- 1Centro Interdisciplinario de Neurociencias de Valparaíso, Wydział Nauk, Universidad de Valparaíso, Valparaíso, Chile
- 2Departamento de Neurociencia, Facultad de Medicina, Universidad de Chile, Santiago, Chile
- 3Núcleo Milenio NUMIND Biologia zaburzeń neuropsychiatrycznych, Universidad de Valparaíso, Valparaíso, Chile
- 4Katedra Biologii Komórkowej i Molekularnej, Wydział Nauk Biologicznych, Pontificia Universidad Católica de Chile, Santiago, Chile
- 5Katedra Farmacji i Interdyscyplinarne Centrum Neuronauki, Wydział Chemii, Pontificia Universidad Católica de Chile, Santiago, Chile
Siła zachowań ukierunkowanych na cel jest regulowana przez neurony dopaminy śródmózgowia. Dysfunkcje obwodów dopaminergicznych obserwuje się w uzależnieniu od narkotyków i zaburzeniu obsesyjno-kompulsyjnym. Zachowanie kompulsywne jest cechą, która łączy oba zaburzenia, co jest związane ze zwiększoną neurotransmisją dopaminy. Aktywność neuronów dopaminy śródmózgowia jest regulowana głównie przez działanie homeostatyczne dopaminy poprzez receptory D2 (D2R), które zmniejszają odpalanie neuronów, a także syntezę i uwalnianie dopaminy. Transmisja dopaminy jest również regulowana między innymi przez heterologiczne układy neuroprzekaźników, takie jak układ opioidowy kappa. Znaczna część naszej obecnej wiedzy o systemie opioidowym kappa i jego wpływie na przekazywanie dopaminy pochodzi z przedklinicznych zwierzęcych modeli chorób mózgu. W 1988 r., Stosując mikrodializę mózgową, wykazano, że ostra aktywacja receptorów opioidowych Kappa (KOR) obniża poziom synaptyczny dopaminy w prążkowiu. Ten hamujący wpływ KOR przeciwstawia się ułatwianiu wpływu nadużywanych leków na uwalnianie dopaminy, co prowadzi do zaproponowania zastosowania agonistów KOR jako terapii farmakologicznej dla kompulsywnego przyjmowania leków. Co zaskakujące, 30 lat później proponuje się antagonistów KOR w leczeniu uzależnienia od narkotyków. Co mogło się wydarzyć w ciągu tych lat, które spowodowały drastyczną zmianę paradygmatu? Zebrane dowody sugerują, że wpływ KOR na poziomy synaptycznej dopaminy jest złożony, w zależności od częstotliwości aktywacji KOR i czasu z innymi przychodzącymi bodźcami do neuronów dopaminy, a także różnic płci i gatunków. W przeciwieństwie do jego ostrego działania, przewlekła aktywacja KOR wydaje się ułatwiać neurotransmisję dopaminy i zachowania zależne od dopaminy. Przeciwstawne działania wywierane przez ostrą kontra przewlekłą aktywację KOR były związane z początkowym efektem awersyjnym i opóźnionym nagradzaniem podczas narażenia na narkotyki. Zachowania kompulsywne indukowane przez powtarzaną aktywację D2R są również nasilane przez przedłużoną koaktywację KOR, która koreluje ze zmniejszonymi poziomami synaptycznymi dopaminy i uczulonym D2R. Zatem zależna od czasu aktywacja KOR wpływa bezpośrednio na poziomy dopaminy, wpływając na dostrajanie zachowań motywowanych. W tym przeglądzie analizuje się wkład systemu opioidowego kappa w korelacje dopaminergiczne zachowań kompulsywnych.
Wprowadzenie
Układ dopaminergiczny w zachowaniach kompulsywnych
Przymus to niemożność samozaparcia w celu wykonania zwykłego działania o znanym wyniku, pomimo niekorzystnych konsekwencji (Robbins i in., 2012). Zachowania kompulsywne są cechami zaburzeń obsesyjno-kompulsyjnych (OCD) i uzależnienia od narkotyków, a także innych chorób psychicznych. Sprawdzanie zachowania jest bardzo częste w zaburzeniach spektrum obsesyjno-kompulsywnego charakteryzujących się ciągłym powtarzaniem określonej rutyny, w stereotypowy lub rytualny sposób (Williams i in., 2013). Szeroki zakres normalnych zachowań (np. Sprawdzanie, czyszczenie, mycie rąk itp.) Może przekształcić się w kompulsywne u pacjentów z OCD i ogólnie pojawia się w odpowiedzi na obsesyjne i niepokojące myśli wywołujące lęk. Podobnie poszukiwanie i spożywanie narkotyków staje się kompulsywne u osób uzależnionych, podobnie jak w przypadku zaburzeń obsesyjno-kompulsyjnych, lęk odgrywa kluczową rolę, wywołując kompulsywne spożywanie narkotyków przez doświadczonych narkomanów. Te same ograniczenia w przetwarzaniu nagród i kar są obserwowane w obu warunkach (Figee i in., 2016), co skłoniło niektórych autorów do dyskusji na temat OCD jako uzależnienia behawioralnego (Holden, 2001).
Jeden z możliwych mechanizmów prowadzących do zachowań kompulsywnych jest ujęty w teorii uzależnienia od bodźca uczulającego, polegającej na tym, że wzmocniona motywacja („pragnienie”) leku rozwija się podczas uzależnienia bez rozwijania wzmocnionego efektu przyjemności („lubienia”) (Berridge i wsp., 1989; Berridge i Robinson, 2016). Trwałe uczulenie obwodu nagrody / motywacji bierze udział w indukcji uczulenia motywacyjnego związanego z poszukiwaniem narkotyków. Obwód nagrody / motywacji składa się z neuronów dopaminy śródmózgowia substantia nigra (SN) i brzuszny obszar nakrywkowy (VTA), które celują odpowiednio w grzbietowe i brzuszne poziomy prążkowia. Neurony dopaminowe, które rzutują na brzuszne prążkowie lub jądro półleżące (NAc) tradycyjnie były związane z zachowaniami ukierunkowanymi na cel, podczas gdy neurony dopaminowe, które rzutują na prążkowate grzbietowe były związane z nabywaniem nawyków (Everitt i Robbins, 2005; Wise, 2009; Yager i in., 2015; Volkow i wsp., 2017).
Uczulenie obwodu nagrody / motywacji obserwuje się u gryzoni jako stopniowy wzrost aktywności ruchowej wywołany powtarzanym podawaniem potencjalnie uzależniającej stałej dawki leku (Pierce i Kalivas, 1997; Robinson i Berridge, 2001). Uczulenie ruchowe jest znośnym zjawiskiem, ponieważ można je zaobserwować po tygodniach, miesiącach, a nawet roku po odstawieniu leku (Robinson i Berridge, 1993). Wcześniej zasugerowano, że uczulenie obwodu nagrody / motywacji przyczynia się do kompulsywnego poszukiwania narkotyków (Robinson i Berridge, 1993). W związku z tym uczulenie ruchowe ułatwia samodzielne podawanie kokainy w celu przywrócenia (De Vries i in., 2002). Ponadto szczury z rozszerzonym dostępem do samodzielnego podawania kokainy wykazują większą odpowiedź ruchową na kokainę niż szczury z ograniczonym dostępem (Ferrario i in., 2005). Ponadto zmiany neurochemiczne leżące u podstaw uczulenia narządu ruchu na psychostymulanty obserwuje się również w kompulsywnym poszukiwaniu leków (Steketee i Kalivas, 2011; Giuliano i in., 2019). Dane te potwierdzają zaproponowaną wcześniej zgodność między uczuleniem narządu ruchu a kompulsywnym poszukiwaniem leku obserwowanym u ludzi (Robinson i Berridge, 1993; Vanderschuren i Kalivas, 2000). Mechanicznie powtarzane podawanie leków uzależniających uwrażliwia mezolimbiczne obwody dopaminowe, zwiększając neurotransmisję dopaminergiczną. Psychostymulanty, takie jak kokaina lub amfetamina, które blokują błonę komórkową transportera dopaminy (DAT), indukują duży wzrost dopaminy w przestrzeni synaptycznej w prążkowiu i NAc, aktywując w ten sposób ruchSteketee i Kalivas, 2011). Podobnie jak w przypadku uzależnienia od narkotyków, uczulenie obwodu nagrody / motywacji dopaminy przyczynia się do zachowań kompulsywnych obserwowanych w OCD. Rzeczywiście, wielokrotna aktywacja receptorów dopaminy D2 (D2R) jest wystarczająca, aby wywołać uczulenie ruchowe i zachowanie sprawdzające zarówno u szczurów, jak i myszy (Szechtman i in., 1998; Szechtman i in., 1999; Sun i wsp., 2019). Powtarzane podawanie chinpirolu, agonisty D2R / D3R, jest przyjętym modelem OCD, ponieważ podsumowuje ważność twarzy, poprzez zwiększenie kompulsywnego sprawdzania i stereotypowych zachowań, trafności prognostycznej, co widać po spadku zachowań kompulsywnych po przewlekłym leczeniu wychwytem serotoniny inhibitory (SRI) i poprawność konstrukcji, ponieważ struktury mózgu zaangażowane w tym modelu są wspólne z tymi w patologii (Stuchlik i in., 2016; Szechtman i in., 2017). Podsumowując, powtarzana aktywacja transmisji dopaminy, albo przez mechanizmy pre-synaptyczne (uwalnianie dopaminy) lub post-synaptyczne (aktywacja D2R) prowadzi do uczulenia narządu ruchu i zachowań kompulsywnych.
System opioidowy kappa jest jednym z najbardziej przeważających systemów kontrolujących transmisję dopaminy w obwodzie nagrody / motywacji. Dowody wskazują, że transmisja opioidów kappa przeciwdziała wpływowi dopaminy; ostra aktywacja receptorów opioidowych kappa (KOR) przeciwdziała aktywności lokomotorycznej indukowanej przez psychostymulanty (Gray i in., 1999). I odwrotnie, powtarzana aktywacja KOR utrzymuje i poprawia kompulsywne i nawykowe poszukiwanie narkotyków (Koob, 2013). Spożywanie narkotyków powoduje nadużywanie homeopatycznego opioidu kappa, prawdopodobnie przyczyniając się do negatywnych stanów emocjonalnych dysforii (Koob, 2013), powodując kompulsywne używanie narkotyków (Chavkin i Koob, 2016). W rzeczywistości blokada KOR zapobiegała przywróceniu nikotyny przez stres, ale nie przez narkotyki (Jackson i in., 2013), kokaina (Beardsley i in., 2005) i etanol (Sperling i in., 2010). Zgodnie z tym odkryciem, blokada KOR przywraca zmiany dopaminergiczne w grzbietowo-bocznym prążkowiu szczurów uczulonych na amfetaminę, bez zmiany ich wzmocnionej odpowiedzi ruchowej na lek (Azocar i in., 2019). Zatem system KOR wydaje się zwiększać wzmocnienie ujemne, zwiększając wartość leku. W OCD wzmocnienie negatywne jest wywoływane przez obsesje, które wzmacniają dany przymus, aby uniknąć tej obsesji. Chociaż nie zostało to bezpośrednio przetestowane, wzmocnienie negatywne może odgrywać rolę w uczuleniu na chinpirol. Rzeczywiście, D2R są zaangażowane w generowanie wzmocnienia ujemnego. Na przykład u myszy pozbawionych długiej izoformy D2R nie opracowano unikania miejsca w obszarze sparowanym z wycofaniem morfiny (Smith i wsp., 2002) i powtarzane leczenie chinpirolem w okresie abstynencji przywraca poszukiwanie kokainy i heroiny w paradygmacie automatycznego podawania, efekt związany z wrażliwą ruchliwością chinpirolu (De Vries i in., 2002), co sugeruje wspólne mechanizmy działania uczulającego na psychostymulant i uczulenie na chinpirol. Ponadto wprowadzenie klatki domowej, ale nie nowej klatki, na arenę otwartego pola zmniejsza uczulenie narządu ruchu i kompulsywne zachowanie kontrolne (Szechtman i in., 2001), wskazując, że wskazówki bezpieczeństwa / znane mogą konkurować z negatywnymi wskazówkami środowiskowymi, które sprzyjają uczuleniu. Podobnie jak w przypadku uczulenia indukowanego psychostymulantem, powtarzana aktywacja KOR ułatwia uczulenie narządu ruchu (Escobar i in., 2017) i obowiązkowe sprawdzanie zachowania (Perreault i in., 2007) wywołane przez wielokrotne podawanie chinpirolu. Nie wiadomo, czy ten efekt wzmacniający jest konsekwencją wzmocnionego wzmocnienia ujemnego.
