990; Damsma et al。、1992; Wenksternら、1993; Fumeroら、1994; Masら、1995b) しかしながら、性欲と性行動の再開との神経化学的相関に関するデータは比較的少ない。 のアプリケーション インビボの クーリッジ効果中の中辺縁系DA神経伝達をモニターするための微量透析は、交尾、性的満腹感、および交尾の再開におけるNAC DAの役割を調べるためのユニークな機会を提供します。
微量透析実験を実施して、以下を決定した:(XNUMX)性的満腹の開始がNAC中の細胞外DA濃度の前母集団値以下への復帰を伴うかどうか、および(XNUMX)性的に満腹した雄性ラットと新規の受容性雌性ラットは、NAC DA流出の増加と相関しています。
材料および方法
科目。 Animal Care Center(British Columbia大学)から入手した雄のSprague Dawleyラット、およびCharles River Canada(St. Constant、ケベック、カナダ)から入手した雌のLong-Evansラットを金網ケージ(18)に収容した。 ×25×65 cm;ケージごとに5つ)別々のコロニー部屋。 コロニー室を、逆の20時間明/暗サイクルで〜12℃の温度に維持した。 ラットは食物(Purina Rat Chow)と水を無制限に摂取した。
脳微小透析前の手術および行動試験試験の少なくとも4週前に、雌ラットをハロタンガス麻酔下(Fluothane、Ayerst Laboratories)で両側に卵巣摘出した。 刺激試験女性における性的受容性は、各試験セッションの前に、それぞれエストラジオールベンゾエート(XNUMX μg)およびプロゲステロン(XNUMX μg)、XNUMXおよびXNUMX hrの皮下注射によって誘発された。 金網の床を有するプレキシグラスチャンバー(XNUMX×XNUMX×XNUMXcm)において、雄ラットを性行動について2回、XNUMX d離れてスクリーニングした。 2回のスクリーニング試験の間に、女性の提示のXNUMX分以内の挿入および最初の挿入のXNUMX分以内の射精を含む成績基準に達したオスのラットのみに微小透析プローブガイドカニューレを移植した。
雄ラット(n 定位手術の前に、塩酸ケタミン(XNUMX mg / kg、腹腔内)およびキシラジン(XNUMX mg / kg、腹腔内)で麻酔した。 微小透析プローブガイドカニューレ(XNUMXゲージ)をNAC上に両側に植え込み(ブレグマからの座標:前部、+ XNUMX mm;内側、±XNUMX mm;腹側、−XNUMX mm;平らな頭蓋骨)、歯科用アクリルで頭蓋骨に固定した。宝石商のネジ。 両側ガイドカニューレインプラントを使用して、微小透析実験を成功させる機会を最大にした。 幸いなことに、本実験では、各ラットに必要なカニューレは1つだけでした。 実験の残りの間、オスのラットをトウモロコシの穂軸付きの大きなプラスチック製ケージに個別に収容した。 手術の1週間後、ラットの性的行動について試験した。 訓練のこの部分の間、試験室は、試験室を大きい区画と小さい区画に分割するスライド式プレキシグラススクリーンを備えていた。 雄ラットを大きな区画に入れ、そしてXNUMX分後に雌をスクリーンの後ろに置いた。 5分の準備期間の後、スクリーンを取り除き、そしてラットに100分の間交尾させた。 10ごとに1回、合計3回のトレーニングセッションが実施された。 全てのラットは各セッション中に成績基準に達した。
クーリッジ効果実験 ラットに、Coolidge効果実験のX時間前にマイクロダイアリシスプローブ12〜18を片側から移植し、食物と水を自由に摂取させて試験室の大きな区画に入れた。 実験の朝に、微量透析サンプルを15分ごとに収集した。 実験は次の7つの連続した段階から成っていた:(XNUMX)ベースライン(少なくともXNUMX分)。 (1)女性の60が画面の後ろにあります(2 min)。 マウントなしで1の最小期間が経過するまで(15)雌の3との交尾。 (1)女性の30をスクリーンの後ろに再導入する(4分)。 (1)マウントがなかった場合、最小15期間の間、メス5へのアクセス(マウントが行われた場合、このフェーズはフェーズ1として扱われました)。 (15)女性の3をスクリーンの後ろに紹介する(6分)。 2)メス15との交尾
行動は、JVCビデオシステムを使用して低照度下で撮影し、試験室の外にあるビデオモニターで観察した。 性行動の標準的な尺度は、コンピュータと適切なソフトウェアを使用して記録された(Holmesら、1987).
