Neurosci Biobehav Rev. 2011 Jan; 35 (3): 939-55. doi: 10.1016 / j.neubiorev.2010.10.014. Epub 2010 Nov 2.
- 1Bernard B. Brodie Departamento de Neurociência, Universidade de Cagliari, Cittadella Universitaria, 09042 Monserrato, CA, Itália. [email protected]
ABSTRATO
A ocitocina é um potente indutor da ereção do pênis quando injetada no sistema nervoso central. Em ratos machos, a área cerebral mais sensível para o efeito pró-erétil da ocitocina é o núcleo paraventricular do hipotálamo. Este núcleo e as regiões circundantes contêm os corpos celulares de todos os neurônios oxitocinérgicos que se projetam para áreas do cérebro extra-hipotalâmicas e para a medula espinhal. Esta revisão mostra que a ocitocina induz a ereção peniana também quando injetada em algumas dessas áreas (por exemplo, área tegmentar ventral, subículo do hipocampo ventral, núcleo cortical posteromedial da amígdala e medula espinal toraco-lombar). Estudos de microinjeção combinados com microdiálise intracerebral e estudos de dupla imunofluorescência sugerem que a ocitocina nessas áreas ativa direta ou indiretamente (principalmente através do ácido glutâmico) neurônios dopaminérgicos mesolímbicos. A dopamina liberada no núcleo accumbens, por sua vez, ativa as vias neurais que levam à ativação de neurônios dopaminérgicos no núcleo paraventricular. Isso ativa não apenas os neurônios oxitocinérgicos que se projetam para a medula espinhal e media a ereção peniana, mas também aqueles que se projetam para as áreas extra-hipotalâmicas acima, modulando direta ou indiretamente (através do ácido glutâmico) a atividade dos neurônios dopaminérgicos mesolímbicos controlando a motivação e a recompensa. Juntas, essas vias neurais podem constituir um circuito hipotético complexo, que desempenha um papel não apenas na fase consumatória da atividade sexual (função erétil e cópula), mas também nos aspectos motivacionais e recompensadores da fase antecipatória do comportamento sexual.
1.Introduction
A ereção peniana é uma resposta sexual masculina que desempenha um papel fundamental na reprodução de mamíferos, incluindo o homem, e que também pode ser observada em contextos diferentes daqueles estritamente relacionados à reprodução. Dependendo do contexto em que a ereção peniana ocorre, diferentes mecanismos neurais e / ou humorais centrais e periféricos participam de sua regulação (veja Meisel e Sachs, 1994; Argiolas e Melis, 1995, 2004, 2005; Sachs, 2000, 2007; McKenna, 2000; Giuliano e Rampin, 2000, 2004, Andersson, 2001, Melis e Argiolas, 1995a, 2003, Hull e outros, 2002). Entre os neurotransmissores centrais e neuropeptídeos que controlam a ereção peniana, os mais conhecidos são dopamina, serotonina, aminoácidos excitatórios, óxido nítrico, adrenocorticotropina, ocitocina e peptídeos opióides. Eles podem facilitar ou inibir a ereção peniana, agindo em várias áreas do cérebro, isto é, a área pré-óptica medial, o núcleo paraventricular do hipotálamo, a área tegmentar ventral, o hipocampo, a amígdala, o núcleo do leito da estria terminal, o núcleo accumbens, a medula oblonga e a medula espinhal (Tabela 1) (veja Meisel e Sachs, 1994; Witt e Insel, 1994; Stancampiano e outros, 1994; Argiolas e Melis, 1995, 2005; Argiolas, 1999; Bancila e outros, 2002; Giuliano e Rampin, 2000, McKenna, 2000; Andersson, 2001; Hull e outros, 2002; Coolen e outros, 2004).
A ocitocina, o peptídeo neuro-hipofisário bem conhecido por seu papel hormonal na lactação e no parto, está presente em mulheres e homens, não apenas em neurônios com corpos celulares localizados nos núcleos paraventriculares e supra-ópticos do hipotálamo que se projetam para a neuro-hipófise, mas também em neurônios do núcleo paraventricular e estruturas adjacentes a áreas cerebrais extra-hipotalâmicas (isto é, o septo, a área tegmentar ventral, o hipocampo, a amígdala, a medula oblonga e a medula espinhal). Acredita-se que esses neurônios estejam envolvidos em numerosas funções centrais, como memória, aprendizagem, comportamentos afiliativos e sócio-sexuais, incluindo ereção peniana e comportamento copulatório (ver Buijs, 1978; Sofroniew, 1983; Argiolas e Gessa, 1991; Pedersen e cols. 1992; Carter, 1992; Wagner e Clemens, 1993; Ivell e Russel, 1995; Carter e outros, 1997; Tang e outros, 1998; Veronneau-Longueville e outros, 1999). De fato, a oxitocina facilita a função erétil e o comportamento sexual masculino em camundongos, ratos, coelhos e macacos (ver Argiolas e Gessa, 1991; Carter, 1992; Pedersen e outros, 1992; Argiolas e Melis, 1995, 2004; Argiolas, 1999). Isso pode ocorrer também em humanos, uma vez que a ocitocina plasmática é aumentada por estímulos sexuais, especialmente à ejaculação (Carmichael et al., 1987; Murphy et al., 1987) e pela manipulação da mama e da genitália, que geralmente ocorrem durante a atividade sexual. relação sexual (Tindall, 1974).
Um efeito facilitador da oxitocina no comportamento sexual masculino foi demonstrado pela primeira vez pela capacidade da ocitocina intravenosa de diminuir a latência para a primeira ejaculação e para retardar a exaustão sexual de coelhos machos emparelhados com fêmeas receptivas (Melin e Kihlstrom, 1963). No entanto, os efeitos sexuais da ocitocina foram definitivamente reconhecidos apenas nos anos oitenta. A ocitocina dada centralmente em quantidades de nanogramas foi então capaz de induzir a ereção peniana (Argiolas et al., 1985, 1986) e melhorar o comportamento copulatório (Arletti et al., 1985) em ratos machos e aumentar a lordose em ratas (Arletti e Bertolini, 1985; Caldwell e outros, 1986), aparentemente agindo em receptores oxitocinérgicos do tipo uterino (ver Argiolas e Melis, 1995, 2004; Argiolas, 1999; Melis e Argiolas, 2003; e referências nele). A oxitocina melhora o comportamento sexual não apenas em ratos machos sexualmente potentes (Arletti et al., 1985) mas também em ratos machos idosos (Arletti et al., 1990) e em macacos-esquilo dominantes mas não subordinados (Winslow e Insel , 1991).
O efeito pró-erétil da ocitocina é dependente de testosterona, uma vez que é abolido por hipofisectomia e castração, e restaurado pela suplementação com testosterona ou seus metabólitos, estradiol e 5-di-hidro-testosterona administrados juntos. (Melis et al., 1994a). A área cerebral mais sensível para a indução da ereção peniana pela ocitocina é o núcleo paraventricular do hipotálamo (Melis et al., 1986), do qual todas as projeções oxitocinérgicas extra-hipotalâmicas se originam (ver acima). Aqui, descobriu-se que a ocitocina é capaz de induzir a ereção peniana (e bocejar) quando injetada em doses tão baixas quanto 3 pmol (veja a Seção 2.1 abaixo). A ereção peniana induzida pela ocitocina também quando injetada bilateralmente no campo CA1 do hipocampo, mas não no subículo dorsal (veja a Seção 2.3 abaixo), o septo lateral, o núcleo caudado, a área pré-óptica medial, o núcleo ventromedial do hipotálamo e núcleo supra-óptico (Melis et al., 1986). Quanto ao mecanismo pelo qual a oxitocina atua no núcleo paraventricular para induzir essa resposta sexual, numerosos estudos sugerem que a ocitocina ativa seus próprios neurônios. Em consonância com essa hipótese, a interação sexual aumenta o FOS, o produto gênico do gene precoce imediato c-fos em neurônios paraventriculares oxitocinérgicos projetados para a medula espinhal, que estão envolvidos no controle da ereção peniana (ver Witt e Insel, 1994 e referências a impotência sexual (por exemplo, a incapacidade de um rato macho adulto de copular com uma fêmea receptiva estrogênio-progesterona ovariectomizada) foi associada no rato masculino com baixos níveis de mRNA de ocitocina no núcleo paraventricular do hipotálamo (Arletti et al. al., 1997).
Se a ocitocina influencia a fase de antecipação ou a fase consumatória do comportamento sexual não está claro no momento. Como a ocitocina induz a ereção peniana e o principal efeito da ocitocina no comportamento copulatório é uma diminuição no intervalo pós-ejaculatório em ratos machos (Arletti et al., 1985), é razoável supor que o peptídeo melhora o desempenho sexual. No entanto, como a ocitocina também aumenta a interação sócio-sexual (ver Pedersen et al., 1992; Carter et al., 1997; Ivell e Russel, 1995), e antagonistas do receptor de ocitocina impedem a ereção sem contato (Melis et al., 1999a), são considerados como um índice de excitação sexual (ver Sachs, 1997, 2000, 2007; Melis et al., 1998, 1999b e suas referências), um possível papel da ocitocina na excitação sexual e na motivação sexual não pode ser descartado.
Esta revisão resume os resultados publicados e não publicados de estudos recentes, que mostram que a ocitocina induz a ereção peniana não só quando injetada no núcleo paraventricular do hipotálamo, mas também em outras áreas cerebrais extra-hipotalâmicas, como a área tegmentar ventral (Melis e outros, 2007, 2009a; Succu e outros, 2008), to subículo do ventre do hipocampo e o núcleo posterior da amígdala (Melis et al., 2009b, 2010), que são constituintes importantes do sistema límbico e são pensados para desempenhar um papel fundamental nos processos de motivação e recompensa. Esses estudos revelam que a oxitocina participa de circuitos neurais, que incluem outros neurotransmissores, como dopamina e ácido glutâmico, e outras áreas do cérebro que não o núcleo paraventricular, por exemplo, a área tegmental ventral, o núcleo accumbens, o hipocampo e áreas ainda a serem identificado. Esses circuitos provavelmente mediarão uma interação entre o sistema dopaminérgico mesolímbico e o incerto-hipotalâmico e desempenharão um papel não apenas na fase consumatória do comportamento sexual masculino (por exemplo, ereção e cópula peniana), mas também na motivação sexual e sexual. excitação, portanto, fornecendo um substrato neural para explicar as propriedades motivacionais e recompensadoras da atividade sexual.
2. A ocitocina influencia a ereção peniana, atuando em diferentes áreas do cérebro
2.1. O núcleo paraventricular do hipotálamo
Como lembrado acima, o núcleo paraventricular do hipotálamo foi logo identificado como a área do cérebro mais sensível para o efeito pró-erétil da ocitocina. Quando injetada unilateralmente neste núcleo, a ocitocina foi encontrada ativa em doses tão baixas quanto 3 ng (3 pmol) (Melis et al., 1986). Estudos de relação estrutura-atividade revelaram que a ereção peniana induzida pela oxitocina era mediada por receptores de oxitocina de tipo uterino, acoplados a um influxo de Ca2 + nos corpos celulares de neurônios oxitocinérgicos projetando-se para áreas do cérebro extra-hipotalâmicas e à ativação da óxido nítrico sintase. O óxido nítrico, por sua vez, agindo como um mensageiro intracelular com um mecanismo ainda desconhecido (não envolvendo a guanilato ciclase) leva à ativação de neurônios ocitocinérgicos que se projetam para a medula espinhal e áreas extra-hipotalâmicas do cérebro, induzindo a ereção peniana (Fig. 1) abaixo e Argiolas e Melis, 1995, 2004, 2005 e referências nele). A capacidade da oxitocina para ativar seus próprios neurônios
Fig. 1. (FALTA) Representação esquemática dos neurônios oxitocinérgicos, que se originam no núcleo paraventricular do hipotálamo e se projetam para áreas do cérebro extra-hipotalâmicas, como a medula espinhal, a VTA, o hipocampo, a amígdala, etc. A ativação desses neurônios por dopamina, aminoácidos excitatórios, a própria oxitocina, peptídeos análogos da hexarelina e peptídeos derivados do VGF levam à ereção peniana, que pode ser reduzida e / ou abolida pela estimulação dos receptores GABAérgicos, opioides e canabinóides CB1. A ativação de neurônios oxitocinérgicos é secundária à ativação da óxido nítrico sintase presente nesses neurônios. De fato, o óxido nítrico endógeno formado pela estimulação de dopamina, aminoácidos excitatórios ou receptores de oxitocina ou óxido nítrico exógeno, como o derivado de doadores de óxido nítrico administrados diretamente no núcleo paraventricular, ativa os neurônios oxitocinérgicos por um mecanismo ainda não identificado, aparentemente não relacionado à estimulação da guanilato ciclase. Isso causa, por sua vez, a liberação de ocitocina na medula espinhal e em áreas cerebrais extra-hipotalâmicas. Alguns detalhes sobre os mecanismos pelos quais a ocitocina induz a ereção peniana quando liberada nessas áreas, por exemplo, a ATV, o subículo e a amigdala, são descritos nas respectivas seções da área do cérebro. Aqui, a ocitocina atua em seus próprios receptores e aumenta a produção de NO, o que leva à ereção peniana como encontrada no PVN. No entanto, em variância com o PVN, no VTA NO caudal ativa guanilato ciclase. Isso causa um aumento na concentração de cGMP levando à ativação dos neurônios dopaminérgicos mesolímbicos e à ereção peniana. No VS NO ativa neurônios glutamatérgicos projetando-se para áreas extra-hipocampais, incluindo o VTA. O ácido glutâmico na VTA ativa, por sua vez, os neurônios dopaminérgicos mesolímbicos encontrados com a ocitocina. Mecanismos semelhantes aos descritos acima provavelmente também funcionarão quando a ereção peniana ocorre em contextos fisiológicos, como quando ratos machos são colocados na presença de uma fêmea receptiva inacessível (por exemplo, ereções sem contato) ou durante a cópula.
no núcleo paraventricular foi apoiado por estudos mostrando que: (1) receptores de ocitocina estão presentes neste núcleo hipotalâmico (Freund-Mercier e outros, 1987; Freund-Mercier e Stoeckel, 1995); (2) oxitocina facilita sua própria liberação in vitro e in vivo (Freund-Mercier e Richard, 1981, 1984; Moos et al., 1984); e (3) oxitocina excita seus próprios neurônios agindo no núcleo paraventricular (Yamashita et al., 1987). Além disso, as sinapses oxitocinérgicas que colidem com os corpos celulares dos neurônios ocitocinérgicos magnocelulares também foram identificadas nos núcleos paraventricular e supra-óptico do hipotálamo. (Theodosis, 1985). Finalmente, a destruição de neurônios oxitocinérgicos centrais por lesões eletrolíticas ou excitotóxicas químicas do núcleo paraventricular, que depletam completamente o conteúdo de oxitocina através do sistema nervoso central e da medula espinhal, abole não apenas o efeito pró-erétil da ocitocina, mas também prejudica o efeito induzido por drogas. ereção peniana e ereções sem contacto (ver abaixo e Argiolas et al., 1987a, b; Liu et al., 1997 e referências). Resultados semelhantes aos encontrados com lesões do núcleo paraventricular são encontrados com antagonistas potentes e seletivos dos receptores de ocitocina. De fato, esses compostos injetados no núcleo paraventricular em quantidades nanométricas impediram a ereção peniana induzida por oxitocina, enquanto que, quando administrados nos ventrículos laterais, impediam não apenas ereções penianas induzidas pela ocitocina em si, mas também ereção peniana induzida por drogas (veja a Seção 3 abaixo). e Argiolas e Melis, 1995, 2004, 2005 e suas referências) e ereções sem contato (Melis et al., 1999a), além de serem extremamente eficazes em prejudicar o comportamento copulatório de ratos machos sexualmente potentes (Argiolas et al., 1988). Além disso, a interação sexual aumenta o FOS, o produto gênico do gene precoce imediato c-fos nos neurônios oxitocinérgicos paraventriculares que se projetam para a medula espinhal envolvidos no controle da ereção peniana (ver Witt e Insel, 1994 e referências lá contidas). Finalmente, a impotência sexual (por exemplo, a incapacidade de um rato macho adulto para copular com uma fêmea receptiva estrogênio-progesterona)como também foi associado no rato macho com baixos níveis de mRNA de ocitocina e de óxido nítrico sintase no núcleo paraventricular do hipotálamo (Benelli e outros, 1995; Arletti e outros, 1997) (para uma revisão extensiva destes estudos veja Argiolas, 1999; Argiolas e Melis, 2004, 2005).
2.2. A área tegmentar ventral
A área tegmentar ventral foi descoberta apenas recentemente como um sítio cerebral no qual a ocitocina induz a ereção peniana. Esta área contém terminações nervosas oxitocinérgicas originadas no núcleo paraventricular e receptores de ocitocina (Freund-Mercier e col., 1987; Vaccari et al., 1998). Mais precisamente, a ocitocina foi encontrada capaz deindução da ereção peniana quando injetada unilateralmente na região caudal, mas não na região tegmentar ventral rostral de uma maneira dependente da dose (Melis et al., 2007). As doses ativas foram maiores do que as necessárias quando injetadas no núcleo paraventricular e similares àquelas que induzem ereção peniana quando injetadas no subículo do hipocampo ventral ou no núcleo cortical posteromedial da amígdala (veja abaixo). Aparentemente, o efeito pró-erétil é mediado pela ativação de neurônios dopaminérgicos mesolímbicos que se projetam para a casca do nucleus accumbens, que por sua vez ativa vias neurais desconhecidas projetando-se para os neurônios dopaminérgicos incerto-hipotalâmicos que invadem os neurônios oxitocinérgicos paraventriculares mediando a ereção peniana (Melis et al., 2007, 2009a).
