OBSERVAȚII: Unul dintre puținele studii care acoperă modul în care abstinența afectează nivelurile de receptori ai dopaminei la primate.
- Receptorii D2 revin destul de repede - mai puțin de o lună
- Receptoarele D1 sunt prea ridicate la o lună, dar săriți înapoi în zilele 90.
- Receptorii D1 de mare sau mică pot fi chei pentru retragerea și pofta acută
Thomas JR Beveridge1, Hilary R Smith1, Michael A Nader1 și Linda J Porrino1
1Departamentul de Fiziologie și Farmacologie, Centrul pentru Investigarea Neurobiologică a Abuzului de Droguri, Universitatea Wake Forest School of Medicine, Winston-Salem, NC, SUA
Corespondență: Dr. LJ Porrino, Departamentul de Fiziologie și Farmacologie, Centrul pentru Investigarea Neurobiologică a Abuzului de Droguri, Wake Forest University School of Medicine, Centrul Medical Boulevard, Winston-Salem, NC 27157-1083, SUA. Tel: + 1 336 716 8575; Fax: + 1 336 716 8501; E-mail: [e-mail protejat]
Primit la 29 aprilie 2008; Revizuit la 25 iulie 2008; Acceptat la 30 iulie 2008; Publicat online 3 septembrie 2008.
Abstract
Deși dysregularea în sistemul de dopamină (DA) este o caracteristică distinctivă a expunerii cocainice cronice, întrebarea dacă aceste modificări persistă în abstinență rămâne în mare parte fără răspuns. Primatele non-umane reprezintă un model ideal în care să se evalueze efectele abstinenței asupra sistemului DA după expunerea cocaină cronică. În acest studiu, maimuțele rhesus de sex masculin autoadministrate de cocaină (0.3 mg/kg pe injecție, agenți de întărire 30 per sesiune) în cadrul unui interval fix cu 3-min schedule pentru zilele 100 urmate de abstinență 30 sau 90 zile. Această durată a autoadministrării cu cocaină a demonstrat anterior scăderea densităților receptorilor de tip DA D2 și creșterea nivelurilor de receptori asemănători cu D1 și transportoarele DA (DAT). Răspunsul la maimuțe de control a fost menținut prin prezentarea alimentelor în cadrul unui protocol identic și în aceleași perioade de abstinență. [3H]SCH 23390 se leagă la Receptorii DA D1 după zilele de abstinență 30 au fost semnificativ mai mari în toate porțiunile striatumului, comparativ cu animalele de control, în timp ce [3H]legarea raclopridei la receptorii DA D2 nu a fost diferită între grupuri. [3H]WIN 35 428 legat de DAT a fost, de asemenea, semnificativ mai mare pe parcursul practic al tuturor porțiunilor striaturii dorsale și ventrale după 30 zile de abstinență. După zilele de abstinență 90, totuși, nivelurile receptorilor DA D1 și DAT nu au fost diferite de valorile de control. Deși aceste rezultate indică faptul că există o eventuală recuperare a elementelor separate ale sistemului DA, ele evidențiază, de asemenea, natura dinamică a acestor componente în timpul fazelor inițiale de abstinență de la auto-administrarea cocainei cronice.
Cuvinte cheie:
cocaină, dopamină, autoradiografie, abstinență, striatum
INTRODUCERE
Utilizarea cocainei cronice în rândul persoanelor dependente de om a fost asociată cu neuroadaptări în sistemul dopaminic (DA) (Malison et al, 1998; Volkow et al, 1993, 1997). Acestea includ creșterea densității transportatorilor DA (DAT) și scăderea concentrațiilor de receptori de tip DA D2 (Mic et al, 1999; borceag et al, 2002; Volkow et al, 1993). În plus, s-au observat și alterarea eliberării DA. De exemplu, anchetatorii care utilizează tomografia cu emisie de pozitroni (PET) studiază cu [11C]racloprida și metilfenidatul au demonstrat scăderi ale eliberării DA în striatumul utilizatorilor cronici de cocaină (Volkow et al, 1997; Wong et al, 2006). O problemă însă este că adesea este dificil să excludem influența altor factori, cum ar fi utilizarea altor droguri ilegale și legale, diferențele în ceea ce privește aportul și utilizarea de droguri anterioare și diferențele în stilul de viață. Aceste diferențe, precum și existența unor condiții care pot duce la consumul de droguri, pot limita interpretarea studiilor la pacienții umani.
Modelele primate non-umane, în care variabilele pot fi manipulate sistematic, reprezintă o abordare alternativă a studiului consecințelor administrării cronice de cocaină și abstinenței ulterioare. Studiile anterioare au demonstrat că expunerea cocaină cronică este însoțită de scăderi semnificative ale concentrațiilor de receptori DA D2, precum și de creșterea nivelurilor receptorilor D1 și de densitatea DAT (Letchworth et al, 2001; Moore et al, 1998a, 1998b; Nader et al, 2002, 2006). Aceste efecte reflectă cele observate la om, dovedind astfel utilitatea acestor modele de expunere la medicament.
Deși există dovezi considerabile privind reglementarea sistemelor de DA, sa dovedit mai dificilă evaluarea dacă există dovezi de redresare după încetarea consumului de droguri (Malison et al, 1998; Jacobsen et al, 2000; Volkow et al, 1993) sau dacă aceste schimbări persistă dincolo de perioada de expunere continuă a cocainei. Din nou, modelele de primate non-umane pot oferi perspective în această fază a dependenței. Farfel et al (1992) a raportat scăderea concentrațiilor de receptori de tip DAT și D1 în striatum de maimuțe după abstinența de la expunerea cronică la riscul de neconcordanță. Cu toate acestea, rolul specific al abstinenței a fost dificil de determinat din cauza lipsei măsurătorilor într-un grup fără perioadă de abstinență. În mod similar, Melega et al (2008) a raportat scăderea semnificativă a nivelului de DAT în striatum de maimuțe vervet după săptămâni de abstinență 3 de la un regim de metamfetamină în creștere. Administrarea stimulentelor în ambele studii, cu toate acestea, nu era contrară. Calea de administrare (contingent vs noncontingent) sa dovedit a afecta creierul diferențiat atât în ceea ce privește eliberarea DA (Hemby et al, 1997) și metabolismul glucozei (Graham și Porrino, 1995; Porrino et al, 2002). Astfel, utilizarea autoadministrării în prezentul studiu eludează această problemă. În plus, efectele autoadministrării pe termen lung a cocainei asupra sistemelor DA cerebrale au fost extensiv studiate utilizând acest model de autoadministrare la maimuțele rhesus, oferind astfel o linie de bază pentru evaluarea neuroadaptărilor care apar în timpul abstinenței.
Scopul acestor studii a fost, prin urmare, de a determina dacă modificările în concentrațiile de receptori DAT și DA D1 și D2 care au fost demonstrate anterior la animale expuse la autoadministrarea cocaineiLetchworth et al, 2001; Moore et al, 1998a, 1998b; Nader et al, 2002) ar fi inversat după perioade îndelungate de abstinență. Pe baza studiilor efectuate la utilizatorii de droguri umane (cf. Volkow et al, 1993), am emis ipoteza că aceste schimbări în sistemul DA ar persista chiar și după 3 luni de abstinență. În acest scop, maimuțele auto-administrate cocaina pentru sesiunile 100, cu aportul total de 900 mg/kg, urmat de 30 sau 90 zile de abstinență de la medicament. Receptorii DA D1 și D2, precum și DAT, au fost măsurați cu cantitative in vitro receptor autoradiografie.