Dokładna analiza przeprowadzona niedawno pokazuje, że wpływ układu kappa-opioidowego na transmisję dopaminergiczną jest złożony: zależy od zaangażowanego szlaku dopaminowego (Margolis i in., 2006; Margolis i in., 2008) oraz czas między aktywacją receptora KOR a aktywacją receptora dopaminy (Chartoff i in., 2016). Zgodnie z tą złożonością szeroko dyskutowano o potencjalnym zastosowaniu terapeutycznym ligandów KOR. Zaproponowano, że agonista KOR może być klinicznie użyteczny podczas fazy zażywania narkotyków, łagodząc hiperdopaminergię wywołaną przez lek (Shippenberg i wsp., 2007). Z drugiej strony, antagonista KOR może być przydatny w leczeniu zespołu odstawienia wywołanego wzrostem ekspresji dynorfin po wielokrotnym spożywaniu leku (Wee and Koob, 2010). W związku z tym zaproponowano, aby częściowy agonista KOR (Béguin i in., 2012) może być terapeutyczną opcją leczenia zarówno kompulsywnego przyjmowania narkotyków, jak i objawów odstawienia u osób uzależnionych (Chartoff i in., 2016; Callaghan i in., 2018). W tym przeglądzie analizujemy modulację zależną od czasu / kontekstu dopaminergicznych korelatów uczulenia behawioralnego i kompulsywności.
Anatomiczny i funkcjonalny przesłuch między opioidami Kappa a układami dopaminergicznymi w regionach prążkowia i śródmózgowia
Regiony prążkowia
KOR to receptory sprzężone z białkami Gi / o silnie wyrażane w układzie dopaminowym śródmózgowia (Mansour i in., 1996). Receptory te należą do rodziny receptorów opioidowych złożonych z mu (MOR), delta (DOR) i kappa (KOR). Endogennymi agonistami tych receptorów są odpowiednio endorfiny, enkefalina i dynorfina. W prążkowiu dynorfina jest syntetyzowana przez średniej wielkości neurony (MSN) zawierające receptor dopaminy D1 (D1R), które mają nawracające aksony aktywujące KOR z tych samych jąder (Mansour i in., 1995). Obrazy mikroskopii elektronowej szczurzego NAc pokazują, że KOR znajdują się głównie w strukturach presynaptycznych zawierających DAT, podczas gdy niewielka część KOR lokalizuje się na dendrytach w połączeniu z DAT (Svingos i in., 2001; Kivell i in., 2014). Badania immunofluorescencyjne charakteryzujące preparaty presynaptyczno-synaptosomalne z NAc pokazują, że KOR i D2R preferencyjnie współistnieją w synaptosomach zawierających enzym syntetyzujący dopaminę, hydroksylazę tyrozynową (TH) (Escobar i in., 2017). Ponadto KOR są obfite w ciałach komórkowych NAc i prążkowia i kolokalizują z D2R w subpopulacji komórek (Escobar i in., 2017). Na podstawie wglądu genetycznego i molekularnego zasugerowano, że 20% całkowitego wiązania KOR w prążkowiu obserwuje się w terminalach DA (Van't Veer i in., 2013). Co więcej, Tejeda i in. (2017) wykazał, że zarówno MSN D1R, jak i D2R wyrażają KOR z wyższą preferencją wobec MSN zawierających D1R (Tejeda i in., 2017). Te dane anatomiczne wskazują, że KOR są obecne przed i postsynaptycznie, regulując neurotransmisję dopaminy w obwodzie nagrody / motywacji.
Kilka podejść eksperymentalnych pokazuje, że aktywacja KOR hamuje uwalnianie dopaminymi. Ostra aktywacja KOR przez wstrzyknięcie ogólnoustrojowe lub miejscowy wlew agonistów zmniejsza pozakomórkowe poziomy dopaminy w NAc (Di Chiara i Imperato, 1988; Spanagel i in., 1992; Fuentealba i in., 2006) i prążkowia grzbietowego (Gehrke i in., 2008). Wspierając toniczne hamujące działanie KOR na neurotransmisję dopaminy, bezpośredni wlew długotrwałego i selektywnego antagonisty KOR nor-binaltorfiminy (nor-BNI) (Broadbear i in., 1994) zwiększa podstawowe poziomy dopaminy w NAc (Spanagel i in., 1992) i uwalnianie dopaminy w prążkowiu grzbietowym (Azocar i in., 2019). Ostateczne dowody tonicznego hamowania KOR dopaminy pokazano u myszy z nokautem KOR, które wykazywały zwiększone poziomy zewnątrzkomórkowego poziomu dopaminy w prążkowiu i NAc (Chefer i in., 2005). Mechanizmy odpowiedzialne za hamowanie uwalniania dopaminy przez KOR nie są do końca wyjaśnione. Jednak dobrze wiadomo, że aktywacja KOR prowadzi do wzrostu K + i spadku przewodnictwa Ca2 +, indukując w ten sposób hiperpolaryzację komórek i blokadę uwalniania neuroprzekaźników pęcherzykowych (Bruchas i Chavkin, 2010; Margolis i Karkhanis, 2019).
do tegoin vitro i in vivo dane funkcjonalne sugerują, że KOR modyfikują poziomy zewnątrzkomórkowe dopaminy przez modulowanie aktywności DAT. Na przykład aktywacja KOR w komórkach EM4, które współeksprymują KOR i DAT, prowadzi do zwiększonego wychwytu dopaminy mierzonego woltammetrią (Kivell i in., 2014). Na ex vivo analiza wykorzystująca woltametrię w zdezagregowanych tkankach wykazała, że ogólnoustrojowe wstrzyknięcie agonisty KOR U-69593 zwiększało wychwyt dopaminy w NAc (Thompson i in., 2000). Podobny niedawny artykuł pokazuje, że nor-BNI blokuje wzrost wychwytu dopaminy w prążkowiu brzusznym i grzbietowym, wywołany ostrym układowym wstrzyknięciem MP1104, mieszanego agonisty receptora opioidowego Kappa / Delta (Atigari i in., 2019). Niemniej jednak wpływ aktywacji KOR na pobieranie dopaminy nie został jeszcze w pełni wyjaśniony. Układowe podawanie częściowego agonisty KOR, nalmefenu, zmniejszało zależnie od prążkowia dawkę wychwytu dopaminy zależnie, co określono ilościowo za pomocą cyklicznego woltamperometrii szybkiego skanowania (FSCV) (Rose i wsp., 2016). Stosując mikrodializę w strumieniu bez netto u dorosłych samców szczurów, blokowaniu KOR towarzyszył wzrost frakcji ekstrakcyjnej (Ed), który jest pośrednią miarą wychwytu dopaminy (Chefer i in., 2006; Azocar i in., 2019), co sugeruje, że toniczna aktywacja KOR wywiera hamującą kontrolę nad aktywnością DAT (wychwyt dopaminy). Wyniki te podkreślają złożoną rolę endogennej aktywności KOR w pobraniu dopaminy w kontrolowaniu poziomów zewnątrzkomórkowych dopaminy. Podejścia o wyższej rozdzielczości czasowej, takie jak FSCV, nie wykazały wpływu KOR na pobieranie dopaminy (Ebner i in., 2010; Ehrich i in., 2015; Hoffman i in., 2016), co sugeruje, że zwiększenie aktywności DAT w KOR w regionach prążkowia wymaga okresu inkubacji. Zwiększenie aktywności DAT za pośrednictwem KOR można wyjaśnić wzrostem liczby DAT na błonach komórkowych indukowanym przez aktywację KOR, jak opisano w synaptosomach i komórkach prążkowia linie (Kivell i in., 2014).
Regiony śródmózgowia
Testy autoradiograficzne przeprowadzone w śródmózgowiu szczura wykazują znaczące wiązanie KOR na osi rostokaudalnej SN i VTA (Speciale i wsp., 1993). Z drugiej strony dane z mikroskopii elektronowej pokazują, że terminale zawierające dynorfiny synapsują bezpośrednio na dendrytach dodatnich TH w SN i VTA (Sesack i Pickel, 1992), co sugeruje, że KOR lokalizują się w przedziałach somatodendrytycznych neuronów dopaminowych. MSN zawierające prążkowia D1R są jednym z dynorfinowych sygnałów wejściowych do neuronów dopaminy śródmózgowia. Co ciekawe, blokowanie KOR nie modyfikuje działania hamującego D1R-MSN na neurony dopaminy VTA, co wskazuje, że w tym hamowaniu pośredniczy GABA (Edwards i in., 2017). KOR modulują odpowiedzi somatodendrytyczne neuronów śródmózgowia dopaminy. Badania elektrofizjologiczne pokazują, że aktywacja KOR w VTA hiperpolaryzuje i zmniejsza spontaniczną szybkość wyzwalania neuronów dopaminowych (Margolis i in., 2003). W konsekwencji infuzja agonistów KOR zmniejsza somatodendrytyczny wypływ dopaminy (Smith i wsp., 1992; Dalman i O'Malley, 1999). Jednak ten hamujący wpływ KOR na neurony dopaminy wydaje się być zależny od obwodu. Wlew agonistów opioidowych kappa do VTA zmniejsza uwalnianie dopaminy w środkowej korze przedczołowej (mPFC) (Margolis i in., 2006), ale nie w NAc (Devine i in., 1993; Margolis i in., 2006). Co więcej, Margolis i in. (2006) odkrył, że KOR hamują neurony dopaminy VTA, które rzutują na mPFC i jądro migdałkowo-podstawne, ale nie te, które rzutują na NAc. W tym samym roku Ford i in. (2006) wykazał, że zastosowanie agonistów KOR w kąpieli w mysich skrawkach VTA indukowało większy prąd zewnętrzny w neuronach dopaminy, które rzutują na NAc w porównaniu z tymi, które rzutują na bazolateralne ciało migdałowate, wskazując, że KOR wywierają większe hamowanie neuronów dopaminowych, które rzutują na NAc niż do ciała migdałowatego. Ponadto aktywacja KOR zmniejsza amplitudę pobudzenia (Margolis i in., 2005) i hamujący (Ford i in., 2007) prądy postsynaptyczne w neuronach dopaminy śródmózgowia. Różnice między gatunkami i złożonymi efektorami prowokacji VTA na mPFC i NAc (Van Bockstaele i Pickel, 1995; Carr i Sesack, 2000) utrudniają ustalenie, czy KOR wybiórczo hamują niektóre neuronalne populacje dopaminy w VTA. Niemniej jednak podsumowane tutaj dane wskazują, że KOR znajdują się w somie i terminalach neuronów dopaminowych, a także w wejściach, które je regulują, dzięki czemu są doskonale ustawione do kontrolowania aktywności synaptycznej neuronów dopaminy śródmózgowia.
Rola KOR w kontrolowaniu neurotransmisji dopaminowej w uczuleniu i zachowaniach kompulsywnych wywołanych przez psychostymulatory
Uzależnienie od narkotyków to proces, który obejmuje początkowo impulsywne poszukiwanie narkotyków związane z ich pozytywnymi efektami wzmacniającymi. Z drugiej strony kompulsywność jest cechą osobowości obserwowaną u osób uzależnionych od narkotyków. Zaproponowano kilka neuroadaptacji w szlakach dopaminergicznych w celu uwzględnienia kompulsywnego poszukiwania i przyjmowania narkotyków po wielokrotnym narażeniu na narkotyki (Everitt i Robbins, 2005; Koob i Volkow, 2016). Jedną z proponowanych hipotez napędzających kompulsywne przyjmowanie leków jest uwrażliwienie na jego negatywne działanie wzmacniające (Koob, 2013). Hamująca kontrola układu opioidowego kappa w zakresie uwalniania dopaminy może przyczyniać się do wzmacniających negatywne właściwości nadużywania narkotyków. Jednak konsekwencje aktywacji KOR dla neurotransmisji dopaminy i poszukiwania kompulsywnych leków wydają się złożone i pozornie sprzeczne. Rzeczywiście, uwalnianie dopaminy indukowane przez amfetaminę i kokainę jest osłabiane przez jednoczesne podawanie agonistów KOR (Heidbreder i Shippenberg, 1994; Maisonneuve i in., 1994; Thompson i in., 2000), a nawet zmniejszyć samozastawianie kokainy (Negus i in., 1997). Ponadto KOR wywierają hamujące sprzężenie zwrotne na uwalnianie dopaminy szlaku mezolimbicznego w odpowiedzi na przedłużoną aktywację postsynaptycznej D1R, co ma miejsce przy wielokrotnym narażeniu na psychostymulanty (Cole i in., 1995; Nestler, 2001). Paradoksalnie, aktywacja KOR może również ułatwić uwalnianie dopaminy na ścieżce nagrody / motywacji (Fuentealba i in., 2006; Fuentealba i in., 2007) i konsumpcja psychostymulantów (Wee i in., 2009). Fuentealba i in. (2007) wykazał, że po czterech dniach podawania U69593, agonista KOR, zwiększył indukowane amfetaminą uwalnianie dopaminy w NAc. Niedawno wykazano, że blokowanie KOR odwraca zmiany uwalniania i wychwytu dopaminy w prążkowiu grzbietowym, które mają miejsce podczas uczulenia narządu ruchu indukowanego przez amfetaminę (Azocar i in., 2019). Podsumowując, dane te sugerują, że aktywacja KOR może również przyczynić się do pozytywnie wzmacniających właściwości narkotyków (Chartoff i in., 2016).
