Neurosci Biobehav Rev. 2011 Jan; 35 (3): 939-55. doi: 10.1016 / j.neubiorev.2010.10.014. Epub 2010 Nov 2.
- 1Bernard B. Brodie Neurologijos katedra, Cagliari universitetas, Cittadella Universitaria, 09042 Monserrato, CA, Italija. [apsaugotas el. paštu]
APRAŠYMAS
Oksitocinas yra stiprus varpos erekcijos induktorius, suleidžiamas į centrinę nervų sistemą. Žiurkėms patinams, jautriausias oksitocino efekto poveikio smegenų plotas yra hipotalamo paraventrikulinis branduolys. Šiame branduolyje ir aplinkiniuose regionuose yra visų oksitocinerginių neuronų ląstelių kūnai, išskiriantys į hipotalamines smegenų sritis ir nugaros smegenis. Ši peržiūra rodo, kad oksitocinas sukelia varpos erekciją, kai švirkščiamas kai kuriose iš šių sričių (pvz., Ventralinis tegmentalinis plotas, hippokampo ventralinis subiculumas, amygdalos ir torako-juosmens nugaros smegenų posteromedialinis žievės branduolys). Mikroinjekciniai tyrimai kartu su smegenų mikrodializės ir dvigubų imunofluorescencinių tyrimų duomenimis rodo, kad šiose srityse oksitocinas tiesiogiai arba netiesiogiai aktyvuoja (daugiausia per glutamo rūgštį) mezolimbines dopaminergines neuronas. Iš branduolio accumbens išsiskyręs dopaminas savo ruožtu suaktyvina neuroninius kelius, dėl kurių paraventriculiniame branduolyje aktyvuojasi hipertalaminiai dopaminerginiai neuronai. Tai aktyvuoja ne tik oksitocinerginius neuronus, nukreipiančius į stuburo smegenis ir tarpininkaujant varpos erekciją, bet ir tuos, kurie nukreipia į pirmiau minėtas hipotalamines sritis, tiesiogiai arba netiesiogiai (per glutamino rūgštį) mesolimbinių dopaminerginių neuronų aktyvumą, kontroliuojantį motyvaciją ir atlygį. Kartu šie nerviniai takai gali sudaryti sudėtingą hipotetinę grandinę, kuri vaidina ne tik seksualinės veiklos (erekcijos ir kopuliacijos) vartojimo etapą, bet ir motyvuojančius bei naudingus lytinio elgesio etapo aspektus.
1.Introduction
Pūslės erekcija yra vyrų lytinis atsakas, kuris vaidina svarbų vaidmenį žinduolių, įskaitant žmogų, reprodukcijai, ir tai taip pat galima pastebėti skirtingose situacijose nei tie, kurie griežtai susiję su reprodukcija. Priklausomai nuo konteksto, kuriame atsiranda varpos erekcija, jos reguliavime dalyvauja įvairūs centriniai ir periferiniai nervų ir (arba) humoraliniai mechanizmai (žr. Meisel ir Sachs, 1994; Argiolas ir Melis, 1995, 2004, 2005; Sachs, 2000, 2007; McKenna, 2000, Giuliano ir Rampin, 2000, 2004, Andersson, 2001, Melis ir Argiolas, 1995a, 2003, Hull ir kt., 2002). Tarp centrinių neurotransmiterių ir neuropeptidų, kurie kontroliuoja varpos erekciją, geriausiai žinoma dopamino, serotonino, eksitacinių aminorūgščių, azoto oksido, adrenokortikotropino, oksitocino ir opioidinių peptidų \ t. Jie gali palengvinti ar slopinti varpos erekciją, veikdami keliose smegenų srityse. ty medialinė preopticinė zona, hipotalamo paraventrikulinis branduolys, ventralinis tegmentalinis plotas, hipokampas, amygdala, stria terminalo lovos branduolys, branduolys accumbens, medulio oblongata ir nugaros smegenys. (1 lentelė) (žr. Meisel ir Sachs, 1994; Witt ir Insel, 1994; Stancampiano ir kt., 1994; Argiolas ir Melis, 1995, 2005; Argiolas, 1999; Bancila ir kt., 2002; Giuliano ir Rampin, 2000; McKenna, 2000, Andersson, 2001, Hull ir kt., 2002, Coolen ir kt., 2004).
Oksitocinas, neurohypofizės peptidas, gerai žinomas dėl savo hormoninio vaidmens laktacijos ir gimdymo metu, randamas moterims ir vyrams, ne tik neuronuose, turinčiuose ląstelių kūnus, esančius hipoglikemijos paraventrikuliniuose ir supraoptiniuose branduoliuose, o taip pat ir neuronuose. nuo paraventriculiarinio branduolio ir aplinkinių struktūrų iki extrahipaloalinių smegenų sričių (pvz., pertvaros, ventralinio tegmentalės, hipokampo, amygdalos, medulla oblongata ir nugaros smegenų). Manoma, kad šie neuronai dalyvauja daugelyje centrinių funkcijų, tokių kaip atmintis, mokymasis, affiliatyvus ir socialinis-seksualinis elgesys, įskaitant varpos erekciją ir kopuliacinį elgesį (žr. Buijs, 1978; Sofroniew, 1983; Argiolas ir Gessa, 1991; Pedersen et al. 1992; Carter, 1992; Wagner ir Clemens, 1993; Ivell ir Russel, 1995; Carter ir kt., 1997; Tang ir kt., 1998; Veronneau-Longueville ir kt., 1999; Iš tiesų, oksitocinas palengvina erekcijos funkciją ir vyrų seksualinį elgesį pelėms, žiurkėms, triušiams ir beždžionėms (žr. Argiolas ir Gessa, 1991; Carter, 1992; Pedersen ir kt., 1992; Argiolas ir Melis, 1995, 2004; Argiolas, 1999, 1987; Argiolas, 1987). Tai gali atsitikti ir žmonėms, nes oksitocino koncentraciją plazmoje didina seksualiniai stimulai, ypač ejakuliacijos metu (Carmichael ir kt., 1974; Murphy ir kt., XNUMX), taip pat manipuliuojant krūtimi ir lytiniais organais, kurie paprastai atsiranda seksualinės veiklos metu. lytinių santykių (Tindall, XNUMX).
Oksitocino palengvinantį poveikį vyrų seksualiniam elgesiui pirmiausia parodė intraveninio oksitocino gebėjimas sumažinti pirmosios ejakuliacijos vėlavimą ir lėtinti vyrų triušių išnaudojimą, susietą su imliomis moterimis (Melin ir Kihlstrom, 1963). Tačiau seksualinis oksitocino poveikis buvo galutinai pripažintas tik aštuntajame dešimtmetyje. Tada nustatyta, kad oksitocinas, gautas centralizuotai nanogramų kiekiu, gali sukelti varpos erekciją (Argiolas ir kt., 1985, 1986) ir patobulinti kopuliacinį elgesį (Arletti ir kt., 1985) žiurkėms ir padidinti lordozę žiurkių patelėms (Arletti ir Bertolini, 1985; Caldwell ir kt., 1986), matyt, veikiant gimdos tipo oksitocinerginiams receptoriams (žr. Argiolas ir Melis, 1995, 2004; Argiolas, 1999; Melis ir Argiolas, 2003; Oksitocinas gerina lytinį elgesį ne tik lytiniu būdu stipriai patinusioms žiurkėms (Arletti et al., 1985), bet ir vyresnio amžiaus patinams (Arletti ir kt., 1990), o vyraujantiems, bet ne pavaldiniams, vyriškiems voverės beždžionėms (Winslow ir Insel , 1991).
Oksitocino pro-erekcijos poveikis priklauso nuo testosterono, nes jis panaikinamas hipofizektomija ir kastracija, ir atkuriamas papildant kartu su testosteronu arba jo metabolitais, estradioliu ir 5_-dihidro-testosteronu kartu (Melis ir kt., 1994a). Labiausiai jautrus smegenų plotas, skirtas oksitocino varpos erekcijai, yra hipotalamo paraventrikulinis branduolys (Melis ir kt., 1986), iš kurių kyla visos hipotalaminės oksitocinerginės projekcijos (žr. Aukščiau). Čia nustatyta, kad oksitocinas gali sukelti varpos erekciją (ir žvėrioti), kai jis švirkščiamas tokiomis mažomis dozėmis kaip 3 pmol (žr. Skyrių 2.1 žemiau). Oksitocino sukeltos varpos erekcija taip pat, kai ji švirkščiama dvišaliai į hipokampo CA1 lauką, bet ne dorsaliniame subiculume (žr. Skyrių 2.3 žemiau), šoninis pertvaros, caudato branduolys, medialinis preopticinis plotas, hipotalamo ventromedinis branduolys ir supraopticinis branduolys (Melis ir kt., 1986). Dėl mechanizmo, kuriuo oksitocinas veikia paraventriculiniame branduolyje, kad sukeltų šį lytinį atsaką, daugelis tyrimų rodo, kad oksitocinas aktyvuoja savo neuronus. Pagal šią hipotezę seksualinė sąveika didina FOS, artimiausio geno c-fos geno produktą paraventrikuliniuose oksitocinerginiuose neuronuose, kurie išsikiša į nugaros smegenis, kurie yra susiję su varpos erekcijos kontrole (žr. Witt ir Insel, 1994 ir nuorodos jame) ir seksualinė impotencija (pvz., suaugusių vyrų žiurkių nesugebėjimas susikaupti su ovariektomizuota estrogeno-progesteroneprimedine moterimi) buvo susijęs su žiurkių žiurkėmis, kurių hipotalamo paraventrikuliniame branduolyje yra mažas oksitocino mRNR kiekis (Arletti et. al., 1997).
Šiuo metu nėra aišku, ar oksitocinas veikia lytinį elgesį, nei numatomasis etapas. Kadangi oksitocinas sukelia varpos erekciją, o pagrindinis oksitocino poveikis kopuliaciniam elgesiui yra žiurkių patinų post-ejakuliacijos intervalo sumažėjimas (Arletti ir kt., 1985), yra protinga manyti, kad peptidas pagerina seksualinę veiklą. Tačiau, kadangi oksitocinas taip pat didina socialinę ir seksualinę sąveiką (žr. Pedersen ir kt., 1992; Carter ir kt., 1997; Ivell ir Russel, 1995) ir oksitocino receptorių antagonistai užkerta kelią nekontaktinei erekcijai (Melis ir kt., 1999a), kurie yra laikomi seksualinio susijaudinimo rodikliu (žr. Sachs, 1997, 2000, 2007; Melis ir kt., 1998, 1999b ir jame esančios nuorodos), negalima atmesti galimo oksitocino vaidmens seksualinio susijaudinimo ir seksualinės motyvacijos.
Šioje apžvalgoje apibendrinami paskelbti ir nepaskelbti naujausių tyrimų rezultatai, kurie rodo, kad oksitocinas sukelia varpos erekciją ne tik tada, kai jis švirkščiamas į hipotalamo paraventriculinį branduolį, bet ir kitose papildomose hipotalaminėse smegenų srityse, pvz. (Melis ir kt., 2007, 2009a; Succu ir kt., 2008), tjis yra hippokampo ventralinis subiculumas ir užpakalinis amygdalos branduolys (Melis ir kt., 2009b, 2010), kurios yra svarbios limbinės sistemos sudedamosios dalys ir kurios, kaip manoma, vaidina pagrindinį vaidmenį motyvacijos ir atlygio procesuose. Šie tyrimai atskleidė, kad oksitocinas dalyvauja neuroninėse grandinėse, tarp jų ir kitų neurotransmiterių, pvz., Dopamino ir glutamo rūgšties, ir kitų smegenų sričių, išskyrus paraventriculinį branduolį, pvz., Ventralinė tegmentalinė sritis, branduolys accumbens, hipokampas ir sritys, kurios dar turi būti nustatyti. Šios grandinės gali tarpininkauti tarp mezolimbinės ir incerto-hipotalaminės dopaminerginės sistemos sąveikos ir vaidinti ne tik vyriškos lyties elgesio (pvz., Varpos erekcijos ir kopuliacijos), bet ir seksualinės motyvacijos bei seksualinės motyvacijos metu. susijaudinimas, tokiu būdu suteikiant neuroninį substratą, skirtą motyvacinėms ir naudingoms seksualinės veiklos savybėms paaiškinti.
2. Oksitocinas veikia varpos erekciją, veikdamas skirtingose smegenų srityse
2.1. Hipotalamies paraventrikulinis branduolys
Kaip priminta aukščiau, hipotalamo paraventrikulinis branduolys netrukus buvo nustatytas kaip smegenų sritis, jautriausia oksitocino pro erekcijos poveikiui. Vienu metu švirkščiant šį branduolį, oksitocinas buvo aktyvus tokiomis mažomis dozėmis kaip 3 ng (3 pmol) (Melis ir kt., 1986). Struktūrinio aktyvumo santykių tyrimai parodė, kad oksitocino sukeltą varpos erekciją tarpininkaujo uterinetype oksitocino receptoriai, susieti su Ca2 + įsiurbimu į oksitocinerginių neuronų ląstelių kūnus, išskleidžiančius į hipotalamines smegenų sritis, ir į azoto oksido sintezės aktyvavimą. Azoto oksidas savo ruožtu veikdamas kaip intracelulinis pasiuntinys su dar nežinomu mechanizmu (nesusijusiu su guanilato ciklaze) veda prie oksitocinerginių neuronų, kurie išsikiša į nugaros smegenis ir išorines hipotalamines smegenų sritis, aktyvavimo (1 pav.) (Žr. žemiau ir Argiolas ir Melis, 1995, 2004, 2005 ir jame esančios nuorodos). Oksitocino gebėjimas aktyvuoti savo neuronus
1. (MISSING) Scheminis oksitocinerginių neuronų, kurie yra kilę iš hipotalamo paraventrikulinio branduolio, schema ir projektavimas į hipotalamines smegenų sritis, tokias kaip nugaros smegenys, VTA, hipokampas, amygdala ir kt. dopaminas, eksitacinės aminorūgštys, pats oksitocinas, heksarelino analoginiai peptidai ir VGF-gauti peptidai sukelia varpos erekciją, kurią galima sumažinti ir (arba) panaikinti stimuliuojant GABAerginius, opioidinius ir kanabinoidinius CB1 receptorius. Oksitocinerginių neuronų aktyvavimas yra antrinis dėl šių neuronų oksidų sintezės aktyvinimo. Iš tiesų endogeninis azoto monoksidas, susidaręs stimuliuojant dopamino, eksitacinių aminorūgščių arba oksitocino receptorių arba egzogeninio azoto oksido, kaip gaunamas iš azoto oksido donorų, tiesiogiai į paraventriculinį branduolį, aktyvuoja oksitocinerginius neuronus dar nenustatytu mechanizmu, kuris, matyt, nėra susijęs su guanilato ciklazės stimuliacija. Tai savo ruožtu sukelia oksitocino išsiskyrimą nugaros smegenyse ir papildomose hipotalaminėse smegenų srityse. Išsami informacija apie mechanizmus, kuriais oksitocinas sukelia varpos erekciją, kai jie išleidžiami šiose srityse, pvz., VTA, ventralinis subiculumas ir amygdala, aprašyti atitinkamose smegenų srities dalyse. Čia oksitocinas veikia savo receptorius ir didina NO gamybą, o tai sukelia varpos erekciją, kaip nustatyta PVM. Tačiau, skirtingai nuo PVM, caudal VTA NO aktyvina guanilato ciklazę. Tai sukelia cGMP koncentracijos padidėjimą, kuris lemia mezolimbinių dopaminerginių neuronų aktyvumą ir varpos erekciją. VS NO aktyvina glutamaterginius neuronus, kurie išsikiša į hipokampo sritis, įskaitant VTA. VTA glutamo rūgštis savo ruožtu aktyvuoja mezolimbines dopaminergines neuronas, kaip nustatyta oksitocinu. Mechanizmai, panašūs į pirmiau aprašytus mechanizmus, gali veikti ir tada, kai varpos erekcija vyksta fiziologiniame kontekste, būtent tada, kai žiurkių žiurkės patenka į neprieinamą moterį (pvz., Nekontaktinę erekciją) arba koluliacijos metu.
paraventriculiniame branduolyje buvo patvirtinti tyrimai, rodantys, kad: (1) oksitocino receptoriai yra šiame hipotalaminiame branduolyje (Freund-Mercier ir kt., 1987; Freund-Mercier ir Stoeckel, 1995); (2) oksitocinas palengvina jo išsiskyrimą in vitro ir in vivo (Freund-Mercier ir Richard, 1981, 1984; Moos ir kt., 1984); ir (3) oksitocinas sužadina savo neuronus, veikdamas paraventrikuliniame branduolyje (Yamashita ir kt., 1987). Be to, hipotalamo paraventrikuliniame ir supraoptiniame branduolyje taip pat buvo nustatyti oksitocinerginiai sinapsijos, sukeliančios magnoceliulinių oksitocinerginių neuronų ląstelių kūnus. (Theodosis, 1985). Galiausiai, centrinių oksitocinerginių neuronų naikinimas elektrolitiniais arba cheminiais paraventriculinio branduolio pažeidimais, kurie visiškai naikina oksitocino kiekį per centrinę nervų sistemą ir nugaros smegenis, panaikina ne tik eritilinį oksitocino poveikį, bet ir mažina vaistų sukeltą poveikį. varpos erekcija ir nekontaktinė erekcija (žr. toliau ir Argiolas ir kt., 1987a, b; Liu ir kt., 1997 ir nuorodos jame). Su stipriais ir selektyviais oksitocino receptorių antagonistais randami panašūs rezultatai, lyginant su paraventriculinio branduolio pažeidimais. Iš tiesų, šie junginiai, įšvirkšti į paraventriculinį branduolį nanogramų kiekiais, neleido visiškai oksitocino sukeltai varpos erekcijai, o kai į šoninius skilvelius buvo užkirstas kelias ne tik pačios oksitocino sukeltai varpos erekcijai, bet ir vaistų sukeltai varpos erekcijai (žr. Toliau 3 skyrių) ir Argiolas ir Melis, 1995, 2004, 2005 ir nuorodos jame) ir nekontaktinės erekcijos (Melis ir kt., 1999a), be to, buvo labai veiksmingos mažinant lytiniu būdu stipriai veikiančių žiurkių kopuliacinį elgesį (Argiolas ir kt., 1988). Be to, lytinė sąveika didina FOS, artimiausio ankstyvojo geno c-fos geno produktą paraventrikuliniuose oksitocinerginiuose neuronuose, kurie išsikiša į nugaros smegenis, dalyvaujančius kontroliuojant varpos erekciją (žr. Witt ir Insel, 1994 ir nuorodos jame). Galiausiai, seksualinė impotencija (pvz., suaugusių patinų žiurkių nesugebėjimas susikaupti su estrogeno-progesteroneprimediniu imituojančia moterimi) hkaip ir vyrų žiurkėms, turinčioms mažą oksitocino mRNR kiekį ir azoto oksido sintazę hipotalamo paraventrikuliniame branduolyje. (Benelli ir kt., 1995; Arletti ir kt., 1997) (išsamioms šių tyrimų apžvalgoms žr. Argiolas, 1999; Argiolas ir Melis, 2004, 2005).