Quanto ao mecanismo pelo qual a ocitocina ativa a neurotransmissão dopaminérgica na área tegmentar ventral, os dados disponíveis sugerem que A oxitocina estimula os receptores oxitocinérgicos localizados nos corpos celulares dos neurônios dopaminérgicos mesolímbicos. Isso aumenta o influxo de Ca2 + dentro dos corpos celulares dos neurônios dopaminérgicos, ativando assim a óxido nítrico sintase (Succu et al., 2008). Em desacordo com o núcleo paraventricular (veja a Seção 3 abaixo), o óxido nítrico, por sua vez, ativa a guanilato ciclase, aumentando assim a concentração de GMP cíclico. Em consonância com este mecanismo, quer d (CH2) 5Tyr (Me) 2-Orn8-vasotocina, um potente antagonista da oxitocina, ou S-metil-tio-l-citrulina, um potente inibidor da óxido nítrico-sintase neuronal, injetado na área tegmentar ventral caudal antes da ocitocina, aboliu a ereção peniana e o aumento da concentração de dopamina extracelular na carapaça do nucleus accumbens induzida pela ocitocina. Além disso, GMP 8-bromo-cíclico, um análogo GMP cíclico resistente à fosfodiesterase ativa, induz a ereção peniana quando injetado na área tegmentar ventral caudal e aumenta a concentração de dopamina extracelular na carapaça do nucleus accumbens, como encontrado com ocitocina injetada a área tegmentar ventral caudal (Succu et al., 2008; Melis e outros, 2009a) (veja também Fig. 2).
Sempre alinhado com este mecanismo, o haloperidol, um potente antagonista do receptor de D2 da dopamina, injetado na concha do núcleo accumbens, reduz a ereção peniana induzida pela ocitocina injetada na área tegmentar ventral (Melis et al., 2007). O mecanismo acima também é apoiado por estudos de imunofluorescência dupla, mostrando que as fibras de oxitocina atingem os corpos celulares de neurônios dopaminérgicos na área tegmentar ventral caudal, que foram previamente marcados com o traçador retrógrado Fluorogold injetado na casca do núcleo accumbens ( Melis et al., 2007; Succu et al., 2008). As ativações desses neurônios dopaminérgicos e dos receptores dopaminérgicos no nucleus accumbens levam, por sua vez, à ativação de vias neurais ainda não identificadas, que estimulam os neurônios dopaminérgicos hipotalâmicos a liberar dopamina no núcleo paraventricular, ativando neurônios oxitocinérgicos que se projetam medula espinhal e mediação da ereção peniana (veja acima e Melis e outros, 2007; Succu e outros, 2007, 2008). De fato, a ocitocina injetada na área tegmentar ventral caudal na dose que induziu a ereção peniana, aumentou a concentração extra-celular de dopamina no dialisato obtido não apenas do núcleo accumbens, mas também do núcleo paraventricular (Succu et al., 2007).
2.3. O hipocampo
O campo CA1 do hipocampo foi a outra área do cérebro rica em fibras oxitocinérgicas e receptores identificados pelos estudos anteriores em que a injeção de ocitocina induziu a ereção peniana (ver Bujis, 1978; Sofroniew, 1983). Entretanto, em desacordo com o núcleo paraventricular, aqui a ocitocina foi encontrada capaz de induzir a ereção peniana apenas quando injetada bilateralmente e em doses mais altas do que aquelas encontradas ativas no núcleo paraventricular (Melis et al., 1986; Chen et al., 1992). Injeções de ocitocina no subículo foram encontradas inativas nesses estudos anteriores. No entanto, estudos recentes e mais cuidadosos da microinjeção levam à identificação de região do subiculum ventral em que a injeção de ocitocina foi capaz de induzir a ereção peniana de maneira dose-dependente (Melis et al., 2009b). O efeito pró-erétil da ocitocina injetada nesta área do cérebro foi observado em doses similares àquelas encontradas ativas na área tegmentar ventral após a injeção unilateral (Melis et al., 2007), como encontrado no núcleo paraventricular. Aparentemente, a oxitocina injetada no subículo do ventre induz a ereção peniana pela ativação de receptores oxitocinérgicos em neurônios contendo óxido nítrico sintase, causando um aumento na produção de óxido nítrico. O óxido nítrico, por sua vez, atuando como mensageiro intercelular, ativa a neurotransmissão do ácido glutâmico, levando à ereção peniana, possivelmente através de projeções eferentes neurais (glutamatérgicas) do subículo para áreas do cérebro extra-hipocampais que modulam a atividade de neurônios dopaminérgicos mesolímbicos (por exemplo, a área tegmental ventral, o córtex pré-frontal, o núcleo paraventricular) (veja abaixo e Melis, 2007, 2009b; Succu et al., 2008).
Esse mecanismo de ação é apoiado por experimentos de microdiálise intracerebral, que mostram que a oxitocina injetada no subículo ventral em doses que induzem a ereção peniana, aumenta a produção de óxido nítrico e a concentração de ácido glutâmico extracelular no dialisado do subículo do ventre (Melis et al. , 2010) e da dopamina extracelular no nucleus accumbens (Melis et al., 2007). Estas respostas foram antagonizadas não só pelo antagonista do receptor de ocitocina d (CH2) 5Tyr (Me) 2-Orn8-vasotocina, mas também pelo inibidor neuronal do óxido nítrico sintase S-metil-tio-litritrina e pela hemoglobina captadora de óxido nítrico dada no subículo valvar poucos minutos antes da ocitocina (Melis et al., 2010).
Além disso, de acordo com este mecanismo de ação, a ativação da neurotransmissão glutamatérgica por NMDA injetada no subículo do ventre induz a ereção peniana (Melis et al., 2010). O fenótipo de projeções eferentes do subículo ventral, que causa a ativação de neurônios dopaminérgicos mesolímbicos e o aumento da dopamina extracelular no nucleus accumbens, é desconhecido no momento. No entanto, como a ereção peniana induzida pela ocitocina injetada no subículo valvar ocorre concomitantemente a um aumento do ácido glutâmico extra-celular no dialisato da área tegmentar ventral, mas não do núcleo accumbens, é antagonizado por (+) MK-801, um potente antagonista não competitivo de receptores de aminoácidos excitatórios do subtipo NMDA (Woodruff et al., 1987), injetado na área tegmentar ventral, mas não no nucleus accumbens (veja Fig. 2 e Melis et al., 2009b) , É provável que essas projeções levem à ativação da neurotransmissão glutamatérgica na área tegmentar ventral, que por sua vez ativa os neurônios dopaminérgicos mesolímbicos que se projetam para o núcleo accumbens. Se a concentração aumentada de ácido glutâmico encontrada na área tegmentar ventral após a injeção de ocitocina no subículo do ventre é liberada de neurônios originários do subículo ou em outras áreas do cérebro (por exemplo, o córtex pré-frontal) é desconhecido no momento. Não obstante, isso causa a ativação de neurônios dopaminérgicos mesolímbicos e uma liberação aumentada de dopamina no nucleus accumbens. Aqui, a ativação de receptores de dopamina leva à ativação de neurônios dopaminérgicos incerto-hipotalâmicos, liberando dopamina no núcleo paraventricular, ativando neurônios oxitocinérgicos que se projetam para a medula espinhal e mediam a ereção peniana (ver acima e Melis e outros, 2007, 2009a; Succu e outros, 2008).
2.4. Amígdala
A amígdala é outra área do cérebro rica em fibras e receptores de ocitocina (ver Freund-Mercier et al., 1987; Vaccari et al., 1998; Uhl-Bronner et al., 2005). Acredita-se que a ocitocina esteja envolvida em diferentes funções, desde ansiólise, memória social e cognição, aprendizado socialmente reforçado, empatia emocional, processamento facial emocional e medo em humanos à função erétil e comportamento sexual (ver Kondo et al., 1998; Dominguez et al. 2001; Ebner et al., 2005; Huber e outros, 2005; Domes e outros, 2007; Petrovic e outros, 2008; Lee e outros 2009; Donaldson e Young, 2009; Hurlemann e outros. , 2010). No entanto, a capacidade de a oxitocina induzir a ereção peniana em ratos machos quando induzida no núcleo cortical posteromedial da amígdala foi descoberta apenas recentemente (Melis et al., 2009b). Esta resposta ocorreu concomitantemente com um aumento da concentração de dopamina extra-celular no dialisato obtido a partir do invólucro do nucleus accumbens, como encontrado após a injeção de ocitocina no subículo do ventre (Melis et al., 2009b). O mecanismo pelo qual a oxitocina injetada no núcleo cortical posteromedial da amígdala induz a ereção peniana é desconhecido no momento. Os dados disponíveis mostram que tanto a ereção peniana quanto o aumento da concentração de dopamina extra-celular no dialisato obtido do nucleus accumbens são mediados pela ativação de receptores oxitocinérgicos, já que ambas as respostas foram abolidas pelo antagonista do receptor de ocitocina d (CH2) 5Tyr ( Me) 2-Orn8-vasotocina injetada no núcleo da amígdala poucos minutos antes da ocitocina (Melis et al., 2009b).
Seja qual for o mecanismo que a ocitocina ative no núcleo cortical posteromedial da amígdala, a resposta sexual induzida pelo peptídeo é abolida pelo bloqueio de todos os receptores dopaminérgicos com cisflupentixol injetados no invólucro do nucleus accumbens e pelo bloqueio dos receptores NMDA com ( +) MK-801 injetado na área tegmentar ventral, mas não no núcleo accumbens, como encontrado para a ereção peniana induzida pela oxitocina injetada no subículo do ventre (Melis et al., 2009b). Isto sugere que a oxitocina injetada no núcleo póstero-medial da amígdala ativa a neurotransmissão do ácido glutâmico na área tegmentar ventral. Isto provoca, por sua vez, a ativação dos neurônios dopaminérgicos mesolímbicos, levando à ereção peniana. Em vista de estudos mostrando vias neurais interconectando esse núcleo da amígdala com o subículo do ventre (Canteras et al., 1995; French e Totterdell, 2003), esses achados levantam a possibilidade de existir uma interação entre essas duas áreas do cérebro, embora vias da amígdala, tanto para o nucleus accumbens como para a área tegmentar ventral, foram descritas (Kelley e Domesick, 1982; Witter, 2006).
2.5. A medula espinhal
A medula espinhal é outra área do sistema nervoso central que contém fibras e receptores oxitocinérgicos (Freund-Mercier e outros, 1987; Uhl-Bronner e outros, 2005), nos quais a ocitocina induz a ereção peniana (Tang et al., 1998 Veronneau-Longueville e outros, 1999; Giuliano e Rampin, 2000; Giuliano e outros, 2001). Como lembrado acima, essas fibras oxitocinérgicas originam-se no núcleo paraventricular do hipotálamo e contribuem para as vias descendentes que controlam os neurônios autonômicos da medula espinhal, mediando a ereção peniana. De fato, essas fibras fazem contatos sinápticos nas colunas de células simpáticas e parassimpáticas pré-ganglionares do corno dorsal no trato tóraco-lombar e lombo-sacral com neurônios espinhais inervando corpos cavernosos penianos (Marson e McKenna, 1996; Giuliano e Rampin, 2000; Giuliano et al. , 2001). Esses contatos sinápticos foram demonstrados pela marcação de neurônios espinhais originados no pênis e atingindo a medula espinhal com traçadores retrógrados específicos injetados em corpos cavernosos, combinados com estudos de dupla microscopia de imunofluorescência e laser confocal (Tang et al., 1998; Veronneau-Longueville et al., 1999). Em consonância com estes estudos, em ratos machos anestesiados, a injeção intratecal de doses cumulativas de ocitocina no nível lombo-sacral, mas não no nível tóraco-lombar, provocou aumento da pressão intracavernosa de maneira dose-dependente. Estes efeitos foram abolidos pelo bloqueio do receptor oxitocinérgico com d (CH2) 5Tyr (Me) 2-Orn8-vasotocina e por secção dos nervos pélvicos (Giuliano e Rampin, 2000; Giuliano et al., 2001). Esses resultados demonstram que a ocitocina, que atua na medula espinhal lumbo-sacra, aumenta a pressão intracavernosa e sugere que a ocitocina, liberada durante a ativação fisiológica do núcleo paraventricular, é um potente ativador de neurônios pró-eréteis da coluna que se projetam para os corpos cavernosos. Curiosamente, esses neurônios espinhais pró-eréteis, nos quais a oxitocina atua para exercer seu efeito pró-erétil, também recebem contatos sinápticos de neurônios serotoninérgicos originários do núcleo paragigantocelular da formação reticular da medula oblonga (Marson e McKenna, 1992; Tang et al. ., 1998). A destruição desses neurônios serotoninérgicos facilita a ejaculação e os reflexos penianos em ratos machos (Marson e McKenna, 1992; Yells et al., 1992). Como os fármacos que estimulam os receptores 5HT2C facilitam a ereção peniana quando administrados intracerebroventricularmente, mas não no núcleo paraventricular, e os fármacos que bloqueiam os receptores 5HT2C reduzem também a ereção peniana induzida pelo agonista dopaminérgico e oxitocina, enquanto os antagonistas dopaminérgicos não reduzem a ereção peniana induzida pelo agonista 5HT2C ver Stancampiano et al., 1994 e suas referências), também foi sugerido que a ocitocina facilita a ação dos receptores 5HT2C pró-eréteis ao nível da medula espinhal lombo-sacral (Stancampiano et al., 1994). Alternativamente, a ocitocina pode influenciar a atividade dos neurônios serotoninérgicos descendentes da coluna, atuando diretamente no núcleo paragigantocelular, onde esses neurônios se originam (ver Stancampiano et al., 1994). Em consonância com essa possibilidade, foi descrita uma via oxitocinérgica originada no núcleo paraventricular e que atinge o núcleo paragigantocelular da formação reticular na medula oblonga (Bancila et al., 2002), e a ativação dos receptores 5HT2C localizados na medula espinhal, a jusante dos receptores de dopamina e oxitocina, é considerada um mecanismo comum do efeito pró-erétil desses compostos e até mesmo dos peptídeos ACTH-MSH (Kimura et al., 2008).
3. Interações entre ocitocina, dopamina e ácido glutâmico no sistema nervoso central e ereção peniana
Como lembrado na Seção 1, todos os neurônios oxitocinérgicos presentes no sistema nervoso central se originam no núcleo paraventricular e nas estruturas adjacentes. A atividade desses neurônios está sob o controle de diferentes neurotransmissores e / ou neuropeptídeos. Entre os mais estudados no nível paraventricular estão dopamina, ácido glutâmico, ácido gama-aminobutírico (GABA), óxido nítrico, endocanabinóides, peptídeos opióides, peptídeos liberadores do hormônio do crescimento, peptídeos relacionados ao VGF e a própria ocitocina. Dopamina, ácido glutâmico, peptídeos liberadores de hormônio de crescimento, peptídeos derivados de VGF e oxitocina são estimulatórios, por exemplo, esses compostos e seus agonistas facilitam a ereção peniana quando injetados no núcleo paraventricular, enquanto GABA, peptídeos opióides e endocanabinóides são inibitórios, por exemplo, esses compostos ou seus agonistas inibem a ereção peniana (veja Meisel e Sachs, 1994; Witt e Insel, 1994; Argiolas e Melis, 1995, 2004, 2005; Giuliano e Rampin, 2000, 2004; McKenna, 2000; Andersson, 2001; Hull e outros, 2002).
Várias linhas de evidências experimentais sugerem que esses neurônios oxitocinérgicos e os neurotransmissores e neuropeptídeos acima estão envolvidos no controle da função erétil e do comportamento sexual em diferentes contextos fisiológicos. Além disso, a ocitocina liberada em áreas cerebrais extra-hipotalâmicas, como a área tegmental ventral, o hipocampo e suas regiões, a amígdala e a medula espinhal pode influenciar a atividade desses neurônios nos quais as sinapses oxitocinérgicas atingem. No momento, os únicos neurônios importantes para a ereção peniana, nos quais as sinapses oxitocinérgicas se chocam, identificados com certeza, são os corpos celulares de neurônios dopaminérgicos mesolímbicos da área tegmentar ventral caudal projetando-se para a casca do núcleo accumbens (Melis et al., 2007 ; Succu et al., 2008) e os neurônios espinhais pró-eréteis que se projetam do trato lombo-sacral aos corpos cavernosos (ver Giuliano e Rampin, 2000; Giuliano e outros, 2001) (ver também Seções 2.2 e 2.5) . De fato, embora as sinapses e os receptores oxitocinérgicos também tenham sido identificados no subículo, na amígdala e na medula espinhal, áreas que são importantes para a ereção peniana (veja acima), nessas áreas o tipo de neurotransmissor presente nos neurônios em quais terminações nervosas oxitocinérgicas colidem, ainda são desconhecidas.