Ponadto wydaje się, że aktywacja KOR przyczynia się do kompulsywnego poszukiwania narkotyków; Blokada KOR zmniejsza kokainę (Wee i in., 2009), heroina (Schlosburg i in., 2013) i metamfetamina (Whitfield i in., 2015) spożycie u szczurów z nieograniczonym dostępem do leku (Wee i in., 2009). Efekt ten znajduje również potwierdzenie w poszukiwaniach leków wywołanych stresem. Na przykład myszy z nokautem KOR nie wykazywały preferencji miejsca kokainy po przymusowym stresie pływania (McLaughlin i in., 2006a). Blokowanie KOR osłabia preferencje dotyczące miejsca nikotynowego wywołane narażeniem na przymusowe pływanie (Smith i wsp., 2012). Co ciekawe, blokowanie KOR osłabia poszukiwanie kokainy i nikotyny wywołane stresem, ale nie wpłynęło na poszukiwanie wywołane prowokacją narkotykową (Beardsley i in., 2005; Jackson i in., 2013). Ten efekt KOR ułatwiający wywołany przez stres wydaje się pośredniczyć w obwodzie nagrody / motywacji (Shippenberg i wsp., 2007; Wee and Koob, 2010). W eleganckim badaniu przeprowadzonym przez dr Kauer i jej grupę wykazano, że blokowanie KOR w VTA, wcześniej lub po ostrym stresie, hamuje ponowne poszukiwania kokainy, efekt związany z ratowaniem długotrwałych nasilenie synaps hamujących w neuronach dopaminy (Graziane i in., 2013; Polter i in., 2014).
Wydaje się, że ułatwienie przyjmowania środków psychostymulujących przez KOR zależy od przedziału czasowego dotyczącego ekspozycji na narkotyki. Podanie agonisty KOR U50488 1 godzinę przed ekspozycją na kokainę zwiększa zarówno preferencję miejsca kokainy, jak i względne uwalnianie dopaminy wywołane przez kokainę w NAc, podczas gdy przeciwne efekty obserwuje się po podaniu 15 minut wcześniej (McLaughlin i in., 2006a; Ehrich i in., 2014). Korzystanie z samostymulacji wewnątrzczaszkowej Chartoff i in. (2016) zaobserwował, że agonista KOR, Salvinorin A, wykazuje początkowe działanie awersyjne i opóźnione nagradzanie, któremu towarzyszy odpowiednio spadek i wzrost stymulowanego uwalniania dopaminy odpowiednio w NAc. Wszystkie te dane wskazują na zależny od czasu wpływ aktywacji KOR na satysfakcjonujące właściwości kokainy i wskazują na aktywowaną przez stres aktywację KOR jako kluczowego gracza w rozwoju kompulsywnego poszukiwania narkotyków.
Indukowane chinpirolem uczulenie ruchowe i kompulsywne zachowanie
Fakty, że układ dopaminowy bierze udział w wytwarzaniu uczulenia i kompulsywności, są wzmacniane przez zachowanie obserwowane u gryzoni leczonych agonistą D2R, chinpirolem. W skrócie, D2R są receptorami sprzężonymi z Gi, szeroko wyrażanymi w obwodzie nagrody / motywacji; są wyrażane somatodendrytycznie i na końcach aksonowych neuronów dopaminowych (Sesack i in., 1994), a jego aktywacja obniża poziomy zewnątrzkomórkowe dopaminy (Imperato i Di Chiara, 1988). W prążkowiu D2R są również zlokalizowane postsynaptycznie na średnich neuronach kolczastych (Sesack i in., 1994), a jego aktywacja hamuje szlak pośredni umożliwiający aktywność ruchową.
Dr Henry Szechtman zaczął badać wpływ chinpirolu na zachowanie szczurów pod koniec dekady 1980. Ich wstępne odkrycia wykazały, że ostre podanie chinpirolu ma zależny od dawki wpływ na aktywność lokomotoryczną. Przy małych dawkach (0.03 mg / kg) obniża aktywność lokomotoryczną, a przy wyższych (> 0.5 mg / kg) zwiększa. (Eilam i Szechtman, 1989). Efekty te związane są odpowiednio z aktywacją presynaptycznych D2R o wysokim powinowactwie i postsynaptycznych D2R o niskim powinowactwie (Usiello i in., 2000). Nieoczekiwanie powtarzane (co drugi dzień) podawanie chinpirolu powoduje stopniowy i trwały wzrost ruchliwości, przypominający uczulenie narządu ruchu wywołane przez psychostymulanty (Szechtman i in., 1993; Szechtman i in., 1994). Wykazano, że działanie uczulające na ruch zależy od D2R, ponieważ myszy z niedoborem tego receptora nie rozwijają uczulenia na ruch na chinpirol (Escobar i in., 2015).
Na początku lat 90. Szechtman i Eilam donosili, że wraz z uczuleniem narządów ruchu u szczurów rozwinęło się stereotypowe zachowanie, które jest wzmacniane po każdym podaniu chinpirolu (Eilam i Szechtman, 1989; Szechtman i in., 1993). Obecnie wielokrotne podawanie chinpirolu jest sprawdzonym modelem OCD (Szechtman i in., 1999; Szechtman i in., 2001; Eilam i Szechtman, 2005; Stuchlik i in., 2016; Szechtman i in., 2017), w oparciu o spostrzeżenie, że zachowanie szczurów staje się coraz bardziej ustrukturyzowane i nieelastyczne, co przypomina rytualne zachowanie charakterystyczne dla zachowania nałogowego sprawdzania (Szechtman i in., 1998; Szechtman i in., 2017). Ostatnie badania pokazują, że powtarzany chinpirol wywołuje również zachowania kompulsywne u myszy, takie jak kompulsywne sprawdzanie (Sun i wsp., 2019), sztywność behawioralna i kompulsywne żucie (Asaoka i in., 2019), ten ostatni został cofnięty przez blokadę D2R w prążkowiu, co dodatkowo potwierdza, że wielokrotna aktywacja D2R jest konieczna do wywołania zachowań kompulsywnych. Razem dane wskazują na kluczową rolę D2R w szlakach dopaminy śródmózgowia w indukowaniu uczulenia narządu ruchu i kompulsywności. Powtarzane podawanie chinpirolu powoduje stereotypowe zachowanie wywołane kokainą (Thompson i in., 2010) i efekty ruchowe amfetaminy (Cope i wsp., 2010), wzmacniając ideę, że aktywacja D2R leży u podstaw uczulenia indukowanego psychostymulantem i sugeruje wspólny mechanizm między chinpirolem i uczuleniem indukowanym psychostymulantami. Co ciekawe, efekt uczulający powtarzanej aktywacji D2R wydaje się być silniejszy niż efekt wywołany przez psychostymulanty, ponieważ u każdego szczura leczonego chinpirolem występuje uczulenie ruchowe (Escobar i in., 2015), podczas gdy około sześćdziesiąt procent szczurów uczula na amfetaminę (Escobar i in., 2012; Casanova i in., 2013).
Uczuleniu behawioralnemu indukowanemu przez powtarzaną aktywację D2R towarzyszą dostosowania w obwodzie nagrody / motywacji. Szczury uczulone chinpirolem mają niższy ton dopaminergiczny w NAc, obserwowany jako obniżony poziom podstawowy (Koeltzow i in., 2003) oraz stymulowane toniczne i fazowe uwalnianie dopaminy (Escobar i in., 2015), co wskazuje na zmniejszoną zdolność uwalniania dopaminy w dopaminoprądowym obwodzie śródmózgowia. Synaptyczne poziomy dopaminy w NAc są kontrolowane przez aktywność zarówno DAT, jak i neuronów dopaminy (Goto and Grace, 2008), który in vivo składa się z wypalania tonikiem i seriami (Wilson i wsp., 1977; Grace i Bunney, 1980). Poprzednie raporty pokazują, że szczury uczulone na chinpirol wykazują mniejszą liczbę neuronów dopaminy w tonice i strzelaniu serią w VTA (Sesia i in., 2013). Razem te dane wskazują, że spadek uwalniania dopaminy obserwowany po uczuleniu na chinpirol jest wynikiem zmniejszenia ogólnej aktywności neuronów dopaminowych. Zachowanie kompulsywne i uwrażliwiona aktywność lokomotoryczna wywołana powtarzanym leczeniem chinpirolem może być konsekwencją uczulenia D2R z powodu obniżonego napięcia dopaminergicznego w NAc. Rzeczywiście, uczulone na chinpirol szczury wykazują wzrost wiązania dopaminy D2R (Culver i in., 2008) i wzrost stanu powinowactwa tych receptorów (Perreault i in., 2007), potwierdzając tę hipotezę.
Interakcje KOR-dopamina w zachowaniach kompulsyjnych wywołanych chinpirolem
Wstępne badania dotyczące roli KOR w zachowaniach kompulsywnych wywołanych przez D2R również pochodziły z laboratorium Szechtmana. Ta grupa badała jednoczesne podawanie agonisty KOR U69593 z chinpirolem na aktywność lokomotoryczną. W szczególności autorzy podawali szczurom zastrzyki podskórne z mieszaniną U69593 i chinpirolu, aż do zakończenia 8 do 10 wstrzyknięć. W przeciwieństwie do hipolokomotorycznego działania samego U69593, hiperlokomocję obserwowano podczas jednoczesnego podawania z niskimi (presynaptycznymi) i dużymi (postsynaptycznymi) dawkami chinpirolu. U69593 zmienił hipolokomotoryczny efekt presynaptycznej dawki chinpirolu na hiperlokomocję i wzmocnił efekt hiperlokomotoryczny postsynaptycznej dawki chinpirolu (Perreault i in., 2006). Jednoczesna aktywacja KOR przyspieszyła również indukcję uczulenia narządu ruchu i potęgowała efekt aktywacji D2R, ponieważ maksymalna lokomocja osiągnięta dzięki podwójnemu leczeniu duplikuje efekt lokomotoryczny wywołany przez sam chinpirol (Perreault i in., 2006; Escobar i in., 2017). Jednoczesna aktywacja KOR przyspiesza również zachowanie kompulsywnego sprawdzania (Perreault i in., 2007). Te wzmacniające działanie KOR na zachowania wywołane chinpirolem wymagają powtarzanej aktywacji KOR. W rzeczywistości, ostre wstrzyknięcie agonisty KOR U69593 nie zmieniło dalej aktywności lokomotorycznej u szczurów uczulonych chinpirolem (Escobar i in., 2017). Mechanizm nasilania KOR uczulenia indukowanego D2R jest nieznany. Jedną z możliwości jest to, że sam endogenny system opioidowy kappa pośredniczy w uczulaniu zależnym od D2R. Jednak możliwość tę odrzucono, pokazując, że wcześniejsze podanie norBNI nie zmodyfikowało uczulenia ruchowego na chinpirol, co sugeruje, że dynorfina nie jest uwalniana po aktywacji D2R (Escobar i in., 2017). Dane te nie wykluczają, że dynorfina może odgrywać rolę w uczulaniu zachowań kompulsywnych, na przykład stres indukuje uwalnianie dynorfin i aktywację KOR, co ułatwia zachowania kompulsywne (McLaughlin i in., 2003; McLaughlin i in., 2006a; McLaughlin i in., 2006b).