2.2. Ventralinė tegmentalinė sritis
Ventralinė tegmentalinė zona buvo aptikta tik kaip smegenų vieta, kurioje oksitocinas sukelia varpos erekciją. Šioje srityje yra oksitocinerginių nervų galų, kilusių iš paraventrikulinių branduolių ir oksitocino receptorių (Freund-Mercier ir kt., 1987; Vaccari ir kt., 1998). Tiksliau sakant, oksitocinas galėjo ipailginant varpos erekciją, kai jis švirkščiamas vienašališkai į caudalį, bet ne rostralinio ventralinio tegmentalio zonoje priklausomai nuo dozės (Melis ir kt., 2007). Aktyviosios dozės buvo didesnės už tas, kurios reikalingos injekuojant į paraventriculinį branduolį ir panašios į tuos, kurie sukelia varpos erekciją, švirkščiant į hippokampo ventralinį subiculumą arba į amygdalos posteromedialinį žievės branduolį (žr. Toliau). Akivaizdu, kad pro-erekcijos efektą skatina mezolimbinių dopaminerginių neuronų, nukreiptų į branduolio accumbens korpusą, aktyvinimas, kuris savo ruožtu aktyvuoja dar nežinomus neuroninius kelius, nukreipiančius į inkaverio hipotalaminius dopaminerginius neuronus, sukeliančius paraventricularis oksitocinerginius neuronus, tarpininkaujančius varpos erekciją (Melis ir kt., 2007, 2009a).
Kiek tai susiję su mechanizmu, kuriuo oksitocinas aktyvuoja dopaminerginį neurotransmisiją ventralinio apvalkalo srityje, turimi duomenys rodo, kad Oksitocinas stimuliuoja oksitocinerginius receptorius, esančius ląstelių, turinčių mezolimbinių dopaminerginių neuronų. Tai padidina Ca2 + antplūdį dopaminerginių neuronų ląstelių kūnuose, tokiu būdu aktyvindamas azoto oksido sintezę (Succu ir kt., 2008). Skirtingai nuo paraventriculinio branduolio (žr. Skyrių 3 žemiau), azoto monoksidas savo ruožtu aktyvuoja guanilato ciklazę, taigi didina ciklinės GMP koncentraciją. Pagal šį mechanizmą, d (CH2) 5Tyr (Me) 2-Orn8-vazotocinas, stiprus oksitocino antagonistas arba S-metil-tio-l-citrulinas, stiprus neuronų azoto oksido sintezės inhibitorius, švirkščiamas į caudalinė ventralinė tegmentalinė zona prieš oksitociną, panaikinta varpos erekcija ir padidėjęs ląstelių dopamino koncentracijos padidėjimas oksitocino sukeltame branduolio accumbens lukšte. Be to, 8-bromciklinis GMP, aktyvus fosfodiesterazei atsparus ciklinis GMP analogas, sukelia varpos erekciją, kai jis švirkščiamas į caudalinį ventralinį tegmentalą, ir padidina nešūnų dopamino koncentraciją branduolio accumbens lukšte, kaip nustatyta oksitocino injekcijose. Caudal ventralinis tegmental area (Succu ir kt., 2008; Melis ir kt., 2009a) (taip pat žr. 2).
Visada, laikantis šio mechanizmo, haloperidolis, stiprus dopamino D2 receptorių antagonistas, švirkščiamas į branduolio akmenų apvalkalą, sumažina penitų erekciją, kurią sukelia oksitocinas, švirkščiantis į ventralinį tegmentalą (Melis ir kt., 2007). Pirmiau minėtą mechanizmą taip pat palaiko dvigubi imunofluorescencijos tyrimai, rodantys, kad oksitocino pluoštai paveikia dopaminerginių neuronų ląstelių kūninės ventralinės tegmentacijos srityje, kurie anksčiau buvo paženklinti retrograciniu žymekliu Fluorogold, švirkščiant į branduolio akumbensą ( Melis ir kt., 2007; Succu ir kt., 2008). Šių dopaminerginių neuronų ir dopamino receptorių aktyvavimas branduolyje accumbens savo ruožtu lemia dar nenustatytų nervų takų aktyvavimą, kuris stimuliuoja incerto-hipotalaminius dopaminerginius neuronus išleisti dopaminą paraventrikuliniame branduolyje, tokiu būdu aktyvuojant oksitocinerginius neuronus, nukreipiančius į stuburo smegenys ir tarpininkaujanti varpos erekcija (žr. aukščiau ir Melis ir kt., 2007; Succu ir kt., 2007, 2008). Iš tiesų, oksitocinas, švirkščiantis į caudalinį ventralinį tegmentalą, dozę, sukeliančią varpos erekciją, padidėjusi išorinė dopamino koncentracija dializate, gaunama ne tik iš branduolių accumbens, bet ir iš paraventrikulinės branduolio (Succu ir kt., 2007).
2.3. Hipokampas
Hippokampo CA1 laukas buvo kitas smegenų plotas, turintis daug oksitocinerginių pluoštų ir ankstesniais tyrimais nustatyti receptoriai, kuriuose oksitocino sukelta varpos erekcija (žr. Bujis, 1978; Sofroniew, 1983). Tačiau, skirtingai nuo paraventriculinio branduolio, čia nustatyta, kad oksitocinas gali sukelti varpos erekciją tik tada, kai jis švirkščiamas dvišaliu būdu ir didesnes dozes, nei buvo nustatyta paraventriculiniame branduolyje (Melis ir kt., 1986; Chen ir kt., 1992). Šiuose ankstesniuose tyrimuose oksitocino injekcijos į subiculumą buvo neaktyvios. Tačiau naujausi ir kruopščiai atliekami mikroinjekcijos tyrimai leido nustatyti a ventralinio subiculumo regionas kai oksitocino injekcija buvo pajėgi sukelti varpos erekciją priklausomai nuo dozės (Melis ir kt., 2009b). Šiam smegenų plotui įšvirkšto oksitocino pro erekcinis poveikis buvo stebimas, kai dozės buvo panašios į tas, kurios buvo aktyvios ventralinio tegmentalio srityje po vienašalės injekcijos (Melis ir kt., 2007), kaip nustatyta paraventrikuliniame branduolyje. Matyt, oksitocinas, švirkščiantis į ventralinę subiculumą, sukelia varpos erekciją aktyvuodamas oksitocinerginius receptorius neuronuose, turinčiuose azoto oksido sintezę, todėl padidėja azoto oksido gamyba. Azoto oksidas, veikdamas kaip tarpląstelinis pasiuntinys, suaktyvina glutamo rūgšties neurotransmisiją, kuri sukelia varpos erekciją, galbūt per nervinio (glutamaterginio) efferentines projekcijas iš ventralinio subiculo papildomų hipokampo smegenų zonų, moduliuojančių mezolimbinių dopaminerginių neuronų aktyvumą (pvz., ventralinį tegmentalinį plotą, prefrontalinę žievę, paraventriculinį branduolį) (žr. toliau ir Melis, 2007, 2009b; Succu ir kt., 2008).
Šį veikimo mechanizmą patvirtina vidiniai smegenų mikrodializės eksperimentai, rodantys, kad oksitocinas, švirkščiantis į ventralinę subiculumą dozėmis, kurios sukelia varpos erekciją, padidina azoto oksido gamybą ir ekstraląstelinį glutamo rūgšties koncentraciją dializate iš ventralinio subiculumo (Melis et al. , 2010) ir ekstraląstelinė dopamino koncentracija branduolyje accumbens (Melis ir kt., 2007). Šiuos atsakymus priešinosi ne tik oksitocino receptorių antagonistas d (CH2) 5Tyr (Me) 2-Orn8-vazotocinas, bet ir neuroninio azoto oksido sintezės inhibitorius S-metil-tio-litritrinas ir azoto oksido šalinimo hemoglobinas. į ventralinę subiculumą keletą minučių prieš oksitociną (Melis ir kt., 2010).
Be to, atsižvelgiant į šį veikimo mechanizmą, gliutamaterginio neurotransmisijos aktyvavimas NMDA, įšvirkštus į ventralinę subiculumą, sukelia varpos erekciją (Melis ir kt., 2010). Šiuo metu nežinomas ventralinio subiculumo efferentinių projekcijų fenotipas, dėl kurio atsiranda mezolimbinių dopaminerginių neuronų aktyvacija ir padidėjęs ląstelių dopamino kiekis branduolyje accumbens. Tačiau, kadangi varpos erekcija, kurią sukelia oksitocinas, švirkščiantis į ventralinę subiculumą, atsiranda kartu su padidėjusiu gliuzamino rūgšties padidėjimu dializate nuo ventralinio tegmentalio srities, bet ne iš branduolio accumbens ir yra antagonizuotas (+) MK-801, potencialus nekonkurencinis NMDA subtieto (Woodruff ir kt., 1987) eksitacinių aminorūgščių receptorių antagonistas, švirkščiamas į ventralinę tegmentalinę sritį, bet ne į branduolį accumbens (žr. 2 ir Melis ir kt., 2009b) , tikėtina, kad šios projekcijos sukelia glutamaterginės neurotransmisijos aktyvavimą ventralinio tegmentalo srityje, kuri savo ruožtu aktyvuoja mesolimbinius dopaminerginius neuronus, nukreipiančius į branduolį accumbens. Šiuo metu nežinoma, ar padidėjusi glutamino rūgšties koncentracija, atsiradusi ventralinio tegmentalioje zonoje po oksitocino injekcijos į ventralinę subiculumą, iš neuronų, kilusių iš subikuliato ar kitų smegenų sričių (pvz., Prefrontalinė žievė). Nepaisant to, tai sukelia mezolimbinių dopaminerginių neuronų aktyvaciją ir padidėjusį dopamino išsiskyrimą į branduolį.. Čia dopamino receptorių aktyvacija sukelia hipertalaminių dopaminerginių neuronų, kurie atleidžia dopaminą paraventrikuliniame branduolyje, aktyvavimą, tokiu būdu aktyvindami oksitocinerginius neuronus, nukreipiančius į nugaros smegenis ir tarpininkaujant varpos erekcijai (žr. aukščiau ir Melis ir kt., 2007, 2009a; Succu ir kt., 2008).
2.4. Amygdala
Amygdala yra dar viena smegenų sritis, turinti daug oksitocino pluoštų ir receptorių (žr. Freundą-Mercierį ir kt., 1987; Vaccari ir kt., 1998; Uhl-Bronner ir kt., 2005). Manoma, kad oksitocinas dalyvauja skirtingose funkcijose: nuo anksiolizės, socialinės atminties ir pažinimo, socialiai sustiprinto mokymosi, emocinės empatijos, emocinio veido apdorojimo ir baimės dėl erekcijos ir seksualinio elgesio (žr. Kondo ir kt., 1998; Dominguez et aln., 2001; Ebner ir kt., 2005; Huber ir kt., 2005; Domes ir kt., 2007; Petrovic ir kt., 2008; Lee ir kt., 2009; Donaldson ir Young, 2009; Hurlemann ir kt. , 2010). Tačiau oksitocino gebėjimas sukelti vyriškos lyties žiurkių erekciją, atsiradusį amigdalos posteromedialiniame žievės branduolyje, buvo aptiktas tik neseniai (Melis ir kt., 2009b). Šis atsakas tuo pačiu metu pasireiškė padidėjus išorinei dopamino koncentracijai dializate, gautame iš branduolio accumbens lukšto, kaip nustatyta po oksitocino injekcijos į ventralinę subiculumą (Melis ir kt., 2009b). Šiuo metu nežinomas mechanizmas, kuriuo oksitocinas švirkščiamas į amygdalos posteromedialinį žievės branduolį, todėl yra nežinoma penio erekcija. Turimi duomenys rodo, kad tiek varpos erekcija, tiek ir padidėjusi ląstelių dopamino koncentracija dializate, gautame iš branduolio accumbens, yra oksitocinerginių receptorių aktyvavimo, nes abu atsakai buvo panaikinti oksitocino receptorių antagonistu d (CH2) 5Tyr ( Me) 2-Orn8-vazotocinas, švirkščiamas į amygdalos branduolį prieš kelias minutes prieš oksitociną (Melis ir kt., 2009b).
Nepriklausomai nuo mechanizmo oksitocino amygdalos posteromedialinio žievės branduolio, seksualinis atsakas, kurį sukelia peptidas, panaikinamas blokuojant visus dopaminerginius receptorius, turinčius cis-flupentiksolį, švirkščiant į branduolio accumbens korpusą ir blokuojant NMDA receptorius su: +) MK-801, švirkščiantis į ventralinį tegmentalinį plotą, bet ne į branduolį, kaip nustatyta penitos erekcijai, sukeltai oksitocino, injekuoto į ventralinę subiculumą (Melis ir kt., 2009b). Tai rodo, kad oksitocinas, švirkščiantis į amygdalos posteromedialinę branduolį, aktyvina glutamo rūgšties neurotransmisiją ventralinėje apykaitinėje srityje. Tai savo ruožtu sukelia mezolimbinių dopaminerginių neuronų aktyvaciją, sukeldama varpos erekciją. Atsižvelgiant į tyrimus, rodančius neuroninius kelius, kurie sujungia šį amygdalos branduolį su ventraliniu subiculumu (Canteras ir kt., 1995; prancūzų ir Totterdell, 2003), šie rezultatai rodo, kad tarp šių dviejų smegenų sričių gali egzistuoti sąveika, nors tai yra tiesioginė buvo aprašyti keliai iš amygdalos arba į branduolį, arba į ventralinį tegmentalą (Kelley ir Domesick, 1982; Witter, 2006).
2.5. Nugaros smegenys
Nugaros smegenys yra kita centrinės nervų sistemos sritis, kurioje yra oksitocinerginių pluoštų ir receptorių (Freund-Mercier ir kt., 1987; Uhl-Bronner ir kt., 2005), kuriuose oksitocinas sukelia varpos erekciją (Tang et al., 1998 Veronneau-Longueville ir kt., 1999, Giuliano ir Rampin, 2000, Giuliano ir kt., 2001). Kaip minėta aukščiau, šie oksitocininiai pluoštai yra kilę iš hipoglikamo paraventrikulinio branduolio ir prisideda prie mažėjančių takų, kontroliuojančių stuburo autonominius neuronus, tarpininkaujančius varpos erekciją. Iš tiesų šie pluoštai sintetinius kontaktus dorinio rago preganglioninėse simpatinėse ir parazimpatinėse ląstelių kolonose torako-juosmens ir lumbos sakraliniame takte su nugaros neuronais, įkvepiančiais varpos ertmę (Marson ir McKenna, 1996; Giuliano ir Rampin, 2000; Giuliano et al. , 2001). Šiuos sinapinius kontaktus parodė varpų kilmės stuburo neuronai ir pasiekė stuburo smegenis specifiniais retrogradiniais žymenimis, įšvirkštais į cavernous korpusus, kartu su dvigubais imunofluorescenciniais ir konfokaliniais lazerio mikroskopijos tyrimais (Tang et al., 1998; Veronneau-Longueville et al., 1999). Remiantis šiais tyrimais, anestezuoto žiurkių žiurkių intratekaliniu būdu injekcijos oksitocino kumuliacinės dozės lumbo sakralinėje, bet ne torako-juosmens lygyje, sukėlė intrakaverninį spaudimą priklausomai nuo dozės. Šį poveikį panaikino oksitocinerginio receptoriaus blokavimas d (CH2) 5Tyr (Me) 2-Orn8-vazotocinu ir dubens nervų sekcija (Giuliano ir Rampin, 2000; Giuliano et al., 2001). Šie rezultatai rodo, kad oksitocinas, veikiantis lumbos-sakralinio stuburo smegenų, padidina intrakaverninį spaudimą ir rodo, kad oksitocinas, išsiskiriantis fiziologiniu paraventrikulinio branduolio aktyvavimu, yra stiprus spinalinės erekcijos neuronų aktyvatorius, išsiskleidęs į cavernous korpusą. Įdomu tai, kad šie pro-erekcijos spinaliniai neuronai, dėl kurių oksitocinas veikia savo erekcinį poveikį, taip pat gauna sinaptinius kontaktus iš serotoninerginių neuronų, kilusių iš branduolio paragigantocellularis retikulinio formavimosi (Marson ir McKenna, 1992; Tang et al. ., 1998). Šių serotoninerginių neuronų naikinimas palengvina ejakuliaciją ir varpos refleksus žiurkėms (Marson ir McKenna, 1992; Yells et al., 1992). Kadangi vaistai, stimuliuojanti 5HT2C receptorius, palengvina varpos erekciją, kai vartojami intracerebroventrikuliniai, bet ne į paraventriculinį branduolį, ir vaistai, kurie blokuoja 5HT2C receptorius, taip pat mažina dopamino agonistą ir oksitocino sukeltą varpos erekciją, o dopamino antagonistai nesumažina 5HT2C agonisto sukeltos varpos erekcijos ( žr. Stancampiano ir kt., 1994 ir jame esančias nuorodas), taip pat buvo pasiūlyta, kad oksitocinas palengvina pro-erekcijos 5HT2C receptorių veikimą stuburo nugaros smegenų smegenų lygyje (Stancampiano et al., 1994). Arba oksitocinas gali turėti įtakos stuburo mažėjimo serotoninerginių neuronų aktyvumui, veikdamas tiesiogiai branduolyje paragigantocellularis, kur šie neuronai yra kilę (žr. Stancampiano et al., 1994).
3. Oksitocino, dopamino ir glutamo rūgšties sąveika centrinėje nervų sistemoje ir varpos erekcija
Kaip priminta 1 skyriuje, visi centrinėje nervų sistemoje esantys oksitocinerginiai neuronai yra kilę iš paraventrikulinių branduolių ir aplinkinių struktūrų. Šių neuronų aktyvumas yra kontroliuojamas įvairių neurotransmiterių ir / arba neuropeptidų. Tarp labiausiai ištirtų paraventrikulinio lygio yra dopaminas, glutamo rūgštis, gama-aminovo rūgštis (GABA), azoto oksidas, endokannabinoidai, opioidiniai peptidai, augimo hormono atpalaiduojantys peptidai, su VGF susiję peptidai ir oksitocinas. Dopamino, glutamo rūgšties, augimo hormono atpalaiduojančių peptidų, VGF gautų peptidų ir oksitocino stimuliatoriai, pvz., Šie junginiai ir jų agonistai palengvina varpos erekciją, kai jie švirkščiami į paraventriculinį branduolį, o GABA, opioidiniai peptidai ir endokannabinoidai yra slopinami, pvz. arba jų agonistai slopina varpos erekciją (žr. Meisel ir Sachs, 1994; Witt ir Insel, 1994; Argiolas ir Melis, 1995, 2004, 2005; Giuliano ir Rampin, 2000, 2004; McKenna, 2000; Andersson, 2001; Hull ir kt., 2002;
Keletas eksperimentinių įrodymų linijų rodo, kad šie oksitocinerginiai neuronai ir pirmiau minėti neurotransmiteriai bei neuropeptidai yra susiję su erekcijos funkcijos ir seksualinės elgsenos kontrole įvairiose fiziologinėse situacijose. Be to, oksitocinas, išsiskyręs į hipotalamines smegenų sritis, tokias kaip ventralinis tegmentalinis plotas, hipokampas ir jo regionai, amygdala ir nugaros smegenys, gali paveikti tų neuronų, kuriems kyla oksitocinerginių sinapsų, aktyvumą. Šiuo metu vieninteliai neuronai, svarbūs varpos erekcijai, dėl kurių oksitocinerginiai sinapsai sukelia tikrumą, yra caudalinio ventralio tegmentalinės srities mezolimbinių dopaminerginių neuronų ląstelių kūnai, išsikišantys į branduolio akumbensą (Melis ir kt., 2007 Succu ir kt., 2008) ir pro-erekcijos spinaliniai neuronai, išsikišę iš lumbos-sakralinio trakto į urvas (žr. Giuliano ir Rampin, 2000; Giuliano ir kt., 2001) (taip pat žr. 2.2 ir 2.5 skyriai) . Iš tiesų, nors oksitocinerginiai sinapsijos ir receptoriai taip pat buvo nustatyti ventraliniame subiculume, amygdaloje ir nugaros smegenyse, visose srityse, kurios yra svarbios varpos erekcijai (žr. Aukščiau), šiose srityse neuronų transmiterių tipas yra neuronuose Oksitocinerginių nervų galūnės dar nesuprantamos.
