Dev Rev. 2008; 28(1): 62-77. doi: 10.1016 / j.dr.2007.08.003
abstraktné
Dospievanie je vývojové obdobie charakterizované suboptimálnymi rozhodnutiami a činmi, ktoré vedú k zvýšenému výskytu neúmyselných zranení a násilia, zneužívaniu alkoholu a drog, neúmyselnému tehotenstvu a pohlavne prenosným chorobám. Tradičné neurobiologické a kognitívne vysvetlenia správania adolescentov nezohľadnili nelineárne zmeny v správaní pozorované počas dospievania v porovnaní s detstvom a dospelosťou. Tento prehľad poskytuje biologicky prijateľnú konceptualizáciu neurálnych mechanizmov, ktoré sú základom týchto nelineárnych zmien v správaní, ako zvýšenú citlivosť na stimuly, zatiaľ čo kontrola impulzov je v tomto období stále relatívne nezrelá. Nedávne zobrazovanie na ľuďoch a štúdie na zvieratách poskytujú biologický základ pre tento pohľad, čo naznačuje rozdielny vývoj systémov limbických odmien v porovnaní s kontrolnými systémami zhora nadol v období adolescencie v porovnaní s detstvom a dospelosťou. Tento vývojový model môže byť exacerbovaný u adolescentov s predispozíciou k riskovaniu, čím sa zvyšuje riziko zlých výsledkov.
Podľa Národného centra pre zdravotnú štatistiku sú v Spojených štátoch každoročne úmrtia adolescentov 13,000. Približne 70% týchto úmrtí je dôsledkom havárie motorového vozidla, neúmyselného zranenia, vraždy a samovraždy (Eaton a kol., 2006). Výsledky prieskumu Národného mládežníckeho prieskumu rizika (YRBS) spoločnosti 2005 ukazujú, že adolescenti sa zapájajú do správania, ktoré zvyšuje ich pravdepodobnosť úmrtia alebo choroby riadením vozidla po pití alebo bez bezpečnostného pásu, prevozu zbraní, používania nelegálnych látok a zapájania sa do nechráneného sexu. tehotenstva a pohlavne prenosných chorôb vrátane infekcie HIV (\ tEaton a kol., 2006). Tieto štatistiky podčiarkujú význam chápania rizikových rozhodnutí a činností u adolescentov.
Predpokladá sa množstvo kognitívnych a neurobiologických hypotéz, prečo sa adolescenti angažujú v suboptimálnom správaní. V nedávnom prehľade literatúry o vývoji ľudského adolescentného mozgu, Yurgelun-Todd (2007) naznačuje, že kognitívny vývoj v adolescentných rokoch je spojený s progresívne vyššou efektivitou kognitívnych kontrolných kapacít. Táto účinnosť je opísaná ako závislá na dozrievaní prefrontálneho kortexu, čoho dôkazom je zvýšená aktivita v ohniskových prefrontálnych oblastiach (Rubia a kol., 2000; Tamm, Menon a Reiss, 2002) a znížená aktivita v irelevantných oblastiach mozgu (\ tBrown et al., 2005; Durston a kol., 2006).
Tento všeobecný model zlepšenej kognitívnej kontroly s dozrievaním prefrontálneho kortexu naznačuje lineárny nárast vývoja od detstva do dospelosti. Napriek tomu suboptimálne voľby a akcie pozorované počas adolescencie predstavujú nelineárnu zmenu v správaní, ktorá sa dá odlíšiť od detstva a dospelosti, o čom svedčí Národné centrum pre zdravotnú štatistiku o adolescentnom správaní a úmrtnosti. Ak bola kognitívna kontrola a nezrelá prefrontálna kôra základom pre suboptimálne správanie pri výbere, potom by deti mali vyzerať pozoruhodne podobné alebo dokonca horšie ako dospievajúci, vzhľadom na ich menej vyvinutú prefrontálnu kôru a kognitívne schopnosti. Samotná nezrelá prefrontálna funkcia teda nemôže zodpovedať za adolescentné správanie.
Presná koncepcia kognitívnych a neurobiologických zmien v období adolescencie musí liečiť adolescenciu ako prechodné vývojové obdobie (Spear, 2000) namiesto jednej snímky v čase (Casey, Tottenham, Liston a Durston, 2005). Inými slovami, na pochopenie tohto vývojového obdobia sú pre rozlišovanie odlišných atribútov tohto štádia vývoja potrebné prechody do adolescencie az neho. Stanovenie vývojových trajektórií pre kognitívne a nervové procesy je nevyhnutné pre charakterizáciu týchto prechodov a obmedzenie interpretácií zmien v správaní počas tohto obdobia. Na kognitívnej alebo behaviorálnej úrovni sú adolescenti charakterizovaní ako impulzívni (tj chýba im kognitívna kontrola) a riskujú, že tieto konštrukty sa používajú synonymne a bez uznania odlišných vývojových trajektórií každého z nich. Na neurobiologickej úrovni, zobrazovanie na ľuďoch a štúdie na zvieratách poukazujú na odlišné neurobiologické bázy a vývojové trajektórie pre neurálne systémy, ktoré sú základom týchto samostatných konštruktov riadenia impulzov a rizikových rozhodnutí.
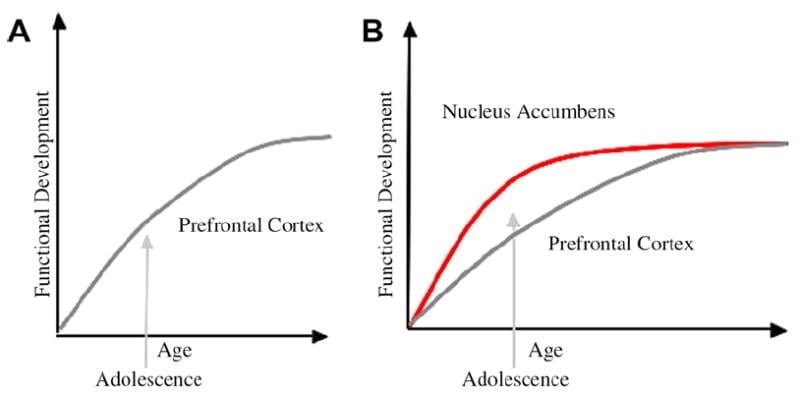
V tomto rámci sme vyvinuli neurobiologický model vývoja adolescentov, ktorý vychádza z modelov hlodavcov (Laviola, Adriani, Terranova a Gerra, 1999; Spear, 2000) a nedávne zobrazovacie štúdie adolescencie (Ernst a spol., 2005; Galvan, Hare, Voss, Glover a Casey, 2007; Galvan a kol., 2006). Obr. 1 Tento model je znázornený nižšie. Na ľavej strane je tradičná charakteristika adolescencie, ktorá sa vzťahuje takmer výlučne na nezrelosť prefrontálneho kortexu. Vpravo je náš navrhovaný neurobiologický model, ktorý ilustruje, ako sa musia posudzovať limbické subkortikálne a prefrontálne oblasti zhora nadol. Karikatúra ilustruje rôzne vývojové trajektórie pre tieto systémy, pričom limbické systémy sa vyvíjajú skôr ako regióny prefrontálnej kontroly. Podľa tohto modelu je jedinec viac ovplyvnený funkčne zrelými limbickými oblasťami v období adolescencie (tj nerovnováha limbickej v porovnaní s predfrontálnou kontrolou) v porovnaní s deťmi, pre ktoré sa tieto systémy (tj limbické a prefrontálne) stále vyvíjajú; v porovnaní s dospelými, pre ktorých sú tieto systémy plne zrelé. Táto perspektíva poskytuje základ pre nelineárne posuny v správaní naprieč vývojom v dôsledku skoršieho dozrievania tejto limbickej relatívne k menej vyspelej top-down prefrontálnej kontrolnej oblasti. S rozvojom a skúsenosťami, funkčné prepojenie medzi týmito regiónmi poskytuje mechanizmus na kontrolu zhora nadol v týchto regiónoch (Hare, Voss, Glover a Casey, 2007a). Okrem toho model zosúlaďuje rozpor medzi zdravotnými štatistikami rizikového správania počas dospievania s pozorným pozorovaním Reyna a Farley (2006) že dospievajúci sú schopní rozumieť a chápať riziká správania, v ktorých sa angažujú. Podľa nášho modelu, v emocionálne vystupujúcich situáciách, limbický systém vyhrá nad riadiacimi systémami vzhľadom na jeho zrelosť v porovnaní so systémom prefrontálneho riadenia. Dôkazy z behaviorálnych a ľudských zobrazovacích štúdií na podporu tohto modelu sa poskytujú v kontexte akcií v odmeňujúcich a emocionálnych kontextoch (Galvan a kol., 2006, 2007; Hare, Voss, Glover a Casey, 2007b; Hare a kol., 2007a). Okrem toho špekulujeme o tom, prečo sa mozog môže vyvíjať týmto spôsobom a prečo niektorí tínedžeri môžu byť vystavení väčšiemu riziku, že urobia suboptimálne rozhodnutia vedúce k horším dlhodobým výsledkom (Galvan a kol., 2007; Hare a kol., 2007b).
