Z przodu. Behav. Neurosci., 23 March 2016 | http://dx.doi.org/10.3389/fnbeh.2016.00054

 Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2,
Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2, 

- 1Translational Research Institute i Instytut Zdrowia i Innowacji Biomedycznych, Queensland University of Technology, Brisbane, QLD, Australia
- 2Szkoła Nauk Biomedycznych, Uniwersytet Queensland, Brisbane, QLD, Australia
Współczesna dieta stała się bardzo słodzona, co skutkuje niespotykanym poziomem spożycia cukru, szczególnie wśród młodzieży. Podczas gdy wiadomo, że przewlekłe długotrwałe spożywanie cukru przyczynia się do rozwoju zaburzeń metabolicznych, w tym otyłości i cukrzycy typu II, niewiele wiadomo na temat bezpośrednich konsekwencji długotrwałego, podobnego do upijania się cukru w mózgu. bponieważ cukier może powodować uwalnianie dopaminy w jądrze półleżącym (NAc), podobnie jak leki nadużywające, badaliśmy zmiany morfologii neuronów w tym regionie mózgu po krótkim (4 tygodniach) i długotrwałym (12 tygodniach) jak konsumpcja sacharozy przy użyciu przerywanego paradygmatu wyboru dwóch butelek Zastosowaliśmy barwienie Golgiego-Coxa do nasycenia średnich neuronów kolczastych (MSN) z rdzenia i powłoki NAc krótko- i długoterminowych szczurów spożywających sacharozę i porównano je z dopasowanymi do wieku kontrolami wody. Pokazujemy, że długotrwałe spożywanie sacharozy typu „binge-like” znacząco zmniejszało całkowitą długość dendrytyczną MSN muszli NAc w porównaniu z dopasowanymi wiekowo szczurami kontrolnymi. Odkryliśmy również, że restrukturyzacja tych neuronów wynikała przede wszystkim ze zmniejszonej dystalnej złożoności dendrytycznej. Odwrotnie, obserwowaliśmy zwiększone gęstości kręgosłupa w odległych rzędach rozgałęzień MSN muszli NAc z długotrwałych szczurów spożywających sacharozę. Łącznie wyniki te podkreślają efekty neuronalne przedłużonego upijania się sacharozy na morfologię MSN powłoki NAc.
Wprowadzenie
W ciągu ostatnich 40 odnotowano wzrost spożycia napojów słodzonych cukrem i żywności zawierającej dodatek cukru (Nielsen i in., 2002; Popkin, 2010; Ng i wsp., 2012), z raportami szacującymi, że do 75% całej żywności i napojów zawiera duże ilości dodanych cukrów (Ford i Dietz, 2013; Bray i Popkin, 2014). W tym okresie nastąpił równoczesny wzrost częstości występowania otyłości i cukrzycy typu II, szczególnie wśród młodzieży (Arslanian, 2002; Reinehr, 2013; Dabelea i in., 2014; Fryar i in., 2014). Ostatnie badania wykazały, że dzieci z nadwagą i otyłością często spożywają duże ilości dodanego cukru, jednak kontrybucja diet o wysokiej zawartości cukru do zwiększonej częstości występowania dzieci z nadwagą i otyłością pozostaje kontrowersyjna (Hu, 2013; Bray i Popkin, 2014; Bucher Della Torre i in., 2015).
Chociaż coraz więcej dowodów wskazuje, że spożywanie diet o wysokiej zawartości cukru może częściowo przyczynić się do zwiększenia masy ciała wśród dzieci i młodzieży (Malik i in., 2010; Te Morenga i in., 2013; Bray i Popkin, 2014), mniej uwagi poświęcono niekorzystnym konsekwencjom niemetabolicznym wynikającym z nadmiernego spożycia cukru. Co ciekawe, niektóre wspólne wzorce behawioralne i psychologiczne często pojawiają się w podgrupie osób, które jedzą i utrzymują diety zawierające dużo cukru. Najbardziej godne uwagi są rozwój zaburzeń odżywiania się, w tym objadanie się, połączone z jednoczesnym wystąpieniem objawów psychologicznych, w tym brakiem motywacji i depresji (przegląd w Sheehan i Herman, 2015). Ponadto, ponieważ osoby jedzące objadanie się często wykazują utratę kontroli i niezdolność do samodzielnego ograniczenia spożycia cukru, jest prawdopodobne, że te zachowania powstają w wyniku adaptacji neurologicznych w regionach mózgu, które oceniają wartość hedoniczną wysoce smacznego jedzenia (Saper i in., 2002; Lutter i Nestler, 2009; Kenny, 2011). To uzasadnienie jest również poparte dowodami u ludzi wykazującymi, że cukier i słodycz mogą wywoływać łaknienie podobne do tych wywołanych przez uzależniające leki, takie jak alkohol i nikotyna (Volkow i wsp., 2012).
Chociaż uzależniające właściwości cukru są nadal spekulacyjne, obserwacje te w połączeniu z badaniami wykazującymi udział nadmiernego spożycia cukru w zmianach w obwodach nagrody i rozwoju uzależniających zachowań i stanów emocjonalnych w modelach zwierzęcych (Avena i wsp., 2008; Benton, 2010; Ventura i in., 2014), gwarantuje potrzebę dalszego dochodzenia. Wcześniejsze badania na gryzoniach wykazały, że okresowy dostęp do sacharozy zmienia aktywność kilku neuroprzekaźników w układzie mezolimbicznym, w tym dopaminę, opioidy i acetylocholinę (przegląd w Avena i wsp., 2008). Wykazano, że konsumpcja sacharozy typu „binge” ułatwia uwalnianie dopaminy w jądrze półleżącym (NAc), podobnie jak narkotyki (Avena i wsp., 2008). Ponadto wykazaliśmy, że długotrwałe spożywanie sacharozy za pomocą paradygmatu wyboru dwóch butelek 24 h z przerywanym dostępem (Simms i in., 2008) moduluje ekspresję nikotynowego receptora acetylocholinowego (nAChR) w NAc (Shariff i in., W druku). Co ciekawe, zaobserwowaliśmy również, że związki nAChR, o których wiadomo, że modulują aktywność dopaminy i acetylocholiny w NAc, mają różny wpływ na spożycie sacharozy po krótko- i długoterminowym spożyciu (Shariff i in., W druku).
Podczas gdy badania te wykazały podobieństwa w zmianach behawioralnych i neurochemicznych spowodowanych przerywanym dostępem do cukru i narkotyków, nie wiadomo, czy te efekty ułatwiają zmiany w morfologii neuronów w NAc. Tw przeciwieństwie do substancji uzależniających, w tym kokainy, amfetaminy i nikotyny, które powodują dobrze scharakteryzowane zmiany w morfologii średnich kolczastych neuronów (MSN) w NAc, w tym zwiększoną gęstość kręgosłupa i zmienioną złożoność dendrytyczną (Robinson i Kolb, 1999, 2004; Li i wsp., 2003; Crombag i in., 2005). Ponieważ wcześniej wykazaliśmy, że długotrwała ekspozycja (tydzień 12) na alkohol i sacharozę za pomocą paradygmatu przerywanego wyboru dwóch butelek powoduje zróżnicowaną reakcję na interwencje farmakoterapeutyczne w porównaniu z krótkotrwałym przyjmowaniem (4 tygodnie; Steensland i in., 2007; Shariff i in., W druku), oceniliśmy wpływ krótko- i długoterminowego zużycia sacharozy na morfologię MSN w NAc. Pozwoliliśmy dorastającym szczurom spożywać sacharozę w podobny sposób jak dla 4 (krótkookresowe) lub 12 (długoterminowe) tygodnie, a następnie analizowano morfologię MSN z NAc z krótko- i długoterminowych sacharozy spożywających szczury i porównano to z dopasowani do wieku kontrole, którym udzielono dostępu tylko do wody. Nasze wyniki pokazują, że MSN z powłoki NAc ulegają zmianie po długim, ale nie krótkotrwałym spożyciu sacharozy, o zmniejszonej długości dendrytycznej, ale zwiększonej dystalnej gęstości dendrytycznej kręgosłupa. Ponadto, odkryliśmy, że morfologia MSN z rdzenia NAc pozostała względnie nienaruszona po krótkotrwałym i długotrwałym spożyciu sacharozy. Wyniki te podkreślają bezpośrednią neurologiczną konsekwencję długotrwałego spożywania sacharozy w sposób upalny. Ponadto dane te wskazują na potrzebę dalszych badań mających na celu wyjaśnienie zmian molekularnych i neurochemicznych, które towarzyszą morfologicznej restrukturyzacji powłoki MSN skorupy NAc indukowanej przez przedłużone, podobne do obżarstwa pobranie sacharozy.
Materiały i Metody
Oświadczenie o etykiecie
Wszystkie procedury eksperymentalne przeprowadzono zgodnie z australijskim kodeksem opieki i używania zwierząt do celów naukowych, 8th Edition (National Health and Medical Research Council, 2013). Protokoły zostały zatwierdzone przez Komisję Etyki Zwierząt Uniwersytetu w Queensland oraz Komisję Etyki Zwierząt Uniwersytetu Queensland.
Zwierzęta i mieszkania
Pięciotygodniowe (dorastające) samce szczurów Wistar (kontrola: 176.4 ± 4.8 g; sacharoza: 178.3 ± 5.0 g) (ARC, WA, Australia) były indywidualnie trzymane w wentylowanym dwupoziomowym pleksi® klatki. Szczury aklimatyzowano do indywidualnych warunków mieszkaniowych, obchodzenia się z nimi i cyklu 5 w świetle wstecznym przed rozpoczęciem eksperymentów. Wszystkie szczury trzymano w klimatyzowanym 12-hr odwróconym cyklu światło / ciemność (wyłączono światła w 9 am) w pokoju ze standardową karmą dla szczurów i dostępną wodą ad libitum.