Šiame peržiūros skyriuje trumpai apibendrinama naujausia literatūra apie mechanizmus, kuriais remiamasi oksitocino pro-erekcijos poveikiu, suleistu į caudalinį ventralinį tegmentalą, hippocampus ventralinį subiculum ir stuburo smegenis. Ypatingas dėmesys skiriamas peptido ir dopamino bei glutamo rūgšties sąveikai šiose srityse ir dėl šios sąveikos vaidmens centrinėje erekcijos funkcijos kontrolės. Pirmiausia pateikiamas trumpas dopamino ir glutamino rūgšties poveikio paraventriculinio branduolio oksitocinerginiams neuronams, kurie taip pat vaidina svarbų vaidmenį erekcijos funkcijoje, santrauka, kad skaitytojas žinotų apie ankstyvą šios srities mokslinių tyrimų būklę, kaip šie tyrimai jau buvo išsamiai peržiūrimi (žr. Argiolas ir Melis, 1995, 2004, 2005; Melis ir Argiolas, 2003). Be to, šiuo atveju ypatingas dėmesys skiriamas naujausiems rezultatams, kurie rodo, kad dopamino ir oksitocino ryšys ir glutamo rūgšties-oksitocino ryšys yra svarbūs ne tik seksualinio pasirodymo (varpos erekcijos ir kopuliacijos), bet ir seksualinės veiklos metu. susijaudinimas ir seksualinė motyvacija.
3.1. Dopamino ir oksitocino sąveika paraventrikuliniame branduolyje
Šios dopamino agonistų gebėjimas sukelti varpos erekciją aktyvuojant centrinius oksitocinerginius neuronus iš karto po atradimo buvo pasiūlyta, kad apomorfinas sukelia varpos erekciją, kai švirkščiamas į paraventriculinį branduolį (Melis ir kt., 1987), kai buvo nustatyta oksitocino receptorių antagonisto d (CH2) 5Tyr (Me) 2-Orn8-vazotocino injekcija intracerebroventrikuliniu būdu (icv). gali sumažinti beveik visiškai varpos erekciją, kurią sukelia ne tik oksitocinas, vartojamas icv, bet ir apomorfino, skirto po oda (Argiolas ir kt., 1987b). Šiems rezultatams sekė kiti tyrimai, kurie parodė panašius rezultatus, kai d (CH2) 5Tyr (Me) 2-Orn8-vazotocinas buvo skiriamas icv ir apomorfinas buvo įvedamas tiesiai į paraventrikulinį branduolį (Melis ir kt., 1989b), lmanydamas, kad dopamino agonistai sukelia varpos erekciją, aktyvuodami paraventrikulinius oksitocinerginius neuronus, kurie išsikiša į hipotalamines smegenų sritis ir ypač į nugaros smegenis. (žr. Argiolas ir Melis, 1995, 2004, 2005). Atsižvelgiant į šią hipotezę, anestezuotose žiurkėse neseniai nustatyta, kad nehepeptidinio oksitocino receptoriaus antagonisto lumbokromatinių oksitocinerginių receptorių blokada gali panaikinti apomorfino sukeltą dopamino agonisto apomorfino sukelto intrakaverninio spaudimo padidėjimą, suteikiant įrodymų, kad paraventriculo-spinalinis oksitocinerginis kelias, susijęs su varpos erekcija (Baskerville ir kt., 2009).
Tyrimai, skirti. \ T Dopamino receptoriaus, atsakingo už varpos erekcijos indukciją, nustatymas parodė, kad dopamino receptorių agonistai taip pat sukelia varpos erekciją, veikdami ant D2 šeimos dopamino receptorių., kaip nustatyta sistemiškai dopamino receptorių agonistams (žr. Melis ir kt., 1987; Eaton ir kt., 1991; Melis ir Argiolas, 1995a). Atitinkamai apomorfinas, stiprus mišrus D1 / D2 receptorių agonistas ir chinpirolis, stiprus selektyvus D2receptoriaus agonistas, bet ne SKF 38393, selektyvus D1 receptorių agonistas, švirkščiamas į šią hipotalaminę branduolį, nustatė, kad gali sukelti varpos erekciją priklausomai nuo dozės. ir šių D2 receptorių agonistų sukeltas seksualinis atsakas buvo panaikintas D2 receptorių antagonistų, pvz., haloperidolio ir l-sulpirido, bet ne SCH 23390, selektyvaus D1 receptoriaus antagonisto (Melis ir kt., 1987). Apomorfino gebėjimą indukuoti varpos erekciją, švirkščiant į paraventriculinį branduolį, taip pat patvirtino telemetriniai tyrimai, rodantys, kad dopamino agonistas, įvedamas į paraventriculinį branduolį, gali padidinti intrakaverninį spaudimą žindančiose žiurkėms, nekeičiant sisteminio kraujospūdžio (Chen et al. , 1999; Giuliano ir Allard, 2001), kaip nustatyta po sisteminės injekcijos (Bernabè ir kt., 1999). Šie tyrimai taip pat patvirtino pagrindinį D2 receptorių vaidmenį, nes nustatyta, kad D1 receptorių agonistai negali padidinti intrakaverninio spaudimo, kai jie švirkščiami į paraventrikulinį branduolį. (Chen ir kt., 1999).
Tada buvo prieinamos kelios eksperimentinių įrodymų eilutės, rodančios, kad paraventrikuliniai D2 receptoriai, kurių stimuliacija sukelia varpos erekciją, yra oksitocinerginių neuronų ląstelių kūnuose. Pirma, paraventriculiniame branduolyje yra dopaminerginių nervų terminalų, kurie priklauso vadinamiesiems incertohypothalamic dopaminerginiams neuronams. Šių neuronų ląstelių organai yra A13 ir A14 grupėje Dahlstrom ir Fuxe (1964), plačiai ir innervatuoti keletą hipotalaminių struktūrų, įskaitant paraventriculines oksitocinergines neuronas, nukreiptas į neurohypofizę ir (arba) papildomas hipotalamines smegenų sritis (Buijs ir kt., 1984; Lindvall ir kt., 1984).
Šių dopaminerginių neuronų dalyvavimas paraventrikuliniame lygyje kontroliuojant varpos erekciją ir kopuliaciją yra pagrįstas mikrodializės tyrimais, rodančiais, kad dializate, gautame iš dializato, padidėja ekstraląstelinio dopamino ir 3,4-dihidroksifenilacto rūgšties (DOPAC) koncentracija. seksualiai stipriai patinų žiurkių paraventriculinis branduolys, rodantis nekontaktinę erekciją, kai jis buvo patekęs į neprieinamą ovariektomizuotą estrogeno + progesterono imituojančią moterį (Melis ir kt., 2003).
Dopamino ir DOPAC koncentracijos padidėjimas buvo dar didesnis, kai buvo leidžiama kopuliacija su imliąja moterimi (Melis ir kt., 2003), kaip nustatyta medialinėje preopticinėje zonoje (Hull ir kt., 1995) ir branduolyje accumbens (Pfaus). ir Everitt, 1995). Antra, keletas tyrimų parodė, kad varpos erekcija, kurią sukelia paraventrikulinių D2 receptorių stimuliacija, yra tarpininkauja šiuose rajonuose išsiskyręs oksitocinas. Atitinkamai, apomorfinas, vartojamas dozėmis, kurios sukelia varpos erekciją, buvo pajėgi padidinti oksitocino koncentraciją, ne tik žiurkių ir beždžionių plazmoje (Melis ir kt., 1989a; Cameron ir kt., 1992), bet ir papildomoje hipotalaminėje smegenyse sritis, pvz., hipokampus (Melis ir kt., 1990). Remiantis šiais rezultatais, neseniai buvo įrodyta, kad apomorfinas, švirkščiantis į paraventriculinį branduolį, sukelia varpos erekciją, gali padidinti ir išorinę dopamino koncentraciją branduolyje, nes tai sumažina oksitocino receptorių antagonisto d (CH2). 5Tyr (Me) 2-Orn8-vazotocinas, suleidžiamas į ventralinį tegmentalą (Succu ir kt., 2007; Melis ir kt., 2009a) (taip pat žr. 4 skyrių). Trečia, dvišaliai paraventriculinio branduolio elektrolitiniai pažeidimai, kurie beveik visiškai pašalina oksitociną iš papildomų hipotalaminių smegenų sričių (Hawthorn ir kt., 1985), panaikina apomorfino sukeltą varpos erekciją (Argiolas ir kt., 1987a) ir selektyvius oksitocino receptorių antagonistus į šoninius skilvelius, bet ne į paraventriculinį branduolį, mažina nuo dozės priklausomą apomorfino sukeltą varpos erekciją, kuri yra lygiagreti šių junginių veiksmingumui blokuojant oksitocino receptorius (Melis ir kt., 1989b). Oksitocino receptorių antagonistai taip pat labai stipriai padeda sumažinti ne tik oksitocino, bet ir apomorfino (Argiolas ir kt., 1988, 1989) sukeltą vyrų lytinį elgesį.
Dėl mechanizmo, kuriuo D2 receptoriai aktyvuojami dopamino arba dopamino receptorių agonistų, didina oksitocinerginių neuronų aktyvumą, tokiu būdu atleidžiant oksitociną ekstremipotalaminėse smegenų srityse ir stuburo smegenyse, daug eksperimentinių duomenų.remti hipotezę, kad D2 receptorių stimuliavimas didina ląstelių ląstelių Ca2 + jonų koncentraciją oksitocinerginių neuronų ląstelių kūnuose, o tai lemia azoto oksido sintezės aktyvaciją, Ca2 + -kalmodulino priklausomas fermentas, kuris yra šiuose ląstelių kūnuose (Vincent ir Kimura, 1992; Torres ir kt., 1993; Sanchez ir kt., 1994; Sato-Suzuki ir kt., 1998). Padidėjusi azoto oksido gamyba sukelia oksitocinerginių neuronų aktyvaciją. Atitinkamai (1) apomorfino sukeltos varpos erekcija buvo užkirstas kelias organiniams kalcio kanalų blokatoriams ir „_-conotoxin GVIA“, stipriam ir selektyviam N tipo tipo Ca2 + kanalų blokatoriui (McCleskey ir kt., 1987), kurie buvo pateikti į paraventrikulinę branduolį (žr. Argiolas ir kt., 1990, ir jame esančias nuorodas); (2) apomorfino sukeltą varpos erekciją neleido slopinti azoto oksido sintezės inhibitoriais, skirtais į paraventriculinį branduolį (Melis ir kt., 1994c); ir (3) apomorfinas ir kiti D2 receptorių agonistai, vartojami dozėmis, kurios sukelia varpos erekciją, padidindamos azoto oksido gamybą paraventrikulinėje dializėje, gautą per smegenų mikrodializę, padidėjimą, kurį sumažino paraventrikulinės azoto oksido sintezės inhibitoriai, vartojant dozes, mažinančias dozę. D2 receptoriaus agonistinis sukeltas varpos erekcija (Melis ir kt., 1996). Mechanizmas, kuriuo azoto oksidas aktyvuoja paraventrikulinius oksitocinerginius neuronus, vis dar nežinomas, nors turimi duomenys rodo, kad azoto oksidas veikia kaip intracelulinis pasiuntinys ir kad guanilato ciklazė nedalyvauja. Iš tiesų, nustatyta, kad aktyvus ciklinio GMP, 8-bromo-ciklinio GMP, fosfodiesterazeristinis analogas nesugeba sukelti varpos erekcijos, kai jis buvo duotas į paraventriculinį branduolį (2 pav.) (Žr. Melis ir Argiolas, 1995b ir jame esančias nuorodas).
Pirmiau pateiktas aiškinimas dažnai buvo laikomas neįtikinančiu, daugiausia dėl to, kad dopamino D2 receptorių stimuliacija paprastai yra susieta su inhibicija, o ne neuronų, turinčių šiuos receptorius, ląstelių kūnų sužadinimu, naudojant skirtingus G baltymų sujungtus mechanizmus. (žr. Sokoloff ir Schwartz, 1995). Tačiau galimas šio nesutapimo, kuris atitinka tiesioginį paraventrikulinių oksitocinerginių neuronų stimuliavimą dopamino, paaiškinimą neseniai buvo pasiūlytas aptinkant G baltymų sujungtas dopaminas D4 receptorius, D2 receptorių šeimos narys (D2, D3 ir D4), kurių stimuliacija padidina Ca2 + įsiurbimą ląstelių preparatuose, turinčiuose klonuotą šio receptoriaus potipio versiją (Moreland ir kt., 2004). Dar svarbiau, kad selektyvus D4 receptorių agonistas (pvz., ABT 724) (N-metil-4- (2-cianofenil) piperazinil-3metilbenzamido maleaatas) buvo rastas gebėjimas sukelti vyriškos lyties žiurkių erekciją, kai vartojama sistemiškai (Brioni ir kt., 2004). Šis poveikis nebuvo nustatytas su selektyviu D2 receptoriaus potipio agonistu PNU-95666E (R-5,6-dihidro-N, N-dimetil-4imidazo [4,5,1-i] chinolin-5-aminu) (Hsieh et al., 2004), kuris taip pat negalėjo padidinti Ca2 + antplūdžio į ląstelių preparatus, turinčius klonuotą D4 receptorių potipio versiją (Brioni ir kt., 2004; Moreland ir kt., 2004). Atitinka pirmiau minėtą hipotezę ir išvadas, PD 168,077 (N-metil-4- (2-cianofenil) piperazinil-3metilbenzamido maleato), PIP-3EA (2- [4- (2-metoksifenil) piperazin-1-ilmetil] imidazo [1,2-a] piridinas) ir kiti selektyvūs D4 receptorių agonistai (Heier ir kt., 1997; Melis ir kt., 2006b; Löber ir kt., 2009), buvo nustatyta, kad jie gali sukelti varpos erekciją, suleidžiant sistemiškai, icv ir į paraventriculinis branduolys, nors ir mažiau veiksmingas nei apomorfinas. Šių D4 receptorių agonistų proektekstinis poveikis buvo išvengtas L-745,870 (3- (4- [chlorfenil] piperazin-1-il) -metil-1H-pirolo [2,3-B] piridino trihidrochlorido), selektyvaus D4 receptoriaus. antagonistas (Patel ir kt., 1997; Melis ir kt., 2005, 2006b; Löber ir kt., 2009).
Galiausiai, aukščiau minėtų D4 receptorių agonistų proektekstinis poveikis sumažėjo ir azoto oksido sintezės inhibitoriais, kurie buvo įnešti į paraventriculinį branduolį, ir d (CH2) 5Tyr (Me) 2-Orn8-vazotocinas, selektyvus oksitocino receptorių antagonistas vartojant icv, bet ne paraventrikuliniame branduolyje. Šie rezultatai atitinka hipotezę, kad D4 receptorių agonistai taip pat stimuliuoja oksitocinerginius neuronus, aktyvuodami azoto oksido sintezę, ir išskiria oksitociną extrahipalališkose smegenų srityse, o tai savo ruožtu palengvina varpos erekciją, kaip parodyta apomorfino ir klasikinių D2 agonistų (Melis et al. 2005, 2006b; Löber ir kt., 2009).
Minėti rezultatai taip pat patvirtina hipotezę, kad dopaminas sukelia varpos erekciją, veikdamas D4 receptorius, esančius paraventrikulinių oksitocinerginių neuronų ląstelių kūnuose, ir dėl to padidėja Ca2 + įtekėjimas į oksitocinerginių neuronų ląstelių kūnus, todėl padidėja azoto oksido kiekis. Azoto oksidas savo ruožtu aktyvuoja oksitocinerginius neuronus, kad išskleistų oksitociną papildomose hipotalaminėse smegenų srityse ir stuburo smegenyse, kaip jau minėta. Šiuo atžvilgiu Pažymėtina, kad dopamino receptoriai buvo nustatyti oksitocinerginių neuronų ląstelių kūnuose paraventrikuliniame branduolyje. tik neseniai atlikus dvigubus imunofluorescencinius tyrimus su didelio selektyvumo D2, D3 ir D4 receptorių antikūnais ir su oksitocino antikūnais. Šie tyrimai parodė visų trijų D2receptor potipių (D2, D3 ir D4) ekspresiją, kurie atskirai lokalizuojasi paraventriculinio branduolio oksitocinerginių neuronų ląstelių kūnuose (taip pat supraoptiniame branduolyje ir medialiniame preopticiniame rajone) (Baskerville). ir Douglas, 2008, Baskerville ir kt., 2009).
Tai suteikia stiprią neuroanatominę paramą galimybei, kad D2 tipo dopamino ir dopamino receptorių agonistai sukelia varpos erekciją, aktyvuodami tiesiogiai oksitocinerginius neuronus, kurie išsikiša į aukščiau paminėtas smegenų sritis.pvz., nugaros smegenys, ventralinė tegmentalinė sritis, hipokampas ir amygdala. Tačiau šie rezultatai nepadeda nustatyti D2 receptorių potipių, kurių stimuliacija sukelia erekcijos atsaką. Deja, pagalbos negalima gauti net iš tyrimų, kuriais siekiama nustatyti dopamino receptorių agonistų aktyvuotus oksitocinerginius neuronus paraventrikuliniame branduolyje. Iš tiesų, nepaisant skirtingo įvairių dopamino receptorių potipių aktyvumo, mišrūs dopamino receptorių agonistai (pvz., Apomorfinas) arba selektyvūs D2 receptorių agonistai (pvz., Chinpirolis, veikiantis visus D2 receptorių potipius) arba selektyvūs D4 receptorių agonistai sukelia oksitocinerginių neuronų aktyvinimas, matuojamas pagal FOS baltymo padidėjimą paraventrikulinių branduolių parvoceliuliniuose oksitocinerginiuose neuronuose (Bitner ir kt., 2006). Tačiau ši pastaba neseniai buvo apklausta, nes FOS baltymų padidėjimas paraventrikuliniuose oksitocinerginiuose neuronuose buvo aptiktas tik tada, kai chinololinas sukėlė varpos erekciją, kuri daugiausia veikia D2 ir D3 receptorių potipius, bet ne PD 168077, D4 receptorių agonistas, nepaisant abiejų junginių gebėjimo sukelti seksualinį atsaką (Baskerville ir kt., 2009).