Predpokladá sa, že tradičné vysvetlenie správania adolescentov je spôsobené predĺženým vývojom prefrontálneho kortexu (A). Náš model berie do úvahy vývoj prefrontálneho kortexu spolu s subkortikálnymi limbickými oblasťami (napr. Nucleus accumbens), ktoré sa podieľajú na rizikových voľbách a akciách (B).
Vývoj cieleného správania
Základným kameňom kognitívneho vývoja je schopnosť potlačiť nevhodné myšlienky a činy v prospech cieľovo orientovaných myšlienok, najmä v prítomnosti presvedčivých stimulov (Casey, Galvan a Hare, 2005; Casey a kol., 2000b; Casey, Thomas, David-son, Kunz, & Franzen, 2002a; Casey, Tottenham a Fossella, 2002b). Niekoľko klasických vývojových štúdií ukázalo, že táto schopnosť sa vyvíja počas detstva a dospievania (Case, 1972; Flavell, Feach a Chinsky, 1966; Keating a Bobbitt, 1978; Pascual-Leone, 1970). Niekoľko teoretikov tvrdilo, že kognitívny vývoj je spôsobený zvýšením rýchlosti spracovania a efektívnosti a nie zvýšením mentálnej kapacity (napr. Bjorkland, 1985; Bjorkland, 1987; Case, 1985). Ďalší teoretici zahrnuli konštrukt „inhibičných“ procesov do svojho kognitívneho vývoja (Harnishfeger & Bjorkland, 1993). Podľa tohto účtu je nezrelá kognícia charakterizovaná citlivosťou na interferenciu z konkurenčných zdrojov, ktoré musia byť potlačené (napr. Brainerd & Reyna, 1993; Casey, Thomas, Davidson, Kunz a Franzen, 2002a; Dempster, 1993; Diamond, 1985; Munakata & Yerys, 2001). Preto cielené správanie si vyžaduje kontrolu impulzov alebo oneskorenie potešenia pre optimalizáciu výsledkov a táto schopnosť sa zdá byť vyspelá v detstve a dospievaní.
Chovanie adolescentov bolo opísané ako impulzívne a riskantné, takmer synonymne, avšak tieto konštrukty sa opierajú o rôzne kognitívne a nervové procesy, ktoré naznačujú odlišné konštrukty s rôznymi vývojovými trajektóriami. Konkrétne prehľad literatúry naznačuje, že impulzivita sa s vekom znižuje v detstve a dospievaní (Casey a kol., 2002a; Casey, Galvan a kol., 2005; Galvan a kol., 2007) a je spojený s predĺženým vývojom prefrontálneho kortexu (Casey, Galvan a kol., 2005), hoci existujú rozdiely v miere, do akej je daný jedinec impulzívny alebo nie, bez ohľadu na vek.
Na rozdiel od kontroly impulzov / kognitívnych funkcií sa zdá, že riskovanie sa zvyšuje v období adolescencie v porovnaní s detstvom a dospelosťou a je spojené so subkortikálnymi systémami, o ktorých je známe, že sa podieľajú na hodnotení odmien. Štúdie zobrazovania na ľuďoch, ktoré budú preskúmané, naznačujú zvýšenie subkortikálnej aktivácie (napr. Accumbens) pri rizikových voľbách (Kuhnen a Knutson, 2005; Matthews a kol., 2004; Montague & Berns, 2002), ktorá je nadsadená u adolescentov v porovnaní s deťmi a dospelými (\ tErnst a spol., 2005; Galvan a kol., 2006). Tieto zistenia poukazujú na rôzne trajektórie pre správanie založené na odmeňovaní alebo stimuloch, pričom skorší vývoj týchto systémov je v porovnaní s kontrolnými systémami, ktoré vykazujú zdĺhavý a lineárny vývojový priebeh, z hľadiska prevládajúcich nevhodných rozhodnutí a činností v prospech cieľovo orientovaných.
Dôkazy z neuroimagingových štúdií vývoja ľudského mozgu
Nedávne výskumy vývoja adolescentného mozgu boli založené na pokrokoch v neuroimagingových metodikách, ktoré sa dajú ľahko použiť pri vývoji ľudských populácií. Tieto metódy sa spoliehajú na metódy magnetickej rezonancie (MRI) (pozri Obr. 2) a zahŕňajú: štrukturálnu MRI, ktorá sa používa na meranie veľkosti a tvaru štruktúr; funkčná MRI, ktorá sa používa na meranie vzorcov mozgovej aktivity; a difúzne tenzorové zobrazovanie (DTI), ktoré sa používa na indexovanie konektivity vlákien z bielej hmoty. Dôkazy pre náš vývojový model kompetície medzi kortikálnymi a subkortikálnymi oblasťami sú podporené nezrelým štruktúrnym a funkčným prepojením, ako je merané pomocou DTI a fMRI.
MRI štúdie vývoja ľudského mozgu
Niekoľko štúdií využilo štrukturálnu MRI na zmapovanie anatomického priebehu normálneho vývoja mozgu (pozri prehľad Durston a kol., 2001). Napriek tomu, že celková veľkosť mozgu je približne šesťnásobná v porovnaní s veľkosťou dospelého jedinca vo veku šiestich rokov, podzložky šedej a bielej hmoty v mozgu naďalej podliehajú dynamickým zmenám počas dospievania. Údaje z nedávnych dlhodobých štúdií MRI ukazujú, že objem šedej hmoty má invertovaný vzor tvaru U, s väčšou regionálnou variabilitou ako biela hmota (Giedd, 2004; Gogtay a kol., 2004; Sowell a kol., 2003; Sowell, Thompson a Toga, 2004). Všeobecne platí, že regióny podriadené primárnym funkciám, ako sú motorické a senzorické systémy, dozrievajú najskôr; oblasti vyššieho rádu, ktoré integrujú tieto primárne funkcie, dozrievajú neskôr (Gogtay a kol., 2004; Sowell, Thompson a Toga, 2004). Štúdie využívajúce opatrenia na báze MRI napríklad ukazujú, že strata kortikálnej šedej hmoty sa vyskytuje najskôr v primárnych senzorimotorických oblastiach a najnovšie v dorzolaterálnej prednej a laterálnej časovej kortikle (Gogtay a kol., 2004). Tento model je v súlade s nehumánnymi primátmi a ľudskými postmortemovými štúdiami, ktoré ukazujú, že prefrontálny kortex je jednou z posledných oblastí mozgu, ktoré dozrievajú (Buržoázia, Goldman-Rakic a Rakic, 1994; Huttenlocher, 1979). Na rozdiel od sivej hmoty sa objem bielej hmoty zväčšuje zhruba lineárnym spôsobom, pričom sa v priebehu vývoja zvyšuje až do dospelosti (Gogtay a kol., 2004). Tieto zmeny pravdepodobne odrážajú pokračujúcu myelináciu axónov oligodendrocytmi, ktoré zvyšujú vedenie a komunikáciu neurónov.