微量透析実験の後、動物に過剰摂取の抱水クロラールを与え、生理食塩水およびホルマリン(XNUMX%)を心臓内に灌流した。 脳をスライスして凍結し、続いて冠状切片をクレシルバイオレットで染色して微小透析プローブの配置を決定した。 NAC内にプローブを配置したラットのみを行動分析および神経化学分析に使用した。
微小透析とHPLC‐電気化学的検出 微小透析プローブは、遠位端に半透性中空繊維膜(露出した2 mm膜、340μm外径、65000分子量カットオフ、Filtral 12、Hospal)と設計上同心であった。 プローブを修飾リンゲル液(XNUMX mリン酸ナトリウム緩衝液、pH XNUMX、XNUMX mmCaCl)でXNUMX μl /分で灌流した。2、3.0 mm KCl、X NUMX mm MgCl2、気密注射器(Hamilton、Reno、NV)および注射器ポンプ(model 147、Harvard Apparatus、South Natick、MA)を用いて測定した。 マイクロ透析プローブガイドカラーを使用して、マイクロ透析プローブをガイドカニューレの内側に固定した。 試験チャンバの上部に取り付けられた液体旋回装置(Instech 22)に取り付けられたスチールコイルを使用して、プローブ管を保護した(Fiorino et al。、1993).
DAおよびその代謝産物であるジヒドロキシフェニル酢酸(DOPAC)およびホモバニリン酸(HVA)を含む微小透析物分析物を、逆相クロマトグラフィー(Ultrasphere column;カリフォルニア州フラートン、カリフォルニア州)、ODS XNUMXμm、XNUMXcm、XNUMXmm、内径によって分離した。 )XNUMX(商標)酢酸ナトリウム緩衝液、pH XNUMX(XNUMX%メタノール)を使用する。 分析物濃度は電気化学的(EC)検出により定量した。 装置は、Bio − Rad(カリフォルニア州リッチモンド)ポンプ、Valco Instruments(テキサス州ヒューストン)ECXNUMXW 2位置インジェクター、ESA(マサチューセッツ州ベッドフォード)クーロックヘムIIEC検出器、およびデュアルチャンネルチャートレコーダー(Kipp)からなる。ゾーネ、ボヘミア、ニューヨーク州)。 電気化学検出器パラメータは以下の通りであった:電極X NUMX、+ X NUMX mV。 電極X NUMX、−X NUMX mV。 ガードセル、−XNUMX mV。 実施された典型的なプローブ回収率 ビトロ 室温では、DAは22%、DOPACは18%、HVAは18%であった。
結果
行動
クーリッジ効果実験からの行動尺度を表に示す。 1。 最初の射精後の発射後の間隔と同様に、据え付け、挿入、および射精までの待ち時間は、前の訓練セッションのものと同様であった(データは示さず)。 これは、微量透析法が通常の性行動を変えなかったことを示唆しています。 基準を満たす前の平均射精回数(7.8±0.5)、各射精の前の挿入数の漸進的減少、および射精後間隔の漸進的増加によって示されるように、性的飽食の発症(データは示さず)。は以前の研究で報告されたものと類似していた(ビーチとヨルダン、1956; ファウラーとウェーレン、1961; フィッシャー、1962; バーマント他、1966; ロドリゲス - マンゾとフェルナンデス - グースティ、1994; Masら、1995d) 女性1で達成された射精の数、女性1と交尾するのに費やされた時間、および満足基準に達するのに必要とされる女性1の提示の数に関して、個人差が観察された(表)。 1、下)。 一部のラットは、フェーズ1が完了するまで、メス5の多数の再導入を必要とした(n = 3) スクリーンの後ろに女性の1を配置し、仕切りを取り除くという行為は、交尾をもたらす主な食欲の手がかりとなっているかもしれません。 以前に使用されていましたが、マウントなしの最小30の満足基準にも注意する必要があります(Beach and Jordan、1965; Masら、1995b)は恣意的なもので、ネズミがもっと長い時間与えられなかったことを保証するものではありません。 そうであっても、遅延または除去および交換手順は、女性1との新たな交尾を確実にもたらさなかった(例えば、フェーズ4および5)。

クーリッジ効果実験中の挙動
