 Fabrizio Sanna1*,
Fabrizio Sanna1*,  Jessica Bratzu1,
Jessica Bratzu1,  Maria A. Piludu2,
Maria A. Piludu2,  Maria G. Corda2,
Maria G. Corda2,  Maria R. Melis1,
Maria R. Melis1,  Osvaldo Giorgi2 a
Osvaldo Giorgi2 a  Antonio Argiolas1,3
Antonio Argiolas1,3- 1Katedra biomedicínskych vied, Oddelenie neurovedy a klinickej farmakológie a Centrum excelentnosti pre neurobiológiu závislostí, University of Cagliari, Cagliari, Taliansko
- 2Katedra vied o živote a životnom prostredí, Oddelenie farmaceutických, farmakologických a nutraceutických vied, Univerzita v Cagliari, Cagliari, Taliansko
- 3Neurovedný ústav, Národná rada pre výskum, Sekcia Cagliari, Cittadella Universitaria, Cagliari, Taliansko
Potkany rasy Roman High- (RHA) a Low-Avoidance (RLA), ktoré sa líšia v prípade jednotlivo rýchleho alebo slabého získania odpovede na aktívne vyhýbanie sa v kyvadlovej skrini, vykazujú rozdiely v sexuálnej aktivite, keď sú umiestnené v prítomnosti sexuálne vnímavá samica potkana. Potkany RHA skutočne vykazujú vyššiu úroveň sexuálnej motivácie a kopulačného výkonu ako potkany RLA, ktoré pretrvávajú aj po opakovanej sexuálnej aktivite. Tieto rozdiely boli korelované s vyšším tónom mezolimbického dopaminergného systému RHA potkanov oproti RLA potkanom, odhaleným vyšším zvýšením dopamínu zisteného v dialyzáte získanom z jadra accumbens RHA ako RLA potkanov počas sexuálnej aktivity. Táto práca ukazuje, že extracelulárny dopamín a noradrenalín (NA) tiež zvyšujú dialyzát zo stredného prefrontálneho kortexu (mPFC) samcov potkanov RHA a RLA, ktorý je prítomný v neprístupnej samici potkana a výraznejšie počas priamej sexuálnej interakcie. Takéto zvýšenia dopamínu (a jeho hlavného metabolitu 3,4-dihydroxyfenyloctová kyselina, DOPAC) a NA sa našli u sexuálne naivných aj skúsených zvierat, ale boli vyššie: (i) u RHA ako u potkanov RLA; a (ii) u sexuálne skúsených potkanov RHA a RLA ako u ich naivných náprotivkov. Nakoniec, rozdiely v dopamíne a NA v mPFC sa vyskytli súbežne s rozdielmi v sexuálnej aktivite, pretože RHA potkany vykazovali vyššiu hladinu sexuálnej motivácie a kopulačnej výkonnosti ako RLA potkany, a to tak v sexuálne naivných, ako aj v skúsených podmienkach. Tieto výsledky naznačujú, že v mPFC sa vyskytuje aj vyšší dopaminergný tonus, spolu so zvýšeným noradrenergickým tonom, ktorý sa môže podieľať na rôznych kopulačných obrazcoch nájdených u potkanov RHA a RLA, ako sa navrhuje pre mezolimbický dopaminergný systém.
úvod
Rímske línie potkanov rímskej vysokej (RHA) a nízkej eliminácie (RLA), pôvodne vybrané pre rýchle, resp. Slabé získanie aktívnej reakcie na vyhýbanie sa v kyvadlovej skrini (Bignami, 1965; Broadhurst a Bignami, 1965; Driscoll a Bättig, 1982; Fernández-Teruel a kol., 2002; Giorgi a kol., 2007) vykazujú významné rozdiely v sexuálnej aktivite, keď sú umiestnené v prítomnosti sexuálne vnímavej samice potkana. Potkany RHA skutočne vykazujú vyššiu úroveň sexuálnej motivácie ako potkany RLA, ako bolo odhalené vyšším počtom bezkontaktných erekcií penisu, ktoré tieto potkany vykazujú, keď sú umiestnené v neprístupnej vnímavej žene, a lepšie odhalené kopulačné výkony ako potkany RLA. výraznými rozdielmi v početných kopulačných parametroch zistených hlavne v prvom kopulačnom teste medzi dvoma potkanými líniami, ale aj po piatich kopulačných testoch, hoci oslabené (Sanna a kol., 2014a). Medzi najvýraznejšie rozdiely zistené medzi potkanmi RHA a RLA je percento potkanov, ktoré iniciujú montáž a intromitting a ktoré sa ejakulujú v prvom kopulačnom teste, 80% RHA potkanov proti 40% RLA potkanov, v danom poradí (Sanna a kol., 2014a). Veľké množstvo experimentálnych dôkazov naznačuje, že do kopulačných rozdielov medzi potkanmi RHA a RLA je zapojený funkčne odlišný dopaminergný tón. V skutočnosti bolo zistené, že rôzne kopulačné vzorce RHA a RLA potkanov sú rôzne modifikované apomorfínom, zmiešaným agonistom dopamínových receptorov D1 / D2 podávaným v dávkach, ktoré uľahčujú sexuálne správanie, a haloperidolom, antagonistom dopamínových receptorov D2, podávaným na nízke dávky, ktoré potláčajú sexuálne správanie (Sanna a kol., 2014b). V súlade s tým sa zistilo, že RLA potkany sú citlivejšie na uľahčenie a inhibíciu sexuálneho správania vyvolaného apomorfínom a haloperidolom, ako je demonštrované väčšími modifikáciami vyvolanými týmito dvoma liečivami hlavne pri podávaní nižších dávok pri rôznych kopulačných parametroch v Potkany RLA s ohľadom na potkany RHA (Sanna a kol., 2014b). Možno, že pre túto štúdiu je dôležitejšie, rôzne kopulačné vzorce RHA a RLA potkanov súvisia tiež s rozdielmi v aktivite mezolimbických dopaminergných neurónov, ktorých aktivita sa počas sexuálneho správania zvyšuje (Pfaus a kol., 1990; Pleim a kol., 1990; Pfaus a Phillips, 1991; Damsma a kol., 1992; Wenkstern a kol., 1993; Balfour a spol., 2004; Pitchers a kol., 2010, 2013; Beloate a kol., 2016). Napriek tomu, že potkany RHA a RLA, ktoré majú pohlavný styk a sexuálne skúsenosti, majú podobné podobné základné hodnoty extracelulárneho dopamínu v dialyzáte z jadra accumbens, koncentrácie extracelulárneho dopamínu a kyseliny 3,4-dihydroxyfenyloctovej (DOPAC, jedného z hlavných metabolitov) ), sa zistilo, že sa dialyzáty získané z plášťa jadra accumbens naivných a sexuálne skúsených potkanov RHA a RLA rozdielne zvyšujú tak v predikčnej, ako aj konzumnej fáze sexuálneho správania. Vyššie uvedené rozdiely boli výraznejšie medzi sexuálne naivnými potkanmi RHA a RLA, ale pretrvávali medzi sexuálne skúsenými potkanmi RHA a RLA, hoci u týchto potkanov mali tendenciu sa zmenšovať, ako sa zistilo pri rozdieloch v sexuálnom správaní (Sanna a kol., 2015).
Funkčná úloha zvýšenia dopaminergnej aktivity v jadre accumbens pozorovaná počas sexuálnej aktivity (buď v apetitívnej a konzumnej fáze sexuálneho správania) je stále predmetom diskusie, ako aj úloha pozorovaná počas kŕmenia, ktorá má, podobne ako sexuálna aktivita, silný motivačná valencia. Aj keď sa dopamín v jadre accumbens podieľa na motivácii a mezolimbické dopaminergné neuróny sa zvyčajne označujú ako obohacujúce neuróny, v posledných 15 rokoch, nedávne štúdie podporujú hypotézu, že mezolimbický dopamín sa nezúčastňuje primárneho vyjadrenia motivovaného alebo odmeňujúceho správania, ale skôr s učením a pamäťou združení stimulov a odmien (Agmo et al., 1995; Berridge a Robinson, 1998; Ikemoto a Panksepp, 1999; Pitchers a kol., 2013, 2014; Beloate a kol., 2016; Salamone a kol., 2016). V súlade s touto hypotézou sa nedávno zistilo, že blokáda dopamínových receptorov v nucleus accumbens alebo inaktivácia dopaminergných neurónov vo ventrálnej tegmentálnej oblasti nie je schopná zmeniť expresiu chutných a konzumných aspektov kopulačného správania samcov potkanov (Pitchers a kol., 2013, 2014; Beloate a kol., 2016).
Nemožno však vylúčiť úlohu iných oblastí mozgu obsahujúcich dopamín vo vyššie uvedených rozdieloch v sexuálnom správaní medzi potkanmi RHA a RLA. Preto je dobre známe, že dopamín má podporný účinok na predvídaciu a konzumnú fázu sexuálneho správania u laboratórnych zvierat a tiež u ľudí nielen v nucleus accumbens (Everitt, 1990; Pfaus a kol., 1990; Hull a kol., 1991; Pfaus a Everitt, 1995; Melis a Argiolas, 2011), ale aj v iných oblastiach mozgu, ako je stredná predoptická oblasť, hypotalamus a jeho jadrá (tj paraventrikulárne jadro (PVN)); Pfaus a Phillips, 1991; Argiolas a Melis, 1995, 2005, 2013; Hull a kol., 1995, 1999; Melis a Argiolas, 1995; Melis a kol., 2003; Succu a kol., 2007; Pfaus, 2010). Ďalšou oblasťou, ktorá obsahuje dopamín a môže hrať úlohu pri sexuálnom správaní, je stredná prefrontálna kôra (mPFC; Fernández-Guasti a kol., 1994; Agmo a Villalpando, 1995; Agmo et al., 1995; Hernández-Gonzáles a kol., 1998, 2007; Kakeyama a kol., 2003; Balfour a spol., 2006; Afonso a kol., 2007; Davis a kol., 2010; Febo, 2011). V skutočnosti sa v tejto oblasti mozgu nachádzajú nervové zakončenia mezokortikálnych dopamínových neurónov, ktorých bunkové telá sú lokalizované vo ventrálnej oblasti tegmentálu ako mezolimbické dopamínové neuróny. Pokiaľ ide o jadro accumbens, presná úloha tejto oblasti mozgu pri sexuálnom správaní nie je zďaleka jasná. Skutočne sa zistilo, že lézie mPFC zvyčajne nedokážu zmeniť sexuálne správanie samcov potkanov so sexuálne vnímavými samicami (Fernández-Guasti a kol., 1994; Agmo a Villalpando, 1995; Agmo et al., 1995; Hernández-Gonzáles a kol., 1998, 2007; Kakeyama a kol., 2003; Balfour a spol., 2006; Afonso a kol., 2007), ani vyjadrenie podmieneného preferencie miesta pre sexuálnu odmenu (Davis a kol., 2010). Lézie mPFC, ktoré nezmenili vzhľad podmieneného preferencie miesta pre sexuálnu odmenu, však u tých istých zvierat zrušili schopnosť vytvárať podmienenú averziu voči sexuálnej aktivite, keď sú spárované s averzívnymi stimulmi (Davis a kol., 2010) a v mPFC samcov potkanov sa zmerali selektívne bunkové strieľanie počas priblíženia sa samcom potkana k neprístupnej sexuálne vnímavej samici (Febo, 2011). Tieto zistenia viedli k náznaku, že aktivácia mPFC počas sexuálneho správania zohráva skôr úlohu pri integrácii externých a interných informácií na vykonávanie a kontrolu správania zameraného na cieľ, ako pri vyjadrovaní vrodených reakcií na prírodné zosilňovače (pozri pozri časť 4.4). Goto a Grace, 2005). Podobne, spolu s nucleus accumbens, mPFC je súčasťou komplexného nervového systému zapojeného do modulácie motivovaného správania (cielené správanie), čo vyžaduje integráciu kognitívnych informácií z mPFC, emocionálnych informácií z amygdaly a kontextuálnych súvisiace informácie z hippocampu v nucleus accumbens (Goto a Grace, 2005).