Tada reikia atlikti papildomus kitų D2 receptorių potipių (daugiausia D2 ir D3) agonistų tyrimus, kad būtų galima tiksliai nustatyti kiekvieno dopamino receptorių potipio vaidmenį kontroliuojant erekcijos funkciją paraventrikuliniame lygyje. Šiuo atžvilgiu, kaip jau buvo priminta anksčiau, apomorfinas, kuris veikia stipriai ant visų dopamino receptorių potipių (žr. Brioni ir kt., 2004, ir jame esančios nuorodos), yra daug veiksmingesnis už D4 receptorių agonistus indukuojant varpos erekciją, kai jis švirkščiamas į paraventrikulinę. branduolys. Tai gali būti paaiškinta padidėjusiu apomorfino afinitetu D4 receptorių atžvilgiu, palyginti su išbandytų D4 receptorių agonistų, arba, alternatyviai, D4 receptorių agonistai gali veikti kaip D4 receptorių daliniai agonistai, arba tuo pačiu metu skirtingų dopamino receptorių potipių aktyvacija apomorfinu gali sukelia didesnį oksitocinerginių neuronų aktyvavimą, kuris skatina varpos erekciją, nei tik D2 receptorių agonistų aktyvavimas tik D4 receptorių potipyje.
Dopamino D1 ir D2 receptorių sąveika jau aprašyta kontroliuojant seksualinį elgesį medialinės preoptinės zonos lygiu (žr. Hull ir kt., 1989). Jei bus patvirtinta, kad selektyvūs D2 receptorių agonistai nesugeba sukelti varpos erekcijos (Hsieh et al., 2004) (žr. Depoortère ir kt., 2009), pavyzdžiui, net ir po šių junginių injekcijos į paraventrikulinį branduolį turėtų būti išsamiai išnagrinėtas D3 receptorių vaidmuo atskirai arba kartu su D4 receptorių oksitocinerginių neuronų aktyvavimu tarpininkaujant varpos erekcijai (žr. Baskerville ir kt., 2009). Deja, šiuo metu nėra selektyvių D2 ir D3 receptorių agonistų (pvz., Kurie skiriasi savo afinitetu šiems dviems receptorių potipiams mažiausiai keturių / penkių dydžių in vitro). Dėl šios priežasties neseniai pateiktas teiginys, kad D3 receptoriai tarpininkauja klasikinių D2 receptorių agonistų sukeltai varpos erekcijai, daugiausia pagrįstas galimų D3 receptorių antagonistų, būdingų in vitro eksperimentams, gebėjimu sumažinti klasikinių D2 agonistų sukeltą varpos erekciją, pvz. apomorfinas, chinpirolis ir pramipeksolis, kurie stipriai aktyvuoja visus dopamino D2 receptorių potipius (Collins ir kt., 2009), turi būti tikrai patvirtinti kitais eksperimentais. Šis patvirtinimas būtinas ir todėl, kad šiame tyrime nenustatyta jokio D4 receptorių agonistų poveikio varpos erekcijai, priešingai nei pirmiau minėtų tyrimų rezultatai, kurie rodo, kad D4 receptorių agonistų poveikis yra erekcinis. Iš tiesų, netgi apomorfino gebėjimas sukelia varpos erekciją (ir yawning) D4 receptorių išjungimo pelėse, kurių stiprumas yra toks pat, kaip ir laukinio tipo D4 receptorių išjungimo pelėse, ir D3 receptorių antagonistų gebėjimas panaikinti šių gyvūnų apomorfino atsaką ( Collins et al., 2009) negalima laikyti galutiniu D3 receptorių potipio vaidmens D2 receptorių agonisto sukeltos varpos erekcijos įrodymu. Rūšių skirtumai, neurotransmiterio / neuropeptido ir (arba) neurotransmiterio / neuropeptido receptorių geno abliacijos tyrimai (neurotransmiterio / neuropeptido ir / arba neurotransmiterio / neuropeptido receptorių išstūmimo gyvūnai) paprastai pridėjo dar daugiau painiavos ir komplikacijų patvirtinant įtariamą neurotransmiterių ir seksualinio transplantato vaidmenį. arba neuropeptidai ir jų receptoriai. Būtent, oksitocino geno abliacija gamina oksitociną išnykusioms pelėms, kurios normalizuojasi ir susimaišo, lyg oksitocinas nebūtų būtinas poravimuisi ir kopuliacijai. Be to, homozigotinės moteriškos oksitocino nulaužtos pelės rodo normalų poravimąsi ir atsivedimą, nors ir žymiai sumažėjo pieno išsiskyrimas (Nishimori ir kt., 1996; Young ir kt., 1996). Neuroninio azoto oksido sintezę koduojančio geno abliacija taip pat gamina azoto oksido sintezę, pašalindama peles, kurios normaliai susitinka ir kopijuoja (Huang ir kt., 1993). Tačiau šie rezultatai greičiausiai rodo svarbų reprodukcinės fiziologijos bruožą, ty sistemų, dalyvaujančių jos kontrolėje centriniame ir periferiniame lygyje, atleidimą.
Toks atleidimas iš tikrųjų yra evoliucinė kilmė, nes tai garantuoja genų judėjimą į kitą kartą, kad išliktų šios rūšies išlikimas. Todėl faktas, kad D4 receptoriaus geno abliacija nepakeičia pro-erekcijos poveikio apomorfinui, rodo, kad D4 receptoriai, tokie kaip oksitocinas ir azoto oksidas, yra tik keletas tarpininkų, dirbančių sistemose, kontroliuojančiose erekcijos funkciją. rodo, kad šių receptorių vaidmuo kontroliuojant varpos erekciją ir seksualinį elgesį nėra svarbus. D4 agonistų nesugebėjimas sukelia varpos erekciją, kai sistemiškai buvo skiriamas skirtingų padermių žiurkėms, neseniai buvo pranešta kitame tyrime (Depoortère ir kt., 2009). Tačiau, priešingai nei Collins et al. (2009) ir dar labiau apsvaiginantį vaizdą apie skirtingų D2 receptorių potipių vaidmenį valdant varpos erekciją, šis tyrimas taip pat rodo, kad galimai pasirinktiniai D3 receptorių antagonistai, vartojami sistemiškai, nesugebėjo sumažinti apomorfino sukeltos varpos erekcijos. kamieninės patelės, jautresnės pro-erektiliniam apomorfino poveikiui, o seksualinis atsakas (ir žvėris) buvo antagonizuotas selektyviu D2 antagonistu L-741,626 (3 - [[4- (4-chlorfenil) -4-hidroksipiperidin -1-il] metil-1H-indolas), todėl autoriai teigia, kad D2 receptoriai, o ne D3 ir D4 receptoriai, yra tie, kurie vaidina svarbų vaidmenį D2 agonisto sukeltame varpos erekcijoje (Depoortère ir kt., 2009). Galiausiai, galimybė, kad dopamino receptorių agonistų eksitacinis poveikis oksitocinerginiams neuronams, tarpininkaujančiam varpos erekciją, bent jau iš dalies yra netiesioginis, o ne tiesioginis, pvz., Tarpininkauja ar įtakoja kitų neurotransmiterių neuropeptidų aktyvumo pokyčiai, galintys moduliuoti aktyvumą. Oksitocinerginiai neuronai paraventrikuliniame branduolyje negali būti visiškai atmesti.
3.2. Glutamo rūgšties-oksitocino sąveika paraventrikuliniame branduolyje
Hipotalamies paraventrikulinis branduolys yra labai gausus sinapsėse, turinčiose neurotransmiterio sužadinimo aminorūgštį (pvz., Glutamo rūgštis ir aspartino rūgštis).) (Van Den Pol, 1991). Šioje branduolio virškinimo aminorūgštys dalyvauja daugelyje funkcijų, įskaitant varpos erekciją ir seksualinį elgesį (Roeling ir kt., 1991; Melis ir kt., 1994b, 2000, 2004b). Atitinkamai N-metil-d-asparagino rūgštis (NMDA), selektyvus NMDA receptorių potipio agonistas, bet ne (±) -_-amino-3-hidroksi-5-metil-izoksazol-4-propiono rūgštis (AMPA) buvo nustatyta, kad AMPA receptoriaus potipio arba (±) -trans (1) -amino-1,3-ciklopentano dikarboksilo rūgšties (ACPD) selektyvus agonistas, selektyvus metabotropinio receptoriaus potipio agonistas, gali paskatinti varpos erekciją, švirkščiant į laisvai judančių žiurkių paraventrikulinis branduolys (Melis ir kt., 1994b). 948 MR Melis, A. Argiolas / Neuroscience ir Biobehavioral Reviews 35 (2011) 939-955 NMDA pro erekcinį poveikį neleido (+) MK-801, nekonkurencinis NMDA receptorių antagonistas (Woodruff et al., 1987 ), švirkščiamas į paraventrikulinį branduolį (Melis ir kt., 1994b). Pagal šiuos rezultatus telemetriniuose tyrimuose, kuriais siekiama stebėti intrakaverninį spaudimą, NMDA buvo nustatyta daug aktyvesnė nei kitų eksitacinių aminorūgščių receptorių potipių agonistai, kai jie švirkščiami į paraventriculinį branduolį, didinant intrakavernozinį spaudimą pabudusiems ar anestezuotiems patinams (Zahran et. 2000; Chen ir Chang, 2003).
Kaip buvo pasiūlyta aukščiau dėl oksitocino ir dopamino, tikėtina, kad NMDA receptoriai, jungiantys varpos erekciją, yra oksitocinerginių neuronų ląstelių kūnuose, nes eritacinės aminorūgšties nervų galūnės pažeidžia oksitocinerginių ląstelių kūnus paraventrikuliniame branduolyje. (Van Den Pol, 1991). Analogiškai, kaip ir su dopamino receptorių agonistais, NMDA pro erekcinis poveikis, matyt, yra oksitocinerginio neurotransmisijos aktyvavimo, kurį panaikina oksitocino antagonistas d (CH2) 5Tyr (Me) 2-Orn8-vazotocinas, tačiau ne į paraventrikulinį branduolį (žr. Argiolas ir Melis, 1995, 2004, 2005 ir jame esančias nuorodas). Lygiai taip pat, NMDA sukeltas oksitocinerginio neurotransmisijos aktyvinimas taip pat yra antrinis dėl azoto oksido sintezės aktyvacijos, nes NMDA sukeltą varpos erekciją slopina azoto oksido sintezės inhibitoriai (N-nitro-N-metil-arginino metilesteris ir N- metilo-tio-l-citrulino), skirto į paraventriculinį branduolį, ir NMDA, suleidžiama į paraventriculinį branduolį dozėmis, kurios sukelia varpos erekciją, padidina azoto oksido gamybą hipotalaminėje branduolyje (žr. Argiolas ir Melis, 1995, 2004, 2005 ir nuorodos joje) ). Kaip ir dopamino receptorių agonistams, NMDA sukeltas azoto oksidinazės aktyvavimas taip pat gali būti antrinis dėl padidėjusio Ca2 + įsiurbimo oksitocinerginių ląstelių kūnuose per Ca2 + kanalų sujungtus NMDA receptorius, kaip parodyta keliuose nerviniuose preparatuose (žr. Snyder, 1992; Southam ir Garthwaite, 1993, Schuman ir Madison, 1994 ir jame esančios nuorodos). Azoto oksidas savo ruožtu aktyvuoja oksitocinerginę neurotransmisiją (žr. Aukščiau). Glutamaterginių projekcijų, aktyvuojančių paraventrikulinius oksitocinerginius neuronus, tarpininkaujančius varpos erekciją, kilmė nežinoma, nors kai kurie neuroanatominiai ir elektrofiziologiniai įrodymai rodo, kad jie gali kilti bent iš dalies hipokampe (Saphier ir Feldman, 1987; Chen ir kt., 1992) . Nors reikia toliau dirbti, kad geriau apibūdintume glutamaterginių projekcijų į paraventrikulinę branduolį kilmę, glutamo rūgšties įtraukimas į paraventriculinį branduolį kontroliuojant varpos erekciją ir seksualinį elgesį yra aiškiai pagrįstas mikrodializės tyrimais. Atitinkamai dializate, gautoje iš patinų žiurkių paraventriculinės branduolio, turinčio nekontaktinę erekciją, padidėjusios glutamo rūgšties ir asparagino rūgšties ekstensyviosios koncentracijos padidėjo, kai buvo patekę į nepasiekiamus estrogeno + progesterono gruntuojančius moteriškus žiurkes (Melis ir kt., 2004b), varpos erekcija, kurią taip pat skatina centrinės oksitocinerginės transmisijos aktyvacija (Melis ir kt., 1999a, b). Tokie padidėjimai buvo nustatyti dar didesni, kai buvo leidžiama kopuliacija su imliąja moterimi (Melis ir kt., 2004a). Atsižvelgiant į hipotezę, kad paraventriculiniame branduolyje atsiranda padidėjęs sužadinimo aminorūgščių aktyvumas varpos erekcijos ir kopuliacijos metu, tiek nesusijusios erekcijos, tiek kopuliacinis elgesys (kurio metu atsiranda kopulų varpos erekcija) sumažėja dėl NMDA receptorių blokavimo paraventrikuliniame branduolyje. po to sumažėja azoto oksido kiekio padidėjimas, kuris vyksta šioje hipotalaminėje branduolyje šiuose fiziologiniuose kontekstuose (Melis ir kt., 2000). Padidėjusi GABA išsiskyrimo iš GABAerginių nervų galūnių, sukeliančių eritacinių amino rūgščių sinapsių, lygiagrečiai su oksitocinerginiais ląstelių organais, padidėjimas padidėjo ir paraventrikuliniame branduolyje, kai CB1 antagonistas SR 1A blokavo kanabinoidinius CB141761 receptorius. į šoninius skilvelius arba tiesiogiai į paraventriculinį branduolį dozėmis, kurios sukelia varpos erekciją (žr. Succu ir kt., 2006; Castelli ir kt., 2007). Toks padidėjimas paskatino azoto oksido sintezės aktyvaciją oksitocinerginių neuronų ląstelių kūnuose, didindamas azoto oksido gamybą. Azoto oksidas savo ruožtu suaktyvina oksitocinerginius neuronus, skatinančius varpos erekciją, kaip aprašyta aukščiau. Laikantis tokio mechanizmo, SR 141761A sukeltas varpos erekcija sumažėjo dėl NMDA receptorių blokavimo ir azoto oksido sintezės inhibitorių, bet ne dėl dopamino ar oksitocino receptorių blokavimo paraventriculiniame branduolyje, o tai buvo užkirstas užblokuota. centriniai oksitocino receptoriai, vartojami oksitocino receptorių antagonistų, vartojančių icv
3.3. Oksitocino ir dopamino sąveika ventralinio apvalkalo srityje
Oksitocinas sukelia varpos erekciją, kai dozė priklauso nuo dozės priklausomybės nuo skilvelio ventralinės tegmentalinės dalies. (Melis ir kt., 2007). Mažiausia aktyvi dozė, švirkščiama vienašališkai, buvo 20 ng, o didžiausia tiriamoji dozė buvo 100 ng. Oksitocino poveikį sąlygoja oksitocinerginių receptorių aktyvacija, nes seksualinis atsakas panaikinamas prieš injekuojant oksitocino antagonistą d (CH2) 5Tyr (Me) 2-Orn8-vazotociną kraujagyslių ventraliniam tegmentalui. Šie receptoriai yra lokalizuoti dopaminerginių neuronų ląstelių kūnuose, kurie daugiausia nukreipiami į branduolio akumbensą. Atitinkamai (1) dvigubi imunofluorescencijos tyrimai rodo, kad caudalinių ventralių tegmentalinių zonų oksitocinerginiai pluoštai yra glaudžiai susiję su dopaminerginių neuronų ląstelių kūnais, kurių dauguma buvo teigiamai paženklinti tirozino hidroksilaze ir turintys retrogradinį žymeklį Fluorogold anksčiau švirkščiant į branduolio akumbensą (Melis ir kt., 2007), ir (2) ventralinę tegmentalinę zoną oksitocino sukelta penio erekcija atsiranda kartu su padidėjusiu ląstelių dopamino koncentracija dializate, gautame iš lukšto. branduolys accumbens (Melis ir kt., 2007). Oksitocino sukeltas varpos erekcija taip pat atsiranda kartu su padidėjusiu azoto oksido susidarymu ventraliniame tegmental srityje, nes abu atsakai yra ne tik d (CH2) 5Tyr (Me) 2-Orn8-vazotocino, bet ir azoto oksido sintezės inhibitoriaus S- metil-tio-1-citrulinas, bet taip pat ir _-conotoxin, įtampos priklausomas Ca2 + kanalų blokatorius ir ODQ (1H- [1,2,4] oksadiazolo [4,3-a] chinoksalin-1-one), stiprus guanilato inhibitorius ciklazė, visi duodami caudalinėje ventralio tegmentalio srityje prieš oksitociną (Succu ir kt., 2008). Kadangi daugelis fluorogoldo žymėtų dopaminerginių ląstelių, su kuriais susiduria oksitocinerginiai pluoštai, kurie, kaip nustatyta, yra teigiami tirozino hidroksilazei, yra fiksuojami azoto oksido sintezės ir guanilato ciklazei (Succu ir kt., 2008), oksitocinas. sukeltą varpos erekciją gali paskatinti toks mechanizmas. Oksitocinerginių receptorių aktyvavimas dopaminerginių ląstelių kūnuose padidina Ca2 + antplūdį dopaminerginių neuronų ląstelių kūnuose. Tai aktyvina azoto oksido sintezę, priklausomą nuo Ca2 + -kalmodulino priklausomo fermento, taip padidindama azoto oksido gamybą. Azoto oksidas savo ruožtu aktyvuoja guanilato ciklazę, todėl padidėja ciklinės GMP koncentracija. Ciklinis GMP aktyvuoja dopaminerginius neuronus, nukreipiančius į branduolį. Į caudalinio ventralinio tegmentalio sritį įpurškiamo oksitocino sukeltų ciklinių GMP vaidmenį taip pat patvirtina 8-bromo-ciklinio GMP, aktyvaus fosfodiesterazės atsparaus ciklinio GMP analogo, gebėjimas sukelti varpos erekciją, kai jis švirkščiamas į caudalinė ventralinė tegmentalinė zona, ir didinti nešūnų dopaminą dializate iš branduolio accumbens (Succu ir kt., 2008; Melis ir kt., 2009a). Tai prieštarauja mechanizmui, kuriuo azoto oksidas aktyvuoja oksitocinerginius neuronus paraventrikuliniame branduolyje, nes tai yra 8-bromciklinis GMP, negalintis sukelti varpos erekcijos, švirkščiant į šį branduolį (Melis ir Argiolas, 1995b). 2). Kalbant apie nervų takus, kuriuos dopaminas aktyvina branduolyje accumbens ir sukelia varpos erekciją, jie vis dar nežinomi. Tačiau turimi duomenys rodo, kad šie keliai aktyvuoja dopamino neurotransmisiją hipotalamo paraventrikuliniame branduolyje. Atitinkamai, oksitocino sukelta varpos erekcija atsiranda kartu su padidėjusiu ląstelių dopamino kiekiu ne tik \ t branduolį, bet ir paraventriculinį branduolį, ir yra antagonizuojamas dopamino receptorių antagonisto haloperidoliu, suleistu į paraventrikulinį branduolį (Melis ir kt., 2007). AKartu šie rezultatai patvirtina idėją, kad oksitocinerginiai neuronai, kilę iš paraventriculinio branduolio ir išsikišę į caudalinę ventralinę tegmentalinę sritį, aktyvuojant išleidimo oksitociną šioje srityje, taip aktyvuojant NO-ciklinę GMP signalizavimo sistemą, kuris savo ruožtu aktyvuoja mezolimbinius dopaminerginius neuronus (Melis ir kt., 2007, 2009a; Succu ir kt., 2008). Iš branduolio accumbens išsiskyręs dopaminas savo ruožtu suaktyvina neuroninius kelius, kurie veda prie hipertalaminių dopaminerginių neuronų, kurie stimuliuoja paraventrikulinius oksitocinerginius neuronus, nukreipiančius į nugaros smegenų erekciją, aktyvaciją. Tuo pačiu metu paraventrikuliniame branduolyje išsiskyręs dopaminas taip pat gali suaktyvinti oksitocinerginius neuronus, nukreipiančius į papildomas hipotalamines smegenų sritis, tokias kaip ventralinė tegmentalinė sritis, hipokampas, amygdala ir galbūt kitos smegenų sritys.