全てのラットがクーリッジ効果を示した。 スクリーンの後ろへの女性の2の配置、そして特にパーティションの削除に関連した活動はこの結果に貢献したかもしれません、しかし、やはり、これらの出来事はそれ自身で実験の早い段階で交尾を更新するのに十分ではありませんでした。 女性1と女性2との性行動の尺度の比較は、 t Bonferroni補正でテストします。 女性の2に対するマウント潜伏潜時と最初の交尾潜時は、女性の1との最初の交尾期の潜伏期と有意に異ならなかったが、一般に、著しく少ない射精によって示されるように、女性2の性行動はそれほど強くなかった(平均、0.6対4.2。 F = 49.86;p <0.01)および導入(平均= 11.2 vs 37.0;F = 20.17; p <0.05)最初の1時間。 雌2と雌XNUMXの最初のXNUMX時間のマウント数は、有意差はありませんでした。
実験の満腹部分に使用された女性(すなわち、女性の1)は、男性との完全な接触期間の間、依然として強い前向き(すなわち、ホッピングおよびダーツ)および受け入れ的(すなわち、ロードシス)行動を示したことに留意することが重要である。 。
神経化学
最初の3つのベースラインサンプルの平均±SEMとして提示された、微小透析液中のDAおよびその代謝産物の基礎ナノモル濃度は以下の通りであった:DA、XNUMX±XNUMX。 DOPAC、3.0±0.7。 HVA、XNUMX±XNUMX(プローブ回収については補正されていない。n = 5) これらの値は100%ベースラインスコアを表した。
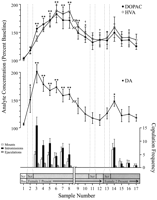
実験の各段階に対応し、すべてのラットに共通の行動的に定義されたデータ点を神経化学的分析に使用した。 これらには以下が含まれた:(XNUMX)女性XNUMXの最初の導入後の7サンプル、(XNUMX)女性XNUMXとの交尾行動の欠如を伴う4サンプル、および(XNUMX)女性XNUMXの提示後の5サンプル。 図 1 DA濃度の変化(折れ線グラフ、 真ん中)およびDA代謝物(折れ線グラフ、 top)交尾行動の平行性(棒グラフ) ボトム)Coolidge効果のテスト中
大きなバージョンを見る:
図 1。
側坐核神経化学はCoolidge効果中の性行動と相関する 最初の8つのサンプルは、フェーズ1から3までの時間的に連続したデータポイントを表します。 サンプル1は、4番目以降の事前入力ベースラインサンプルです(ロー) サンプル2は、画面の後ろに女性用の1を導入したものです(SCR) XNUMX分後、スクリーンを取り除き、ラットに交尾させた(サンプルXNUMX〜XNUMX)。 の破る x軸は、長期間にわたって最初の雌と交尾した3匹のラットからのデータの排除に対応する。 最後の9つのサンプルも年代順に連続していました。 サンプル9および10は、フェーズ3の満了期間(つまり、マウントなしの30 min)に対応します。 次いで、女性のXNUMXをスクリーンの後ろに再挿入し(サンプルXNUMX)、そしてXNUMX分後にスクリーンを取り外した(サンプルXNUMX)。 交尾のない1分後、女性の11をスクリーンの後ろに置いた(サンプル15)。 サンプル12〜15は、女性の2との交尾に対応しています。 各13 minマイクロダイアリシスサンプルに関連するマウント数、挿入数、または射精数は、 下の棒グラフ。 神経化学的データはベースライン濃度の百分率で表される。 NAC DAの変更点(閉じた正方形)、DOPAC(黒丸)、およびHVA(白丸)流出量は 折れ線グラフ。 次の比較が行われました。ベースラインサンプル1とサンプル2〜10。 サンプルXNUMX対サンプルXNUMXおよびXNUMX。 新しいベースラインサンプル10とサンプル11 – 12(*p <0.05; **** p <0.01)。 独立 t テストはベースライン値(サンプル1、10、および12)間で行われました。 最初のベースライン(サンプル1)との大きな違いについては、†p <0.05。
女性の1(サンプル1〜12)と女性の2(サンプル12〜17)に関連する神経化学データに対して、別々の一方向の反復測定ANOVAを実施しました。 アプリオリ 比較はDunnの多重比較検定(Bonferroni)を使って行われました。 t) 次の3つの主な比較が行われた:(1)初期ベースライン(サンプル1)対サンプル2〜10(最初の女性1への曝露)、(2)2番目のベースライン(サンプル10)対サンプル11および12(女性1への再曝露)そして、(3)第3ベースライン(サンプル12)とサンプル13〜17(女性2へのばく露)。