Experimentálne dôkazy naznačujú, že uvoľňovanie dopamínu v mPFC je zapojené do adaptívnej regulácie motivovaného správania a predpokladá sa, že deregulácia týchto mechanizmov hrá úlohu v patologických alebo maladaptívnych stavoch, ako sú psychiatrické poruchy, ako je schizofrénia, deficit pozornosti a hyperaktívne poruchy ( ADHD), depresia (Dunlop a Nemeroff, 2007; Masana a kol., 2011) alebo zneužívanie návykových látok a hazardné hry (Everitt a Robbins, 2005). Zaujímavé je, že aktivitu dopamínu uvoľňovaného v mPFC môže ovplyvniť noradrenalín (NA), ktorý je v mPFC prítomný vo vyšších hladinách ako dopamín, a najmä transportér NA (NET), ktorý nie je len hojnejší ako dopamínový transportér (DAT) v mPFC (Carboni a kol., 1990, 2006; Gresch a kol., 1995; Westernik a kol., 1998), ale tiež vykazuje afinitu k dopamínu ešte vyššiu ako pre NA (Horn, 1973). Pretože dopamín a NA často spolupracujú pri mnohých funkciách mPFC, od formovania pracovnej pamäte a pozornosti, od prechodu k reverznému učeniu, inhibícii odozvy a reakcii na stres (pozri Robbins a Arnsten, 2009) sa tým zvyšuje možnosť, že dopamín, samotný alebo spolu s NA, v mPFC môže tiež hrať úlohu v rozdieloch v správaní medzi RHA a RLA potkanmi vrátane tých, ktoré sa nachádzajú v sexuálnom správaní.
Za účelom overenia tejto hypotézy sa študovala aktivita mezokortikálneho dopaminergného systému a noradrenergického systému v mPFC v dvoch líniách potkanov RHA a RLA pomocou intracerebrálnej mikrodialýzy. V stručnosti, dopamín (a jeho hlavný metabolit DOPAC) a NA sa merali v dialyzátoch získaných z prelimbických (PrL) a infralimbických (IL) kompartmentov mPFC sexuálne naivných (napr. Nikdy nevystavených vnímavej žene) a sexuálne skúsených RHA a Potkany RLA (napr. Ktoré sa podrobili piatim predbežným kopulačným testom a vykazujú konštantné úrovne kopulačnej aktivity), keď sa dostali do neprístupnej vnímavej ženy a počas priamej sexuálnej interakcie pomocou vysokotlakovej kvapalinovej chromatografie spojenej s elektrochemickou detekciou (HPLC-ECD).
Materiály a metódy
zver
Plemenné samce potkanov RHA a RLA (N = 30 pre každú líniu s hmotnosťou ≈300 g na začiatku experimentálnej práce) boli všetky z kolónie založenej v 1998 na univerzite v Cagliari v Taliansku (Giorgi a kol., 2007). Postupy použité na selektívne šľachtenie sardínskych kolónií už boli podrobne opísané (Giorgi a kol., 2005).
Ovariektomizované stimuly samice potkanov SD (250 – 300 g na začiatku experimentálnej práce) použité vo všetkých experimentoch boli získané od spoločnosti Envigo (San Pietro al Natisone, Taliansko). Zvieratá boli aklimatizované štyri na klietku (38 cm x 60 cm x 20 cm) do ustajňovacích zariadení Katedry biomedicínskych vied univerzity Cagliari minimálne 10 dní pred začiatkom experimentov, pri teplote 24 ° C, vlhkosti. 60%, reverzný cyklus 12 h svetlo / tma (svetlá vypnuté z 08: 00 h do 20: 00 h), voda a štandardné laboratórne jedlo podľa chuti, Zvieratá boli denne liečené 1-2 min po celú dobu habituácie, aby sa počas pokusov obmedzil manipulačný stres; okrem toho bol kontakt s personálom údržby zvierat obmedzený na jediného sprievodcu a posteľná bielizeň v domácich klietkach sa nikdy nezmenila ani deň pred, ani v deň experimentov. Všetky experimenty sa uskutočňovali medzi 10: 00 h a 18: 00 h. Táto štúdia sa uskutočnila v súlade s odporúčaniami usmernení Európskych spoločenstiev, smernice zo septembra 22, 2010 (2010 / 63 / EU) a talianskych právnych predpisov (DL March 4, 2014, n. 26). Protokol bol schválený Etickou komisiou pre experimenty na zvieratách univerzity Cagliari (autorizačné č. 361 / 2016-PR, apríl 08, 2016 k FS).
Experimentálne skupiny
Boli použité sexuálne naivné a sexuálne skúsené samce potkanov RHA a RLA. Sexuálne naivné potkany boli potkany, ktoré neboli nikdy vystavené pohlavne vnímavej ovariektomizácii a samici primovanej estradiolom + progesterónom; potkany, ktoré majú pohlavný styk, boli potkany, ktoré už podstúpili päť po sebe nasledujúcich kopulačných testov 60 min v intervaloch 3 dní s vnímavou ženou (Sanna a kol., 2014a,b). Samice sa priviedli do estru ošetrením subkutánnym estradiol benzoátom (200 μg / potkan v arašidovom oleji) a progesterónom (0.5 mg / potkan v arašidovom oleji), 48 ha 6 h pred kopulačnými testami. Oestrus sa pred experimentmi overil sfarbením May-Grunwald-Giemsa a mikroskopickým vyšetrením vaginálnych náterov 1 h. V súlade s predchádzajúcimi štúdiami (Sanna a kol., 2014a,b, 2015) sa zistilo, že päť predbežných kopulačných testov postačuje na to, aby samce rímskych potkanov obidvoch línií vykazovali konštantné úrovne kopulačnej aktivity: napr. potkany RHA a RLA so sexuálnou skúsenosťou spĺňajú kritérium najmenej jednej ejakulácie dosiahnuté v každom z posledných dvoch testov (jedna krysa RHA a dve krysy RLA, ktoré nespĺňajú toto kritérium, boli v tejto fáze vyradené). Dva dni po týchto predbežných kopulačných testoch podstúpili rímske potkany, ktoré boli sexuálne skúsení, stereotaxický chirurgický zákrok na implantáciu mikrodialyzačnej sondy do mPFC, ako je opísané nižšie (Sanna a kol., 2015).
Mikrodialýza v mPFC počas sexuálneho správania
Deň pred mikrodialýzou boli sexuálne naivné alebo skúsené RHA a RLA potkany umiestnené do stereotaxického aparátu (Stoelting Co., Wood Dale, IL, USA) pod anestéziou izofluránom (1.5% - 2%; Harvard Apparatus, Holliston, MA, USA). a implantované vertikálnou domácou mikrodialyzačnou sondou (dialyzačná membrána ≈3 mm voľného povrchu; Melis a kol., 2003) a smerované jednostranne do kompartmentov mPFC, PrL a IL (súradnice: 3.0 mm predná a 0.7 mm laterálne k bregma a 5.5 mm ventrálne k dura; Paxinos a Watson, 2004). V deň experimentu boli zvieratá počas temnej fázy cyklu premiestnené do párovej klietky (45 cm x 30 cm x 24 cm), ktorá bola umiestnená v miestnosti zabezpečenej proti hluku, ktorá bola osvetlená slabým červeným svetlom a obsahovala vnútri iného malá klietka z plexiskla (15 cm × 15 cm × 15 cm) s otvormi 25 (Ø 2 mm) vo zvislých stenách, ktoré umožňujú vizuálnu, čuchovú a akustickú, ale nie priamu interakciu. Po habituačnej perióde 2 h sa mikrodialyzačná sonda pripojila k mikroinfúznej pumpe CMA / 100 (Harvard Apparatus, Holliston, MA, USA) s polyetylénovou hadičkou a perfundovala sa Ringerovým roztokom (147 mM NaCl, 3 mM KCl a 1.2 mM). CaCl2, pH 6.5), pri prietoku 2.5 μl / min. Po ekvilibračnej perióde 2 h perfúzneho média s extracelulárnou tekutinou sa odoberali alikvóty dialyzátu 37.5 μL každých 15 min. Počas experimentu v ľadom chladených polyetylénových skúmavkách na meranie koncentrácie dopamínu, DOPAC a NA, ako je opísané. nižšie. Po odobratí najmenej štyroch alikvót dialyzátu sa do malej klietky umiestnenej vo vnútri párovacej klietky na 30 min zaviedla receptívna samica potkana. Počas týchto 30 min sa odobrali ďalšie dva dialyzátové alikvoty. V týchto podmienkach samce potkanov nemôžu interagovať priamo so samičkou, ale vykazujú bezkontaktnú erekciu (pozri nižšie). Po tomto období bola malá klietka odstránená, kopulácia bola ponechaná 75 min a boli odobraté alikvóty ďalších päť dialyzátov. Na konci tohto obdobia sa samica vybrala z párenia a odobrala sa ďalšia alikvotná časť dialyzátu (Pfaus a Everitt, 1995; Melis a kol., 2003; Sanna a kol., 2015). Počas experimentu boli zaznamenané sexuálne parametre týkajúce sa predvídacej a konzumnej fázy sexuálneho správania (pozri nižšie).
Sexuálne správanie
V priebehu experimentu zaznamenal pozorovateľ niekoľko parametrov sexuálnej motivácie a kopulačného výkonu, ktoré sa týkali predvídacej a konzumnej fázy sexuálneho správania, a nevedel o konkrétnych experimentálnych podmienkach, napríklad nepoznal líniu a úroveň sexuálnej skúsenosti. zvierat použitých v tomto experimente (pozri nižšie). Stručne povedané, latencia k prvej bezkontaktnej erekcii (NCPEL, časovaná od zavedenia vnímavej samice do vnútornej malej klietky) a ich frekvencia (NCPEF, počet erekcií penisu bez dotyku, ktoré sa vyskytujú v období, v ktorom je samica) prítomné vo vnútornej klietke). Tieto feromónom sprostredkované erekcie penisu, ktoré sa vyskytujú u sexuálne silných samcov potkanov v prítomnosti neprístupnej vnímavej ženy, sa považujú za dôležitý index sexuálneho vzrušenia (Sachs a kol., 1994; Sachs, 2000; Melis a kol., 2003). Keď bola povolená sexuálna interakcia, napr. Počas kopulácie, latencia k vrcholu a intromit (ML a IL, časovanie od odstránenia vnútornej malej klietky až po prvý vrchol alebo prvý intromisia); frekvencia montáže a intromittingu (MF a IF, počet pripojení a intromisií v prvej sérii kopulačnej aktivity a počas celého obdobia kopulácie); latencia k ejakulácii (EL, časovaná od prvého intromisie prvej série do ejakulácie); bola zaznamenaná frekvencia ejakulácie (EF, celkový počet ejakulácií počas kopulačného testu) a po ejakulačnom intervale (PEI, časovaný od prvej ejakulácie do nasledujúcej intromisie). Ďalej, kopulačná účinnosť (CE; počet intromisií danej série vydelený súčtom počtu pripojení a intromisií v tej istej sérii) a interval medzi intromisiami (III; pomer medzi latenciou ejakulácie danej série). a počet intromisií v tejto sérii) sa tiež vypočítali pre prvú sériu kopulačnej aktivity (Sachs a Barfield, 1976; Meisel a Sachs, 1994; Melis a kol., 2003; Sanna a kol., 2014a,b, 2015).