Kaip minėta pirmiau, atsižvelgiant į šią hipotezę, apomorfinas, suleidžiamas į paraventriculinį branduolį doze, kuri sukelia varpos erekciją, taip pat padidina ląstelių dopamino koncentraciją branduolyje accumbens, tai sumažina oksitocino receptorių antagonistas d (CH2) 5Tyr (Me ) 2-Orn8-vazotocinas, švirkščiamas į ventralinį tegmentalą (Succu ir kt., 2007; Melis ir kt., 2009a). Kartu pirmiau minėti neuroniniai keliai gali būti hipotetinė neuroninė grandinė, apimanti dopaminą, oksitociną ir kitus neurotransmiterius (pvz., Glutamo rūgštį, žr. Toliau), turinčius įtakos ne tik seksualinei veiklai, bet ir seksualinei motyvacijai bei seksualinei naudai (žr. skyrių 4).
3.4. Oksitocino-glutamo rūgšties sąveika hipokampo ventraliniame subiculume
Oksitocinas sukelia varpos erekciją, kai jis švirkščiamas į ventralį, bet ne dorsaliniame subiculume, priklausomai nuo dozės (Melis ir kt., 2009b). Lytinį atsaką skatina oksitocino receptorių stimuliavimas, panaikinus išankstinę d (CH2) 5Tyr (Me) 2-Orn8-vazotocino injekciją į tą pačią oksitocino vietą, kaip nustatyta kitose smegenų srityse (žr. Aukščiau) . Kalbant apie šių receptorių lokalizavimą, turimi duomenys rodo, kad jie yra lokalizuoti neuronų, turinčių daug azoto oksido sintezės, ląstelių kūnuose.
Atitinkamai, mikrodializės tyrimai rodo, kad oksitocino sukelta varpos erekcija atsiranda kartu su padidėjusiu azoto oksido susidarymu ventraliniame subiculume, ir šis padidėjimas panaikinamas ne tik iš anksto injekavus azoto oksidinazės inhibitorių S-metil-tio-l-citruliną ir azoto oksido šalinimo hemoglobino, bet taip pat ir d (CH2) 5Tyr (Me) 2-Orn8-vazotocino, visų į tą pačią oksitocino vietą dozėmis, kurios priešinasi varpos erekcijai (Melis ir kt., 2010). Dar svarbiau yra tai, kad oksitocino sukelta varpos erekcija atsiranda kartu su padidėjusiu neporeliniu glutamino rūgšties koncentracija ventraliniame subiculume, kurį tik iš dalies antagonizuoja nekonkurencinis NMDA receptorių antagonistas (+) MK-801. ventralinis subiculumas (Melis ir kt., 2010). Kartu šie rezultatai rodo, kad naujai suformuotas azoto oksidas, veikdamas kaip tarpląstelinis pasiuntinys, suaktyvina glutamo rūgšties neurotransmisiją, kuri sukelia varpos erekciją, galbūt per nervų efferentines projekcijas iš skilvelio. subiculum į papildomų hipokampo smegenų sritis. Pagal šią hipotezę, NMDA, suleista į ventralinę subiculumą, sukelia varpos erekciją, priklausomai nuo dozės, ir šis poveikis yra visiškai priešingas injekcijai į tą pačią (+) MK-801, bet ne S-metil-tio-l-citrulino vietą , hemoglobino arba d (CH2) 5Tyr (Me) 2-Orn8-vazotocino (Melis ir kt., 2010). Kalbant apie nervų efferentinius kelius, kurie išsikiša į hippokampo smegenų sritis, suaktyvintas eksitacinių aminorūgščių (ty glutamo rūgšties), esant ventraliniam subiculumui, tikėtina, kad jos yra glutamaterginės, kaip ir dauguma hipokampo efferentinių projekcijų. Šiuo metu galima tik pasiūlyti, kad šios projekcijos moduliuoja mezolimbinių dopaminerginių neuronų aktyvumą, kuris savo ruožtu moduliuoja hipertalaminių dopaminerginių neuronų aktyvumą paraventrikuliniame branduolyje, o tai lemia oksitocinerginių neuronų aktyvaciją. tarpininkaujant varpos erekcijai, kaip jau aptarta (pažiūrėkite aukščiau).
Atitinkamai ventralinio subiculumo oksitocino sukeltas varpos erekcija atsiranda kartu su papildomo ląstelinio dopamino koncentracijos padidėjimu branduolio accumbens lukštuose, ir šis padidėjimas, kaip ir varpos erekcija, panaikinamas d (CH2) 5Tyr (Me) 2- Orn8-vazotocinas, duodamas į ventralinę subiculumą prieš oksitociną (Melis ir kt., 2009b). Be to, kadangi ventralinis subiculumo oksitocino sukeltas varpos erekcija taip pat sumažėja (+) MK-801, švirkščiant į ventralinį tegmentalą, bet ne į branduolį accumbens (Melis ir kt., 2009b) ir atsiranda kartu su padidėjusiu ląstelių skaičiumi. glutamo rūgštis ventraliniame tegmentiniame plote, bet ne branduolyje accumbens, nes abu atsakai buvo panaikinti d (CH2) 5Tyr (Me) -Orn8-vazotocino, švirkščiant į ventralinę subiculumą prieš oksitociną (žr. 3), tikėtina, kad mezolimbinių dopaminerginių neuronų aktyvacija yra antrinė, nes padidėjusi glutamaterginė neurotransmisija yra ventralinio tegmentalo srityje. Tai leidžia manyti, kad ventralinio tegmentalio srityje yra sąveika, kontroliuojanti varpos erekciją. Reikia atlikti tolesnius tyrimus, siekiant nustatyti, ar pro-erekciniai efferentiniai glutameterginiai keliai nuo subikuliato iki ventralinio tegmentalio srities yra tiesioginiai ar netiesioginiai, ty per prefrontalinę žievę arba kitas smegenų sritis (žr. Melis ir kt., 2009b ir jame esančias nuorodas). Kadangi paraventriculinis branduolys taip pat gauna glutamatergines projekcijas iš hipokampo (žr. Aukščiau ir Saphier ir Feldman, 1987), ir glutamo rūgštis aktyvina paraventrikulinius oksitocinerginius neuronus, įskaitant tuos, kurie išsikiša į ventralinį (žr. Argiolas ir Melis, 2005 ir jame esančias nuorodas), ir oksitocinas ventraliniame tegmentalyje sukelia varpos erekciją ir padidina mezolimbinio dopaminerginio neurono aktyvumą.s (žr. aukščiau), yra viliojanti spėlioti, kad paraventrikuliniai oksitocinerginiai neuronai taip pat gali būti bent jau iš dalies įtraukti į mezolimbinių dopaminerginių neuronų aktyvavimą oksitocinu, švirkštu į ventralinę subiculumą. (žr. skyrių 4).
4. baigiamasis žodis
Pirmiau apžvelgti tyrimai patvirtina ir išplečia ankstyvus tyrimus, rodančius, kad žiurkių patinams oksitocinas atlieka pagrindinį vaidmenį atliekant centrinę varpos erekcijos kontrolę hipotalamo ir nugaros smegenų paraventrikulinio branduolio lygyje. Visų pirma, labiausiai iš naujocentų tyrimai rodo, kad oksitocinas daro įtaką varpos erekcijai, taip pat veikdamas kitose smegenų srityse, t. y. ventraliniame tegmentaliniame plote, ventraliniame subiculume ir posteromedialiniame žievės branduolyje..
Paraventrikuliniame lygyje svarbiausias naujas atradimas galbūt yra visų D2 šeimos (D2, D3 ir D4) dopamino receptorių ekspresijos paraventriculinio branduolio (ir supraoptinio branduolio) ir oksitocinerginių neuronų ląstelių kūnuose ekspresija. medialinė preopticinė zona) (Baskerville ir Douglas, 2008; Baskerville ir kt., 2009). Tai suteikia stiprią neuroanatominę parama hipotezei, kad dopamino ir dopamino receptorių agonistai gali suaktyvinti tiesiogiai oksitocinerginius neuronus, dalyvaujančius erekcijos funkcijoje ir projektuojant ne tik stuburo smegenis, bet ir į hipotalamines smegenų sritis. Atsižvelgiant į tai, taip pat svarbu nustatyti, kad dopamino receptorių agonisto sukeltas intrakaverninio spaudimo padidėjimas yra sumažintas oksitocinerginių receptorių blokavimu nugaros sakraliniame stuburo smegenyje (Baskerville ir kt., 2009). Iš tiesų, nors tokie įrodymai buvo gauti anestezuojamose žiurkių patelėse, nustatymas patvirtina paraventriculo-spinalinio oksitocinerginio mažėjančio kelio, susijusio su dopamino receptorių agonisto sukelta varpos erekcija, aktyvaciją. Hvis dėlto dar reikia išsiaiškinti, ar penitų erekcija, kurią sukelia oksitocinerginių ląstelių kūnuose esančių dopamino receptorių stimuliavimas, yra antrinis dėl specifinio D2 šeimos dopamino receptorių potipio (D2, D3 arba D4) aktyvacijos arba jei šie receptorių potipiai bendradarbiauja moduliuodami erekcijos atsaką, galbūt skirtingais būdais, priklausomai nuo konteksto, kuriame atsiranda varpos erekcija (žr. Moreland ir kt., 2004; Enguehard-Gueiffier ir kt., 2006; Melis ir kt., 2006a, b; Löber ir kt., 2009, Collins ir kt., 2009, Depoortère ir kt., 2009, Baskerville ir kt., 2009).
Kita svarbi nauja išvada yra ta, kad oksitocinas sukelia varpos erekciją, kai švirkščiamas ne tik į paraventriculinį branduolį ar hipokampo CA1 lauką, bet ir į ventralinę tegmentalinę sritį, ventralinį subiculumą ir posteromedialinį žievės branduolį. Ankstesniuose tyrimuose šios smegenų sritys nebuvo ištirtos, rodančios, kad oksitocinas padidino spontaninių varpos erekcijos epizodų žiurkių patinams, nors jie patyrė panašias į nugarinės sakralinės nugaros smegenų oksitocinergines projekcijas iš paraventriculinio branduolio. Iš tiesų nustatyta, kad oksitocinas gali padidinti spontaninės varpos erekcijos epizodus, atsiradusius suaugusiems vyrams, nesant jokių seksualinių stimulų, tokių kaip tie, kurie kilę iš prieinamų ar neprieinamų imuninių (estrogeno-progesterono gruntuojamų) kiaušidžių žiurkių ar genitalijų manipuliacija, kai ji išplaukiama į paraventriculinį branduolį ir hipokampo CA1 lauką, bet ne dorsaliniame subiculume, šoniniame pertvara, caudate branduolyje, medialiniame preopticiniame plote, ventromedialiniame branduolyje ir supraopticiniame branduolyje (Melis et al. , 1986). Visuose šiuose tyrimuose varpos erekcija paprastai buvo skaičiuojama, kai penis atsirado iš varpos apvalkalo stebėtojo, kuris nežinojo apie gydymą tiesiogiai eksperimento metu arba vėliau, stebėdamas eksperimentą, įrašytą į vaizdo juostą su vaizdo kamera. Kiekvienas varpos erekcijos epizodas trunka 0.5-1 min ir paprastai lydi pūslę ir (arba) šlaunikaulius. Šių žiurkių eksperimentai paprastai nenaudojami, siekiant nustatyti seksualinės patirties, amžiaus poveikį arba jei šios žiurkės gali būti suskirstytos į mažą ar didelį atsaką į eritilinį oksitocino poveikį, suleidžiamą į skirtingas smegenų sritis. Tai taip pat taikoma daugumai kitų neuropeptidų ir vaistų, kurie didina spontaninio varpos erekcijos epizodus, įskaitant dopamino agonistus, eksitacines aminorūgštis, ACTH-MSH, heksareliną ir VGF peptidus, proektekstinio poveikio tyrimams. Tačiau šių junginių pro-erekcijos efektas buvo pakartotinai patvirtintas telemetrijos metodais, kurie lemia varpos erekcijos atsiradimą, didinant intrakaverninį spaudimą, kuris atsiranda spontaniškai arba po šių junginių įvedimo įvairiais būdais, ty sistemiškai, intracerebroventrikuliariai arba tiesiai į specifinius smegenų branduolius, po slėgio mikrotransduktoriaus implanto tiesiogiai į ertmę (žr. Bernabè ir kt., 1999). Ventralinio aspirato srityje ventralinis subiculumas ir amygdalos posteromedialinis branduolys, taip pat oksitocinas sukelia varpos erekciją veikiant oksitocinerginius receptorius. Tai veda prie mezolimbinių dopaminerginių neuronų, atsiradusių iš ventralinio tegmentalio srities, ir išsikišusių į branduolio accumbens korpusą, aktyvacijos. kaip matuojama pagal padidėjusį dopamino koncentraciją ląstelėje, esančią dializate, gautame iš branduolio accumbens apvalkalo, ir erekcijos atsako, kurį sukėlė peptidas, švirkščiantis į šias papildomas hipotalamines sritis, sumažėjimas, rastas po dopaminerginių receptorių blokados. branduolys (žr. toliau). Kalbant apie mechanizmus, aktyvuotus oksitocinerginių receptorių stimuliacija šiuose smegenų plotuose, kurie sukelia mezolimbinių dopaminerginių neuronų aktyvavimą ir varpos erekciją, geriausia išsiaiškinti tuos, kurie atsiranda caudalinėje ventralio tegmentalio srityje. Iš tiesų, šiame farmakologiniame ir imunofluorescenciniame rezultate matyti, kad oksitocino nervų galai veikia dopaminerginių neuronų ląstelių kūnus, nukreipiančius į branduolio accumbens korpusą (Melis ir kt., 2007, 2009a; Succu ir kt., 2008). Daugelis šių neuronų turi daug azoto oksido sintezės ir guanilato ciklazės. Oksitocinerginių receptorių stimuliavimas šių dopaminerginių neuronų ląstelių kūnuose sukelia azoto oksido sintezės aktyvaciją, dėl kurios padidėja azoto oksido gamyba. Azoto oksidas savo ruožtu aktyvuoja guanilato ciklazę, taip padidindamas ciklinio GMP koncentraciją, kuris sukelia mezolimbinių dopaminerginių neuronų aktyvaciją ir dopamino išsiskyrimą branduolyje accumbens, matuojant iš ląstelių dopamino padidėjimą dializate iš branduolių akumbenų, gautų per smegenų mikrodializę (Succu ir kt., 2008 ). Iš branduolio accumbens išsiskyręs dopaminas savo ruožtu suaktyvina nervų takus, kurie sukelia varpos erekciją. Tai patvirtina dopamino receptorių antagonistų haloperidolio ir (arba) cis-flupentiksolio, švirkšto į branduolį, gebėjimas sumažinti ventralinio tegmentalio oksitocino sukeltą varpos erekciją (Succu ir kt., 2008). Vienas iš pro-erekcijos būdų, atrodo, aktyvuoja incerto-hipotalaminius dopaminerginius neuronus, ypač tuos, kurie išsivysto į paraventrikulinių oksitocinerginių neuronų ląstelių kūnus. Iš tiesų oksitocinas, švirkščiantis į caudalinį ventralinį tegmentalą, didina ekstraląstelinį dopaminą ne tik branduoliuose, bet ir paraventrikuliniame branduolyje, o dopamino receptorių blokada paraventriculiniame branduolyje žymiai sumažina ventralinę tegmentalinę oksitocino sukeltą varpos erekciją (Succu et al. 2007, 2008, Melis ir kt., 2007, 2009a). Šių branduolių egzistavimas dopamino-paraventrikulinių dopamino-paraventrikulinių oksitocino-ventralinių tegmentalinių zonų oksitocino ir dopamino ryšys taip pat rodo pro-erekcijos dozės apomorfino ir D4 receptorių agonisto PD 168077, švirkščiamų į paraventriculinį branduolį, gebėjimas didinti ne-ląstelių dopaminą branduolio accumbens lukšte (Succu ir kt., 2007), atsakas kuris yra panaikintas d (CH2) 5Tyr (Me) -Orn8-vazotocino, skirto į ventralinį tegmentalą (Melis ir kt., 2009a, taip pat žr. toliau). Tačiau reikia atlikti tolesnį darbą, norint nustatyti neuroninius kelius, jungiančius branduolį accumbens su incerto-hypothalamic dopaminergine sistema.