Aj keď sa subkortikálnym oblastiam venovala menšia pozornosť pri skúmaní štrukturálnych zmien, niektoré z najväčších zmien v mozgu v priebehu vývoja sa pozorujú v týchto regiónoch, najmä v bazálnych gangliách (Sowell a kol., 1999, Pozri Obr. 3) a najmä u mužov (\ tGiedd a kol., 1996). Vývojové zmeny v štrukturálnom objeme v bazálnych gangliách a prefrontálnych oblastiach sú zaujímavé vo svetle známych vývojových procesov (napr. Dendritická arborizácia, bunková smrť, synaptické prerezávanie, myelinizácia), ktoré sa vyskytujú počas detstva a dospievania. Tieto procesy umožňujú jemné doladenie a posilnenie prepojení medzi prefrontálnymi a subkortikálnymi oblasťami s rozvojom a učením, ktoré sa môžu zhodovať s väčšou kognitívnou kontrolou. Ako sa tieto štrukturálne zmeny týkajú kognitívnych zmien? Viaceré štúdie sa týkali štruktúrneho dozrievania a kognitívnych funkcií frontálneho laloku pomocou neuropsychologických a kognitívnych opatrení (napr. Sowell a kol., 2003). Konkrétne boli hlásené asociácie medzi prefrontálnymi kortikálnymi a bazálnymi gangliálnymi regionálnymi objemami založenými na MRI a mierami kognitívnej kontroly (tj schopnosť prekonať nevhodnú reakciu v prospech iného alebo potlačiť pozornosť k irelevantnému stimulačnému atribútu v prospech relevantného atribútu stimulu (Casey, Trainor a kol., 1997). Tieto zistenia naznačujú, že kognitívne zmeny sa odrážajú v štrukturálnych zmenách mozgu a podčiarkujú význam subkortikálneho (bazálneho ganglia), ako aj vývoja kortikálneho (napr. Prefrontálneho kortexu).

DTI štúdie vývoja ľudského mozgu
Preskúmané morfometrické štúdie na základe MRI naznačujú, že kortikálne spojenia sú jemne doladené odstránením prebytku synapsií a posilnením relevantných spojení s rozvojom a skúsenosťami. Nedávne pokroky v technológii MRI, ako napríklad DTI, predstavujú potenciálny nástroj na skúmanie úlohy špecifických útvarov bielej hmoty k rozvoju mozgu a správania s väčšou podrobnosťou. Relevantné pre tento dokument sú neuroimaging štúdie, ktoré spojili vývoj vláknitých ciest so zlepšením kognitívnych schopností. Konkrétne boli preukázané asociácie medzi opatreniami založenými na DTI prefrontálneho vývoja bielej hmoty a kontrolou kognitívnych funkcií u detí. V jednej štúdii vývoj tejto kapacity pozitívne koreloval s predfrontálnymi parietálnymi vláknitými traktmi (Nagy, Westerberg a Klingberg, 2004) v súlade s funkčnými štúdiami zobrazujúcimi neuroimaging, ktoré ukázali rozdielny nábor týchto oblastí u detí v porovnaní s dospelými.
Pomocou podobného prístupu, \ t Liston a kol. (2005) ukázali, že trakty bielej hmoty medzi prefrontálnym bazálnym gangliom a -posteriorným vláknitým traktom sa naďalej vyvíjajú v detstve až do dospelosti, ale len tie oblasti medzi prefrontálnym kortexom a bazálnymi gangliami korelujú s kontrolou impulzov, ako sa meria výkonom na go / nogo úloha. Prefrontálne vláknité trakty boli definované oblasťami záujmov identifikovanými v štúdii fMRI s použitím tej istej úlohy. V obidvoch vývojových štúdiách DTI boli opatrenia vláknového traktu korelované s vývojom, ale špecifickosť jednotlivých vláknitých úsekov s kognitívnou výkonnosťou bola preukázaná disociáciou konkrétneho traktu (Liston a kol., 2005) alebo kognitívnych schopností (Nagy a kol., 2004). Tieto zistenia podčiarkujú dôležitosť skúmania nielen regionálnych zmien, ale aj zmien súvisiacich s obvodmi, keď sa tvrdia o zmenách v nervových substrátoch kognitívneho vývoja závislých od veku.
Funkčné MRI štúdie správania a vývoja mozgu
Hoci štrukturálne zmeny merané pomocou MRI a DTI boli spojené so zmenami správania počas vývoja, priamejším prístupom pri skúmaní asociácie štruktúry a funkcie je meranie zmien v mozgu a správaní súčasne, ako pri fMRI. Schopnosť merať funkčné zmeny vo vyvíjajúcom sa mozgu s MRI má významný potenciál pre oblasť vývojovej vedy. V kontexte súčasného článku, fMRI poskytuje prostriedok na obmedzenie interpretácie správania adolescentov. Ako už bolo uvedené, predpokladá sa, že vývoj prefrontálneho kortexu hrá dôležitú úlohu pri dozrievaní vyšších kognitívnych schopností, ako je rozhodovanie a kognitívna kontrola (Casey, Tottenham a Fossella 2002b; Casey, Trainor a kol., 1997). Na posúdenie neurobiologického základu týchto schopností, vrátane úloh flanker, Stroop a go / nogo, boli použité mnohé paradigmy spolu s fMRI.Casey, Castellanos a kol., 1997; Casey, Giedd a Thomas, 2000a; Durston a kol., 2003). Tieto štúdie spoločne ukazujú, že deti pri vykonávaní týchto úloh prijímajú odlišné, ale často väčšie, viac rozptýlené prefrontálne regióny ako dospelí. Štruktúra aktivity v oblastiach mozgu, ktorá je centrálna pre výkon úloh (tj koreluje s kognitívnou výkonnosťou), sa stáva viac zameranou alebo jemne doladenou s vekom, zatiaľ čo regióny, ktoré nesúvisia s výkonom úlohy, sa znižujú s aktivitou. Tento vzor bol pozorovaný v oboch prierezových (Brown et al., 2005) a dlhodobých štúdií (\ tDurston a kol., 2006) a naprieč rôznymi paradigmami. Hoci neuroimagingové štúdie nedokážu definitívne charakterizovať mechanizmus takýchto vývojových zmien (napr. Dendritická arborizácia, synaptické prerezávanie), zistenia odrážajú vývoj v rámci a zjemňovanie projekcií do a z aktivovaných oblastí mozgu s dozrievaním a naznačujú, že tieto zmeny sa vyskytujú počas predĺženého obdobia. doba (Brown et al., 2005; Bunge, Dudukovic, Thomason, Vaidya a Gabrieli, 2002; Casey, Trainor a kol., 1997; Casey a kol., 2002a; Crone, Donohue, Honomichl, Wendelken a Bunge, 2006; Luna a kol., 2001; Moses a kol., 2002; Schlaggar a kol., 2002; Tamm a kol., 2002; Thomas a kol., 2004; Turkeltaub, Gareau, Flowers, Zeffiro a Eden, 2003).