Stanovenie koncentrácie dopamínu, DOPAC a noradrenalínu v dialyzáte z mPFC
Koncentrácie dopamínu, DOPAC a NA sa merali v 20 μl toho istého dialyzátového alikvotu z mPFC pomocou vysokotlakovej kvapalinovej chromatografie (HPLC) spojenej s elektrochemickou detekciou s použitím duálnej bunky 4011 (Coulochem II, ESA, Cambridge, MA, USA) ako už popísané (Melis a kol., 2003). Detekcia sa uskutočnila v redukčnom režime pri + 350 a -180 mV. HPLC bola vybavená kolónou Supelcosil C18 (7.5 cm x 3.0 mm id, 3 μm veľkosť častíc; Supelco, Supelchem, Miláno, Taliansko), eluovaná 0.06M citrát / acetát pH 4.2, obsahujúci metanol 20% v / v, 0.1 mM EDTA, 1 uM trietylamín a 0.03 mM dodecylsulfát sodný ako mobilná fáza, s prietokom 0.6 ml / min a laboratórnou teplotou. Citlivosť testu bola 0.125 pg pre dopamín, 0.1 pg pre DOPAC a 0.2 pg pre NA.
histológia
Na konci experimentov boli potkany usmrtené dekapitáciou, mozgy boli okamžite odstránené z lebky a ponorené do 4% vodného formaldehydu na 12-15 dni. Po uplynutí tohto obdobia boli pripravené 40 um koronálne rezy mozgu zmrazovacím mikrotómom, farbené neutrálnou červeňou a skontrolované mikroskopom s fázovým kontrastom. Poloha špičky sondy sa potom lokalizovala v mPFC sledovaním traktu sondy cez sériu rezov mozgu (pozri obrázok). 1). Na štatistické vyhodnotenie výsledkov sa zvážili iba potkany, u ktorých sa zistilo, že aktívna časť dialyzačnej membrány je správne umiestnená v priečinkoch PrL a IL mPFC (v tejto fáze bol vyradený jeden potkan z každej experimentálnej skupiny).
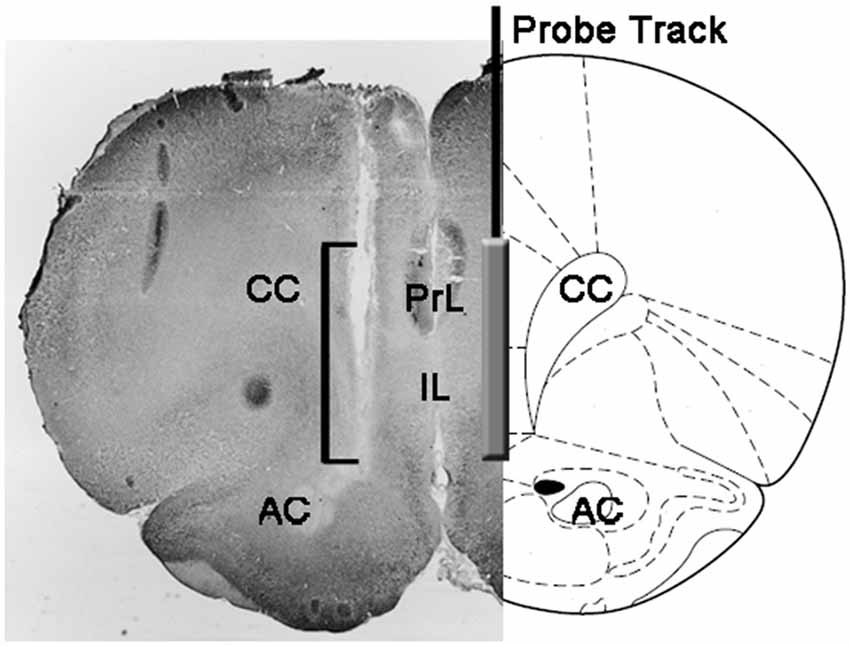 Obrázok 1, Schematické znázornenie koronálneho rezu mozgu potkana, ktorý ukazuje stopu mikrodialýznej sondy v PrL a IL častiach stredného prefrontálneho kortexu (mPFC; Paxinos a Watson, 2004). Štvorcová hranatá zátvorka na mikrofotografii ukazuje časť neutrálnej červenej farby, ktorá ukazuje aktívnu časť dialyzačnej membrány sondy na mikrodialýzu. Skratky: PrL, predimbická oblasť; IL, infralimbická oblasť; AC, predná komissura; CC, corpus callosum.
Obrázok 1, Schematické znázornenie koronálneho rezu mozgu potkana, ktorý ukazuje stopu mikrodialýznej sondy v PrL a IL častiach stredného prefrontálneho kortexu (mPFC; Paxinos a Watson, 2004). Štvorcová hranatá zátvorka na mikrofotografii ukazuje časť neutrálnej červenej farby, ktorá ukazuje aktívnu časť dialyzačnej membrány sondy na mikrodialýzu. Skratky: PrL, predimbická oblasť; IL, infralimbická oblasť; AC, predná komissura; CC, corpus callosum.
štatistika
Štatistické analýzy biochemických (dopamínových, DOPAC a NA) a behaviorálnych (NCPEL a NCPEF, ML, IL a EL, MF, IF a EF a PEI) údajov sa vykonali buď vrátane všetkých experimentálnych subjektov (ako sa tradične robí v štúdiách na potkanoch samcov). kopulačné správanie) alebo vylúčenie tých jedincov, ktorí sa počas mikrodialýzneho experimentu neaplikovali na ejakuláciu. V prvom prípade, keď boli do analýzy zahrnuté všetky zvieratá, zvieratám, ktoré nevykazovali NCPE v čase, keď bola samica neprístupná alebo sa nezainteresovala alebo neprichytila alebo ejakulovala k dispozícii samici, sa pridelilo príslušné skóre v celom rozsahu: 1800 s, ak samec nevykazoval NCPE v 30 min, v ktorom bola samica neprístupná; 900 pre ML a IL, ak sa samec nenamontoval alebo neprichytil k dispozícii samici; 1800 pre EL, ak samec nedosiahol ejakuláciu a 600 pre PEI, ak samec po prvej ejakulácii intromitoval. V druhom prípade boli tie zvieratá, ktoré sa k ejakulácii nekopírovali, vylúčené z analýz. Toto sa uskutočnilo s cieľom posúdiť, či rozdiely v sexuálnom správaní môžu korelovať s rozdielmi v koncentráciách extracelulárneho dopamínu, DOPAC a NA v mPFC línií rímskych potkanov počas dvoch fáz sexuálnej aktivity, čím sa vylúčia možné mätúce interferencie v dôsledku začlenenia. subjektov, ktoré nevykazovali sexuálne správanie tým, že im priradili fixné hodnoty pre analyzované sexuálne parametre.
Vykonalo sa niekoľko štatistických analýz s ANOVA, aby sa zistili a lepšie charakterizovali možné rozdiely medzi použitými experimentálnymi podmienkami. Pred vykonaním ANOVA sa v jednotlivých prípadoch skúmali údaje o súboroch každej z rôznych experimentálnych premenných z hľadiska homogenity variantov medzi štyrmi experimentálnymi skupinami (uskutočnené so všetkými zvieratami alebo potkanmi kopulujúcimi sa iba k ejakulácii) pomocou Bartlettovho alebo Levinovho testu. Keď sa zistili významné rozdiely v odchýlkach, súbory údajov sa logaritmus transformovali (tj experimentálne hodnoty Y sa zmenili na hodnoty Log Y), opätovne sa skontrolovala homogenita variantov a potom sa analyzovali pomocou ANOVA. Stručne povedané, behaviorálne parametre získané pred (napr. S neprístupnou ženou) a počas prvej série kopulačnej aktivity (od prvého kopca / intromisie po prvý kopec / intromisiu po prvej ejakulácii) naivných a sexuálne skúsených samcov potkanov RHA a RLA. počas mikrodialýzy sa analyzovali dvojsmernými ANOVA, použitím potkanej línie a úrovne sexuálnej skúsenosti medzi faktormi subjektu (priemerné hodnoty parametrov správania sú uvedené v tabuľke 1 a F hodnoty a úrovne významnosti analýz ANOVA v tabuľke 1 2).
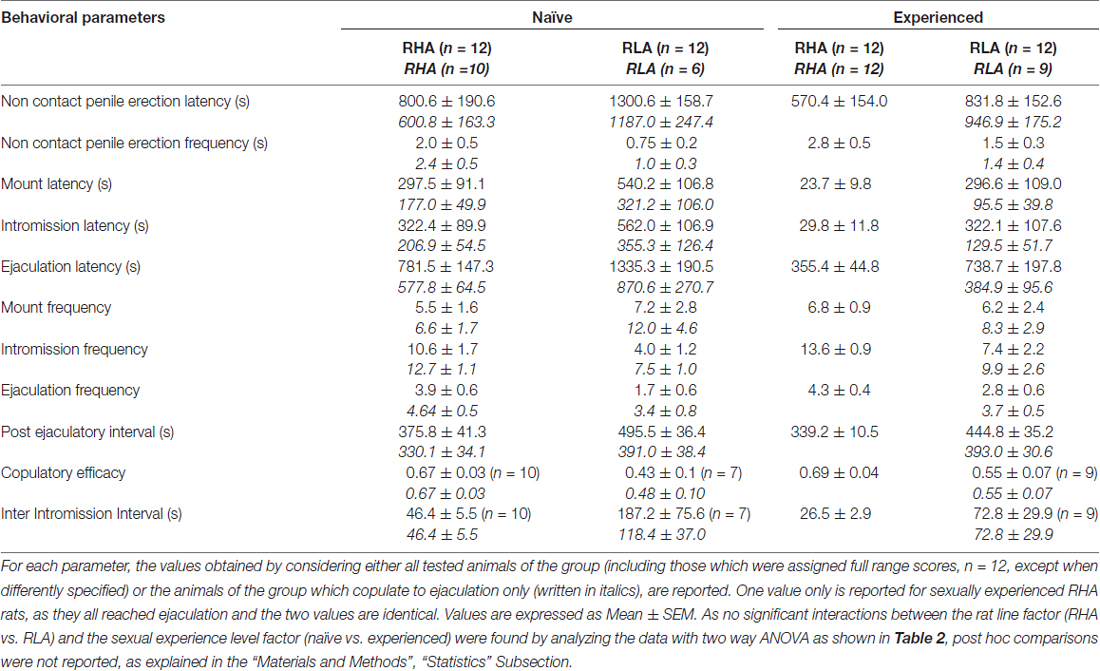 Tabuľka 1, Kopulačné parametre merané v prvej sérii kopulačnej aktivity (ktorá sa začína prvou horou / intromisiou, keď je samica sprístupnená samcovi, a končí po intervale po ejakulácii s prvou horou / intromisiou druhej série) a bezkontaktným penisom erekcie sexuálne naivných a skúsených RHA a RLA potkanov.
Tabuľka 1, Kopulačné parametre merané v prvej sérii kopulačnej aktivity (ktorá sa začína prvou horou / intromisiou, keď je samica sprístupnená samcovi, a končí po intervale po ejakulácii s prvou horou / intromisiou druhej série) a bezkontaktným penisom erekcie sexuálne naivných a skúsených RHA a RLA potkanov.