METODE
Subiecții
Un total de maimuțe rhesus masculi adulți proveniți din experiment 17 (Macaca mulatta) cântărind între 7.7 și 13 kg (media ± SD, 10.2 ± 1.32) la începutul studiului au servit ca subiecți. Toate procedurile au fost efectuate în conformitate cu practicile stabilite astfel cum sunt descrise în National Institutes of Health Ghid pentru îngrijirea și utilizarea animalelor de laborator. În plus, toate procedurile au fost revizuite și aprobate de Comitetul pentru îngrijirea și utilizarea animalelor de la Universitatea Wake Forest. Maimuțele erau adăpostite individual în cuști din oțel inoxidabil cu apă ad libitum; animalele au avut contact fizic și vizual unul cu celălalt. Greutatea corporală a acestora a fost menținută la aproximativ 90-95% a greutăților libere de hrănire cu peleți cu aromă de banană câștigați în timpul sesiunilor experimentale și prin hrănirea suplimentară cu Lab Diet Monkey Chow, cu condiția ca nu mai devreme de 30 min post-sesiune. În plus, li s-au dat fructe proaspete sau arahide de cel puțin trei ori pe săptămână. Fiecare maimuță a fost cântărită o dată pe săptămână, iar dacă este necesar, dietele au fost ajustate pentru a menține greutățile stabile.
Aparatură comportamentală
Sesiunile experimentale s-au desfășurat în camere ventilate și atenuate fonic (1.5 × 0.74 × 0.76 m; Med Associates Inc., East Fairfield, VT) concepute pentru a găzdui un scaun primat (Model R001; Primate Products, Redwood City, CA). Camera conținea un panou de informații (48 × 69 cm), care consta din două pârghii retractabile (5 cm lățime) și trei lumini de stimulare. Pârghiile erau poziționate la mică distanță de maimuță care stătea în scaunul primatului. Peletele alimentare cu aromă de banane (1 g; Bio-Serv, Frenchtown, NJ) au fost livrate de la un alimentator situat în partea superioară a camerei. O pompă de perfuzie peristaltică (7531-10; Cole-Parmer Co., Chicago, IL) a fost utilizată pentru a furniza injecții de droguri la o rată de aproximativ 1 ml pe 10 la acele animale care administrează cocaină. Funcționarea camerelor și achiziția de date a fost realizată cu un sistem informatic Power Macintosh cu o interfață (Med Associates Inc.).
Proceduri chirurgicale
Toate maimuțele, inclusiv martorii, au fost preparate chirurgical, în condiții sterile, cu catetere intravenoase și porturi de acces vascular (Model GPV, Access Technologies, Skokie, IL). Maimuțele au fost anesteziate cu o combinație de ketamină (15 mg/kg, im) și butorfanol (0.03 mg/kg, im) și sa făcut o incizie în apropierea venei femurale. După disecția și izolarea venei, capătul proximal al cateterului a fost introdus în venă pentru o distanță calculată pentru a se termina în vena cavă inferioară. Capătul distal al cateterului a fost filetat subcutanat la o incizie făcută ușor de pe linia mediană a spatelui. Portul de acces vascular a fost plasat într-un buzunar format printr-o disecție tangentă în apropierea acestei incizii. Maimuțele au primit 24-48 h înainte de a reveni la răspunsul armat cu alimente. Aproximativ 5 de zile înainte de procedura terminală, fiecare maimuță a fost implantată cu un cateter cronic în interiorul arterei femurale adiacente pentru colectarea probelor de sânge arteriale temporizate. Procedurile chirurgicale au fost identice cu cele descrise pentru cateterii venosi. În ziua sesiunii finale, a fost efectuat un studiu terminal de metabolizare a glucozei cerebrale în care maimuțele au fost injectate cu 2-[14C]deoxiglucoză (2-DG) aproximativ 2 min după sfârșitul sesiunii și probele de sânge au fost obținute prin cateterul arterial pe un 45 min (a se vedea Beveridge et al, 2006 pentru detalii). Datele metabolice din aceste studii nu sunt prezentate aici.
Proceduri de auto-administrare
Maimuțele au fost inițial instruite să răspundă pe una din cele două pârghii prin întărirea fiecărui răspuns pe pârghia corectă cu o peletă alimentară. Peste aproximativ o perioadă de 3 săptămâni intervalul dintre disponibilitatea de pelete de alimente a fost crescută treptat până la un interval de 3-min a fost atins (adică intervalul fix-program-3 min de armare; FI 3-min). În condițiile de programare finală, primul răspuns pe pârghie după 3 min a dus la livrarea unei pelete alimentare; ședințele s-au încheiat după prezentările de produse alimentare 30. La sfârșitul fiecărei sesiuni, pârghiile de răspuns au fost retrase, lumina de fundal și luminile de stimulare au fost stinse, iar animalele au rămas în camera întunecată pentru aproximativ 30 min înainte de a fi întorși în cuștile lor. Toate maimuțele au răspuns la programul de prezentare a alimentelor FI 3-min pentru cel puțin sesiuni 20 și până la obținerea unei performanțe stabile (± 20% din media timp de trei sesiuni consecutive, fără tendințe de rate de răspuns). Atunci când răspunde întreținut alimentare a fost stabil, alimentatorul a fost scos și efectele extincție asupra au fost examinate pentru care au raspuns cinci sesiuni consecutive, după care să răspundă a fost restabilit și întreținute de prezentarea produselor alimentare.
După ce a fost stabilită performanța inițială, toți maimuțele au fost preparate chirurgical cu catetere venoase, așa cum s-a descris mai sus, și repartizate aleatoriu într-una din cele trei grupuri. Un grup de maimuțe a servit drept control și a continuat să răspundă în cadrul programului de prezentare a alimentelor FI 3-min pentru un total de sesiuni 100 (N=6). Maimuțele 11 rămase au fost atribuite grupurilor de autoadministrare a cocainei (0.3 mg/kg pe injecție). Deoarece 0.3 mg/kg de cocaină pe injecție a fost considerată o doză mare pentru maimuțele anterioare cu cocaină, pentru majoritatea animalelor această doză a fost atinsă în două sesiuni, permițând mai întâi maimuței să se auto-administreze 0.1 mg/kg de cocaină. Întreținerea alimentară a fost lăsată să se stabilizeze după o intervenție chirurgicală (aproximativ 4-6 zile) înainte de începerea sesiunilor de auto-administrare a cocainei. Înainte de fiecare sesiune experimentală, spatele animalului a fost curățat cu 95% etanol și frecarea betadină și un indicator de 22 Huber Ac (model PG20-125) a fost introdus în portul care duce la cateter venos, conectarea unei pompe de perfuzie conținând soluția de cocaină la cateter. Înainte de începerea sesiunii, pompa a fost operată pentru aproximativ 3 s, umplând portul cu doza de cocaină disponibilă în timpul sesiunii experimentale. Sesiunile s-au încheiat după injecțiile 30; deoarece în condiții de control, maimuțele au rămas în camera întunecată pentru aproximativ 30 min. La sfârșitul fiecărei sesiuni, portul a fost umplut cu soluție salină heparinizată (100 U/ml) pentru a preveni coagularea.