Przesłuch między D2R i KOR jest złożony i wydaje się zależeć od tego, czy aktywacja obu receptorów jest przypadkowa, czy czasowo oddzielona. Dane anatomiczne wskazują, że przesłuch między D2R i KOR może występować presynaptycznie w aksonach i somie neuronów dopaminowych, a także postsynaptycznie w MSN prążkowia. Chociaż nie wyklucza to roli KOR zlokalizowanych na aksonach innych układów neurochemicznych, dane anatomiczne silnie wskazują na bezpośrednią rolę KOR regulujących D2R. Aktywacja KOR, ostra lub powtarzana, zmniejsza hamującą funkcję D2R na neuronach dopaminy. Badania elektrofizjologiczne wykazały, że ostra aktywacja KOR w neuronach dopaminowych VTA i SN hamuje hamujący prąd postynaptyczny za pośrednictwem D2R, efekt za pośrednictwem mechanizmów pre i postsynaptycznych, gdy KOR zmniejsza uwalnianie dopaminy, a dynorfina blokuje hamujące działanie dopaminy stosowanej w kąpieli (Ford i in., 2007). Badania neurochemiczne wykazały, że powtarzająca się aktywacja KOR blokuje indukowane przez D2R hamowanie uwalniania dopaminy w NAc (Fuentealba i in., 2006). Co więcej, jednoczesna aktywacja ostra D2R i KOR zmniejsza hamowanie uwalniania dopaminy w NAc w porównaniu z działaniem każdego receptora osobno (Escobar i in., 2017). Zatem presynaptyczne KOR nie działają addytywnie lub synergicznie z presynaptycznymi D2R, i odwrotnie, KOR albo hamują, albo blokują działanie hamujące D2R. Mechanizm ten może wyjaśnić działanie aktywujące narządu ostrej dawki agonistów KOR jednocześnie z niską dawką chinpirolu (Perreault i in., 2006).
Ostatnie badanie pokazuje, że aktywacja KOR w VTA pośredniczy w zachowaniach kompulsywnych mierzonych jako hamowanie behawioralne i zakopywanie marmuru (Abraham i in., 2017), wzmacniając pogląd, że aktywacja KOR jest rzeczywiście wyzwalaczem kompulsywności. Dane opublikowane przez Margolis i in. (2006; 2008) wskazują, że interakcje KOR i D2R powinny mieć miejsce na neuronach dopaminy atakujących mPFC (Margolis i in., 2006; Margolis i in., 2008). Jednakże, Ford i in. (2006; 2007) stwierdzono, że hamowanie KOR IPSC za pośrednictwem D2R zachodzi na neuronach dopaminy celujących w NAc (Ford i in., 2006; Ford i in., 2007). Razem dane te pokazują, że interakcja KOR z D2R w przedziale somatodendrytycznym neuronów dopaminy może powstać w wyniku przesłuchu w tym samym neuronie dopaminy. To, czy dzieje się tak w projekcjach mezolimbicznych, czy mezokortykalnych, jest nadal kontrowersyjne.
Co zaskakujące, KOR znaleziono w MSN NAc (Escobar i in., 2017; Tejeda i in., 2017), wskazując w ten sposób, że nasilenie zachowań kompulsywnych indukowanych przez D2R może również wynikać z bezpośrednich działań na komórki docelowe neuronów dopaminowych. W tym względzie warto wspomnieć, że wielokrotne podawanie U69593 zwiększa ilość D2R w stanie wysokiego powinowactwa (Perreault i in., 2007). Dane neurochemiczne wskazują, że obniżony poziom zewnątrzkomórkowej dopaminy jest związany z uczuleniem na D2R. Koaktywacja KOR nie zmniejsza dalszego poziomu zewnątrzkomórkowego dopaminy w NAc już obniżonego przez powtarzaną aktywację D2R (Escobar i in., 2017), wykluczając rolę presynaptycznych KOR przyspieszających lub wzmacniających uczulenie D2R w NAc poprzez ten mechanizm. Dlatego KOR wyzwalają powolne mechanizmy molekularne, które dodatkowo uczulają neurochemiczne i behawioralne efekty D2R, sugerując, że zwiększenie uczulenia na ruch może być spowodowane raczej adaptacyjnym efektem postsynaptycznym niż presynaptycznym. W związku z tym powtarzana aktywacja KOR może wywołać hamowanie pośredniego szlaku prążkowia D2R przełączającego równowagę D1R / D2R na kompulsywność D1R (Rysunek 1).
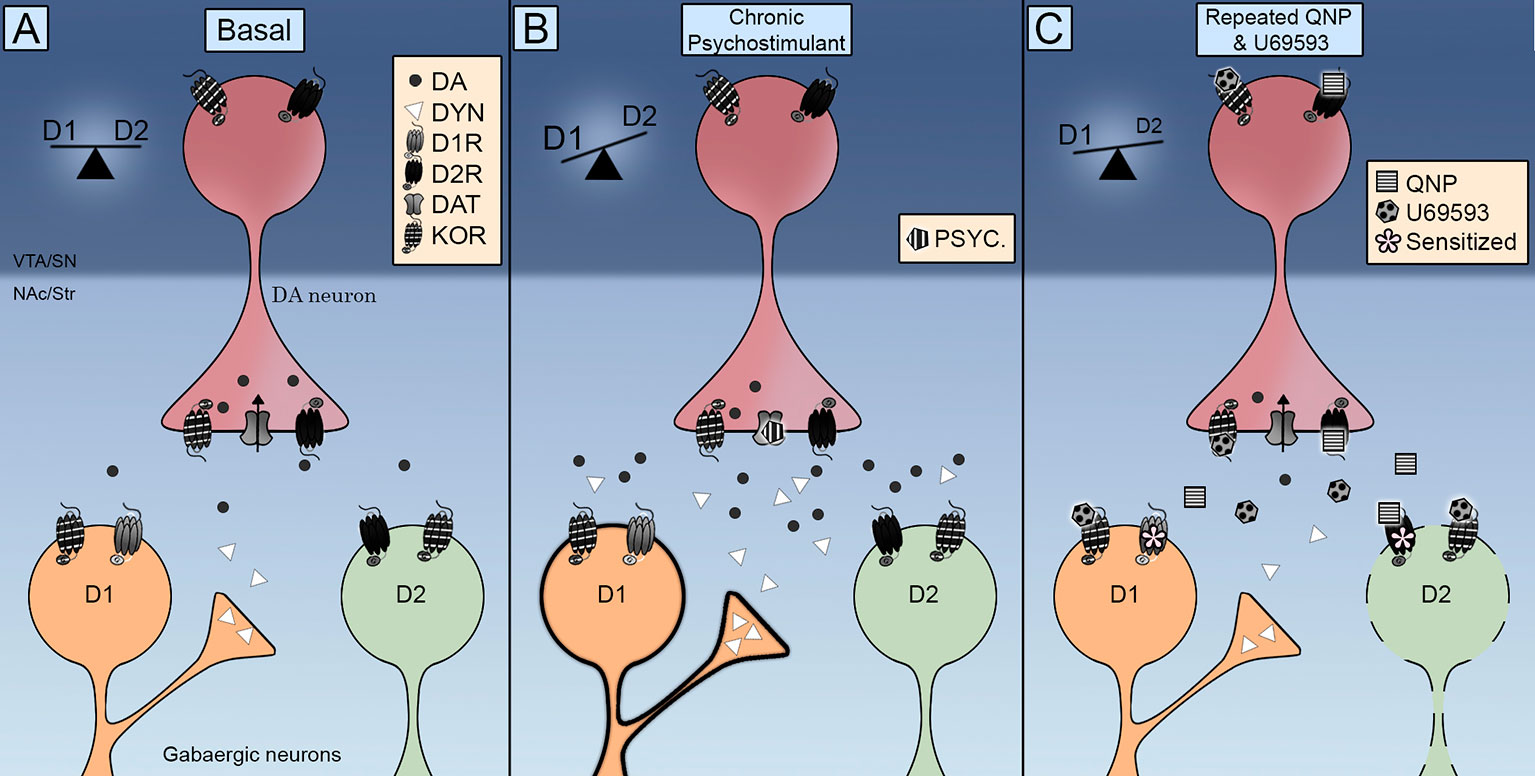 Rysunek 1 Schemat integracji receptorów opioidowych Kappa (KOR) na bezpośrednich (D1R) i pośrednich (D2R) przejściach prążkowia. () KOR są umiejscowione przedsynaptycznie na terminalach dopaminowych i postsynaptycznie w średnich neuronach (MSN). Jego aktywacja kontroluje poziomy zewnątrzkomórkowe dopaminy, a jego lokalizacja promuje interakcję z transporterem dopaminy (DAT) i receptorami dopaminy D2. (B) Powtarzającej się ekspozycji na psychostymulant towarzyszy wzrost zarówno poziomu zewnątrzkomórkowego dopaminy, jak i dynorfiny. Aktywacja receptorów D1 i D2 przełącza równowagę na bezpośredni szlak D1R, promując uczulenie ruchowe. (DO) Jednoczesne podawanie chinpirolu i U69593 towarzyszy obniżeniu poziomu zewnątrzkomórkowego dopaminy. Jednoczesna aktywacja receptorów KOR i D2 osłabia pośredni szlak D2 indukując kompulsywne zachowanie.
Rysunek 1 Schemat integracji receptorów opioidowych Kappa (KOR) na bezpośrednich (D1R) i pośrednich (D2R) przejściach prążkowia. () KOR są umiejscowione przedsynaptycznie na terminalach dopaminowych i postsynaptycznie w średnich neuronach (MSN). Jego aktywacja kontroluje poziomy zewnątrzkomórkowe dopaminy, a jego lokalizacja promuje interakcję z transporterem dopaminy (DAT) i receptorami dopaminy D2. (B) Powtarzającej się ekspozycji na psychostymulant towarzyszy wzrost zarówno poziomu zewnątrzkomórkowego dopaminy, jak i dynorfiny. Aktywacja receptorów D1 i D2 przełącza równowagę na bezpośredni szlak D1R, promując uczulenie ruchowe. (DO) Jednoczesne podawanie chinpirolu i U69593 towarzyszy obniżeniu poziomu zewnątrzkomórkowego dopaminy. Jednoczesna aktywacja receptorów KOR i D2 osłabia pośredni szlak D2 indukując kompulsywne zachowanie.
Różnice płciowe interakcji KOR-dopamina w zachowaniach kompulsywnych
Badania kliniczne wykazały różnice płci w zachowaniu kompulsywnym, w tym kompulsywnym poszukiwaniu narkotyków. Wcześniejsze objawy OCD obserwuje się u mężczyzn w porównaniu z kobietami (Mathis i in., 2011), przy czym kobiety częściej wykazują objawy zanieczyszczenia i czyszczenia (Labad i in., 2008). Jeśli chodzi o różnice płci w uzależnieniu od narkotyków, dowody kliniczne wskazują, że chociaż stosowanie narkotyków jest bardziej rozpowszechnione u mężczyzn, kobiety wykazują szybszy postęp niż mężczyźni w kompulsywnym poszukiwaniu narkotyków (Hernandez-Avila i in., 2004; Fattore i Melis, 2016).
Ostatnio dane przedkliniczne silnie podkreśliły podstawy neurobiologiczne leżące u podstaw różnic płci w nadużywaniu narkotyków zaobserwowanych w badaniach klinicznych (Becker i Chartoff, 2019). Wczesne obserwacje z wykorzystaniem mikrodializy z przepływem bez sieci wykazały, że zewnątrzkomórkowe stężenie dopaminy w prążkowiu grzbietowym zmienia się podczas cyklu rujowego z wyższymi poziomami w proestrus i rui w porównaniu z diestrusem. Ponadto, podczas gdy wycięcie jajników zmniejsza pozakomórkowe stężenie prążkowia dopaminy u samic szczurów, kastracja samców szczurów nie modyfikuje pozakomórkowego stężenia prążkowia zewnątrzkomórkowego (Xiao i Becker, 1994), co sugeruje ważną rolę hormonów jajnikowych w aktywności dopaminy. Ponadto hormony żeńskie regulują odpowiedź na środki psychostymulujące. Wcześnie in vitro eksperymenty wykazały, że estradiol i progesteron przywracają indukowane amfetaminą uwalnianie dopaminy z tkanki prążkowia uzyskanej od samic szczurów po usunięciu jajników (Becker i Ramirez, 1981). Niedawno szybkie cykliczne badania woltammetrii z szybkim skanowaniem wykazały, że kobiety wykazują większe stymulowane elektrycznie uwalnianie i pobieranie dopaminy w porównaniu do mężczyzn (Walker i wsp., 2000). Te różnice płciowe w neuroprzekaźnictwie dopaminy mogą tłumaczyć większe poszukiwanie kokainy i amfetaminy obserwowane u kobiet. (Roberts i wsp., 1989; Cox i in., 2013).