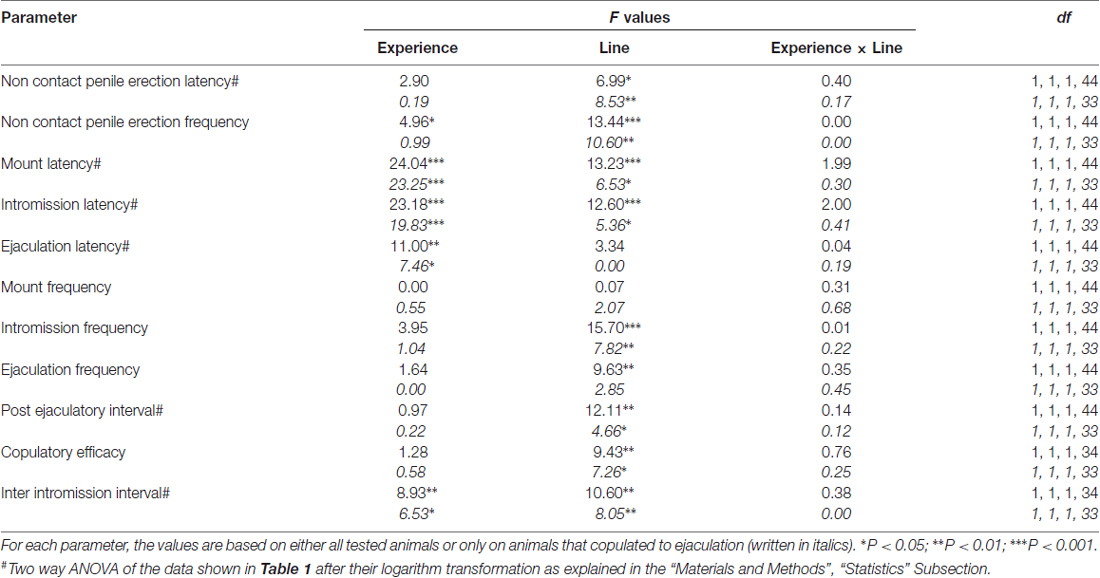 Tabuľka 2. F hodnoty a úrovne významnosti obojsmernej ANOVA vykonané na údajoch uvedených v tabuľke č 1 použitím línie potkanov (RHA verzus RLA) a úrovne sexuálnej skúsenosti (naivný vs. skúsený) medzi faktormi subjektu.
Tabuľka 2. F hodnoty a úrovne významnosti obojsmernej ANOVA vykonané na údajoch uvedených v tabuľke č 1 použitím línie potkanov (RHA verzus RLA) a úrovne sexuálnej skúsenosti (naivný vs. skúsený) medzi faktormi subjektu.
Okrem toho sa uskutočnila celková analýza údajov získaných od každého potkana počas mikrodialýzy vypočítaním najskôr AUC získaných vynesením hodnôt koncentrácií dopamínu, DOPAC alebo NA alebo počtu bezkontaktných erekcií, pripojení, intromisií a ejakulácií vs - čas (začínajúci 1 h po zavedení samca potkana do párovej klietky do konca experimentu - 180 min rozdelený do frakcií 15 min pre neurochemické hodnoty - alebo v čase, keď bola samica zavedená do párovej klietky) pre parametre správania) a potom porovnaním vypočítaných hodnôt pomocou dvojitých ANOVA s použitím potkanej línie a úrovne sexuálnej skúsenosti medzi faktormi subjektu (priemerné hodnoty AUC neurochemických a behaviorálnych parametrov sú uvedené v tabuľke). 4 a F hodnoty a úrovne významnosti analýz ANOVA v tabuľke 1 5). Nakoniec bola pomocou faktoriálov vykonaná podrobnejšia analýza bod za bodom každej sady údajov (tj hodnoty koncentrácií dopamínu, DOPAC, NA a počet nekontaktných erekcií, pripojení, intromisií a ejakulácií v závislosti od času). ANOVA pre opakované merania pomocou línie potkanov a úrovne sexuálnej skúsenosti medzi faktormi subjektu a časom (tj dialyzátové frakcie) ako v rámci faktora subjektu ( F hodnoty a úrovne významnosti týchto analýz ANOVA sú uvedené v tabuľke č 6). Podobné analýzy sa vykonali aj pri zohľadnení iba základných hodnôt neurochemických parametrov (posledné štyri dialyzátové alikvoty zozbierané pred zavedením do párovej klietky receptívnej ženy), ktorých priemerné hodnoty sú uvedené v tabuľke 3, Pri vykonávaní všeobecných faktoriálnych ANOVA pri opakovaných meraniach sa zistili interakcie prvého rádu, ale nie interakcie druhého rádu, najmä neurčemické parametre, najmä línia × čas a skúsenosť × časové interakcie (pozri tabuľku). 6) sa na týchto súboroch údajov vykonali priame ANOVA s líniou alebo úrovňou sexuálnej skúsenosti medzi faktorom subjektu a časom ako v rámci faktora subjektu priamym porovnaním sexuálne naivných alebo skúsených RHA verzus potkanov RLA alebo sexuálne skúsených vs. naivné RHA potkany alebo sexuálne skúsenosti oproti naivným RLA potkanom. Výsledky post hoc párové múdre kontrasty uskutočňované pomocou Tukeyovho HSD testu na významné interakcie odhalené týmito obojsmernými ANOVA sú uvedené na obrázku 2, Vo všetkých ostatných prípadoch post hoc porovnania neboli uvedené, pretože ANOVA neodhalili žiadnu významnú interakciu medzi líniou potkanov, úrovňou sexuálnej skúsenosti a časom. Všetky štatistické analýzy sa uskutočňovali s Graph Pad 5 (PRISM, San Diego, CA, USA) a STATISTICA 12 (Statsoft, Tulsa, OK, USA) s hladinou významnosti stanovenou na P <0.05.
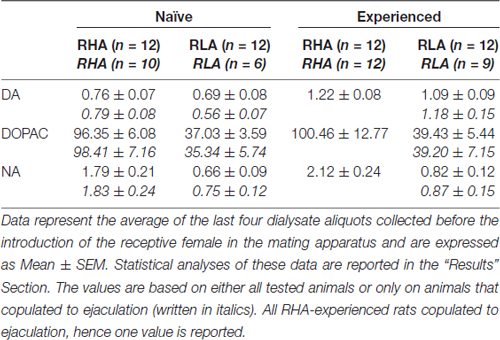 Tabuľka 3, Koncentrácie bazálneho dopamínu (DA), kyseliny 3,4-dihydroxyfenyloctovej (DOPAC) a noradrenalínu (NA) (nM) v dialyzáte zo stredných prefrontálnych kortexov (mPFC) sexuálne naivných a skúsených potkanov RHA a RLA.
Tabuľka 3, Koncentrácie bazálneho dopamínu (DA), kyseliny 3,4-dihydroxyfenyloctovej (DOPAC) a noradrenalínu (NA) (nM) v dialyzáte zo stredných prefrontálnych kortexov (mPFC) sexuálne naivných a skúsených potkanov RHA a RLA.
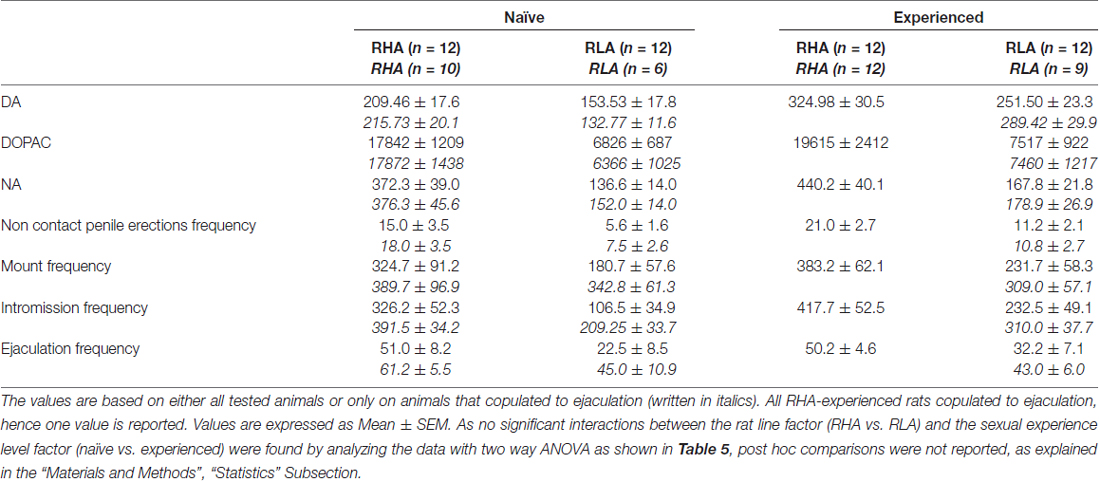 Tabuľka 4, Celkové hodnotenie rozdielov v koncentráciách DA, DOPAC a NA a v sexuálnom správaní medzi sexuálne naivnými a skúsenými RHA a RLA potkanmi analýzou spriemerovaných AUC získaných z výsledkov uvedených na obrázkoch 2 a 3.
Tabuľka 4, Celkové hodnotenie rozdielov v koncentráciách DA, DOPAC a NA a v sexuálnom správaní medzi sexuálne naivnými a skúsenými RHA a RLA potkanmi analýzou spriemerovaných AUC získaných z výsledkov uvedených na obrázkoch 2 a 3.
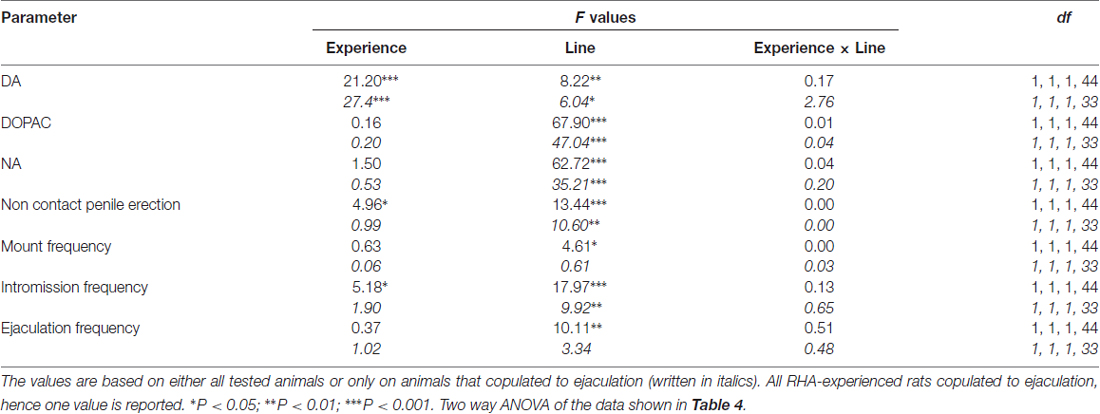 Tabuľka 5. F hodnoty a úrovne významnosti obojsmerných ANOVA vykonané na údajoch uvedených v tabuľke č 4 použitím línie potkanov (RHA verzus RLA) a úrovne sexuálnej skúsenosti (naivný vs. skúsený) medzi faktormi subjektu.
Tabuľka 5. F hodnoty a úrovne významnosti obojsmerných ANOVA vykonané na údajoch uvedených v tabuľke č 4 použitím línie potkanov (RHA verzus RLA) a úrovne sexuálnej skúsenosti (naivný vs. skúsený) medzi faktormi subjektu.