Ako nás môže táto metodika informovať o tom, či adolescentom skutočne chýba dostatočná kognitívna kontrola (impulzívna) alebo či sú riskantní vo svojich voľbách a činnostiach? Kontrola impulzov meraná úlohami kognitívnej kontroly, ako je úloha go / nogo, ukazuje lineárny priebeh vývoja v detstve a adolescencii, ako je opísané vyššie. Nedávne štúdie zobrazujúce neuroimaging však začali skúmať spracovanie súvisiace s odmenami, ktoré je špecifické pre riziko u adolescentov (Bjork a kol., 2004; Ernst a spol., 2005; May a kol., 2004). Tieto štúdie sa zameriavali primárne na oblasť akumulovaných oblastí, čo je časť bazálnych ganglií, ktoré sa podieľajú na predpovedaní odmeny, skôr než na charakterizácii vývoja tejto oblasti v spojení s kontrolnými oblasťami zhora nadol (prefrontálny kortex). Hoci sa v nedávnej správe o menovej rozhodovacej úlohe o rizikovom správaní ukázala nedávna správa o menšej ventrálnej prefrontálnej aktivite u adolescentov v porovnaní s dospelými (Eshel, Nelson, Blair, Pine a Ernst, 2007).
Celkovo možno konštatovať, že niekoľko štúdií skúmalo, ako sa vývoj obvodov odmeňovania v subkortikálnych oblastiach (napr. Akumulácie) mení v súvislosti s rozvojom kortikálnych prefrontálnych regiónov. Okrem toho, ako sa tieto nervové zmeny zhodujú s vyhľadávaním odmeny, impulzívnosťou a rizikovým správaním, zostáva relatívne neznáma. Náš neurobiologický model navrhuje, aby kombinácia zvýšenej citlivosti na odmeňovanie a nezrelosť v oblastiach kontroly správania mohla zaujať adolescentov k tomu, aby hľadali okamžité a nie dlhodobé zisky, čo by mohlo vysvetliť ich nárast rizikového rozhodovania a impulzívneho správania. Sledovanie subkortikálneho (napr. Accumbens) a kortikálneho (napr. Prefrontálneho) vývoja rozhodovania v detstve prostredníctvom dospelosti poskytuje ďalšie obmedzenia, či zmeny hlásené v adolescencii sú špecifické pre toto obdobie vývoja, alebo odrážajú dozrievanie, ktoré sa neustále vyskytuje v trochu lineárny vzor od detstva až po dospelosť.
Empirické dôkazy z nedávnej štúdie fMRI pomáhajú podporovať náš neurobiologický model a využívajú prechodný prístup k pochopeniu adolescencie skúmaním zmien pred a po dospievaní. V tejto štúdii (Galvan a kol., 2006), skúmali sme behaviorálne a nervové reakcie na odmeňovanie manipulácií v rámci vývoja, pričom sa zameriavame na oblasti mozgu, ktoré sa podieľajú na vzdelávaní a správaní v súvislosti s odmeňovaním (Hikosaka a Watanabe, 2000; Pecina, Cagniard, Berridge, Aldridge a Zhuang, 2003; Schultz, 2006) a štúdie pre dospelých (napr. Knutson, Adams, Fong a Hommer, 2001; O, Doherty, Kringelbach, Rolls, Hornak, Andrews, 2001; Zald a kol., 2004) av štúdiách závislosti (Hyman & Malenka, 2001; Volkow & Li, 2004). Na základe modelov hlodavcov (Laviola a kol., 1999; Spear, 2000) a predchádzajúce zobrazovacie práce (Ernst a spol., 2005), predpokladali sme, že vo vzťahu k deťom a dospelým by adolescenti ukázali prehnanú aktiváciu akumulov, v zhode s menej zrelým náborom oblastí zhora nadol. Nedávna práca, ktorá ukazuje oneskorenú funkčnú konektivitu medzi týmito prefrontálnymi a limbickými subkortikálnymi oblasťami v adolescencii v porovnaní s dospelými, poskytuje mechanizmus pre nedostatok kontroly zhora nadol v týchto oblastiach (Hare a kol., 2007a).
Naše zistenia boli v súlade s modelmi hlodavcov (Laviola, Macri, Morley-Fletcher a Adriani, 2003) a predchádzajúcich zobrazovacích štúdií (\ tErnst a spol., 2005) naznačujúce zvýšenú aktivitu akumulovania odmien počas dospievania. V porovnaní s deťmi a dospelými adolescenti skutočne prejavili prehnanú odozvu accumbens v očakávaní odmeny. Deti aj dospievajúci však vykazovali menej zrelú reakciu v prefrontálnych kontrolných oblastiach ako dospelí. Tieto zistenia naznačujú, že rôzne vývojové trajektórie pre tieto regióny môžu byť základom pre zvýšenie aktivity akumulovaných pacientov vo vzťahu k deťom alebo dospelým, čo môže súvisieť so zvýšeným impulzívnym a rizikovým správaním pozorovaným počas tohto obdobia vývoja (pozri Obr. 4).

Rozdielny nábor prefrontálnych a subkortikálnych oblastí bol zaznamenaný v mnohých vývojových štúdiách fMRI (Casey a kol., 2002b; Monk a kol., 2003; Thomas a kol., 2004). Typicky boli tieto zistenia interpretované skôr z hľadiska nedospelých prefrontálnych oblastí než nerovnováhy medzi prefrontálnym a subkortikálnym regionálnym rozvojom. Vzhľadom na dôkazy o prefrontálnych regiónoch v riadení vhodných opatrení v rôznych kontextoch (Miller & Cohen, 2001) nezrelá predpredajná činnosť by mohla brániť vhodnému odhadu budúcich výsledkov a hodnoteniu rizikových rozhodnutí a mohla by byť teda menej ovplyvnená hodnotením odmeny ako akumulovaním. Tento model je v súlade s predchádzajúcim výskumom, ktorý ukazuje zvýšenú subkortikálnu, v porovnaní s kortikálnou aktivitou, keď sú rozhodnutia ovplyvnené okamžitými dlhodobými ziskami (McClure, Laibson, Loewenstein a Cohen, 2004). Okrem toho sa ukázalo, že akumulovaná aktivita s fMRI pozitívne koreluje s následným rizikovým správaním (Kuhnen a Knutson, 2005). Počas dospievania, v porovnaní s detstvom alebo dospelosťou, nezrelá ventrálna prefrontálna kôra nemusí poskytovať dostatočnú kontrolu zhora nadol v silne aktivovaných oblastiach spracovania odmien (napr. Accumbens), čo vedie k menšiemu vplyvu prefrontálnych systémov (orbitofrontálnej kôry) v porovnaní s akumulovanými odmenami ocenenie.
Prečo by mal byť mozog naprogramovaný tak, aby sa vyvíjal týmto spôsobom?
Dospievanie je prechodné obdobie medzi detstvom a dospelosťou, ktoré sa často vyskytuje spoločne s pubertou. Puberta označuje začiatky sexuálneho dozrievania (Graber & Brooks-Gunn, 1998) a môže byť definovaný biologickými markermi. Dospievanie možno opísať ako progresívny prechod do dospelosti s neurčitým ontogenetickým časovým priebehom (Spear, 2000). Evolučne je obdobie dospievania obdobím, v ktorom sa získavajú zručnosti nezávislosti na zvýšenie úspechu po oddelení od ochrany rodiny, hoci sa zvyšuje šanca na škodlivé okolnosti (napr. Zranenie, depresia, úzkosť, užívanie drog a závislosť).Kelley, Schochet a Landry, 2004). Správanie zamerané na nezávislosť prevláda naprieč druhmi, ako je napr. Zvýšenie sociálnych interakcií zameraných na rovesníkov a intenzívnosť pri hľadaní nových a hľadaní rizika. Psychosociálne faktory ovplyvňujú adolescentný sklon k rizikovému správaniu. Rizikové správanie je však výsledkom biologicky poháňanej nerovnováhy medzi zvýšeným hľadaním novosti a vnímania v spojení s nezrelými „samoregulačnými schopnosťami“ (Steinberg, 2004). Naše neurobiologické údaje naznačujú, že k tomu dochádza prostredníctvom diferenciálneho vývoja týchto dvoch systémov (limbic a control).