Regulacja KOR w zakresie poziomów zewnątrzkomórkowych dopaminy pokazuje również różnice płci (Chartoff i Mavrikaki, 2015). Stosując wewnątrzczaszkową samostymulację i cykliczną woltametrię, Conway i in. (2019) wykazał, że niższej wrażliwości na ostry efekt anhedoniczny agonisty KOR obserwowanej u samic szczurów w porównaniu do samców szczurów towarzyszy osłabione hamowanie stymulowanego uwalniania dopaminy w NAc (Conway i in., 2019). Sugeruje się, że estradiol przyczynia się do stępionego hamowania uwalniania dopaminy obserwowanego u samic szczurów po aktywacji KOR (Abraham i in., 2018). Podczas gdy przesłuchy między KOR a sygnalizacją dopaminową badano u mężczyzn (Tejeda i Bonci, 2019), brakuje badań dotyczących tej interakcji i jej wpływu na proces uzależnienia u kobiet (Chartoff i Mavrikaki, 2015). U samic szczurów ostre podanie agonisty KOR U69593 osłabiło hiperlokomocję indukowaną kokainą zarówno u szczurów kontrolnych, jak i jajników. Co ciekawe, wielokrotne podawanie U69593 osłabiło indukowaną kokainą hiperlokomocję w sposób zależny od estradiolu (Puig-Ramos i in., 2008). Dane te sugerują, że estradiol stymuluje działania KOR u samic szczurów, co może być związane z różnicami płciowymi w reakcji na stres (Puig-Ramos i in., 2008). To, czy u samic szczurów powtarzana aktywacja KOR ułatwia prążkowskie uwalnianie dopaminy, jak zaobserwowano u samców, jest pytaniem bez odpowiedzi.
Chociaż ułatwienie w indukowanym przez psychostymulanty uwalnianiu dopaminy obserwuje się u samic w porównaniu z samcami szczurów, różnice płciowe w mechanizmach dopaminy leżących u podstaw uczulenia na ruch amfetaminy nie zostały w pełni wyjaśnione (Becker, 1999). Powtarzająca się ekspozycja na amfetaminę indukuje większą aktywność ruchową u obu nastolatków (Mathews i McCormick, 2007) i dorosłe samice szczurów (Milesi-Hallé i in., 2007), przy czym samice szczurów młodocianych wykazują silniejsze uczulenie ruchowe po wielokrotnym narażeniu na amfetaminę. Aktywacja receptora D2 u noworodków nasilała uczulenie behawioralne indukowane amfetaminą tylko u samic szczurów (Brown i wsp., 2011). Jak wspomniano wcześniej, zaobserwowano u samców szczurów, że wielokrotne narażenie na agonistę D2 indukuje uczulenie ruchowe i zachowanie podobne do kompulsywnego (Dvorkin i in., 2006). Co więcej, koaktywacja KOR nasila uczulenie ruchowe indukowane przez powtarzane narażenie na chinpirol, ułatwiając kontrolę hamowania receptorów D2 podczas uwalniania DA w NAc (Escobar i in., 2017). Różnice płciowe, takie jak zaobserwowana niższa wrażliwość na hamujący wpływ KOR na uwalnianie dopaminy u kobiet (Conway i in., 2019) może uwzględniać zróżnicowany wkład KOR w kompulsywne poszukiwanie narkotyków.
wnioski
W jaki sposób KOR modulują sygnalizację dopaminową w celu opracowania zachowań zmotywowanych i kiedy prowadzi to do wrażliwych zachowań kompulsywnych? Dane anatomiczne pokazują, że KOR są doskonale ustawione do kontrolowania aktywności synaptycznej neuronów dopaminy śródmózgowia. Dane funkcjonalne wskazują, że KOR kontrolują funkcjonowanie DAT i D2R, a także szybkość wyzwalania neuronów dopaminowych. Wstępne dowody wskazujące, że ostra aktywacja KOR zmniejsza uwalnianie dopaminy wywołane przez leki uzależniające, zostały uzupełnione danymi wskazującymi, że powtarzana aktywacja KOR ułatwia uwalnianie dopaminy i kompulsywne poszukiwanie narkotyków. Sygnały dopaminowe równoważą bezpośrednie i pośrednie ścieżki wyjściowe z obszarów prążkowia (Rysunek 1A). Chroniczna stymulacja za pomocą psychostymulatorów, która zwiększa uwalnianie dopaminy aktywując zarówno D1R, jak i D2R (Rysunek 1B) lub chinpirol, które aktywują tylko D2R (Rysunek 1C) powoduje uczulenie ruchowe i zachowania kompulsywne przez osłabioną ścieżkę pośrednią D2R, tym samym przełączając równowagę na ścieżkę bezpośrednią D1R. Transmisja KOR jest zwiększona podczas długotrwałego przyjmowania psychostymulantów przez wzrost dynorfiny w prążkowatych neuronach D1 (Rysunek 1B). Zwiększona transmisja KOR jest emulowana w modelu farmakologicznym OCD poprzez podanie U69593. Ta jednoczesna aktywacja KOR dodatkowo osłabia pośrednią ścieżkę D2 (Rysunek 1C). Należy przeprowadzić przyszłe badania, aby w pełni wyjaśnić konsekwencje aktywacji KOR dla aktywności DAT, zrozumieć rolę endogennego układu KOR w kompulsywności indukowanej chinpirolem i określić wkład systemu KOR w różnice płci obserwowane w zachowaniach kompulsywnych.
Autorskie Wkłady
AE, MA i JF przyczyniły się do powstania manuskryptu. AE i JF napisali pierwszy szkic manuskryptu przy udziale MA. MA i JC przyczynili się do krytycznej recenzji i edycji manuskryptu. Wszyscy autorzy zatwierdzili go do publikacji.
Finansowanie
Praca autorów cytowanych w tym przeglądzie została poparta numerami grantów FONDECYT: 1110352 i 1150200 dla MA; 1141088 na rzecz JF; Dotacja DIPOG 391340281 dla JF; FONDECYT Stypendysta podoktorancki 3170497 do JC i 3190843 do AE.
Konflikt interesów
Autorzy oświadczają, że badanie zostało przeprowadzone przy braku jakichkolwiek powiązań handlowych lub finansowych, które mogłyby być interpretowane jako potencjalny konflikt interesów.
Redaktor ds. Obsługi organizuje obecnie temat badań z jednym z autorów JF i potwierdza brak jakiejkolwiek innej współpracy.
Referencje
Abraham, AD, Fontaine, HM, Song, AJ, Andrews, MM, Baird, MA, Kieffer, BL, i in. (2017). Aktywacja receptora opioidowego Kappa w neuronach dopaminy zakłóca hamowanie behawioralne. Neuropsychopharmacology 43 (2), 362–372. doi: 10.1038 / npp.2017.133
Abraham, AD, Schattauer, SS, Reichard, KL, Cohen, JH, Fontaine, HM, Song, AJ, i in. (2018). Regulacja estrogenów GRK2 inaktywuje sygnalizację receptora opioidowego kappa pośrednicząc w analgezji, ale nie awersji. J. Neurosci. 38 (37), 8031–8043. doi: 10.1523 / JNEUROSCI.0653-18.2018
Asaoka, N., Nishitani, N., Kinoshita, H., Nagai, Y., Hatakama, H., Nagayasu, K., i in. (2019). Antagonista receptora adenozynowego A2A poprawia wiele objawów powtarzanej psychozy wywołanej chinpirolem. eNeuro 6 (1), 1–16. ENEURO.0366-18.2019. doi: 10.1523 / ENEURO.0366-18.2019
Atigari, DV, Uprety, R., Pasternak, GW, Majumdar, S., Kivell, BM (2019). MP1104, mieszany agonista receptora opioidowego kappa-delta ma właściwości przeciw kokainie ze zmniejszonymi skutkami ubocznymi u szczurów. Neuropharmacology 150, 217 – 228. doi: 10.1016 / j.neuropharm.2019.02.010
Azocar, VH, Sepúlveda, G., Ruiz, C., Aguilera, C., Andrés, ME, Fuentealba, JA (2019). Blokowanie receptora opioidowego kappa odwraca zmiany dynamiki dopaminy prążkowia grzbietowo-boczna podczas uczulenia na amfetaminę. J. Neurochem. 148, 348–358. doi: 10.1111 / jnc.14612
Béguin, C., Potuzak, J., Xu, W., Liu-Chen, LY, Streicher, JM, Groer, CE, i in. (2012). Różnicowe właściwości sygnalizacyjne na receptorze opioidowym kappa 12-epi-salwinoryny A i jej analogów. Bioorg. Med. Chem. Łotysz. 15; 22 (2), 1023–1026. doi: 10.1016 / j.bmcl.2011.11.128
Beardsley, PM, Howard, JL, Shelton, KL, Carroll, FI (2005). Różnicowy wpływ nowego antagonisty receptora opioidowego kappa, JDTic, na przywrócenie poszukiwania kokainy wywołanego przez stresory w wyniku wstrząsów w stosunku do liczb pierwszych kokainy i jego działanie podobne do antydepresyjnego u szczurów. Psychofarmakol. (Berl) 183, 118–126. doi: 10.1007/s00213-005-0167-4
Becker, JB, Chartoff, E. (2019). Różnice płciowe w mechanizmach neuronalnych pośredniczących w nagradzaniu i uzależnieniu. Neuropsychopharmacology 44, 166–183. doi: 10.1038/s41386-018-0125-6
Becker, JB, Ramirez, VD (1981). Różnice płciowe w amfetaminie stymulowały uwalnianie katecholamin z tkanki prążkowanej szczura in vitro. Brain Res. 204, 361–372. doi: 10.1016/0006-8993(81)90595-3
Becker, JB (1999). Różnice płci w funkcji dopaminergicznej w prążkowiu i jądrze półleżącym. Pharmacol. Biochem. Behav. 64, 803–812. doi: 10.1016/S0091-3057(99)00168-9
Berridge, KC, Robinson, TE (2016). Lubienie, pragnienie i teoria uzależnienia uczulająco-motywująca. Rano. Psychol. 71, 670–679. doi: 10.1037 / amp0000059
Berridge, KC, Venier, IL, Robinson, TE (1989). Analiza reaktywności smaku afazji indukowanej 6-hydroksydopaminą: implikacje dla hipotez pobudzenia i anhedonii dotyczących funkcji dopaminy. Behav. Neurosci. 103, 36-45. doi: 10.1037 / 0735-7044.103.1.36
Broadbear, JH, Negus, SS, Butelman, ER, de Costa, BR, Woods, JH (1994). Różnicowy wpływ ogólnoustrojowej norbinaltorfiminy (nor-BNI) na agonistów kappa-opioidowych w teście skręcania myszy. Psychopharmacology 115, 311 – 319. doi: 10.1007 / BF02245071
Brown, RW, Perna, MK, Noel, DM, Whittemore, JD, Lehmann, J., Smith, ML (2011). Uczulenie ruchowe na amfetaminę i warunkowane preferencje miejsca u dorastających samców i samic szczurów leczonych noworodkiem chinpirolem. Behav. Pharmacol. 22, 374–378. doi: 10.1097/FBP.0b013e328348737b
Bruchas, MR, Chavkin, C. (2010). Kaskady kinaz i sygnalizacja kierowana ligandem na receptorze opioidowym kappa. Psychofarmakol. (Berl) 210, 137–147. doi: 10.1007/s00213-010-1806-y
Callaghan, CK, Rouine, J., O'Mara, SM (2018). Potencjalne role receptorów opioidowych w motywacji i dużym zaburzeniu depresyjnym. Wałówka. Brain Res. 239, 89–119. doi: 10.1016 / bs.pbr.2018.07.009
Carr, DB, Sesack, SR (2000). Neurony zawierające GABA w obszarze brzusznej części brzusznej szczura skierowane są do kory przedczołowej. Synapse 38 (2), 114–123. doi: 10.1002/1098-2396(200011)38:2<114::AID-SYN2>3.0.CO;2-R
Casanova, JP, Velis, GP, Fuentealba, JA (2013). Uczuleniu ruchowemu na amfetaminę towarzyszy zwiększone stymulowane K + uwalnianie dopaminy w środkowej korze przedczołowej szczura. Behav. Brain Res. 237, 313 – 317. doi: 10.1016 / j.bbr.2012.09.052
