KOMENTÁRE: Jedna z mála štúdií, ktorá sa zaoberá tým, ako abstinencia ovplyvňuje hladiny dopamínových receptorov u primátov.
- D2 receptory sa odrazia späť pomerne rýchlo - menej ako mesiac
- Receptory D1 sú príliš vysoké za mesiac, ale do 90 dní sa odrazia.
- Receptory s vysokým alebo nízkym obsahom D1 môžu byť kľúčom k akútnemu vysadeniu a chute
Thomas JR Beveridge1, Hilary R Smith1, Michael A Nader1 a Linda J. Porrino1
1Oddelenie fyziologie a farmakológie, Centrum neurobiologického vyšetrovania zneužívania drog, Lekárska fakulta Univerzity Wake Forest, Winston-Salem, NC, USA
Korešpondencia: Dr LJ Porrino, Oddelenie fyziologie a farmakológie, Centrum neurobiologického vyšetrovania zneužívania drog, Lekárska fakulta Univerzity Wake Forest, Lekárske centrum Boulevard, Winston-Salem, NC 27157-1083, USA. Tel: + 1 336 716 8575; Fax: + 1 336 716 8501; E-mail: [chránené e-mailom]
Prijaté 29. apríla 2008; Revidované 25. júla 2008; Prijaté 30. júla 2008; Publikované online 3. septembra 2008.
abstraktné
Aj keď dysregulácia v dopamínovom (DA) systéme je charakteristickým znakom chronickej expozície kokaínu, otázka, či tieto zmeny pretrvávajú do abstinencie, zostáva do značnej miery nezodpovedaná. Neľudské primáty predstavujú ideálny model na hodnotenie účinkov abstinencie na DA systém po chronickej expozícii kokaínu. V tejto štúdii sa u samcov opíc rhesus, ktorým sa podáva kokaín (0.3 mg/kg na injekciu, zosilňovače 30 na reláciu) v pevnom intervale 3-minchedule na dni 100 nasledované abstinenciou na dni 30 alebo 90. Ukázalo sa, že toto trvanie kokaínového samopodania už skôr znižuje hustotu receptorov podobných DA D2 a zvyšuje hladiny receptorov podobných D1 a transportérov DA (DAT)., Reakcie kontrolných opíc sa udržiavali prezentáciou potravy podľa rovnakého protokolu a rovnakých období abstinencie. [3H]SCH 23390 sa viaže na Receptory DA D1 po 30 dňoch abstinencie boli významne vyššie vo všetkých častiach striata v porovnaní s kontrolnými zvieratami, zatiaľ čo [3H]Väzba raclopridu na DA D2 receptory sa medzi skupinami nelíšila. [3H]Viazanie WIN 35 428 na DAT bolo tiež významne vyššie počas takmer všetkých častí dorzálneho a ventrálneho striata po 30 dňoch abstinencie. Po 90 dňoch abstinencie sa hladiny DA D1 receptorov a DAT nelíšili od kontrolných hodnôt. Aj keď tieto výsledky naznačujú, že dôjde k regenerácii jednotlivých prvkov systému DA, zdôrazňujú aj dynamický charakter týchto zložiek v počiatočných fázach abstinencie od chronického podania kokaínu.
Kľúčové slová:
kokaín, dopamín, autorádiografia, abstinencia, striatum
ÚVOD
Chronické užívanie kokaínu medzi závislými na ľuďoch je spojené s neuroadaptáciami v dopamínovom (DA) systéme (kliatba et al, 1998; Volkow et al, 1993, 1997). Tieto zahŕňajú zvýšenie hustoty DA transportérov (DAT) a zníženie koncentrácií DA D2 podobných receptorov (málo et al, 1999; Kaša et al, 2002; Volkow et al, 1993). Okrem toho boli pozorované aj zmeny v uvoľňovaní DA. Napríklad výskumní pracovníci, ktorí používajú štúdie pozitrónovej emisnej tomografie (PET) s [11C]racloprid a metylfenidát preukázali zníženie uvoľňovania DA v striatu chronických užívateľov kokaínu (Volkow et al, 1997; Wong et al, 2006). Jedným z problémov je však to, že je často ťažké vylúčiť vplyv iných faktorov, ako je užívanie iných nelegálnych a legálnych drog, rozdiely v predchádzajúcom spôsobe užívania a užívania drog a rozdiely v životnom štýle. Tieto rozdiely, ako aj existencia stavov, ktoré môžu predchádzať užívaniu drog, môžu obmedziť interpretáciu štúdií na ľudských pacientoch.
Nehumánne primátové modely, v ktorých možno systematicky manipulovať s premennými, predstavujú alternatívny prístup k štúdiu dôsledkov chronického podania kokaínu a následnej abstinencie. Predchádzajúce štúdie preukázali, že chronická expozícia kokaínu je sprevádzaná výrazným znížením koncentrácií DA D2 receptorov, ako aj zvýšením hladín D1 receptorov a hustoty DAT (Letchworth et al, 2001; Moore et al, 1998a, 1998b; Nader et al, 2002, 2006). Tieto účinky odzrkadľujú účinky pozorované u ľudí, a tak zdôvodňujú užitočnosť týchto modelov expozície lieku.
Aj keď existujú značné dôkazy o dysregulácii DA systémov, ukázalo sa, že je zložitejšie vyhodnotiť, či existujú nejaké dôkazy na zotavenie po ukončení užívania drog (kliatba et al, 1998; Jacobsen et al, 2000; Volkow et al, 1993) alebo či tieto zmeny pretrvávajú aj po časovom rámci pokračujúcej expozície kokaínu. Modely nehumánnych primátov môžu opäť poskytnúť pohľad do tejto fázy závislosti. Farfel et al (1992) hlásili znížené koncentrácie DAT a receptorov podobných D1 v striatu opíc po abstinencii od chronického nekontrolovaného vystavenia kokaínu. Špecifickú úlohu abstinencie však bolo ťažké určiť z dôvodu chýbajúcich meraní v skupine bez obdobia abstinencie. Podobným spôsobom Melega et al (2008) hlásili významne znížené hladiny DAT v striatu opíc jeleňov po 3 týždňoch abstinencie od eskalujúceho režimu metamfetamínu. Podávanie stimulantov v obidvoch štúdiách však nebolo podmienené. Cesta podania (podmienená vs ukázalo sa, že nemá vplyv na mozog odlišne, pokiaľ ide o obidve uvoľnenia DA (Hemby et al, 1997) a metabolizmus glukózy (Graham a Porrino, 1995; Porrino et al, 2002). Použitie samosprávy v tejto štúdii teda obchádza tento problém. Okrem toho sa intenzívne študovali účinky dlhodobého podania kokaínu na mozgové DA systémy pomocou tohto modelu samopodania u opíc rhesus, čím sa poskytuje základ pre stanovenie neuroadaptácií, ku ktorým dochádza počas abstinencie.
Účelom týchto štúdií bolo preto zistiť, či sa zmeny v koncentráciách receptorov DAT a DA D1 a D2, ktoré sa predtým preukázali u zvierat vystavených samopodaniu kokaínu (Letchworth et al, 2001; Moore et al, 1998a, 1998b; Nader et al, 2002) by sa po predĺžení zdržania zdržalo. Na základe štúdií s užívateľmi drog u ľudí (porovn Volkow et al, 1993), predpokladali sme, že tieto zmeny v systéme DA budú pretrvávať aj po 3 mesiacoch abstinencie. Za týmto účelom si opice podávajú kokaín na relácie 100 s úplným príjmom 900u. mg/kg, nasledované 30 alebo 90 dňami abstinencie od drogy. Receptory DA D1 a D2, ako aj DAT boli merané kvantitatívne in vitro autorádiografia receptorov.