Esta seção da revisão resume brevemente a literatura recente sobre os mecanismos subjacentes ao efeito pró-erétil da ocitocina injetada na área tegmentar ventral caudal, no subículo do hipocampo ventral e na medula espinhal. Particular atenção é dada à interação do peptídeo com dopamina e ácido glutâmico nessas áreas e ao papel que essa interação pode desempenhar no controle central da função erétil. Um breve resumo dos efeitos da dopamina e do ácido glutâmico sobre os neurônios oxitocinérgicos no núcleo paraventricular, que também desempenham um papel fundamental na função erétil, é fornecido em primeiro lugar, a fim de conscientizar o leitor sobre o estado inicial da pesquisa neste campo, como esses estudos já foram revisados extensivamente (ver Argiolas e Melis, 1995, 2004, 2005; Melis e Argiolas, 2003). Também neste caso, atenção particular é dada aos resultados mais recentes, que sugerem um papel importante tanto para uma ligação dopamina-ocitocina quanto para uma ligação ácido glutâmico-oxitocina não apenas no desempenho sexual (ereção e cópula peniana), mas também na sexualidade. excitação e motivação sexual.
3.1. Interação dopamina-ocitocina no núcleo paraventricular
As capacidade dos agonistas dopaminérgicos de induzir a ereção peniana pela ativação de neurônios ocitocinérgicos centrais foi sugerido imediatamente após a descoberta de que a apomorfina induz a ereção peniana quando injetada no núcleo paraventricular (Melis et al., 1987) quando o antagonista do receptor de ocitocina d (CH2) 5Tyr (Me) 2-Orn8-vasotocina injetado intracerebroventricularmente (icv) capaz de reduzir quase completamente a ereção peniana induzida não só pela ocitocina dada icv, mas também pela apomorfina, administrada por via subcutânea (Argiolas et al., 1987b). Estes resultados foram seguidos pelos de outros estudos que mostraram resultados semelhantes quando d (CH2) 5Tyr (Me) 2-Orn8-vasotocina foi administrado icv e a apomorfina foi administrada diretamente no núcleo paraventricular (Melis et al., 1989b), lsugerindo que os agonistas dopaminérgicos induziriam a ereção peniana pela ativação de neurônios oxitocinérgicos paraventriculares projetando-se para áreas do cérebro extra-hipotalâmicas e, em particular, para a medula espinhal (veja Argiolas e Melis, 1995, 2004, 2005). Em consonância com essa hipótese, em ratos anestesiados, o bloqueio dos receptores oxitocinérgicos lumbo-sacrais por um antagonista não-peptídico do receptor de ocitocina foi recentemente encontrado capaz de abolir os aumentos induzidos pela apomorfina na pressão intracavernosa induzida pelo apomorfina agonista da dopamina, fornecendo evidência de via oxitocinérgica paraventriculo-espinhal envolvida na ereção peniana (Baskerville et al., 2009).
Estudos voltados para o A identificação do receptor dopaminérgico responsável pela indução da ereção peniana revelou que também no núcleo paraventricular os agonistas do receptor de dopamina induzem a ereção peniana atuando sobre os receptores dopaminérgicos da família D2., como encontrado com os agonistas do receptor de dopamina administrados sistemicamente (ver Melis et al., 1987; Eaton et al., 1991; Melis e Argiolas, 1995a). Assim, a apomorfina, um potente agonista do receptor D1 / D2, e quinpirole, um potente agonista seletivo dos receptores D2, mas não o SKF 38393, um agonista seletivo do receptor D1, foram encontrados capazes de induzir a ereção peniana de forma dose-dependente e a resposta sexual induzida por estes agonistas do receptor D2 foi abolida pelos antagonistas do receptor D2, tais como o haloperidol e a l-sulpirida, mas não pelo SCH 23390, um antagonista selectivo do receptor D1 (Melis et al., 1987). A capacidade da apomorfina para induzir a ereção peniana quando injetada no núcleo paraventricular também foi confirmada por estudos de telemetria mostrando que o agonista dopaminérgico administrado no núcleo paraventricular é capaz de aumentar a pressão intracavernosa em ratos machos acordados sem modificar a pressão arterial sistêmica (Chen et al. , 1999; Giuliano e Allard, 2001), como encontrado após injeção sistêmica (Bernabè et al., 1999). Esses estudos também confirmou um papel principal dos receptores D2, uma vez que os agonistas do receptor D1 eram normalmente incapazes de aumentar a pressão intracavernosa quando injetados no núcleo paraventricular (Chen et al., 1999).
Várias linhas de evidências experimentais estavam disponíveis, sugerindo que os receptores D2 paraventriculares, cuja estimulação induz a ereção peniana, estão localizados nos corpos celulares dos neurônios oxitocinérgicos. Primeiro, o núcleo paraventricular contém terminais nervosos dopaminérgicos que pertencem aos chamados neurônios dopaminérgicos incertohipotalâmicos. Os corpos celulares desses neurônios estão situados no grupo A13 e A14 de Dahlstrom e Fuxe (1964), arborizam extensivamente e inervam várias estruturas hipotalâmicas, incluindo neurônios ocitocinérgicos paraventriculares projetando-se para a neuro-hipófise e / ou para áreas cerebrais extra-hipotalâmicas (Buijs et al., 1984; Lindvall et al., 1984).
O envolvimento destes neurónios dopaminérgicos ao nível paraventricular no controlo da ereção e cópula peniana é apoiado por estudos de microdiálise que mostram que as concentrações de dopamina extracelular e ácido 3,4-dihidroxifenilacético (DOPAC), o seu principal metabolito, estão aumentadas no dialisado obtido a partir de o núcleo paraventricular de ratos machos sexualmente potentes apresentando ereções sem contato quando colocados na presença de uma fêmea receptora de primo ovariectomizado estrogênio + progesterona (Melis et al., 2003).
O aumento das concentrações de dopamina e DOPAC foi ainda maior quando a cópula com a fêmea receptiva foi permitida (Melis et al., 2003), como encontrado na área pré-óptica medial (Hull et al., 1995) e no nucleus accumbens (Pfaus e Everitt, 1995). Em segundo lugar, vários estudos mostram que a ereção peniana induzida pela estimulação dos receptores D2 paraventriculares é mediada pela ocitocina liberada nessas áreas. Consequentemente, a apomorfina administrada em doses que induzem a ereção peniana foi encontrada capaz de aumentar a concentração de oxitocina, não apenas no plasma de ratos e macacos (Melis et al., 1989a; Cameron et al., 1992), mas também no cérebro extra-hipotalâmico. áreas, como o hipocampo (Melis et al., 1990). Em consonância com estes resultados, a apomorfina injetada no núcleo paraventricular a uma dose que induz a ereção peniana foi recentemente demonstrada como capaz de aumentar também a concentração de dopamina extracelular no nucleus accumbens, um efeito reduzido pelo antagonista do receptor de oxitocina d (CH2) 5Tyr (Me) 2-Orn8-vasotocina injetado na área tegmentar ventral (Succu et al., 2007; Melis e outros, 2009a) (ver também Seção 4). Terceiro, lesões eletrolíticas bilaterais do núcleo paraventricular, que quase eliminam completamente a ocitocina de áreas cerebrais extra-hipotalâmicas (Hawthorn et al., 1985), abolem a ereção peniana induzida pela apomorfina (Argiolas et al., 1987a) e antagonistas seletivos do receptor de ocitocina administrados nos ventrículos laterais, mas não no núcleo paraventricular, reduzem a ereção peniana induzida pela apomorfina, dose-dependente, com uma potência paralela àquela desses compostos no bloqueio dos receptores de ocitocina (Melis et al., 1989b). Os antagonistas do receptor de ocitocina também são extremamente potentes na redução da facilitação do comportamento sexual masculino induzido não apenas pela ocitocina, mas também pela apomorfina (Argiolas et al., 1988, 1989).
Quanto ao mecanismo pelo qual os receptores D2, ativados pela dopamina ou pelos agonistas do receptor de dopamina, aumentam a atividade dos neurônios oxitocinérgicos, liberando a oxitocina em áreas cerebrais extra-hipotalâmicas e, na medula espinhal, inúmeros dados experimentaissustenta a hipótese de que a estimulação dos receptores D2 aumenta a concentração de íons Ca2 + intracelulares dentro dos corpos celulares dos neurônios oxitocinérgicos, levando à ativação da óxido nítrico-sintase, uma enzima Ca2 + dependente de calmodulina, que está presente nesses corpos celulares (Vincent e Kimura, 1992; Torres et al., 1993; Sanchez et al., 1994; Sato-Suzuki et al., 1998). O aumento da produção de óxido nítrico provoca, por sua vez, a ativação de neurônios oxitocinérgicos. Consequentemente, a ereção peniana induzida por apomorfina (1) foi impedida pelos bloqueadores dos canais de cálcio orgânico e pela GVIA por conotoxina, um potente e seletivo bloqueador dos canais Ca2 + dependentes de voltagem do tipo N (McCleskey et al., 1987), administrados no ventrículo paraventricular. núcleo (ver Argiolas et al., 1990, e referências nele); (2) A ereção peniana induzida pela apomorfina foi prevenida pelos inibidores da óxido nítrico sintase administrados no núcleo paraventricular (Melis et al., 1994c); e (3) apomorfina e outros agonistas do receptor D2 administrados em doses indutoras de ereção peniana aumentaram a produção de óxido nítrico no dialisado paraventricular obtido por microdiálise intracerebral, um aumento que foi reduzido por inibidores da óxido nítrico sintase paraventricular administrados em doses que reduzem Ereção peniana induzida por agonista do receptor D2 (Melis et al., 1996). O mecanismo por meio do qual o óxido nítrico ativa os neurônios oxitocinérgicos paraventriculares ainda é desconhecido, embora os dados disponíveis sugiram que o óxido nítrico atua como um mensageiro intracelular e que a guanilato ciclase não está envolvida. De fato, o fosfodiéster ativo análogo do GMP cíclico, GMP cíclico 8-bromo, foi encontrado incapaz de induzir a ereção peniana quando administrado no núcleo paraventricular (Fig. 2) (ver Melis e Argiolas, 1995b e suas referências).
A interpretação acima tem sido freqüentemente considerada não convincente, principalmente porque a estimulação dos receptores dopaminérgicos D2 é geralmente associada à inibição, e não à excitação, dos corpos celulares dos neurônios que contêm esses receptores através de diferentes mecanismos acoplados à proteína G. (veja Sokoloff e Schwartz, 1995). No entanto, uma possível explicação para essa discrepância, que está de acordo com a estimulação direta dos neurônios oxitocinérgicos paraventriculares pela dopamina, foi sugerida recentemente pela descoberta de uma Dopamina acoplada a proteína G D4 receptor, um membro da famia de receptores D2 (D2, D3 e D4), cuja estimulao aumenta o influxo de Ca2 + em preparaes celulares contendo uma vers clonada deste subtipo de receptor (Moreland et al., 2004). Mais importante, um agonista seletivo do receptor D4 (por exemplo, ABT 724) (maleato de N-metil-4- (2-cyanophenyl) piperazinil-3metilbenzamida) foi encontrado capaz de induzir a ereção peniana em ratos machos quando dado sistemicamente (Brioni et al., 2004). Este efeito não foi encontrado com o agonista selectivo do subtipo do receptor D2 PNU-95666E (R-5,6-di-hidro-N, N-dimetil-4Himidazo [4,5,1-i] quinolin-5-amina) (Hsieh et al., 2004), que também foi incapaz de aumentar o influxo de Ca2 + nas preparações de células contendo a versão clonada do subtipo de receptor D4 (Brioni et al., 2004; Moreland e outros, 2004). De acordo com a hipótese e os resultados acima, PD 168,077 (maleato de N-metil-4- (2-cianofenil) piperazinil-3metilbenzamida), PIP-3EA (2- [4- (2-Metoxifenil) piperazina-1-ilmetil] imidazo [1,2-a] piridina) e outros agonistas do receptor D4 (Heier et al., 1997; Melis et al., 2006b; Löber et al., 2009), foram encontrados capazes de induzir a ereção peniana quando injetados sistemicamente, icv e em o núcleo paraventricular, embora com menor eficácia que a apomorfina. O efeito pró-eréctil destes agonistas do receptor D4 foi evitado pelo L-745,870 (tricloridrato de 3H-pirrolo [4-B] piridina) 1- (1- [clorofenil] piperazin-2,3-il) -metil-4H-pirrolo), um receptor D1997 selectivo antagonista (Patel e outros, 2005; Melis e outros, 2006, 2009b; Löber e outros, XNUMX).
Finalmente, o efeito pró-erétil dos agonistas do receptor D4 acima também foi reduzido pelos inibidores da sintase do óxido nítrico, administrados no núcleo paraventricular, e pelo d (CH2) 5Tyr (Me) 2-Orn8-vasotocina, um antagonista seletivo do receptor de oxitocina dado icv mas não no núcleo paraventricular. Esses resultados estão de acordo com a hipótese de que os agonistas do receptor D4 também estimulam os neurônios oxitocinérgicos ativando a óxido nítrico sintase e liberando a ocitocina em áreas cerebrais extra-hipotalâmicas, o que facilita a ereção peniana, como mostrado para apomorfina e agonistas D2 clássicos (Melis et al. ., 2005, 2006b; Löber et al., 2009).
Os achados acima também suportam a hipótese de que a dopamina induz a ereção peniana atuando sobre os receptores D4 localizados nos corpos celulares dos neurônios oxitocinérgicos paraventriculares, e que causam um influxo aumentado de Ca2 + nos corpos celulares dos neurônios oxitocinérgicos, levando ao aumento da produção de óxido nítrico. O óxido nítrico, por sua vez, ativa os neurônios oxitocinérgicos para liberar a ocitocina em áreas do cérebro extra-hipotalâmicas e na medula espinhal, como já discutido. A respeito disso, Vale ressaltar que os receptores de dopamina foram identificados nos corpos celulares dos neurônios oxitocinérgicos no núcleo paraventricular. apenas recentemente por estudos de imunofluorescência dupla com alta seletividade de anticorpos D2, D3 e D4 e com anticorpos para oxitocina. Estes estudos mostraram a expressão de todos os três subtipos de receptores D2 (D2, D3 e D4), que co-localizam separadamente nos corpos celulares dos neurônios oxitocinérgicos no núcleo paraventricular (e também no núcleo supra-óptico e na área pré-óptica medial) (Baskerville). e Douglas, 2008; Baskerville e outros, 2009).
Isso proporciona um forte apoio neuroanatômico à possibilidade de que os agonistas dos receptores dopaminérgicos e dopaminérgicos do tipo D2 induzam a ereção peniana ativando diretamente os neurônios oxitocinérgicos que se projetam para as áreas cerebrais extra-hipotalâmicas descritas acimapor exemplo, a medula espinhal, a área tegmentar ventral, o hipocampo e a amígdala. No entanto, esses achados não fornecem nenhuma ajuda para a identificação do subtipo de receptores D2, cuja estimulação causa a resposta erétil. Infelizmente, nenhuma ajuda pode ser obtida mesmo de estudos que visem identificar neurônios oxitocinérgicos ativados por agonistas de receptores de dopamina no núcleo paraventricular. De fato, apesar da atividade diferente nos vários subtipos de receptores dopaminérgicos, agonistas mistos do receptor da dopamina (por exemplo, apomorfina) ou agonistas seletivos do receptor D2 (por exemplo, quinpirole, que atua em todos os subtipos do receptor D2) ou agonistas seletivos do receptor D4 a ativação de neurônios oxitocinérgicos, medida pelo aumento da proteína FOS em neurônios ocitocinérgicos parvocelulares do núcleo paraventricular (Bitner et al., 2006). No entanto, este achado tem sido questionado recentemente, pois o aumento da proteína FOS nos neurônios para-ventriculares foi encontrado apenas quando a ereção peniana foi induzida pelo quinerolano, que atua principalmente nos subtipos dos receptores D2 e D3, mas não pelo PD 168077, um agonista do receptor D4. apesar da capacidade de ambos os compostos induzirem a resposta sexual (Baskerville et al., 2009).
Outras experiências com agonistas seletivos dos outros subtipos de receptores D2 (principalmente D2 e D3) são então necessárias para identificar o papel exato de cada subtipo de receptor de dopamina no controle da função erétil no nível paraventricular. A esse respeito, como já lembrado anteriormente, a apomorfina, que atua de forma potente em todos os subtipos de receptores de dopamina (ver Brioni et al., 2004 e suas referências), é muito mais eficaz do que os agonistas dos receptores D4 na indução da ereção peniana quando injetada no ventrículo paraventricular. núcleo. Isto pode ser explicado por uma maior afinidade da apomorfina nos receptores D4 quando comparada com a dos agonistas do receptor D4 testados, ou alternativamente, os agonistas do receptor D4 podem actuar como agonistas parciais do receptor D4, ou a activação concomitante de diferentes subtipos do receptor da dopamina pela apomorfina pode produzir uma maior activação dos neurónios oxitocinérgicos que medeiam a ereção peniana, do que a activação apenas pelos agonistas do receptor D2 do subtipo do receptor D4.