Špekulácie naznačujú, že tento vývojový model je evolučnou vlastnosťou. Musíte sa angažovať vo vysoko rizikovom správaní, aby ste opustili svoju rodinu a dedinu, aby ste našli partnera a riskovať v rovnakom čase, ako hormóny poháňajú adolescentov, aby vyhľadávali sexuálnych partnerov. V dnešnej spoločnosti, keď sa dospievanie môže predĺžiť donekonečna, pričom deti žijú s rodičmi a majú finančnú závislosť a neskôr si vyberajú kamarátov, tento vývoj možno považovať za nevhodný.
Existujú dôkazy naprieč druhmi o zvýšenom vyhľadávaní a riskovaní počas adolescentných rokov. Hľadanie rovesníkov v rovnakom veku a boj s rodičmi, ktoré pomáhajú dospievajúcemu odtrhnúť od domáceho územia na párenie, sa vyskytujú u iných druhov vrátane hlodavcov, primátov a iných vtákov (Spear, 2000). V porovnaní s dospelými vykazujú periadolescentné potkany zvýšené správanie pri hľadaní novosti, ktoré sa zameriava na novosť (Laviola a kol., 1999). Neurochemické dôkazy naznačujú, že rovnováha v adolescentnom mozgu medzi kortikálnymi a subkortikálnymi dopamínovými systémami sa začína presúvať smerom k vyšším hladinám kortikálneho dopamínu počas dospievania (Spear, 2000). Podobná protrahovaná dopaminergná enzervencia prostredníctvom adolescencie do dospelosti sa ukázala aj v prekulovom kortexe neľudského primáta (Rosenberg a Lewis, 1995). Zdá sa, že toto zvýšené zdanlivé riskovanie je naprieč druhmi a má dôležité adaptívne účely.
Biologické predispozície, vývoj a riziko
Individuálne rozdiely v kontrole impulzov a prijímaní rizík boli v psychológii už nejaký čas uznané (Benthin, Slovic a Severson, 1993). Jedným z klasických príkladov individuálnych rozdielov uvedených v týchto schopnostiach v sociálnej, kognitívnej a vývojovej psychologickej literatúre je zrejme oneskorenie potešenia (Mischel, Shoda a Rodriguez, 1989). Oneskorenie potešenia sa zvyčajne posudzuje u batoľat 3- až 4-ročných. Batoľa sa pýta, či by radšej malú odmenu (jednu cookie) alebo veľkú odmenu (dve cookies). Potom je dieťaťu povedané, že experimentátor opustí miestnosť, aby sa pripravil na nadchádzajúce aktivity a vysvetľuje dieťaťu, že ak zostane na svojom mieste a nebude jesť cookie, dostane veľkú odmenu. Ak dieťa neurobí, alebo nemôže počkať, mala by zazvoniť, aby privolala experimentátora a získala tak menšiu odmenu. Akonáhle je jasné, že dieťa chápe úlohu, sedí pri stole s dvomi odmenami a zvončekom. Rozptýlenie v miestnosti je minimalizované, bez hračiek, kníh alebo obrázkov. Experiment sa vracia po 15 min alebo po tom, čo dieťa zazvonilo zvonček, zjedol odmenu alebo prejavil akékoľvek známky úzkosti. Mischel ukázal, že deti sa zvyčajne správajú jedným z dvoch spôsobov: (1) buď zvonia takmer okamžite, aby mali cookie, čo znamená, že ho dostanú len jeden; alebo (2) čakajú a optimalizujú svoje zisky a prijímajú obidva cookies. Toto zistenie naznačuje, že niektorí jednotlivci sú lepší ako iní v ich schopnosti kontrolovať impulzy tvárou v tvár silným stimulom a táto zaujatosť sa dá zistiť v ranom detstve (Mischel a kol., 1989) a zdá sa, že zostávajú v období dospievania a mladej dospelosti (Eigsti a kol., 2006).
Čo môže vysvetliť jednotlivé rozdiely v optimálnom rozhodovaní a správaní? Niektorí teoretici predpokladali, že dopaminergné mezolimbické obvody, ktoré sa podieľajú na spracovaní odmien, sú základom rizikového správania. Jednotlivé rozdiely v tomto obvode, ako sú alelické varianty v génoch súvisiacich s dopamínom, čo vedie k príliš malému alebo príliš veľkému množstvu dopamínu v subkortikálnych oblastiach, by mohli súvisieť so sklonom k riskantnému správaniu (O'Doherty, 2004). Ukázalo sa, že nucleus accumbens zvyšuje aktivitu bezprostredne pred riskantným výberom paradigiem peňažného rizika (Kuhnen a Knutson, 2005; Matthews a kol., 2004; Montague & Berns, 2002) a ako je opísané vyššie, adolescenti vykazujú nadsadenú aktivitu akumulovania na odmeňovanie výsledkov vo vzťahu k deťom alebo dospelým (Ernst a spol., 2005; Galvan a kol., 2006). Tieto údaje kolektívne naznačujú, že adolescenti môžu byť náchylnejší na riskantné voľby ako skupina (Gardener & Steinberg, 2005), ale niektorí adolescenti budú náchylnejší ako iní, aby sa zapojili do rizikového správania, čím by sa potenciálne ohrozili negatívne výsledky. Preto je dôležité zvážiť individuálnu variabilitu pri skúmaní komplexných vzťahov medzi mozgom a správaním súvisiacich s podstupovaním rizika a odmeňovaním vo vývojových populáciách.
Preskúmať individuálne rozdiely v rizikovom správaní, Galvan a kol. (2007) nedávno skúmal súvislosť medzi aktivitou v odmeňovaní súvisiacou nervovou sústavou v očakávaní veľkej peňažnej odmeny s mierou osobnostných charakteristík riskovania a impulzívnosti v adolescencii. Funkčné zobrazovanie magnetickou rezonanciou a anonymné hodnotiace škály rizikového správania, vnímanie rizika a impulzívnosť boli získané u jedincov vo veku 7 a 29. Medzi aktivitou akumulovania a pravdepodobnosťou zapojenia sa do rizikového správania naprieč vývojom bola pozitívna súvislosť. Táto aktivita sa líšila ako funkcia hodnotenia predpokladaných pozitívnych alebo negatívnych dôsledkov takéhoto správania jednotlivcami. Tí jednotlivci, ktorí vnímali riskantné správanie ako vedúce k zúfalým následkom, aktivovali akumulátory menej na odmenu. Toto združenie bolo poháňané prevažne deťmi, pričom dospelí hodnotili dôsledky takéhoto správania. Hodnoty impulzivity neboli spojené s aktivitou akumulátorov, ale skôr s vekom. Tieto zistenia naznačujú, že v období dospievania môžu byť niektorí jedinci náchylnejší na riskantné správanie v dôsledku vývojových zmien v zhode s variabilitou predispozície daného jedinca k riskantnému správaniu, a nie k jednoduchým zmenám v impulzivite (pozri Obr. 5).

Chovanie adolescentov bolo opakovane charakterizované ako impulzívne a riskantné (Steinberg, 2004, 2007), ale táto revízia zobrazovacej literatúry naznačuje rôzne neurobiologické substráty a rôzne vývojové trajektórie pre tieto správanie. Konkrétne, impulzivita je spojená s nezrelým ventrálnym prefrontálnym vývojom a postupne sa znižuje od detstva do dospelosti (Casey, Galvan a kol., 2005). Negatívna korelácia medzi hodnotami impulzivity a vekom v štúdii Galvan a kol. (2007) ďalej podporuje tento pojem. Na rozdiel od toho je riziko spojené so zvýšením aktivity akumulovaných pacientov (Kuhnen a Knutson, 2005; Matthews a kol., 2004; Montague & Berns, 2002), čo je v prípade adolescentov v porovnaní s deťmi a dospelými prehnané (Ernst a spol., 2005; Galvan a kol., 2006). Voľby a správanie adolescentov teda nemožno vysvetliť impulzívnosťou alebo dlhotrvajúcim vývojom samotného prefrontálneho kortexu, pretože deti by potom mali byť predpovedané ako väčšie riziko. Zistenia poskytujú nervový základ pre to, prečo sú niektorí adolescenti vystavení väčšiemu riziku ako iní, ale ďalej poskytujú základ pre to, ako sa správanie adolescentov odlišuje od detí a dospelých v riziku.