METÓDY
Predmety
Celkom 17 experimentálne naivných dospelých samcov makak rhesus (Macaca mulatta) s hmotnosťou medzi 7.7 a 13 kg (priemer ± SD, 10.2 ± 1.32) na začiatku štúdie slúžili ako subjekty. Všetky postupy sa vykonali v súlade so zavedenými postupmi opísanými v Sprievodca národnými inštitútmi zdravia pre starostlivosť a používanie laboratórnych zvierat, Okrem toho boli všetky postupy preskúmané a schválené Výborom pre starostlivosť o zvieratá a ich využitie na Wake Forest University. Opice boli jednotlivo umiestnené v klietkach z nehrdzavejúcej ocele s vodou podľa chuti; zvieratá mali medzi sebou fyzický a vizuálny kontakt. Ich telesná hmotnosť sa udržiavala na približne 90-95% kŕmnych závaží z peliet s príchuťou banánu získaných počas experimentov a doplnkovým kŕmením Lab Diet Monkey Chow, pokiaľ nie skôr ako 30 min. po relácii. Okrem toho dostávali čerstvé ovocie alebo arašidy najmenej trikrát týždenne. Každá opica bola vážená raz týždenne, a ak to bolo potrebné, jej strava bola upravená tak, aby sa udržala stabilná hmotnosť.
Behaviorálne prístroje
Experimentálne sedenia sa uskutočňovali vo vetraných a zvukovo tlmených operačných komorách (1.5 × 0.74 × 0.76 m; Med Associates Inc., East Fairfield, VT) navrhnutý na umiestnenie stoličky pre primátov (Model R001; Primate Products, Redwood City, CA). Komora obsahovala spravodajský panel (48 × 69) cm), ktorý pozostával z dvoch zatiahnuteľných pák (5 cm šírka) a tri stimulačné svetlá. Páky boli umiestnené v dosahu opice, ktorá sedela na primátovom kresle. Potravinové pelety s príchuťou banánu (1 g; Bio-Serv, Frenchtown, NJ) boli dodané z podávača umiestneného na hornej časti komory. Peristaltická infúzna pumpa (7531-10; Cole-Parmer Co., Chicago, IL) sa použila na dodávanie injekcií liečiva rýchlosťou približne 1. ml na 10 s tými zvieratami, ktoré si sami podávajú kokaín. Činnosť komôr a získavanie údajov sa uskutočňovali počítačovým systémom Power Macintosh s rozhraním (Med Associates Inc.).
Chirurgické postupy
Všetky opice, vrátane kontrol, sa chirurgicky pripravili za sterilných podmienok pomocou zavedených intravenóznych katétrov a vaskulárnych prístupových portov (Model GPV; Access Technologies, Skokie, IL). Opice sa anestetizovali kombináciou ketamínu (15 mg/kg, im) a butorfanolu (0.03 mg/kg, im) a urobil sa rez v femorálnej žile. Po tupej disekcii a izolácii žily sa proximálny koniec katétra vložil do žily na vzdialenosť vypočítanú tak, aby končila v dolnej dutej vene. Distálny koniec katétra bol subkutánne navlečený do zárezu, ktorý sa mierne urobil mimo stredovej línie chrbta. Vaskulárny prístupový port bol umiestnený vo vrecku vytvorenom tupou disekciou v blízkosti tohto rezu. Opiciam bol podaný 24 – 48 h čas regenerácie pred návratom k reakcii zosilnenej potravou. Približne 5 dní pred terminálnym zákrokom sa každej opici implantoval chronický katéter do okolitej femorálnej artérie, aby sa odobrali načasované vzorky arteriálnej krvi. Chirurgické postupy boli rovnaké ako pri žilových katétroch. V deň poslednej relácie sa vykonala terminálna štúdia metabolizmu glukózy v mozgu, pri ktorej sa opiciam injekčne podala 2-[14C]deoxyglukóza (2-DG) približne 2 min. po ukončení relácie a krvné vzorky sa získali arteriálnym katétrom cez 45 min. obdobie (pozri Beveridge et al, 2006 pre viac informácií). Údaje o metabolizme z týchto štúdií sa tu neuvádzajú.
Postupy samosprávy
Opice boli spočiatku trénované tak, aby reagovali na jednu z dvoch pák zosilnením každej reakcie na správnej páke potravou. Počas približne 3-týždňového obdobia sa interval medzi dostupnosťou potravinových peliet postupne zvyšoval, až kým sa nedosiahol interval 3-min (tj časový plán zosilnenia 3-min v pevnom intervale; FI 3-min). Za podmienok konečného harmonogramu je prvá reakcia na páke po 3 min viedlo k dodaniu potravinovej pelety; relácie sa skončili po potravinových prezentáciách 30. Na konci každej relácie boli páčky odozvy zasunuté, zhasnuté svetlá a stimulačné svetlá a zvieratá zostali v tmavej komore približne 30. min. predtým, ako sa vrátili do svojich domácich klietok. Všetky opice reagovali podľa harmonogramu FI 3-min prezentácie jedla najmenej na 20 reláciách, kým sa nedosiahla stabilná výkonnosť (± 20)% priemer za tri po sebe nasledujúce relácie, bez trendov v miere odozvy). Keď bola reakcia na udržiavanie potravy stabilná, napájač bol odpojený a účinky zániku na reakciu boli skúmané počas piatich po sebe nasledujúcich relácií, po ktorých bola reakcia obnovená a udržiavaná prezentáciou potravy.
Po stanovení základnej výkonnosti boli všetky opice chirurgicky pripravené venóznymi katétrami, ako je opísané vyššie, a náhodne rozdelené do jednej z troch skupín. Jedna skupina opíc slúžila ako kontrola a naďalej reagovala podľa harmonogramu prezentácie jedla FI 3-min na celkový počet relácií 100 (N=6). Zostávajúce opice 11 boli priradené ku skupinám, kde sa podáva kokaín samostatne (0.3 mg/kg na injekciu). Pretože 0.3 mg/kg kokaínu na injekciu sa považovala za vysokú dávku pre opice, ktoré predtým neboli na kokaíne, pre väčšinu zvierat sa táto dávka dosiahla do dvoch relácií, keď sa opici umožnilo samoaplikáciu 0.1u. mg/kg kokaínu. Po chirurgickom zákroku (približne 4 - 6 dní) sa výkonnosť udržiavaná na jedle mohla stabilizovať pred začatím relácií samoaplikácie kokaínu. Pred každým pokusom bol chrbát zvieraťa vyčistený pomocou 95% Do otvoru vedúceho k venóznemu katétru sa vložil etanol a betadín a do čela vedúceho k venóznemu katéteru sa vložila meracia ihla 22 (model PG20-125), pričom k katétru sa pripojila infúzna pumpa obsahujúca roztok kokaínu. Pred začiatkom relácie bolo čerpadlo prevádzkované približne 3 s, naplnenie portu dávkou kokaínu, ktorá bola k dispozícii počas experimentálnej relácie. Relácie sa skončili po injekciách 30; ako v kontrolných podmienkach, opice zostali v tmavej komore približne 30 min. Na konci každej relácie bol prístav naplnený heparinizovaným soľným roztokom (100 U/ml), aby sa zabránilo zrážaniu.
Experimentálne relácie sa uskutočňovali každý deň približne v rovnakom čase a pokračovali celkovo 100 relácií. Po ukončení relácií 100 sa zaviedla abstinenčná perióda dní 30 alebo 90, počas ktorých sa katétre denne prepláchli heparinizovaným soľným roztokom, ale neuskutočňovali sa žiadne relácie samoaplnenia kokaínu alebo jedla. Pre kontrolnú skupinu boli u štyroch zvierat uložené obdobia abstinencie 30 dní a u ostatných dvoch 90 dní. V prípade kokaínovej skupiny sa osem zvierat uložilo obdobie abstinencie 30 dní a tri 90 dni. Na konci obdobia abstinencie sa uskutočnilo jedno záverečné samo-podávanie (kontrola potravín alebo kokaín) a postup 2-DG sa začal bezprostredne po tomto zasadnutí. U dvoch kontrolných zvierat a štyroch zvierat, ktoré si podali kokaín v skupine s abstinenciou za deň 30, sa na záverečnej schôdzi nedostal žiadny kokaín. Zvieratá boli humánne usmrtené predávkovaním pentobarbitalom (100 mg/kg, iv) na konci 45 min. doba vychytávania indikátorov.