As interações entre os receptores dopaminérgicos D1 e D2 já foram descritas no controle do comportamento sexual no nível da área pré-óptica medial (ver Hull et al., 1989). No caso em que a incapacidade de agonistas do receptor D2 para induzir a ereção peniana (Hsieh et al., 2004) será confirmada (ver Depoortère et al., 2009), por exemplo, mesmo após a injeção desses compostos no núcleo paraventricular , um papel importante para os receptores D3 sozinhos ou em conjunto com os receptores D4 na ativação de neurônios oxitocinérgicos mediando a ereção peniana deve ser analisado em detalhe (ver Baskerville et al., 2009). Infelizmente, os agonistas seletivos dos receptores D2 e D3 (por exemplo, que diferem em sua afinidade por esses dois subtipos de receptores por pelo menos quatro / cinco ordens de magnitude in vitro) não estão disponíveis no momento. Por esta razão, a recente sugestão de que os receptores D3 medeiam a ereção peniana induzida pelos agonistas clássicos do receptor D2, que se baseia principalmente na capacidade de antagonistas putativos do receptor D3 caracterizados em experiências in vitro, de reduzir a ereção peniana induzida por agonistas D2 clássicos, tais como apomorfina, quinpirole e pramipexole, que ativam potencialmente todos os subtipos de receptores dopaminérgicos D2 (Collins et al., 2009), precisam certamente ser validados com outros experimentos. Esta validação é necessária também porque nenhum efeito dos agonistas do receptor D4 na ereção peniana foi encontrado neste estudo, em contraste marcante com os resultados dos estudos citados acima, que demonstram um efeito pró-erétil dos agonistas do receptor D4. De fato, mesmo a capacidade da apomorfina de induzir a ereção peniana (e bocejar) em camundongos knockout para o receptor D4 com uma potência idêntica àquela observada em camundongos knockout do receptor D4 do tipo selvagem e a capacidade dos antagonistas do receptor D3 de abolir a resposta da apomorfina nestes animais ( Collins et al., 2009) não pode ser considerado como uma evidência definitiva para um papel seletivo do subtipo de receptor D3 na ereção peniana induzida por agonista do receptor D2. Diferenças entre as espécies, estudos com neurotransmissores / neuropeptídeos e / ou ablação de neurotransmissores / receptores de neuropeptídeos (neurotransmissores / neuropeptídeos e / ou neurotransmissores / animais knockout para receptores de neuropeptídeos) geralmente adicionam mais confusão e complicações na confirmação do papel sexual putativo de neurotransmissores e / ou neuropeptídeos e seus receptores. Ou seja, a ablação do gene da ocitocina produz ocitocina knock out de camundongos que acasalam e copulam normalmente, como se a ocitocina fosse desnecessária para acasalamento e cópula. Além disso, os camundongos knockout de oxitocina fêmea homozigotos apresentam acasalamento e parto normais, embora com um acentuado comprometimento da descida do leite (Nishimori et al., 1996; Young et al., 1996). A ablação do gene que codifica a sintase do óxido nítrico neuronal também produz camundongos knockout da sintase do óxido nítrico que acasalam e copulam normalmente (Huang et al., 1993). Isso ocorre apesar de a óxido nítrico sintase produzir óxido nítrico, que é um dos principais mediadores fisiológicos da ereção peniana no nível peniano (Burnett et al., 1992; Rajfer et al., 1992) e no nível central, no núcleo paraventricular do hipotálamo (Argiolas, 1994; Benelli et al., 1995; Melis et al., 1998).No entanto, esses achados provavelmente indicam uma característica importante da fisiologia reprodutiva, ou seja, a redundância dos sistemas envolvidos em seu controle a nível central e periférico.
Tal redundância certamente tem origem evolutiva, pois garante a passagem de genes para a próxima geração para a sobrevivência da espécie. Portanto, o fato de que a ablação do gene do receptor D4 não altera o efeito pró-erétil da apomorfina, sugere que os receptores D4, como ocitocina e óxido nítrico, são apenas alguns dos mediadores que atuam nos sistemas que controlam a função erétil, em vez de sugerindo que não há papel para esses receptores no controle da ereção peniana e do comportamento sexual. A falha dos agonistas D4 em induzir a ereção peniana quando administrada sistemicamente a ratos machos de diferentes estirpes foi recentemente relatada por outro estudo (Depoortère et al., 2009). No entanto, em contraste com o trabalho de Collins et al. (2009), e para tornar a imagem sobre o papel dos diferentes subtipos de receptores D2 no controle da ereção peniana ainda mais intrigante, este estudo também mostra que antagonistas seletivos de receptores D3 putativos sistemicamente foram incapazes de reduzir a ereção peniana induzida pela apomorfina em ratos machos da linhagem mais sensíveis ao efeito pró-erétil da apomorfina, enquanto a resposta sexual (e bocejar) foi antagonizada pelo antagonista seletivo de D2 L-741,626 (3 - [[4- (4-clorofenil) -4-hidroxipiperidina] -1-yl] metil-1H-indole), levando os autores a sugerir que os receptores D2, em vez dos receptores D3 e D4, são os que desempenham um papel importante na ereção peniana induzida pelo agonista D2 (Depoortère et al., 2009). Finalmente, a possibilidade de que o efeito excitatório dos agonistas do receptor de dopamina nos neurônios oxitocinérgicos mediando a ereção peniana, pelo menos em parte, seja indireta e não direta, por exemplo, mediada ou influenciada por mudanças na atividade de outros neurotransmissores neuropeptídeos capazes de modular a atividade de neurônios oxitocinérgicos no núcleo paraventricular, não pode ser completamente descartada.
3.2. Interação ácido glutâmico-ocitocina no núcleo paraventricular
O núcleo paraventricular do hipotálamo é muito rico em sinapses contendo um aminoácido excitatório como neurotransmissor (por exemplo, ácido glutâmico e ácido aspártico).) (Van Den Pol, 1991). Aminoácidos excitatórios neste núcleo estão envolvidos em inúmeras funções, incluindo ereção peniana e comportamento sexual (Roeling et al., 1991; Melis et al., 1994b, 2000, 2004b). Por conseguinte, o ácido N-metil-d-aspártico (NMDA), um agonista selectivo do subtipo de receptor de NMDA, mas não o ácido (±) -_- amino-3-hidroxi-5-metil-isoxazole-4-propiónico (AMPA) , um agonista seletivo do subtipo de receptor de AMPA ou ácido (±) -trans (1) -amino-1,3-ciclopentano dicarboxílico (ACPD), um agonista seletivo do subtipo de receptor metabotrópico, foi encontrado capaz de induzir ereção peniana quando injetado no núcleo paraventricular de ratos em movimento livre (Melis et al., 1994b). O efeito pró-eréctil de 948 MR Melis, A. Argiolas / Neurociências e Revisões BioComportamentais 35 (2011) 939-955 NMDA foi prevenido por (+) MK-801, um antagonista não competitivo do receptor de NMDA (Woodruff et al., 1987 ), injetado no núcleo paraventricular (Melis et al., 1994b). Em consonância com esses resultados, em estudos de telemetria para monitorar a pressão intracavernosa, o NMDA foi muito mais ativo do que os agonistas dos outros subtipos de receptores de aminoácidos excitatórios quando injetados no núcleo paraventricular em aumento da pressão intracavernosa em ratos acordados ou anestesiados (Zahran et al. al., 2000; Chen e Chang, 2003).
Como sugerido acima para a ocitocina e a dopamina, é provável que os receptores NMDA mediadores da ereção peniana estejam localizados nos corpos celulares dos neurônios oxitocinérgicos, uma vez que as terminações nervosas dos aminoácidos excitatórios colidem com os corpos celulares oxitocinérgicos no núcleo paraventricular. (Van Den Pol, 1991). Em analogia ao encontrado com os agonistas do receptor de dopamina, o efeito pró-erétil do NMDA é aparentemente mediado pela ativação da neurotransmissão oxitocinérgica, sendo abolido pelo antagonista da oxitocina d (CH2) 5Tyr (Me) 2-Orn8-vasotocina dado icv, mas não no núcleo paraventricular (ver Argiolas e Melis, 1995, 2004, 2005 e referências nele). Da mesma forma, a ativação da neurotransmissão oxitocinérgica induzida por NMDA também é secundária à ativação da sintase do óxido nítrico, uma vez que a ereção peniana induzida pelo NMDA é prevenida pelos inibidores da óxido nítrico sintase (N-Nitro-N-metil-arginina metil éster e N- metil-tio-l-citrulina) administrada no núcleo paraventricular, e NMDA injetada no núcleo paraventricular em doses que induzem a ereção peniana, aumenta a produção de óxido nítrico no núcleo hipotalâmico (ver Argiolas e Melis, 1995, 2004, 2005 e as referências nele contidas) ). Quanto aos agonistas dos receptores dopaminérgicos, a ativação da óxido nítrico induzida por NMDA também pode ser secundária a um influxo aumentado de Ca2 + em corpos celulares oxitocinérgicos através dos receptores NMDA acoplados ao canal Ca2 +, como mostrado em várias preparações neurais (para uma revisão ver Snyder, 1992; Southam e Garthwaite, 1993; Schuman e Madison, 1994 e suas referências). O óxido nítrico, por sua vez, ativa a neurotransmissão oxitocinérgica (ver acima). A origem das projeções glutamatérgicas que ativam neurônios ocitocinérgicos paraventriculares mediando a ereção peniana é desconhecida, embora algumas evidências neuroanatômicas e eletrofisiológicas sugiram que elas possam ter origem, pelo menos em parte, no hipocampo (Saphier e Feldman, 1987; Chen et al., 1992) . Embora seja necessário um trabalho adicional para caracterizar melhor a origem das projeções glutamatérgicas no núcleo paraventricular, o envolvimento do ácido glutâmico no núcleo paraventricular no controle da ereção peniana e do comportamento sexual é claramente apoiado por estudos de microdiálise. Consequentemente, as concentrações extra-celulares de ácido glutâmico e ácido aspártico foram aumentadas no dialisado obtido do núcleo paraventricular de ratos machos mostrando ereções sem contato quando colocadas na presença de ratos fêmeas receptivos com estrogênio + progesterona inacessíveis (Melis et al., 2004b), ereções penianas que também são mediadas pela ativação da transmissão central oxitocinérgica (Melis et al., 1999a, b). Tais aumentos foram encontrados ainda mais quando a cópula com a fêmea receptiva foi permitida (Melis et al., 2004a). Em consonância com a hipótese de que uma atividade aumentada de aminoácidos excitatórios ocorre no núcleo paraventricular durante a ereção e cópula peniana, ambas as ereções sem contato e o comportamento copulatório (durante os quais ocorrem as ereções penianas) são reduzidos pelo bloqueio dos receptores NMDA no núcleo paraventricular. e esta redução é seguida por uma diminuição no aumento da produção de óxido nítrico que ocorre neste núcleo hipotalâmico nestes contextos fisiológicos (Melis et al., 2000). Um aumento na concentração de ácido glutâmico extracelular secundário à diminuição da liberação de GABA das terminações nervosas GABAérgicas que colidem com sinapses aminoacidérgicas excitatórias justapostas a corpos celulares oxitocinérgicos, também foi encontrado no núcleo paraventricular após o bloqueio dos receptores canabinóides CB1 pelo antagonista CB1 SR 141761A, dada nos ventrículos laterais ou diretamente no núcleo paraventricular em doses que induzem a ereção peniana (ver Succu et al., 2006; Castelli et al., 2007). Esse aumento levou à ativação da óxido nítrico sintase nos corpos celulares dos neurônios oxitocinérgicos, aumentando a produção de óxido nítrico. O óxido nítrico, por sua vez, ativa os neurônios oxitocinérgicos mediando a ereção peniana como descrito acima. Em consonância com esse mecanismo, a ereção peniana induzida pelo SR 141761A foi reduzida pelo bloqueio dos receptores NMDA e pelos inibidores da óxido nítrico sintase, mas não pelo bloqueio dos receptores de dopamina ou ocitocina no núcleo paraventricular, enquanto foi evitado pelo bloqueio de receptores de ocitocina central por antagonistas do receptor de ocitocina administrados por ICV (Melis et al., 2004a, 2006a).
3.3. Interação ocitocina-dopamina na região tegmentar ventral
A ocitocina induz a ereção peniana quando injetada na parte caudal da área tegmentar ventral de maneira dose-dependente(Melis et al., 2007). A dose activa mais baixa injectada unilateralmente foi de 20 ng, enquanto a dose mais elevada testada foi de 100 ng. O efeito da ocitocina é mediado pela ativação de receptores oxitocinérgicos, pois a resposta sexual é abolida pela injeção prévia do antagonista da oxitocina d (CH2) 5Tyr (Me) 2-Orn8-vasotocina na área tegmentar ventral caudal. Esses receptores estão localizados nos corpos celulares dos neurônios dopaminérgicos, que se projetam principalmente para a casca do núcleo accumbens. Assim, estudos de dupla imunofluorescência (1) mostram que na área tegmentar ventral caudal as fibras oxitocinérgicas estão em íntimo contato com os corpos celulares dos neurônios dopaminérgicos, a maioria dos quais foram positivamente marcados para tirosina-hidroxilase e contendo o traçador retrógrado Fluorogold anteriormente injetada na carapaça do nucleo accumbens (Melis et al., 2007), e (2) ereção peniana induzida pela oxitocina na área tegmentar ventral ocorre concomitantemente com um aumento na concentração de dopamina extra-celular no dialisato obtido da casca de o núcleo accumbens (Melis et al., 2007). A ereção peniana induzida pela oxitocina também ocorre concomitantemente com um aumento na produção de óxido nítrico na área tegmentar ventral, sendo ambas as respostas antagonizadas não apenas pelo d (CH2) 5Tyr (Me) 2-Orn8-vasotocina e pelo inibidor da óxido nítrico sintase S- metil-tio-l-citrulina, mas também por _-conotoxina, um bloqueador de canais Ca2 + dependente de voltagem e por ODQ (1H- [1,2,4] oxadiazole [4,3-a] quinoxalin-1-ona), um potente inibidor de guanilato ciclase, todas dadas na área tegmentar ventral caudal antes da ocitocina (Succu et al., 2008). Como muitos dos corpos celulares dopaminérgicos marcados com Fluorogold contatados por fibras oxitocinérgicas, considerados positivos para tirosina hidroxilase na área tegmentar ventral caudal, também foram marcados positivamente para óxido nítrico-sintase e guanilato ciclase (Succu et al., 2008), oxitocina A ereção peniana induzida pode ser mediada pelo seguinte mecanismo. A ativação de receptores oxitocinérgicos em corpos celulares dopaminérgicos pelo peptídeo aumenta o influxo de Ca2 + dentro dos corpos celulares de neurônios dopaminérgicos. Isso ativa a óxido nítrico sintase, uma enzima dependente de calmodulina Ca2 +, aumentando assim a produção de óxido nítrico. O óxido nítrico, por sua vez, ativa a guanilato ciclase, levando a uma concentração aumentada de GMP cíclico. O GMP cíclico ativa os neurônios dopaminérgicos que se projetam para o nucleus accumbens. O papel do GMP cíclico na ereção peniana induzida pela ocitocina injetada na área tegmentar ventral caudal também é sustentado pela capacidade do GMP 8-bromo-cíclico, um análogo ativo do GMP cíclico, resistente à fosfodiesterase, para induzir a ereção peniana quando injetado no área tegmentar ventral caudal, e para aumentar a dopamina extracelular no dialisato do nucleus accumbens (Succu et al., 2008; Melis e outros, 2009a). Isto está em desacordo com o mecanismo pelo qual o óxido nítrico ativa os neurônios oxitocinérgicos no núcleo paraventricular, sendo o GMP 8-bromo-cíclico incapaz de induzir a ereção peniana quando injetado neste núcleo (Melis e Argiolas, 1995b) (Fig. O papel da dopamina liberada no núcleo accumbens na ereção peniana induzida pela oxitocina injetada na área tegmental ventral caudal é corroborado pela capacidade do haloperidol, um potente antagonista do receptor de dopamina injetado no núcleo accumbens, de reduzir a resposta à oxitocina (Melis et al., 2007).Quanto às vias neurais ativadas pela dopamina no núcleo accumbens levando à ereção peniana, estas ainda são desconhecidas. No entanto, os dados disponíveis sugerem que essas vias ativam a neurotransmissão da dopamina no núcleo paraventricular do hipotálamo. Consequentemente, a ereção peniana induzida pela ocitocina ocorre concomitantemente a um aumento da dopamina extracelular, nucleus accumbens, mas também no núcleo paraventricular, e é antagonizado pelo antagonista do receptor de dopamina haloperidol injetado no núcleo paraventricular (Melis et al., 2007). UMATodos estes resultados suportam a ideia de que os neurónios oxitocinérgicos que se originam no núcleo paraventricular e se projectam para a área tegmentar ventral caudal, quando activados, libertam oxitocina nesta área, activando assim um sistema de sinalização de GMP cíclico de NO., que, por sua vez, ativa neurônios dopaminérgicos mesolímbicos (Melis e outros, 2007, 2009a; Succu e outros, 2008). A dopamina liberada no nucleus accumbens, por sua vez, ativa as vias neurais que levam à ativação dos neurônios dopaminérgicos incerto-hipotalâmicos, que estimulam os neurônios oxitocinérgicos paraventriculares que se projetam para a medula espinhal, mediando a ereção peniana. Ao mesmo tempo, a dopamina liberada no núcleo paraventricular também pode ativar os neurônios oxitocinérgicos que se projetam para áreas do cérebro extra-hipotalâmicas, como a área tegmental ventral, o hipocampo, a amígdala e talvez outras áreas do cérebro.