Paradygmat picia w dwóch butelkach z przerywanym dostępem
Przerywany dostęp 5% sacharozy wybór dwóch butelek do picia (Simms i in., 2008) został dostosowany z Wise (1973). Wszystkie płyny były prezentowane w plastikowych butelkach miarowych 300 ml z dziobkami do picia ze stali nierdzewnej wkładanymi przez dwa przelotki z przodu klatki po rozpoczęciu cyklu ciemnego światła. Ciężary każdej butelki rejestrowano przed prezentacją butelki. Dwie butelki były prezentowane jednocześnie: jedna butelka zawierająca wodę; druga butelka zawierająca 5% (w / v) sacharozy. Umieszczenie butelki sacharozy 5% (w / v) było naprzemiennie z każdą ekspozycją na kontrolę preferencji bocznych. Butelki ważono 24 h po przedstawieniu płynów, a pomiary prowadzono do najbliższego 0.1 g. Mierzono także masę każdego szczura w celu obliczenia gramów spożycia sacharozy na kilogram masy ciała. W dniu 1 okresu picia, szczury (n = 6 – 9) uzyskano dostęp do jednej butelki 5% (w / v) sacharozy i jednej butelki wody. Po 24 h butelkę z sacharozą zastąpiono drugą butelką z wodą, która była dostępna w następnym 24 h. Ten wzór powtórzył się w środy i piątki. Szczury miały nieograniczony dostęp do wody przez wszystkie pozostałe dni. Zużycie sacharozy podobne do binge powodowało eskalację całkowitego spożycia sacharozy (ml) w czasie (rysunek uzupełniający 1) i towarzyszyły mu stabilne wyjściowe poziomy picia w oparciu o masę ciała [20 ± 5 g / kg 5% (w / v)] w krótkim okresie [~ 4 tygodni (sesje picia 13)] i długoterminowe [ ~ 12 tygodnie (sesje picia 37)] okresy picia. Oddzielna grupa szczurów kontrolnych (n = 6 – 9) uzyskano dostęp do wody w obu butelkach (tj. Bez sacharozy) w tych samych warunkach opisanych powyżej. Średnia masa ciała kontrolnych i szczurów spożywających sacharozę pod koniec krótkotrwałej ekspozycji wynosiła odpowiednio 405.7 ± 40.8 g i 426.4 ± 31.2 g. Pod koniec długotrwałej ekspozycji średnia masa ciała dla grup kontrolnych i sacharozy wynosiła 578.8 ± 53.4 g i 600.2 ± 45.2 g.
Barwienie Golgi-Cox
Po ostatniej sesji picia szczury przeniesiono z ośrodka dla zwierząt, aby umożliwić przetwarzanie próbek mózgu w zakładzie histologicznym w School of Biomedical Sciences, University of Queensland (St Lucia, Australia). Wszystkie zatwierdzone środki zostały podjęte w celu zmniejszenia stresu podczas transportu, po czym szczurom pozwolono na regenerację przez noc. Następnego dnia szczury uśmiercano przez przedawkowanie pentobarbitalu sodu (60-80 mg / kg, ip Vetcare, Brisbane, Australia) i perfundowano dosercowo za pomocą ~ 300 ml sztucznego płynu mózgowo-rdzeniowego, który zawierał (w mM): 130 NaCl, 3 KCl, 26 NaHCO3, 1.25 NaH2PO4, 5 MgCl2, 1 CaCl2i 10 D-glukoza. Następnie każde zwierzę poddano dekapitacji, a mózg usunięto i inkubowano w ciemności w roztworze Golgi-Cox, który zawierał 5% dichromian potasu, 5% chromian potasu i 5% chlorek rtęci (wszystkie substancje chemiczne z Sigma-Aldrich), który był świeżymi dniami 3 przed poświęceniem, jak opisano wcześniej (Rutledge i in., 1969). Zmodyfikowano inkubację barwnika Golgiego-Coxa i metody przetwarzania końcowego Ranjan i Mallick (2010). Mózgi zwierząt krótkoterminowych spożywających sacharozę inkubowano przez 6 dni w 37 ° C, podczas gdy mózgi zwierząt długotrwale spożywających sacharozę inkubowano przez 10 dni, z jedną zmianą na świeży roztwór Golgi-Cox po 4 dniach inkubacji.
Po inkubacji wycinki koronalne 300 μm wycięto przy użyciu mikrotomu wibracyjnego Zeiss Hyrax V50 (Carl Zeiss, Niemcy). Następnie plasterki umieszczano kolejno w płytkach do studzienek 24 wypełnionych sacharozą 30% (wag./obj.) W soli fizjologicznej buforowanej fosforanem 0.1 M i przetwarzano jak opisano w (Ranjan i Mallick, 2010). Pokrótce, skrawki odwodniono w 50% etanolu przez 5 min, a następnie umieszczono w 0.1 M NH4Roztwór OH dla 30 min, przepłukano dwukrotnie wodą destylowaną dla min 5 i umieszczono w utrwalaczu do folii Fujihunt (Fujifilm, Singapur) dla 30 min w ciemności. Następnie plastry płukano dwukrotnie w wodzie destylowanej dla każdej 2 min i odwadniano dwukrotnie w 70, 90, 95 i 100% etanolu dla każdej 5 min. Skrawki następnie oczyszczono w roztworze CXA (1: 1: 1 chloroform: ksylen: alkohol) dla 10 min i zamontowano w DPX (Sigma-Aldrich) na szkiełkach Superfrost Plus (Menzel-Glaser, Lomb Scientific, Australia) i zsuwano (Menzel-Glaser, Niemcy). Szkiełka pozostawiono w ciemności do wyschnięcia w temperaturze pokojowej przez noc.
Wybór neuronów i śledzenie w Nucleus Accumbens
Koronalne plasterki między bregma + 2.8 i + 1.7 badano pod kątem MSN w rdzeniu i powłoce NAc, wykorzystując boczną komorę i spoidło przednie jako punkty orientacyjne za pomocą atlasu mózgu szczura (Paxinos i Watson, 2007) (Rysunek 1). Funkcja konturów w Neurolucida 7 (MBF Bioscience, VT, USA) została wykorzystana do wyznaczenia rdzenia NAc i powłoki NAc w każdym przekroju (rysunek 2). Pomiędzy neuronami 2 i 9 na region na zwierzę śledzono parametry długości dendrytycznej przy użyciu celu 63x lub gęstości kręgosłupa (zgłaszane jako kolce na 100 μm) przy użyciu obiektywu 100x na Zeiss Axioskop II (Carl Zeiss, Niemcy) przy użyciu automatycznego xyz etap napędzany przez Neurolucida® Oprogramowanie 7 (MBF Biosciences, VT, USA). Wszystkie śledzenie przeprowadzono w sposób ślepy w odniesieniu do leczenia. Parametry morfologiczne neuronów nasyconych Golgi-Coxem analizowano w sposób podobny do poprzednich raportów (Klenowski i in., 2015).
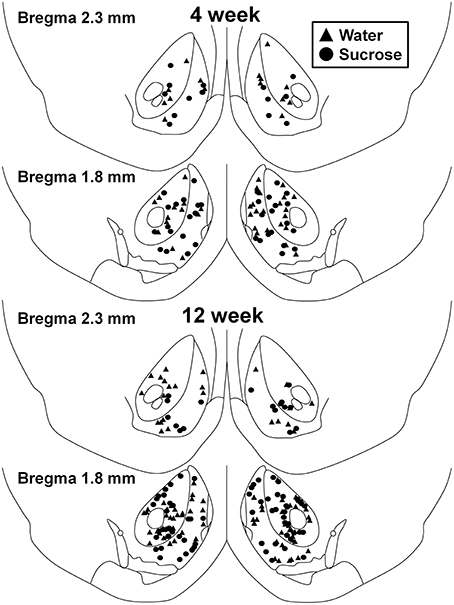
Rysunek 1. Mapa pokazująca lokalizacje średnich neuronów kolczastych pobranych z jądra półleżącego półkuli i skorupy sacharozy 4 i 12 zużywających szczury i kontrole dobrane pod względem wieku. Dwa górne panele pokazują lokalizacje neuronów próbkowanych z jądra półleżącego półkuli i skorupy 4 tygodniowej kontroli (trójkąty) i sacharozy (koła) zwierząt. Dwa dolne panele pokazują pozycje neuronów próbkowanych ze zwierząt kontrolnych 12 (trójkąty) i zwierząt sacharozy (kółka).
Analiza statystyczna
Średni i standardowy błąd średniej (SEM) obliczono dla każdego zestawu danych ze zwierzęciem jako n, używając średnich danych morfometrycznych z wszystkich rdzeniowych lub powłokowych numerów NAN MSN (n = 7 dla powłoki NAc i n = 6 dla rdzenia NAC 4-tydzień, n = 9 dla grup 12-tygodniowych). Tam, gdzie jest to wskazane, niesparowany, dwugonowy Student t-testy lub dwukierunkowe ANOVA z dodatkowymi testami Bonferroni przeprowadzono dla wszystkich analiz obejmujących porównanie średnich grupowych, przy użyciu GraphPad Prism w wersji 6.02 (GraphPad Software, San Diego, CA). Istotność statystyczna została zaakceptowana na P <0.05. Wszystkie dane w sekcji wyników przedstawiono jako średnie ± SEM. Procentowe zmiany są obliczane w odniesieniu do wartości kontrolnej.