Spracovanie tkanív
Po usmrtení boli mozgy okamžite odstránené, blokované a zmrazené v izopentáne pri -35 až -55 ° C a potom uložené pri -80 ° C. Tkanivové bloky obsahujúce striatum sa potom narezali v kryostate pri -20 ° C v koronálnej rovine na 20 μm rezy, zhromaždené na elektrostaticky nabitých podložných sklíčkach, vysušené vo vákuu cez noc pri teplote 4 ° C, potom skladované pri teplote -80 ° C až do spracovania na autorádiografiu. Z častí caudátového jadra sa odobrali rezy mozgu, putamény a jadrá accumbens, ktoré ležia rostrálne k prednej komisii. Táto oblasť sa označuje ako predkompurálne striatum. Ďalej boli s ohľadom na jadro accumbens označené rostrálne a kaudálne hladiny prekomissurálneho striatu. Rostrálne predkompurálne striatum je oblasť, v ktorej nie je jadro accumbens diferencované na odlišné podsúbory škrupiny a jadra. Kaudálny precommissurálny striatum je oblasť zhodná s výskytom škrupiny a jadra nucleus accumbens, ktoré je pozadu pred vznikom čuchového tuberkulu. Pre každú z väzobných štúdií sa odobrali dve susediace rezy na každej z piatich úrovní (dve rostrálne a tri kaudálne) cez predkompurálne striatum, čo bolo celkom 10 rezov na zviera.
Väzba na receptor D1
Hustoty väzbového miesta DA D1 receptora boli stanovené pomocou [3H]SCH 23390 (špecifická aktivita - 85 Ci/mmol; PerkinElmer, Boston, MA), kvantitatívne in vitro autorádiografia receptorov podľa postupov upravených v Lidow et al (1991) a Nader et al (2002), Rezy boli preinkubované pre 20 min v pufri (50 mM Tris, 120 mM NaCl, 5 mM KCl, 2 mM CaCl2, 1 mM MgCl2, pH 7.4, 25 ° C) na odstránenie endogénneho DA, kokaínu a [14C] z postupu 2-DG. Rezy sa potom inkubovali pre 30 min. v rovnakom pufri, pH 7.4, 25 ° C, obsahujúci 1 mM kyselina askorbová, 40 nM ketanserín a 1 nM [3H]SCH 23390. Po inkubácii boli rezy dvakrát premyté na 20 v pufri obsahujúcom 1 mM kyselina askorbová pri pH 7.4, 4 ° C, potom sa ponorila do destilovanej vody pri teplote 4 ° C a vysušila sa pod prúdom chladného vzduchu. Nešpecifická väzba bola definovaná inkubáciou susedných rezov v inkubačnom roztoku v prítomnosti 5 μM (+) -Butaclamol. Profily spolu s kalibrovanými [3H] autorádiografické štandardy (Amersham, Piscataway, NJ) boli nanesené na film Kodak Biomax MR (Fisher Scientific, Pittsburgh, PA) na 6 týždne.
Väzba na receptor D2
Hustota a distribúcia väzobných miest pre receptor DA D2 sa stanovili pomocou [3H]racloprid (špecifická aktivita, 87 Ci/mmol; PerkinElmer) podľa postupov upravených v Lidow et al (1991) a Nader et al (2002), Rezy boli preinkubované pre 20 min v pufri (50 mM Tris, 120 mM NaCl, 5 mM KCl, pH 7.4, 25 ° C) na odstránenie endogénneho DA, kokaínu a [14C] z postupu 2-DG. Sklíčka sa potom inkubovali pre 30 min v rovnakom tlmivom roztoku obsahujúcom 5 mM kyselina askorbová a 2 nM [3H]Raclopride. Rezy sa prepláchli 3 x 2 min. v pufri pri pH 7.4, 4 ° C, potom sa ponoril do destilovanej vody pri teplote 4 ° C a vysušil sa pod prúdom chladného vzduchu. Nešpecifická väzba bola definovaná inkubáciou susedných rezov v inkubačnom roztoku v prítomnosti 1 μM (+) -Butaclamol. Profily spolu s kalibrovanými [3H] autorádiografické štandardy boli nanesené na film Kodak Biomax MR po dobu 8 týždňov.
Väzba dopamínového transportéra
Hustota DAT väzbových miest bola stanovená pomocou [3H]VYHRAJTE 35,428 (špecifická aktivita, 87 Ci/mmol; Autoradiografia podľa postupov upravených v Canfield et al (1990) a Letchworth et al (2001), Tkanivové rezy boli preinkubované v pufri (50) mM Tris, 100 mM NaCI, pH 7.4, 4 ° C) pre 20 min na odstránenie zvyškov DA, kokaínu a [14C] z postupu 2-DG. Rezy sa potom inkubovali pre 2 h pri 4 ° C v rovnakom pufri obsahujúcom 5 nM [3H]VYHRAJTE 35 428. Rezy boli opláchnuté celkovo 2 min. v pufri pri 4 ° C, potom ponorený do destilovanej vody pri 4 ° C a vysušený pod prúdom studeného vzduchu. Nešpecifická väzba bola definovaná inkubáciou susedných rezov v inkubačnom roztoku v prítomnosti 30 μM kokaín. Profily spolu s kalibrovanými [3H] autorádiografické štandardy boli nanesené na film Kodak Biomax MR po dobu 6 týždňov.
Denzitometria a analýza údajov
Filmy sa vyvíjali s vývojárom Kodak GBX, stopbath a Rapid Fixer (VWR, West Chester, PA) a potom sa opláchli. Analýza autorádiogramov sa uskutočňovala pomocou kvantitatívnej denzitometrie s počítačovým systémom spracovania obrazu (MCID, Imaging Research; InterFocus Imaging Ltd, Cambridge, UK). Hodnoty optickej hustoty boli prevedené na fmol/mg (vlhkého tkaniva) vo vzťahu k kalibrovanému [3H] normy. Špecifická väzba sa stanovila digitálnym odčítaním obrazov nešpecifickej väzby od superponovaných susedných obrazov celkovej väzby. Štruktúry boli identifikované Nissl zafarbením rezov susediacich so sekciami analyzovanými na väzbu na receptor. Dáta z každého testu boli analyzované nezávisle pomocou jednosmernej analýzy rozptylu nasledovanej najmenšími štvorcami post hoc testy na viacnásobné porovnania. Každá oblasť pozostávala zo samostatnej analýzy. Pretože údaje o väzbe získané od kontrolných zvierat, ktoré sa zdržiavali počas 30 a 90 dní, sa navzájom významne nelíšili, podobne ako v predchádzajúcich štúdiách (Nader et al, 2002), boli údaje z kontrolných skupín kombinované. Okrem toho sa nezistili žiadne významné rozdiely medzi údajmi od tých zvierat, ktoré dostali kokaín, a tými, ktoré nedostali na svojom poslednom zasadnutí, takže údaje z týchto skupín boli tiež kombinované.
VÝSLEDKY
Účinky abstinencie od chronického podania kokaínu počas 30 dní
Koncentrácie [3H]Väzba SCH 23390 na receptory DA D1 v predkomurálnom striatu je uvedená v Tabuľka 1, Špecifická väzba [3H]SCH 23390 predstavoval viac ako 90% z celkovej väzby. V súlade s predchádzajúcimi správami (Moore et al, 1998a; Nader et al, 2002), väzba [3H]Receptory SCH 23390 na D1 u kontrolných zvierat, ktoré neboli vystavené liečivu, boli heterogénne s výraznými rozdielmi v stupni väzby medzi podoblasťami striata. Značenie bolo hustejšie vo viac rostrálnych a stredných častiach v celom striate.