Como lembrado acima, de acordo com essa hipótese, a apomorfina injetada no núcleo paraventricular em uma dose que induz a ereção peniana também aumenta a concentração de dopamina extracelular no nucleus accumbens, um efeito reduzido pelo antagonista do receptor de oxitocina d (CH2) 5Tyr (Me ) 2-Orn8-vasotocina injetada na área tegmentar ventral (Succu et al., 2007; Melis et al., 2009a). Juntas, as vias neurais acima podem constituir um circuito neural hipotético envolvendo dopamina, ocitocina e outros neurotransmissores (por exemplo, ácido glutâmico, ver abaixo), influenciando não apenas o desempenho sexual, mas também a motivação sexual e a recompensa sexual. (veja a seção 4).
3.4. Interação ácido oxitocina-glutâmico no subículo do hipocampo ventral
A ocitocina induz a ereção peniana quando injetada na região ventral, mas não no subículo dorsal, de maneira dose-dependente (Melis et al., 2009b). A resposta sexual é mediada pela estimulação dos receptores de oxitocina, sendo abolida pela injeção prévia de d (CH2) 5Tyr (Me) 2-Orn8-vasotocina administrada no mesmo sítio de ocitocina, como encontrado em outras áreas do cérebro (ver acima) . Quanto à localização desses receptores, os dados disponíveis sugerem que eles estão localizados nos corpos celulares de neurônios ricos em óxido nítrico-sintase.
Assim, estudos de microdiálise mostram que a ereção peniana induzida pela ocitocina ocorre concomitantemente com um aumento na produção de óxido nítrico no subículo do ventrículo, e esse aumento é abolido não apenas pela injeção prévia do inibidor da óxido nítrico sintase S-metil-tio-l-citrulina. e pela hemoglobina captadora de óxido nítrico, mas também por d (CH2) 5Tyr (Me) 2-Orn8-vasotocina, todos administrados no mesmo local da ocitocina em doses que antagonizam a ereção peniana (Melis et al., 2010). Mais importante, a ereção peniana induzida pela oxitocina ocorre também concomitantemente com um aumento na concentração de ácido glutâmico extracelular no subículo do ventre, que é apenas parcialmente antagonizado pelo antagonista não competitivo do receptor de NMDA (+) MK-801 administrado no subículo ventral (Melis et al., 2010). Juntos, esses resultados sugerem que o óxido nítrico recém-formado, agindo como um mensageiro intercelular, ativa a neurotransmissão do ácido glutâmico levando à ereção peniana, possivelmente através de projeções neurais eferentes a partir da veia ventral. subículo para áreas do cérebro extra-hipocampais. De acordo com esta hipótese, O NMDA injetado no subículo do ventre induz a ereção peniana de maneira dose-dependente, e este efeito é completamente antagonizado pela injeção prévia no mesmo local de (+) MK-801, mas não por S-metil-tio-l-citrulina. , hemoglobina ou d (CH2) 5Tyr (Me) 2-Orn8-vasotocina (Melis et al., 2010). Quanto às vias neurais eferentes que se projetam para áreas do cérebro extra-hipocampais ativadas por aminoácidos excitatórios (isto é, ácido glutâmico) no subículo do ventre, é provável que sejam glutamatérgicas, assim como a maioria das projeções eferentes do hipocampo. No momento, pode-se sugerir apenas que essas projeções modulem a atividade dos neurônios dopaminérgicos mesolímbicos, que por sua vez modulam a atividade dos neurônios dopaminérgicos orto-hipotalâmicos no núcleo paraventricular, levando à ativação de neurônios ocitocinérgicos. mediação ereção peniana como já foi discutido (Veja acima).
Por conseguinte, a ereção peniana induzida pelo subículo ictínico ocitocina ocorre concomitantemente com um aumento na concentração de dopamina extra-celular na concha do núcleo accumbens, e esse aumento, como a ereção peniana, é abolido por d (CHÂNUMX) 2Tyr (Me) 5- Orn2-vasotocina administrada no subículo do ventre antes da ocitocina (Melis et al., 8b). Além disso, uma vez que a ereção peniana induzida pela ocitocina ventricular é também reduzida por (+) MK-2009 injetada na área tegmentar ventral, mas não no núcleo accumbens (Melis et al., 801b) e ocorre concomitantemente a um aumento extracelular ácido glutâmico na área tegmentar ventral, mas não no nucleus accumbens, sendo ambas as respostas abolidas por d (CH2009) 2Tyr (Me) -Orn5-vasotocina, injetadas no subículo do ventre antes da ocitocina (veja Fig. 8), É provável que a ativação de neurônios dopaminérgicos mesolímbicos seja secundária a uma maior neurotransmissão glutamatérgica na área tegmentar ventral. Isto sugere que existe uma interação ácido glutâmico-dopamina que controla a ereção peniana na área tegmentar ventral. Mais estudos são necessários para verificar se as vias glutametérgicas eferentes pró-eréteis do subículo para a área tegmentar ventral são diretas ou indiretas, ou seja, através do córtex pré-frontal ou de outras áreas do cérebro (ver Melis et al., 2009b e referências). Como o núcleo paraventricular também recebe projeções glutamatérgicas do hipocampo (ver acima e Saphier e Feldman, 1987), e O ácido glutâmico ativa neurônios oxitocinérgicos paraventriculares, incluindo aqueles que se projetam para a área ventral tegmental (ver Argiolas e Melis, 2005 e referências nele), e A ocitocina na região tegmentar ventral induz a ereção peniana e aumenta a atividade do neurônio dopaminérgico mesolímbicos (veja acima), é tentador especular que pneurônios oxitocinérgicos araventriculares também podem estar envolvidos, ao menos em parte, na ativação dos neurônios dopaminérgicos mesolímbicos pela ocitocina injetada no subículo (veja a seção 4).
4. Observações finais
Os estudos revisados acima confirmam e ampliam os achados iniciais, mostrando que nos ratos machos a ocitocina desempenha um papel fundamental no controle central da ereção peniana no nível do núcleo paraventricular do hipotálamo e da medula espinhal. Em particular, a maioriacento estudos mostram que a oxitocina influencia a ereção peniana, agindo também em outras áreas do cérebro, isto é, na área tegmentar ventral, no subículo do ventre e no núcleo cortical posteromedial da amígdala..
No nível paraventricular, o novo achado mais importante talvez seja a descoberta da expressão de todos os receptores de dopamina da família D2 (D2, D3 e D4) nos corpos celulares dos neurônios oxitocinérgicos no núcleo paraventricular (e no núcleo supra-óptico e área pré-óptica medial) (Baskerville e Douglas, 2008; Baskerville et al., 2009). Isso fornece forte neuroanatomia apoio à hipótese de que os agonistas dos receptores dopaminérgicos e dopaminérgicos podem ativar diretamente os neurônios oxitocinérgicos envolvidos na função erétil, projetando-se não apenas para a medula espinhal, mas também para áreas cerebrais extra-hipotalâmicas. A este respeito, é também importante a descoberta de que o aumento da pressão intracavernosa induzida pelo agonista do receptor da dopamina é reduzido pelo bloqueio dos receptores oxitocinérgicos na medula espinal lombo-sacral (Baskerville et al., 2009). De fato, embora tal evidência tenha sido obtida em ratos machos anestesiados, o achado confirma a ativação de uma via descendente ocitocinérgica paraventriculo-espinhal envolvida na ereção peniana induzida por agonista de receptor de dopamina. HNo entanto, ainda precisa ser averiguado se a ereção peniana induzida pela estimulação dos receptores de dopamina presentes nos corpos celulares oxitocinérgicos é secundária à ativação de um subtipo específico de receptor de dopamina da família D2 (D2, D3 ou D4). ou se esses subtipos de receptores cooperarem na modulação da resposta erétil, possivelmente de maneiras diferentes, dependendo do contexto no qual a ereção peniana ocorre (ver Moreland e outros, 2004; Enguehard-Gueiffier e outros, 2006; Melis e outros, 2006a, b; Löber e outros, 2009; Collins e outros, 2009; Depoortère e outros, 2009; Baskerville e outros, 2009).
Outro novo achado importante é que a ocitocina induz a ereção peniana quando injetada não apenas no núcleo paraventricular ou no campo CA1 do hipocampo, mas também na área tegmentar ventral, no subículo do ventre e no núcleo cortical posteromedial da amígdala. Estas áreas do cérebro não foram testadas nos estudos anteriores, mostrando que a oxitocina aumentou os episódios de ereção peniana espontânea em ratos machos, embora eles recebam como as projeções oxitocinérgicas lombo-sacrais da medula espinhal do núcleo paraventricular. A oxitocina foi de fato encontrada capaz de aumentar episódios espontâneos de ereção peniana, que ocorrem em ratos adultos machos na ausência de qualquer estímulo sexual, como aqueles originados da presença de um rato fêmea ovariectomizado receptivo acessível ou inacessível (estimulada por progesterona) manipulação da genitália, quando injetada no núcleo paraventricular e no campo CA1 do hipocampo, mas não no subículo dorsal, no septo lateral, no núcleo caudado, na área pré-óptica medial, no núcleo ventromedial e no núcleo supra-óptico (Melis et al. , 1986). Em todos esses estudos, a ereção peniana era geralmente contada quando o pênis emergia da bainha peniana por um observador que desconhecia os tratamentos administrados diretamente durante o experimento ou, mais tarde, observava a experiência registrada em um videotape com um aparelho de câmera de vídeo. Cada episódio de ereção peniana dura 0.5-1 min e é geralmente acompanhado por aliciamento peniano e / ou flexões do quadril. Nenhum experimento é feito nesses ratos para determinar o efeito da experiência sexual, idade ou se esses ratos podem ser divididos em respondedores baixos ou altos ao efeito pró-erétil da ocitocina injetada nas diferentes áreas do cérebro. Isto também se aplica à maioria dos estudos sobre o efeito pró-erétil de outros neuropeptídeos e drogas que aumentam os episódios espontâneos de ereção peniana, incluindo os agonistas da dopamina, aminoácidos excitatórios, ACTH-MSH, hexarelina e peptídeos VGF. No entanto, o efeito pró-erétil desses compostos tem sido repetidamente confirmado por métodos de telemetria, que determinam a ocorrência da ereção peniana pelo aumento da pressão intracavernosa que ocorre espontaneamente ou após a administração desses compostos por diferentes vias, ou seja, sistemicamente, intracerebroventricularmente ou diretamente em núcleos cerebrais específicos, após o implante de um microtransdutor de pressão diretamente nos corpora cavernosos (ver Bernabè et al., 1999). Na área ventral tegmentar, o subículo ventral e o núcleo póstero-medial da amígdala também a ocitocina induz a ereção peniana, agindo nos receptores oxitocinérgicos. Isso leva à ativação de neurônios dopaminérgicos mesolímbicos originados na área tegmentar ventral e projetando-se para a casca do núcleo accumbens, medida pelo aumento da concentração de dopamina extra-celular no dialisato obtido do invólucro do nucleus accumbens e pela redução da resposta erétil induzida pelo peptídeo injetado nessas áreas extra-hipotalâmicas, encontradas após o bloqueio dos receptores dopaminérgicos no nucleus accumbens (veja abaixo). Quanto aos mecanismos ativados pela estimulação de receptores oxitocinérgicos nessas áreas cerebrais, que levam à ativação dos neurônios dopaminérgicos mesolímbicos e à ereção peniana, os mais esclarecidos são os que ocorrem na área tegmentar ventral caudal. De fato, aqui os resultados farmacológicos e de imunofluorescência mostram que as terminações nervosas da oxitocina atingem os corpos celulares dos neurônios dopaminérgicos que se projetam para o núcleo do nucleus accumbens (Melis et al., 2007, 2009a; Succu et al., 2008). Muitos desses neurônios são ricos em óxido nítrico sintase e em guanilato ciclase. A estimulação de receptores oxitocinérgicos nos corpos celulares desses neurônios dopaminérgicos causa a ativação da óxido nítrico sintase levando a um aumento da produção de óxido nítrico. O óxido nítrico, por sua vez, ativa a guanilato ciclase, aumentando assim a concentração de GMP cíclico, que leva à ativação dos neurônios dopaminérgicos mesolímbicos e à liberação de dopamina no nucleus accumbens, medida pelo aumento da dopamina extracelular no dialisato do nucleus accumbens obtido por microdiálise intracerebral (Succu et al., 2008 ). A dopamina liberada no núcleo accumbens, por sua vez, ativa as vias neurais que levam à ereção peniana. Isto é suportado pela capacidade dos antagonistas do receptor de dopamina, haloperidol e / ou cis-flupentixol, serem injetados no núcleo accumbens para reduzir a ereção peniana induzida pela ocitocina na área tegmentar ventral (Succu et al., 2008). Uma das vias pró-eréteis parece ativar os neurônios dopaminérgicos orto-hipotalâmicos, em particular aqueles que se projetam para os corpos celulares dos neurônios oxitocinérgicos paraventriculares. De fato, a ocitocina injetada na área tegmentar ventral caudal aumenta a dopamina extracelular não apenas no núcleo accumbens, mas também no núcleo paraventricular, e o bloqueio dos receptores dopaminérgicos no núcleo paraventricular reduz significativamente a ereção peniana induzida pela ocitocina na área tegmentar ventral (Succu et al. , 2007, 2008; Melis et al., 2007, 2009a). A existência desses núcleos accumbens dopamina-paraventricular dopamina-paraventricular ocitocina-ventral área tegmentária ocitocina-dopamina links também é sugerido pela capacidade de uma dose pró-erétil de apomorfina e do agonista do receptor D4 PD 168077 injetado no núcleo paraventricular para aumentar a dopamina extracelular na carapaça do nucleus accumbens (Succu et al., 2007), resposta que é abolida por d (CH2) 5Tyr (Me) -Orn8-vasotocina administrada na área tegmentar ventral (Melis et al., 2009a, ver também abaixo). Entretanto, mais trabalhos são necessários para identificar as vias neurais que conectam o núcleo accumbens ao sistema dopaminérgico incerto-hipotalâmico.
O mecanismo pelo qual a ocitocina induz a ereção peniana e ativa os neurônios dopaminérgicos mesolímbicos quando injetada no subículo valvular ou no núcleo póstero-medial da amígdala é apenas parcialmente entendido no momento. Nessas áreas também, a ocitocina ativa seus próprios receptores que levam à ativação da óxido nítrico sintase, aumentando assim a produção de óxido nítrico. O óxido nítrico, por sua vez, ativa projeções eferentes desconhecidas, que aparentemente aumentam a neurotransmissão glutamatérgica na área tegmentar ventral. O ácido glutâmico, então, estimula os neurônios dopaminérgicos mesolímbicos, levando à ereção peniana.Esta hipótese é apoiada principalmente pela capacidade de oxitocina injetada no subículo do ventrículo para aumentar o ácido glutâmico extra-celular na área tegmentar ventral (Fig. 3), e do antagonista não-competitivo do receptor de NMDA (+) MK-801 injetado na área tegmentar ventral, mas não no núcleo accumbens, para reduzir a ereção peniana induzida pela ocitocina injetada no subículo adipital ou no núcleo póstero-medial da amígdala (Melis et al., 2009b). No momento, mais detalhes estão disponíveis para a ereção peniana induzida pela ocitocina do subiculum ventral. Neste caso, a ereção peniana induzida pela ocitocina parece secundária à ativação de receptores oxitocinérgicos localizados nos corpos celulares de neurônios contendo óxido nítrico sintase. Isso causa um aumento na produção de óxido nítrico, que ativa a neurotransmissão glutamatérgica atuando como um mensageiro intercelular com um mecanismo semelhante ao descrito para a potenciação de longo prazo (ver Snyder, 1992; Southam e Garthwaite, 1993; Schuman e Madison, 2004) . De acordo com esta hipótese, a ereção peniana induzida pela ocitocina ocorre concomitantemente com um aumento do ácido glutâmico extracelular no dialisado do subículo valvar, e a estimulação de receptores de aminoácidos excitatórios no subículo do ventrículo esquerdo por NMDA induz a ereção peniana. O ácido glutâmico, por sua vez, ativa as projeções neurais eferentes, que levam à ativação de neurônios dopaminérgicos mesolímbicos na área tegmentar ventral, como relatado acima. Se esses mecanismos estão operando também no núcleo póstero-medial da amígdala é desconhecido no momento. Além disso, mais estudos são necessários para provar que (1) terminações nervosas oxitocinérgicas e receptores no subículo do ventre e no núcleo póstero-medial da amígdala estão localizados em corpos celulares de neurônios contendo óxido nítrico-sintase, (2) se esses neurônios forem eventualmente sob o controle de sinapses de aminoácidos excitatórios (glutamatérgicos) e (3) para identificar o sistema de sinalização ativado pelo óxido nítrico. A esse respeito, vale ressaltar que a capacidade da oxitocina em aumentar a atividade da óxido nítrico sintase nos corpos celulares de neurônios dopaminérgicos na área tegmentar ventral caudal e de neurônios ainda desconhecidos no subículo ventral e no núcleo póstero-medial da amígdala, lembra a capacidade da ocitocina para ativar a óxido nítrico sintase nos corpos celulares dos neurônios oxitocinérgicos no núcleo paraventricular (Melis et al., 1997). No entanto, enquanto na área tegmentar ventral o aumento da produção de óxido nítrico nos corpos celulares dos neurônios dopaminérgicos leva à ativação desses neurônios pela ativação da guanilato ciclase e aumento do GMP cíclico, isso não ocorre no núcleo paraventricular. Consequentemente, o GMP 8-bromo-cíclico injetado no núcleo paraventricular não induz a ereção peniana, ao mesmo tempo em que é injetado na área tegmentar ventral. Outra via de sinalização diferente do sistema GMP oxidicíclico nítrico é então envolvida no nível paraventricular na ativação de neurônios oxitocinérgicos mediando a ereção peniana por óxido nítrico endógeno e / ou exógeno (Melis e Argiolas, 1995b; Melis et al., 1997) (Fig. . Por outro lado, o GMP cíclico na área tegmentar ventral parece desempenhar também um papel fundamental na ativação dos neurônios dopaminérgicos mesolímbicos e no aumento da dopamina extracelular que ocorre no dialisato obtido da casca do núcleo acumbente de ratos machos. selecionado para mostrar ou não mostrar ereções penianas sem contato quando colocado na presença de um rato fêmea receptivo ovariectomizado (estrogênio + progesterona tratado) inacessível. Nestas condições experimentais, em ratos machos mostrando ereções penianas sem contato, um aumento na concentração de dopamina extracelular é encontrado como esperado, e este aumento é ainda maior, embora apenas modestamente, pelos inibidores de fosfodiesterase administrados na área tegmentar ventral caudal (Sanna et al., 2009). Em contraste, em ratos machos que não apresentam ereções sem contato, nos quais não se observa aumento apreciável de dopamina extracelular no núcleo accumbens, a concentração de dopamina extracelular aumenta acentuadamente com a administração de inibidores da fosfodiesterase na área tegmental ventral caudal, e observam-se ereções sem contato (Sanna et al., 2009).