Sesiunile experimentale s-au desfășurat aproximativ în aceeași zi în fiecare zi și au continuat pentru un total de sesiuni 100. În urma finalizării sesiunilor 100, a fost introdusă o perioadă de abstinență de 30 sau 90 zile în care catetere de timp au fost spălate zilnic cu soluție salină heparinizată, dar nici cocaina sau produsele alimentare sesiuni de autoadministrare au fost efectuate. Pentru grupul de control, perioadele de abstinență a zilelor 30 au fost impuse în patru animale și 90 zile în celelalte două. Pentru grupul de cocaină, au fost impuse perioade de abstinență în zilele 30 pe opt animale și 90 zile pe trei animale. La sfârșitul perioadei de abstinență a fost efectuată o sesiune de autoadministrare finală (control al alimentelor sau cocaină) și procedura 2-DG a fost inițiată imediat după sesiune. În două loturi de control și patru animale de autoadministrare a cocainei în grupul de abstinență 30, în sesiunea finală nu a fost primit cocaină. Animalele au fost omorâte cu supradozaj de pentobarbital (100 mg/kg, iv) la sfârșitul 45 min perioada de absorbție a trasorului.
Prelucrarea țesuturilor
După ucidere, creierul a fost imediat îndepărtat, blocat și înghețat în izopentan la -35 până la -55 ° C și apoi păstrat la -80 ° C. Blocurile de țesut care conțin striatum au fost apoi tăiate într-un criostat la -20 ° C în planul coronal în 20 μm, colectate pe diapozitive încărcate electrostatic, deshidratate sub vid peste noapte la 4 ° C, apoi depozitate la -80 ° C până la prelucrare pentru autoradiografie. Secțiunile creierului au fost colectate din porțiunile nucleului caudat, putamenului și nucleului accumbens care se situează între rostul anterior al comisiei anterioare. Această regiune este denumită striatum precommisural. Mai mult, nivelele rostral și caudal ale striatumului precommissional au fost desemnate cu referire la nucleul accumbens. Striatul preremiscular rostral este regiunea în care nucleul accumbens nu este diferențiat în subcomponente separate ale cochiliei și ale nucleului. Caudală striatum precommissural este regiunea congruentă cu aspectul cojii și miezul nucleului accumbens, care este posterior apariția tuberculul olfactiv. Pentru fiecare dintre studiile de legare, două secțiuni adiacente au fost luate la fiecare dintre cele cinci nivele (două rostrale și trei caudale) prin striatum precommisural pentru un total de secțiuni 10 pe animal.
Legarea receptorului D1
Densitățile locului de legare a receptorului DA D1 au fost determinate cu [3H]SCH 23390 (activitate specifică-85 Ci/mmol; PerkinElmer, Boston, MA) prin cantitative in vitro receptor de autoradiografie conform procedurilor adaptate de la Lidow et al (1991) și Nader et al (2002). Secțiunile au fost preincubate pentru 20 min în soluție tampon (50 mM Tris, 120 mM NaCI, 5 mM KCI, 2 mM CaCI2, 1 mM MgCI2, pH 7.4, 25 ° C) pentru a elimina DA endogen, cocaină și [14C] din procedura 2-DG. Secțiunile au fost apoi incubate pentru 30 min în același tampon, pH 7.4, 25 ° C, conținând 1 mM acid ascorbic, 40 nM ketanserin și 1 nM [3H]SCH 23390. După incubare, secțiunile au fost clătite de două ori pentru 20 s în tampon conținând 1 mM acid ascorbic la pH 7.4, 4 ° C, apoi se scurge în apă distilată la 4 ° C și se usucă sub un curent de aer rece. Legarea nespecifică a fost definită prin incubarea secțiunilor adiacente în soluția de incubare în prezența 5 μM (+) -Butaclamol. Secțiuni, împreună cu calibrate [3H] au fost aplicate standardelor autoradiografice (Amersham, Piscataway, NJ) pentru filmul Kodak Biomax MR (Fisher Scientific, Pittsburgh, PA) pentru săptămâni 6.
Legarea receptorului D2
Densitatea și distribuția situsurilor de legare a receptorului DA D2 au fost determinate cu [3H]racloprida (activitate specifică, 87 Ci/mmol; PerkinElmer) conform procedurilor adaptate de la Lidow et al (1991) și Nader et al (2002). Secțiunile au fost preincubate pentru 20 min în soluție tampon (50 mM Tris, 120 mM NaCI, 5 mM KCI, pH 7.4, 25 ° C) pentru a îndepărta DA endogen, cocaină și [14C] din procedura 2-DG. Diapozitivele au fost apoi incubate pentru 30 min în același buffer, conținând 5 mM acid ascorbic și 2 nM [3H]racloprid. Secțiunile au fost clătite 3 × 2 min în tampon la pH 7.4, 4 ° C, apoi se înmoaie în apă distilată la 4 ° C și se usucă sub un curent de aer rece. Legarea nespecifică a fost definită prin incubarea secțiunilor adiacente în soluția de incubare în prezența 1 μM (+) -Butaclamol. Secțiuni, împreună cu calibrate [3H] autoradiografice, au fost aplicate filmului Kodak Biomax MR pentru săptămâni 8.
Dopaminarea transportatorului dopaminei
Densitatea siturilor de legare DAT a fost determinată folosind [3H]WIN 35,428 (activitate specifică, 87 Ci/mmol; PerkinElmer) autoradiografie conform procedurilor adaptate de la Canfield et al (1990) și Letchworth et al (2001). Secțiunile de țesut au fost preincubate în tampon (50 mM Tris, 100 mM NaCI, pH 7.4, 4 ° C) pentru 20 min pentru a elimina orice DA rezidual, cocaină și [14C] din procedura 2-DG. Secțiunile au fost apoi incubate pentru 2 h la 4 ° C în același tampon care conține 5 nM [3H]WIN 35 428. Secțiunile au fost clătite pentru un total de 2 min în tampon la 4 ° C, apoi se înmoaie în apă distilată la 4 ° C și se usucă sub un curent de aer rece. Legarea nespecifică a fost definită prin incubarea secțiunilor adiacente în soluția de incubare în prezența 30 μM cocaină. Secțiuni, împreună cu calibrate [3H] autoradiografice, au fost aplicate filmului Kodak Biomax MR pentru săptămâni 6.
Densitometria și analiza datelor
Filmele au fost dezvoltate cu dezvoltatorul Kodak GBX, stopbath și Rapid Fixer (VWR, West Chester, PA) și apoi clătite. Analiza autoradiogramelor a fost efectuată prin densitometrie cantitativă cu un sistem computerizat de procesare a imaginii (MCID, Imaging Research, InterFocus Imaging Ltd, Cambridge, UK). Valorile densității optice au fost convertite în fmol/mg (de țesut cu greutate umedă) în funcție de calibrat [3H] standarde. Legarea specifică a fost determinată prin scăderea digitală a imaginilor de legare nespecifică din imaginile adiacente superioare ale legării totale. Structurile au fost identificate prin colorarea Nissl a secțiunilor adiacente acelora analizate pentru legarea receptorului. Datele din fiecare analiză au fost analizate independent printr-o analiză unică a varianței, urmată de diferențele dintre cele mai mici pătrate post hoc teste pentru comparații multiple. Fiecare regiune a constat într-o analiză separată. Deoarece datele obligatorii obținute de la animalele de control abstinente pentru zilele 30 și 90 nu au fost semnificativ diferite unul de celălalt, similar cu studiile anterioare (Nader et al, 2002), datele din grupurile de control au fost combinate. În plus, nu au existat diferențe semnificative între datele de la acele animale care au primit cocaină și cele care nu au avut loc la ultima lor sesiune, astfel încât datele din aceste grupuri au fost, de asemenea, combinate.