![Tabuľka 1 - Účinok dní 30 alebo 90 po abstinencii po chronickom podávaní kokaínu na hustoty [lsqb] 3H [rsqb] SCH 23390 Väzba na receptory D1 v predkomurálnom striatu opíc Rhesus - Bohužiaľ nie sme schopní poskytnúť prístupný alternatívny text pre tento. Ak potrebujete pomoc s prístupom k tomuto obrázku, kontaktujte help@nature.com alebo autora](https://yourbrainonporn.com/wp-content/uploads/2020/01/table_thumb.gif)
Po 30 dňoch abstinencie od vystavenia kokaínu bola väzba na receptory D1 charakterizovaná rozsiahlym zvýšením v celom rostrálno-kaudálnom rozsahu predkomurálneho striata v porovnaní s väzbovými hustotami u kontrolných zvierat, ktoré neboli vystavené pôsobeniu liečiva (Tabuľka 1; Obrázok 1). Vo viac rostrálnom striatu boli koncentrácie v jadre kaudátu vrátane dorsolaterálu významne vyššie (+27%), centrálna (+27%), dorzomedial (+27%) a ventromedial (+23%), ako aj v chrbtovej (+17%), centrálna (+22%) a ventrálne (+23%) časti putamenu v porovnaní s hustotou kontrolných zvierat bez vystavenia liečivu. Významné zvýšenia boli tiež zrejmé v nucleus accumbens (+23%) na tejto úrovni. Na úrovni striata, kde je jadro a obal jadra accumbens najviac diferencované, boli hustoty receptorov podobných D1 tiež významne vyššie v dorsolaterálnom (+31%), centrálna (+29%), dorzomedial (+30%) a ventromedial (+18%) jadro caudate, ako aj dorzálne (+23%), centrálna (+29%) a ventrálne (+28%) putamen v porovnaní s hustotou kontrol, ktoré neboli vystavené účinkom liečiv. V rámci ventrálneho striata na tejto úrovni je koncentrácia D1 väzbové miesta pre receptory boli vyššie v jadre accumbens (+45%) a shell (+20%), ako aj v čuchovom tuberkule (+26%) v porovnaní s hustotou kontrol.
Obrázok 1.
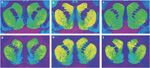 Reprezentatívne autorádiogramy [3H] Schéma SCH 23390 na receptory D1 (horný panel) a [3H]VYHRAJTE väzbu 35428 na dopamínové transportéry (dolný panel) v koronálnych rezoch striatum opíc rhesus. a, d) kontrolné zviera reagujúce na zosilnenie potravy. (b, e) Zviera, ktoré si samo podáva kokaín, s 30 dňovou zdržanlivosťou. (c, f) Zviera, ktoré sa podáva kokaín s 90 dňovou abstinenciou.
Reprezentatívne autorádiogramy [3H] Schéma SCH 23390 na receptory D1 (horný panel) a [3H]VYHRAJTE väzbu 35428 na dopamínové transportéry (dolný panel) v koronálnych rezoch striatum opíc rhesus. a, d) kontrolné zviera reagujúce na zosilnenie potravy. (b, e) Zviera, ktoré si samo podáva kokaín, s 30 dňovou zdržanlivosťou. (c, f) Zviera, ktoré sa podáva kokaín s 90 dňovou abstinenciou.
Koncentrácie [3H]Väzba raclopridu na receptory DA D2 v predkomurálnom striatu je uvedená v Tabuľka 2, Špecifická väzba s [3H]racloprid predstavoval viac ako 90% z celkovej väzby. Distribúcia [3H]Väzba raclopridu na receptory D2 bola tiež heterogénna v rámci subregiónov dorzálneho a ventrálneho striata, ako v predchádzajúcich správach (Moore et al, 1998b; Nader et al, 2002). U kontrol, ktoré neboli vystavené liečivu, boli v dorzálnom porovnaní s ventrálnym striatom prítomné vyššie koncentrácie väzbových miest D2. Okrem toho sa preukázal stredný až bočný gradient s vyššími koncentráciami väzobných miest prítomných vo viac bočných častiach striata.
Po 30 dňoch abstinencie sa hladiny väzby receptora D2 u zvierat vystavených kokaínu a zvieratám posilnených potravou vo väčšine oblastí striata navzájom významne nelíšili. Vo ventrálnych putamenoch boli pozorované vyššie koncentrácie väzbových miest (+10%) a predné jadro accumbens (+12%) v tkanive opíc vystavených kokaínu v porovnaní s kontrolami. Neboli zaznamenané žiadne ďalšie významné rozdiely.
Koncentrácie [3H]Väzba WIN 35428 na DAT v predkomurálnom striatume je uvedená v Tabuľka 3, V súlade s predchádzajúcimi správami (Letchworth et al, 2001), väzba k miestam DAT u zvierat, ktoré neboli vystavené pôsobeniu liečiva, bola vyššia v dorzálnej v porovnaní s ventrálnym striatom. V jadre accumbens boli pozorované vyššie hustoty v jadre v porovnaní s delením škrupiny. Nakoniec nešpecifická väzba predstavuje menej ako 10% z celkového počtu.
Po 30 dňoch abstinencie od vystavenia kokaínu bola väzba na DAT významne väčšia vo väčšine oblastí striatum rostrálneho k prednej komisii v porovnaní s väzbou na kontrolných zvieratách, ktoré neboli vystavené liečivu (Obrázok 1). Konkrétne boli koncentrácie väzbových miest DAT na rostrálnych hladinách v centre (+22%), dorzomedial (+25%) a ventromedial (+28%) jadro caudate a v dorzálnom (+16%) a centrálne (+23%) putamen v porovnaní s kontrolami posilnenými potravinami. Okrem toho sa tiež významne zvýšila väzba na DAT v prednom jadre accumbens (+37%) v porovnaní s kokaínom v porovnaní s opicami posilnenými potravinami. Vo viac kaudálnych častiach predkomurálneho striata boli hustoty väzbových miest DAT významne vyššie v jadre centrálneho kaudátu (+21%) a putamen, centrálny (+20%) a ventrálne (+19%; Obrázok 1). V rámci ventrálneho striatu na tejto úrovni bola väzba na DAT v jadre nucleus accumbens významne vyššia (+20%) a čuchový tuberkul (+24%) v tkanive z kokaínu- vs opice zosilnené potravinami.
Účinky abstinencie od chronického podania kokaínu počas 90 dní
Na rozdiel od rozšírených rozdielov v hustote väzobných miest receptora D1 pozorovaných u zvierat exponovaných kokaínu po 30 dňoch abstinencie, po 90 dňoch abstinencie neboli žiadne významné rozdiely v porovnaní s kontrolami posilnenými potravou v ktorejkoľvek časti precommissurálnej liečby. striatum (Tabuľka 1; Obrázok 1). Podobne koncentrácie [3H]Väzba raclopridu na DA D2 receptory v precommissurálnom striatu po 90 dňoch abstinencie sa tiež významne nelíšila od väzby u zvierat, ktoré neboli vystavené drogám (Tabuľka 2).
Koncentrácie [3H]Väzba WIN 35428 na DAT vykazovala podobný obrazec ako v prípade receptorov D1 a D2. Po 90 dňoch abstinencie neboli pozorované významné rozdiely v hustote DAT medzi opicami vystavenými kokaínu po XNUMX dňoch abstinencie v porovnaní s hladinami v kontrolách, ktoré neboli vystavené účinkom liečiva (Tabuľka 3; Obrázok 1), hoci treba poznamenať, že v prednom jadre accumbens bol trend k vyšším úrovniam väzby.