A capacidade de oxitocina injetada na área tegmentar ventral, no subículo valvar e no núcleo cortical posteromedial da amígdala, juntamente com os agonistas dopaminérgicos injetados no núcleo paraventricular, para induzir a ereção peniana e ativar os neurônios dopaminérgicos mesolímbicos merece algum comentário. . Primeiro, mecanismos semelhantes aos lembrados acima podem ser operativos quando a ereção peniana ocorre em contextos fisiológicos, como durante a cópula (quando em ereções penianas do cópula ocorrem) ou durante as ereções penianas sem contato. Essas ereções são ereções penianas mediadas por feromônios indistinguíveis daquelas induzidas por drogas ou ocitocina, que ocorrem quando ratos machos sexualmente potentes são colocados na presença de um rato fêmea receptivo inacessível (estimulado ovariectomizado com estrogênio + progesterona) e são considerados um índice de excitação (Sachs, 1997, 2007). De fato, embora esses resultados não demonstrem que a ocitocina nessas áreas desempenha um papel na ereção peniana que ocorre em contextos fisiológicos ou após a administração do medicamento, eles adicionam mais força aos achados iniciais, sugerindo que essas áreas cerebrais pertencem àquelas onde a ocitocina administrada centralmente age para aumentar não apenas episódios espontâneos de ereção peniana observados após drogas pró-eréteis, mas também para melhorar o comportamento sexual masculino (e feminino) (ver Argiolas e Melis, 2004 e as referências nele contidas). Assim, a concentração de oxitocina aumenta no hipocampo de ratos machos tratados com uma dose pró-erétil de apomorfina, um agonista dopaminérgico clássico (Melis et al., 1990) e d (CH2) 5Tyr (Me) -Orn8-vasotocina, que bloqueia a ocitocina receptores, é extremamente eficaz, não só em prejudicar o comportamento copulatório (Argiolas et al., 1987a), mas também o efeito facilitador da apomorfina no comportamento copulatório masculino (Argiolas et al., 1987b) em ratos machos sexualmente potentes, durante o qual na cópula ereção peniana ocorrer. d (CH2) A 5Tyr (Me) -Orn8-vasotocina também é extremamente potente na redução de ereções sem contato em ratos machos sexualmente potentes, quando administrada em quantidades nanogramas nos ventrículos laterais, mas não no PVN (Melis et al., 1999a).
Segundo, os neurônios dopaminérgicos mesolímbicos desempenham um papel fundamental nas propriedades motivacionais e recompensadoras dos estímulos naturais de reforço, como alimentos, água e atividade sexual (Fibiger e Phillips, 1988; Wise e Rompre, 1989; Everitt, 1990). Em particular, pensa-se que a dopamina libertada por estes neurónios medeia a transposição dos aspectos motivacionais dos estímulos naturais em comportamentos dirigidos por objectivos, por exemplo, no caso da actividade sexual, a procura de um parceiro sexual e de relações sexuais para alcançar recompensa e satisfação. (Goto e Grace, 2005). Consequentemente, a concentração de dopamina extracelular aumenta no dialisado do núcleo accumbens de ratos machos sexualmente potentes durante a exposição a uma ratazana fêmea receptiva com estrogênio + progesterona ovariectomizados inacessíveis, e tal aumento foi ainda maior quando o rato macho foi autorizado a copular com a fêmea receptiva (Pfaus e Everitt, 1995).
Terceiro, os presentes resultados corroboram a hipótese de que um circuito neural conecta o núcleo paraventricular com a área tegmentar ventral direta ou indiretamente (através do subículo e / ou do núcleo cortical posteromedial da amígdala) e do núcleo accumbens, e daqui através de desconhecidos. percorre novamente o núcleo paraventricular para controlar a atividade de neurônios oxitocinérgicos que se projetam para a medula espinhal, mediando a ereção peniana e de neurônios oxitocinérgicos que se projetam para a área ventral tegmentar, o subículo ventral e o núcleo cortical posteromedial da amígdala, modulando assim atividade de neurônios dopaminérgicos mesolímbicos (Fig. 4). Este circuito neural complexo pode desempenhar um papel na integração de atividades neurais envolvidas no controle dos aspectos consumatório (erétil-ejaculatório) e antecipatório (motivacional e recompensador) do comportamento sexual masculino em contextos fisiológicos. De fato, a dopamina extracelular aumenta no nucleus accumbens (Pfaus e Everitt, 1995) e no núcleo paraventricular de ratos machos sexualmente potentes durante a exposição a uma fêmea receptora inacessível, quando ocorrem ereções sem contato, e ainda mais quando a cópula é permitida, por exemplo, quando nas ereções penianas do cópula ocorrem (Melis et al., 2003). Assim, embora mais estudos sejam necessários para esclarecer o papel da ocitocina endógena na área tegmentar ventral, no subículo ventral e na amígdala durante ereções sem contato e comportamento sexual, pode ser razoável supor que esse circuito neural hipotético, enquanto contribui para a aspectos consumadores do comportamento sexual, ao mesmo tempo, também podem ativar o sistema dopaminérgico mesolímbico fornecendo um substrato neural para explicar as propriedades recompensadoras da atividade sexual (Everitt, 1990; Pfaus e Everitt, 1995). A esse respeito, vale ressaltar que o sistema dopaminérgico mesolímbico ativado pela ocitocina injetada na área tegmentar ventral é o mesmo ativado por drogas de abuso, como opiáceos, canabinóides, anfetaminas, cocaína e álcool (Tanda et al., 1997), e que a ocitocina foi encontrada capaz de reduzir a tolerância e dependência à cocaína, morfina, álcool e canabinóides (Kovacs et al., 1998; Cui et al., 2001). Em conclusão, parece que a ocitocina liberada não apenas na área ventral tegmentar, mas também no subículo ventral e no núcleo cortical posteromedial da amígdala, pode ativar os neurônios dopaminérgicos mesolímbicos, que podem estar envolvidos nos efeitos apetitivos e recompensadores da atividade sexual . A ativação dos neurônios dopaminérgicos mesolímbicos pode ser direta, através de receptores oxitocinérgicos nos corpos celulares dos neurônios dopaminérgicos mesolímbicos, ou indiretos, através da ativação da neurotransmissão do ácido glutâmico na região tegmentar ventral.
A dopamina liberada na camada núcleo accumbens modula a atividade dos neurônios dopaminérgicos incerto-hipotalâmicos no núcleo paraventricular causando ou ereção peniana (via ativação de neurônios oxitocinérgicos que se projetam para a medula espinhal), ou motivação e recompensa sexual (via ativação de ocitocinérgicos). neurônios que se projetam para a área ventral tegmentar, o subículo ventral ou o núcleo cortical posteromedial da amígdala). Como a dopamina também é liberada na concha do núcleo accumbens e no núcleo paraventricular quando a ereção peniana ocorre em contextos fisiológicos (por exemplo, ereções sem contato e cópula) (Succu et al., 2007; Melis e outros, 2003, 2007), É provável que os neurônios oxitocinérgicos centrais participem dos circuitos neurais, mediando uma interação entre o sistema mesolímbico e o sistema dopaminérgico incerto-hipotalâmico. Esses circuitos neurais podem desempenhar um papel não apenas na fase consumatória do comportamento sexual (por exemplo, ereção e cópula peniana), mas também na motivação sexual, na excitação sexual e na recompensa sexual.
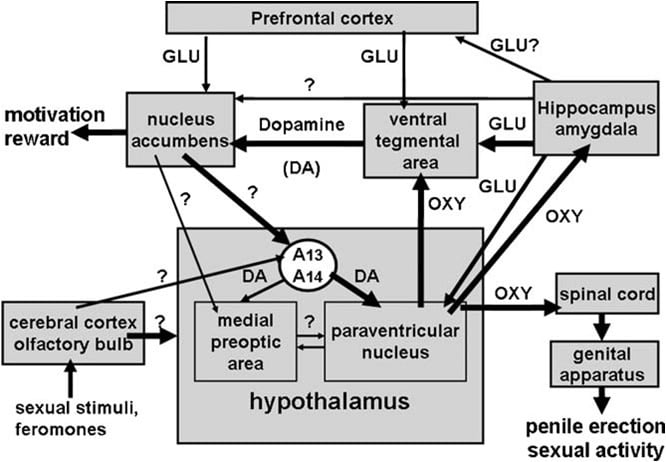
Representação esquemática de um circuito neural hipotético envolvendo a ocitocina, influenciando a motivação sexual, recompensando e desempenho sexual, como sugerido pelos resultados deste capítulo e relatórios anteriores. Neurônios oxitocinérgicos originados no núcleo paraventricular e projetados para a medula espinhal quando ativados, por exemplo, pela dopamina e ácido glutâmico (mas também por outros neurotransmissores e / ou neuropeptídeos), facilitam a ereção peniana e o desempenho sexual pela ativação de neurônios oxitocinérgicos que se projetam para a medula espinhal. A dopamina e o ácido glutâmico (mas também neurotransmissores e neuropeptídeos) no núcleo paraventricular também ativam os neurônios oxitocinérgicos que se projetam para a área tegmentar ventral, ativando os neurônios dopaminérgicos mesolímbicos que se projetam para o núcleo accumbens, modulando a motivação e a recompensa sexual. A dopamina liberada no nucleus accumbens (NAs) ativa por sua vez ainda vias neurais desconhecidas, que aumentam a atividade dos neurônios dopaminérgicos orto-hipotalâmicos (originários dos grupos A13-A14 de Dahlstrom e Fuxe) colidindo entre outros neurônios oxitocinérgicos, incluindo aqueles que se projetam a medula espinhal, levando à ereção peniana. Este circuito também pode ser ativado pela ocitocina injetada não apenas na área tegmentar ventral caudal, mas também no subículo valvar e na amígdala, que também recebem uma inervação oxitocinérgica do núcleo paraventricular, possivelmente por glutamatérgicas direta ou indireta eferente à veia ventral. área tegmentar, levando à modulação da motivação sexual e ereção peniana. Finalmente, os circuitos acima também podem ser ativados por estímulos sexuais e feromônios, uma vez que a dopamina extracelular e o ácido glutâmico aumentam no núcleo paraventricular (e na área pré-óptica medial) durante as ereções e cópulas não contatadas mediadas por feromônio (para referências apropriadas, ver Lista de referências).
Agradecimentos Este trabalho foi parcialmente apoiado por uma bolsa do Ministério Italiano da Universidade e de Pesquisa para AA e MRM.
Referências
Andersson, KE, 2001. Farmacologia da ereção peniana. Pharmacol. Rev. 53, 417-450. Argiolas, A., 1994. O óxido nítrico é um mediador central da ereção peniana. Neurofarmacologia 33, 1339-1344. Argiolas, A., 1999. Neuropeptídeos e comportamento sexual. Neurosci. Biobehav. Rev. 23, 1127-1142. Argiolas, A., Gessa, GL, 1991. Funções centrais da ocitocina. Neurosci. Biobehav. Rev. 15, 217-231. Argiolas, A., Melis, MR, 1995. Neuromodulação da ereção peniana: uma visão geral do papel dos neurotransmissores e neuropeptídeos. Prog. Neurobiol. 47, 235-255. Argiolas, A., Melis, MR, 2004. O papel da ocitocina e do núcleo paraventricular no comportamento sexual de mamíferos masculinos. Physiol. Behav. 83, 309-317. Argiolas, A., Melis, MR, 2005. Controle central da ereção peniana: papel do núcleo paraventricular do hipotálamo. Prog. Neurobiol. 76, 1-21. Argiolas, A., Collu, M., Gessa, GL, Melis, MR, Serra, G., 1988. O antagonista da oxitocina d (CH2) 5Tyr (Me) -Orn8-vasotocina antagoniza o comportamento copulatório masculino em ratos. E. J. Pharmacol. 149, 389-392. Argiolas, A., Collu, M., D'Aquila, P., Gessa, GL, Melis, MR, Serra, G., 1989. A estimulao da apomorfina do comportamento copulatio masculino evitada pelo antagonista da oxitocina d (CH2) 5Tyr (Me) -Orn8-vasotocina em ratos. Pharmacol. Biochem. Behav. 33, 81-83. Argiolas, A., Melis, MR, Gessa, GL, 1985. A ocitocina intraventricular induz a ereção do pênis e do pênis em ratos. E. J. Pharmacol. 117, 395-396. Argiolas, A., Melis, MR, Gessa, GL, 1986. Ocitocina: um indutor extremamente potente da ereção peniana e bocejando em ratos machos. E. J. Pharmacol. 130, 265-272. Argiolas, A., Melis, MR, Mauri, A., Gessa, GL, 1987a. A lesão do núcleo paraventricular impede o bocejo e a ereção peniana induzida pela apomorfina e ocitocina, mas não pelo ACTH em ratos. Cérebro Res. 421, 349-352. Argiolas, A., Melis, MR, Vargiu, L., Mauri, A., Gessa, GL, 1987b. d (CH2) 5Tyr (Me) -Orn8-vasotocina, um potente antagonista da oxitocina, antagoniza a ereção peniana e o bocejo induzido pela ocitocina e apomorfina, mas não pelo ACTH 1-24. E. J. Pharmacol. 134, 221-224. Argiolas, A., Melis, MR, Stancampiano, R., Gessa, GL, 1990. A conotoxina previne a ereção peniana induzida pela apomorfina e da oxitocina e o bocejo em ratos machos. Pharmacol. Biochem. Behav. 37, 253-257. Arletti, R., Bertolini, A., 1985. A oxitocina estimula o comportamento de lordose em ratas. Neuropeptídeos 6, 247-255. Arletti, R., Bazzani, C., Castelli, M., Bertolini, A., 1985. A ocitocina melhora o desempenho copulatório masculino em ratos. Horm. Behav. 19, 14-20. Arletti, R., Benelli, A., Bertolini, A., 1990. O comportamento sexual de ratos machos envelhecidos é estimulado pela ocitocina. E. J. Pharmacol. 179, 377-382. Arletti, R., Calzà, L., Giardino, L., Benelli, A., Cavazzutti, E., Bertolini, A., 1997. A impotência sexual está associada a uma produção reduzida de ocitocina e ao aumento da produção de peptídeos opióides no núcleo paraventricular do hipotálamo. Neurosci. Lett. 233, 65-68. Bancila, M., Giuliano, F., Rampin, O., Mailly, P., Brisorgueil, MJ, Clas, A., Verge, D., 2002. Evidência de uma projeção direta do núcleo paraventricular do hipotálamo a putativos neurônios serotoninérgicos do núcleo paragigantocelular envolvidos no controle da ereção em ratos. E. J. Neurosci. 16, 1240-1249. Burnett, AL, Lowenstein, CJ, Bredt, DS, Chang, TSK, Snyder, SH, 1992. Óxido nítrico: um mediador fisiológico da ereção peniana. Ciência 257, 401-403. Baskerville, TA, Douglas, AJ, 2008. Interação entre dopamina e ocitocina no controle do comportamento sexual. Prog. Cérebro Res. 170, 277-289. Baskerville, TA, Allard, J., Wayman, C., Douglas, AJ, 2009. Interação dopamina-ocitocina na ereção peniana. E. J. Neurosci. 30, 2151-2164. Benelli, A., Bertolini, A., Poggioli, R., Cavazzutti, E., Calzà, L., Giardino, L., Arletti, R., 1995. O óxido nítrico está envolvido no comportamento sexual masculino de ratos. E. J. Pharmacol. 294, 505-510. Bernabè, J., Rampin, O., Sachs, BD, Giuliano, F., 1999. Pressão intracavernosa durante a ereção em ratos: uma abordagem integrativa baseada no registro telemétrico. Sou. J. Physiol. 