REZULTATE
Efectele abstinenței de la administrarea cocainei cronice pentru zilele 30
Concentrațiile de [3H]SCH 23390 care se leagă la receptorii DA D1 în striatum precommisural sunt arătați în Tabelul 1. Legarea specifică a [3H]SCH 23390 a reprezentat mai mult decât 90% din totalul obligatoriu. În conformitate cu rapoartele anterioare (Moore et al, 1998a; Nader et al, 2002), legarea lui [3H]SCH 23390 la receptorii D1 la animalele de control expuse non-medicament a fost eterogen cu diferențe apreciabile în gradul de legare între subregiuni ale striatumului. Etichetarea a fost mai densă în porțiunile mai mult rostrale și mediale pe întreg striatumul.

După zilele de 30 de abstinență din cauza expunerii la cocaină, legarea la receptorii D1 a fost caracterizată prin creșteri foarte răspândite pe întreaga extindere rostral-caudală a striatumului precommissional, comparativ cu densitățile de legare la animalele de control neexprimate (Tabelul 1; Figura 1). În striatumul mai rostral, concentrațiile au fost semnificativ mai mari în nucleul caudat, inclusiv în nivelul dorsolateral (+27%), central (+27%), dorsomedial (+27%) și ventromedial (+23%), precum și în partea dorsală (+17%), central (+22%) și ventral (+23%) porțiuni de putamen, în comparație cu densitățile animalelor controlate expuse non-medicament. Cresteri semnificative au fost de asemenea evidente in nucleul accumbens (+23%) la acest nivel. La nivelul striatumului în care miezul și coaja nucleului accumbens sunt cele mai diferențiate, densitățile receptorilor asemănători cu D1 au fost, de asemenea, semnificativ mai mari pe toată suprafața dorsolateralului (+31%), central (+29%), dorsomedial (+30%) și ventromedial (+18%) nucleu caudat, precum și nucleul dorsal (+23%), central (+29%) și ventral (+28%) putamen, comparativ cu densitățile controalelor expuse non-medicament. În interiorul striatumului ventral la acest nivel, concentrația de D1 receptori a fost mai mare în nucleul nucleus accumbens (+45%) și coajă (+20%), precum și în tuberculul olfactiv (+26%), comparativ cu densitățile controalelor.
Figura 1.
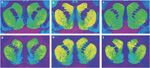 Autoradiograme reprezentative din [3H] SCH 23390 se leagă de receptorii D1 (panoul de sus) și [3H]WIN 35428 se leagă de transportoarele de dopamină (panou de fund) în secțiunile coronale ale striatum de maimuță rhesus. (a, d) Animalul de control care răspunde pentru armarea produselor alimentare. (b, e) Animalul de autoadministrare a cocainei cu abstinență de zile 30. (c, f) Animalul de auto-administrare a cocainei cu abstinență de zile 90.
Autoradiograme reprezentative din [3H] SCH 23390 se leagă de receptorii D1 (panoul de sus) și [3H]WIN 35428 se leagă de transportoarele de dopamină (panou de fund) în secțiunile coronale ale striatum de maimuță rhesus. (a, d) Animalul de control care răspunde pentru armarea produselor alimentare. (b, e) Animalul de autoadministrare a cocainei cu abstinență de zile 30. (c, f) Animalul de auto-administrare a cocainei cu abstinență de zile 90.
Concentrațiile de [3H]legarea raclopridei la receptorii DA D2 în striatum precommisual este prezentată în Tabelul 2. Legarea specifică cu [3H]racloprida a reprezentat mai mult decât 90% din totalul obligatoriu. Distribuția lui [3H]legarea raclopridei la receptorii D2 a fost, de asemenea, eterogenă în subregiunile striatului dorsal și ventral, ca în rapoartele precedente (Moore et al, 1998b; Nader et al, 2002). În controalele expuse non-medicament, în dorsal s-au prezentat concentrații mai mari de situsuri de legare D2 comparativ cu striatul ventral. În plus, au existat dovezi ale unui gradient medial spre lateral cu concentrații mai mari de situsuri de legare prezente în porțiunile mai laterale ale striatumului.
După zilele de abstinență 30, nivelurile de legare a receptorilor de D2 la animalele expuse cocainei și alimentele armate nu au fost semnificativ diferite între ele în cele mai multe regiuni ale striatumului. Au fost observate concentrații mai mari de situsuri de legare în putamen ventral (+10%) și nucleul anterior accumbens (+12%) în țesuturi de la maimuțele expuse cocainei comparativ cu martorii. Nu au fost observate alte diferențe semnificative.
Concentrațiile de [3H]WIN 35428 se leagă de DAT în striatum precommisural sunt prezentate în Tabelul 3. În conformitate cu rapoartele anterioare (Letchworth et al, 2001), legând la locurile DAT la animale expuse non-medicament a fost mai mare în dorsal în comparație cu striatum ventral. În interiorul nucleului accumbens, s-au observat densități mai mari în miez în comparație cu diviziunile cochiliei. În final, legarea nespecifică a reprezentat mai puțin decât 10% din total.
După zilele de 30 de abstinență de la expunerea la cocaină, legarea la DAT a fost semnificativ mai mare în majoritatea regiunilor striatumului rostral până la comisia anterioară în comparație cu legarea la animalele de control expuse non-medicamentoaseFigura 1). Concret, concentrațiile de situsuri de legare DAT la nivelele rostrale au fost semnificativ mai mari la nivel central (+22%), dorsomedial (+25%) și ventromedial (+28%) nucleu caudat și în zona dorsală (+16%) și central (+23%) putamen, comparativ cu controalele armate cu alimente. În plus, a existat, de asemenea, o legare semnificativ mai mare la DAT în nucleul anterior accumbens (+37%) în cocaină - comparativ cu maimuțele armate cu alimente. În cadrul mai multor porțiuni caudale ale striatumului precommissural, densitățile situsurilor de legare a DAT au fost semnificativ mai mari în nucleul caudat central (+21%) și putamen, central (+20%) și ventral (+19%; Figura 1). În interiorul striatumului ventral la acest nivel, legarea la DAT a fost semnificativ mai mare în nucleul nucleus accumbens (+20%) și tubercul olfactiv (+24%) în țesutul din cocaină- vs maimuțe armate cu alimente.
Efectele abstinenței de la administrarea cocainei cronice pentru zilele 90
Spre deosebire de diferențele foarte răspândite în densitatea situsurilor de legare la receptorii D1 observate la animalele expuse cocainei după zile de abstinență 30, după zilele de abstinență 90, nu au existat diferențe semnificative în comparație cu controalele armate cu alimente în orice porțiune a celulelor precommisurale striatum (Tabelul 1; Figura 1). În mod similar, concentrațiile de [3H]legarea de raclopridă la receptorii DA D2 în striatum precommisual după zilele de abstinență 90 nu au fost, de asemenea, semnificativ diferite de cele ale animalelor expuse non-medicament (Tabelul 2).
Concentrațiile de [3H]WIN Legarea 35428 la DAT a prezentat un model similar celui observat cu receptorii D1 și D2. Nu au existat diferențe semnificative în densitatea DAT între maimuțele expuse cocainei după zilele de abstinență 90 în comparație cu valorile din controalele expuse non-medicamentoase (Tabelul 3; Figura 1), deși trebuie remarcat faptul că există o tendință spre niveluri mai ridicate de legare în nucleul accumbens anterior.