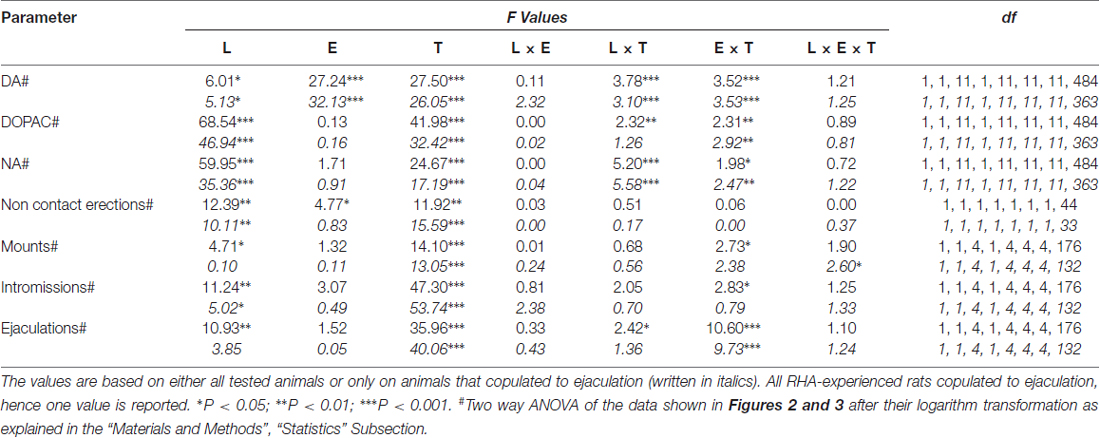 Tabuľka 6. F hodnoty a úrovne významnosti všeobecných faktorových ANOVA pre opakované merania vykonané na výsledkoch znázornených na obrázkoch č 2 a 3 použitím línie potkanov (L; RHA vs. RLA) a úrovne sexuálnej skúsenosti (E) (naivné vs. zažívané) medzi faktormi subjektu a časom (T) (frakcie dialyzátu) ako v rámci faktora subjektu.
Tabuľka 6. F hodnoty a úrovne významnosti všeobecných faktorových ANOVA pre opakované merania vykonané na výsledkoch znázornených na obrázkoch č 2 a 3 použitím línie potkanov (L; RHA vs. RLA) a úrovne sexuálnej skúsenosti (E) (naivné vs. zažívané) medzi faktormi subjektu a časom (T) (frakcie dialyzátu) ako v rámci faktora subjektu.
 Obrázok 2, Koncentrácie dopamínu (DA), DOPAC a noradrenalínu (NA) v dialyzátoch mPFC zo sexuálne naivných (A, C, E) a skúsených samcov potkanov RHA a RLA (B, D, F) počas sexuálnej aktivity s vnímavou ženou. Sexuálne naivné (tj nikdy predtým nevystavené sexuálne vnímavej žene) a sexuálne skúsené (ktoré podstúpili päť kopulačných testov) oboch línií s mikrodialyzačnou sondou stereotaxicky implantovanou do mPFC sa umiestnili jednotlivo do párovej klietky. Mikrodialyzačná sonda sa premývala dialyzačným pufrom, ako je opísané v časti „Materiály a metódy“. Po odobratí štyroch dialyzátových alikvotov na stanovenie bazálnych hodnôt bola do malej klietky párovacieho prístroja umiestnená neprístupná recepčná samica (čas = 0, neprístupný). Po 30 min. Bola malá klietka odstránená a kopulácia bola ponechaná 75 min (kopulácia), potom bola samica odstránená z párenia klietky (fe.remov). Počas experimentu sa merali bezkontaktné erekcie a kopulačné parametre a dialyzátové alikvoty sa zbierali každých 15 min a analyzovali sa na dopamín, DOPAC a NA, ako je opísané v časti „Materiály a metódy“. Hodnoty sú stredné hodnoty ± SEM hodnôt získaných u všetkých potkanov 12 na skupinu (RHA_all = plné zelené čiary, RLA_all = plné hnedé čiary) alebo tých, ktoré sa kopulujú iba na ejakuláciu (RHA_cop = prerušované červené čiary; RLA_cop = prerušované modré čiary). U skúsených potkanov RHA dosiahli všetky zvieratá kopulácie a hodnoty sú rovnaké. *P <0.05 vzhľadom na bazálne hodnoty (bez samice) skupiny (zelená pre RHA_all; červená pre RHA_cop, hnedá pre RLA_all, modrá pre RLA_cop); #P <0.05 vzhľadom na zodpovedajúce hodnoty skupiny RLA (červená, RHA_all vs. RLA_all; zelená, RHA_cop vs. RLA_cop); §P <0.05 vzhľadom na časovo porovnané hodnoty sexuálne naivných potkanov (zelená, skúsená RHA_all vs. naivná RHA_all; červená, skúsená RHA_cop vs. naivná RHA_cop; hnedá, skúsená RLA_all vs. naivná RLA_all; modrá, skúsená RLA_cop vs. naivná RLA_cop) (obojsmerné ANOVA vykonané na zobrazených dátach po ich logaritmickej transformácii, ako je vysvetlené v pododdieloch „Materiály a metódy“, „Štatistika“, po ktorých nasledujú testy HSD spoločnosti Tukey).
Obrázok 2, Koncentrácie dopamínu (DA), DOPAC a noradrenalínu (NA) v dialyzátoch mPFC zo sexuálne naivných (A, C, E) a skúsených samcov potkanov RHA a RLA (B, D, F) počas sexuálnej aktivity s vnímavou ženou. Sexuálne naivné (tj nikdy predtým nevystavené sexuálne vnímavej žene) a sexuálne skúsené (ktoré podstúpili päť kopulačných testov) oboch línií s mikrodialyzačnou sondou stereotaxicky implantovanou do mPFC sa umiestnili jednotlivo do párovej klietky. Mikrodialyzačná sonda sa premývala dialyzačným pufrom, ako je opísané v časti „Materiály a metódy“. Po odobratí štyroch dialyzátových alikvotov na stanovenie bazálnych hodnôt bola do malej klietky párovacieho prístroja umiestnená neprístupná recepčná samica (čas = 0, neprístupný). Po 30 min. Bola malá klietka odstránená a kopulácia bola ponechaná 75 min (kopulácia), potom bola samica odstránená z párenia klietky (fe.remov). Počas experimentu sa merali bezkontaktné erekcie a kopulačné parametre a dialyzátové alikvoty sa zbierali každých 15 min a analyzovali sa na dopamín, DOPAC a NA, ako je opísané v časti „Materiály a metódy“. Hodnoty sú stredné hodnoty ± SEM hodnôt získaných u všetkých potkanov 12 na skupinu (RHA_all = plné zelené čiary, RLA_all = plné hnedé čiary) alebo tých, ktoré sa kopulujú iba na ejakuláciu (RHA_cop = prerušované červené čiary; RLA_cop = prerušované modré čiary). U skúsených potkanov RHA dosiahli všetky zvieratá kopulácie a hodnoty sú rovnaké. *P <0.05 vzhľadom na bazálne hodnoty (bez samice) skupiny (zelená pre RHA_all; červená pre RHA_cop, hnedá pre RLA_all, modrá pre RLA_cop); #P <0.05 vzhľadom na zodpovedajúce hodnoty skupiny RLA (červená, RHA_all vs. RLA_all; zelená, RHA_cop vs. RLA_cop); §P <0.05 vzhľadom na časovo porovnané hodnoty sexuálne naivných potkanov (zelená, skúsená RHA_all vs. naivná RHA_all; červená, skúsená RHA_cop vs. naivná RHA_cop; hnedá, skúsená RLA_all vs. naivná RLA_all; modrá, skúsená RLA_cop vs. naivná RLA_cop) (obojsmerné ANOVA vykonané na zobrazených dátach po ich logaritmickej transformácii, ako je vysvetlené v pododdieloch „Materiály a metódy“, „Štatistika“, po ktorých nasledujú testy HSD spoločnosti Tukey).
výsledky
Krysy RHA a RLA vykazujú rôzny počet kontaktných erekcií a rôzne vzorce kopulačného správania
V súlade s predchádzajúcimi štúdiami (Sanna a kol., 2014a,b, 2015), rôzny počet sexuálne naivných samcov potkanov RHA a RLA sa zapojil do sexuálnej aktivity, keď sa spojil s receptívnou samicou počas odberu alikvót dialyzátu z mPFC intracerebrálnou mikrodialýzou. Stručne, v tejto štúdii sa 10 z 12 sexuálne naivných samcov RHA (83%) kopulovalo k ejakulácii v ich prvom kopulačnom teste, iba proti 6 z 12 naivných RLA potkanov (50%) počas experimentu s mikrodialýzou. Tento rozdiel sa tiež zistil u sexuálne skúsených samcov potkanov RHA a RLA (po piatich kopulačných testoch), hoci boli oslabené, pričom všetky potkany 12 RHA dosiahli ejakuláciu proti 9 z potkanov 12 RLA počas mikrodialýzy. Vždy v zhode s predchádzajúcimi štúdiami, aj v tejto štúdii, dve línie rímskych potkanov vykazovali rôzne latencie a frekvencie nekontaktných erekcií penisu, keď boli vystavené neprístupnej vnímavej žene, ako aj rôzne vzorce kopulačného správania počas sexuálnej interakcie. Tieto rozdiely sa našli buď pri posudzovaní údajov všetkých experimentálnych zvierat danej skupiny, bez ohľadu na to, či sa kopulovali k ejakulácii alebo nie (napr. Pridelením úplného skóre tým zvieratám, ktoré nevykazovali správanie, tj potkanov, ktoré sa kopulovali do ejakulácia) alebo pri posudzovaní údajov o zvieratách, ktoré ukazujú správanie danej skupiny (napr. zvieratá, ktoré sa pripájali k ejakulácii; tabuľka) 1). Štatistická analýza hodnôt bezkontaktných erekcií (NCPE) a kopulačných parametrov meraných v prvej sérii kopulačnej aktivity pomocou dvojsmerných ANOVA odhalila významné rozdiely medzi dvoma líniami potkanov, buď pri posudzovaní všetkých experimentálnych potkanov, alebo iba pri kopulovaní potkanoch. v štyroch experimentálnych skupinách (pozri tabuľku č 2). V skutočnosti, keď sa brali do úvahy údaje získané zo všetkých pokusných zvierat, bol počet bezkontaktných erekcií vyšší a NCPEL, ML, IL a PEI boli významne nižšie u RHA krýs v porovnaní s RLA potkanmi. Ďalej, EF a CE boli vyššie, zatiaľ čo III boli významne kratšie, u RHA potkanov v porovnaní s RLA potkanmi. Niektoré z vyššie uvedených rozdielov medzi dvoma líniami rímskych potkanov mali tendenciu zmenšovať sa alebo vymiznúť pri opakovaných kopulačných testoch. Niektoré z týchto rozdielov však stále pretrvávali aj po stabilizácii sexuálneho správania opakovaným sexuálnym zážitkom. Podobné výsledky sa získali, keď sa zvažovali iba hodnoty zvierat zo štyroch experimentálnych skupín, ktoré sa kopulovali k ejakulácii, s výnimkou EF (pozri tabuľky). 1, 2).