女性の1に反応して、DA排出量に有意な全体的変化があった。F (11,44) = 8.48; p <0.001]および女性2 [F (5,20) = 2.83;p <0.05]。 女性1が画面の後ろにいると、DA流出の有意な増加が見られました(+ 44%、p <0.05; サンプル2)。 交尾中、DA濃度はさらに増加し、最大値(+ 95%;p <0.01)最初の性交中(サンプル3)。 DAは交尾中ずっと上昇したままであり、マウントが発生しなかった30分間でのみベースライン濃度に戻りました(サンプル9および10)。 画面の後ろに女性1を再導入することも(サンプル11)、物理的に相互作用する機会もありませんが、マウントせずに(サンプル12)、10番目のベースライン値(サンプル2)と比較してDA濃度を上昇させました。 画面の後ろに女性13がいると(サンプル12)、統計的有意性に達していない12番目のベースライン値(サンプル2)からDA流出がわずかに増加しました(34%)。 女性XNUMXとの新たな交尾は、有意な(XNUMX%)増加をもたらしました(p <0.05)最初の交尾サンプル(サンプル14)中のDA流出。 次の15つのサンプルでは弱い交尾行動が続きましたが、DA濃度はベースライン値まで減少しました(サンプル17〜XNUMX)。 独立 t 「ベースライン」サンプル(すなわち、1、10、および12)の間で行われたテストは、これらの値が有意に異ならないことを示しました。
メスの1が再導入されたときに交尾を再開した3匹のラットでは、メスの1がスクリーンの後ろ(範囲、25〜47%)および交尾中(範囲、13〜37%)に存在したとき女性の再紹介の前に。 しかし、これらの増加は性的行動が活発で射精につながったときにのみ起こりました。
DOPACにおける大幅な全体的な変更[F (11,44) = 9.57; p <0.001]およびHVA [F (11,44) = 12.47; p <0]濃度は、女性001に反応して見られましたが、女性1には反応しませんでした。代謝物濃度は、画面の後ろに女性2が表示されている間(両方の場合で+ 15%)わずかに増加しました(サンプル1)が、これは重要ではありませんでした。統計的に。 しかし、交尾中にDOPACとHVAの濃度が大幅に増加し(サンプル2〜3)、最大値(それぞれ、+ 8と+ 80%)に達しました。 p <0.01)60分後(どちらの場合もサンプル6)。 代謝物濃度は、女性1との接触の終わりの性的不活動の期間中に減少したが(サンプル9および10)、濃度は依然としてベースラインに対して上昇したままであった(p どちらの場合も<0.05)。 スクリーンの後ろのメス1の再導入(サンプル11)、スクリーンの取り外し後のメス1へのアクセス(サンプル12)、およびメス2の導入(サンプル13)は、代謝物濃度に変化をもたらしませんでした。 ベースライン(サンプル23)と比較したDOPACおよびHVA濃度のわずかではあるが統計的に重要でない増加(両方の場合で+ 12%)は、女性2(サンプル14)との交尾の最初の発作に対応しました。 ただし、この増加は短命であり、残りの15つのサンプル(17〜XNUMX)のベースライン値まで低下しました。 独立 t「ベースライン」サンプル(すなわち、1、10、および12)の間で行われたテストは、2番目と3番目のベースライン値(それぞれサンプル10と12)は、最初のベースラインサンプルと比較して有意に上昇したままであることを示したDOPACとHVA(p どちらの場合も<0.05)。
組織学
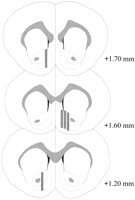
微小透析プローブはNAC内に位置していた(図1)。2)ブレグマ(平らな頭蓋骨)から+ 1.20から+ 1.70 mmの範囲。 内側外側面にも変動性がありました。 データは、NACのシェルとコアのサブリージョンからのサンプリングを反映しています。
大きなバージョンを見る:
図 2。
Coolidge効果実験に用いた雄ラットのNAC内の微小透析プローブの位置 影付きの長方形 微小透析プローブの露出膜面積に対応する。 連続冠状脳切片をから再描画した。パキシノスとワトソン(1986).
考察
以前の報告と一致して、本結果は、以下によって評価されるように、雄ラットの性行動の食欲および完了成分に関連した中辺縁DA伝達の増強を実証する。インビボの マイクロ透析(Masら、1990; Pfausら、1990;Pleimら、1990; Damsma et al。、1992; Wenksternら、1993; Fumeroら、1994; Masら、1995a,b,d) さらに、これらの結果は、新規の受容的な女性に応答した性的飽食およびそれに続く交尾の再開に対する神経化学的相関を提供する(クーリッジ効果)。 本データは、新規の受容性雌の刺激特性が性的に満足している雄性ラットにおけるNAC DA伝達を増加させるのに役立ち得、それは今度は性行動の再開に関連し得ることを示唆する。 これは、スクリーンの向こう側にある新しい女性の発表中にNAC DAがわずかに増加したことで最初に明らかになり、女性2との交尾交替中のより顕著な増加として最も説得力があるように起こる(図2)。1).