DISCUŢIE
Studiile anterioare din partea grupului nostru au arătat că expunerea cronică la autoadministrarea cocainei este însoțită de o dysreglare semnificativă a sistemului DA al primatelor neumane (Letchworth et al, 2001; Moore et al, 1998a, 1998b; Nader et al, 2002, 2006). Rezultatele studiului prezent demonstrează că această dysreglementare rămâne evidentă după întreruperea expunerii la cocaină. După zilele de abstinență de la 30, concentrațiile receptorilor DA D1 și DAT au fost semnificativ crescute în întregul striat al maimuțelor cu istorii de auto-administrare cronică cronică în comparație cu controalele armate cu alimente. Cu toate acestea, studiul de față furnizează, de asemenea, dovezi clare privind recuperarea în cadrul sistemului DA după perioade mai lungi de abstinență (zile 90), evidențiate prin lipsa unor diferențe semnificative între expușii de cocaină și animalele controlate în acest moment. Aceste date sugerează că expunerea la cocaină nu poate produce modificări permanente în sistemul DA, dar că recuperarea poate apărea odată cu abstinența prelungită de la consumul de droguri.
Dizreglementarea concentrațiilor DAT prezentate aici după abstinență este în concordanță cu rapoartele anterioare din primatele neumane (Letchworth et al, 2001), care au prezentat creșteri semnificative în densitățile situsurilor de legare DAT atât în striatum ventral cât și dorsal. Deși nu au fost testate în mod explicit, se pare că, după încetarea expunerii la medicament, creșterile în densitatea situsului de legare DAT sunt cel puțin la fel de mari și sunt mai răspândite în toate regiunile striatum decât cele raportate fără nici o perioadă de așteptare (Letchworth et al, 2001). În mod similar, concentrațiile crescute ale locurilor de legare a receptorului D1 observate aici după zilele de abstinență 30 sunt, de asemenea, în concordanță cu studiile anterioare care au arătat densități de legare a receptorilor de tip D1 în striatum de primate neumane expuse la un regim identic de autoadministrare a cocaineiNader et al, 2002). În contrast, nu s-au observat diferențe semnificative între nivelele densităților de legare ale receptorilor de tip D2 în striatumul animalelor expuse cocainelor și al animalelor de control. Această lipsă de reglaj a fost prezentă, în ciuda scăderii mari a concentrațiilor de receptori D2 care au fost raportate la ambii dependenți umani (Volkow et al, 1993) și modele animale de autoadministrare a cocainei (Moore et al, 1998a, 1998b; Nader et al, 2002, 2006). Datele de față sugerează o normalizare mai rapidă pentru controlul nivelurilor în acest sistem în comparație cu receptorii tip D1 și DAT. Luate împreună, schimbările în receptorii DA și DAT arată în mod clar că perioada imediat următoare încetării autoadministrării cocainei este foarte instabilă, cu modificări considerabile în ceea ce privește reglementarea sistemului DA, dar că aceasta este urmată de re-reglementarea sistemul abordând o distribuție mai normală a receptorilor DA și DAT după o abstinență mai prelungită.
Modificările receptorului D1
Creșterile răspândite în densitatea receptorilor D1 observate aici după încetarea consumului de cocaină sunt în concordanță cu rapoartele care demonstrează o sensibilitate crescută a receptorilor D1 în timpul retragerii. Măsurarea sensibilității D1 a fost efectuată într - un studiu realizat de Henry și White (1991), în care s-a constatat că înregistrările de unități unice ale neuronilor din nucleul accumbens sunt mai sensibile la agonistul receptorului D1 SKF 38393 ca urmare a injectărilor cronice cronice zilnice în comparație cu martorii tratați cu soluție salină. Acest efect a fost persistent, deoarece sensibilitatea crescută a rămas evidentă până la o lună de retragere. Autorii au emis ipoteza că sensibilizarea receptorului D1 se datorează subsensibilității autoreceptorului D2 în zona somatodendritică A10, reducând astfel fluxul de impuls inhibitor în întreg sistemul mesoaccumbens DA (Henry și White, 1991). Datele din studiul de față sugerează creșterea abuzului de receptori de receptori D1, care ar putea explica această sensibilitate crescută a neuronilor dopaminergici la un agonist al receptorului D1 cu acțiune directă. Mai mult, efectul îmbunătățit al SKF 38393 asupra neuronilor nucleului accumbens nu a fost aparent la două luni după retragere, sugerând că a existat recuperarea sensibilității receptorului D1 (Henry și White, 1991); un rezultat consecvent cu recuperarea densităților receptorilor D1 observate în studiul de față după zile de abstinență 90. Alte rapoarte susțin, de asemenea, un rol important pentru receptorii D1 în recădere. Stimularea directă a receptorilor D1 în coaja nucleului accumbens poate readuce în căutarea cocainei în rozătoare abstinente (Schmidt et al, 2006). Cu toate acestea, literatura de specialitate este oarecum inconsecventă prin faptul că ambii agoniști și antagoniști D1 pot atenua căutarea de droguri provocată de primă de cocaină sau de stimuli de cocaină (Alleweireldt et al, 2002; De Vries et al, 1999; Khroyan et al, 2000; Auto et al, 1996; Alb et al, 2001). Recent, Khroyan et al (2003) a raportat că agoniștii și antagoniștii D1 reduc recaderea într-un model de primate non-umane de căutare a cocainei. Acești autori au sugerat că poate exista o gamă critică de activitate a receptorului D1 necesară pentru căutarea cocainei și că atât antagoniștii, cât și agonistul ar putea să-și schimbe activitatea din această fereastră. Concentrațiile crescute ale receptorilor D1 care însoțesc abstinența ar putea modifica acest interval, ducând la o modificare a sensibilității acestui sistem. O altă considerație este că activitatea D1 poate acționa pentru a modula activitatea la receptorii D2 (Nolan et al, 2007; Ruskin et al, 1999; Walters et al, 1987). Datele prezente sugerează că raportul dintre receptorii D1 și receptorii D2 se schimbă în cursul abstinenței și poate modifica astfel eficacitatea acestei modulații.
Deși, spre deosebire de datele actuale, s-au raportat scăderi ale nivelurilor de receptori D1 ca urmare a auto-administrării cocainei cronice (Moore et al, 1998a), există diferențe considerabile între aceste studii, cum ar fi doza și durata expunerii la cocaină, aportul total și grupurile de control comparativ. Luate împreună, prin urmare, dovezile convergente arată că sistemul D1 este în flux considerabil după retragerea din administrarea cocainei cronice.
Modificările transportatorului de dopamină
Constatările concentrațiilor crescute de DAT din striatul animalelor expuse cocainei după întreruperea consumului de droguri se extind pe cele din studiile noastre anterioare, care arată niveluri crescute de situsuri de legare a DAT care însoțesc autoadministrarea cocainei în primatele neumane. Datele actuale demonstrează că această dysreglementare persistă în timpul fazelor inițiale de abstinență. În plus, ele sugerează că valorile de recuperare la nivel de control urmează un curs de timp relativ lung (până la 90 zile în studiul de față). În studiile noastre anterioare am arătat că, deși inițial s-au limitat la regiunile striatale ventrale, schimbările în densitatea situsurilor de legare DAT s-au extins pentru a cuprinde mai multe porțiuni dorsale și rostrale ale striatumului cu perioade mai lungi de expunere la cocainăLetchworth et al, 2001; Porrino et al, 2004). În studiul de față revenirea la nivelurile de control ale concentrațiilor de DAT în timpul abstinenței părea să fie mai mare și mai rapidă în striatum dorsal decât în striatum ventral și astfel părea să urmeze o traiectorie anatomică inversă la modelul efectelor induse de expunerea cocaină cronică .