Chartoff, EH, Mavrikaki, M. (2015). Różnice płciowe w funkcji receptora opioidowego kappa i ich potencjalny wpływ na uzależnienie. Z przodu. Neurosci. 9, 466. doi: 10.3389 / fnins.2015.00466
Chartoff, EH, Ebner, SR, Sparrow, A., Potter, D., Baker, PM, Ragozzino, ME, i in. (2016). Względny czas między aktywacją receptora opioidowego kappa a kokainą determinuje wpływ na nagrodę i uwalnianie dopaminy. Neuropsychopharmacology 41, 989 – 1002. doi: 10.1038 / npp.2015.226
Chavkin, C., Koob, GF (2016). Dynorfina, dysforia i uzależnienie: stres uzależnienia. Neuropsychopharmacology 41, 373 – 374. doi: 10.1038 / npp.2015.258
Chefer, VI, Czyzyk, T., Bolan, EA, Moron, J., Pintar, JE, Shippenberg, T. S. (2005). Endogenne systemy receptorów opioidowych kappa regulują dynamikę dopaminy w mezoakumbie i wrażliwość na kokainę. J. Neurosci. 25, 5029 – 5037. doi: 10.1523 / JNEUROSCI.0854-05.2005
Chefer, VI, Zapata, A., Shippenberg, TS, Bungay, PM (2006). Ilościowa mikrodializa bez przepływu netto umożliwia wykrycie wzrostu i spadku wychwytu dopaminy w miąższu jądra mętnego. J. Neurosci. Metody 155, 187 – 193. doi: 10.1016 / j.jneumeth.2005.12.018
Cole, RL, Konradi, C., Douglass, J., Hyman, SE (1995). Adaptacja neuronalna do amfetaminy i dopaminy: molekularne mechanizmy regulacji genu prodynorfiny w prążkowiu szczura. Neuron 14, 813–823. doi: 10.1016/0896-6273(95)90225-2
Conway, SM, Puttick, D., Russell, S., Potter, D., Roitman, MF, Chartoff, E. H. (2019). Kobiety są mniej wrażliwe niż mężczyźni na motywacyjne i hamujące działanie dopaminy działanie aktywacji receptora opioidowego kappa. Neuropharmacology 146, 231 – 241. doi: 10.1016 / j.neuropharm.2018.12.002
Cope, ZA, Huggins, KN, Sheppard, AB, Noel, DM, Roane, DS, Brown, RW (2010). Leczenie chinpirolem noworodków zwiększa aktywację ruchową i uwalnianie dopaminy w rdzeniu jądra półleżącego w odpowiedzi na leczenie amfetaminą w wieku dorosłym. Synapse 64, 289 – 300. doi: 10.1002 / syn.20729
Cox, BM, Young, AB, See, RE, Reichel, CM (2013). Różnice płciowe w poszukiwaniu metamfetaminy u szczurów: wpływ oksytocyny. Psychoneuroendocrinology 38, 2343 – 2353. doi: 10.1016 / j.psyneuen.2013.05.005
Culver, KE, Szechtman, H., Levant, B. (2008). Zmienione wiązanie receptora dopaminopodobnego D2 u szczurów z uczuleniem behawioralnym na chinpirol: skutki wstępnego leczenia Ro 41-1049. Eur. J. Pharmacol. 592, 67-72. doi: 10.1016 / j.ejphar.2008.06.101
Dalman, FC, O'Malley, KL (1999). Tolerancja i uzależnienie od kappa-opioidów w kulturach neuronów dopaminergicznych śródmózgowia. J. Neurosci. 19, 5750–5757. doi: 10.1523/JNEUROSCI.19-14-05750.1999
De Vries, TJ, Schoffelmeer, AN, Binnekade, R., Raasø, H., Vanderschuren, LJ (2002). Nawrót do zachowań związanych z poszukiwaniem kokainy i heroiny za pośrednictwem receptorów dopaminy D2 jest zależny od czasu i związany z uczuleniem behawioralnym. Neuropsychopharmacology 26, 18–26. doi: 10.1016/S0893-133X(01)00293-7
Devine, DP, Leone, P., Pocock, D., Wise, RA (1993). Różnicowe zaangażowanie brzusznych czwartorzędowych receptorów opioidowych mu, delta i kappa w modulację podstawowego mezolimbicznego uwalniania dopaminy: badania mikrodializy in vivo. J. Pharmacol. Exp. Ther. 266, 1236-1246.
Di Chiara, G., Imperato, A. (1988). Przeciwne działanie agonistów opioidów mu i kappa na uwalnianie dopaminy w jądrze półleżącym i w grzbietowym ogonie swobodnie poruszających się szczurów. J. Pharmacol. Exp. Ther. 244, 1067 – 1080. doi: 10.1073 / pnas.85.14.5274
Dvorkin, A., Perreault, ML, Szechtman, H. (2006). Rozwój i czasowa organizacja kompulsywnego sprawdzania indukowanego przez wielokrotne wstrzykiwanie agonisty dopaminy chinpirolu w zwierzęcym modelu zaburzenia obsesyjno-kompulsyjnego. Behav. Brain Res. 169, 303 – 311. doi: 10.1016 / j.bbr.2006.01.024
Ebner, SR, Roitman, MF, Potter, DN, Rachlin, AB, Chartoff, EH (2010). Depresyjne działanie agonisty receptora opioidowego kappa, salvinorin A, wiąże się ze zmniejszonym fazowym uwalnianiem dopaminy w jądrze półleżącym. Psychofarmakol. (Berl) 210, 241–252. doi: 10.1007/s00213-010-1836-5
Edwards, NJ, Tejeda, HA, Pignatelli, M., Zhang, S., McDevitt, RA, Wu, J., i in. (2017). Specyficzność obwodu w architekturze hamującej VTA reguluje zachowanie wywołane kokainą. Nat. Neurosci. 20 (3), 438–448. doi: 10.1038 / nn.4482
Ehrich, JM, Phillips, PEM, Chavkin, C. (2014). Aktywacja receptora opioidowego Kappa nasila indukowany kokainą wzrost wywołanego uwalniania dopaminy zarejestrowanego in vivo w miąższu jądra móżdżkowego. Neuropsychopharmacology 39, 3036 – 3048. doi: 10.1038 / npp.2014.157
Ehrich, JM, Messinger, DI, Knakal, CR, Kuhar, JR, Schattauer, SS, Bruchas, MR, i in. (2015). Awersja indukowana przez receptory opioidowe Kappa wymaga aktywacji p38 MAPK w neuronach dopaminy VTA. J. Neurosci. 35, 12917 – 12931. doi: 10.1523 / JNEUROSCI.2444-15.2015
Eilam, D., Szechtman, H. (1989). Dwufazowy efekt agonisty D-2 chinpirolu na ruchliwość i ruchy. Eur. J. Pharmacol. 161, 151–157. doi: 10.1016/0014-2999(89)90837-6
Eilam, D., Szechtman, H. (2005). Zachowanie wywołane psychostymulantami jako zwierzęcy model zaburzeń obsesyjno-kompulsyjnych: etologiczne podejście do formy kompulsywnych rytuałów. CNS Spectr. 10, 191–202. doi: 10.1017 / S109285290001004X
Escobar, AP, Cornejo, FA, Andrés, ME, Fuentealba, JA (2012). Powtarzane leczenie agonistą receptora opioidowego kappa U69593 odwraca zwiększone indukowane K + uwalnianie dopaminy w jądrze półleżącym, ale nie wyraża wrażliwości ruchowej u szczurów uczulonych na amfetaminę. Neurochem. Int. 60 (4), 344–349. doi: 10.1016 / j.neuint.2012.01.014
Escobar, AP, Cornejo, FA, Olivares-Costa, M., González, M., Fuentealba, JA, Gysling, K., i in. (2015). Zmniejszona neurotransmisja dopaminy i glutaminianu w jądrze półleżącym szczurów uczulonych na chinpirol wskazuje na hamującą funkcję autoreceptora D2. J. Neurochem. 134, 1081–1090. doi: 10.1111 / jnc.13209
Escobar, AP, González, MP, Meza, RC, Noches, V., Henny, P., Gysling, K., i in. (2017). Mechanizmy wzmocnienia receptora opioidowego kappa funkcji receptora dopaminergicznego D2 w indukowanym chinpirolem uczuleniu ruchowym u szczurów. Int. J. Neuropsychopharmacol. 20, 660–669. doi: 10.1093 / ijnp / pyx042
Everitt, BJ, Robbins, TW (2005). Neurologiczne systemy wzmacniania uzależnienia od narkotyków: od działań, nawyków, po przymus. Nat. Neurosci. 8, 1481 – 1489. doi: 10.1038 / nn1579
Fattore, L., Melis, M. (2016). Różnice płciowe w zachowaniach impulsywnych i kompulsywnych: koncentracja na uzależnieniu od narkotyków. Nałogowiec. Biol. 21 (5), 1043–1051. doi: 10.1111 / adb.12381
Ferrario, CR, Gorny, G., Crombag, HS, Li, Y., Kolb, B., Robinson, TE (2005). Plastyczność neuronowa i behawioralna związana z przejściem od kontrolowanego do eskalowanego używania kokainy. Biol. Psy 58 (9) 751–9.
Figee, M., Pattij, T., Willuhn, I., Luigjes, J., van den Brink, W., Goudriaan, A., i in. (2016). Kompulsywność w zaburzeniach obsesyjno-kompulsyjnych i nałogach. Eur. Neuropsychopharmacol. 26, 856 – 868. doi: 10.1016 / j.euroneuro.2015.12.003
Ford, CP, Mark, GP, Williams, JT (2006). Właściwości i hamowanie opioidowe mezolimbicznych neuronów dopaminowych różnią się w zależności od docelowej lokalizacji. J. Neurosci. 26, 2788 – 2797. doi: 10.1523 / JNEUROSCI.4331-05.2006
Ford, CP, Beckstead, MJ, Williams, JT (2007). Hamowanie opioidowe przez kappa prądów postsynaptycznych hamujących somatodendrytyczne dopaminy. J. Neurophysiol. 97, 883 – 891. doi: 10.1152 / jn.00963.2006
Fuentealba, JA, Gysling, K., Magendzo, K., Andrés, ME (2006). Wielokrotne podawanie selektywnego agonisty receptora kappa-opioidowego U-69593 zwiększa stymulowane poziomy zewnątrzkomórkowej dopaminy w jądrze szczura półleżącego. J. Neurosci. Res. 84, 450 – 459. doi: 10.1002 / jnr.20890
Fuentealba, JA, Gysling, K., Andrés, ME (2007). Zwiększona odpowiedź ruchowa na amfetaminę indukowana przez wielokrotne podawanie selektywnego agonisty receptora kappa-opioidowego U-69593. Synapse 61, 771 – 777. doi: 10.1002 / syn.20424
Gehrke, BJ, Chefer, VI, Shippenberg, TS (2008). Wpływ ostrego i powtarzanego podawania salwinoryny A na funkcję dopaminy w prążkowiu grzbietowym szczura. Psychofarmakol. (Berl) 197, 509–517. doi: 10.1007/s00213-007-1067-6
Giuliano, C., Belin, D., Everitt, BJ (2019). Kompulsywne poszukiwanie alkoholu wynika z braku rozłączenia grzbietowo-bocznej kontroli prążkowia nad zachowaniem. J. Neurosci. 39 (9), 1744–1754. doi: 10.1523 / JNEUROSCI.2615-18.2018
Goto, Y., Grace, AA (2008). Limbiczne i korowe przetwarzanie informacji w jądrze półleżącym. Trendy Neurosci. 31, 552 – 558. doi: 10.1016 / j.tins.2008.08.002
Grace, AA, Bunney, BS (1980). Nigral neurony dopaminowe: rejestracja i identyfikacja wewnątrzkomórkowa za pomocą iniekcji L-dopa i histofluorescencji. nauka 210, 654 – 656. doi: 10.1126 / science.7433992
Gray, AM, Rawls, SM, Shippenberg, TS, McGinty, JF (1999). Agonista opioidów κ, U-69593, zmniejsza ostre zachowania wywoływane przez amfetaminę oraz zależne od wapnia poziomy dopaminy i glutaminianu w prążkowiu brzusznym. J. Neurochem. 73, 1066-1074. doi: 10.1046 / j.1471-4159.1999.0731066.x
Graziane, NM, Polter, AM, Briand, LA, Pierce, RC, Kauer, JA (2013). Receptory opioidowe Kappa regulują wywołane stresem poszukiwanie kokainy i plastyczność synaptyczną. Neuron 77, 942 – 954. doi: 10.1016 / j.neuron.2012.12.034
Heidbreder, CA, Shippenberg, TS (1994). U-69593 zapobiega uczuleniu na kokainę poprzez normalizację podstawowej półleżącej dopaminy. Neuroreport 5, 1797–1800. doi: 10.1097/00001756-199409080-00028