Efekt
Średnie kolczaste neurony ze skorupy Nucleus Accumbens mają zmniejszoną długość dendrytyczną, zmniejszoną złożoność dendrytyczną, ale zwiększoną średnią gęstość kręgosłupa w odległych rzędach gałęzi po długim, ale nie krótkotrwałym spożyciu sacharozy
Po krótkotrwałym (4 tygodniach) zużyciu sacharozy, nie było istotnych różnic w parametrach morfometrycznych powłoki MSN NAc (Tabela 1). Nie było również istotnych różnic między krótkotrwałymi konsumpcjami MSN w sacharozie i kontrolnymi MSN w analizach związanych z odśrodkowym porządkiem gałęzi. Mianowicie segmenty dendrytyczne przypadające na kolejność gałęzi (P = 0.4111), średnia długość dendrytyczna na kolejność rozgałęzień (P = 0.5581) i średnia gęstość kręgosłupa na kolejność gałęzi (P = 0.2977, dwukierunkowe ANOVA) nie różniły się znacząco między grupami. Mapa lokalizacji pokazująca przybliżone pozycje próbkowanych neuronów jest pokazana na rysunku 1.
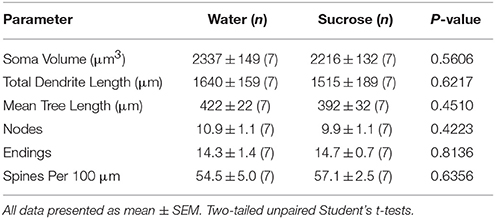
Tabela 1. Ogólne parametry morfologiczne średnich kolczastych neuronów z jądra półleżącego skorupki krótkookresowej sacharozy spożywającej szczury i dopasowane do wieku kontrole wody.
Po długotrwałym (12 tygodniach) zużyciu sacharozy, całkowita długość dendrytycznej altany powłoki MSN powłoki NAc została zmniejszona o 21% w porównaniu z kontrolami zużywającymi wodę (Woda: 1827 ± 148 μm, n = 9; Sacharoza 1449 ± 78 μm, n = 9, *P = 0.0384, dwustronne niesparowane studenta t-test, ryc 2, Stół 2). Porównanie średniej liczby bifurkacji dendrytycznych (węzłów) i zakończeń dendrytycznych między grupami wody i sacharozy ujawniło obniżony (choć nieistotny) poziom złożoności dendrytycznej w MSN powłoki NAc (węzły: woda 12.9 ± 1.4 n = 9, sacharoza 10.1 ± 0.8 n = 9, P = 0.0879; zakończenia: Water 17.9 ± 1.4 n = 9, sacharoza 14.8 ± 0.7 n = 9, P = 0.0657, dwustronne niesparowane studenta t-test, Tabela 2). Nie było zmiany objętości somy (P = 0.9400), średnia długość drzewa dendrytycznego (P = 0.1646) lub całkowita gęstość kręgosłupa (P = 0.3662) w skorupkach NAc MSN od szczurów długotrwale spożywających sacharozę w porównaniu z próbami kontrolnymi wody. Te parametry morfometryczne są wyszczególnione w tabeli 2.
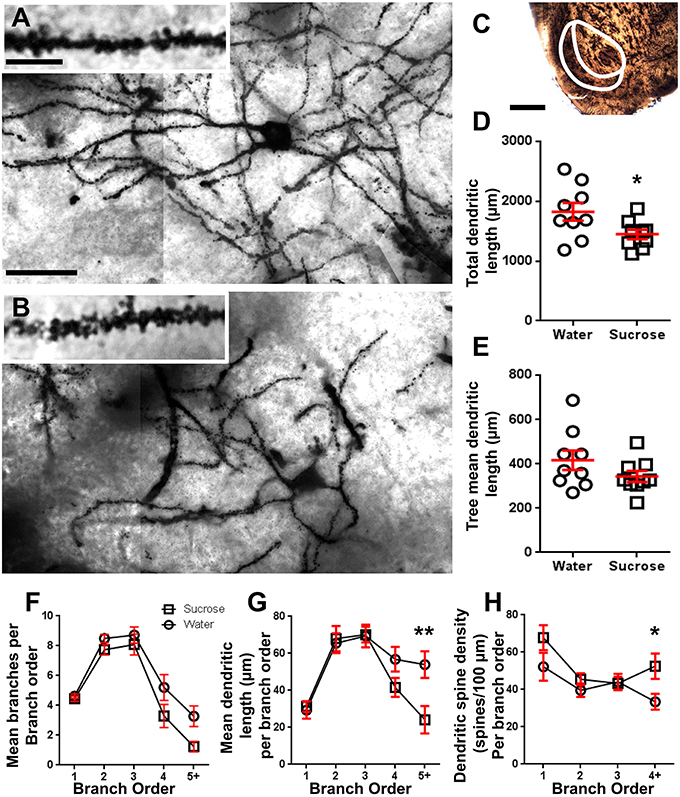 Rycina 2. Zmniejszona długość trzpienia dendrytycznego i zwiększona dystalna gęstość dendrytycznego kręgosłupa średnich neuronów kolczastych (MSN) z powłoki jądra półleżącego (NAc) szczurów długo leczonych sacharozą w porównaniu do szczurów kontrolnych. (A, B) pokazują reprezentacje kontrolnego (u góry) i długoterminowego (tydzień 12) sacharozy (u dołu) traktowanego jasnego pola z-stak mozaiki impregnowanych Golgim MSN z powłoki NAc (powiększenie 63x). Wstawka (A, B) pokazuje kontrolne i długoterminowe obrazy jasnego pola traktowane sacharozą impregnowanych Golgim dendrytów MSN i kolców dendrytycznych z powłoki NAc (powiększenie 100x). (DO) pokazuje regiony anatomiczne, z których próbki MSN pobrano w tym badaniu. (D) pokazuje wykres punktowy zmniejszonej całkowitej altany dendrytycznej MSN (średnia ± SEM) ze skorupy NAc u długoterminowych zwierząt z sacharozy (kwadraty) w porównaniu do kontroli (koła), niesparowanych uczniów t-test *P <0.05, n = 9; kontrola i n = 9; 12 tydzień sacharoza. (MI) pokazuje wykres punktowy niezmienionej średniej długości drzewa dendrytycznego MSN (średnia ± SEM) ze skorupy NAc u długoterminowych zwierząt z sacharozy (kwadraty) w porównaniu do kontroli (koła), niesparowanych uczniów t-test, P > 0.05, n = 9; kontrola i n = 9; 12 tydzień sacharoza. Analiza kolejności gałęzi (średnia ± SEM) numeru segmentu dendrytycznego na zamówienie gałęzi (F), średnia długość dendrytyczna na zamówienie oddziału (G) i gęstość kręgosłupa dendrytycznego według rzędu gałęzi (H). Długotrwałe spożycie sacharozy zmniejszyło długość dendrytów przy dystalnych rzędach gałęzi (5 +) i zwiększyło gęstość kręgosłupa dendrytycznego przy dystalnych rzędach gałęzi (4 +) w porównaniu do kontroli (G, H), dwukierunkowe ANOVA z posttestami Bonferroniego, *P <0.05, **P <0.01, n = 9; kontrola i n = 9; długoterminowa sacharoza. Paski skali: (A, B) = 20 μm; wstawka z (A, B) = 10 μm; (DO) = 1 mm.
Rycina 2. Zmniejszona długość trzpienia dendrytycznego i zwiększona dystalna gęstość dendrytycznego kręgosłupa średnich neuronów kolczastych (MSN) z powłoki jądra półleżącego (NAc) szczurów długo leczonych sacharozą w porównaniu do szczurów kontrolnych. (A, B) pokazują reprezentacje kontrolnego (u góry) i długoterminowego (tydzień 12) sacharozy (u dołu) traktowanego jasnego pola z-stak mozaiki impregnowanych Golgim MSN z powłoki NAc (powiększenie 63x). Wstawka (A, B) pokazuje kontrolne i długoterminowe obrazy jasnego pola traktowane sacharozą impregnowanych Golgim dendrytów MSN i kolców dendrytycznych z powłoki NAc (powiększenie 100x). (DO) pokazuje regiony anatomiczne, z których próbki MSN pobrano w tym badaniu. (D) pokazuje wykres punktowy zmniejszonej całkowitej altany dendrytycznej MSN (średnia ± SEM) ze skorupy NAc u długoterminowych zwierząt z sacharozy (kwadraty) w porównaniu do kontroli (koła), niesparowanych uczniów t-test *P <0.05, n = 9; kontrola i n = 9; 12 tydzień sacharoza. (MI) pokazuje wykres punktowy niezmienionej średniej długości drzewa dendrytycznego MSN (średnia ± SEM) ze skorupy NAc u długoterminowych zwierząt z sacharozy (kwadraty) w porównaniu do kontroli (koła), niesparowanych uczniów t-test, P > 0.05, n = 9; kontrola i n = 9; 12 tydzień sacharoza. Analiza kolejności gałęzi (średnia ± SEM) numeru segmentu dendrytycznego na zamówienie gałęzi (F), średnia długość dendrytyczna na zamówienie oddziału (G) i gęstość kręgosłupa dendrytycznego według rzędu gałęzi (H). Długotrwałe spożycie sacharozy zmniejszyło długość dendrytów przy dystalnych rzędach gałęzi (5 +) i zwiększyło gęstość kręgosłupa dendrytycznego przy dystalnych rzędach gałęzi (4 +) w porównaniu do kontroli (G, H), dwukierunkowe ANOVA z posttestami Bonferroniego, *P <0.05, **P <0.01, n = 9; kontrola i n = 9; długoterminowa sacharoza. Paski skali: (A, B) = 20 μm; wstawka z (A, B) = 10 μm; (DO) = 1 mm.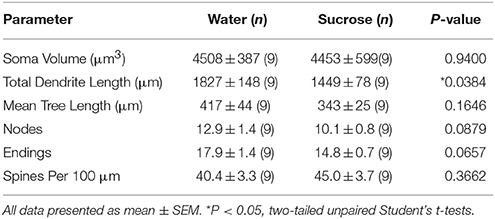
Tabela 2. Ogólne parametry morfologiczne średnich neuronów kolczastych z jądra półleżącego skorupę długoterminowych szczurów spożywających sacharozę i dobrane do wieku kontrole wodne.