Datele actuale sunt, de asemenea, în concordanță cu rapoartele privind utilizatorii de cocaină umană (Mic et al, 1999; Malison et al, 1998; borceag et al, 2002; Staley et al, 1994) care au prezentat niveluri ridicate de legare la situsurile DAT din striat în comparație cu martorii, cu cele mai semnificative creșteri localizate în striatum ventral. Recent, aceste creșteri s-au dovedit a fi însoțite de scăderi semnificative ale legării 2 (VMAT2) a transportorului monoamin vezicular (Mic et al, 2003), sugerând o pierdere reală a neuronilor DA. Autorii au concluzionat că DAT ridicată a fost probabil datorată în mod direct unui răspuns compensator la blocarea farmacologică a cocainei, în timp ce scăderea VMAT2 reflectă cel mai probabil schimbările globale ale metabolismului DA, rezultând în funcția hipodopaminergică.
Persoanele dependente de cocaină umană au raportat o scădere a concentrațiilor de DA în striatum ventral, măsurată cu PET, ca răspuns la provocarea cu metilfenidat în comparație cu martorii sănătoși (Volkow et al, 1997). Recent, Martinez et al (2007) a raportat că utilizatorii de cocaină au avut un răspuns blunt la o provocare de amfetamină din striatum ventral și putamen. Mai mult, această scădere a eliberării DA provocată de amfetamină a fost corelată cu alegerea pentru cocaină în sesiuni de autoadministrare separate, astfel încât acei utilizatori cu cel mai scăzut grad de eliberare DA ca răspuns la amfetamină au fost cel mai probabil să aleagă cocaina pe un agent de întărire alternativMartinez et al, 2007). Studiile recente efectuate pe modele de rozătoare de consum de cocaină susțin această idee. Mateo et al (2005), de exemplu, a raportat că expunerea la auto-administrarea cocaină cronică este asociată cu alterarea funcției DAT. Acești investigatori au arătat că absorbția inițială a DA a fost crescută, ducând la o mai rapidă clearance-ul DA sinaptic și, prin urmare, scăderea nivelelor bazale ale DA extracelulare sau a unei stări hipodopaminergice. Astfel, este probabil ca concentrațiile crescute ale DAT după retragerea din administrarea cocainei cronice, observate în studiul prezent, să reprezinte un răspuns compensator, ceea ce duce la scăderea nivelurilor inițiale ale DA extracelulare.
Modificările receptorului D2
Un rezultat al studiului prezentat a fost acela că concentrațiile de niveluri ale receptorilor D2 s-au reîntors la valorile de control după zilele de abstinență 30, comparativ cu scăderile semnificative observate la animalele fără perioadă de așteptare (Nader et al, 2002). Spre deosebire de prezenta anchetă, studiile privind imagistica umană au constatat, în general, că nivelurile receptorilor D2 sunt mai scăzute decât cele ale controalelor după o abstinență prelungită de la expunerea cocaină cronică (Martinez et al, 2004; Volkow et al, 1993). Explicațiile potențiale ale diferențelor dintre aceste studii umane și ancheta prezenței primate neumane în prezent includ diferențe în modelul și durata consumului de cocaină, precum și posibilitatea existenței unor niveluri mai scăzute de receptori D2 la dependenții umani.
În concordanță cu această din urmă idee, există dovezi care sugerează că nivelurile bazale inferioare ale receptorilor D2 la oamenii sănătoși prezic o creștere a eficacității de întărire a stimulanților, cum ar fi methylphenidatși (Volkow et al, 1999) și, similar, la maimuțe, nivelele de bază ale receptorilor D2 au prezis propensitatea de autoadministrare a cocainei (Morgan et al, 2002; Nader et al, 2006). În concordanță cu natura paralelă a acestor constatări în rândul speciilor, atât cele umane (Volkow et al, 1993) și primat non-uman (Nader et al, 2006) studiile imagistice au demonstrat niveluri mai scăzute de disponibilitate a receptorilor D2 după abstinența de la expunerea la cocaină. În special, programul de autoadministrare a cocainei în ultimul experiment (Nader et al, 2006) a fost similar cu programul de armare utilizat în acest studiu. Astfel, este puțin probabil ca rezultatele diferite ale acestor două studii să se fi datorat unor diferențe metodologice, cum ar fi programul de întărire sau de absorbție cumulativă în timpul autoadministrării cocainei.
O explicație mai probabilă implică dinamica funcțională a sistemului DA. Măsurile privind disponibilitatea receptorului D2 cu PET au fost descrise ca fiind "funcționale", deoarece semnalul este legat de cantitatea de proteină (în acest caz densitățile receptorilor D2) și de nivelurile de neurotransmițător circulant (A se vedea Laruelle, 2000; Nader și Czoty, 2008 pentru discuții ulterioare). eun contrast, autoradiografia receptorului este necontaminată prin nivelurile de circulație ale DA. Astfel, studiul prezent, împreună cu activitatea noastră anterioară, sugerează că densitățile receptorilor D2 sunt scăzute prin administrarea cocainei, dar nivelul receptorului pare să se redreseze în timpul abstinenței. Într-un studiu realizat în mod similar în PET (Nader et al, 2006), recuperarea a fost observată la trei din cinci maimuțe. Constatările prezente sugerează că acești maimuțe nu au fost probabil diferiți în densitățile receptorilor D2, dar probabil diferența dintre sistemul de DA (adică nivelurile DA care circulă în timpul abstinenței) a fost diferențiat între subiecții "recuperați" și "nerecuperați".
Limitări
O limitare importantă a studiilor actuale este că studiile noastre nu pot aborda funcționalitatea receptorilor D1 și D2 sau a DAT. Mai degrabă am examinat doar modificările în densitatea proteinelor receptorilor. Deși rezultatele au implicații asupra rolurilor potențiale ale acestor sisteme, vor fi necesare studii suplimentare pentru a arăta consecințele comportamentale ale schimbărilor prezentate aici. O altă limitare a studiului prezent este că liganzii autoradiografici nu reușesc adesea să disocieze între locurile intracelulare și citoplasmatice ale țintelor lor, deoarece antagoniștii radiomarcați sunt adesea permeabili la membrană. Mic et al (2002) a demonstrat că expunerea cronică la cocaină duce la o reglaj semnificativ a DAT la suprafața membranei, concomitent cu o scădere a concentrației DAT intracelulare, în celulele transfectate. Recent, Samuvel et al (2008) a raportat o descoperire similară în preparatele sinaptozomale striatale de șobolan. Aceste rezultate sugerează că modificările distribuției DAT observate în studiul de față pot reprezenta schimbări la nivelul suprafeței membranei, mai degrabă decât în locurile intracelulare.
În cele din urmă, trebuie acordată o atenție deosebită interpretării studiilor privind abstinența prelungită (90 zile), deoarece aceste constatări se bazau pe un grup relativ mic de animale (N=3). În ciuda numărului redus de subiecți, datele obținute de la acest grup au fost mai degrabă consecvente, așa cum se poate observa într-un grafic de scatter prezentat în Figura 2. Concentrația situsurilor de legare a receptorilor D1 pe striat a arătat o variabilitate redusă în cadrul grupurilor, sugerând fiabilitatea acestor constatări. Consistența similară a fost, de asemenea, evidentă în datele obținute din analizele de legare a receptorului D2 și DAT. Deși trebuie prudentă, aceste date sugerează că restaurarea concentrațiilor de receptori DAT și DA în striat poate avea loc cu abstinență prelungită.
Figura 2.