スクリーンの後ろの最初の受容性女性の存在は、同様のデザインを用いた以前の実験において報告されたものと大きさにおいて同様の大きさにおいて、NAC DA流出(ベースラインからの44%)の強い食欲増進をもたらした。 Pfausら、1990; 35%、Damsma et al。、1992)。 また、これらの研究と一致して、NAC DAの流出が交尾中にさらに増強されたという観察結果がありました(現在の実験ではベースラインを95%以上上回っています)。 消費行動は、強化されたNAC DAリリースに関連していると見なすことができますが(Wenksternら、1993; Wilsonら、1995性的行動の文脈の中で、「食欲」と「完ぺき」という用語を検討することが重要です。 女性がスクリーンの後ろにいる段階はもっぱら食欲または準備段階ですが、交尾段階の間の行動は純粋に完ぺきなものと考えることはできません。 「食欲」は動機づけられた行動(交尾)の完結につながるすべての行動を記述するために使用することができるので、「完結」段階で活動している間男性が示す主な行動は食欲として最もよく記述されます。 男性は自分の時間と努力の大部分を費やして女性を交尾させます。 これに関して、最大のNAC DA送信を消費者と相関させることができる。 と同様 雄ラットの性行動の強烈な食欲の要素。
2番目の、新しい女性へのアクセスはすべての科目で新たな交尾をもたらしました。 以前の研究は、本実験で使用されたものと同様の行動プロトコルを使用して満腹に交尾することを許されたラットの大多数は、XNUMX時間後に試験されたとき交尾を再開しなかったことを示した。ビーチとヨルダン、1956) 嗅覚的ならびに視覚的および聴覚的合図を含み得る、女性の2の新規刺激特性の存在が、新たな交尾交尾をもたらした可能性が高い。 まだ答えが残されている興味深い質問は、雄のラットが彼が最近交配した女性から新しい雌を区別するメカニズムによってです。 そのメカニズムのための場所は主要な嗅覚系にあるかもしれません。 このシステムの完全性はハムスターのクーリッジ効果に不可欠であると報告されています(ジョンストンとラスムッセン、1984) しかし、鋤鼻 - 副嗅覚系では、最近マウスでフェロモン記憶プロセスが報告されています(Kaba他、1994)も素数の候補です。 この点に関して、NAC DA伝送の増加が以下を使用して測定されたことは注目に値する。 インビボの 発情期に雌ラットに曝露した寝具を呈した雄ラットのボルタンメトリー(Louillot他、1991; ミッチェルとグラットン、1992) なお、Kの適用+ 副嗅球それ自体と同様に、副嗅球の鋤鼻神経層に直接、NAC DA伝達を増加させるのに十分でした(ミッチェルとグラットン、1992).
女性の15との交尾の最初の2最小の発作は、NAC DAの有意な増加と関連していた。 女性の1とは対照的に、女性の2との相互作用は、食欲(12%)または完了(34%)のいずれの段階でも同じ大きさのNAC DAの増加をもたらさなかった。 しかしながら、NAC DAのこれらのわずかな増加は、女性の2と比較して女性の1で示された低下したレベルの性行動とよく相関しています。 満腹期の間、代謝産物濃度は上昇したままであり、初期ベースライン値(サンプル10)から有意に上昇した新しいベースライン濃度(サンプル12および1)をもたらした。
交尾中のDOPACおよびHVA濃度の増加の時間的遅れは、親化合物DAの代謝産物としてのそれらの形成と一致している。 マイクロダイアリシス代謝産物濃度は、少なくとも薬理学的には駆動されない自然な行動の間に、神経活動の有用な指標を提供することが示唆されている。Damsma et al。、1992; Fumeroら、1994) DA濃度が試験前のベースライン値に戻ったとき、代謝物濃度がこの実験における性的不活動期間中でさえ上昇したままであったという事実は、この示唆に疑問を投げかける。
この実験で見られたDA代謝産物濃度の持続的な上昇は、満腹まで交尾した最初の日にラットで観察されたDA代謝産物の内側視索前野(mPOA)プロファイルを反映している(Masら、1995a,b) NACまたはmPOAにおけるDOPACおよびHVA濃度の持続的な上昇は、交配期間が一定期間である場合には必ずしも観察されず、満腹に達するのに必要な時間よりはるかに短い。 例えば、多くの研究は、DOPAC濃度は交尾中に増加しそして上昇したままであるが、女性が除かれた直後にベースライン値に減少することを示しました(Pfausら、1990; Pleimら、1990; Damsma et al。、1992;ハル他、1993; Wenksternら、1993; ハル他、1995) による研究では Mas et al。 (1995b)すなわち、mPOA中のDOPACおよびHVAの基礎細胞外濃度は、性的不活動の期間に対応する4連続日にわたって上昇したままであった。 4日目までに、動物が交尾を再開する直前に、代謝産物の基礎濃度はプレステーション値に近かった。 著者らは、神経化学的変化のパターンを、DA受容体遮断薬投与後に見られるものと類似させた(Zetterströmら、1984; インペラトとディキアラ、1985そして、性的不活動の状態は「内因性神経弛緩薬」として作用するかもしれないプロラクチン放出によって媒介されるかもしれないことを示唆しました(Masら、1995a,b,d) 神経弛緩薬投与が細胞外代謝産物濃度およびDA流出の増加を伴うことは明らかである(Zetterströmら、1984; インペラトとディキアラ、1985) 残念ながら Mas et al。 (1995a,b)mPOA DA濃度を検出できなかった。 本研究では、NAC中のDA濃度は前人口値に戻ったが、DOPACおよびHVA濃度は上昇したままであった。 このパターンは、性的満腹を誘発するためにNACで作用する内因性神経弛緩薬の役割と矛盾している。