Po scharakteryzowaniu ogólnej morfologii dendrytycznej długookresowych MSN-ów z powłoką NAc, sacharozą, przeanalizowaliśmy arborizacje dendrytyczne i gęstość kręgosłupa w odniesieniu do ich charakterystyk rzędu gałęzi. Nasza kompleksowa ocena drzew dendrytycznych określiła ilościowo liczbę segmentów dendrytycznych na rząd gałęzi, średnią długość segmentów dendrytycznych na rząd gałęzi i średnią gęstość kręgosłupa na rząd gałęzi MSN powłoki NAc kontrolującej wodę i długookresowych szczurów spożywających sacharozę. Podsumowanie danych i analiz zamówień w oddziale znajduje się w tabeli 3.
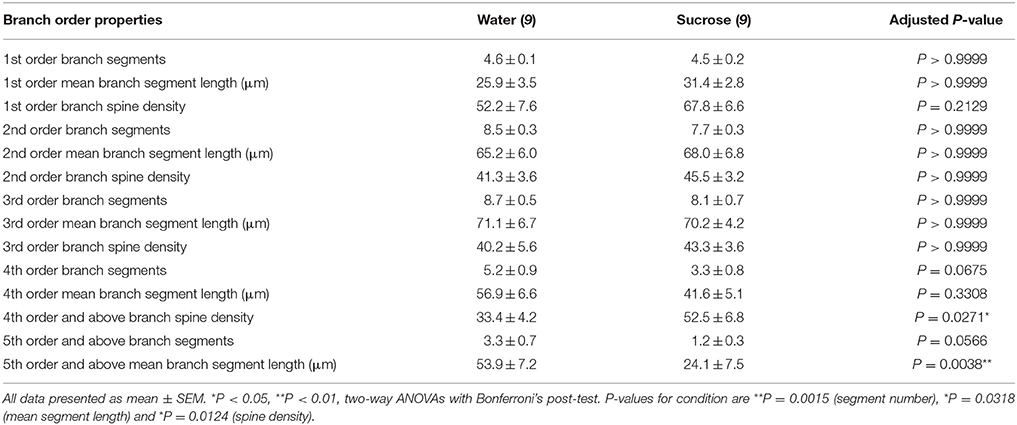 Tabela 3. Charakterystyka rzędów średnich neuronów kolczastych od szczurów pijących sacharozę długo i pijących wodę.
Tabela 3. Charakterystyka rzędów średnich neuronów kolczastych od szczurów pijących sacharozę długo i pijących wodę.Średnia liczba segmentów gałęzi dendrytycznych na rząd gałęzi MSN powłoki NAc była znacznie zmniejszona u szczurów spożywających sacharozę w długim okresie w porównaniu do kontroli wody (**P = 0.0015, dwukierunkowa ANOVA). Post-testy Bonferroni wykazały trend w kierunku zmniejszenia liczby segmentów gałęzi na 4th (woda: 5.2 ± 0.9, n = 9; Sacharoza 3.3 ± 0.8, n = 9, P = 0.0675, rysunek 2F, Stół 3) oraz 5-ty rząd i powyżej zamówień oddziału (woda: 3.3 ± 0.7, n = 9; Sacharoza 1.2 ± 0.3, n = 9, P = 0.0566, rysunek 2F, Stół 3). Średnia długość segmentu dendrytycznego na rząd gałęzi MSN powłoki NAc została również znacznie zmniejszona u szczurów spożywających sacharozę w długim okresie w porównaniu do kontroli wody (*P = 0.0444, dwukierunkowa ANOVA). Post-testy Bonferroniego wykazały zmniejszenie 55% w gałęziach 5 rzędu i później (woda: 53.9 ± 7.2 μm, n = 9; Sacharoza 24.1 ± 7.5 μm, n = 9, **P = 0.0038, rysunek 2G, Stół 3).
Analiza kolejności gałęzi wykazała znaczny wzrost gęstości dendrytycznego kręgosłupa MSN powłoki NAc u szczurów długotrwale spożywających sacharozę w porównaniu do kontroli (*P = 0.0124, dwukierunkowa ANOVA). Post-testy Bonferroniego wykazały wzrost gęstości kręgosłupa o 57% w dystalnych gałęziach 4 rzędu i dalej (woda: 33.4 ± 4.2, n = 9; Sacharoza 52.5 ± 6.8, n = 9, P = 0.0271 *, wstawka z rysunkami 2A, B, H, Stół 3). Reprezentatywne obrazy ogólnej architektury MSN i dystalnej gęstości kręgosłupa (wstawki) przedstawiono na rycinach 2A, B.
Podsumowując, wyniki te wskazują, że krótkotrwałe spożycie sacharozy ma niewielki wpływ na parametry morfologiczne MSN w powłoce NAc. Jednak po długotrwałym spożywaniu następuje znaczny spadek długości i złożoności trzonu neuronu, szczególnie w dystalnych gałęziach dendrytycznych. Równoczesny wzrost gęstości dystalnej kręgosłupa jest również widoczny w skorupkach MSN skorup NAc długoterminowych szczurów spożywających sacharozę.
Średnie kolczaste neurony z jądra jądra półleżącego zmniejszyły złożoność rozgałęzień po długim, ale nie krótkotrwałym zużyciu sacharozy
Po krótkotrwałym spożyciu sacharozy nie stwierdzono istotnych różnic w parametrach morfometrycznych rdzenia MSN NAc (Tabela 4). Nie stwierdzono również istotnych różnic między zużyciem sacharozy w tygodniu 4 a podstawowymi MSN kontrolującymi wodę w analizach związanych z porządkiem gałęzi odśrodkowej. Mianowicie, segmenty dendrytyczne na zamówienie oddziału (P = 0.7717), średnia długość dendrytyczna na kolejność rozgałęzień (P = 0.2096) i średnia gęstość kręgosłupa na kolejność gałęzi (P = 0.3521, dwukierunkowe ANOVA) nie różniły się między grupami.
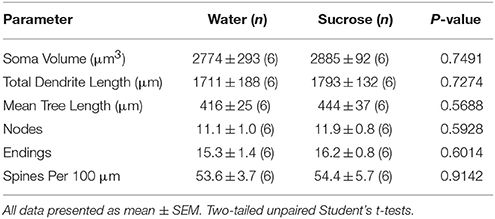 Tabela 4. Ogólne parametry morfologiczne średnich kolczastych neuronów z jądra półleżącego jądra półleżącego sacharozy spożywających szczury i kontrolowane wiekiem kontrole wody.
Tabela 4. Ogólne parametry morfologiczne średnich kolczastych neuronów z jądra półleżącego jądra półleżącego sacharozy spożywających szczury i kontrolowane wiekiem kontrole wody.Przedłużone spożycie sacharozy nie miało również istotnego wpływu na parametry morfometryczne rdzenia MSN NAc (tabela 5). Średnia liczba segmentów gałęzi dendrytycznej przypadająca na kolejność rozgałęzień MSN rdzenia NAc była znacznie zmniejszona w długotrwałych szczurach spożywających sacharozę w porównaniu do kontroli wody (*P = 0.0416, dwukierunkowa ANOVA), jednak nie było istotnych różnic w średniej długości dendrytycznej na kolejność rozgałęzień (P = 0.0995) i średnia gęstość kręgosłupa na kolejność gałęzi (P = 0.4888, dwukierunkowe ANOVA) między MSN w rdzeniu NAc długotrwałych szczurów spożywających sacharozę w porównaniu z kontrolami wody. Podsumowując, nasze dane pokazują, że rdzeń NAc nie jest tak wrażliwy na długotrwałe zużycie sacharozy w porównaniu z MSN z regionu powłoki NAc.
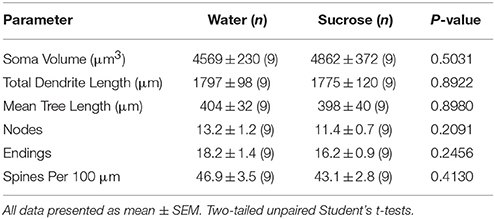 Tabela 5. Ogólne parametry morfologiczne średnich kolczastych neuronów z jądra półleżącego jądra półleżącego długotrwałej sacharozy spożywających szczury i kontrolowane wiekiem kontrole wody.