Tieto údaje spoločne naznačujú, že hoci dospievajúci ako skupina sú považovaní za osoby, ktoré prijímajú riziko (Gardener & Steinberg, 2005), niektorí adolescenti budú náchylnejší ako iní, aby sa zapojili do rizikového správania, čím by sa potenciálne ohrozili negatívne výsledky. Tieto zistenia podčiarkujú dôležitosť zváženia individuálnej variability pri skúmaní komplexných vzťahov medzi mozgom a správaním súvisiacich s podstupovaním rizika a spracovaním odmien vo vývojových populáciách. Okrem toho, tieto individuálne a vývojové rozdiely môžu pomôcť vysvetliť zraniteľnosť niektorých jedincov rizikom spojeným s užívaním látok a nakoniec závislosť.
Závery
Štúdie zobrazovania u ľudí ukazujú štrukturálne a funkčné zmeny vo frontostriatálnych oblastiach (Giedd a kol., 1996, 1999; Jernigan a kol., 1991; Sowell a kol., 1999; na preskúmanie, Casey, Galvan a kol., 2005), ktoré súbežne zvyšujú kognitívnu kontrolu a samoreguláciu (Casey, Trainor a kol., 1997; Luna & Sweeney, 2004; Luna a kol., 2001; Rubia a kol., 2000; Steinberg, 2004; pozri tiež Steinberg, 2008, táto záležitosť). Zdá sa, že tieto zmeny poukazujú na posun v aktivácii prefrontálnych regiónov z difúzneho na viac zamerané na nábor v priebehu času (Brown et al., 2005; Bunge a kol., 2002; Casey, Trainor a kol., 1997; Durston a kol., 2006; Moses a kol., 2002) a zvýšeného náboru subkortikálnych oblastí počas dospievania (Casey a kol., 2002a; Durston a kol., 2006; Luna a kol., 2001). Hoci neuroimagingové štúdie nedokážu definitívne charakterizovať mechanizmus takýchto vývojových zmien, tieto zmeny v objeme a štruktúre môžu odrážať vývoj v rámci a zjemňovanie projekcií do az týchto oblastí mozgu počas dozrievania, čo naznačuje jemné doladenie systému s vývojom.
Celkovo vzaté, zistené zistenia naznačujú, že zvýšené správanie s rizikom v adolescencii je spojené s rôznymi vývojovými trajektóriami subkortikálneho potešenia a regiónov s kortikálnou kontrolou. Tieto vývojové zmeny môžu zhoršiť individuálne rozdiely v činnosti systémov odmeňovania. Hoci adolescencia bola rozlíšená ako obdobie charakterizované vyhľadávaním odmien a rizikovým správaním (Gardener & Steinberg, 2005; Spear, 2000) individuálne rozdiely v nervových reakciách na odmeňovanie, predispozíciu niektorých adolescentov k tomu, aby prijali viac rizík ako iné, čím sa vystavujú väčšiemu riziku negatívnych výsledkov. Tieto zistenia poskytujú rozhodujúci základ pre syntézu rôznych zistení súvisiacich s rizikovým správaním v adolescencii a pre pochopenie individuálnych rozdielov a vývojových markerov pre sklony k zapojeniu sa do negatívneho správania.
Poďakovanie
Táto práca bola čiastočne podporená grantmi Národného inštitútu pre zneužívanie drog R01 DA18879 a Národného ústavu duševného zdravia 1P50 MH62196.
Referencie
- Benthin A, Slovic P, Severson H. Psychometrická štúdia vnímania rizika adolescentov. Journal of Adolescence. 1993;16: 153-168. [PubMed]
- Bjork JM, Knutson B, Fong GW, Caggiano DM, Bennett SM, Hommer DW. Incentívna aktivácia mozgu u adolescentov: podobnosti a rozdiely od mladých dospelých. Journal of Neuroscience. 2004;24: 1793-1802. [PubMed]
- Bjorkland DF. Úloha koncepčných poznatkov vo vývoji organizácie v detskej pamäti. In: Brainerd CJ, Pressley M, redaktori. Základné procesy vo vývoji pamäti: Pokrok v oblasti výskumu kognitívneho vývoja. New York: Springer-Verlag; 1985. str. 103-142.
- Bjorkland DF. Ako vekové zmeny v vedomostnej základni prispievajú k rozvoju detskej pamäti: výkladová interpretácia. Vývojová kontrola. 1987;7: 93-130.
- Bourgeois JP, Goldman-Rakic PS, Rakic P. Synaptogenéza v prefrontálnom kortexe opíc rhesus. Mozgová kôra. 1994;4: 78-96. [PubMed]
- Brainerd CJ, Reyna VF. Nezávislosť pamäte a rušenie pamäte v kognitívnom vývoji. Psychologický prehľad. 1993;100: 42-67. [PubMed]
- Brown TT, Lugar HM, Coalson RS, Miezin FM, Petersen SE, Schlaggar BL. Vývojové zmeny v ľudskej mozgovej funkčnej organizácii pre generovanie slov. Mozgová kôra. 2005;15: 275-290. [PubMed]
- Bunge SA, Dudukovic NM, Thomason ME, Vaidya CJ, Gabrieli JD. Nezrelý frontálny lalok prispieva k kognitívnej kontrole u detí: Dôkazy z fMRI. Neurón. 2002;33: 301-311. [PubMed]
- Prípad R. Validácia neo-piagetovského kapacitného konštruktu. Journal of Experimental Child Psychology. 1972;14: 287-302.
- Prípad R. Duševný rozvoj: Narodenie do dospelosti. New York: Akademická tlač; 1985.
- Casey BJ, Castellanos FX, Giedd JN, Marsh WL, Hamburger SD, Schubert AB. et al. Implikácia pravého frontostriatálneho obvodu pri inhibícii odozvy a poruchy pozornosti / hyperaktivity. Journal of American Academy of Child and Adolescent Psychiatry. 1997;36: 374-383. [PubMed]
- Casey BJ, Galvan A, Hare TA. Zmeny v organizácii mozgových funkcií počas kognitívneho vývoja. Aktuálne stanovisko v neurobiológii. 2005;15: 239-244. [PubMed]
- Casey BJ, Giedd JN, Thomas KM. Štrukturálny a funkčný vývoj mozgu a jeho vzťah k kognitívnemu vývoju. Biologická psychológia. 2000;54: 241-257. [PubMed]
- Casey BJ, Thomas KM, Davidson MC, Kunz K, Franzen PL. Disociácia striatálnej a hipokampálnej funkcie vývojovo s úlohou kompatibility stimulačnej odozvy. Journal of Neuroscience. 2002;22: 8647-8652. [PubMed]
- Casey BJ, Thomas KM, Welsh TF, Badgaiyan RD, Eccard CH, Jennings JR a kol. Disociácia konfliktu odozvy, výber pozornosti a očakávania s funkčným zobrazením magnetickou rezonanciou. Zborník Národnej akadémie vied. 2000b;97: 8728-8733.
- Casey BJ, Tottenham N, Fossella J. Klinické, zobrazovacie, lézné a genetické prístupy k modelu kognitívnej kontroly. Vývojová psychobiológia. 2002b;40: 237-254. [PubMed]
- Casey BJ, Tottenham N, Liston C, Durston S. Zobrazovanie vyvíjajúceho sa mozgu: Čo sme sa dozvedeli o kognitívnom vývoji? Trendy v kognitívnej vede. 2005;9: 104-110.
- Casey BJ, Trainor RJ, Orendi JL, Schubert AB, Nystrom LE, Giedd JN, et al. Vývojová funkčná MRI štúdia prefrontálnej aktivácie počas výkonu úlohy go-no-go. Journal of Cognitive Neuroscience. 1997;9: 835-847.