やる気のある行動における中辺縁DAニューロンの関与を考えると(Fibiger and Phillips、1986; Blackburnら、1992; Kalivasら、1993; レモール、1995)およびそれらの新規環境刺激に対する感受性(Fabreら、1983; シュルツ、1992; MirenowiczとSchultz、1994)、小説の女性に反応して観察されたNAC DAの細胞外濃度の増加は、このDAシステムにおける活性が性行動の再開に重要であるという仮説と一致する。 加えて、食欲増進的および最終的なDA感染の増加の報告(ハル他、1993, 1995;Masら、1995b; 佐藤ら、1995)と神経活動(志村ら、1994性行動中の雄ラットのmPOAにおける)は、この構造もCoolidge効果に特徴的な新たな交尾交尾に寄与することを示唆している。
動機付けられた行動における中辺縁系DAシステムの一般的な役割に沿って、DAの細胞外濃度も食事の消費の前、最中、および直後に上昇し、〜30分後にベースライン値に戻ることが十分に確立されている。Wilsonら、1995) 食物によって引き起こされる満腹感がその官能特性によって影響されることはよく知られている。 人間や動物は、食べ物が満腹になるのを拒否し、食べられなかった他の食べ物を摂取します(ロール、1986) これは、NACにおける細胞外DA流出が新規なタイプの食物の提示によって選択的に増加するが、本研究において以下の文脈で報告されたものと類似の方法で満腹になるように最近消費された食物によってではないかどうかに関する問題を提起する。性的動機 確認されれば、自然な報酬の感覚特性、満腹感、および中脳辺縁系DA伝達の間のこの一般的関係は、動機づけプロセスの調節におけるこの神経系の重要な役割を示唆し、その中断は摂食および性機能の深刻な障害につながり得る。
参考文献
- ↵
- ビーチFA、
- ヨルダンL
(1956)雄ラットにおける性的枯渇および回復。 QJ Exp Psychol 8:121-133。
- ↵
- バーマントG、
- ロットDF、
- アンダーソンL
(1966)雄ラット交尾行動におけるCoolidge効果の時間的特徴 J Comp Physiol精神医学 65:447-452。
- ↵
- ブラックバーンJR、
- Pfaus JG、
- フィリップスAG
(1992)ドーパミンは食欲と防御行動において機能します。 Prog Neurobiol 39:247-279。
- ↵
- ダンマG、
- Pfaus JG、
- ウェンクスターンD、
- フィリップスAG、
- フィビガーHC
(1992)性行動は雄ラットの側坐核および線条体におけるドーパミン伝達を増加させる:新規性および歩行との比較 Behav Neurosci 106:181-191。
- ↵
- ファーブルM、
- ロールET、
- アシュトンJP、
- ウィリアムズG
(1983)行動をしているサルの腹側被蓋野におけるニューロンの活動。 Behav Brain Res 9:213-235。
- ↵
- Fibiger HC、
- フィリップスAG
(1986)報酬、動機、認識:中脳症ドーパミン系の精神生物学。 生理学のハンドブック:神経系IV、edsブルームFE、ガイガーSD(アメリカ生理学協会、ベセスダ、メリーランド州)、pp 647 – 675。
- ↵
- フィオリーノDF、
- Coury AG、
- Fibiger HC、
- フィリップスAG
(1993)腹側被蓋野における報酬部位の電気的刺激はラットの側坐核におけるドーパミン伝達を増加させる。 Behav Brain Res 55:131-141。
- ↵
- フィッシャーA
(1962)雄ラットの性的飽満に対する刺激変動の影響 J Comp Physiol精神医学 55:614-620。
- ↵
- ファウラーH、
- Whalen RE
(1961)雄ラットにおける刺激刺激と性行動の変動 J Comp Physiol精神医学 54:68-71。
- ↵
- フメロB、
- Fernendez-Vera JR、
- ゴンザレスモラJL、
- マスM
(1994)男性の性行動に関連する前脳領域におけるモノアミン代謝回転の変化:マイクロダイアリシス研究。 脳解像度 662:233-239。
- ↵
- ホームズGM、
- ホームズDG、
- サックスBD
(1987)げっ歯類の性行動を記録し、一般的なイベントを記録するためのIBM-PCベースのデータ収集システム。 フィジオールBehav 44:825-828。
- ↵
- ハルEM、
- イートンRC、
- モーセJ、