Tabela 5. Ogólne parametry morfologiczne średnich kolczastych neuronów z jądra półleżącego jądra półleżącego długotrwałej sacharozy spożywających szczury i kontrolowane wiekiem kontrole wody.Dyskusja
Zwiększona dostępność wysoko słodzonych pokarmów w zachodniej diecie nie tylko przyczyniła się do wzrostu częstości występowania i obciążenia ekonomicznego otyłością i cukrzycą typu II, ale także doprowadziła do wystąpienia zaburzeń odżywiania, takich jak objadanie się (Swanson i in., 2011; Kessler i in., 2013; Davis, 2015). Chociaż uzależniające właściwości cukrów, w tym fruktozy i sacharozy, pozostają spekulatywne, istnieje uderzające podobieństwo korelacji behawioralnych i neuronalnych, które objawiają się w wyniku nadmiernego jedzenia i długotrwałego używania narkotyków (Avena i wsp., 2008, 2011). Ponadto cukier aktywuje obwód nagrody w mózgu w sposób podobny do narkotyków (Volkow i wsp., 2012), a wyniki badań na ludziach sugerują, że cukier i słodycz mogą wywoływać łaknienie, które są porównywalne pod względem wielkości z tymi wywoływanymi przez uzależniające leki, takie jak alkohol i nikotyna (Volkow i wsp., 2012). Dlatego wykorzystaliśmy model konsumpcji binge-sacharozy u szczurów w celu określenia wpływu krótkiego (4 tygodnie) i długotrwałego (12 tygodnie) zużycia sacharozy na neuronalną morfologię MSN w NAc, kluczowym składniku nakładających się obwodów nagrody który jest modulowany przez cukier i uzależniające leki. Pokazujemy, że MSN ze skorupy NAc przewlekłych długotrwałych szczurów spożywających sacharozę znacznie zmniejszyły długość i złożoność dendrytyczną, ale zwiększyły gęstość dystalnego kręgosłupa dendrytycznego. Długotrwałe zużycie sacharozy nie miało wpływu na morfologię MSN z rdzenia NAc, podczas gdy krótkotrwałe zużycie sacharozy nie miało również istotnego wpływu na morfologię MSN z rdzenia lub powłoki NAc. Wyniki te nie tylko wskazują na bezpośredni wpływ przedłużonego przyjmowania sacharozy typu „binge-like” na morfologię neuronów w MSN muszli NAc, ale także podkreślają potencjalnie szkodliwe konsekwencje przedłużonego spożywania diet zawierających dużo cukru.
NAc, który stanowi część prążkowia brzusznego, składa się głównie z MSN, które charakteryzują się morfologicznie jako neurony średniej wielkości z rozległymi arborisjami dendrytycznymi i wysoką gęstością kręgosłupa (Kemp i Powell, 1971; Graveland i DiFiglia, 1985; Rafols i in., 1989; Kawaguchi i in., 1990). Neurony glutaminergiczne i dopaminergiczne są dwoma głównymi wejściowymi doprowadzeniami do NAc, głównie stykającymi się z trzonami dendrytycznymi i kolcami MSN (Gaje, 1980; Kaiya i Namba, 1981; Groves i in., 1994). W szczególności powłoka i rdzeń NAc otrzymują wkład glutaminergiczny z funkcjonalnie różnych obszarów korowych (Brog i in., 1993). Powłoka NAc jest również unerwiona przez pobudzające doprowadzające komórki z obszarów podkorowych, takich jak hipokamp, wzgórze i ciało boczno-boczne. (Brog i in., 1993; Wright i Groenewegen, 1995). Poprzednie badania wykazały, że te dane wejściowe glutaminergiczne odgrywają kluczową rolę w motywowaniu i zachowaniach ukierunkowanych na cel, takich jak poszukiwanie żywności i nagroda (Maldonado-Irizarry i in., 1995; Kelley i Swanson, 1997; Reynolds i Berridge, 2003; Richard and Berridge, 2011). Drugi dominujący wkład w MSNs NAc pochodzi z dopaminergicznych aferentów, które wystają z brzusznego obszaru nakrywkowego (Lindvall i Björklund, 1978; Veening i in., 1980; Kalivas i Miller, 1984). Co ciekawe, poprzednie badania wykorzystujące podobne modele przerywanego dostępu do cukru wykazały, że wynikające z tego zjawisko konsumpcji podobnej do obżarstwa powoduje wzrost pozakomórkowej dopaminy w NAc podobnie (choć w mniejszym stopniu) do leków nadużywanych (Rada i in., 2005; Avena i wsp., 2006) i może modulować ekspresję receptora dopaminy (Colantuoni i wsp., 2001, 2002) w rdzeniu i powłoce NAc. Co ciekawe, nadmierne spożywanie sacharozy powoduje eskalację spożycia w miarę upływu czasu, podobnie jak samo-podawanie narkotyków, takich jak kokaina i heroin (Ahmed i Koob, 1998; Ahmed i wsp., 2000, 2003), który jest związany z rozwojem stanu „uzależniającego”.
Nasza analiza morfometrii rzędu rozgałęzień pokazuje, że ogólna redukcja długości dendrytycznej powłoki MSN skorupy NAc spowodowana długotrwałym spożyciem sacharozy, wynika przede wszystkim ze zmniejszenia złożoności dystalnych zamówień gałęzi. Zaobserwowaliśmy zmniejszone rozgałęzienia dystalne (4th i 5th order i powyżej rozkazy rozgałęzień) i znacznie zmniejszoną średnią długość w 5th rzędzie i powyżej dendrytów, w połączeniu ze zwiększoną gęstością kręgosłupa w tych rzędach gałęzi. Wspólny czynnik, który może wpłynąć na ten typ restrukturyzacji dendrytycznej, obejmuje zmiany w łączności synaptycznej i / lub funkcji (Russo i wsp., 2010). Poprzednie badania wykazały, że synapsy glutaminergiczne na MSN powstają głównie na kolcach, szczególnie w dystalnych dendrytach (Groenewegen i in., 1999). Dodatkowo kolokalizacja dopaminy i wejść glutaminergicznych z kory przedczołowej (Sesack i Pickel, 1992), hipokamp (Totterdell and Smith, 1989; Sesack i Pickel, 1990), i ciało migdałowate (Johnson i in., 1994) zaobserwowano na kolcach dendrytycznych MSN. Obserwacje te w połączeniu ze zwiększoną gęstością kręgosłupa po długotrwałym spożyciu sacharozy obserwowane w naszym badaniu, wspierają powstawanie zwiększonych pobudzeń pobudzających. W związku z tym pojawia się możliwość, w której trwałe skutki spowodowane przedłużonym upijaniem się sacharozy mogą ułatwić zwiększoną pobudzającą aktywność synaptyczną w dalszych dendrytach MSN w powłoce NAc. W konsekwencji redukcja i / lub wycofanie dystalnych dendrytów może wynikać z synaptycznego mechanizmu homeostatycznego (Reissner i Kalivas, 2010), jednak pozostaje to do ustalenia.
Warto zauważyć, że Crombag i jego współpracownicy wykazali, że nie nastąpił wzrost gęstości kręgosłupa w powłoce NAc po spożyciu sacharozy w tygodniu 4 przez paradygmat samozaparcia przez nos, pomimo silniejszego przejęcia i wyższego odsetka odpowiedzi na sacharozę w porównaniu z amfetaminą (Crombag i in., 2005). Ich obserwacja braku zmiany gęstości kręgosłupa w tygodniach 4 odzwierciedla nasze odkrycia. W przeciwieństwie do tego, nasze badanie pokazuje, że po długotrwałej (12 tydzień) ekspozycji na przewlekłe spożycie sacharozy, istnieje znaczny wzrost dystalnej gęstości kręgosłupa na MSN szczurów doświadczających sacharozy. Ponadto nasze laboratorium wykazało wcześniej, że długotrwałe (12 tydzień) zużycie sacharozy ułatwia różnicową odpowiedź farmakologiczną na farmakoterapeutyki, które, jak wykazano, modulują reakcje dopaminy i acetylocholiny na poziomie NAc (Shariff i in., W druku). Podsumowując, sugeruje to, że ekspozycja na sacharozę długoterminową (12 tygodni i więcej), która jest dokładniej odzwierciedlona w rzeczywistych scenariuszach, powoduje adaptacje morfologii na poziomie NAc.
Jeśli chodzi o narkotyki, powtarzane narażenie na różne leki powoduje długotrwałe zmiany w strukturze dendrytów i kolców dendrytycznych. Na przykład amfetaminy i kokaina zwiększają gęstość kręgosłupa w NAc zarówno w skorupie, jak i rdzeniu (Robinson i Kolb, 2004). Wykazano również, że ekspozycja na nikotynę zwiększa gęstość kręgosłupa w powłoce NAc. Odwrotnie, ekspozycja na morfinę prowadzi do zmniejszenia gęstości kręgosłupa i złożoności gałęzi dendrytycznej (Robinson i Kolb, 2004). Jeśli chodzi o długotrwałe spożywanie sacharozy, zaobserwowaliśmy wzrost gęstości kręgosłupa podobny do amfetaminy, kokainy i nikotyny i przeciwny do działania morfiny. Jednakże, w przeciwieństwie do amfetaminy i kokainy, ale podobnie jak nikotyna, wzrost gęstości kręgosłupa przy długotrwałej ekspozycji na sacharozę jest ograniczony do powłoki NAc. Interesujące jest również to, że zmiany w obu rozgałęzieniach dendrytycznych (Robinson i Kolb, 1999) i gęstość kręgosłupa (Li i wsp., 2003) wytwarzane przez amfetaminę lub kokainę są ograniczone do dystalnych dendrytów MSN w NAc, co odzwierciedla wyniki naszego badania. Ponadto, potwierdzając opisane powyżej zmiany, wykazano również, że zużycie sacharozy zwiększa pobudzającą siłę synaptyczną na odkładające się neurony dopaminowe (Stuber i in., 2008b), jak również inne składniki ścieżki nagrody mezolimbicznej (Stuber i in., 2008a; Chen i wsp., 2010). Podsumowując, zakłada to, że sacharoza jest silnym modulatorem morfologii neuronów po długotrwałym intensywnym użytkowaniu, co jest podobne do efektów obserwowanych w przypadku narkotyków.