DISKUSIA
Predchádzajúce štúdie z našej skupiny ukázali, že chronická expozícia kokaínovému samovzniknutiu je sprevádzaná významnou dysreguláciou DA systému nehumánnych primátov (Letchworth et al, 2001; Moore et al, 1998a, 1998b; Nader et al, 2002, 2006). Výsledky tejto štúdie ukazujú, že táto deregulácia zostáva zrejmá aj po ukončení vystavenia kokaínu. Po 30 dňoch abstinencie boli koncentrácie DA D1 receptorov a DAT signifikantne zvýšené v celom striatu opíc s históriou chronického podania kokaínu v porovnaní s kontrolami zosilnenými potravou. Táto štúdia však tiež poskytuje jasný dôkaz zotavenia v systéme DA po dlhšej dobe abstinencie (dni 90), o čom svedčí nedostatok významných rozdielov medzi zvieratami vystavenými kokaínu a kontrolnými zvieratami v tomto časovom okamihu. Tieto údaje naznačujú, že expozícia kokaínu nemusí spôsobiť trvalé zmeny v systéme DA, ale že k zotaveniu môže dôjsť pri dlhodobej abstinencii od užívania drog.
Deregulácia koncentrácií DAT tu uvedená po abstinencii je v súlade s predchádzajúcimi správami u primátov (okrem človeka) (Letchworth et al, 2001), ktoré vykazovali významné zvýšenie hustoty väzbových miest DAT vo ventrálnom aj dorzálnom striate. Aj keď to nie je výslovne testované, zdá sa, že po ukončení vystavenia účinkom liečiva je zvýšenie hustoty väzbového miesta DAT prinajmenšom také veľké a rozsiahlejšie v rámci oblastí striata, ako sú tie, ktoré boli zaznamenané bez ochrannej lehoty (Letchworth et al, 2001). Podobne zvýšené koncentrácie väzobných miest receptora D1 pozorované tu po 30 dňoch abstinencie sú tiež konzistentné s predchádzajúcimi štúdiami, ktoré preukázali zvýšené hustoty väzby receptora podobného D1 v striatu primátov iného ako ľudského pôvodu vystaveného rovnakému režimu samoaplikácie kokaínu (Nader et al, 2002). Na rozdiel od toho neboli pozorované žiadne významné rozdiely medzi hladinami väzobných hustôt podobných receptorom D2 v striatu u zvierat vystavených kokaínu a kontrolných zvierat. Tento nedostatok deregulácie bol prítomný, napriek výraznému poklesu koncentrácií receptorov D2, ktoré boli hlásené u oboch závislých (Volkow et al, 1993) a živočíšne modely samopodania kokaínu (Moore et al, 1998a, 1998b; Nader et al, 2002, 2006). Predložené údaje teda naznačujú rýchlejšiu normalizáciu kontrolných hladín v tomto systéme v porovnaní s receptormi podobnými D1 a DAT. Celkovo možno konštatovať, že zmeny v receptoroch DA a DAT jasne ukazujú, že obdobie bezprostredne po ukončení samoaplikácie kokaínu je vysoko labilné a dochádza k značným zmenám v regulácii systému DA, ale po ňom nasleduje opätovná regulácia systém sa blíži k normálnejšej distribúcii DA receptorov a DAT po dlhšej abstinencii.
Zmeny receptora D1
Rozsiahle zvýšenia hustoty D1 receptorov pozorované tu po ukončení užívania kokaínu sú v súlade so správami preukazujúcimi zvýšenú citlivosť D1 receptorov počas vysadenia. Meranie citlivosti D1 sa uskutočnilo v štúdii uskutočnenej spoločnosťou Henry a Biely (1991), pri ktorom sa zistilo, že záznamy jednotkových jednotiek neurónov v nucleus accumbens sú citlivejšie na agonistu D1 receptora SKF 38393 po chronických denných injekciách kokaínu v porovnaní s kontrolami ošetrenými soľným roztokom. Tento účinok bol pretrvávajúci v tom, že zvýšená citlivosť zostala zrejmá až do jedného mesiaca po vysadení. Autori predpokladali, že senzibilizácia receptorov D1 bola spôsobená subcitlivosťou autoreceptorov D2 v somatodendritickej oblasti A10, čím sa znížil tok inhibičných impulzov v celom systéme mezoaccumbens DA (Henry a White, 1991). Údaje z tejto štúdie naznačujú zvýšenie väzobných miest receptora D1 v abstinencii, čo by mohlo vysvetliť túto zvýšenú citlivosť dopaminergných neurónov na priamo pôsobiaceho agonistu receptora D1. Zvýšený účinok SKF 38393 na neuróny nucleus accumbens nebol zrejmý dva mesiace po vysadení, čo naznačuje, že došlo k obnoveniu citlivosti receptora D1 (Henry a White, 1991); výsledok zodpovedajúci obnoveniu hustôt receptora D1, ktorý bol zaznamenaný v tejto štúdii po 90 dňoch abstinencie. Iné správy tiež podporujú dôležitú úlohu receptorov D1 pri relapse. Priama stimulácia receptorov D1 v škrupine nucleus accumbens môže obnoviť hľadanie kokaínu u abstinentných hlodavcov (Schmidt et al, 2006). Literatúra je však trochu nekonzistentná v tom, že tak agonisti, ako aj antagonisti D1u môžu oslabiť hľadanie liekov vyvolaných kokaínovými prvočíslami alebo stimulmi súvisiacimi s kokaínom (Alleweireldt et al, 2002; De Vries et al, 1999; Khroyan et al, 2000; Vlastné et al, 1996; Biely et al, 2001). V poslednej dobe, Khroyan et al (2003) uviedli, že agonisty a antagonisty D1 znižujú relaps v modeli hľadania kokaínu u nehumánnych primátov. Títo autori naznačujú, že môže byť kritický rozsah aktivity receptora D1 potrebný na vyhľadávanie kokaínu, a že tak antagonisti, ako aj agonisti môžu presunúť aktivitu z tohto okna. Zvýšené koncentrácie receptorov D1, ktoré sprevádzajú abstinenciu, by mohli tento rozsah modifikovať, čo by malo za následok zmenu citlivosti tohto systému. Ďalším aspektom je, že aktivita D1 môže pôsobiť tak, že moduluje aktivitu na D2 receptoroch (Nolan et al, 2007; Ruskin et al, 1999; Walters et al, 1987). Predložené údaje naznačujú, že pomer receptorov D1 k D2 sa mení v priebehu abstinencie, a preto môže zmeniť účinnosť tejto modulácie.
Aj keď na rozdiel od súčasných údajov boli hlásené údaje o znížených hladinách D1 receptorov po chronickom podaní kokaínu (Moore et al, 1998a), existujú značné rozdiely medzi týmito štúdiami, ako je dávka a dĺžka expozície kokaínu, celkový príjem a porovnávacie kontrolné skupiny. Celkovo teda konvergenčné dôkazy silne naznačujú, že systém D1 je po ukončení chronického podávania kokaínu značne rozšírený.
Dopamínové transportné zmeny
Nálezy zvýšených koncentrácií DAT naprieč striatom u zvierat vystavených kokaínu po ukončení užívania drog rozširujú zistenia z našich predchádzajúcich štúdií, ktoré ukazujú zvýšené hladiny väzbových miest DAT, ktoré sprevádzajú samopodávanie kokaínu u primátov iných ako človek. Súčasné údaje ukazujú, že táto dysregulácia pretrváva počas počiatočných fáz abstinencie. Ďalej naznačujú, že návrat k kontrolným hladinám prebieha pomerne dlhým časovým priebehom (v tejto štúdii až do 90 dní). V našich predchádzajúcich štúdiách sme ukázali, že hoci sa pôvodne obmedzoval na prevažne ventrálne striatálne oblasti, zmeny hustoty väzbových miest DAT sa rozšírili tak, aby zahŕňali viac dorzálnych a rostrálnych častí striata s dlhšími obdobiami vystavenia kokaínu (Letchworth et al, 2001; Porrino et al, 2004). V tejto štúdii sa návrat k kontrolným hladinám koncentrácií DAT počas abstinencie zdal byť väčší a rýchlejší v dorzálnom striatume ako v ventrálnom striatume, a preto sa zdalo, že sleduje anatomickú trajektóriu späť k vzoru účinkov vyvolaných chronickou expozíciou kokaínu. ,
Súčasné údaje sú tiež v súlade so správami používateľov humánneho kokaínu (málo et al, 1999; kliatba et al, 1998; Kaša et al, 2002; Staley et al, 1994), ktoré vykázali zvýšené úrovne väzby na miesta DAT v striatu v porovnaní s kontrolami, pričom najvýraznejšie zvýšenie sa nachádzalo vo ventrálnom striatu. Nedávno sa ukázalo, že tieto zvýšenia sú sprevádzané významným znížením väzby vezikulárneho monoamínového transportéra 2 (VMAT2) (málo et al, 2003), čo naznačuje skutočnú stratu DA neurónov. Autori dospeli k záveru, že zvýšený DAT bol pravdepodobne priamo spôsobený kompenzačnou odpoveďou na farmakologickú blokádu kokaínom, zatiaľ čo pokles VMAT2 pravdepodobnejšie odrážal celkové zmeny metabolizmu DA, čo malo za následok hypodopaminergnú funkciu.