Bazálne koncentrácie extracelulárneho dopamínu, DOPAC a noradrenalínu v mPFC dialyzátoch od sexuálne naivných a skúsených RHA a RLA potkanov
Za súčasných experimentálnych podmienok boli množstvá dopamínu, DOPAC a NA v dialyzátoch získaných z mPFC všetkých (napr. Bez ohľadu na to, či sa kopulovali do ejakulácie alebo nie) sexuálne naivných RHA a RLA potkanov 2.32 pg a 2.11 pg pre dopamín, 322.56 pg a 124.42 pg pre DOPAC a 6.05 pg a 2.24 pg pre NA v uvedenom poradí, v 20 μl dialyzátu. Podobné množstvá sa merali v dialyzátoch získaných z mPFC u sexuálne skúsených potkanov RHA a RLA (dopamín: 3.73 pg a 3.33 pg, DOPAC: 353.62 pg a 133.32 pg, NA: 7.17 pg a 2.78 pg, v danom poradí). , Tieto hodnoty označujú koncentráciu ≅0.8 – 1.2 nM a ≅95 – 100 nM pre extracelulárny dopamín a DOPAC, respektíve ≅1.8 – 2.2 nM pre extracelulárnu NA, v mPFC RHA potkanov a koncentráciu ≅0.8 – 1.2 nM a ≅35 – 40 nM pre extracelulárny dopamín a DOPAC, respektíve ≅0.6 – 0.9 nM pre extracelulárnu NA, v mPFC potkanov RLA (tabuľka) 3). Vyššie uvedené hodnoty sa získali po ekvilibračnej perióde 2 h dialyzačného pufra s extracelulárnou tekutinou mPFC. Pretože sa odhadovalo, že výťažok autentického dopamínu, DOPAC a NA dialyzačných sond je blízko 20%, koncentrácie extracelulárneho dopamínu, DOPAC a NA sa môžu odhadnúť na blízku ≅4 – 5 nM v oboch líniách pre dopamín, ≅500. a ≅ 200 nM pre DOPAC a UM10 a ≅4 nM pre NA v mPFC potkanov RHA a RLA. Faktorové ANOVA pre opakované merania vyššie uvedených hodnôt po transformácii logaritmu odhalili významné rozdiely v základných hladinách dopamínu medzi sexuálne naivnými a sexuálne skúsenými potkanmi (F(1,44,132) = 26.05, P <0.001) a v bazálnych hladinách DOPAC (F(1,44,132) = 63.36, P <0.001) a NA (F(1,44,132) = 42.14, P <0.001) medzi potkanmi RHA a RLA (posledné štyri vzorky odobraté pred zavedením samice do malej klietky). Podobné hodnoty boli zistené pri uvažovaní iba u potkanov, ktoré kopulovali k ejakulácii počas experimentu s mikrodialýzou (tabuľka 3). Aj v tomto prípade faktorové ANOVA pre opakované merania hodnôt po transformácii logaritmu odhalili významné rozdiely v bazálnych hladinách dopamínu medzi sexuálne naivnými a sexuálne skúsenými potkanmi (F(1,33,99) = 26.97, P <0.001) a v bazálnych hladinách DOPAC (F(1,33,99) = 42.95, P <0.001) a NA (F(1,33,99) = 27.63, P <0.001) medzi RHA a RLA potkanmi.
Koncentrácie extracelulárneho dopamínu, DOPAC a noradrenalínu v mPFC dialyzátoch zo sexuálne naivných a skúsených RHA a RLA potkanov sa počas sexuálnej aktivity menia rozdielne.
Prítomnosť neprístupnej vnímavej ženy vo vnútornej malej klietke a následná priama sexuálna interakcia zvýšila koncentrácie extracelulárneho dopamínu, DOPAC a NA v dialyzátoch mPFC získaných od sexuálne naivných a skúsených samcov potkanov RHA a RLA, hoci s významnými rozdielmi medzi dve línie krýs a úroveň skúseností, buď pri zvažovaní všetkých pokusných zvierat, alebo iba zvierat, ktoré sa pripájajú k ejakulácii (obrázok) 2). V skutočnosti dvojsmerné ANOVA AUC koncentrácií dopamínu, DOPAC a NA ukázali, že celkový obsah dopamínu, DOPAC a NA z celého testu bol významne vyšší u RHA potkanov v porovnaní s RLA potkanmi a sexuálne. v porovnaní s naivnými potkanmi (pozri tabuľku 5) 4 pre hodnoty AUC a tabuľku 5 pre F hodnoty a hladina významnosti). Tieto rozdiely boli ďalej potvrdené bodovo-bodovými faktorovými ANOVA pre opakované merania množstva dopamínu, DOPAC a NA u dvoch sexuálne naivných a skúsených línií potkanov pozdĺž experimentu, ktoré odhalili významné hlavné účinky línií, skúseností, času a významných línií × času a Skúsenosti × Interakcie prvého poriadku v čase (pozri tabuľku 6 pre F hodnoty a hladina významnosti).
Koncentrácie extracelulárneho dopamínu, DOPAC a noradrenalínu v mPFC dialyzátoch získaných zo sexuálne naivných RHA a RLA potkanov sa počas sexuálnej aktivity odlišujú.
U sexuálne naivných potkanov RHA a RLA sa extracelulárny dopamín, ale nie DOPAC, zvýšil najskôr, keď sa dostali do neprístupnej samice, zatiaľ čo NA sa zvýšil iba u RHA potkanov. U naivných RHA potkanov sa extracelulárny dopamín ďalej zvýšil v prvých 15 min. Sexuálnej interakcie, ako sa zistilo pri DOPAC a NA. Prírastky v extracelulárnom dopamíne, DOPAC a NA trvali po celú dobu kopulácie, pričom dopamín, DOPAC a NA dosahovali najvyššiu hodnotu pri 60 min, 75 min a 75 min, potom extracelulárny dopamín a v menšej miere DOPAC a NA , klesla na hodnoty podobné bazálnym hodnotám po odstránení samice.
Na druhej strane k prvému významnému zvýšeniu a maximálnej hodnote extracelulárneho dopamínu, DOPAC a NA došlo u naivných RLA potkanov počas prvých 15 min. Kopulácie. Avšak na rozdiel od RHA krýs, u RLA potkanov extracelulárny dopamín, DOPAC a NA mali tendenciu sa vracať na hodnoty podobné bazálnym hodnotám počas prvých 30 min. Priamej interakcie so samičkou (pozri obrázky). 2, C, E).
Nakoniec počas kopulácie boli prírastky extracelulárneho dopamínu väčšie a perzistentnejšie u sexuálne naivných RHA ako u potkanov RLA (pozri obrázok). 2A), zatiaľ čo v neprítomnosti ženy boli ich koncentrácie podobné, aj keď nie identické, cez dve línie potkanov (pozri vyššie a tabuľku 1) 3). Je zaujímavé, že hoci sa nezistil žiadny rozdiel medzi dvoma rímskymi čiarami v percentuálnom náraste DOPAC a v menšej miere koncentráciách NA počas prítomnosti receptívnej ženy, rozdiel viac ako dvojnásobku absolútnych hodnôt koncentrácií DOPAC a NA medzi základnými potkanými líniami sa pozorovali obidve línie krýs (pozri tabuľku 1) 3) a počas testu s vnímavou ženou (pozri obrázky 1 a 2) 2C, E).
Koncentrácie extracelulárneho dopamínu, DOPAC a noradrenalínu v mPFC dialyzátoch získaných zo sexuálne skúsených potkanov RHA a RLA sa počas sexuálnej aktivity menia rozdielne.
Ako sa zistilo u sexuálne naivných potkanov RHA a RLA, prítomnosť neprístupných samíc a následná priama sexuálna interakcia viedli k zvýšeniu extracelulárneho dopamínu a DOPAC v dialyzáte mPFC získanom od pohlavne skúsených samcov potkanov RHA a RLA. Aj v tomto prípade sa však zistili rozdiely týkajúce sa línie potkanov, buď pri posudzovaní všetkých pokusných zvierat, alebo v menšej miere iba tých, ktoré sa pripájali k ejakulácii (obrázok). 2). U sexuálne skúsených potkanov RHA a RLA sa u neprístupnej samice vyskytlo prvé zvýšenie extracelulárneho dopamínu. Potom sa koncentrácie dopamínu počas kopulácie zvýšili v oboch líniách potkanov. Koncentrácie dopamínu dosiahli svoje maximálne hodnoty po 45 – 60 min. Kopulácie a na konci kopulačného testu pomaly klesali smerom k bazálnym hodnotám (pozri obrázok) 2B). Podobne ako u sexuálne naivných potkanov boli koncentrácie extracelulárneho dopamínu vyššie u sexuálne skúsených potkanov RHA v porovnaní s potkanmi RLA hlavne v alikvótoch dialyzátu získaných v strednej časti kopulačnej fázy (po 30 – 45 min. Kopulácie). Koncentrácie DOPAC sa počas prítomnosti neprístupnej samice zvyšovali iba u potkanov RHA, zatiaľ čo v rovnakom rozsahu sa zvyšovali v oboch líniách potkanov počas kopulácie (najvyššie hodnoty pri 75 – 90 min.) A mali tendenciu sa na konci základnej hodnoty vracať k bazálnym hodnotám. kopulačný test (pozri obrázok č 2D). Koncentrácie NA sa tiež zvýšili v oboch pohlavne skúsených potkaních líniách po zavedení samice dosahujúcej najvyššie hodnoty pri 90 min u RHA potkanov a 45 min u RLA potkanov (pozri obrázok) 2F). Pri odchýlke od dopamínu (pozri vyššie) sa zistil podobný rozdiel vo vzraste koncentrácií DOPAC a NA u sexuálne skúsených potkanov z oboch línií, keď sa zvažovali všetky zvieratá oproti zvieratám, ktoré dosiahli iba ejakuláciu. Ako sa však zistilo u sexuálne naivných krýs, hoci percentuálne zvýšenie koncentrácií DOPAC a NA bolo podobné, v priebehu testu (tj pred, počas a po ňom) sa zistili významné rozdiely v absolútnych hodnotách DOPAC a NA medzi sexuálne skúsenými potkanmi RHA a RLA. prítomnosť receptívnej samice), pričom RHA potkany vykazujú vyššie hodnoty ako RLA potkany (pozri tabuľku 1) 3 pre základné hodnoty a obrázky 2D, F pre hodnoty počas celého testu).
Sexuálne skúsenosti ovplyvňujú zmeny v koncentráciách dopamínu a noradrenalínu v mPFC dialyzátoch získaných z RHA a RLA potkanov počas sexuálnej aktivity
Porovnanie koncentrácií extracelulárneho dopamínu a NA u sexuálne naivných a skúsených potkanov RHA a RLA ukázalo, že sexuálna skúsenosť zmenila koncentráciu extracelulárneho dopamínu a v menšej miere NA v oboch líniách potkanov, a to buď pri posudzovaní všetkých pokusných zvierat. alebo tie, ktoré sa pripájali iba k ejakulácii (obrázky č 2, B, E, F). V súlade s tým sa vyskytli významné rozdiely v bazálnych hodnotách dopamínu medzi sexuálne naivnými a skúsenými potkanmi oboch línií (pozri tabuľku). 3) a hodnoty dopamínu počas kopulačnej aktivity boli všeobecne vyššie u sexuálne skúsených potkanov RHA a RLA v porovnaní s ich naivnými náprotivkami (pozri obrázok) 2B). Okrem toho u sexuálne skúsených potkanov RHA, hoci zvýšenie dopamínu zistené počas prítomnosti neprístupnej samice nasledovalo rovnaký časový profil, aký sa pozoroval u sexuálne naivných krýs, počas kopulácie sa zistili rozdiely v časovom charaktere. V tomto prípade sa na rozdiel od toho, čo sa pozorovalo u sexuálne naivných RHA krýs (jeden hlavný pík pri 60 min), našli dve hlavné hodnoty piku, prvý po 15 min a druhý po 45 min kopulácie. Po tomto poslednom zvýšení mali hodnoty dopamínu tendenciu sa vracať k bazálnym hodnotám (pozri obrázok č 2B).