Chociaż konieczne są dalsze badania, aby odkryć mechanizmy komórkowe i synaptyczne przyczyniające się do zmian morfologicznych obserwowanych w tym badaniu, nasze wyniki wykazują znaczące efekty neuronalne wywołane długotrwałym spożyciem sacharozy. W szczególności, nie przeanalizowano w naszym badaniu, czy obserwowane efekty morfologiczne sacharozy mogą być również wywołane przez niekaloryczne substancje słodzące, takie jak sacharyna. W tym względzie należy zauważyć, że Lenoir i współpracownicy wykazali, że intensywna słodycz przewyższa nagrodę kokainową, czy to przez sacharynę, czy sacharozę (Lenoir i in., 2007). Ponadto ostatnie badanie opublikowane przez nasze laboratorium (Shariff i in., W druku) wykazuje, że wareniklina, nikotynowy częściowy agonista receptora acetylocholinowego, zmniejszyła zarówno spożycie sacharozy, jak i sacharyny u gryzoni, stosując ten sam długoterminowy schemat przerywanego dostępu stosowany w niniejszym badaniu. Co ciekawe, poprzednie badania wykazały podobieństwa w ostrych skutkach niekalorycznych substancji słodzących, takich jak sacharyna i sacharoza, na poziomie NAc (Scheggi i in., 2013; Tukey i in., 2013; Carelli i West, 2014). Konieczne są jednak dalsze badania w celu ustalenia, czy niekaloryczne substancje słodzące mogą wywołać długotrwałe skutki podobne do zmian morfologii powłoki MSN w NAc spowodowanej długotrwałym zgłoszonym tu spożyciem sacharozy.
Brak wpływu na morfologię MSN NAc po krótkotrwałym spożyciu sacharozy, podkreśla znaczenie wdrażania długoterminowych badań w celu oceny wpływu przedłużającego się nadużywania leków lub naturalnych nagród, takich jak sacharoza. Jeśli chodzi o zależność, powtarzane cykle przyjmowania napadów i abstynencji są kluczowymi składnikami cyklu uzależnień, a coraz więcej dowodów ujawnia, że przejście na uzależnienie jest procesem postępującym, który często zachodzi przez dłuższy okres czasu. Chociaż uzależniające właściwości cukrów pozostają niepewne, coraz bardziej bada się prawdopodobieństwo uzależnienia od innych nagród nielekowych, takich jak seks, hazard i żywność. Wyniki tego badania dodają zasługi hipotezie, że cukry, takie jak sacharoza, potencjalnie mają właściwości uzależniające po długotrwałej konsumpcji podobnej do binge. Nasze wyniki mają również wpływ na rosnącą liczbę dzieci i młodzieży, którzy zachowują niezdrowe nawyki żywieniowe (wysokie spożycie cukru i objadanie się) do wieku dorosłego. Zgodnie ze zwiększonym ryzykiem wystąpienia efektów metabolicznych możliwe jest również, że konsekwencje neurologiczne i psychiatryczne wpływające na nastrój i motywację mogą również wynikać z tych zachowań.
Autorskie Wkłady
Udział w projekcie badawczym: PK, SB. Przeprowadzone eksperymenty: PK, MS, AB, MF, EM. Analiza danych: PK, MF, MS. Interpretował dane i przyczynił się do napisania manuskryptu: PK, MS, MF, EM, MB, SB. Wszyscy autorzy przeczytali i zatwierdzili ostateczny tekst do złożenia.
Oświadczenie o konflikcie interesów
Autorzy oświadczają, że badanie zostało przeprowadzone przy braku jakichkolwiek powiązań handlowych lub finansowych, które mogłyby być interpretowane jako potencjalny konflikt interesów.
Recenzenci SC, SA i redaktor zarządzający zadeklarowali wspólną przynależność, a redaktor obsługujący stwierdza, że mimo to proces spełniał standardy uczciwego i obiektywnego przeglądu.
Podziękowanie
Prace te były wspierane finansowaniem z australijskiej Rady ds. Badań Naukowych (FT1110884) na rzecz SB i Krajowej Rady Badań Zdrowia i Badań Medycznych (1061979) na SB i MB.
Materiał uzupełniający
Dodatkowe materiały do tego artykułu można znaleźć w Internecie pod adresem: http://journal.frontiersin.org/article/10.3389/fnbeh.2016.00054
Rysunek uzupełniający 1. Spożycie sacharozy i preferencja z 4 i 12 tydzień sacharozy spożywania szczurów. (A, B) wykazują eskalację całkowitego spożycia sacharozy (ml) przez tygodnie ekspozycji 4 i 12. (PŁYTA CD) wykazują wysoką preferencję dla sacharozy nad wodą podczas okresów prezentacji sacharozy.
Referencje
Ahmed, SH i Koob, GF (1998). Przejście od umiarkowanego do nadmiernego przyjmowania leków: zmiana wartości hedonicznej. nauka 282, 298 – 300. doi: 10.1126 / science.282.5387.298
Ahmed, SH, Lin, D., Koob, GF i Parsons, LH (2003). Eskalacja samopodawania kokainy nie zależy od zmienionych indukowanych kokainą jąder półleżących poziomów dopaminy. J. Neurochem. 86, 102-113. doi: 10.1046 / j.1471-4159.2003.01833.x
Ahmed, SH, Walker, JR i Koob, GF (2000). Utrzymujący się wzrost motywacji do przyjmowania heroiny u szczurów z historią eskalacji narkotyków. Neuropsychopharmacology 22, 413–421. doi: 10.1016/S0893-133X(99)00133-5
Arslanian, S. (2002). Cukrzyca typu 2 u dzieci: aspekty kliniczne i czynniki ryzyka. Horm Res 57 (Suppl. 1), 19 – 28. doi: 10.1159 / 000053308
Avena, NM, Bocarsly, ME, Hoebel, BG i Gold, MS (2011). Nakładanie się na nozologię nadużywania substancji i przejadania się: translacyjne implikacje „uzależnienia od żywności”. Curr. Nadużywanie narkotyków Rev. 4, 133-139. doi: 10.2174 / 1874473711104030133
Avena, NM, Rada, P. i Hoebel, BG (2008). Dowody na uzależnienie od cukru: skutki behawioralne i neurochemiczne przerywanego, nadmiernego spożycia cukru. Neurosci. Biobehav. Obrót silnika. 32, 20-39. doi: 10.1016 / j.neubiorev.2007.04.019
Avena, NM, Rada, P., Moise, N., i Hoebel, BG (2006). Karmienie fałszywą sacharozą w harmonogramie napadowym uwalnia wielokrotnie dopaminę i eliminuje reakcję sytości acetylocholiny. Neuroscience 139, 813 – 820. doi: 10.1016 / j.neuroscience.2005.12.037
Benton, D. (2010). Wiarygodność uzależnienia od cukru i jego rola w otyłości i zaburzeniach odżywiania. Clin. Nutr. 29, 288 – 303. doi: 10.1016 / j.clnu.2009.12.001
Bray, GA i Popkin, BM (2014). Cukier dietetyczny i masa ciała: czy doszło do kryzysu epidemii otyłości i cukrzycy? Wlać cukier. Diabetes Care 37, 950 – 956. doi: 10.2337 / dc13-2085
Brog, JS, Salyapongse, A., Deutch, AY i Zahm, DS (1993). Wzory doprowadzającego unerwienia rdzenia i powłoki w „półleżącej” części prążkowia brzusznego szczura: immunohistochemiczne wykrycie wstecznie transportowanego fluoro-złota. J. Comp. Neurol. 338, 255 – 278. doi: 10.1002 / cne.903380209
Bucher Della Torre, S., Keller, A., Laure Depeyre, J. i Kruseman, M. (2015). Napoje słodzone cukrem i ryzyko otyłości u dzieci i młodzieży: systematyczna analiza wpływu jakości metodologicznej na wnioski. J. Acad. Nutr. Dieta. [Wydanie elektroniczne przed papierowym]. doi: 10.1016 / j.jand.2015.05.020
Carelli, RM i West, EA (2014). Kiedy dobry smak staje się zły: mechanizmy neuronalne leżące u podstaw powstawania negatywnego afektu i związana z tym naturalna dewaluacja nagrody przez kokainę. Neuropharmacology 76 (Pt B), 360 – 369. doi: 10.1016 / j.neuropharm.2013.04.025
Chen, BT, Hopf, FW i Bonci, A. (2010). Plastyczność synaptyczna w układzie mezolimbicznym: implikacje terapeutyczne dla nadużywania substancji. Ann. NY Acad. Sci. 1187, 129-139. doi: 10.1111 / j.1749-6632.2009.05154.x
Colantuoni, C., Rada, P., McCarthy, J., Patten, C., Avena, NM, Chadeayne, A., i in. (2002). Dowody na to, że przerywane, nadmierne spożycie cukru powoduje endogenne uzależnienie od opioidów. Obes. Res. 10, 478 – 488. doi: 10.1038 / oby.2002.66
Colantuoni, C., Schwenker, J., McCarthy, J., Rada, P., Ladenheim, B., Cadet, JL, et al. (2001). Nadmierne spożycie cukru zmienia wiązanie z dopaminą i receptorami opioidowymi mu w mózgu. Neuroreport 12, 3549–3552. doi: 10.1097/00001756-200111160-00035
Crombag, HS, Gorny, G., Li, Y., Kolb, B. i Robinson, TE (2005). Przeciwne skutki samo-podawania amfetaminy na kolce dendrytyczne w przyśrodkowej i oczodołowej korze przedczołowej. Cereb. Kora 15, 341 – 348. doi: 10.1093 / cercor / bhh136
Dabelea, D., Mayer-Davis, EJ, Saydah, S., Imperatore, G., Linder, B., Divers, J., i in. (2014). Częstość występowania cukrzycy typu 1 i 2 wśród dzieci i młodzieży od 2001 do 2009. JAMA 311, 1778 – 1786. doi: 10.1001 / jama.2014.3201
Davis, C. (2015). Epidemiologia i genetyka zaburzeń objadania się (BED). CNS Spectr. 20, 522 – 529. doi: 10.1017 / s1092852915000462
Ford, ES i Dietz, WH (2013). Tendencje w poborze energii wśród dorosłych w Stanach Zjednoczonych: ustalenia z NHANES. Rano. J. Clin. Nutr. 97, 848 – 853. doi: 10.3945 / ajcn.112.052662
Fryar, CD, Carroll, MD i Ogden, CL (2014). Rozpowszechnienie nadwagi, otyłości i skrajnej otyłości wśród dorosłych: Stany Zjednoczone, 1960 – 1962 przez 2011 – 2012. Atlanta, GA: CDC.