U ľudí závislých na kokaíne bolo hlásené, že majú znížené koncentrácie DA vo ventrálnom striatume, merané pomocou PET, v reakcii na expozíciu metylfenidátu v porovnaní so zdravými kontrolami (Volkow et al, 1997). V poslednej dobe, Martinez et al (2007) uviedli, že užívatelia kokaínu mali otupenú odpoveď na amfetamínovú výzvu vo ventrálnom striatume a putamene. Navyše toto zníženie uvoľňovania DA vyvolaného amfetamínom bolo v korelácii s výberom kokaínu v samostatných reláciách samoaplikácie, takže tí používatelia s najnižším stupňom uvoľňovania DA v reakcii na amfetamín si s najväčšou pravdepodobnosťou vybrali kokaín pred alternatívnym zosilňovačom (Martinez et al, 2007). Túto myšlienku podporujú aj nedávne štúdie na modeloch užívania kokaínu u hlodavcov. Mateo et al (2005)napríklad uviedli, že expozícia chronickému kokaínovému podávaniu je spojená so zmenami vo funkcii DAT. Títo výskumníci ukázali, že východisková absorpcia DA sa zvýšila, čo viedlo k rýchlejšiemu klírensu synaptického DA, a preto sa znížili základné hladiny extracelulárneho DA alebo hypodopaminergný stav. Je teda pravdepodobné, že zvýšené koncentrácie DAT po vysadení z chronického podania kokaínu pozorovaného v tejto štúdii predstavujú kompenzačnú odpoveď, ktorá vedie k nižším východiskovým hladinám extracelulárneho DA.
Zmeny receptora D2
Jedným z výsledkov tejto štúdie bolo, že koncentrácie receptorov D2 sa po 30 dňoch abstinencie vrátili na kontrolné hodnoty v porovnaní s významným poklesom pozorovaným u zvierat bez ochrannej lehoty (Nader et al, 2002). Na rozdiel od súčasného výskumu, štúdie zobrazovania u ľudí všeobecne zistili, že hladiny receptorov D2 sú nižšie ako hladiny kontrol po dlhodobej abstinencii pri chronickej expozícii kokaínu (Martinez et al, 2004; Volkow et al, 1993). Možné vysvetlenia rozdielov medzi týmito štúdiami na ľuďoch a súčasným výskumom na primátoch okrem človeka zahŕňajú rozdiely vo vzore a trvaní príjmu kokaínu, ako aj možnosť existencie nižších hladín receptorov D2 u ľudí závislých na drogách.
V súlade s touto poslednou myšlienkou existujú dôkazy, ktoré naznačujú, že nižšie bazálne hladiny receptorov D2 u zdravých ľudí predpovedajú zvýšenú posilňujúcu účinnosť stimulantov, ako sú napr. methylphenidata (Volkow et al, 1999) a podobne u opíc predpovedali východiskové hladiny D2 receptorov náchylnosť k samoinjikovaniu kokaínu (Morgan et al, 2002; Nader et al, 2006). V súlade s paralelným charakterom týchto nálezov medzi druhmi, obaja ľudské (Volkow et al, 1993) a primátov (okrem človeka) (Nader et al, 2006) zobrazovacie štúdie preukázali nižšie úrovne dostupnosti D2 receptorov po abstinencii od vystavenia kokaínu, Je potrebné uviesť, že v poslednom experimente je uvedený časový rozvrh spolužitia kokaínu (Nader et al, 2006) bol podobný harmonogramu zosilnenia použitému v tejto štúdii. Preto je nepravdepodobné, že by rozdielne výsledky z týchto dvoch štúdií boli spôsobené metodologickými rozdielmi, ako je napríklad harmonogram zosilnenia alebo kumulatívny príjem počas samoaplikácie kokaínu.
Pravdepodobnejšie vysvetlenie zahŕňa funkčnú dynamiku systému DA. Merania dostupnosti D2 receptora s PET boli opísané ako „funkčné“, pretože signál súvisí s množstvom proteínu (v tomto prípade hustotou D2 receptorov) a hladinami cirkulujúceho neurotransmitera. (vidieť Laruelle, 2000; Nader a Czoty, 2008 pre ďalšiu diskusiu). jaNaproti tomu autoradiografia receptora je nekontaminovaná cirkulujúcimi hladinami DA. Preto táto štúdia, spolu s našou predchádzajúcou prácou, naznačuje, že hustoty receptorov D2 sa znižujú samoaplikáciou kokaínu, ale zdá sa, že hladiny receptorov sa počas abstinencie obnovujú. V podobne vykonanej štúdii PET (Nader et al, 2006), zotavenie bolo zaznamenané u troch z piatich opíc. Súčasné zistenia naznačujú, že tieto opice sa pravdepodobne nelíšili v hustotách receptorov D2, ale možno aj citlivosť systému DA (tj hladiny cirkulujúceho DA počas abstinencie) rozlišovala medzi „zotavenými“ a „neobnovenými“ jedincami.
Obmedzenia
Jedným dôležitým obmedzením súčasných štúdií je, že naše štúdie nemôžu riešiť funkčnosť receptorov D1 a D2 alebo DAT. Skôr sme skúmali iba zmeny v hustote receptorových proteínov. Aj keď výsledky majú vplyv na potenciálne úlohy týchto systémov, budú sa vyžadovať ďalšie štúdie, aby sa objasnili dôsledky zmeny tu uvedené na správanie. Ďalším obmedzením tejto štúdie je, že autorádiografické ligandy často nedokážu disociovať medzi intracelulárnymi a cytoplazmatickými polohami svojich cieľov, pretože rádioaktívne značené antagonisty sú často membránovo priepustné. málo et al (2002) preukázali, že chronická expozícia kokaínu vedie k signifikantnému zvýšeniu DAT na povrchu membrány, súčasne so znížením intracelulárnej koncentrácie DAT, v transfekovaných bunkách. V poslednej dobe, Samuvel et al (2008) uvádza podobný nález v preparátoch striatálnej synaptozomálnej potkany. Tieto výsledky naznačujú, že zmeny v distribúcii DAT pozorované v tejto štúdii môžu skôr predstavovať zmeny na povrchu membrány, ako vnútrobunkové miesta.
Nakoniec by sa mala venovať zvýšená pozornosť interpretácii štúdií s predĺženou (90-dňovou) abstinenciou, pretože tieto zistenia vychádzajú z relatívne malej skupiny zvierat (N=3). Napriek malému počtu subjektov boli údaje získané z tejto skupiny dosť konzistentné, ako je možné vidieť na rozptylovom grafe znázornenom na obr. Obrázok 2, Koncentrácia väzbových miest receptora D1 cez striatum vykazovala malú variabilitu v rámci skupín, čo svedčí o spoľahlivosti týchto zistení. Podobná konzistencia bola tiež zrejmá v údajoch z D2 receptorov a DAT väzbových testov. Aj keď je potrebné postupovať opatrne, tieto údaje silne naznačujú, že k obnoveniu koncentrácií DAT a DA receptorov v striate môže dôjsť pri dlhodobej abstinencii.