Podobný obrázok bol nájdený u sexuálne naivných vs. skúsených RLA potkanov. V skutočnosti, podobne ako v prípade RHA potkanov, sa tiež u skúsených RLA potkanov zistil časový obraz charakterizovaný tromi hlavnými zvýšeniami extracelulárneho dopamínu: prvé s neprístupnou samicou a ďalšie dve počas kopulácie, tj po 15 min a 60 min. kopulácie (pozri obrázky 2, B). Vo všeobecnosti sa zdá, že sexuálne skúsenosti spôsobujú dlhodobejšie zmeny v časovom priebehu uvoľňovania dopamínu v oboch líniách potkanov pri prechode zo sexuálne naivnej na skúsený stav.
Ako je znázornené na obr 2C-Fboli zistené iba malé rozdiely v koncentrácii DOPAC a NA medzi naivnými a skúsenými potkanmi oboch línií. Avšak v bazálnom podaní nebol pozorovaný významný trend extracelulárnej NA smerom k vyšším hodnotám v porovnaní s naivnými potkanmi (pred zavedením vnímavej samice, pozri tabuľku). 3) a v celkovom objeme NA, ako je uvedené pomocou hodnôt AUC (pozri tabuľku 1) 4) vypočítané na extracelulárne koncentrácie získané z celého testu (pred a počas prítomnosti vnímavej ženy). Nakoniec bol časový priebeh koncentrácií NA veľmi podobný pri porovnaní skúsených potkanov oboch línií potkanov s ich naivnými náprotivkami, čo naznačuje, že sexuálny zážitok môže viesť skôr k všeobecnému zvýšeniu extracelulárnej NA, ako k zmenám v štruktúre jeho zvýšenia (pozri obrázky). 2, F).
Zmeny v koncentráciách extracelulárneho dopamínu a noradrenalínu v mPFC dialyzátoch získaných zo sexuálnych naivných a skúsených RHA a RLA potkanov sa vyskytujú súčasne so zmenami v sexuálnom správaní
Rozdiely v koncentráciách extracelulárneho dopamínu, DOPAC a NA v dialyzátoch získaných z mPFC, ktoré sa našli v teste u sexuálne naivných a skúsených potkanov RHA a RLA (obrázok) 2) sa vyskytli spolu s modifikáciami rôznych sexuálnych parametrov meranými v dvoch hlavných fázach experimentov (tj samica neprístupná a dostupná pre mužov), medzi ktoré patria okrem iného bezkontaktné erekcie penisu (obrázky). 3, B), držiaky (obrázky 3C, D), intromisie (obrázky č 3, F) a ejakulácie (obrázky 3G, H). Tieto rozdiely sa zistili pri posudzovaní všetkých pokusných zvierat alebo zvierat, ktoré sa pripájajú k ejakulácii. V skutočnosti predbežná analýza hodnôt AUC týchto parametrov pomocou obojsmerných ANOVA (pozri tabuľku 1) 4 pre hodnoty AUC a tabuľku 5 pre F hodnoty a hladina významnosti) nasledovaná bodovou analýzou hodnôt týchto parametrov v experimente faktoriálnou ANOVA pri opakovaných meraniach odhalila významné hlavné účinky interakcie medzi linkami, skúsenosťami, časom, významnými prvkami prvého rádu × čas a skúsenosťami x časové interakcie, a významnú interakciu druhého rádu pre držiaky (pozri tabuľku 1) 6 pre F hodnoty a hladina významnosti). V súlade s predchádzajúcimi štúdiami (Sanna a kol., 2015): i) sexuálne naivné RHA potkany vykazovali viac bezkontaktných erekcií penisov, úchyty, intromisie a ejakulácie ako ich náprotivky RLA; ii) sexuálne skúsené potkany RHA vykazovali vyšší počet erekcií, intromisií a ejakulácií bez kontaktu, ako ich náprotivky RLA; a (iii) sexuálne skúsené potkany oboch línií, a najmä potkanov RLA, vykazovali viac bezkontaktných erekcií penisov, intromisií a ejakulácií ako ich sexuálne naivné náprotivky. V skutočnosti, zatiaľ čo u RHA potkanov sa väčšina zmien vyvolaných sexuálnymi skúsenosťami vyskytla skôr v časovom priebehu ako vo frekvencii behaviorálnych parametrov (napr. Skúsené potkany kopulovali hlavne v prvej polovici testu, zatiaľ čo naivné potkany vykázali podobný kopulačný trend v priebehu testu), u potkanov RLA došlo k výraznejšiemu všeobecnému zlepšeniu sexuálneho správania, najmä pri sexuálnom výkone, sexuálnymi skúsenosťami, čo bolo odhalené zmenami hodnôt kopulačných parametrov u skúsených potkanov RLA v porovnaní s ich potkanmi naivné náprotivky.
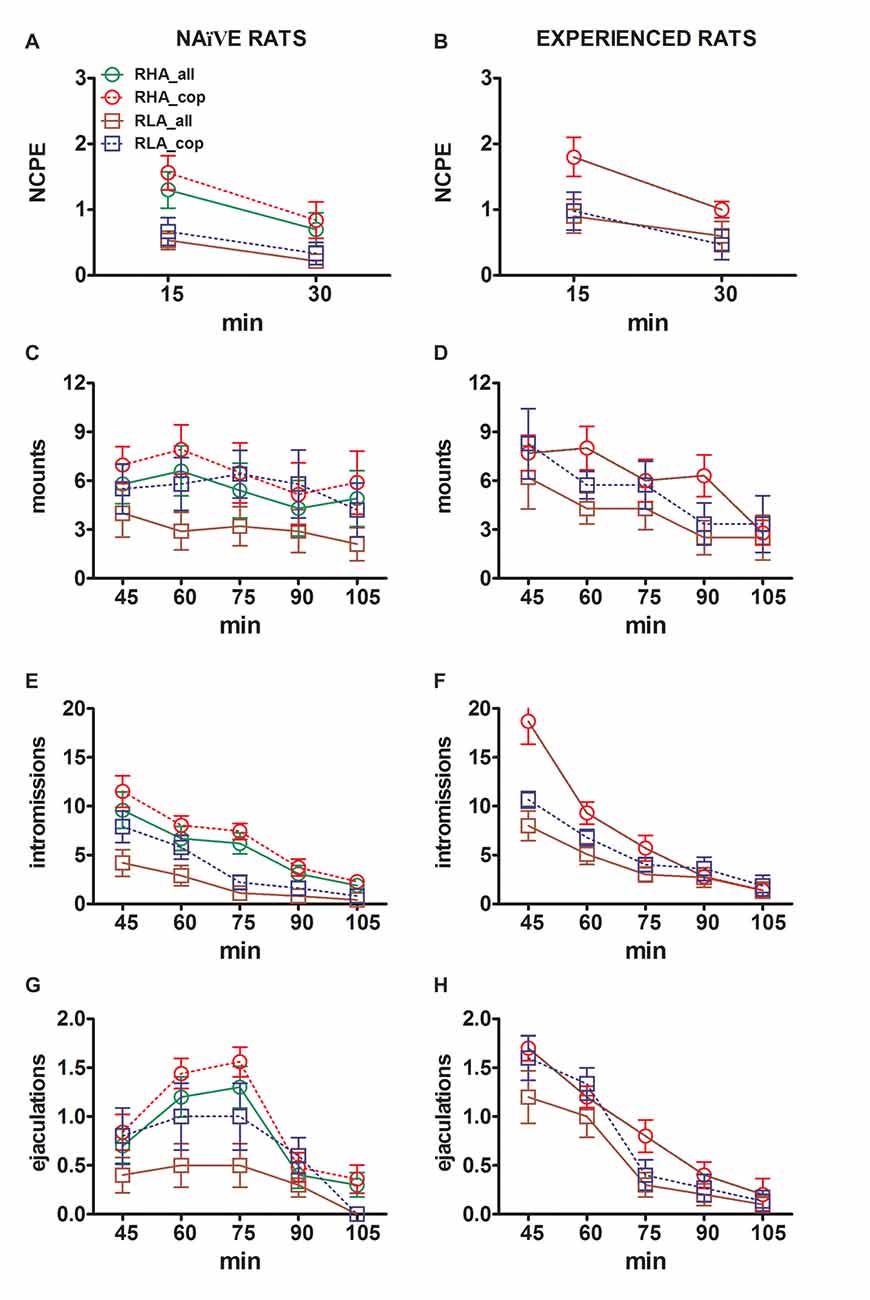 Obrázok 3, Zmeny počtu bezkontaktných erekcií penisu (NCPE), úchyty, intromisie a ejakulácie zaznamenané zo sexuálne naivných (A, C, E, G) a zažili RHA a RLA potkany (B, D, F, H) počas experimentov na mikrodialýze uvedených na obrázku č 2, Všetky experimentálne podmienky boli identické s podmienkami opísanými v legende na obr 2, Hodnoty sú stredné hodnoty ± SEM hodnôt získaných všetkými potkanmi 12 v skupine (vrátane tých, ktorým boli pridelené skóre na plný úväzok; RHA = plné zelené čiary, RLA = plné hnedé čiary) alebo tých, ktoré sa kopírovali iba k ejakulácii (RHA = prerušované červené čiary) ; RLA = prerušované modré čiary). U skúsených potkanov RHA dosiahli všetky zvieratá kopulácie a hodnoty sú rovnaké. Pretože pri analýze údajov s dvojsmernými ANOVA, ako je uvedené v tabuľke, sa nezistili žiadne významné interakcie medzi faktorom potkanej línie (RHA vs. RLA), faktor úrovne sexuálnej skúsenosti (naivný vs. skúsený) a čas. 6, post hoc porovnania neboli uvedené, ako je vysvetlené v pododdiele „Materiály a metódy“, „Štatistika“.
Obrázok 3, Zmeny počtu bezkontaktných erekcií penisu (NCPE), úchyty, intromisie a ejakulácie zaznamenané zo sexuálne naivných (A, C, E, G) a zažili RHA a RLA potkany (B, D, F, H) počas experimentov na mikrodialýze uvedených na obrázku č 2, Všetky experimentálne podmienky boli identické s podmienkami opísanými v legende na obr 2, Hodnoty sú stredné hodnoty ± SEM hodnôt získaných všetkými potkanmi 12 v skupine (vrátane tých, ktorým boli pridelené skóre na plný úväzok; RHA = plné zelené čiary, RLA = plné hnedé čiary) alebo tých, ktoré sa kopírovali iba k ejakulácii (RHA = prerušované červené čiary) ; RLA = prerušované modré čiary). U skúsených potkanov RHA dosiahli všetky zvieratá kopulácie a hodnoty sú rovnaké. Pretože pri analýze údajov s dvojsmernými ANOVA, ako je uvedené v tabuľke, sa nezistili žiadne významné interakcie medzi faktorom potkanej línie (RHA vs. RLA), faktor úrovne sexuálnej skúsenosti (naivný vs. skúsený) a čas. 6, post hoc porovnania neboli uvedené, ako je vysvetlené v pododdiele „Materiály a metódy“, „Štatistika“.