Mechanizmas, kuriuo oksitocinas sukelia varpos erekciją ir aktyvina mezolimbines dopaminergines neuronas, kai jis švirkščiamas į ventralinį subiculumą arba į amigdalos posteromedinį branduolį, yra tik iš dalies suprantamas. Be to, šiose srityse oksitocinas aktyvuoja savo receptorius, kurie veda prie azoto oksido sintezės aktyvinimo, tokiu būdu didindami azoto oksido gamybą. Azoto oksidas savo ruožtu suaktyvina nežinomas efferentines projekcijas, kurios akivaizdžiai padidina glutamaterginį neurotransmisiją ventraliniame apatiniame plote. Tuomet glutamino rūgštis stimuliuoja mezolimbines dopaminergines neuronas, kurios sukelia varpos erekciją. Šią hipotezę visų pirma patvirtina oksitocino, švirkščiamo į ventralinę subiculumą, gebėjimas didinti neporulinį glutamo rūgštį ventraliniame tegmentalje (Fig. 3) ir nekonkurencinio NMDA receptorių antagonisto (+) MK-801, švirkščiamo į ventralinį tegmentalą, bet ne į branduolį, kad sumažėtų oksitocino sukelta penio erekcija, suleista į ventralinį subiculumą arba į posteromedialinį branduolį. (Melis ir kt., 2009b). Šiuo metu galima gauti daugiau informacijos apie ventralinio subiculumo oksitocino sukeltą varpos erekciją. Čia oksitocino sukelta varpos erekcija atrodo antrinė, kai oksitocinerginiai receptoriai yra veikiami azoto oksido sintezės turinčių neuronų ląstelių kūnuose. Tai padidina azoto oksido gamybą, kuri aktyvuoja glutamaterginę neurotransmisiją, veikdama kaip tarpląstelinis pasiuntinys su mechanizmu, panašiu į aprašytą ilgalaikiam potencialui (žr. Snyder, 1992; Southam ir Garthwaite, 1993; Schuman ir Madison, 2004; Schuman ir Madison, XNUMX; . Atsižvelgiant į šią hipotezę, oksitocino sukelta varpos erekcija kartu su ekstraląstelinio glutamo rūgšties padidėjimu dializate nuo ventralinio subiculumo, ir stimuliuojančių amino rūgščių receptorių stimuliavimas ventraliniame subiculume NMDA, sukelia varpos erekciją. Glutamo rūgštis savo ruožtu suaktyvina neuronines efferentines projekcijas, dėl kurių ventralinio apvalkalo zonoje aktyvuojasi mezolimbiniai dopaminerginiai neuronai, kaip nurodyta pirmiau. Jei šie mechanizmai veikia taip pat ir amergos posteromedialiniame branduolyje, šiuo metu nežinoma. Be to, reikia atlikti tolesnius tyrimus, siekiant įrodyti, kad (1) oksitocinerginių nervų galūnės ir receptoriai ventraliniame subiculume ir amygdalos posteromedialiniame branduolyje yra lokalizuoti neuronų, turinčių azoto oksido sintezę (2), ląstelių kūnuose, jei šie neuronai galiausiai yra kontroliuojant eksitacinių aminorūgščių (glutamaterginių) sinapsių ir (3), kad būtų nustatyta azoto oksido aktyvuota signalizavimo sistema. Šiuo atžvilgiu pažymėtina, kad oksitocino gebėjimas didinti azoto oksido-sintazės aktyvumą dopaminerginių neuronų ląstelių kūnuose caudalinėje ventralio tegmentalinėje srityje ir dar nežinomų neuronų ventraliniame subiculume ir amygdalos posteromedialiniame branduolyje primena oksitocino gebėjimas aktyvuoti azoto oksido sintezę oksitocinerginių neuronų ląstelių kūnuose paraventrikuliniame branduolyje (Melis ir kt., 1997). Tačiau, nors ventralinio tegmentalioje zonoje padidėjusi azoto oksido gamyba dopaminerginių neuronų ląstelių kūnuose sukelia šių neuronų aktyvaciją aktyvuodama guanilato ciklazę ir didindama ciklinę GMP, tai neįvyksta paraventrikuliniame branduolyje. Atitinkamai, 8-bromciklinis GMP, suleidžiamas į paraventriculinį branduolį, nesukelia varpos erekcijos, o tai daro, kai jis švirkščiamas į ventralinį tegmentalą. Tuomet kitas signalizacijos kelias, kuris skiriasi nuo azoto okseciklinio GMP sistemos, dalyvauja paraventrikuliniame lygyje aktyvuojant oksitocinerginius neuronus, tarpininkaujančius varpos erekciją endogeninio ir / arba eksogeninio azoto oksido (Melis ir Argiolas, 1995b; Melis ir kt., 1997) (Fig. . 1). Kita vertus, atrodo, kad ciklinis GMP ventraliniame tegmentalyje taip pat vaidina svarbų vaidmenį aktyvuojant mezolimbinius dopaminerginius neuronus ir padidėjusį ląstelių dopaminą, atsirandantį dializate, gautame iš patinų žiurkių branduolių. pasirenkamas rodant ar nerodant nesusijusių varpos erekcijos, kai jis yra patekęs į neprieinamą ovariektomizuotą (estrogeno + progesterono gydytą) žiurkių patelę. Šiomis eksperimentinėmis sąlygomis žiurkių patelėms, turinčioms nesusijusių varpos erekciją, padidėja išorinės ląstelės dopamino koncentracija, kaip tikėtasi, ir šis padidėjimas dar padidėja, nors ir tik nedideliu mastu, fosfodiesterazės inhibitoriais, skirtais caudal ventraliniam tegmentalui (Sanna et. al., 2009).
Oksitocino, švirkščiamo į ventralinį tegmentalą, ventralinį subiculumą ir posteromedialinį amygdalos branduolį, gebėjimas kartu su dopamino agonistais, švirkštais į paraventriculinį branduolį, sukelia varpos erekciją ir aktyvuoti mezolimbinius dopaminerginius neuronus. . Pirma, mechanizmai, panašūs į pirmiau minėtus mechanizmus, gali būti veiksmingi, kai varpos erekcija vyksta fiziologiniuose kontekstuose, pvz., Per kopuliaciją (kai atsiranda kopulų varpos erekcija) arba nesusijusių su varpos erekcija. Šios erekcijos yra feromonų sukeltos varpos erekcijos, kurios skiriasi nuo vaistų ar oksitocino sukeltų erekcijų, kurios atsiranda, kai lytiškai stiprus patinas žiurkėms patenka į neprieinamą moterišką žiurkę (ovariektomizuotą estrogeną ir progesteroną) ir yra laikomi seksualiniu indeksu. susijaudinimas (Sachs, 1997, 2007). Iš tiesų, nors šie rezultatai neįrodo, kad oksitocinas šiose srityse vaidina fiziologiniuose kontekstuose ar po vaisto skyrimo atsirandančią varpos erekciją, jie dar labiau sustiprina ankstyvus rezultatus, rodančius, kad šios smegenų sritys priklauso tiems, kur centralizuotai vartojamas oksitocinas padidina koncentraciją. ne tik spontaniškos varpos erekcijos epizodai, pastebėti po erekcinių vaistų, bet ir siekiant pagerinti vyrų (ir moterų) seksualinį elgesį (žr. Argiolas ir Melis, 2004 ir nuorodos jame). Atitinkamai, oksitocino koncentracija padidėja hippokampe žiurkių patinų, gydytų pro-erekcijos doze apomorfinu, klasikiniu dopamino agonistu (Melis ir kt., 1990) ir d (CH2) 5Tyr (Me) -Orn8-vazotocinu, kuris blokuoja oksitociną receptoriai, yra labai veiksmingi ne tik mažinant kopuliacinį elgesį (Argiolas ir kt., 1987a), bet ir palengvinantis apomorfino poveikis vyrų kopuliaciniam elgesiui (Argiolas ir kt., 1987b) lytiškai stipriai žiurkėms, kurių metu kopulos varpos erekcija atsiranda. d (CH2) 5Tyr (Me) -Orn8-vazotocinas taip pat labai stipriai mažina nesusijusį erekciją lytiniu būdu stipriai patinų žiurkėms, kai jis buvo duotas į nanogramą į šoninius skilvelius, bet ne į PVM (Melis ir kt., 1999a).
Antra, mezolimbiniai dopaminerginiai neuronai vaidina pagrindinį vaidmenį skatinant natūralias stiprinančias paskatas, pavyzdžiui, maistą, vandenį ir seksualinį aktyvumą (Fibiger ir Phillips, 1988; Wise ir Rompre, 1989; Everitt, 1990). Manoma, kad iš šių neuronų išskiriamas dopaminas tarpininkauja motyvacinių natūralių dirgiklių aspektų perkėlimui į tikslą nukreiptą elgesį, pavyzdžiui, seksualinio aktyvumo, seksualinio partnerio ir lytinio akto ieškojimo atveju, siekiant gauti atlygį ir pasitenkinimą (Goto ir Grace, 2005). Atitinkamai, ląstelėje esanti dopamino koncentracija padidėja nuo lytiniu keliu patyrusių patinų žiurkių branduolio akumbenų, kai patenka į neprieinamą ovariektomizuotą estrogeno + progesterono imituojančią moterišką žiurkę, ir toks padidėjimas buvo dar didesnis, kai žiurkių patinams buvo leista susikaupti su imliąja moterimi (Pfaus ir Everitt, 1995).
Trečia, dabartiniai rezultatai patvirtina hipotezę, kad neuroninė grandinė sujungia paraventriculinį branduolį su ventraliu tegmentaliniu rajonu tiesiogiai arba netiesiogiai (per ventralinį subiculum ir / ar amygdalos posteromedialinį branduolį) ir branduolį accumbens, ir iš čia per nežinomą atgal į paraventriculinį branduolį, norint kontroliuoti oksitocinerginių neuronų, nukreipiančių į nugaros smegenų erekciją, ir oksitocinerginių neuronų, išsikišusių į ventralinį tegmentalinį plotą, ventralinį subiculum ir amterdalos posteromedialinį žievės branduolį, aktyvumą. dopaminerginių neurolonų neuropakų aktyvumas (4 pav.). Ši sudėtinga nervų grandinė gali atlikti vaidmenį integruojant nervinę veiklą, susijusią su vyriškojo seksualinio elgesio vartojimo (erekcijos-ejakuliacijos) ir numatomųjų (motyvacinių ir naudingų) aspektų kontrole fiziologiniuose kontekstuose. Iš tiesų, nepageidaujamų patinų žiurkių patekimo metu, kai nėra kontakto erekcijos, ir dar labiau, kai leidžiama susikalbėti, dar labiau padidėja ląstelių dopamino (Pfaus ir Everitt, 1995) ir paraventricular branduolių lytiniu potencialu patyrusių patelių žiurkės. pvz., kai kopuliacija atsiranda varpos erekcija (Melis ir kt., 2003). Taigi, nors reikia atlikti tolesnius tyrimus, siekiant išsiaiškinti endogeninio oksitocino vaidmenį ventraliniame tegmentiniame plote, ventraliniame subiculume ir amygdaloje nesusijusios erekcijos ir seksualinio elgesio metu, gali būti pagrįsta manyti, kad ši hipotetinė nervų grandinė, prisidedant prie seksualinio elgesio vartojimo aspektai, tuo pačiu metu taip pat gali aktyvuoti mezolimbinę dopaminerginę sistemą, suteikiančią neuroninį substratą, skirtą paaiškinti seksualinio aktyvumo naudingas savybes (Everitt, 1990; Pfaus ir Everitt, 1995). Šiuo atžvilgiu pažymėtina, kad miatimbinė dopaminerginė sistema, suaktyvinta oksitocino, suleisto į ventralinį tegmentalą, yra tokia pati, kaip ir opiatų, kanabinoidų, amfetamino, kokaino ir alkoholio (Tanda ir kt., 1997), ir buvo nustatyta, kad oksitocinas sugeba sumažinti toleranciją ir priklausomybę nuo kokaino, morfino, alkoholio ir kanapių (Kovacs ir kt., 1998; Cui et al., 2001). Apibendrinant, atrodo, kad oksitocinas, išsiskyręs ne tik ventralinio tegmentalio srityje, bet ir ventraliniame subiculume bei posteromedialiniame žievės branduolyje, gali aktyvuoti mezolimbinius dopaminerginius neuronus, kurie gali būti susiję su seksualinės veiklos apetitiniu ir naudingu poveikiu. . Mesolimbinių dopaminerginių neuronų aktyvavimas gali būti tiesioginis, per oksitocinerginius receptorius mezolimbinių dopaminerginių neuronų ląstelių kūnuose, arba netiesiogiai, aktyvuojant glutamo rūgšties neurotransmisiją ventralinėje apatinėje zonoje.
Iš branduolio accumbens apvalkalo išsiskyręs dopaminas savo ruožtu moduliuoja paraventriculiniame branduolyje esančius hipertalaminius dopaminerginius neuronus, sukeldamas arba varpos erekciją (aktyvuojant oksitocinerginius neuronus, nukreipiančius į stuburo smegenis), arba seksualinę motyvaciją ir atlygį (aktyvuojant oksitocinerginį). neuronus, nukreipiančius į ventralinę tegmentalinę sritį, ventralinę subiculumą arba amygdalos posteromedialinį žievės branduolį). Kadangi dopaminas taip pat išsiskiria branduolio accumbens apvalkale ir paraventrikuliniame branduolyje, kai varpos erekcija vyksta fiziologiniuose kontekstuose (pvz., Nesusijusios erekcijos ir kopuliacijos) (Succu ir kt., 2007; Melis ir kt., 2003, 2007), tikėtina, kad centriniai oksitocinerginiai neuronai dalyvauja neuroninėse grandinėse, tarpininkaujant sąveikai tarp mezolimbinės ir incerto-hipotalaminės dopaminerginės sistemos. Šios nervinės grandinės gali atlikti ne tik seksualinės elgsenos (pvz., Varpos erekcijos ir kopuliacijos), bet ir seksualinės motyvacijos, seksualinio susijaudinimo ir seksualinio atlygio metu.
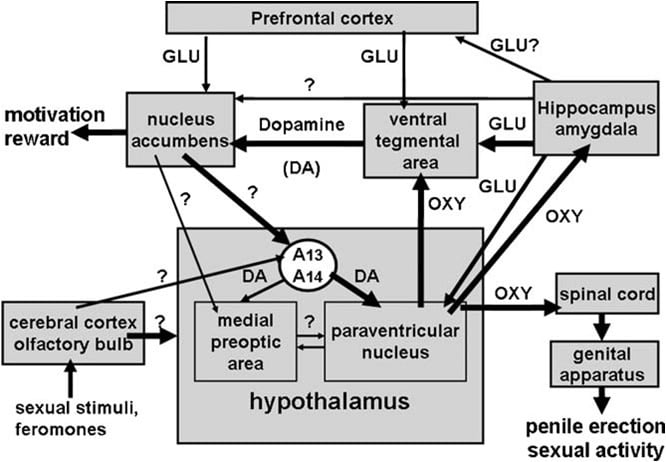
Scheminis hipotetinės neuroninės grandinės, apimančios oksitociną, įtakojantį seksualinę motyvaciją, atlygį ir seksualinę veiklą, vaizdavimas, kaip siūloma šio skyriaus ir ankstesnių ataskaitų rezultatuose. Oksitocinerginiai neuronai, kilę iš paraventrikulinio branduolio ir išsikišę į stuburo smegenis, kai jie aktyvuojami, pvz., Dopamino ir glutamo rūgšties (bet taip pat ir kitų neurotransmiterių ir (arba) neuropeptidų), palengvina varpos erekciją ir seksualinę veiklą, aktyvuojant oksitocinerginius neuronus, nukreiptus į stuburo smegenis. Dopaminas ir glutamo rūgštis (bet taip pat neurotransmiteriai ir neuropeptidai) paraventriculiniame branduolyje taip pat aktyvuoja oksitocinerginius neuronus, nukreipiančius į ventralinę tegmentalinę sritį, tokiu būdu aktyvuodami mezolimbinius dopaminerginius neuronus, nukreipiančius į branduolį accumbens, moduliuodami seksualinę motyvaciją ir atlygį. Iš branduolių accumbens (NAs) išsiskiriantis dopaminas savo ruožtu aktyvuoja vis dar nežinomus neuroninius kelius, kurie padidina hipertalaminių dopaminerginių neuronų, kilusių iš A13-A14 grupių Dahlstrom ir Fuxe, aktyvumą, sukeldami oksitocinerginius neuronus, įskaitant tuos, kurie projektuoja nugaros smegenų, sukelia varpos erekciją. Šią schemą taip pat gali aktyvuoti oksitocinas, švirkščiantis ne tik į caudalinę ventralinę tegmentalinę sritį, bet ir į ventralinę subiculumą ir amygdalą, kuri taip pat gauna oksitocinerginį inervavimą iš paraventrikulinio branduolio, galbūt per tiesioginį ar netiesioginį glutamaterginį efferentą į ventralą. lemia tiek seksualinės motyvacijos, tiek ir varpos erekcijos moduliavimą. Galiausiai, aukščiau minėta schema taip pat gali būti suaktyvinta seksualiniais stimulais ir feromonais, nes padidėjęs dopamino ir glutamo rūgšties padidėjimas paraventrikuliniame branduolyje (ir medialiniame preopticiniame plote) per feromoną sukeltas nesusijusias erekcijas ir kopuliaciją (atitinkamoms nuorodoms žr. Nuorodų sąrašas).