Obrázok 2.
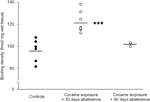 Väzbové hustoty receptorov D1 pre jednotlivé zvieratá boli spriemerované po celom striatu po posilnení jedla (kontroly) alebo po 30 alebo 90 dňoch abstinencie od chronického kokaínového podania. Prostriedky pre skupiny sú znázornené čiernymi pruhmi, ***p<0.001 v porovnaní s kontrolami vystuženými potravinami.
Väzbové hustoty receptorov D1 pre jednotlivé zvieratá boli spriemerované po celom striatu po posilnení jedla (kontroly) alebo po 30 alebo 90 dňoch abstinencie od chronického kokaínového podania. Prostriedky pre skupiny sú znázornené čiernymi pruhmi, ***p<0.001 v porovnaní s kontrolami vystuženými potravinami.
záver
Záverom je možné povedať, že expozícia pri podávaní kokaínu vyvolala významné zmeny v regulácii systémov DA, ktoré pretrvávali počas počiatočných fáz (absencie 30) abstinencie. Toto bolo najzreteľnejšie pri regulácii koncentrácie D1 receptorov a DAT, a to tak z hľadiska rozsahu zmien, ako aj z hľadiska ich topografického rozsahu. Na rozdiel od toho, existujú dôkazy o normalizácii s dlhším trvaním abstinencie z vystavenia kokaínu v tom, že koncentrácie receptorov DAT, D1 a D2 sa po 90 dňoch abstinencie nelíšili od koncentrácií u kontrol, ktoré neboli vystavené účinkom drog. TTieto systémy však nemusia nevyhnutne sledovať rovnaký časový priebeh zotavovania, čo naznačuje, že existuje pravdepodobnosť určitej nestability v regulácii hladín DA, najmä čoskoro v abstinencii., Táto dopaminergná dysregulácia môže ovplyvniť účinnosť akejkoľvek potenciálnej farmakoterapie podávanej abstinujúcim závislým od kokaínu, najmä ak sa liek spolieha na mechanizmus účinku na systém DA.
Poznámky
podstata/KONFLIKT ZÁUJMOV
Autori nemajú žiadny konflikt záujmov, ktorý by mohli zverejniť.
Referencie
- Alleweireldt AT, Weber SM, Kirschner KF, Bullock BL, Neisewander JL (2002). Blokáda alebo stimulácia dopamínových receptorov D1 zmierňuje narastajúce obnovenie zhasnutého správania pri vyhľadávaní kokaínu u potkanov. Psychofarmakológia (Berl) 159: 284–293. | Článok | PubMed | ChemPort |
- Beveridge TJ, Smith HR, Daunais JB, Nader MA, Porrino LJ (2006). Chronické podávanie kokaínu je spojené so zmenenou funkčnou aktivitou v dočasných lalokoch primátov (okrem človeka). Eur J Neurosci 23: 3109–3118. | Článok | PubMed |
- Canfield DR, Spealman RD, Kaufman MJ, Madras BK (1990). Autorádiografická lokalizácia väzbových miest pre kokaín pomocou [3H] CFT ([3H] WIN 35428) v mozgu opice. Synapse 6: 189–195. | Článok | PubMed | ChemPort |
- De Vries TJ, Schoffelmeer AN, Binnekade R, Vanderschuren LJ (1999). Dopaminergné mechanizmy sprostredkujúce motiváciu vyhľadávať kokaín a heroín po dlhodobom ukončení IV podávania liekov. Psychofarmakológia (Berl) 143: 254–260. | Článok | PubMed | ChemPort |
- Farfel GM, Kleven MS, Woolverton WL, Seiden LS, Perry BD (1992). Účinky opakovaných injekcií kokaínu na väzbové miesta pre katecholamínové receptory, väzobné miesta pre transportér dopamínu a správanie u opíc rhesus. Brain Res 578: 235–243. | Článok | PubMed | ChemPort |
- Graham J, Porrino LJ (1995). Neuroanatomické substráty samopodania kokaínu, CRC: Boca Raton, FL.
- Hemby SE, Co C, Koves TR, Smith JE, Dworkin SI (1997). Rozdiely v koncentráciách extracelulárneho dopamínu v jadre sa vyskytujú počas reakcie závislej od kokaínu závislej od reakcie a od reakcie závislej od reakcie. Psychofarmakológia (Berl) 133: 7–16. | Článok | PubMed | ChemPort |
- Henry DJ, White FJ (1991). Opakované podávanie kokaínu spôsobuje trvalé zvýšenie citlivosti dopamínového receptora D1 v jadre potkana. J Pharmacol Exp Ther 258: 882–890. | PubMed | ChemPort |
- Jacobsen LK, Staley JK, Malison RT, Zoghbi SS, Seibyl JP, Kosten TR et al (2000). Zvýšená dostupnosť väzby centrálneho serotonínového transportéra u akútne abstinujúcich pacientov závislých od kokaínu. Am J Psychiatria 157: 1134–1140. | Článok | PubMed | ISI | ChemPort |
- Khroyan TV, Barrett-Larimore RL, Rowlett JK, Spealman RD (2000). Mechanizmy receptorov dopamínu D1 a D2 pri relapse na správanie zamerané na vyhľadávanie kokaínu: účinky selektívnych antagonistov a agonistov. J Pharmacol Exp Ther 294: 680–687. | PubMed | ISI | ChemPort |
- Khroyan TV, Platt DM, Rowlett JK, Spealman RD (2003). Zmiernenie relapsu pri hľadaní kokaínu agonistami a antagonistami receptora dopamínu D1 u primátov (okrem človeka). Psychofarmakológia (Berl) 168: 124–131. | Článok | PubMed | ChemPort |
- Laruelle M (2000). Zobrazovanie synaptickej neurotransmisie pomocou in vivo záväzné súťažné techniky: kritické preskúmanie. J Cereb Blood Flow Metab 20: 423–451. | Článok | PubMed | ISI | ChemPort |
- Letchworth SR, Nader MA, Smith HR, Friedman DP, Porrino LJ (2001). Progresia zmien v hustote väzbového miesta pre dopamínový transportér ako výsledok samoaplikácie kokaínu u opíc rhesus. J Neurosci 21: 2799–2807. | PubMed | ISI | ChemPort |
- Lidow MS, Goldman-Rakic PS, Gallager DW, Rakic P (1991). Distribúcia dopaminergných receptorov v mozgovej kôre primátov: kvantitatívna autorádiografická analýza s použitím [3H] raclopridu, [3H] spiperónu a [3H] SCH 23390. Neurovedy 40: 657–671. | Článok | PubMed | ChemPort |
- Malý KY, Elmer LW, Zhong H, Scheys JO, Zhang L (2002). Indukcia kokaínu transportérom dopamínu na plazmatickú membránu. Mol Pharmacol 61: 436–445. | Článok | PubMed | ISI | ChemPort |
- Malý KY, Krolewski DM, Zhang L, Cassin BJ (2003). Strata striatálneho vezikulárneho monoamínového transportného proteínu (VMAT2) u ľudí s obsahom kokaínu. Am J Psychiatria 160: 47–55. | Článok | PubMed |
- Little KY, Zhang L, Desmond T, Frey KA, Dalack GW, Cassin BJ (1999). Striatálne dopaminergné abnormality u používateľov kokaínu u ľudí. Am J Psychiatria 156: 238–245. | PubMed | ChemPort |
- Malison RT, najlepší SE, van Dyck CH, McCance EF, Wallace EA, Laruelle M. et al (1998). Zvýšené striatálne dopamínové transportéry počas akútnej abstinencie kokaínu merané pomocou [123I] beta-CIT SPECT. Am J Psychiatria 155: 832–834. | PubMed | ChemPort |