- Crone E, Donohue S, Honomichl R, Wendelken C, Bunge S. Oblasti mozgu sprostredkujúce flexibilné použitie pravidiel počas vývoja. Journal of Neuroscience. 2006;26: 11239-11247. [PubMed]
- Dempster FN. Odolnosť proti rušeniu: Vývojové zmeny v základnom mechanizme spracovania. V: Howe ML, Pasnak R, redaktori. Rozvíjajúce sa témy v kognitívnom vývoji Volume 1: Základy. New York: Springer; 1993. s. 3 – 27.
- Diamant A. Vývoj schopnosti používať odvolanie na sprievodnú akciu, ako to naznačuje výkon dojčiat na AB. Detský rozvoj. 1985;56: 868-883. [PubMed]
- Durston S, Davidson MC, Thomas KM, Worden MS, Tottenham N, Martinez A, et al. Parametrická manipulácia s konkurenciou konfliktov a odpovedí pomocou rýchleho fMRI udalostí súvisiacich so zmiešanou skúškou. Neuroimage. 2003;20: 2135-2141. [PubMed]
- Durston S, Davidson MC, Tottenham N, Galvan A, Spicer J, Fossella J a kol. Prechod od difúznej k fokálnej kortikálnej aktivite s vývojom. Vývojová veda. 2006;1: 18-20. [PubMed]
- Durston S, Hulshoff Pol HE, Casey BJ, Giedd JN, Buitelaar JK, van Engeland H. Anatomická MRI vyvíjajúceho sa ľudského mozgu: Čo sme sa naučili? Časopis Americkej akadémie detskej adolescentnej psychiatrie. 2001;40: 1012-1020.
- Eaton LK, Kinchen S, Ross J, Hawkins J, Harris WA, Lowry R a kol. Sledovanie rizikového správania mládeže - Spojené štáty, 2005, prehľady o dohľade. Týždenná správa o chorobnosti a úmrtnosti. 2006;55: 1-108. [PubMed]
- Eigsti IM, Zayas V, Mischel W, Shoda Y, Ayduk O, Dadlani MB a kol. Predpovedanie kognitívnej kontroly od predškolskej do neskorej adolescencie a mladej dospelosti. Psychologické vedy. 2006;17: 478-484. [PubMed]
- Ernst M, Nelson EE, Jazbec S, McClure EB, Monk CS, Leibenluft E a kol. Amygdala a nucleus accumbens v odpovediach na príjem a opomenutie zisku u dospelých a dospievajúcich. Neuroimage. 2005;25: 1279-1291. [PubMed]
- Eshel N, Nelson EE, Blair RJ, borovica DS, Ernst M. Neurálne substráty výberu u dospelých a adolescentov: Vývoj ventrolaterálnych predchodových a predných cingulárnych kortikúl. Neuropsychológie. 2007;45: 1270-1279. [Článok bez PMC] [PubMed]
- Flavell JH, Feach DR, Chinsky JM. Spontánna verbálna skúška v úlohe pamäti ako funkcia veku. Detský rozvoj. 1966;37: 283-299. [PubMed]
- Galvan A, Hare TA, Parra CE, Penn J, Voss H, Glover G a kol. Skorší vývoj akumulovaní v porovnaní s orbitofrontálnym kortexom by mohol byť základom správania sa u adolescentov. Journal of Neuroscience. 2006;26: 6885-6892. [PubMed]
- Galvan A, Hare T, Voss H, Glover G, Casey BJ. Riziko a dospievajúci mozog: Kto je v ohrození? Vývojová veda. 2007;10: F8-F14. [PubMed]
- Gardener M, Steinberg L. Vzájomný vplyv na riskovanie, preferovanie rizík a riskantné rozhodovanie v adolescencii a dospelosti: Experimentálna štúdia. Vývojová psychológia. 2005;41: 625-635. [PubMed]
- Giedd JN. Štrukturálne magnetické rezonančné zobrazovanie adolescentného mozgu. Annals z Newyorskej akadémie vied. 2004;1021: 77-85. [PubMed]
- Giedd JN, Blumenthal J., Jeffries NO, Castellanos FX, Liu H, Zijdenbos A a kol. Vývoj mozgu počas detstva a dospievania: Dlhodobá štúdia MRI. Nature Neuroscience. 1999;2: 861-863.
- Giedd JN, Snell JW, Lange N, Rajapakse JC, Casey BJ, Kozuch PL a kol. Kvantitatívne zobrazovanie vývoja ľudského mozgu pomocou magnetickej rezonancie: Vek 4 – 18. Mozgová kôra. 1996;6: 551-560. [PubMed]
- Gogtay N, Giedd JN, Lusk L, Hayashi KM, Greenstein D, Vaituzis AC a kol. Dynamické mapovanie ľudského kortikálneho vývoja v detstve až do ranej dospelosti. Zborník Národnej akadémie vied Spojených štátov amerických. 2004;101: 8174-8179. [Článok bez PMC] [PubMed]
- Graber JA, Brooks-Gunn J. Puberty. In: Blechman EA, Brownell KD, redaktori. Behaviorálna medicína a ženy komplexnú príručku. New York, NY: Guilford Press; 1998. s. 51 – 58.
- Hare TA, Voss HU, Glover GH, Casey BJ. Dospievajúci mozog a potenciálne riziko úzkosti a depresie. 2007 Odoslané na uverejnenie.
- Hare TA, Voss HU, Glover GH, Casey BJ. Súťaž medzi prefrontálnym a subkortikálnym limbickým systémom je základom emocionálnej reaktivity počas dospievania. 2007b Odoslané na uverejnenie.
- Harnishfeger KK, Bjorkland F. Ongeny inhibičných mechanizmov: Obnovený prístup k kognitívnemu vývoju. In: Howe ML, Pasnek R, redaktori. Nové témy v kognitívnom vývoji. Vol. 1. New York: Springer-Verlag; 1993. s. 28 – 49.
- Hikosaka K, Watanabe M. Oneskorenie aktivity orbitálnych a laterálnych prefrontálnych neurónov opice s rôznymi odmenami. Mozgová kôra. 2000;10: 263-271. [PubMed]
- Huttenlocher PR. Synaptická hustota v ľudskej frontálnej kôre - Vývojové zmeny a účinky starnutia. Brain Research. 1979;163: 195-205. [PubMed]
- Hyman SE, Malenka RC. Závislosť a mozog: neurobiológia nutkania a jeho vytrvalosť. Príroda Recenzie Neuroscience. 2001;2: 695-703.
- Jernigan TL, Zisook S, Heaton RK, Moranville JT, Hesselink JR, Braff DL. Abnormality zobrazovania magnetickou rezonanciou v lentikulárnych jadrách a mozgovej kôre pri schizofrénii. Archívy všeobecnej psychiatrie. 1991;48: 811-823.
- Keating DP, Bobbitt BL. Individuálne a vývojové rozdiely v kognitívnych procesných zložkách mentálnych schopností. Detský rozvoj. 1978;49: 155-167.