Graveland, GA i DiFiglia, M. (1985). Częstotliwość i rozmieszczenie neuronów średniej wielkości z wciętymi jądrami w naczelnym i neostriatum gryzonia. Brain Res. 327, 307–311. doi: 10.1016/0006-8993(85)91524-0
Groenewegen, HJ, Wright, CI, Beijer, AV i Voorn, P. (1999). Zbieżność i segregacja brzusznych wejść i wyjść prążkowia. Ann. NY Acad. Sci. 877, 49–63. doi: 10.1111/j.1749-6632.1999.tb09260.x
Groves, PM (1980). Końcówki synaptyczne i ich postynaptyczne cele w neostriatum: specjalizacje synaptyczne ujawnione na podstawie analizy przekrojów seryjnych. Proc. Natl. Acad Sci. USA 77, 6926 – 6929. doi: 10.1073 / pnas.77.11.6926
Groves, PM, Linder, JC i Young, SJ (1994). Aksony dopaminergiczne znakowane hydroksydopaminą 5: trójwymiarowe rekonstrukcje aksonów, synaps i postynaptycznych celów w neostriatum szczura. Neuroscience 58, 593–604. doi: 10.1016/0306-4522(94)90084-1
Hu, FB (2013). Rozwiązano: istnieją wystarczające dowody naukowe, że zmniejszenie spożycia słodzonych napojów zmniejszy częstość występowania otyłości i chorób związanych z otyłością. Obes. Obrót silnika. 14, 606 – 619. doi: 10.1111 / obr.12040
Johnson, LR, Aylward, RL, Hussain, Z. i Totterdell, S. (1994). Wejście z ciała migdałowatego do jądra półleżącego szczura: jego związek z immunoreaktywnością hydroksylazy tyrozynowej i zidentyfikowanymi neuronami. Neuroscience 61, 851–865. doi: 10.1016/0306-4522(94)90408-1
Kaiya, H. i Namba, M. (1981). Dwa typy dopaminergicznych zakończeń nerwowych w neostriatum szczura. Badanie ultrastrukturalne. Neurosci. Łotysz. 25, 251–256. doi: 10.1016/0304-3940(81)90400-6
Kalivas, PW i Miller, JS (1984). Neurony neuronów w brzusznym obszarze nakrywkowym wystają do przyśrodkowego jądra półleżącego. Brain Res. 300, 157–160. doi: 10.1016/0006-8993(84)91351-9
Kawaguchi, Y., Wilson, CJ i Emson, PC (1990). Podtypy projekcji komórek macierzy neostriatalnej szczura ujawnione przez wewnątrzkomórkową iniekcję biocytiny. J. Neurosci. 10, 3421-3438.
Kelley, AE i Swanson, CJ (1997). Karmienie indukowane przez blokadę receptorów AMPA i kainianowych w prążkowiu brzusznym: badanie mapowania mikroinfuzji. Behav. Brain Res. 89, 107–113. doi: 10.1016/S0166-4328(97)00054-5
Kemp, JM i Powell, TP (1971). Synaptyczna organizacja jądra ogoniastego. Philos. Trans. R. Soc. Lond. B Biol. Sci. 262, 403 – 412. doi: 10.1098 / rstb.1971.0103
Kenny, PJ (2011). Mechanizmy nagród w otyłości: nowe spostrzeżenia i przyszłe kierunki. Neuron 69, 664 – 679. doi: 10.1016 / j.neuron.2011.02.016
Kessler, RC, Berglund, PA, Chiu, WT, Deitz, AC, Hudson, JI, Shahly, V., et al. (2013). Częstość występowania i korelacje zaburzeń związanych z objadaniem się w Światowej Organizacji Zdrowia Światowej Organizacji Zdrowia Psychicznego. Biol. Psychiatria 73, 904-914. doi: 10.1016 / j.biopsych.2012.11.020
Klenowski, PM, Fogarty, MJ, Belmer, A., Noakes, PG, Bellingham, MC i Bartlett, SE (2015). Charakterystyka strukturalna i funkcjonalna altan dendrytycznych i synaptycznych wejść GABAergicznych na interneuronach i komórkach głównych w bazolateralnym ciele migdałowatym szczura. J. Neurophysiol. 114, 942 – 957. doi: 10.1152 / jn.00824.2014
Lenoir, M., Serre, F., Cantin, L. i Ahmed, SH (2007). Intensywna słodycz przewyższa nagrodę kokainową. PLoS ONE 2: e698. doi: 10.1371 / journal.pone.0000698
Li, Y., Kolb, B. i Robinson, TE (2003). Lokalizacja utrzymujących się indukowanych amfetaminą zmian w gęstości kolców dendrytycznych na średnich neuronach kolczastych w jądrze półleżącym i ogoniastym. Neuropsychopharmacology 28, 1082-1085. doi: 10.1038 / sj.npp.1300115
Lindvall, O. i Björklund, A. (1978). Anatomia układów neuronów dopaminergicznych w mózgu szczura. Adv. Biochem. Psychopharmacol. 19, 1-23.
Lutter, M. i Nestler, EJ (2009). Sygnały homeostatyczne i hedoniczne oddziałują w regulacji spożycia żywności. J. Nutr. 139, 629 – 632. doi: 10.3945 / jn.108.097618
Maldonado-Irizarry, CS, Swanson, CJ i Kelley, AE (1995). Receptory glutaminianowe w powłoce jądra półleżącego kontrolują zachowanie żywieniowe poprzez boczne podwzgórze. J. Neurosci. 15, 6779-6788.
Malik, VS, Popkin, BM, Bray, GA, Després, JP i Hu, FB (2010). Napoje słodzone cukrem, otyłość, cukrzyca typu 2 i ryzyko chorób sercowo-naczyniowych. Obieg 121, 1356 – 1364. doi: 10.1161 / CIRCULATIONAHA.109.876185
Ng, SW, Slining, MM i Popkin, BM (2012). Zastosowanie kalorycznych i niecalorycznych substancji słodzących w amerykańskiej żywności pakowanej 2005-2009. J. Acad. Nutr. Dieta. 112, 1828 – 1834 e1821 – e1826. doi: 10.1016 / j.jand.2012.07.009
Nielsen, SJ, Siega-Riz, AM i Popkin, BM (2002). Trendy w poborze energii w USA między 1977 a 1996: podobne przesunięcia widoczne w różnych grupach wiekowych. Obes. Res. 10, 370 – 378. doi: 10.1038 / oby.2002.51
Paxinos, G. i Watson, C. (2007). Mózg szczura w współrzędnych stereotaktycznych. Amsterdam; Boston, MA: Academic Press / Elsevier.