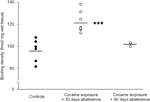 Densitățile de legare a receptorilor D1 pentru animalele individuale au fost medii pe întregul striat, în urma unei armări alimentare (controale) sau a abstinenței 30 sau 90 de zile de la autoadministrarea cocainei cronice. Mijloacele pentru grupuri sunt reprezentate cu bare negre, ***p<0.001 comparativ cu controalele întărite cu alimente.
Densitățile de legare a receptorilor D1 pentru animalele individuale au fost medii pe întregul striat, în urma unei armări alimentare (controale) sau a abstinenței 30 sau 90 de zile de la autoadministrarea cocainei cronice. Mijloacele pentru grupuri sunt reprezentate cu bare negre, ***p<0.001 comparativ cu controalele întărite cu alimente.
CONCLUZII
În concluzie, expunerea la autoadministrarea cocainei a produs modificări semnificative în reglementarea sistemelor DA care au persistat în fazele timpurii (primele 30 zile) de abstinență. Acest lucru a fost cel mai evident în reglementarea concentrației receptorilor D1 și DAT, atât în ceea ce privește amploarea lor de modificări, cât și gradul lor topografic. În contrast, au existat dovezi de normalizare cu durate mai lungi de abstinență față de expunerea la cocaină, deoarece concentrațiile de receptori DAT, D1 și D2 după zilele de abstinență 90 nu au fost diferite de cele ale controalelor expuse non-medicament. TTotuși, aceste sisteme nu urmăresc neapărat același curs temporal de recuperare, sugerând că este probabil să existe o anumită instabilitate în reglementarea nivelurilor DA, în special la începutul abstinenței. Această disreglementare dopaminergică poate influența eficacitatea oricărei posibile farmacoterapii administrate dependenților abstinenți de cocaină, în special dacă medicamentul se bazează pe sistemul DA pentru mecanismul său de acțiune.
notițe
DEZVĂLUIRE/CONFLICTUL DE INTERES
Autorii nu au nici un conflict de interese de dezvăluit.
Referinte
- Alleweireldt AT, Weber SM, Kirschner KF, Bullock BL, Neisewander JL (2002). Blocarea sau stimularea receptorilor de dopamină D1 atenuează reintroducerea comportamentului căutător de cocaină stins în șobolani. Psihofarmacologie (Berl) 159: 284–293. | Articol | PubMed | ChemPort |
- Beveridge TJ, Smith HR, Daunais JB, Nader MA, Porrino LJ (2006). Administrarea cocainei cronice este asociată cu o activitate funcțională modificată în lobii temporali ai primatelor neumane. Eur J Neurosci 23: 3109–3118. | Articol | PubMed |
- Canfield DR, Spealman RD, Kaufman MJ, Madras BK (1990). Localizarea autoradiografică a siturilor de legare a cocainei prin [3H] CFT ([3H] WIN 35428) în creierul maimuței. Synapse 6: 189–195. | Articol | PubMed | ChemPort |
- De Vries TJ, Schoffelmeer AN, Binnekade R, Vanderschuren LJ (1999). Mecanisme dopaminergice care stimulează stimularea cocainei și a heroinei după retragerea pe termen lung a autoadministrării medicamentelor IV. Psihofarmacologie (Berl) 143: 254–260. | Articol | PubMed | ChemPort |
- Farfel GM, Kleven MS, Woolverton WL, Seiden LS, Perry BD (1992). Efectele injecțiilor repetate de cocaină asupra situsurilor de legare a receptorilor de catecolamină, a locurilor de legare a transportatorului de dopamină și a comportamentului la maimuța rhesus. Brain Res 578: 235–243. | Articol | PubMed | ChemPort |
- Graham J, Porrino LJ (1995). Substraturi neuroanatomice ale autoadministrării cocainei. CRC: Boca Raton, FL.
- Hemby SE, Co C, Koves TR, Smith JE, Dworkin SI (1997). Diferențele în concentrațiile de dopamină extracelulare în nucleul accumbens în timpul administrării de cocaină dependentă de răspuns și independent de răspuns la șobolan. Psihofarmacologie (Berl) 133: 7–16. | Articol | PubMed | ChemPort |
- Henry DJ, alb FJ (1991). Administrarea repetată de cocaină determină o creștere persistentă a sensibilității receptorului dopaminei D1 în nucleul accumbens de șobolan. J. Pharmacol Exp Ther 258: 882–890. | PubMed | ChemPort |
- Jacobsen LK, Staley JK, Malison RT, Zoghbi SS, Seibyl JP, Kosten TR et al (2000). A crescut capacitatea de legare a transportorului serotoninei centrale la pacienții dependenți de cocaină abstinenți acută. Am J Psihiatrie 157: 1134–1140. | Articol | PubMed | ISI | ChemPort |
- Khroyan TV, Barrett-Larimore RL, Rowlett JK, Spealman RD (2000). Dopaminele D1-și D2-like mecanisme de receptori în recadere la comportamentul care caută cocaina: efectele antagoniștilor selectivi și agoniști. J. Pharmacol Exp Ther 294: 680–687. | PubMed | ISI | ChemPort |
- Televiziunea Khroyan, Platt DM, Rowlett JK, Spealman RD (2003). Atenuarea recidivei la cocaina caută agoniști și antagoniști ai receptorilor de dopamină D1 în primate neumane. Psihofarmacologie (Berl) 168: 124–131. | Articol | PubMed | ChemPort |
- Laruelle M (2000). Imagistica neurotransmisiei sinaptice cu in vivo tehnici de concurență obligatorii: o revizuire critică. J Cereb Metab de flux sanguin 20: 423–451. | Articol | PubMed | ISI | ChemPort |
- Letchworth SR, Nader MA, Smith HR, Friedman DP, Porrino LJ (2001). Progresia schimbărilor în densitatea situsului de legare a transportatorului de dopamină ca urmare a autoadministrării cocainei la maimuțele rhesus. J Neurosci 21: 2799–2807. | PubMed | ISI | ChemPort |
- Lidow MS, Goldman-Rakic PS, Gallager DW, Rakic P (1991). Distribuția receptorilor dopaminergici în cortexul cerebral al primatelor: analiza autoradiografică cantitativă utilizând racloprida [3H], [3H] spiperona și [3H] SCH 23390. Neuroştiinţe 40: 657–671. | Articol | PubMed | ChemPort |
- Little KY, Elmer LW, Zhong H, Scheys JO, Zhang L (2002). Inducerea de cocaină a traficului de transportator de dopamină către membrana plasmatică. Mol Pharmacol 61: 436–445. | Articol | PubMed | ISI | ChemPort |
- Little KY, Krolewski DM, Zhang L, Cassin BJ (2003). Pierderea proteinei transportoare monoamină striatală veziculară (VMAT2) la utilizatorii de cocaină umană. Am J Psihiatrie 160: 47–55. | Articol | PubMed |
- Micul KY, Zhang L, Desmond T, Frey KA, Dalack GW, Cassin BJ (1999). Anomalii stomatologice dopaminergice la utilizatorii de cocaină umană. Am J Psihiatrie 156: 238–245. | PubMed | ChemPort |
- Malison RT, Cel mai bun SE, van Dyck CH, McCance EF, Wallace EA, Laruelle M et al (1998). Creșteri ale transportorilor de dopamină striatali în timpul abstinenței acute de cocaină măsurată prin [123I] beta-CIT SPECT. Am J Psihiatrie 155: 832–834. | PubMed | ChemPort |