Hernandez-Avila, CA 1., Rounsaville, BJ, Kranzler, HR (2004). Kobiety uzależnione od opioidów, konopi indyjskich i alkoholu wykazują szybszy postęp w leczeniu uzależnień. Drug Alcohol Depend. 74 (3), 265–272. doi: 10.1016 / j.drugalcdep.2004.02.001
Hoffman, AF, Spivak, CE, Lupica, CR (2016). Zwiększone uwalnianie dopaminy przez inhibitory transportu dopaminy opisane przez model ograniczonej dyfuzji i cykliczną woltametrię z szybkim skanowaniem. ACS Chem. Neurosci. 7, 700–709. doi: 10.1021 / acschemneuro.5b00277
Holden, C. (2001). Uzależnienia „behawioralne”: czy istnieją? nauka 294, 980 – 982. doi: 10.1126 / science.294.5544.980
Imperato, A., Di Chiara, G. (1988). Wpływ lokalnie stosowanych agonistów i antagonistów receptorów D-1 i D-2 badano za pomocą dializy mózgu. Eur. J. Pharmacol. 156, 385–393. doi: 10.1016/0014-2999(88)90284-1
Jackson, KJ, McLaughlin, JP, Carroll, FI, Damaj, MI (2013). Wpływ antagonisty receptora opioidowego kappa, norbinaltorfiminy, na stres i wywołane lekami przywrócenie uzależnionych od nikotyny preferencji miejsca u myszy. Psychofarmakol. (Berl) 226, 763–768. doi: 10.1007/s00213-012-2716-y
Kivell, B., Uzelac, Z., Sundaramurthy, S., Rajamanickam, J., Ewald, A., Chefer, V., i in. (2014). Salvinorin A reguluje funkcję transportera dopaminy poprzez receptor opioidowy kappa i mechanizm zależny od ERK1 / 2. Neuropharmacology 86, 228 – 240. doi: 10.1016 / j.neuropharm.2014.07.016
Koeltzow, TE, Austin, JD, Vezina, P. (2003). Uczulenie behawioralne na chinpirol nie jest związane ze zwiększonym przelaniem jądra półleżącego dopaminy. Neuropharmacology 44, 102–110. doi: 10.1016/S0028-3908(02)00328-3
Koob, GF, Volkow, ND (2016). Neurobiologia uzależnienia: analiza układu nerwowego. Lancet Psychiatry 3, 760–773. doi: 10.1016/S2215-0366(16)00104-8
Koob, GF (2013). Uzależnienie jest deficytem nagród i zaburzeniami nadmiernego stresu. Z przodu. Psychiatria 4, 72. doi: 10.3389 / fpsyt.2013.00072
Labad, J. 1., Menchon, JM, Alonso, P., Segalas, C., Jimenez, S., Jaurrieta, N., i in. (2008). Różnice płci w wymiarach objawów obsesyjno-kompulsyjnych. Wciśnij lęk. 25 (10), 832–838. doi: 10.1002 / da.20332
Maisonneuve, IM, Archer, S., Glick, SD (1994). U50,488, agonista receptora opioidowego kappa, osłabia indukowany kokainą wzrost pozakomórkowej dopaminy w jądrze półleżącym szczurów. Neurosci. Łotysz. 181, 57–60. doi: 10.1016/0304-3940(94)90559-2
Mansour, A., Fox, CA, Akil, H., Watson, SJ (1995). Ekspresja mRNA receptora opioidowego w ośrodkowym układzie nerwowym szczura: implikacje anatomiczne i funkcjonalne. Trendy Neurosci. 18 (1), 22–29. doi: 10.1016/0166-2236(95)93946-U
Mansour, A., Burke, S., Pavlic, RJ, Akil, H., Watson, SJ (1996). Immunohistochemiczna lokalizacja sklonowanego receptora kappa 1 w ośrodkowym układzie nerwowym szczura i przysadce mózgowej. Neuroscience 71, 671–690. doi: 10.1016/0306-4522(95)00464-5
Margolis, EB, Karkhanis, AN (2019). Dopaminergiczny udział komórek i obwodów w awersji za pośrednictwem receptora opioidowego kappa. Neurochem. Int. 129, 104504. doi: 10.1016 / j.neuint.2019.104504
Margolis, EB, Hjelmstad, GO, Bonci, A., Fields, HL (2003). Agoniści opioidów kappa bezpośrednio hamują neurony dopaminergiczne śródmózgowia. J. Neurosci. 23, 9981–9986. doi: 10.1523/JNEUROSCI.23-31-09981.2003
Margolis, EB, Hjelmstad, GO, Bonci, A., Fields, HL (2005). Zarówno agoniści opioidów Kappa, jak i Mu hamują wejście glutaminianergiczne do neuronów brzusznej okolicy czołowej. J. Neurophysiol. 93, 3086 – 3093. doi: 10.1152 / jn.00855.2004
Margolis, EB, Lock, H., Chefer, VI, Shippenberg, TS, Hjelmstad, GO, Fields, HL (2006). Opioidy Kappa selektywnie kontrolują neurony dopaminergiczne wystające na korę przedczołową. Proc. Natl. Acad. Sci. USA 103, 2938 – 2942. doi: 10.1073 / pnas.0511159103
Margolis, EB, Mitchell, JM, Ishikawa, J., Hjelmstad, GO, Fields, HL (2008). Neurony dopaminy w śródmózgowiu: cel projekcji określa czas trwania potencjału czynnościowego i hamowanie receptora dopaminy D (2). J. Neurosci. 28, 8908 – 8913. doi: 10.1523 / JNEUROSCI.1526-08.2008
Mathews, IZ, McCormick, CM (2007). Samice i samce szczurów w późnym okresie dojrzewania różnią się od dorosłych aktywnością ruchową indukowaną amfetaminą, ale nie preferują amfetaminy w warunkowym miejscu. Behav. Pharmacol. 18, 641–650. doi: 10.1097/FBP.0b013e3282effbf5
Mathis, MA 1., Pd, A., Funaro, G., RC, T., Moraes, I., AR, T., i in. (2011). Różnice płci w zaburzeniu obsesyjno-kompulsyjnym: przegląd literatury. Braz. J. Psychiatria 33 (4), 390–399. doi: 10.1590 / S1516-44462011000400014
McLaughlin, JP, Marton-Popovici, M., Chavkin, C. (2003). Antagonizm receptora opioidowego Kappa i zaburzenie genu prodynorfiny blokują wywołane stresem odpowiedzi behawioralne. J. Neurosci. 23 (13), 5674–5683. doi: 10.1523 / JNEUROSCI.23-13-05674.2003
McLaughlin, JP, Land, BB, Li, S., Pintar, JE, Chavkin, C. (2006a). Wcześniejsza aktywacja receptorów opioidowych kappa przez U50,488 naśladuje powtarzany stres wymuszony podczas pływania w celu wzmocnienia warunkowania preferencji miejsca kokainy. Neuropsychopharmacology 31, 787-794. doi: 10.1038 / sj.npp.1300860
McLaughlin, JP, Li, S., Valdez, J., Chavkin, TA, Chavkin, C. (2006b). W reakcjach behawioralnych wywołanych stresem społecznym pośredniczy endogenny układ opioidowy kappa. Neuropsychopharmacology 31 (6), 1241–1248. doi: 10.1038 / sj.npp.1300872
Milesi-Hallé, A., McMillan, DE, Laurenzana, EM, Byrnes-Blake, KA, Owens, SM (2007). Różnice płciowe w odpowiedziach behawioralnych wywołanych (+) - amfetaminą i (+) - metamfetaminą u samców i samic szczurów Sprague-Dawley. Pharmacol. Biochem. Behav. 86, 140 – 149. doi: 10.1016 / j.pbb.2006.12.018
Negus, SS, Mello, NK, Portoghese, PS, Lin, CE (1997). Wpływ opioidów kappa na samopodawanie kokainy przez małpy rezus. J. Pharmacol. Exp. Ther. 282, 44-55.
Nestler, EJ (2001). Molekularne podstawy długotrwałej plastyczności leżącej u podstaw uzależnienia. Nat. Wielebny Neurosci. 2, 119-128. doi: 10.1038 / 35053570
Perreault, ML, Graham, D., Bisnaire, L., Simms, J., Hayton, S., Szechtman, H. (2006). Agonista opioidów kappa U69593 nasila uczulenie ruchowe na chinpirol agonisty D2 / D3: mechanizmy pre- i postsynaptyczne. Neuropsychopharmacology 31, 1967-1981. doi: 10.1038 / sj.npp.1300938
Perreault, ML, Seeman, P., Szechtman, H. (2007). Stymulacja receptora kappa-opioidowego przyspiesza patogenezę kompulsywnego sprawdzania w modelu uwrażliwienia chinpirolowego na zaburzenia obsesyjno-kompulsyjne (OCD). Behav. Neurosci. 121, 976-991. doi: 10.1037 / 0735-7044.121.5.976
Pierce, RC, Kalivas, PW (1997). Model obwodów ekspresji uczulenia behawioralnego na psychostymulanty podobne do amfetaminy. Brain Res. Brain Res. Obrót silnika. 25, 192–216. doi: 10.1016/S0165-0173(97)00021-0
Polter, AM, Bishop, RA, Briand, LA, Graziane, NM, Pierce, RC, Kauer, JA (2014). Blok antystresowy receptorów opioidowych Kappa ratuje długotrwałe wzmocnienie synaps hamujących i zapobiega przywróceniu kokainy. Biol. Psychiatria 76, 785-793. doi: 10.1016 / j.biopsych.2014.04.019
Puig-Ramos, A., Santiago, GS, Segarra, AC (2008). U-69593, agonista receptora opioidowego kappa, zmniejsza indukowane kokainą uczulenie behawioralne u samic szczurów. Behav. Neurosci. 122, 151-160. doi: 10.1037 / 0735-7044.122.1.151
Robbins, TW, Gillan, CM, Smith, DG, de Wit, S., Ersche, KD (2012). Endofenotypy neurokognitywne impulsywności i kompulsywności: w kierunku psychiatrii wymiarowej. Trendy Cognit. Sci. 16, 81 – 91. doi: 10.1016 / j.tics.2011.11.009
Roberts, DCS, Bennett, SAL, Vickers, GJ (1989). Cykl rujowy wpływa na samodzielne podawanie kokainy według progresywnego harmonogramu u szczurów. Psychofarmakol. (Berl) 98, 408 – 411. doi: 10.1007 / BF00451696
Robinson, TE, Berridge, KC (1993). Neuralne podstawy głodu narkotyków: teoria uzależnienia od uczulenia motywacyjnego. Brain Res. Brain Res. Obrót silnika. 18, 247–291. doi: 10.1016/0165-0173(93)90013-P
Robinson, TE, Berridge, KC (2001). Bodźce uczulające i uzależnienie. Nałóg 96, 103-114. doi: 10.1046 / j.1360-0443.2001.9611038.x
Rose, JH, Karkhanis, AN, Steiniger-Brach, B., Jones, SR (2016). Wyraźny wpływ nalmefenu na szybkość wychwytu dopaminy i aktywność receptora opioidowego kappa w jądrze półleżącym po przewlekłej przerywanej ekspozycji na etanol. Int. J. Mol. Sci. 17, 1216. doi: 10.3390 / ijms17081216
Schlosburg, JE, Whitfield, TW, Jr., Park, PE, Crawford, EF, George, O., Vendruscolo, LF, i in. (2013). Długotrwały antagonizm receptorów opioidowych κ zapobiega eskalacji i zwiększonej motywacji do przyjmowania heroiny. J. Neurosci. 33 (49), 19384–19392. doi: 10.1523 / JNEUROSCI.1979-13.2013
Sesack, SR, Pickel, VM (1992). Podwójna lokalizacja ultrastrukturalna immunoreaktywności enkefaliny i hydroksylazy tyrozynowej w brzusznej okolicy brzusznej szczura: wiele substratów dla interakcji opiat-dopamina. J. Neurosci. 12, 1335–a1350. doi: 10.1523/JNEUROSCI.12-04-01335.1992
Sesack, SR, Aoki, C., Pickel, VM (1994). Ultrastrukturalna lokalizacja immunoreaktywności podobnej do receptora D2 w neuronach dopaminy śródmózgowia i ich celach prążkowiowych. J. Neurosci. 14, 88–106. doi: 10.1523/JNEUROSCI.14-01-00088.1994
Sesia, T., Bizup, B., Grace, AA (2013). Ocena modeli zwierzęcych zaburzenia obsesyjno-kompulsyjnego: korelacja z fazową aktywnością neuronu dopaminowego. Int. J. Neuropsychopharmacol. 16, 1295–1307. doi: 10.1017 / S146114571200154X
Shippenberg, TS, Zapata, A., Chefer, VI (2007). Dynorfina i patofizjologia uzależnienia od narkotyków. Pharmacol Ther. 116, 306–321. doi: 10.1016 / j.pharmthera.2007.06.011
Smith, JA, Loughlin, SE, Leslie, FM (1992). Hamowanie kappa-opioidowe uwalniania [3H] dopaminy z szczurzych brzusznych zdysocjowanych hodowli komórek mięśniowych. Mol. Pharmacol. 42.