- Kelley AE, Schochet T, Landry C. Annals z Newyorskej akadémie vied. 2004;1021: 27-32. [PubMed]
- Knutson B, Adams CM, Fong GW, Hommer D. Očakávanie zvýšenia peňažnej odmeny selektívne rekrutuje nucleus accumbens. Journal of Neuroscience. 2001;21: RC159. [PubMed]
- Kuhnen CM, Knutson B. Neurónový základ prijímania finančných rizík. Neurón. 2005;47: 763-770. [PubMed]
- Laviola G, Adriani W, Terranova ML, Gerra G. Psychobiologické rizikové faktory pre citlivosť na psychostimulanty u ľudských adolescentov a zvieracích modelov. Neuroscience a Biobehavioral Reviews. 1999;23: 993-1010. [PubMed]
- Laviola G, Macri S, Morley-Fletcher S, Adriani W. Abstraktné rizikové správanie u dospievajúcich myší: Psychobiologické determinanty a včasný epigenetický vplyv. Neuroscience a Biobehavioral Reviews. 2003;27: 19-31. [PubMed]
- Liston C, Watts R, Tottenham N, Davidson MC, Niogi S, Ulug AM a kol. Frontostriatálna mikroštruktúra moduluje účinné získavanie kognitívnej kontroly. Mozgová kôra. 2005;16: 553-560. [PubMed]
- Luna B, Sweeney JA. Vznik spolupráce mozgových funkcií: FMRI štúdie vývoja inhibície odpovede. Annals z Newyorskej akadémie vied. 2004;1021: 296-309. [PubMed]
- Luna B, Thulborn KR, Munoz DP, Merriam EP, Garver KE, Minshew NJ a kol. Zrenie široko distribuovaných funkcií mozgu podporuje kognitívny vývoj. Neuroimage. 2001;13: 786-793. [PubMed]
- Matthews SC a kol. Selektívna aktivácia nucleus accumbens počas rozhodovania o riziku. Neuroreport. 2004;15: 2123-2127. [PubMed]
- Máj JC, Delgado MR, Dahl RE, Stenger VA, Ryan ND, Fiez JA, et al. Funkčné magnetické rezonančné zobrazovanie mozgových obvodov súvisiacich s odmenou u detí a dospievajúcich. Biologická psychiatria. 2004;55: 359-366. [PubMed]
- McClure SM, Laibson DI, Loewenstein G, Cohen JD. Samostatné neurónové systémy oceňujú okamžité peňažné odmeny. Science. 2004;306: 503-507. [PubMed]
- Miller EK, Cohen JD. Integrovaná teória funkcie prefrontálneho kortexu. Ročný prehľad neurovedy. 2001;24: 167-202.
- Mischel W, Shoda Y, Rodriguez MI. Oneskorenie potešenia detí. Science. 1989;244: 933-938. [PubMed]
- Monk CS, McClure EB, Nelson EE, Zarahn E, Bilder RM, Leibenluft E a kol. Nezrelosť dospievajúcich v súvislosti s angažovanosťou mozgu zameranou na pozornosť na emocionálne výrazy tváre. Neuroimage. 2003;20: 420-428. [PubMed]
- Montague PR, Berns GS. Neurónová ekonomika a biologické substráty oceňovania. Neurón. 2002;36: 265-284. [PubMed]
- Moses P, Roe K, Buxton RB, Wong EC, Frank LR, Stiles J. Funkčné MRI globálneho a lokálneho spracovania u detí. Neuroimage. 2002;16: 415-424. [PubMed]
- Munakata Y, Yerys BE. Teraz spolu: Keď zmiznú disociácie medzi vedomosťami a činmi. Pscychologická veda. 2001;12: 335-337.
- Nagy Z, Westerberg H, Klingberg T. Zrenie bielej hmoty je spojené s rozvojom kognitívnych funkcií v detstve. Journal of Cognitive Neuroscience. 2004;16: 1227-1233. [PubMed]
- O'Doherty J, Kringelbach ML, Rolls ET, Hornak J, Andrews C. Abstraktné odmeny a tresty v ľudskej orbitofrontálnej kôre. Nature Neurosci. 2001;4: 95-102. [PubMed]
- O'Doherty JP. Zastúpenie odmeňovania a učenie súvisiace s odmenou v ľudskom mozgu: pohľady na neuroimaging. Aktuálne názory v neurobiológii. 2004;14: 769-776.
- Pascual-Leone JA. Matematický model prechodu v Piagetových vývojových štádiách. Acta Psychologica. 1970;32: 301-345.
- Pecina S, Cagniard B, Berridge KC, Aldridge JW, Zhuang X. Hyperdopaminergné mutantné myši majú vyššie "chcieť", ale nie "sympatie" pre sladké odmeny. Journal of Neuroscience. 2003;23: 9395-9402. [PubMed]
- Reyna VF, Farley F. Riziko a racionálnosť v rozhodovaní adolescentov: Dôsledky pre teóriu, prax a verejnú politiku. Psychologické vedy vo verejnom záujme. 2006;7: 1-44.
- Rosenberg DR, Lewis DA. Postnatálne dozrievanie dopaminergnej inervácie opíc prefrontálnych a motorických kortikúl: imunohistochemická analýza tyrozínhydroxylázy. Journal of Comparative Neurology. 1995;358: 383-400. [PubMed]
- Rubia K, Overmeyer S, Taylor E, Brammer M, Williams SC, Simmons A a kol. Funkčná frontalizácia s vekom: mapovanie trajektórií neurologického vývoja pomocou fMRI. Neuroscience a Biobehavioral Reviews. 2000;24: 13-19. [PubMed]
- Schlaggar BL, Brown TT, Lugar HM, Visscher KM, Miezin FM, Petersen SE. Funkčné neuroanatomické rozdiely medzi dospelými a školopovinnými deťmi pri spracovaní jednotlivých slov. Science. 2002;296: 1476-1479. [PubMed]
- Schultz W. Behaviorálne teórie a neurofyziológia odmeny. Ročné recenzie psychológie. 2006;57: 87-115.
- Sowell ER, Peterson BS, Thompson PM, Vitajte SE, Henkenius AL, Toga AW. Mapovanie kortikálnej zmeny v priebehu celého ľudského života. Nature Neuroscience. 2003;6: 309-315.
- Sowell ER, Thompson PM, Holmes CJ, Jernigan TL, Toga AW. In vivo dôkazy pre dospievanie mozgu po dospievaní v predných a striatálnych oblastiach. Nature Neuroscience. 1999;2: 859-861.
- Sowell ER, Thompson PM, Toga AW. Zmeny v ľudskej kôre počas celého života. Neurológ. 2004;10: 372-392. [PubMed]
- Spear LP. Adolescentný mozog a prejav správania súvisiace s vekom. Neuroscience a Biobehavioral Reviews. 2000;24: 417-463. [PubMed]
- Steinberg L. Riziko v adolescencii: Aké zmeny a prečo? Annals z Newyorskej akadémie vied. 2004;1021: 51-58. [PubMed]
- Steinberg L. Riziko v adolescencii: nové perspektívy v oblasti mozgu a behaviorálnej vedy. Súčasné smery v psychologických vedách. 2007;16: 55-59.
- Steinberg L. Sociálno-neurovedecká perspektíva na riskovanie adolescentov. Vývojová kontrola. 2008;28: 78-106. [Článok bez PMC] [PubMed]
- Tamm L, Menon V, Reiss AL. Zrenie funkcie mozgu spojené s inhibíciou reakcie. Journal of American Academy of Child and Adolescent Psychiatry. 2002;41: 1231-1238. [PubMed]
- Thomas KM, Hunt RH, Vizueta N, Sommer T, Durston S, Yang Y a kol. Evidencia vývojových rozdielov v implicitnom učení sekvencií: Štúdia FMRI u detí a dospelých. Journal of Cognitive Neuroscience. 2004;16: 1339-1351. [PubMed]
- Turkeltaub PE, Gareau L, Flowers DL, Zeffiro TA, Eden GF. Vývoj nervových mechanizmov pre čítanie. Nature Neuroscience. 2003;6: 767-773.
- Volkow ND, Li TK. Drogová závislosť: Neurobiológia správania sa stratila. Príroda Recenzie Neuroscience. 2004;5: 963-970.
- Yurgelun-Todd D. Emocionálne a kognitívne zmeny počas dospievania. Aktuálne stanovisko v neurobiológii. 2007;17: 251-257. [PubMed]
- Zald DH, Boileau I, El-Dearedy W, Gunn R, McGlone F, Dichter GS a kol. Prenos dopamínu v ľudskom striate počas úloh peňažného odmeňovania. Journal of Neuroscience. 2004;24: 4105-4112. [PubMed]