Popkin, BM (2010). Co jest złego w amerykańskim podejściu do otyłości? Wirtualny mentor 12, 316–320. doi: 10.1001/virtualmentor.2010.12.4.pfor2-1004
Rada, P., Avena, NM, i Hoebel, BG (2005). Codzienne objadanie się cukrem wielokrotnie uwalnia dopaminę w skorupie półleżącej. Neuroscience 134, 737 – 744. doi: 10.1016 / j.neuroscience.2005.04.043
Rafols, JA, Cheng, HW i McNeill, TH (1989). Badanie Golgiego prążkowia myszy: związane z wiekiem zmiany dendrytyczne w różnych populacjach neuronów. J. Comp. Neurol. 279, 212 – 227. doi: 10.1002 / cne.902790205
Ranjan, A. i Mallick, BN (2010). Zmodyfikowana metoda spójnego i niezawodnego barwienia Golgiego-Coxa w znacznie krótszym czasie. Z przodu. Neurol. 1: 157. doi: 10.3389 / fneur.2010.00157
Reinehr, T. (2013). Cukrzyca typu 2 u dzieci i młodzieży. World J. Diabetes 4, 270 – 281. doi: 10.4239 / wjd.v4.i6.270
Reissner, KJ i Kalivas, PW (2010). Wykorzystanie homeostazy glutaminianowej jako celu leczenia zaburzeń uzależniających. Behav. Pharmacol. 21, 514–522. doi: 10.1097/FBP.0b013e32833d41b2
Reynolds, SM i Berridge, KC (2003). Zespoły motywacyjne glutaminianowe w jądrze półleżącym: gradienty skorupy strostrocaudalnej strachu i karmienia. Eur. J. Neurosci. 17, 2187-2200. doi: 10.1046 / j.1460-9568.2003.02642.x
Richard, JM i Berridge, KC (2011). Nucleus accumbens interakcja dopaminy / glutaminianu przełącza tryby generowania pożądania w stosunku do strachu: D (1) sam dla apetycznego jedzenia, ale D (1) i D (2) razem ze strachu. J. Neurosci. 31, 12866 – 12879. doi: 10.1523 / JNEUROSCI.1339-11.2011
Robinson, TE i Kolb, B. (1999). Zmiany w morfologii dendrytów i kolców dendrytycznych w jądrze półleżącym i korze przedczołowej po wielokrotnym leczeniu amfetaminą lub kokainą. Eur. J. Neurosci. 11, 1598-1604. doi: 10.1046 / j.1460-9568.1999.00576.x
Robinson, TE i Kolb, B. (2004). Plastyczność strukturalna związana z ekspozycją na narkotyki. Neuropharmacology 47 (Suppl. 1), 33 – 46. doi: 10.1016 / j.neuropharm.2004.06.025
Russo, SJ, Dietz, DM, Dumitriu, D., Morrison, JH, Malenka, RC i Nestler, EJ (2010). Uzależniona synapsa: mechanizmy synaptycznej i strukturalnej plastyczności jądra półleżącego. Trendy Neurosci. 33, 267 – 276. doi: 10.1016 / j.tins.2010.02.002
Rutledge, LT, Duncan, J. i Beatty, N. (1969). Badanie obustronnych aksonów komórek piramidalnych w nienaruszonej i częściowo izolowanej dorosłej korze mózgowej. Brain Res. 16, 15–22. doi: 10.1016/0006-8993(69)90082-1
Saper, CB, Chou, TC i Elmquist, JK (2002). Potrzeba żywienia: homeostatyczna i hedoniczna kontrola jedzenia. Neuron 36, 199–211. doi: 10.1016/S0896-6273(02)00969-8
Scheggi, S., Secci, ME, Marchese, G., De Montis, MG i Gambarana, C. (2013). Wpływ smakowitości na motywację do działania na kaloryczne i niekaloryczne jedzenie u szczurów pozbawionych żywności i pozbawionych żywności. Neuroscience 236, 320 – 331. doi: 10.1016 / j.neuroscience.2013.01.027
Sesack, SR i Pickel, VM (1990). W przyśrodkowym jądrze szczura półleżące, końcówki hipokampa i katecholaminergiczne zbiegają się na neuronach kolczastych i są w stosunku do siebie. Brain Res. 527, 266–279. doi: 10.1016/0006-8993(90)91146-8
Sesack, SR i Pickel, VM (1992). Przedczołowe kory eferentne w synapsie szczura na nieznakowanych celach neuronowych terminali katecholaminowych w jądrze półleżącym septi i neuronach dopaminowych w brzusznym obszarze nakrywkowym. J. Comp. Neurol. 320, 145 – 160. doi: 10.1002 / cne.903200202
Shariff, M., Quik, M., Holgate, JY, Morgan, M., Patkar, OL, Tam, V., i in. (w prasie). Neuronalne nikotynowe modulatory receptora acetylocholiny zmniejszają spożycie cukru. PLoS ONE.
Sheehan, DV i Herman, BK (2015). Czynniki psychologiczne i medyczne związane z nieleczonymi zaburzeniami odżywiania się. Sztywny. Care Companion CNS Disord. 17. doi: 10.4088 / PCC.14r01732
Simms, JA, Steensland, P., Medina, B., Abernathy, KE, Chandler, LJ, Wise, R., i in. (2008). Przerywany dostęp do 20% etanolu powoduje wysokie zużycie etanolu u szczurów Long-Evans i Wistar. Alkohol. Clin. Exp. Res. 32, 1816-1823. doi: 10.1111 / j.1530-0277.2008.00753.x
Steensland, P., Simms, JA, Holgate, J., Richards, JK, i Bartlett, SE (2007). Wareniklina, częściowy agonista receptora acetylocholinowego alfa4beta2, selektywnie zmniejsza zużycie i poszukiwanie etanolu. Proc. Natl. Acad Sci. USA 104, 12518 – 12523. doi: 10.1073 / pnas.0705368104
Stuber, GD, Hopf, FW, Hahn, J., Cho, SL, Guillory, A., i Bonci, A. (2008a). Dobrowolne przyjmowanie etanolu zwiększa pobudzającą siłę synaptyczną w brzusznym obszarze nakrywkowym. Alkohol. Clin. Exp. Res. 32, 1714-1720. doi: 10.1111 / j.1530-0277.2008.00749.x
Stuber, GD, Klanker, M., de Ridder, B., Bowers, MS, Joosten, RN, Feenstra, MG, i in. (2008b). Wskazówki przewidujące nagrody zwiększają pobudzającą siłę synaptyczną neuronów dopaminy śródmózgowia. nauka 321, 1690 – 1692. doi: 10.1126 / science.1160873
Swanson, SA, Crow, SJ, Le Grange, D., Swendsen, J., i Merikangas, KR (2011). Występowanie i korelacje zaburzeń odżywiania u młodzieży. Wyniki krajowego uzupełniającego badania uzupełniającego suplementu dla młodzieży. Łuk. Gen. Psychiatry 68, 714 – 723. doi: 10.1001 / archgenpsychiatry.2011.22
Te Morenga, L., Mallard, S., i Mann, J. (2013). Cukry dietetyczne i masa ciała: przegląd systematyczny i metaanaliza randomizowanych badań kontrolowanych i badań kohortowych. BMJ 346: e7492. doi: 10.1136 / bmj.e7492
Totterdell, S. i Smith, AD (1989). Konwergencja wkładu hipokampa i dopaminergicznego do zidentyfikowanych neuronów w jądrze półleżącym szczura. J. Chem. Neuroanat. 2, 285-298.
Tukey, DS, Ferreira, JM, Antoine, SO, D'Amour, JA, Ninan, I., Cabeza de Vaca, S. i wsp. (2013). Spożycie sacharozy wywołuje szybki transport receptorów AMPA. J. Neurosci. 33, 6123 – 6132. doi: 10.1523 / JNEUROSCI.4806-12.2013
Veening, JG, Cornelissen, FM i Lieven, PA (1980). Miejscowa organizacja aferentów do caudatoputamen szczura. Badanie nad peroksydazą chrzanową. Neuroscience 5, 1253–1268. doi: 10.1016/0306-4522(80)90198-0
Ventura, T., Santander, J., Torres, R. i Contreras, AM (2014). Neurobiologiczne podstawy głodu węglowodanów. Odżywianie 30, 252 – 256. doi: 10.1016 / j.nut.2013.06.010
Volkow, ND, Wang, GJ, Fowler, JS, Tomasi, D. i Baler, R. (2012). Nagroda za jedzenie i narkotyki: nakładające się obwody w otyłości i uzależnieniu od ludzi. Curr. Top. Behav. Neurosci. 11, 1–24. doi: 10.1007/7854_2011_169
Wise, RA (1973). Dobrowolne spożycie etanolu u szczurów po ekspozycji na etanol w różnych harmonogramach. Psychofarmakologia 29, 203 – 210. doi: 10.1007 / BF00414034
Wright, CI i Groenewegen, HJ (1995). Wzory zbieżności i segregacji w przyśrodkowym jądrze półleżącym szczura: relacje przedczołowej kory korowej, środkowej linii wzgórzowej i podstawnych aferentnych jąder migdałowatych. J. Comp. Neurol. 361, 383 – 403. doi: 10.1002 / cne.903610304
Słowa kluczowe: konsumpcja typu binge, długoterminowe, średni kolczasty neuron, jądro półleżące, sacharoza
Cytowanie: PM Klenowski, Shariff MR, Belmer A, Fogarty MJ, Mu EWH, Bellingham MC i Bartlett SE (2016) Przedłużone spożycie sacharozy w obłędny sposób, zmienia morfologię średnich neuronów kolczastych w skorupie jądra półleżącego. Z przodu. Behav. Neurosci. 10: 54. doi: 10.3389 / fnbeh.2016.00054
Otrzymano: 03 December 2015; Akceptowane: 07 March 2016;
Opublikowane: 23 March 2016.
Edytowany przez:
Djoher Nora Abrous, Institut des Neurosciences de Bordeaux, Francja
Zrecenzowany przez:
Serge H. Ahmed, Centre National de la Recherche Scientifique, Francja
Stéphanie Caille, Centre National de la Recherche Scientifique, Francja
Copyright © 2016 Klenowski, Shariff, Belmer, Fogarty, Mu, Bellingham i Bartlett. To jest artykuł o otwartym dostępie dystrybuowany na warunkach Licencja Creative Commons Uznanie autorstwa (CC BY). Używanie, rozpowszechnianie lub powielanie na innych forach jest dozwolone, pod warunkiem, że oryginalni autorzy lub licencjodawcy są uznani i że cytowana jest oryginalna publikacja w tym czasopiśmie, zgodnie z przyjętą praktyką akademicką. Zabronione jest użytkowanie, dystrybucja lub reprodukcja, która nie jest zgodna z niniejszymi warunkami.
* Korespondencja: Selena E. Bartlett, [email chroniony]