276, R441-R449. Bitner, RS, Nikkel, AL, Otte, S., Martino, B., Barlow, EH, Bhatia, P., Stewart, AO, Brioni, JD, Decker, MW, Moreland, RB, 2006. Sinalização do receptor de dopamina D4 no núcleo hipotalâmico paraventricular de rato: evidência de acoplamento natural envolvendo indução gênica precoce imediata e fosforilação de proteína cinase ativada por mitógeno. Neurofarmacologia 50, 521-531. Brioni, JD, Moreland, RB, Cowart, M., Hsieh, GC, Stewart, AO, Hedlund, P., Donnelly-Roberts, DL, Nakane, M., Lynch, 3rd., J., Kolasa, T., Polakowski. JS, Osinski, MA, Marsh, K., Andersson, KE, Sullivan, JP, 2004. A ativação dos receptores dopaminérgicos D4 por ABT-724 induz a ereção peniana em ratos. Proc Natl. Acad. Sci U.S.A. 101, 6758-6763. Buijs, RM, 1978. Vias intra e extra-hipotalamica de vasopressina e ocitocina no rato. Tecido Celular Res. 192, 423-435. Buijs, RM, Geffard, M., Piscina, CW, Hoorneman, EMD, 1984. A inervação dopaminérgica do núcleo supra-óptico e paraventricular. Um estudo microscópico de luz e eletrônica. Cérebro Res. 323, 65-72. Caldwell, JD, Prange, AJ, Pedersen, CA, 1986. A oxitocina facilita a receptividade sexual de ratos fêmeas tratados com estrogênio. Neuropeptídeos 7, 175-189. Cameron, JL, Pomerantz, SM, Layden, LM, Amico, JA, 1992. Estimulação dopaminérgica das concentrações de ocitocina no plasma de macacos machos e fêmeas por apomorfina e um agonista do receptor D2. J. Clin. Endocrinol. Metab. 75, 855-860. Canteras, NS, Simerly, RB, Swanson, LW, 1995. Organização de projeções a partir do núcleo medial da amígdala: um estudo PHAL no rato. J. Comp. Neurol. 360, 213-245. Carmichael, MS, Humbert, R., Dixen, J., Palmisano, G., Greeleaf, W., Davidson, JM, 1987. A oxitocina plasmática aumenta na resposta sexual humana. J. Clin. Endocrinol. Metab. 64, 27-31. Carter, CS, 1992. Ocitocina e comportamento sexual. Neurosci. Biobehav. Rev. 16, 131-144. Carter, CS, Lederhendler, II, Kirkpatrick, B., 1997. A Neurobiologia Interativa da Afiliação, Anais da Academia de Ciências de Nova York, vol. 807. Academia de Ciências de Nova York, Nova York. Castelli, MP, Piras, AP, Melis, T., Succu, S., Sanna, F., Melis, MR, Collu, S., Ennas, MG, Díaz, G., Mackie, K., Argiolas, A. 2007 Receptores canabinóides CB1 no núcleo paraventricular e controle central da ereção peniana: imunocitoquímica, autorradiografia e estudos comportamentais. Neurociência 147, 197-206. Chen, KK, Chang, LS, 2003. Efeito de agonistas do receptor de aminoácidos excitatórios na ereção peniana após administração no núcleo paraventricular do hipotálamo em ratos. J. Urol. 62, 575-580. Chen, KK, Chan, JYH, Chang, LS, Chen, MT, Chang, SHH, 1992. Elicitação da ereção peniana após ativação da formação hipocampal no rato. Neurosci. Lett. 141, 218-222. Chen, KK, Chan, JYH, Chang, LS e 1999. Neurotransmissão dopaminérgica no núcleo paraventricular do hipotálamo em regulação central da ereção peniana no rato. J. Urol. 162, 237-242. Collins, GT, Truccone, A., Haji-Abdi, F., Newman, AH, Grundt, P., Arroz, KC, maridos, SM, ganancioso, BM, Enguehard-Gueiffer, C., Gueiffer, A., Chen , J., Wang, S., Katz, JL, Grandy, NS, Sunahara, RK, Woods, JH, 2009. Os efeitos pró-erécteis dos agonistas dopaminérgicos tipo D2 são mediados pelo receptor D3 em ratos e ratinhos. J. Pharmacol. Exp. Ther. 329, 210-217. Coolen, LM, Allard, J., Truitt, WA, McKenna, KE, 2004. Regulação central da ejaculação. Physiol. Behav. 83, 203-215. Cui, SS, Bowen, RC, Gu, GB, Hannesson, BK, Yu, PH, Zhang, X., 2001. Prevenção da síndrome de abstinência de canabinóides pelo lítio: envolvimento da ativação neuronal oxitocinérgica. J. Neurosci. 21, 9867-9876. Dahlstrom, A., Fuxe, K., 1964. Evidência da existência de neurônios contendo monoamina no sistema nervoso central. I. Demonstração de monoaminas nos corpos celulares dos neurónios do tronco cerebral. Acta Physiol. Scand. 62 (Supl. 232), 1-54. Depoortère, R., Bardin, L., Rodrigues, M., Abrial, E., Aliaga, M., Newman-Tancredi, A., 2009. Ereção e bocejos penianos induzidos por agonistas do receptor tipo D2 da dopamina em ratos: influência da deformação e da contribuição da dopamina D2, mas não dos receptores D3 e D4. Behav. Pharmacol. 20, 303-311. Domes, G., Heinrichs, M., Buchel, C., Braus, DF, Herpertz, SC, 2007. A ocitocina atenua as respostas da amígdala às faces emocionais, independentemente da valência. Biol Psiquiatria 62, 11871190. Dominguez, J., Riolo, JV, Xu, Z., Hull, ME, 2001. Regulação pela amígdala medial da cópula e liberação pré-óptica medial de dopamina. J. Neurosci. 21, 349-355. Donaldson, ZR, Young, LJ, 2009. Oxitocina, vasopressina e neurogenética da sociabilidade. Ciência 322, 900-904. Eaton, RG, Markowski, VF, Lumley, Los Angeles, Thompson, JT, Moisés, J., Hull, EM, 1991. Os receptores D2 no núcleo paraventricular regulam as respostas genitais e a cópula em ratos machos. Pharmacol. Biochem. Behav. 39, 177-181. Ebner, K., Bosch, OJ, Kroder, SA, Singewald, N., Neumann, ID, 2005. A liberação de ocitocina na amígdala central de ratos modula o comportamento de enfrentamento do estresse e a liberação de aminoácidos excitatórios. Neuropsicofarmacologia 30, 223-230. Enguehard-Gueiffier, C., Hübner, H., El Hakmaoui, A., Allouchi, H., Gmeiner, P., Argiolas, A., Melis, MR, Gueiffier, A., 2006. 2 - [(4-Fenilpiperazin-1-il) metil] imidazo (di) azinas como ligandos D4 selectivos. Indução da ereção peniana por 2- [4- (2-metoxifenil) piperazina-1-ilmetil] imidazo [1,2-a] piridina (PIP3EA), um agonista D4 potente e seletivo. J. Med. Chem. 49, 3938-3947. Everitt, BJ, 1990. Motivação sexual: uma análise neural e comportamental dos mecanismos subjacentes às respostas apetitivas e copulatórias de ratos machos. Neurosci. Biobehav. Rev. 14, 217-232. Fibiger, HC, Phillips, AG, 1988. Sistema de dopamina mesocorticolímbica e recompensa. Ann. N. Y. Acad. Sci 5, 206-215. Francês, SJ, Totterdell, S., 2003. Neurônios de projeção de accumbens de núcleo individual recebem tanto amídalas basolaterais quanto subferentes ventriculares sublinguais em ratos. Neurociência 119, 19-31. 954 MR Melis, A. Argiolas / Neurociência e Comentários BioComportamentais 35 (2011) 939-955 Freund-Mercier, MJ, Richard, P., 1981. Efeitos excitatórios de injeções intraventriculares de ocitocina no reflexo de ejeção do leite no rato. Neurosci. Lett. 23, 193-198. Freund-Mercier, MJ, Richard, P., 1984. Evidências eletrofisiológicas para o controle facilitatório de neurônios da ocitocina pela ocitocina durante a sucção no rato. J. Physiol. (Lond.) 352, 447-466. Freund-Mercier, MJ, Stoeckel, ME, 1995. Autorreceptores somatodendríticos em neurônios da ocitocina. In: Ivell, R., Russel, JA (Eds.), Ocitocina, abordagens celulares e moleculares em medicina e pesquisa. Adv. Exp. Med. Biol., 365. Plenum Press, Nova York e Londres, pp. 185-194. Freund-Mercier, MJ, Stoeckel, MIM, Palacios, JM, Pazos, JM, Richard, PH, Porte, A., 1987. Características farmacológicas e distribuição anatómica dos locais de ligação da oxitocina 3H no cérebro de ratos Wistar estudados por autoradiografia. Neurociência 20, 599-614. Giuliano, F., Rampin, O., 2000. Controle central da ereção peniana. Neurosci. Biobehav. Rev. 24, 517-533. Giuliano, F., Allard, J., 2001. Dopamina e função sexual. Int. J. Impotência Res. 13 (Supl. 3), 18-28. Giuliano, F., Rampin, O., 2004. Controle neural da ereção. Physiol. Behav. 83, 189-201. Giuliano, F., Bernabè, J., McKenna, KE, Longueville, F., Rampin, O., 2001. Efeito proeretil espinal de ocitocina em ratos anestesiados. Am J. Physiol. Regul. Integ. Comp. Physiol. 280, R1870-R1877. Goto, Y., Grace, AA, 2005. Modulação dopaminérgica do drive límbico e cortical do nucleus accumbens no comportamento objetivo-dirigido. Nat. Neurosci. 8, 805-812. Hawthorn, J., Ang, VT, Jenkins, JS, 1985. Efeitos das lesões nos núcleos hipotalâmico paraventricular, supra-óptico e supraquiasmático na vasopressina e ocitocina no cérebro e medula espinhal de ratos. Cérebro Res. 346, 51-57. Heier, RF, Dolak, LA, Duncan, JN, HlSlop, DK, Lipton, MF, Martin, LJ, Mauragis, MA, Piercey, MF, Nichols, NF, Schreur, PJ, Smith, MW, Lua, MW, 1997. S�tese e actividades biol�icas de (R) -5,6-di-hidro-N, N-dimetil-4H-imidazo [4,5,1-ij] quinolino-5-amina) e seus metabolitos. J. Med. Chem. 40, 639-646. Hsieh, GC, Hollingsworth, PR, Martino, B., Chang, R., Terranova, MA, O'Neill, AB, Lynch, JJ, Moreland, RB, Donnelly-Roberts, DL, Kolasa, T., Mikusa, JP , McVey, JM, Marsh, KC, Sullivan, JP, Brioni, JD, 2004. Mecanismos centrais que regulam a ereção peniana em ratos conscientes: os sistemas dopaminérgicos relacionados ao efeito proerético da apomorfina. J. Pharmacol. Exp. Ther. 308, 330-338. Huang, PL, Dawson, TM, Bredt, DS, Snyder, SH, Fishman, MC, 1993. Ruptura direcionada do gene neuronal da óxido nítrico sintase. Célula 75, 1273-1286. Huber, D., Veinante, P., Stoop, R., 2005. A vasopressina e a ocitocina excitam populações neuronais distintas na amígdala central. Ciência 308, 245-248. Casco, EM, Warner, RK, Bazzett, TJ, Eaton, RC, Thompson, JT, 1989. A relação D2 / D1 na área pré-óptica medial afeta a cópula de ratos machos. J. Pharmacol. Exp. Ther. 251, 422-427. Casco, EM, Du, J., Lorrain, DS, Matuszewich, L., 1995. Dopamina extracelular na área pré-óptica medial: implicações para a motivação sexual e controle hormonal da cópula. J. Neurosci. 15, 7465-7471. Casco, EM, Meisel, RL, Sachs, BD, 2002. Comportamento sexual masculino Em: Pfaff, DW, Arnold, AP, Etgen, AM, Fahrbach, SE, Rubin, RT (Eds.), Hormônios, cérebro e comportamento. Academic Press, New York, pp. 3-137. Hurlemann, R., Patin, A., Onur, OA, Cohen, MX, Baumgartner, T., Metzler, S., Dziobek, I., Gallinat, J., Wagner, M., Maier, W., Kendrick, KM, 2010. A ocitocina aumenta a aprendizagem socialmente reforçada e a empatia emocional dependente da amígdala em humanos. J. Neurosci. 30, 4999-5007. Ivell, R., Russel, JA, 1995. Oxitocina: abordagens celulares e moleculares em medicina e pesquisa. Avanços em Medicina Experimental e Biologia, vol. 395. Plenum Press, Nova Iorque. Kelley, AE, Domesick, VB, 1982. A distribuição da projeção da formação hipocampal ao nucleus accumbens no rato: um estudo peroxidase anterógrada e retrógrada. Neurociência 7, 2321-2335. Kimura, Y., Naitou, Y., Wanibuchi, F., Yamaguchi, T., 2008. A ativação do receptor 5-HT (2C) é um mecanismo comum nos efeitos proeretileis da apomorfina, da ocitocina e do melanotan-II em ratos. E. J. Pharmacol. 589, 157-162. Kondo, Y., Sachs, BD, Sakuma, Y., 1998. Importância da amígdala medial na ereção peniana de ratos evocada por estímulos remotos de fêmeas de estro. Behav. Cérebro Res. 91, 215-222. Kovacs, GL, Sarnyai, Z., Szabo, G., 1998. Ocitocina e dependência: uma revisão. Psychoneuroendocrinology 23, 945-962. Lee, HJ, Macbeth, AH, Pagani, JH, Scott Young 3rd, W., 2009. A ocitocina: o grande facilitador da vida. Prog. Neurobiol. 88, 127-151. Lindvall, O., Bjorklund, A., Skagerberg, G., 1984. Demonstração isotroquímica seletiva de sistemas terminais dopaminérgicos em diensa e telencéfalo de rato: novas evidências para inervação dopaminérgica de núcleos neurossecretores hipotalâmicos. Cérebro Res. 306, 19-30. Liu, YC, Salamone, JD, Sachs, BD, 1997. Resposta sexual prejudicada após lesões do núcleo paraventricular do hipotálamo em ratos machos. Behav. Neurosci. 111, 1361-1367. Löber, S., Tschammer, N., Hübner, H., Melis, MR, Argiolas, A., Gmeiner, P., 2009. O arcabouço do azuleno como um novo bioisostere: projeto de potentes ligantes do receptor dopaminérgico D4 induzindo a ereção peniana. Chem. Med. Chem. 4, 325-328. McCleskey, EW, Fox, AP, Feldman, DH, Cruz, LJ, Olivera, BM, Tsien, RW, Yoshikami, D., 1987. Conotoxina: bloqueio direto e persistente de tipos específicos de canais de cálcio nos neurônios, mas não nos músculos. Prot. Nat. Acad. Sci U.S.A. 84, 4327-4331. McKenna, KE, 2000. Algumas propostas sobre a organização do controle do sistema nervoso central da ereção peniana. Neurosci. Biobehav. Rev. 24, 535-540. Marson, L., McKenna, KE, 1992. Papel da 5-hidroxitriptamina na inibição descendente dos reflexos sexuais espinais. Exp. Cérebro Res. 88, 313-318. Marson, L., McKenna, KE, 1996. Grupos celulares do SNC envolvidos no controle dos músculos isquiocavernoso e bulbospongioso: estudo de rastreamento transneuronal utilizando o vírus da pseudo-raiva. J. Comp. Neurol. 374, 161-179. Meisel, RL, Sachs, BD, 1994. A fisiologia do comportamento sexual masculino. Em: Knobil, E., Neil, J. (Eds.), The Physiology of Reproduction, vol. 2, segunda edição. Raven Press, Nova Iorque, pp. 3-96. Melin, P., Kihlstrom, JE, 1963. Influência da ocitocina no comportamento sexual de coelhos machos. Endocrinologia 73, 433-435. Melis, MR, Argiolas, A., 1995a. Dopamina e comportamento sexual. Neurosci. Biobehav. Rev. 19, 19-38. Melis, MR, Argiolas, A., 1995b. Os doadores de óxido nítrico induzem ereção peniana e bocejam quando injetados no sistema nervoso central de ratos machos. E. J. Pharmacol. 294, 1-9. Melis, MR, Argiolas, A., 2003. Neurotransmissão oxitocinérgica central: um alvo farmacológico para a terapia da disfunção erétil psicogênica. Curr. Alvos de medicamentos 4, 55-66. Melis, MR, Argiolas, A., Gessa, GL, 1986. Bocejo induzido por oxitocina e ereção peniana: local de ação no cérebro. Cérebro Res. 398, 259-265. Melis, MR, Argiolas, A., Gessa, GL, 1987. Bocejo e ereção peniana induzida pela apomorfina: local de ação no cérebro. Cérebro Res. 415, 98-104. Melis, MR, Argiolas, A., Gessa, GL, 1989a. A apomorfina aumenta os níveis plasmáticos de ocitocina em ratos. Neurosci. Lett. 98, 351-355. Melis, MR, Argiolas, A., Gessa, GL, 1989b. Evidência de que a apomorfina induz a ereção e o bocejo do pênis liberando ocitocina no sistema nervoso central. E. J. Phamacol. 164, 565-570. Melis, MR, Argiolas, A., Stancampiano, R., Gessa, GL, 1990. Efeito da apomorfina nas concentrações de ocitocina em diferentes áreas cerebrais e no plasma de ratos machos. E. J. Pharmacol. 