Padėkos Šis darbas iš dalies buvo paremtas Italijos universiteto ir mokslinių tyrimų ministerijos dotacija AA ir MRM
Nuorodos
Andersson, KE, 2001. Varpos erekcijos farmakologija. Pharmacol. Red 53, 417-450. Argiolas, A., 1994. Azoto oksidas yra centrinis varpos erekcijos tarpininkas. Neurofarmakologija 33, 1339-1344. Argiolas, A., 1999. Neuropeptidai ir seksualinis elgesys. Neurosci. Biobehav. Red 23, 1127-1142. Argiolas, A., Gessa, GL, 1991. Oksitocino centrinės funkcijos. Neurosci. Biobehav. Red 15, 217-231. Argiolas, A., Melis, MR, 1995. Varpos erekcijos neuromoduliacija: neurotransmiterių ir neuropeptidų vaidmens apžvalga. Prog. Neurobiol. 47, 235-255. Argiolas, A., Melis, MR, 2004. Oksitocino ir paraventrikulinio branduolio vaidmuo vyriškų žinduolių elgesyje. Physiol. Behav. 83, 309-317. Argiolas, A., Melis, MR, 2005. Vidutinė varpos erekcijos kontrolė: hipotalamo paraventrikulinio branduolio vaidmuo. Prog. Neurobiol. 76, 1-21. Argiolas, A., Collu, M., Gessa, GL, Melis, MR, Serra, G., 1988. Oksitocino antagonistas d (CH2) 5Tyr (Me) -Orn8-vazotocinas antagonizuoja vyrų kopuliacinį elgesį žiurkėms. E. J. Pharmacol. 149, 389-392. Argiolas, A., Collu, M., D'Aquila, P., Gessa, GL, Melis, MR, Serra, G., 1989. Žiurkėms oksitocino antagonistas d (CH2) 5Tyr (Me) -Orn8-vasotocinas apsaugo vyrų kopuliacinio elgesio apomorfino stimuliavimą. Pharmacol. Biochem. Behav. 33, 81-83. Argiolas, A., Melis, MR, Gessa, GL, 1985. Žiurkėms intraventrikulinė oksitocinas sukelia žandikaulių ir varpos erekciją. E. J. Pharmacol. 117, 395-396. Argiolas, A., Melis, MR, Gessa, GL, 1986. Oksitocinas: labai stiprus varpos erekcijos ir žiaurių žiurkių patinimas. E. J. Pharmacol. 130, 265-272. Argiolas, A., Melis, MR, Mauri, A., Gessa, GL, 1987a. Paraventrikulinis branduolio pažeidimas neleidžia žaizdų ir varpos erekcijai, kurią sukelia apomorfinas ir oksitocinas, bet ne ACTH žiurkėms. Brain Res. 421, 349-352. Argiolas, A., Melis, MR, Vargiu, L., Mauri, A., Gessa, GL, 1987b. d (CH2) 5Tyr (Me) -Orn8-vazotocinas, stiprus oksitocino antagonistas, antagonizuoja varpos erekciją ir žiaurumą, kurį sukelia oksitocinas ir apomorfinas, bet ne ACTH 1-24. E. J. Pharmacol. 134, 221-224. Argiolas, A., Melis, MR, Stancampiano, R., Gessa, GL, 1990. - Konotoksinas apsaugo apomorfiną ir oksitocino sukeltą varpos erekciją ir žiaurumą žiurkėms. Pharmacol. Biochem. Behav. 37, 253-257. Arletti, R., Bertolini, A., 1985. Oksitocinas stimuliuoja lordozės elgesį žiurkių patelėms. Neuropeptidai 6, 247-255. Arletti, R., Bazzani, C., Castelli, M., Bertolini, A., 1985. Oksitocinas pagerina vyrų kopuliacinį poveikį žiurkėms. Horm. Behav. 19, 14-20. Arletti, R., Benelli, A., Bertolini, A., 1990. Oksitocinas stimuliuoja senėjimo žiurkių lytinį elgesį. E. J. Pharmacol. 179, 377-382. Arletti, R., Calzà, L., Giardino, L., Benelli, A., Cavazzutti, E., Bertolini, A., 1997. Seksualinė impotencija siejama su sumažėjusia oksitocino gamyba ir padidėjusia opioidinių peptidų gamyba hipotalamo paraventrikuliniame branduolyje. Neurosci. Lett. 233, 65-68. Bancila, M., Giuliano, F., Rampin, O., Mailly, P., Brisorgueil, MJ, Clas, A., Verge, D., 2002. Įrodymai, kad hipotalamo paraventriculinis branduolys yra tiesioginis projekcija į galimus serotoninerginius branduolio paragigantocellularis neuronus, dalyvaujančius kontroliuojant erekciją žiurkėms. E. J. Neurosci. 16, 1240-1249. Burnett, AL, Lowenstein, CJ, Bredt, DS, Chang, TSK, Snyder, SH, 1992. Azoto oksidas - fiziologinis varpos erekcijos tarpininkas. Mokslas 257, 401-403. Baskerville, TA, Douglas, AJ, 2008. Dopamino ir oksitocino sąveika kontroliuojant seksualinį elgesį. Prog. Brain Res. 170, 277-289. Baskerville, TA, Allard, J., Wayman, C., Douglas, AJ, 2009. Dopamino-oksitocino sąveika varpos erekcijos metu. E. J. Neurosci. 30, 2151-2164. Benelli, A., Bertolini, A., Poggioli, R., Cavazzutti, E., Calzà, L., Giardino, L., Arletti, R., 1995. Azoto oksidas yra susijęs su žiurkių lytiniu elgesiu. E. J. Pharmacol. 294, 505-510. Bernabè, J., Rampin, O., Sachs, BD, Giuliano, F., 1999. Intrakaverninis spaudimas žiurkių erekcijos metu: integruotas požiūris, pagrįstas telemetriniu įrašymu. Pak. J. Physiol. 276, R441-R449. Bitner, RS, Nikkel, AL, Otte, S., Martino, B., Barlow, EH, Bhatia, P., Stewart, AO, Brioni, JD, Decker, MW, Moreland, RB, 2006. Dopamino D4 receptorių signalizavimas žiurkių paraventrikulinėje hipotalaminėje branduolyje: natūralaus susiejimo įrodymas, apimantis neatidėliotiną ankstyvo geno indukciją ir mitogeno aktyvuotą baltymų kinazės fosforilinimą. Neurofarmakologija 50, 521-531. Brioni, JD, Moreland, RB, Cowart, M., Hsieh, GC, Stewart, AO, Hedlund, P., Donnelly-Roberts, DL, Nakane, M., Lynch 3rd., J., Kolasa, T., Polakowski , JS, Osinski, MA, Marsh, K., Andersson, KE, Sullivan, JP, 2004. ABT-4 dopamino D724 receptorių aktyvinimas sukelia varpos erekciją žiurkėms. Proc. Natl. Acad. Sci. JAV 101, 6758-6763. Buijs, RM, 1978. Žiurkių viduje ir ne hipotalaminiuose vazopresino ir oksitocino keliuose. Ląstelių audinių rez. 192, 423-435. Buijs, RM, Geffard, M., Pool, CW, Hoorneman, EMD, 1984. Supraopticinio ir paraventrikulinio branduolio dopaminerginis inervavimas. Šviesos ir elektronų mikroskopinis tyrimas. Brain Res. 323, 65-72. Caldwell, JD, Prange, AJ, Pedersen, CA, 1986. Oksitocinas palengvina estrogenais gydytų žiurkių seksualinį imlumą. Neuropeptidai 7, 175-189. Cameron, JL, Pomerantz, SM, Layden, LM, Amico, JA, 1992. Dopaminerginis oksitocino koncentracijos stimuliavimas vyrų ir moterų beždžionių plazmoje apomorfinu ir D2 receptoriaus agonistu. J. Clin. Endokrinolis. Metab. 75, 855-860. Canteras, NS, Simerly, RB, Swanson, LW, 1995. Amigdalos medialinio branduolio projekcijų organizavimas: PHAL tyrimas su žiurkėmis. J. Comp. Neurolis. 360, 213-245. Carmichael, MS, Humbert, R., Dixen, J., Palmisano, G., Greeleaf, W., Davidson, JM, 1987. Plazmos oksitocinas didina žmogaus lytinį atsaką. J. Clin. Endokrinolis. Metab. 64, 27-31. Carter, CS, 1992. Oksitocinas ir seksualinis elgesys. Neurosci. Biobehav. Red 16, 131-144. Carter, CS, Lederhendler, II, Kirkpatrick, B., 1997. Niujorko mokslų akademijos interaktyvioji neurobiologija, Annals. 807. Niujorko mokslų akademija. Castelli, MP, Piras, AP, Melis, T., Succu, S., Sanna, F., Melis, MR, Collu, S., Ennas, MG, Diaz, G., Mackie, K., Argiolas, A. , 2007. Kannabinoidiniai CB1 receptoriai paraventrikuliniame branduolyje ir centrinė varpos erekcijos kontrolė: imunocitochemija, autoradiografija ir elgesio tyrimai. Neurologija 147, 197-206. Chen, KK, Chang, LS, 2003. Eksitacinių aminorūgščių receptorių agonistų įtaka varpos erekcijai po žiurkių į paraventriculinį branduolį. J. Urol. 62, 575-580. Chen, KK, Chan, JYH, Chang, LS, Chen, MT, Chang, SHH, 1992. Varpos erekcijos atsiradimas po hippokampo susidarymo žiurkėse. Neurosci. Lett. 141, 218-222. Chen, KK, Chan, JYH, Chang, LS, 1999. Dopaminerginė neurotransmisija hipotalamo paraventriculiniame branduolyje centriniame varpos erekcijos reguliavime žiurkėse. J. Urol. 162, 237-242. Collins, GT, Truccone, A., Haji-Abdi, F., Newman, AH, Grundt, P., Rice, KC, Husbands, SM, Greedy, BM, Enguehard-Gueiffer, C., Gueiffer, A., Chen , J., Wang, S., Katz, JL, Grandy, DK, Sunahara, RK, Woods, JH, 2009. DopaminoD2 tipo agonistų proektektinį poveikį medikavo D3 receptorius žiurkėms ir pelėms. J. Pharmacol. Exp. Ther. 329, 210-217. Coolen, LM, Allard, J., Truitt, WA, McKenna, KE, 2004. Centrinis ejakuliacijos reguliavimas. Physiol. Behav. 83, 203-215. Cui, SS, Bowen, RC, Gu, GB, Hannesson, BK, Yu, PH, Zhang, X., 2001. Kannabinoidų nutraukimo sindromo prevencija ličio pagalba: oksitocinerginio neuronų aktyvavimo dalyvavimas. J. Neurosci. 21, 9867-9876. Dahlstromas, A., Fuxe, K., 1964. Įrodymai apie monoaminą turinčių neuronų buvimą centrinėje nervų sistemoje. I. Monoaminų demonstravimas smegenų kamieno neuronų ląstelių kūnuose. Acta Physiol. Scand. 62 (Suppl. 232), 1-54. Depoortère, R., Bardin, L., Rodrigues, M., Abrial, E., Aliaga, M., Newman-Tancredi, A., 2009. Dopamino D2 tipo receptorių agonistų sukeltas pūslinės erekcijos ir žiovulys žiurkėms: dopamino D2, bet ne D3 ir D4 receptorių įtaka ir įtaka. Behav. Pharmacol. 20, 303-311. Domes, G., Heinrichs, M., Buchel, C., Braus, DF, Herpertz, SC, 2007. Oksitocinas susilpnina amygdala atsakymus į emocinius veidus, nepriklausomai nuo valencijos. Biol. Psichiatrija 62, 11871190. Dominguez, J., Riolo, JV, Xu, Z., Hull, ME, 2001. Reguliavimas pagal medialinį ambulatorinį kopuliaciją ir medialinį preopticinį dopamino išsiskyrimą. J. Neurosci. 21, 349-355. Donaldson, ZR, Young, LJ, 2009. Oksitocinas, vazopresinas ir socialumo neurogenetika. Mokslas 322, 900-904. Eaton, RG, Markowski, VF, Lumley, LA, Thompson, JT, Mozė, J., Hull, EM, 1991. Paraventrikuliniame branduolyje esantys D2 receptoriai reguliuoja lyties organų reakciją ir kopuliaciją patinų žiurkėms. Pharmacol. Biochem. Behav. 39, 177-181. Ebner, K., Bosch, OL, Krömer, SA, Singewald, N., Neumann, ID, 2005. Oksitocino išsiskyrimas žiurkės centrinėje amygdaloje moduliuoja streso įveikimo elgesį ir eksitacinių aminorūgščių išsiskyrimą. Neuropsichofarmakologija 30, 223-230. Enguehard-Gueiffier, C., Hübner, H., El Hakmaoui, A., Allouchi, H., Gmeiner, P., Argiolas, A., Melis, MR, Gueiffier, A., 2006. 2 - [(4-fenilpiperazin-1-il) metil] imidazo (di) azinai kaip selektyvūs D4-ligandai. Varpos erekcijos indukcija 2- [4- (2-metoksifenil) piperazin-1-ilmetil] imidazo [1,2-a] piridinas (PIP3EA), stiprus ir selektyvus D4 agonistas. J. Med. Chem. 49, 3938-3947. Everitt, BJ, 1990. Seksualinė motyvacija: vyrų žiurkių apetitinių ir kopuliacinių reakcijų mechanizmų nervų ir elgesio analizė. Neurosci. Biobehav. Red 14, 217-232. Fibiger, HC, Phillips, AG, 1988. Mesokortikolimbinė dopamino sistema ir atlygis. Ann. N. Y. Acad. Sci. 5, 206-215. Prancūzų, SJ, Totterdell, S., 2003. Atskiros branduolinės akumbenso projekcijos neuronai žiurkėms gauna tiek bazolaterinius amygdala, tiek ventralinius subicular afferentus. Neurologija 119, 19-31. 954 MR Melis, A. Argiolas / Neurotyros ir biologinės elgsenos apžvalgos 35 (2011) 939-955 Freund-Mercier, MJ, Richard, P., 1981. Oksitocino intraventrikulinių injekcijų žiurkių patekimo į pieno šalinimo refleksą poveikis. Neurosci. Lett. 23, 193-198. Freund-Mercier, MJ, Richard, P., 1984. Oksitocino oksitocino neuronų palengvinimo kontrolė žindymo metu žiurkėse. J. Physiol. (Lond.) 352, 447-466. Freund-Mercier, MJ, Stoeckel, ME, 1995. Somatodendriniai autoreceptoriai oksitocino neuronuose. In: Ivell, R., Russel, JA (Red.), Oksitocinas, Ląsteliniai ir molekuliniai metodai medicinoje ir moksliniuose tyrimuose. Adv. Exp. Med. Biol., 365. Plenum Press, Niujorkas ir Londonas, pp. 185 194. Freund-Mercier, MJ, Stoeckel, ME, Palacios, JM, Pazos, JM, Richard, PH, Porte, A., 1987. 3H oksitocino jungimosi vietų farmakologinės charakteristikos ir anatominis pasiskirstymas Wistar žiurkių smegenyse tiriami autoradiografija. Neurologija 20, 599-614. Giuliano, F., Rampin, O., 2000. Centrinė varpos erekcijos kontrolė. Neurosci. Biobehav. Red 24, 517-533. Giuliano, F., Allard, J., 2001. Dopamino ir lytinės funkcijos. Žiniasklaida. J. Impotence Res. 13 (Suppl. 3), 18-28. Giuliano, F., Rampin, O., 2004. Erekcijos neuroninė kontrolė. Physiol. Behav. 83, 189-201. Giuliano, F., Bernabè, J., McKenna, KE, Longueville, F., Rampin, O., 2001. Oksitocino nugaros proerektilinis poveikis anestezuotoms žiurkėms. Am J. Physiol. Regul. Integ. Comp. Physiol. 280, R1870-R1877. Goto, Y., Grace, AA, 2005. Branduolinio akumbenso limbinio ir žievės pavojaus dopaminerginis moduliavimas tiksliniame elgesyje. Nat. Neurosci. 8, 805-812. Hawthorn, J., Ang, VT, Jenkins, JS, 1985. Žiurkių smegenų ir nugaros smegenų pakitimų hipotalaminių paraventrikulinių, supraoptinių ir suprachiasmatinių branduolių poveikis vazopresinui ir oksitocinui. Brain Res. 346, 51-57. Heier, RF, Dolak, LA, Duncan, JN, Hyslop, DK, Lipton, MF, Martin, LJ, Mauragis, MA, Piercey, MF, Nichols, NF, Schreur, PJ, Smith, MW, Moon, MW, 1997. (R) -5,6-dihidro-N, N-dimetil-4H-imidazo [4,5,1-ij] chinolin-5-amino ir jo metabolitų sintezė ir biologinis aktyvumas. J. Med. Chem. 40, 639-646. Hsiehas, GC, Hollingsworthas, PR, Martino, B., Changas, R., Terranova, MA, O'Neillas, AB, Lynchas, JJ, Morelandas, RB, Donnelly-Robertsas, DL, Kolasa, T., Mikusa, JP , McVey, JM, Marsh, KC, Sullivan, JP, Brioni, JD, 2004 m. Centriniai mechanizmai, reguliuojanti varpos erekciją sąmoningose žiurkėse: dopaminerginės sistemos, susijusios su apomorfino proerektiniu poveikiu. J. Pharmacol. Exp. Ther. 308, 330-338. Huang, PL, Dawson, TM, Bredt, DS, Snyder, SH, Fishman, MC, 1993. Tikslus neuronų azoto oksido sintezės geno sutrikimas. Ląstelė 75, 1273-1286. Huber, D., Veinante, P., Stoop, R., 2005. Vasopresinas ir oksitocinas sužadina atskiras neuronų populiacijas centrinėje amygdaloje. Mokslas 308, 245-248. Hull, EM, Warner, RK, Bazzett, TJ, Eaton, RC, Thompson, JT, 1989. D2 / D1 santykis medialiniame preopticiniame plote veikia patinų žiurkių kopuliaciją. J. Pharmacol. Exp. Ther. 251, 422-427. Hull, EM, Du, J., Lorrain, DS, Matuszewich, L., 1995. Ekstrakulinė dopaminas medialinėje preopticinėje srityje: poveikis seksualinei motyvacijai ir kopuliacijos hormoninei kontrolei. J. Neurosci. 15, 7465-7471. Korpusas, EM, Meisel, RL, Sachs, BD, 2002. Vyrų seksualinis elgesys. Pfaff, DW, Arnold, AP, Etgen, AM, Fahrbach, SE, Rubin, RT (Red.), Hormonai, Smegenys ir Elgesys. Academic Press, New York, pp. 3 137. Hurlemann, R., Patin, A., Onur, OA, Cohen, MX, Baumgartner, T., Metzler, S., Dziobek, I., Gallinat, J., Wagner, M., Maier, W., Kendrick, KM, 2010. Oksitocinas stiprina priklausomybę nuo amygdos, socialiai sustiprintą mokymąsi ir emocinę empatiją žmonėms. J. Neurosci. 30, 4999-5007. Ivell, R., Russel, JA, 1995. Oksitocinas: ląstelių ir molekuliniai metodai medicinoje ir moksliniuose tyrimuose. Eksperimentinės medicinos ir biologijos pažanga, t. 395. Plenum Press, Niujorkas. Kelley, AE, Domesick, VB, 1982. Projektavimas nuo hippokampo susidarymo iki žiurkės branduolio pasiskirstymo: anterogrado ir retrogradinis-krienų peroksidazės tyrimas. Neurologija 7, 2321-2335. Kimura, Y., Naitou, Y., Wanibuchi, F., Yamaguchi, T., 2008. 5-HT (2C) receptorių aktyvacija yra įprastas apomorfino, oksitocino ir melanotano II proerektilinio poveikio mechanizmas žiurkėms. E. J. Pharmacol. 589, 157-162. Kondo, Y., Sachs, BD, Sakuma, Y., 1998. Medialinės amygdalos reikšmė žiurkių varpos erekcijai, kurią sukelia nuotolinės moterys. Behav. Brain Res. 91, 215-222. Kovacs, GL, Sarnyai, Z., Szabo, G., 1998. Oksitocinas ir priklausomybė: apžvalga. Psychoneuroendocrinology 23, 945-962. Lee, HJ, Macbeth, AH, Pagani, JH, Scott Young 3rd, W., 2009. Oksitocinas: didelis gyvenimo palengvintuvas. Prog. Neurobiol. 88, 127-151. Lindvall, O., Bjorklund, A., Skagerberg, G., 1984. Selektyvus dochamino ir telencephalono dopamino galinių sistemų democheminis demonstravimas: nauji hipotalaminių neurosekordinių branduolių dopaminerginės inervacijos įrodymai. Brain Res. 306, 19-30. Liu, YC, Salamone, JD, Sachs, BD, 1997. Sumažėjęs seksualinis atsakas po žiurkių patogeninių paraventrikulinių branduolių pažeidimų. Behav. Neurosci. 111, 1361-1367. Löber, S., Tschammer, N., Hübner, H., Melis, MR, Argiolas, A., Gmeiner, P., 2009. Azuleno sistema kaip naujas bioisosteris: stiprių dopamino D4 receptorių ligandų, sukeliančių varpos erekciją, projektavimas. Chem. Med. Chem. 4, 325-328. McCleskey, EW, Fox, AP, Feldman, DH, Cruz, LJ, Olivera, BM, Tsien, RW, Yoshikami, D., 1987. - Konotoksinas: tiesioginis ir nuolatinis specifinių kalcio kanalų blokavimas neuronuose, bet ne raumenys. Prot. Nat. Acad. Sci. JAV 84, 4327-4331. McKenna, KE, 2000. Keletas pasiūlymų dėl centrinės nervų sistemos kontrolės varpos erekcijai. Neurosci. Biobehav. Red 24, 535-540. Marson, L., McKenna, KE, 1992. 