- ロレインDS
(1993)交尾は雄ラットの内側視索前野におけるドーパミン活性を増加させる ライフサイ 52:935-940。
- ↵
- ハルEM、
- Jianfang D、
- ロレインDS、
- マトゥゼイッチL
内側視索前野における(1995)細胞外ドーパミン性的動機および交尾のホルモン制御に対する意義 J Neurosci 15:7465-7471。
- ↵
- インペラトA、
- ディキアラG
(1985)経線条体透析で調べた全身性神経弛緩薬投与後の覚醒ラットにおけるドーパミン放出と代謝 J Neurosci 5:297-306。
- ↵
- ジョンストンRE、
- ラスムッセンK
(1984)男性による女性ハムスターの個人認識:化学的合図ならびに嗅覚および鋤鼻系の役割。 フィジオールBehav 33:95-104。
- ↵
- カバH、
- 林Yさん、
- 樋口Tさん、
- 中西S
(1994)代謝型グルタミン酸受容体の活性化による嗅覚記憶の誘導。 科学 265:262-264。
- ↵
- カリバスPW、
- Sorg BA、
- MSをフック
(1993)精神刺激薬に対する感作の薬理学と神経回路。 Behav Pharmacol 4:315-334。
- ↵
- レモールM
(1995)中皮質コルチコイドドーパミン作動性ニューロン。 機能的および規制的役割 精神薬理学で:進歩の第4世代、エドブルームFE、クッパーDJ(レイブン、ニューヨーク)、pp 283 - 294。
- ↵
- ルイロットA、
- ゴンザレスモラJL、
- グアダルーペT、
- マスM
(1991)雄性ラットの側坐核における性関連嗅覚刺激はドーパミン放出の選択的増加を誘導する 脳解像度 553:313-317。
- ↵
- マスM、
- ゴンザレスモラJL、
- ルイロットA、
- ソールC、
- グアダルーペT
(1990)in vivoボルタンメトリーによって証明されるように交尾雄ラットの側坐核におけるドーパミン放出の増加。 神経Lett 110:303-308。
- ↵
- マスM、
- フメロB、
- フェルナンデスヴェラJR、
- ゴンザレスモーラJL
(1995a)神経化学物質は、in vivoマイクロダイアリシスによって評価される性的疲労と回復の相関関係を示します。 脳解像度 675:13-19。
- ↵
- マスM、
- フメロB、
- ゴンザレスモーラJL
(1995b)社会的同性愛相互作用中の脳モノアミン神経伝達物質放出のボルタンメトリーおよび微量透析のモニタリング Behav Brain Res 71:69-79。
- ↵
- マスM、
- フメロB、
- ペレスロドリゲス1世
(1995c)性的に同腹のラットにおけるアポモルヒネによる交尾行動の誘導 Eur J Pharmacol 280:331-334。
- ↵
- マスM、
- フメロB、
- ペレスロドリゲス1世、
- ゴンザレスモーラJL
(1995d)性的満腹感の神経化学。 抑制欲求の実験モデル 性機能および機能障害の薬理学、ed Bancroft J(Raven、ニューヨーク)、pp 115 – 126。
- ↵
- Mirenowicz J、
- シュルツW
(1994)霊長類のドーパミン作動性ニューロンにおける報酬反応の予測不可能性の重要性。 J神経フィジオル 72:1024-1027。
- ↵
- ミッチェルJB、
- グラットンA
副嗅覚系の活性化により誘発される(1992)中辺縁系ドーパミン放出:高速クロノアンペロメトリー研究 神経Lett 140:81-84。
- ↵
- パキシノスG、
- ワトソンC
(1986)定位座標におけるラット脳(2およびed)。 (学術、サンディエゴ)。
- ↵
- Pfaus JG、
- ダンマG、
- ノミコスGG、
- ウェンクスターンD、
- Blaha CD、
- フィリップスAG、
- フィビガーHC
(1990)性的行動は雄ラットの中枢ドーパミン伝達を増強する 脳解像度 530:345-348。
- ↵
- Pfaus JG、
- エブリットBJ
(1995)性行動の精神薬理学。 精神薬理学で:進歩の第4世代、エドブルームFE、クッパーDJ(レイブン、ニューヨーク)、pp 743 - 758。
- ↵
- Pfaus JG、
- ゴルザルカBB
(1987)オピオイドと性行動。 Neurosci Biobehav Rev 11:1-34。
- ↵
- フィリップスAG、
- Blaha CD、
- Pfaus JG、
- ブラックバーンJR
(1992)ポジティブな感情状態の神経生物学的相関:ドーパミン、予想および報酬。 感情に関する研究の国際レビュー、ed Strongman(Wiley、New York)、pp 31 – 50。
- ↵
- Pleim ET、
- Matochik JA、
- バーフィールドRJ、
- アウエルバッハSB
(1990)側坐核におけるドーパミン放出とラットにおける男性の性行動との相関 脳解像度 524:160-163。
- ↵
- ロドリゲスマンゾG、
- フェルナンデス - グアスティA
(1994)セロトニン作動薬とノルアドレナリン作動薬による性的疲労の回復。 Behav Brain Res 62:127-134。
- ↵