182, 101-107. Melis, MR, Mauri, A., Argiolas, A., 1994a. Ereção peniana induzida pela apomorfina e oxitocina e bocejo em ratos machos inteiros e castrados: efeito de esteróides sexuais. Neuroendocrinologia 59, 349-354. Melis, MR, Stancampiano, R., Argiolas, A., 1994b. A ereção peniana e o bocejo induzido por injeções NMDA paraventriculares são mediados pela ocitocina. Pharmacol. Biochem. Behav. 48, 203-207. Melis, MR, Stancampiano, R., Argiolas, A., 1994c. Prevenção pelo éster metílico de NG-nitro-l-arginina da ereção peniana induzida por apomorfina e oxitocina e bocejando: local de ação no cérebro. Pharmacol. Biochem. Behav. 48, 799-804. Melis, MR, Succu, S., Argiolas, A., 1996. Os agonistas dopaminérgicos aumentam a produção de óxido nítrico no núcleo paraventricular do hipotálamo: correlação com a ereção peniana e o bocejo. E. J. Neurosci. 8, 2056-2063. Melis, MR, Succu, S., Iannucci, U., Argiolas, A., 1997. A ocitocina aumenta a produção de óxido nítrico no núcleo paraventricular do hipotálamo: correlação com ereção e bocejo peniano. Reg. Peptídeos 69, 105-112. Melis, MR, Succu, S., Mauri, A., Argiolas, A., 1998. A produção de óxido nítrico é aumentada no núcleo paraventricular do hipotálamo de ratos machos durante a ereção peniana sem contato e a cópula. E. J. Neurosci. 10, 1968-1974. Melis, MR, Spano, MS, Succu, S., Argiolas, A., 1999a. O antagonista da oxitocina d (CH2) 5Tyr (Me) 2-Orn8-vasotocina reduz as ereções penianas sem contato em ratos machos. Neurosci. Lett. 265, 171-174. Melis, MR, Succu, S., Spano, MS, Argiolas, A., 1999b. A morfina injetada no núcleo paraventricular do hipotálamo evita ereções sem contato e prejudica a cópula: envolvimento do óxido nítrico. E. J. Neurosci. 11, 1857-1864. Melis, MR, Spano, MS, Succu, S., Argiolas, A., 2000. Efeito de aminoácidos excitatórios, antagonistas dos receptores de dopamina e oxitocina nas ereções penianas sem contato e produção de óxido nítrico paraventricular em ratos machos. Behav. Neurosci. 114, 849-857. Melis, MR, Succu, S., Mascia, MS, Cortis, L., Argiolas, A., 2003. A dopamina extracelular aumenta no núcleo paraventricular de ratos machos durante a atividade sexual. E. J. Neurosci. 17, 1266-1272. Melis, MR, Succu, S., Mascia, MS, Argiolas, A., 2004a. Antagonismo de canabinóides Os receptores CB1 no núcleo paraventricular de ratos machos induzem ereção peniana. Neurosci. Lett. 359, 17-20. Melis, MR, Succu, S., Mascia, MS, Cortis, L., Argiolas, A., 2004b. Aumento dos aminoácidos excitatórios extracelulares no núcleo paraventricular de ratos machos durante a atividade sexual: papel principal dos receptores NMDA na função erétil. E. J. Neurosci. 19, 2569-2575. Melis, MR, Succu, S., Mascia, MS, Argiolas, A., 2005. O PD-168,077, um agonista seletivo do receptor de D4 da dopamina, induz a ereção peniana quando injetado no núcleo paraventricular de ratos machos. Neurosci. Lett. 379, 59-62. Melis, MR, Succu, S., Mascia, MS, Sanna, F., Melis, T., Succu, S., Castelli, MP, Argiolas, A., 2006a. E ereção peniana induzida por SR 141716A em ratos machos: envolvimento de ácido glutâmico paraventricular e óxido nítrico. Neurofarmacologia 50, 219-228. Melis, MR, Succu, S., Sanna, F., Mascia, MS, Melis, T., Enguehard-Gueiffier, C., Hubner, H., Gmenier, P., Gueiffier, A., Argiolas, A., 2006b. PIP3EA e PD168077, dois agonistas seletivos do receptor de D4 da dopamina, induzem ereção peniana em ratos machos: local e mecanismo de ação no cérebro. E. J. Neurosci. 24, 2021-2030. Melis, MR, Melis, T., Cocco, C., Succu, S., Sanna, F., Pillolla, G., Boi, A., Ferri, GL, Argiolas, A., 2007. A oxitocina injetada na área tegmentar ventral induz a ereção peniana e aumenta a dopamina extracelular no núcleo accumbens MR Melis, A. Análise de Argiolás / Neurociências e BioComportamentais 35 (2011) 939-955 955 e núcleo paraventricular do hipotálamo de ratos machos. E. J. Neurosci. 26, 1026-1035. Melis, MR, Sanna, F., Succu, S., Zarone, P., Boi, A., Argiolas, A., 2009a. O papel da ocitocina nas fases antecipatória e consumatória do comportamento sexual de ratos machos. In: Jastrow, H., Feuerbach, D. (Eds.), Manual de Pesquisa com Ocitocina: Síntese, Armazenamento e Liberação, Ações e Formas de Medicamentos. Nova Publishers Inc, Nova Iorque, EUA, pp. 109-125. Melis, MR, Succu, S., Sanna, F., Boi, A., Argiolas, A., 2009b. A oxitocina injetada no subículo ventral ou no núcleo cortical posteromedial da amígdala induz a ereção peniana e aumenta a dopamina extracelular no núcleo acumbente de ratos machos. E. J. Neurosci. 30, 1349-1357. Melis, MR, Succu, S., Cocco, C., Caboni, E., Sanna, F., Boi, A., Ferri, GL, Argiolas, A., 2010. A ocitocina induz ereção peniana quando injetada no subículo valvar: papel do óxido nítrico e do ácido glutâmico. Neurofarmacologia 58, 1153-1160. Moreland, RB, Nakane, M., Donnelly-Roberts, DL, Miller, LN, Chang, R., Uchic, ME, Terranova, MA, Gubbins, EJ, Helfrich, RJ, Namovic, MT, El-Kouhen, OF, Mestres, JN, Brioni, JD, 2004. Farmacologia comparativa de linhas celulares estelares semelhantes a receptores de dopamina D (2) humana acopladas a fluxo de ccio atrav de Galpha (qo5). Biochem. Pharmacol. 68, 761-772. Moos, F., Freund-Mercier, MJ, Guerne, Y., Guerne, JM, Stoeckel, ME, Richard, P., 1984. Liberação de ocitocina e vasopressina por núcleos magnocelulares in vitro: efeito facilitador específico da ocitocina em sua própria liberação. J. Endocrinol. 102, 63-72. Murphy, MR, Seckl, JR, Burton, S., Checkley, SA, Lightman, SL, 1987. Alterações na secreção de ocitocina e vasopressina durante a atividade sexual em homens. J. Clin. Endocrinol. Metab. 65, 738-741. Nishimori, K., Young, LJ, Guo, Q., Wang, Z., Insel, TR, Matzuk, MM, 1996. A ocitocina é necessária para amamentar, mas não é essencial para o parto ou comportamento reprodutivo. Proc Natl. Acad. Sci U.S.A. 93, 11699-11704. Patel, S., Freedman, S., Chapman, KL, Emms, F., Fletcher, AE, Knowles, M., Marwood, R., Mccallister, G., Myers, J., Curtis, J., Kulagowski, JJ, Leeson, PD, Ridgill, M., Graham, M., Matheson, S., Rathbone, D., Watt, AP, Bristow, LJ, Rupniak, NM, Baskin, E., Lynch, JJ, Ragan, CI. 1997 Perfil biológico de L 745,870, um antagonista seletivo com alta afinidade pelo receptor D4 da dopamina. J. Pharmacol. Exp. Ther. 283, 636-647. Pedersen, CA, Caldwell, JD, Jirikowski, GF, Insel, TR, 1992. Ocitocina em Comportamentos Maternos, Sexuais e Sociais, Anais da Academia de Ciências de Nova York, vol. 652. Academia de Ciências de Nova York, Nova York. Petrovic, P., Kalisch, R., Singer, T., Dolan, RJ, 2008. A ocitocina atenua as avaliações afetivas das faces condicionadas e da atividade da amígdala. J. Neurosci. 28, 6607-6615. Pfaus, JG, Everitt, BJ, 1995. A psicofarmacologia do comportamento sexual. Em: Knobil, FE, Kupfer, DJ (Eds.), Psicofarmacologia: a quarta geração de progresso. Raven Press, Nova Iorque, pp. 742-758. Rajfer, J., Aronson, WJ, Bush, PA, Dorey, FJ, Ignarro, LJ, 1992. Óxido nítrico como mediador do relaxamento do corpo cavernoso em resposta à neurotransmissão não-adrenérgica e não-colinérgica. N. Engl. J. Med. 326, 90-94. Roeling, TAP, Van Erp, AMM, Meelis, W., Kruk, MR, Veening, JG, 1991. Efeitos comportamentais do NMDA injetado no núcleo paraventricular hipotalâmico do rato. Cérebro Res. 550, 220-224. Sachs, BD, 1997. Ereção evocada em ratos machos pelo cheiro no ar de fêmeas de cio. Physiol. Behav. 62, 921-924. Sachs, BD, 2000. Abordagens contextuais da fisiologia e classificação da função erétil, disfunção erétil e excitação sexual. Neurosci. Biobehav. Rev. 24, 541-560. Sachs, BD, 2007. Uma definição contextual de excitação sexual masculina. Horm. Behav. 51, 569-578. Sanchez, F., Alonso, JR, Arevalo, R., Blanco, E., Aijon, J., Vazquez, R., 1994. Coexistência de NADPH-diaforase com vasopressina e ocitocina nos núcleos neuro-secretórios magnocelulares hipotalâmicos do rato. Tecido Celular Res. 276, 31-34. Sanna, F., Succu, S., Boi, A., Melis, MR, Argiolas, A., 2009. Inibidores da fosfodiesterase tipo 5 facilitam a ereção sem contato em ratos machos: local de ação no cérebro e mecanismo de ação. J. Sexo. Med. 6, 2680-2689. Saphier, D., Feldman, S., 1987. Efeitos dos estímulos septal e hipocampal nos neurônios do núcleo paraventricular. Neurociência 20, 749-755. Sato-Suzuki, I., Kita, I., Oguri, M., Arita, H., 1998. Respostas de bocejar estereotipadas induzidas por estimulação elétrica e química do núcleo paraventricular do rato. J. Neurofisiol. 80, 2765-2775. Schuman, EM, Madison, DV, 1994. Óxido nítrico e função sináptica. Ann. Rev. Neurosci. 17, 153-183. Snyder, SH, 1992. Óxido nítrico: primeiro em uma nova classe de neurotransmissores? Ciência 254, 494-496. Sofroniew, MV, 1983. Vasopressina e ocitocina no cérebro e medula espinhal de mamíferos. Tendências Neurosci. 6, 467-472. Sokoloff, P., Schwartz, JC, 1995. Novos receptores de dopamina meia década depois. Trends Pharmacol. Sci 16, 270-275. Southam, E., Garthwaite, J., 1993. A via de sinalização GMP de óxido nítrico-cíclico no cérebro de ratos. Neurofarmacologia 32, 1267-1277. Stancampiano, R., Melis, MR, Argiolas, A., 1994. Ereção e bocejo peniano induzidos por agonistas 5-HT1c em ratos machos: relação com a transmissão dopaminérgica e oxitocinérgica. E. J. Pharmacol. 261, 149-155. Succu, S., Mascia, MS, Sanna, F., Melis, T., Argiolas, A., Melis, MR, 2006. O antagonista do receptor canabinóide CB1 SR 141716A induz a ereção peniana pelo aumento do ácido glutâmico extracelular no núcleo paraventricular de ratos machos. Behav. Cérebro Res. 169, 274-281. Succu, S., Sanna, F., Melis, T., Boi, A., Argiolas, A., Melis, MR, 2007. A estimulação de receptores de dopamina no núcleo paraventricular do hipotálamo de ratos machos induz a ereção peniana e aumenta a dopamina extracelular no núcleo accumbens: envolvimento da ocitocina central. Neurofarmacologia 52, 1034-1043. Succu, S., Sanna, F., Cocco, C., Melis, T., Boi, A., Ferri, GL, Argiolas, A., Melis, MR, 2008. A ocitocina induz ereção peniana quando injetada na área tegmentar ventral de ratos machos: papel do óxido nítrico e GMP cíclico. E. J. Neurosci. 28, 813-821. Tanda, G., Pontieri, FE, Di Chiara, G., 1997. Ativação de canabinóides e heroína da transmissão da dopamina mesolímbica por um mecanismo comum do receptor opióide mu1. Ciência 276, 2048-2050. Tang, Y., Rampin, O., Calas, A., Facchinetti, P., Giuliano, F., 1998. Inervação ocitocinérgica e serotoninérgica de núcleos lombossacrais identificados controlando a ereção peniana em ratos machos. Neurociência 82, 241-254. Teodose, DT, 1985. Os terminais imunorreativos à oxitocina fazem sinapse nos neurónios da ocitocina nos núcleos supra-ópticos. Natureza (Londres) 313, 682-684. Tindall, JS, 1974. Estímulos que causam a liberação de ocitocina. Em: Geiger, SR, Knobil, E., Sawyer, WH, Greef, R., Astwood, EB (Eds.), Manual de Fisiologia. Seita. 7, Endocrinology, vol. IV. American Physiology Society, Washington DC, pp. 257-267. Torres, G., Lee, S., Rivier, C., 1993. Ontogenia da sintase do óxido nítrico hipotalâmico de rato e colocalização com neuropeptídeos. Mol. Célula Neurosci. 4, 155-163. Uhl-Bronner, S., Waltisperger, E., Martinez-Lorenzana, G., Condes, LM, Freund-Mercier, MJ, 2005. Expressão sexualmente dimórfica dos sítios de ligação da ocitocina no prosencéfalo e medula espinhal do rato. Neurociência 135, 147-154. Vaccari, C., Lolait, SJ, Ostrowski, NL, 1998. Distribuição comparativa dos ácidos ribonucléicos vasopressina V1b e receptor de ocitocina no cérebro. Endocrinologia 139, 5015-5033. Van Den Pol, A., 1991. Imunorreatividade de glutamato e aspartato em axônios pré-sinápticos hipotalâmicos. J. Neurosci. 11, 2087-2101. Veronneau-Longueville, F., Rampin, O., Freund-Mercier, MJ, Tang, Y., Calas, A., Marson, L., McKenna, KE, Stoeckel, ME, Benoit, G., Giuliano, F. 1999 Inervação ocitocinérgica de núcleos autonômicos controlando a ereção peniana no rato. Neurociência 93, 1437-1447. Vincent, SR, Kimura, H., 1992. Mapeamento histoquímico da óxido nítrico sintase no cérebro de ratos. Neurociência 46, 755-784. Wagner, CK, Clemens, LG, 1993. Via contendo neurofisina desde o núcleo paraventricular do hipotálamo até um núcleo motor dimórfico sexual na medula espinhal lombar. J. Comp. Neurol. 336, 106-116. Winslow, JT, Insel, TR, 1991. O status social em pares de macacos-esquilo macho determina a resposta comportamental à administração central de ocitocina. J. Neurosci. 11, 2032-2038. Wise, RA, Rompre, P.-P., 1989. Dopamina cerebral e recompensa. Ann. Rev. Psychol. 40, 191-225. Witt, DM, Insel, TR, 1994. O comportamento sexual masculino ativa a proteína c-fos-like nos neurônios da ocitocina no núcleo paraventricular do hipotálamo. J. Neuroendocrinol. 6, 13-18. Witter, MP, 2006. Conexões do subículo do rato: topografia em relação à organização colunar e laminar. Behav Brain Res. 174, 251-264. Woodruff, GN, Foster, CA, Gill, R., Kemp, JA, Wong, EH, Iversen, LL, 1987. A interação entre MK-801 e receptores para N-metil-d-aspartato: consequências funcionais. Neurofarmacologia 26, 903-909. Yamashita, H., Shigeru, O., Inenaga, K., Kasai, M., Uesugi, S., Kannan, H., Kaneko, T., 1987. A oxitocina excita predominantemente os neurônios putativos de ocitocina no núcleo supra-óptico de ratos in vitro. Cérebro Res. 416, 364-368. Grita, DP, Hendricks, SE, Prendergast, MA, 1992. Lesões do núcleo paragigantocelular: efeitos no comportamento de acasalamento em ratos machos. Cérebro Res. 596, 73-79. Young, WS, Shepard, E., Amico, J., Hennighausen, L., LaMarca, ME, McKinney, C., Ginns, EI, 1996. Deficiência na ocitocina do rato previne a ejeção do leite, mas não a fertilidade ou o parto. J. Neuroendocrinol. 8, 847-854. Zahran, AR, Vachon, P., Courtois, F., Carrier, S., 2000. Aumento da pressão peniana intracavernosa após injeções de agonistas do receptor de aminoácidos excitatórios no núcleo paraventricular hipotalâmico de ratos anestesiados. J. Urol. 164, 1793-1797.