- Martinez D, Broft A, Foltin RW, Slifstein M, Hwang DR, Huang Y et al (2004). Závislosť od kokaínu a dostupnosť d2 receptora vo funkčných podtriedach striata: vzťah k správaniu sa pri vyhľadávaní kokaínu. neuropsychofarmakologie 29: 1190–1202. | Článok | PubMed | ISI | ChemPort |
- Martinez D, Narendran R, Foltin RW, Slifstein M, Hwang DR, Broft A et al (2007). Uvoľňovanie dopamínu vyvolané amfetamínom: výrazne oslabené v závislosti od kokaínu a predpovedá výber vlastného podania kokaínu. Am J Psychiatria 164: 622–629. | Článok | PubMed | ISI |
- Mash DC, Pablo J, Ouyang Q, Hearn WL, Izenwasser S (2002). U užívateľov kokaínu je zvýšená transportná funkcia dopamínu. J Neurochem 81: 292–300. | Článok | PubMed | ChemPort |
- Mateo Y, nedostatok CM, Morgan D, Roberts DC, Jones SR (2005). Znížená terminálna funkcia dopamínu a necitlivosť na kokaín po samopodávaní a deprivácii kokaínu. neuropsychofarmakologie 30: 1455–1463. | Článok | PubMed | ISI | ChemPort |
- Melega WP, Jorgensen MJ, Lacan G, Way BM, Pham J, Morton G et al (2008). Pri dlhodobom podávaní metamfetamínu u opice mačiek sa sledujú aspekty expozície človeka: neurotoxicita mozgu a profily správania. neuropsychofarmakologie 33: 1441–1452. | Článok | PubMed | ChemPort |
- Moore RJ, Vinsant SL, Nader MA, Porrino LJ, Friedman DP (1998a). Vplyv podania kokaínu na receptory striatálneho dopamínu D1 u opíc rhesus. Synapse 28: 1–9. | Článok | PubMed | ChemPort |
- Moore RJ, Vinsant SL, Nader MA, Porrino LJ, Friedman DP (1998b). Účinok samopodania kokaínu na dopamínové D2 receptory u opíc rhesus. Synapse 30: 88–96. | Článok | PubMed | ChemPort |
- Morgan D, Grant KA, Gage HD, Mach RH, Kaplan JR, Prioleau O et al (2002). Sociálna dominancia u opíc: dopamínové receptory D2 a samopodávanie kokaínu. Nat Neurosci 5: 169–174. | Článok | PubMed | ISI | ChemPort |
- Nader MA, Czoty PW (2008). Zobrazovanie mozgu u primátov (okrem človeka): vhľad do drogovej závislosti. ILAR J 49: 89–102. | PubMed | ChemPort |
- Nader MA, Daunais JB, Moore T, Nader SH, Moore RJ, Smith HR et al (2002). Účinky samopodania kokaínu na striatálne dopamínové systémy u opíc rhesus: počiatočná a chronická expozícia. neuropsychofarmakologie 27: 35–46. | Článok | PubMed | ISI | ChemPort |
- Nader MA, Morgan D, Gage HD, Nader SH, Calhoun TL, Buchheimer N et al (2006). Zobrazovanie PET dopamínových D2 receptorov počas chronického kokaínového podania opiciam. Nat Neurosci 9: 1050–1056. | Článok | PubMed | ChemPort |
- Nolan EB, Harrison LM, Lahoste GJ, Ruskin DN (2007). Behaviorálna synergia medzi dopamínovými receptormi D (1) a D (2) nezávisí od spojov medzier. Synapse 61: 279–287. | Článok | PubMed | ChemPort |
- Porrino LJ, Lyons D, Miller MD, Smith HR, Friedman DP, Daunais JB et al (2002). Metabolické mapovanie účinkov kokaínu počas počiatočných fáz samoaplikácie u primátov. J Neurosci 22: 7687–7694. | PubMed | ChemPort |
- Porrino LJ, Lyons D, Smith HR, Daunais JB, Nader MA (2004). Podávanie kokaínu vedie k progresívnemu zapojeniu limbických, asociačných a senzorimotorických striatálnych domén. J Neurosci 24: 3554–3562. | Článok | PubMed | ISI | ChemPort |
- Ruskin DN, Bergstrom DA, Walters JR (1999). Viacsekundové oscilácie rýchlosti spaľovania v globus pallidus: synergická modulácia dopamínovými receptormi D1 a D2. J Pharmacol Exp Ther 290: 1493–1501. | PubMed | ISI | ChemPort |
- Samuvel DJ, Jayanthi LD, Manohar S, Kaliyaperumal K, pozri RE, Ramamoorthy S (2008). Dysregulácia obchodovania s dopamínovými transportérmi a ich funkcia po abstinencii od podania kokaínu samcom potkanov: dôkaz rozdielnej regulácie v putách putamenu a jadre accumbens. J Pharmacol Exp Ther 325: 293–301. | Článok | PubMed | ChemPort |
- Schmidt HD, Anderson SM, Pierce RC (2006). Stimulácia dopamínových receptorov podobných D1 alebo D2 v škrupine, ale nie v jadre jadra accumbens, obnovuje správanie pri vyhľadávaní kokaínu u potkanov. Eur J Neurosci 23: 219–228. | Článok | PubMed |
- Self DW, Barnhart WJ, Lehman DA, Nestler EJ (1996). Proti modulácii správania pri vyhľadávaní kokaínu agonistami dopamínového receptora podobného D1 a D2. veda 271: 1586–1589. | Článok | PubMed | ISI | ChemPort |
- Staley JK, Hearn WL, Ruttenber AJ, Wetli CV, Mash DC (1994). Miesta na rozpoznávanie kokaínu s vysokou afinitou na dopamínovom transportéri sú zvýšené u obetí predávkovania smrteľnými kokaínmi. J Pharmacol Exp Ther 271: 1678–1685. | PubMed | ChemPort |
- Volkow ND, Fowler JS, Wang GJ, Hitzemann R, Logan J, Schlyer DJ et al (1993). Znížená dostupnosť dopamínového D2 receptora je spojená so zníženým frontálnym metabolizmom u užívateľov kokaínu. Synapse 14: 169–177. | Článok | PubMed | ISI | ChemPort |
- Volkow ND, Wang GJ, Fowler JS, Logan J, Gatley SJ, Gifford A et al (1999). Predpoveď posilňujúcich reakcií na psychostimulanty u ľudí pomocou hladín receptora dopamínu D2 v mozgu. Am J Psychiatria 156: 1440–1443. | PubMed | ISI | ChemPort |
- Volkow ND, Wang GJ, Fowler JS, Logan J, Gatley SJ, Hitzemann R et al (1997). Znížená striatálna dopaminergná citlivosť u pacientov závislých od detoxifikovaného kokaínu. príroda 386: 830–833. | Článok | PubMed | ISI | ChemPort |
- Walters JR, Bergstrom DA, Carlson JH, Chase TN, Braun AR (1987). Aktivácia dopamínového receptora D1 potrebná na postsynaptickú expresiu agonistických účinkov D2. veda 236: 719–722. | Článok | PubMed | ChemPort |
- Weiss F, Martin-FardonR, CiccocioppoR, KerrTM, Smith DL, Ben-Shahar O (2001). Pretrvávajúca rezistencia voči vyhynutiu správania pri vyhľadávaní kokaínu vyvolaného narážkami súvisiacimi s drogami. neuropsychofarmakologie 25: 361–372. | Článok | PubMed | ISI | ChemPort |
- Wong DF, Kuwabara H, Schretlen DJ, Bonson KR, Zhou Y, Nandi A. et al (2006). Zvýšená obsadenosť dopamínových receptorov v ľudskom striate počas túžby po vyvolaní kokaínu. neuropsychofarmakologie 31: 2716–2727. | Článok | PubMed | ChemPort |