- Martinez D, Broft A, Foltin RW, Slifstein M, Hwang DR, Huang Y et al (2004). Dependența de cocaină și disponibilitatea receptorului d2 în subdiviziunile funcționale ale striatumului: relația cu comportamentul care caută cocaina. Neuropsychopharmacology 29: 1190–1202. | Articol | PubMed | ISI | ChemPort |
- Martinez D, Narendran R, Foltin RW, Slifstein M, Hwang DR, Broft A et al (2007). Eliberarea de dopamină indusă de amfetamină: puternic afectată de dependența de cocaină și predicția alegerii de autoadministrare a cocainei. Am J Psihiatrie 164: 622–629. | Articol | PubMed | ISI |
- Mash DC, Pablo J, Ouyang Q, Hearn WL, Izenwasser S (2002). Funcția de transport al dopaminei este crescută la utilizatorii de cocaină. J Neurochem 81: 292–300. | Articol | PubMed | ChemPort |
- Mateo Y, Lipsa CM, Morgan D, Roberts DC, Jones SR (2005). Reducerea funcției terminale a dopaminei și insensibilitatea față de cocaină după administrarea de cocaină și deprivarea. Neuropsychopharmacology 30: 1455–1463. | Articol | PubMed | ISI | ChemPort |
- Melega WP, Jorgensen MJ, Lacan G, Way BM, Pham J, Morton G et al (2008). Administrarea pe termen lung a metamfetaminei în modelele de maimuțe vervet ale unei expuneri umane: neurotoxicitatea creierului și profilele comportamentale. Neuropsychopharmacology 33: 1441–1452. | Articol | PubMed | ChemPort |
- Moore RJ, Vinsant SL, Nader MA, Porrino LJ, Friedman DP (1998a). Efectul autoadministrării cocainei asupra receptorilor D1 ai dopaminei striatale la maimuțele rhesus. Synapse 28: 1–9. | Articol | PubMed | ChemPort |
- Moore RJ, Vinsant SL, Nader MA, Porrino LJ, Friedman DP (1998b). Efectul autoadministrării cocainei asupra receptorilor dopaminergici D2 la maimuțele rhesus. Synapse 30: 88–96. | Articol | PubMed | ChemPort |
- Morgan D, Grant KA, Gage HD, Mach RH, Kaplan JR, Prioleau O et al (2002). Dominanța socială la maimuțe: receptorii dopaminei D2 și autoadministrarea cocainei. Nat Neurosci 5: 169–174. | Articol | PubMed | ISI | ChemPort |
- Nader MA, Czoty PW (2008). Imagistica creierului în primatele non-umane: perspective asupra dependenței de droguri. ILAR J 49: 89–102. | PubMed | ChemPort |
- Nader MA, Daunais JB, Moore T, Nader SH, Moore RJ, Smith HR et al (2002). Efectele autoadministrării cocainei asupra sistemelor de dopamină striatală la maimuțele rhesus: expunerea inițială și cronică. Neuropsychopharmacology 27: 35–46. | Articol | PubMed | ISI | ChemPort |
- Nader MA, Morgan D, Gage HD, Nader SH, Calhoun TL, Buchheimer N et al (2006). Imagistica PET a receptorilor de dopamină D2 în timpul administrării cronice de cocaină la maimuțe. Nat Neurosci 9: 1050–1056. | Articol | PubMed | ChemPort |
- Nolan EB, Harrison LM, Lahoste GJ, Ruskin DN (2007). Sinergismul comportamental între receptorii D (1) și D (2) ai dopaminei la șoareci nu depinde de joncțiunile gap. Synapse 61: 279–287. | Articol | PubMed | ChemPort |
- Porrino LJ, Lyons D, Miller MD, Smith HR, Friedman DP, Daunais JB et al (2002). Cartografierea metabolică a efectelor cocainei în timpul fazelor inițiale de autoadministrare în primatul non-uman. J Neurosci 22: 7687–7694. | PubMed | ChemPort |
- Porrino LJ, Lyons D, Smith HR, Daunais JB, Nader MA (2004). Administrarea de cocaină autoadministrează o implicare progresivă a domeniilor striate, limbic, de asociere și senzorimotor. J Neurosci 24: 3554–3562. | Articol | PubMed | ISI | ChemPort |
- Ruskin DN, Bergstrom DA, Walters JR (1999). Multitudine de oscilații în rata de ardere în globus pallidus: modularea sinergică a receptorilor de dopamină D1 și D2. J. Pharmacol Exp Ther 290: 1493–1501. | PubMed | ISI | ChemPort |
- Samuvel DJ, Jayanthi LD, Manohar S, Kaliyaperumal K, vezi RE, Ramamoorthy S (2008). Dysregularea traficului de transportator de dopamină și a funcției după abstinența de la autoadministrarea cocainei la șobolani: dovezi pentru o reglementare diferențiată în caudate putamen și nucleus accumbens. J. Pharmacol Exp Ther 325: 293–301. | Articol | PubMed | ChemPort |
- Schmidt HD, Anderson SM, Pierce RC (2006). Stimularea receptorilor de dopamină D1 sau D2 în cochilie, dar nu și în nucleul nucleului accumbens, reintroduce comportamentul cocainei în șobolan. Eur J Neurosci 23: 219–228. | Articol | PubMed |
- Self DW, Barnhart WJ, Lehman DA, Nestler EJ (1996). Modularea opusă a comportamentului căutător de cocaină de către agoniștii receptorilor de dopamină D1 și D2. Ştiinţă 271: 1586–1589. | Articol | PubMed | ISI | ChemPort |
- Staley JK, Hearn WL, Ruttenber AJ, Wetli CV, Mash DC (1994). Site-urile de recunoaștere a cocainei cu afinitate ridicată pe transportorul de dopamină sunt ridicate în cazul victimelor de supradozaj cu cocaină fatală. J. Pharmacol Exp Ther 271: 1678–1685. | PubMed | ChemPort |
- Volkow ND, Fowler JS, Wang GJ, Hitzemann R, Logan J, Schlyer DJ et al (1993). Scăderea disponibilității receptorilor de dopamină D2 este asociată cu un metabolism frontal redus la persoanele care abuză de cocaină. Synapse 14: 169–177. | Articol | PubMed | ISI | ChemPort |
- Volkow ND, Wang GJ, Fowler JS, Logan J, Gatley SJ, Gifford A et al (1999). Predictii de consolidare a raspunsurilor la psihostimulanti la om prin nivelurile receptorilor D2 de dopamina creierului. Am J Psihiatrie 156: 1440–1443. | PubMed | ISI | ChemPort |
- Volkow ND, Wang GJ, Fowler JS, Logan J, Gatley SJ, Hitzemann R et al (1997). S-a redus receptivitatea dopaminergică striatală la subiecții dependenți de cocaină. Natură 386: 830–833. | Articol | PubMed | ISI | ChemPort |
- Walters JR, Bergstrom DA, Carlson JH, Chase TN, Braun AR (1987). Activarea receptorului de dopamină D1 necesară pentru exprimarea postsynaptică a efectelor agoniste D2. Ştiinţă 236: 719–722. | Articol | PubMed | ChemPort |
- Weiss F, Martin-Fardon R, Ciccocioppo R, Kerr TM, Smith DL, Ben-Shahar O (2001). Rezistența durabilă la dispariția comportamentului de căutare a cocainei indus de indiciile legate de consumul de droguri. Neuropsychopharmacology 25: 361–372. | Articol | PubMed | ISI | ChemPort |
- Wong DF, Kuwabara H, Schretlen DJ, Bonson KR, Zhou Y, Nandi A et al (2006). Creșterea gradului de ocupare a receptorilor dopaminergici în striatum uman în timpul poftei de cocaină provocată de către consumatori. Neuropsychopharmacology 31: 2716–2727. | Articol | PubMed | ChemPort |