- ロドリゲスマンゾG、
- フェルナンデス - グアスティA
(1995a)ヨヒンビン、ナロキソン、および8-OH-DPATによる性的消耗ラットの交尾行動の再確立における中枢ノルアドレナリン作動性システムの関与。 ブレインレスブル 38:399-404。
- ↵
- ロドリゲスマンゾG、
- フェルナンデス - グアスティA
(1995b)オピオイドきっ抗薬と性的飽食現象 精神薬局方 122:131-136。
- ↵
- ロールスロイスBJ
(1986)感覚特有の満腹感。 ニュートラルレビュ 44:93-101。
- ↵
- サラモンJD
動機づけの行動神経化学:側坐核ドーパミンの動的活性の研究における方法論的および概念的問題 J神経科学の方法 64:137-149。
- ↵
- 佐藤Y、
- 和田H、
- 堀田さん、
- スズキN、
- 渋谷A、
- あだちHさん、
- 加藤R、
- 塚本Tさん、
- 熊本Y
(1995)ラットの雄性交尾行動中の内側視索前野におけるドーパミン放出 脳解像度 692:66-70。
- ↵
- シュルツW
(1992)行動中の霊長類におけるドーパミンニューロンの活動。 Semin Neurosci 4:129-138。
- ↵
- 志村T、
- 山本Tさん、
- シモコチM
(1994)視索前内側領域は、雄ラットにおける性的覚醒とパフォーマンスの両方に関与している:自由に動く動物におけるニューロン活動の再評価。 脳解像度 640:215-222。
- ↵
- ウェンクスターンD、
- Pfaus JG、
- フィビガーHC
(1993)ドーパミン伝達は、性的に受容的な雌性ラットへの最初の曝露時に雄性ラットの側坐核で増加する。 脳解像度 618:41-46。
- ↵
- ウィルソンC、
- ノミコスGG、
- Collu M、
- フィビガーHC
動機づけられた行動の(1995)ドーパミン作動性相関:ドライブの重要性 J Neurosci 15:5169-5178。
- ↵
- ウィルソンJR、
- カーンRE、
- ビーチFA
(XNUMX)雌性刺激を変化させることにより生じた雄性ラットの性行動の変化。 J Comp Physiol精神医学 56:636-644。
- ↵
- ZetterströmT、
- シャープT、
- Ungerstedt U
(1984)覚醒ラットにおける線条体ドーパミン放出と代謝に対する神経遮断薬の効果の脳内透析による研究 Eur J Pharmacol 106:27-37。
この記事を引用している記事
-
腹側被蓋野におけるドーパミン作動性ニューロンの内因性オピオイド誘発神経可塑性は自然および鎮静剤報酬に影響する Journal of Neuroscience、25 6月2014、34(26):8825-8836
-
病的ギャンブルにおける異なる種類の報酬に対する感受性の不均衡 脳、1 8月2013、136(8):2527-2538
-
重要な仲介者としてのΔFosBを有する一般的な神経可塑性メカニズムに及ぼす天然および薬物報酬法 ジャーナルオブニューロサイエンス、20 2月2013、33(8):3434-3442
-
雄ラットの性行動中の側坐核殻における神経反応 ジャーナルオブニューロサイエンス、1 2月2012、32(5):1672-1686
-
ロマンチックな愛:配偶者選択のための哺乳類の脳システム 王立協会の哲学的取引B:生物学、29 12月2006、361(1476):2173-2186
-
脳の快楽のホットスポット 神経科学者、1 12月2006、12(6):500-511
-
ラット側坐核および内側前頭前皮質における満腹までの摂餌と相関するドーパミン作動性の中枢および側底扁桃核による調節 Journal of Neuroscience、15 12月2002、22(24):10958-10965
-
ドーパミン濃度一過性の頻度は同系の導入中に雄ラットの背側および腹側線条体で増加する Journal of Neuroscience、1 12月2002、22(23):10477-10486
-
書評:コカインの急性および慢性作用における細胞外ドーパミンの動態 神経科学者、1 8月2002、8(4):315-322
-
パブロフアプローチアプローチ行動の獲得および遂行における側坐核コアにおけるNMDA、AMPA / Kainate、およびドーパミン受容体の異なる関与 Journal of Neuroscience、1 12月2001、21(23):9471-9477
-
ラットにおける薬物関連刺激によるコカイン探索行動の制御:扁桃体および側坐核における消失したオペラント反応および細胞外ドーパミンレベルの回復に対する影響 PNAS、11 4月2000、97(8):4321-4326
-
D‐アンフェタミン誘発行動感作後の雄ラットの側坐核における性行動の促進とドーパミン流出の促進 Journal of Neuroscience、1 1月1999、19(1):456-463
-
神経科学:脳の注意を引く サイエンス、3 10月1997、278(5335):35-37
-
ラットの内側前頭前野および側坐核における感覚特異的満腹感のドーパミン作動性相関 Journal of Neuroscience、1 10月1999、19(19):RC29