Smith, JW, Fetsko, LA, Xu, R., Wang, Y. (2002). Myszy z nokautem receptora dopaminy D2L wykazują deficyty w dodatnich i ujemnych właściwościach wzmacniających morfiny oraz w uczeniu się unikania. Neuroscience 113 (4), 755–765. doi: 10.1016/S0306-4522(02)00257-9
Smith, JS, Schindler, AG, Martinelli, E., Gustin, RM, Bruchas, MR, Chavkin, C. (2012). Wywołana stresem aktywacja układu receptorowego dynorfiny / κ-opioidu w ciele migdałowatym potęguje preferencję miejsca uwarunkowaną nikotyną. J. Neurosci. 32, 1488 – 1495. doi: 10.1523 / JNEUROSCI.2980-11.2012
Spanagel, R., Herz, A., Shippenberg, TS (1992). Przeciwstawne tonicznie aktywne endogenne układy opioidowe modulują mezolimbiczny szlak dopaminergiczny. Proc. Natl. Acad. Sci. 89, 2046 – 2050. doi: 10.1073 / pnas.89.6.2046
Speciale, SG, Manaye, KF, Sadeq, M., niemiecki, DC (1993). Receptory opioidowe w dopaminergicznych obszarach śródmózgowia szczura. II. Autoradiografia receptora Kappa i delta. J. Przek. neuron. Gen. Sek. 91, 53 – 66. doi: 10.1007 / BF01244918
Sperling, RE, Gomes, SM, Sypek, EI, Carey, AN, McLaughlin, JP (2010). Endogenne pośrednictwo kappa-opioidowe wywołanego stresem wzmocnienia uwarunkowanego etanolem preferencji miejsca i samodzielnego podawania. Psychofarmakol. (Berl) 210 (2), 199–209. doi: 10.1007/s00213-010-1844-5
Steketee, JD, Kalivas, PW (2011). Drug Wanting: uczulenie behawioralne i nawrót do zachowań związanych z poszukiwaniem narkotyków sibley DR, red. Pharmacol. Obrót silnika. 63, 348–365. doi: 10.1124 / pr.109.001933
Stuchlik, A., Radostová, D., Hatalova, H., Vales, K., Nekovarova, T., Koprivova, J., i in. (2016). Ważność szczurzego modelu OCD uczulającego na chinpirol: powiązanie dowodów z badań na zwierzętach i badaniach klinicznych. Z przodu. Behav. Neurosci. 10, 209. doi: 10.3389 / fnbeh.2016.00209
Sun, T., Song, Z., Tian, Y., Tian, W., Zhu, C., Ji, G., i in. (2019). Bazolateralne wejście ciała migdałowatego do przyśrodkowej kory przedczołowej kontroluje zachowanie sprawdzające obsesyjno-kompulsywne. Proc. Natl. Acad. Sci. USA 116, 3799 – 3804. doi: 10.1073 / pnas.1814292116
Svingos, AL, Chavkin, C., Colago, EEO, Pickel, VM (2001). Główna koekspresja receptorów - - opioidowych i transportera dopaminy w jądrze obciąża profile aksonalne. Synapse 42, 185 – 192. doi: 10.1002 / syn.10005
Szechtman, H., Talangbayan, H., Eilam, D. (1993). Środowiskowe i behawioralne składniki uczulenia indukowane przez agonistę dopaminy, chinpirol. Behav. Pharmacol. 4, 405–410. doi: 10.1097/00008877-199308000-00014
Szechtman, H., Talangbayan, H., Canaran, G., Dai, H., Eilam, D. (1994). Dynamika sensytyzacji behawioralnej indukowanej przez chinpirol będący agonistą dopaminy oraz proponowany mechanizm centralnej kontroli energii. Psychofarmakol. (Berl) 115, 95 – 104. doi: 10.1007 / BF02244757
Szechtman, H., Sulis, W., Eilam, D. (1998). Chinopirol wywołuje u szczurów kompulsywne sprawdzanie: potencjalny zwierzęcy model zaburzeń obsesyjno-kompulsyjnych (OCD). Behav. Neurosci. 112, 1475-1485. doi: 10.1037 / 0735-7044.112.6.1475
Szechtman, H., Culver, K., Eilam, D. (1999). Rola układów dopaminowych w zaburzeniu obsesyjno-kompulsyjnym (OCD): implikacje nowego modelu zwierzęcego indukowanego psychostymulantem. pol. J. Pharmacol. 51, 55-61.
Szechtman, H., Eckert, MJ, Tse, WS, Boersma, JT, Bonura, Ca, JZ, M., i in. (2001). Zachowanie kompulsywne u szczurów uczulonych na chinpirol jako zwierzęcy model zaburzenia obsesyjno-kompulsyjnego (OCD): forma i kontrola. BMC Neurosci. 2, 4. doi: 10.1186/1471-2202-2-4
Szechtman, H., Ahmari, SE, Beninger, RJ, Eilam, D., Harvey, BH, Edemann-Callesen, H., i in. (2017). Zaburzenie obsesyjno-kompulsyjne: spostrzeżenia z modeli zwierzęcych. Neurosci. Biobehav. Obrót silnika. 76, 254-279. doi: 10.1016 / j.neubiorev.2016.04.019
Tejeda, HA, Bonci, A. (2019). Kontrola dynamiki dopaminy przez receptor dynorfin / kappa-opioidowy: Implikacje dla negatywnych stanów afektywnych i zaburzeń psychicznych. Brain Res. 1713, 91 – 101. doi: 10.1016 / j.brainres.2018.09.023
Tejeda, HA, Wu, J., Kornspun, AR, Pignatelli, M., Kashtelyan, V., Krashes, M. J., et al. (2017). Specyficzna dla szlaku i komórki modulacja receptora kappa-opioidowego równowagi pobudzenia-hamowania różnicuje aktywność neuronów d1 i d2 półleżących. Neuron 93 (1), 147–163. doi: 10.1016 / j.neuron.2016.12.005
Thompson, AC, Zapata, A., Justice, JB, Vaughan, RA, Sharpe, LG, Shippenberg, TS (2000). Aktywacja receptora kappa-opioidowego modyfikuje wychwyt dopaminy w jądrze półleżącym i przeciwdziała działaniu kokainy. J. Neurosci. 20, 9333–9340. doi: 10.1523/JNEUROSCI.20-24-09333.2000
Thompson, D., Martini, L., Whistler, JL (2010). Zmieniony stosunek receptorów dopaminy D1 i D2 w prążkowiu myszy jest związany z uczuleniem behawioralnym na kokainę. PloS One 5, e11038. doi: 10.1371 / journal.pone.0011038
Usiello, A., Baik, J.-H., Rougé-Pont, F., Picetti, R., Dierich, A., LeMeur, M., i in. (2000). Różne funkcje dwóch izoform receptorów dopaminy D2. Natura 408, 199-203. doi: 10.1038 / 35041572
Van Bockstaele, EJ, Pickel, VM (1995). Neurony zawierające GABA w brzusznym obszarze nakrywkowym skierowane są na jądro półleżące w mózgu szczura. Brain Res. 682 (1-2), 215–221. doi: 10.1016/0006-8993(95)00334-M
Van't Veer, A., Bechtholt, AJ, Onvani, S., Potter, D., Wang, Y., Liu-Chen, LY, et al. (2013). Ablacja receptorów kappa-opioidowych z neuronów dopaminowych mózgu ma działanie podobne do przeciwlękowych i zwiększa plastyczność wywołaną kokainą. Neuropsychopharmacology 38 (8), 1585–1597. doi: 10.1038 / npp.2013.58
Vanderschuren, LJ, Kalivas, PW (2000). Zmiany w transmisji dopaminergicznej i glutaminergicznej w indukcji i ekspresji uczulenia behawioralnego: krytyczny przegląd badań przedklinicznych. Psychofarmakol. (Berl) 151 (2-3), 99–120. doi: 10.1007 / s002130000493
Volkow, ND, Wise, RA, Baler, R. (2017). System motywacyjny dopaminy: implikacje uzależnienia od narkotyków i żywności. Nat. Wielebny Neurosci. 18, 741–752. doi: 10.1038 / nrn.2017.130
Walker, QD, Rooney, MB, Wightman, RM, Kuhn, CM (2000). Uwalnianie i wychwyt dopaminy jest większe u prążkowia samic niż samców szczura, co zmierzono za pomocą szybkiej woltamperometrii cyklicznej. Neuroscience 95, 1061–1070. doi: 10.1016/S0306-4522(99)00500-X
Wee, S., Koob, GF (2010). Rola systemu opioidowego dynorfin – κ we wzmacniającym działaniu narkotyków. Psychofarmakol. (Berl) 210, 121–135. doi: 10.1007/s00213-010-1825-8
Wee, S., Orio, L., Ghirmai, S., Cashman, JR, Koob, GF (2009). Hamowanie receptorów opioidowych kappa osłabiało zwiększone spożycie kokainy u szczurów z rozszerzonym dostępem do kokainy. Psychofarmakol. (Berl) 205, 565–575. doi: 10.1007/s00213-009-1563-y
Whitfield, TW, Jr., Schlosburg, JE, Wee, S., Gould, A., George, O., Grant, Y., i in. (2015). Receptory opioidowe w jądrze półleżącym pośredniczą w zwiększaniu spożycia metamfetaminy. J. Neurosci. 35 (10), 4296–4305. doi: 10.1523 / JNEUROSCI.1978-13.2015
Williams, MT, Mugno, B., Franklin, M., Faber, S. (2013). Wymiary objawów w zaburzeniu obsesyjno-kompulsyjnym: fenomenologia i wyniki leczenia z ekspozycją i profilaktyką rytualną. Psychopatologia 46, 365-376. doi: 10.1159 / 000348582
Wilson, CJ, Young, SJ, Groves, PM (1977). Właściwości statystyczne neuronalnych łańcuchów szczytowych w istocie czarnej: typy komórek i ich interakcje. Brain Res. 136, 243–260. doi: 10.1016/0006-8993(77)90801-0
Wise, RA (2009). Role nigrostriatalnej - nie tylko mezokortykolimbicznej - dopaminy w nagrodach i uzależnieniu. Trendy Neurosci. 32, 517 – 524. doi: 10.1016 / j.tins.2009.06.004
Xiao, L., Becker, JB (1994). Ilościowe oznaczanie mikrodializy pozakomórkowego prążkowia stężenia dopaminy u samców i samic szczurów: skutki cyklu rujowego i gonadektomii. Neurosci. Łotysz. 180, 155–158. doi: 10.1016/0304-3940(94)90510-X
Słowa kluczowe: receptor opioidowy kappa, dopamina, kompulsywność, amfetamina, chinpirol, uczulenie ruchowe
Cytowanie: Escobar AdP, Casanova JP, Andrés ME i Fuentealba JA (2020) Crosstalk Between Kappa Opioid and Dopamine Systems in Compulsive Behaviors. Z przodu. Pharmacol. 11: 57. doi: 10.3389 / fphar.2020.00057
Otrzymano: 16 października 2019 r .; Przyjęte: 22 stycznia 2020 r .;
Opublikowane: 18 February 2020.
Edytowany przez:
Gonzalo E. Yevenes, Uniwersytet Poczęcia, Chile
Zrecenzowany przez:
Luisa Gerardo Aguayo, Uniwersytet Poczęcia, Chile
Hugo Tejedy, National Institute on Drug Abuse (NIDA), Stany Zjednoczone
Cecylia Skorza, Instituto de Investigaciones Biológicas Clemente Estable (IIBCE), Urugwaj
Prawa autorskie © 2020 Escobar, Casanova, Andrés i Fuentealba. To jest artykuł o otwartym dostępie dystrybuowany na warunkach Licencja Creative Commons Uznanie autorstwa (CC BY). Dozwolone jest używanie, dystrybucja lub powielanie na innych forach, pod warunkiem że oryginalny autor (autorzy) i właściciel (właściciele) praw autorskich są uznani i że pierwotna publikacja w tym czasopiśmie jest cytowana, zgodnie z przyjętą praktyką akademicką. Dozwolone jest używanie, dystrybucja lub powielanie niezgodne z tymi warunkami.
* Korespondencja: José Antonio Fuentealba, [email chroniony]
†ORCID: José Antonio Fuentealba, orcid.org/0000-0003-0775-0675