5-hidroksitriptamino vaidmuo mažėjant stuburo lytinių refleksų slopinimui. Exp. Brain Res. 88, 313-318. Marson, L., McKenna, KE, 1996. CNS ląstelių grupės, dalyvaujančios kontroliuojant izchiocavernosus ir bulbospongiosus raumenis: transneuronalinis tyrimas, naudojant pseudorabies virusą. J. Comp. Neurolis. 374, 161-179. Meisel, RL, Sachs, BD, 1994. Vyrų lytinio elgesio fiziologija. In: Knobil, E., Neil, J. (Red.), Reprodukcijos fiziologija, t. 2, antrasis leidimas. Raven Press, Niujorkas, p. 3 96. Melin, P., Kihlstrom, JE, 1963. Oksitocino įtaka vyrų triušių seksualiniam elgesiui. Endokrinologija 73, 433-435. Melis, MR, Argiolas, A., 1995a. Dopaminas ir seksualinis elgesys. Neurosci. Biobehav. Red 19, 19-38. Melis, MR, Argiolas, A., 1995b. Azoto oksido donorai sukelia varpos erekciją ir žaizdą, kai švirkščiami į centrinės nervų sistemos žiurkių patinus. E. J. Pharmacol. 294, 1-9. Melis, MR, Argiolas, A., 2003. Centrinė oksitocinerginė neurotransmisija: vaistas, skirtas psichogeninės erekcijos disfunkcijos gydymui. Curr. Narkotikų tikslai 4, 55-66. Melis, MR, Argiolas, A., Gessa, GL, 1986. Oksitocinų sukelta žandikaulio ir varpos erekcija: veikimo vieta smegenyse. Brain Res. 398, 259-265. Melis, MR, Argiolas, A., Gessa, GL, 1987. Apomorfino sukeltas žvėrys ir varpos erekcija: veikimo vieta smegenyse. Brain Res. 415, 98-104. Melis, MR, Argiolas, A., Gessa, GL, 1989a. Apomorfinas padidina oksitocino koncentraciją plazmoje žiurkėms. Neurosci. Lett. 98, 351-355. Melis, MR, Argiolas, A., Gessa, GL, 1989b. Įrodymai, kad apomorfinas sukelia varpos erekciją ir žiaurumą, atleidžiant oksitociną centrinėje nervų sistemoje. E. J. Phamacol. 164, 565-570. Melis, MR, Argiolas, A., Stancampiano, R., Gessa, GL, 1990. Apomorfino poveikis oksitocino koncentracijai skirtingose smegenų srityse ir patinų žiurkių plazmoje. E. J. Pharmacol. 182, 101-107. Melis, MR, Mauri, A., Argiolas, A., 1994a. Apomorfino ir oksitocino sukeltos varpos erekcija ir žvėrys žiurkių patekusiuose ir kastruotuose žiurkėse: seksualinių steroidų poveikis. Neuroendokrinologija 59, 349-354. Melis, MR, Stancampiano, R., Argiolas, A., 1994b. Paraventrikulinių NMDA injekcijų sukeltą varpos erekciją ir žvėrimą medijuoja oksitocinas. Pharmacol. Biochem. Behav. 48, 203-207. Melis, MR, Stancampiano, R., Argiolas, A., 1994c. Apomorfino ir oksitocino sukeltos varpos erekcijos ir žnyplės profilaktika NG-nitro-l-arginino metilesteriu: veikimo vieta smegenyse. Pharmacol. Biochem. Behav. 48, 799-804. Melis, MR, Succu, S., Argiolas, A., 1996. Dopamino agonistai padidina azoto oksido gamybą hipotalamo paraventrikuliniame branduolyje: koreliacija su varpos erekcija ir žiaurumu. E. J. Neurosci. 8, 2056-2063. Melis, MR, Succu, S., Iannucci, U., Argiolas, A., 1997. Oksitocinas didina azoto oksido gamybą hipotalamo paraventrikuliniame branduolyje: koreliacija su varpos erekcija ir žiaurumu. Reg. Peptidai 69, 105-112. Melis, MR, Succu, S., Mauri, A., Argiolas, A., 1998. Vyriškų žiurkių hipotalamijos paraventriculiniame branduolyje azoto oksido gamyba didėja nešvarios varpos erekcijos ir kopuliacijos metu. E. J. Neurosci. 10, 1968-1974. Melis, MR, Spano, MS, Succu, S., Argiolas, A., 1999a. Oksitocino antagonistas d (CH2) 5Tyr (Me) 2-Orn8-vazotocinas sumažina kontakto varpos erekciją žiurkių patinams. Neurosci. Lett. 265, 171-174. Melis, MR, Succu, S., Spano, MS, Argiolas, A., 1999b. Morfinas, švirkščiantis į hipotalamo paraventrikulinį branduolį, neleidžia susidurti su erekcija ir sutrikdo kopuliaciją: azoto oksido įtraukimas. E. J. Neurosci. 11, 1857-1864. Melis, MR, Spano, MS, Succu, S., Argiolas, A., 2000. Eksitacinių aminorūgščių, dopamino ir oksitocino receptorių antagonistų įtaka nevartojančiam varpos erekcijai ir paraventrikulinei azoto oksido gamybai žiurkių patinams. Behav. Neurosci. 114, 849-857. Melis, MR, Succu, S., Mascia, MS, Cortis, L., Argiolas, A., 2003. Vyresnio amžiaus žiurkių paraventriculiniame branduolyje padidėja nejautrinis dopaminas lytinio aktyvumo metu. E. J. Neurosci. 17, 1266-1272. Melis, MR, Succu, S., Mascia, MS, Argiolas, A., 2004a. Kannabinoidų CB1 receptorių antagonizmas patinų žiurkių paraventrikuliniame branduolyje sukelia varpos erekciją. Neurosci. Lett. 359, 17-20. Melis, MR, Succu, S., Mascia, MS, Cortis, L., Argiolas, A., 2004b. Ekstrahacinės aminorūgštys padidėja patinų žiurkių paraventrikuliniame branduolyje lytinio aktyvumo metu: pagrindinis NMDA receptorių vaidmuo erekcijos funkcijoje. E. J. Neurosci. 19, 2569-2575. Melis, MR, Succu, S., Mascia, MS, Argiolas, A., 2005. PD-168,077, selektyvus dopamino D4 receptorių agonistas, sukelia varpos erekciją, kai švirkščiamas į patinų paraventriculinį branduolį. Neurosci. Lett. 379, 59-62. Melis, MR, Succu, S., Mascia, MS, Sanna, F., Melis, T., Succu, S., Castelli, MP, Argiolas, A., 2006a. SR 141716A sukeltos varpos erekcijos žiurkėms: paraventrikulinės glutamo rūgšties ir azoto oksido dalyvavimas. Neurofarmakologija 50, 219-228. Melis, MR, Succu, S., Sanna, F., Mascia, MS, Melis, T., Enguehard-Gueiffier, C., Hubner, H., Gmenier, P., Gueiffier, A., Argiolas, A. 2006b. PIP3EA ir PD168077, du selektyvūs dopamino D4 receptorių agonistai, sukelia varpos erekciją patinų žiurkėms: veikimo vieta ir mechanizmas smegenyse. E. J. Neurosci. 24, 2021-2030. Melis, MR, Melis, T., Cocco, C., Succu, S., Sanna, F., Pillolla, G., Boi, A., Ferri, GL, Argiolas, A., 2007. Oksitocinas, švirkščiantis į ventralinę tegmentalinę sritį, sukelia varpos erekciją ir padidina ląstelių dopamino kiekį branduolyje. Melis, A. Argiolas / Neurotyros ir biologinės elgsenos apžvalgos 35 (2011) 939-955 955 ir vyrų žiurkių hipotalamo branduolys. E. J. Neurosci. 26, 1026-1035. Melis, MR, Sanna, F., Succu, S., Zarone, P., Boi, A., Argiolas, A., 2009a. Oksitocino vaidmuo numatant ir vartojant lyties žiurkių lytinio elgesio fazes. In: Jastrow, H., Feuerbach, D. Oksitocino tyrimų vadovas: sintezė, saugojimas ir išleidimas, veiksmai ir vaistų formos. Nova Publishers Inc, Niujorkas, JAV, pp. 109 125. Melis, MR, Succu, S., Sanna, F., Boi, A., Argiolas, A., 2009b. Oksitocinas, švirkščiantis į ventralinę subiculumą arba amterdalos posteromedialinį žievės branduolį, sukelia varpos erekciją ir padidina ekstraląstelinį dopaminą žiurkių patinų branduolyje. E. J. Neurosci. 30, 1349-1357. Melis, MR, Succu, S., Cocco, C., Caboni, E., Sanna, F., Boi, A., Ferri, GL, Argiolas, A., 2010. Oksitocinas sukelia varpos erekciją, kai švirkščiamas į ventralinį subiculum: azoto oksido ir glutamo rūgšties vaidmuo. Neurofarmakologija 58, 1153-1160. Moreland, RB, Nakane, M., Donnelly-Roberts, DL, Miller, LN, Chang, R., Uchic, ME, Terranova, MA, Gubbins, EJ, Helfrich, RJ, Namovic, MT, El-Kouhen, OF, Meistrai, JN, Brioni, JD, 2004. Žmogaus dopamino D (2) panašių receptorių stabilių ląstelių linijų, prijungtų prie kalcio srauto per Galpha (qo5), farmakologija. Biochem. Pharmacol. 68, 761-772. Moos, F., Freund-Mercier, MJ, Guerne, Y., Guerne, JM, Stoeckel, ME, Richard, P., 1984. Oksitocino ir vazopresino išsiskyrimas magnoceliuliniais branduoliais in vitro: specifinis oksitocino palengvinimo poveikis. J. Endokrinolis. 102, 63-72. Murphy, MR, Seckl, JR, Burton, S., Checkley, SA, Lightman, SL, 1987. Oksitocino ir vazopresino sekrecijos pokyčiai lytinio aktyvumo metu vyrams. J. Clin. Endokrinolis. Metab. 65, 738-741. Nishimori, K., Young, LJ, Guo, Q., Wang, Z., Insel, TR, Matzuk, MM, 1996. Oksitocinas reikalingas žindymui, bet nėra būtinas gimdymui ar reprodukciniam elgesiui. Proc. Natl. Acad. Sci. JAV 93, 11699-11704. Patel, S., Freedman, S., Chapman, KL, Emms, F., Fletcher, AE, Knowles, M., Marwood, R., Mccallister, G., Myers, J., Curtis, J., Kulagowski, JJ, Leeson, PD, Ridgill, M., Graham, M., Matheson, S., Rathbone, D., Watt, AP, Bristow, LJ, Rupniak, NM, Baskin, E., Lynch, JJ, Ragan, CI , 1997. L 745,870, selektyvaus antagonisto, turinčio didelį afinitetą dopamino D4 receptoriui, biologinis profilis. J. Pharmacol. Exp. Ther. 283, 636-647. Pedersen, CA, Caldwell, JD, Jirikowski, GF, Insel, TR, 1992. Oksitocinas motininėje, seksualinėje ir socialinėje veikloje, Niujorko mokslų akademijos leidiniai, t. 652. Niujorko mokslų akademija. Petrovic, P., Kalisch, R., Singer, T., Dolan, RJ, 2008. Oksitocinas susilpnina afektinius sąlyginių veidų ir amygdalos aktyvumo vertinimus. J. Neurosci. 28, 6607-6615. Pfaus, JG, Everitt, BJ, 1995. Lytinio elgesio psichofarmakologija. In: Knobil, FE, Kupfer, DJ Psichofarmakologija: ketvirtoji pažangos karta. Raven Press, Niujorkas, p. 742 758. Rajfer, J., Aronsonas, WJ, Bušas, PA, Dorey, FJ, Ignarro, LJ, 1992. Azoto oksidas kaip korpuso cavernosum atsipalaidavimo tarpininkas, reaguojant į neadrenerginį, nekolinerginį neurotransmisiją. N. Engl. J. Med. 326, 90-94. Roeling, TAP, Van Erp, AMM, Meelis, W., Kruk, MR, Veening, JG, 1991. NMDA elgsenos poveikis, suleistas į žiurkės hipotalaminę paraventrikulinę branduolį. Brain Res. 550, 220-224. Sachs, BD, 1997. Erekcija, atsiradusi žiurkių patinėlėse, įkvepia kvapą iš estrous patelių. Physiol. Behav. 62, 921-924. Sachs, BD, 2000. Kontekstiniai požiūriai į erekcijos funkcijos, erekcijos disfunkcijos ir seksualinio susijaudinimo fiziologiją ir klasifikaciją. Neurosci. Biobehav. Red 24, 541-560. Sachs, BD, 2007. Kontekstinis vyrų seksualinio susijaudinimo apibrėžimas. Horm. Behav. 51, 569-578. Sanchez, F., Alonso, JR, Arevalo, R., Blanco, E., Aijon, J., Vazquez, R., 1994. NADPH-diaforazės ir vazopresino ir oksitocino sambūvis žiurkės hipotalaminiuose magnoceluliniuose neurosekretoriniuose branduoliuose. Ląstelių audinių rez. 276, 31-34. Sanna, F., Succu, S., Boi, A., Melis, MR, Argiolas, A., 2009. Fosfodiesterazės tipo 5 inhibitoriai palengvina patekimo į erekciją patinų žiurkėms: veikimo vieta smegenyse ir veikimo mechanizmas. J. Lytis. Med. 6, 2680-2689. Saphier, D., Feldman, S., 1987. Sekalinių ir hipokampinių stimulų įtaka paraventrikuliniams branduolių neuronams. Neurologija 20, 749-755. Sato-Suzuki, I., Kita, I., Oguri, M., Arita, H., 1998. Stereotipiniai žandikaulio atsakai, atsiradę dėl žiurkės paraventrikulinės branduolio elektrinės ir cheminės stimuliacijos. J. Neurofiziolis. 80, 2765-2775. Schuman, EM, Madison, DV, 1994. Azoto oksido ir sinaptinės funkcijos. Ann. Red Neurosci. 17, 153-183. Snyder, SH, 1992. Azoto oksidas: pirmiausia naujoje neurotransmiterių klasėje? Mokslas 254, 494-496. Sofroniew, MV, 1983. Vasopresinas ir oksitocinas žinduolių smegenyse ir nugaros smegenyse. Tendencijos neurosci. 6, 467-472. Sokoloff, P., Schwartz, JC, 1995. Nauji dopamino receptoriai po pusės dešimtmečio. Trends Pharmacol. Sci. 16, 270-275. Southam, E., Garthwaite, J., 1993. Azoto oksido-ciklinis GMP signalizacijos kelias žiurkių smegenyse. Neurofarmakologija 32, 1267-1277. Stancampiano, R., Melis, MR, Argiolas, A., 1994. 5-HT1c agonistų sukeltas pūslinės erekcijos ir žvėrys žiurkių patelėms: ryšys su dopaminerginiu ir oksitocinerginiu transmisija. E. J. Pharmacol. 261, 149-155. Succu, S., Mascia, MS, Sanna, F., Melis, T., Argiolas, A., Melis, MR, 2006. Kannabinoidinis CB1 receptorių antagonistas SR 141716A sukelia varpos erekciją, padidindamas nešūnų glutamino rūgštį patinų žiurkių paraventrikuliniame branduolyje. Behav. Brain Res. 169, 274-281. Succu, S., Sanna, F., Melis, T., Boi, A., Argiolas, A., Melis, MR, 2007. Dopamino receptorių stimuliavimas vyrų žiurkių hipotalamijos paraventrikuliniame branduolyje sukelia varpos erekciją ir padidina išorinį dopaminą branduolyje accumbens: centrinio oksitocino dalyvavimas. Neurofarmakologija 52, 1034-1043. Succu, S., Sanna, F., Cocco, C., Melis, T., Boi, A., Ferri, GL, Argiolas, A., Melis, MR, 2008. Oksitocinas sukelia varpos erekciją, kai švirkščiamas į patinų žiurkių ventralinį apetitą: azoto oksido ir ciklinio GMP vaidmuo. E. J. Neurosci. 28, 813-821. Tanda, G., Pontieri, FE, Di Chiara, G., 1997. Cannabinoidai ir heroino aktyvacija mezolimbine dopamino transmisija, naudojant įprastą mu1 opioidų receptorių mechanizmą. Mokslas 276, 2048-2050. Tang, Y., Rampin, O., Calas, A., Facchinetti, P., Giuliano, F., 1998. Nustatyta lumbosakralinių branduolių oksitocinerginė ir serotoninerginė inervacija, kontroliuojanti varpos erekciją patinų žiurkėse. Neurologija 82, 241-254. Teodozė, DT, 1985. Oksitocino-imunoreaktyvūs terminalai sinapse apie oksitocino neuronus supraoptiniuose branduoliuose. Gamta (Londonas) 313, 682-684. Tindall, JS, 1974. Stimuliai, sukeliantys oksitocino išsiskyrimą. Geiger, SR, Knobil, E., Sawyer, WH, Greef, R., Astwood, EB (Red.), Fiziologijos vadovas. Sektas. 7, Endocrinology, t. IV. Amerikos fiziologijos draugija, Vašingtonas, p. 257 267. Torres, G., Lee, S., Rivier, C., 1993. Žiurkės hipotalaminio azoto oksido sintezės ontogenija ir kolokalizacija su neuropeptidais. Mol. Ląstelė Neurosci. 4, 155-163. Uhl-Bronner, S., Waltisperger, E., Martinez-Lorenzana, G., Condes, LM, Freund-Mercier, MJ, 2005. Oksitocino surišimo vietų seksualinė dimorfinė ekspresija žiurkių priešakyje ir nugaros smegenyse. Neurologija 135, 147-154. Vaccari, C., Lolait, SJ, Ostrowski, NL, 1998. Lyginamasis vazopresino V1b ir oksitocino receptorių ribonukleino rūgščių pasiskirstymas smegenyse. Endokrinologija 139, 5015-5033. Van Den Pol, A., 1991. Glutamatas ir aspartato imunoreaktyvumas hipotalaminių presinaptinių axonų atveju. J. Neurosci. 11, 2087-2101. Veronneau-Longueville, F., Rampin, O., Freund-Mercier, MJ, Tang, Y., Calas, A., Marson, L., McKenna, KE, Stoeckel, ME, Benoit, G., Giuliano, F. , 1999. Autonominių branduolių oksitocinerginė inervacija, kontroliuojanti varpos erekciją žiurkėse. Neurologija 93, 1437-1447. Vincent, SR, Kimura, H., 1992. Azoto oksido sintezės histocheminis žemėlapis žiurkių smegenyse. Neurologija 46, 755-784. Wagner, CK, Clemens, LG, 1993. Neurofizino turintis kelias nuo hipotalamo paraventrikulinio branduolio iki luminio nugaros smegenų lytiškai dimorfinio branduolio. J. Comp. Neurolis. 336, 106-116. Winslow, JT, Insel, TR, 1991. Vyrų voverės beždžionių porų socialinė padėtis lemia elgesio atsaką į centrinį oksitocino vartojimą. J. Neurosci. 11, 2032-2038. Wise, RA, Rompre, P.-P., 1989. Smegenų dopaminas ir atlygis. Ann. Red Psychol. 40, 191-225. Witt, DM, Insel, TR, 1994. Vyrų seksualinis elgesys aktyvuoja c-fos panašų baltymą oksitocino neuronuose hipotalamo paraventrikuliniame branduolyje. J. Neuroendokrinolis. 6, 13-18. Witter, MP, 2006. Žiurkės subiculumo jungtys: topografija, susijusi su kolonos ir laminarine organizacija. Behav Brain Res. 174, 251-264. Woodruff, GN, Foster, AC, Gill, R., Kemp, JA, Wong, EH, Iversen, LL, 1987. MK-801 ir N-metil-d-aspartato receptorių sąveika: funkcinės pasekmės. Neurofarmakologija 26, 903-909. Yamashita, H., Shigeru, O., Inenaga, K., Kasai, M., Uesugi, S., Kannan, H., Kaneko, T., 1987. Oksitocinas in vitro dažniausiai sužadina galimus oksitocino neuronus žiurkių supraoptiniame branduolyje. Brain Res. 416, 364-368. Yells, DP, Hendricks, SE, Prendergast, MA, 1992. Branduolio paragigantocellularis pažeidimai: poveikis patinų elgesiui su žiurkėmis. Brain Res. 596, 73-79. Young, WS, Shepard, E., Amico, J., Hennighausen, L., LaMarca, ME, McKinney, C., Ginns, EI, 1996. Pelės oksitocino trūkumas užkerta kelią pieno išmetimui, bet ne vaisingumui ar gimdymui. J. Neuroendokrinolis. 8, 847-854. Zahran, AR, Vachon, P., Courtois, F., Carrier, S., 2000. Padidėjęs pūslelinio spaudimo padidėjimas po stimuliuojančių aminorūgščių receptorių agonistų injekcijos anestezuotų žiurkių hipotalaminėje branduolinėje branduolyje. J. Urol.