Diskusia
Táto štúdia potvrdzuje a rozširuje zistenia predchádzajúcich štúdií, ktoré ukazujú, že RHA a RLA potkany, ktoré vykazujú výrazne odlišné štýly zvládania reakcie v závislosti na averzívnych podmienkach (Driscoll a Bättig, 1982; Giorgi a kol., 2003a; Steimer a Driscoll, 2003), vykazujú odlišné vzorce správania v prítomnosti neprístupných vnímavých žien a počas klasických kopulačných testov (Sanna a kol., 2014a,b, 2015). Preto táto štúdia tiež ukazuje, že sexuálne naivné RHA potkany (nikdy predtým vystavené vnímavej žene) vykazovali vyššiu sexuálnu motiváciu a lepšiu sexuálnu výkonnosť v porovnaní so sexuálne naivnými RLA potkanmi, napr. RHA potkany vykazujú vyšší počet bezkontaktných erekcií penisu (feromónové indukované erekcie penisu považované za index sexuálneho vzrušenia; Sachs a kol., 1994; Sachs, 2000) a rôzne pro-sexuálne zmeny v niekoľkých kopulačných parametroch, ako sa zistilo v predchádzajúcich štúdiách (Sanna a kol., 2014a,b, 2015). Tieto rozdiely stále pretrvávajú v oboch líniách rímskych potkanov po získaní sexuálnej skúsenosti, to znamená po stabilizácii sexuálneho správania s piatimi expozíciami a kopulačnými testami s receptívnou ženou. Táto štúdia tiež prvýkrát ukazuje, že v obidvoch RHA a RLA potkaních líniách, buď v naivnom alebo skúsenom stave, bezdotykovej erekcie penisu a sexuálnej interakcie, došlo súčasne so zvýšením koncentrácií extracelulárneho dopamínu (a jeho hlavného metabolitu DOPAC). ) a NA v dialyzátoch získaných intracerebrálnou mikrodialýzou z mPFC oboch línií rímskych potkanov. Je zaujímavé, že tieto zvýšenia extracelulárneho dopamínu a NA v mPFC sa zdajú byť spojené s rozdielmi vo fenotype potkanov alebo v úrovni sexuálneho zážitku. V súlade s tým, ako je podrobne uvedené v časti Výsledky, tak sexuálne naivné, ako aj skúsené RHA potkany vykazovali vyššie a dlhotrvajúce zvýšenia extracelulárneho dopamínu a NA v porovnaní s ich RLA náprotivkami v apetitívnej aj konzumnej fáze sexuálneho správania. Navyše, sexuálne skúsené potkany oboch rímskych línií vykazovali vyššie a dlhotrvajúce zvýšenia extracelulárneho dopamínu a NA v mPFC v oboch fázach sexuálneho správania v porovnaní so svojimi naivnými náprotivkami. Nakoniec všetky vyššie uvedené rozdiely v extracelulárnom dopamíne a NA v mPFC zjavne súviseli s rozdielmi v behaviorálnych parametroch sexuálneho správania, napr. Sexuálne skúsené potkany oboch rímskych línií vykazovali vyššiu sexuálnu motiváciu a lepšie kopulačné výkony v porovnaní so svojimi naivnými náprotivkami. a sexuálne naivné a skúsené RHA potkany vykazovali vyššiu sexuálnu motiváciu a lepšie kopulačné výkony v porovnaní so svojimi sexuálne naivnými a skúsenými náprotivkami RLA.
Pokiaľ ide o zvýšenie extracelulárneho dopamínu zisteného v dialyzáte mPFC počas sexuálnej aktivity, k prvému zvýšeniu došlo v prvej 15 min po zavedení sexuálne vnímavej ženy do párovacieho prístroja, keď bola samica neprístupná samcovi, čo zvyčajne ukazuje v tomto stave bezkontaktná erekcia penisu. K druhému a väčšiemu zvýšeniu extracelulárneho dopamínu došlo v priebehu prvého 15 min. Po tom, ako bola samica sprístupnená samcovi na kopuláciu. Je zaujímavé, že zatiaľ čo v naivnom stave sa extracelulárny dopamín zväčšil hlavne v prvej časti testu s prístupnou ženou so sklonom k návratu k bazálnym hodnotám v druhej časti, v sexuálne skúsenom stave dopamín zostal počas testu zvýšený a dosiahol najvyššie hodnoty. po 45 – 60 min. párovania v oboch líniách rímskych potkanov.
Okrem vyššie uvedených rozdielov v extracelulárnom dopamíne počas rôznych fáz sexuálnej aktivity medzi RHA a RLA potkanmi v sexuálne naivnom a skúsenom stave, táto štúdia tiež ukazuje neočakávané dôležité a vysoko významné rozdiely v extracelulárnej DOPAC koncentrácii medzi dvoma rímskymi potkanými líniami predtým (tj. (základné hodnoty) a po zavedení vnímavej ženy do párovacieho zariadenia. Preto sa zistilo, že bazálne hodnoty extracelulárneho DOPAC v mPFC dialyzáte sú u RHA potkanov asi X-násobne vyššie ako hodnoty zistené u potkanov RLA. Tento rozdiel medzi RHA a RLA potkanmi nastal a pretrvával v sexuálne naivných a skúsených podmienkach napriek veľmi miernym rozdielom v bazálnych hodnotách extracelulárneho dopamínu v mPFC dialyzátoch získaných z dvoch línií rímskych potkanov za týchto experimentálnych podmienok. Celkovo by tieto rozdiely mohli naznačovať rozdielny obrat dopamínu na úrovni mPFC medzi dvoma líniami rímskych potkanov. V súlade s tým koncentrácie DOPAC zvyčajne odrážajú množstvo uvoľneného dopamínu a potom znovu zachytené pomocou dopaminergných nervových terminálov a konvertované na DOPAC pomocou monoaminooxydázy (MAO; Carlsson, 1975). Vyššie bazálne koncentrácie DOPAC nájdené u potkanov RHA teda naznačujú, že vyššie množstvá dopamínu sa uvoľňujú a znovu zachytávajú pomocou dopaminergných nervových zakončení v mPFC potkanov RHA, napr. Vyššia bazálna aktivita mezokortikálneho dopaminergného systému potkanov RHA v porovnaní s RLA. krysy. Vyšší mezokortikálny dopaminergný tonus u RHA krýs než u RLA krýs je tiež podporovaný vyššími extracelulárnymi koncentráciami dopamínu zistenými počas sexuálnej aktivity v mPFC dialyzáte od sexuálne naivných a skúsených RHA krýs, pokiaľ ide o ich RLA náprotivky. V tejto súvislosti je potrebné pripomenúť, že vyšší dopaminergný tón sa považuje za aspoň čiastočne zodpovedný za mnoho rôznych a dokonca opačných behaviorálnych znakov prítomných u potkanov RHA a RLA. V súlade s tým sú RHA potkany aktívnymi kopírovateľmi, vysoko impulzívnymi hľadačmi novosti a senzácie a sú náchylné na príjem a zneužívanie niekoľkých skupín drog závislosti, zatiaľ čo RLA potkany sú reaktívne kopolyméry, hyperemotionické a sú náchylné na rozvoj symptómov podobných depresii (Zeier a kol., 1978; Giorgi a kol., 1994, 2003b, 2007; Corda a kol., 1997, 2014; Escorihuela a kol., 1999; Steimer a Driscoll, 2003; Lecca a kol., 2004; Giménez-Llort a kol., 2005; Carrasco a kol., 2008; Fattore a kol., 2009; Moreno a kol., 2010; Coppens a kol., 2012; Díaz-Morán a kol., 2012; Sabariego a kol., 2013; Manzo a kol., 2014a,b; Oliveras a kol., 2015), vrátane vyššej sexuálnej motivácie a lepšieho kopulačného výkonu sexuálne naivných a skúsených RHA potkanov oproti RHA potkanom (Sanna a kol., 2015). Vyšší nárast koncentrácií extracelulárneho dopamínu zistený v dialyzáte z mPFC sexuálne naivných a skúsených RHA potkanov sa podobá vyšším koncentráciám extracelulárneho dopamínu zisteným v dialyzáte z jadra accumbens sexuálne naivných a zaznamenaných RHA potkanov, pokiaľ ide o ich RLA náprotivky. (Sanna a kol., 2015). Avšak na rozdiel od mPFC sa bazálne hladiny extracelulárneho dopamínu a DOPAC v dialyzáte z jadra accumbens sexuálne naivných a skúsených RHA a RLA krýs významne nelíšili (Sanna a kol., 2015). Vyšší mezolimbický dopaminergný tonus v jadre accumbens sexuálne naivných a skúsených RHA potkanov môže byť odvodený iba z vyšších extracelulárnych koncentrácií dopamínu zistených v dialyzátoch z jadra accumbens RHA potkanov v porovnaní s RLA potkanmi v prítomnosti receptívnych. samice, a nie odlišnou bazálnou dopaminergnou aktivitou odhalenou vyššími a nižšími bazálnymi hladinami DOPAC u potkanov RHA a RLA, v tomto poradí, ako sa zistilo v mPFC v tejto štúdii. To je tiež v súlade s výsledkami predchádzajúcich správ, ktoré ukazujú, že hladiny extracelulárneho dopamínu v dialyzáte z mPFC potkanov RHA boli nájdené vyššie ako hladiny potkanov RLA, zatiaľ čo podobné hladiny dopamínu boli nájdené v dialyzáte z obalu jadra accumbens oboch RHA a RLA potkanov za rôznych experimentálnych podmienok (D'Angio a kol., 1988; Scatton a kol., 1988; Willig a kol., 1991; Giorgi a kol., 2003a, 2007). Napriek vyššie uvedeným rozdielom medzi mPFC a nucleus accumbens sa zdá, že sexuálna skúsenosť vyvoláva v mPFC dlhšie trvajúce zvýšenia časovej schémy uvoľňovania dopamínu v oboch líniách rímskych potkanov pri prechode zo sexuálne naivnej na skúsený stav, hoci sa takéto zvýšenia zvyšujú boli obvykle vyššie u potkanov RHA ako u potkanov RLA. Je to trochu odlišné od toho, čo sa nachádza v nucleus accumbens, kde sa sexuálna skúsenosť tendenciu posunúť extracelulárny dopamín zvyšuje na prvú časť kopulačného testu u RHA potkanov, zatiaľ čo sa zvyšuje počas celého kopulačného testu u RLA potkanov (Sanna a kol., 2015).
Podľa našich vedomostí táto štúdia tiež prvýkrát ukazuje, že nielen extracelulárny dopamín, ale aj extracelulárny NA sa zvyšuje počas sexuálnej aktivity v dialyzáte z mPFC oboch RHA a RLA potkanov, ako sa zistilo u dopamínu, ale aj v tomto prípade s rozdiely súvisiace s líniou rímskych potkanov a so sexuálnymi skúsenosťami. Najmä pri odchýlke od bazálnych hladín dopamínu, ktoré sa zistili veľmi podobné v dialyzáte z mPFC oboch RHA a RLA krýs, boli základné hladiny extracelulárnej NA v dialyzáte z mPFC RHA krýs významne vyššie (približne 2,5-krát ) ako u potkanov RLA. Dôvody týchto rozdielov nie sú známe, ale ako už bolo uvedené vyššie pre dopamín a DOPAC, tieto výsledky môžu naznačovať, že vyšší bazálny noradrenergický tonus existuje prinajmenšom v mPFC kôre RHA potkanov v porovnaní s RLA potkanmi. Taký vyšší noradrenergický tón môže byť tiež zapojený do rôznych a často opačných behaviorálnych znakov prítomných u RHA a RLA potkanov, ako je uvedené vyššie pre dopamín. Ďalšia podpora vyššieho noradrenergického tónu u RHA potkanov ako u RLA potkanov v mPFC vychádza z výsledkov, ktoré ukazujú, že rozdiely v bazálnych extracelulárnych hladinách NA pretrvávali aj pri sexuálnej aktivite tak v sexuálne naivných, ako aj v skúsených podmienkach, napr. Hladiny extracelulárnych NA sa zvýšili v mPFC dialyzát u RHA potkanov, keď bol vystavený neprístupnej samici a počas celého obdobia kopulácie oveľa viac ako u potkanov RLA, v ktorých hladiny NA stúpli iba v prvých 15 min z obdobia kopulácie, potom sa hladiny NA vrátili späť na základné hodnoty , Tieto zmeny v extracelulárnych hladinách NA sa pozorovali tak v sexuálne naivnom, ako aj v skúsenom stave, ale zvýšenie hladín NA bolo vyššie u sexuálne skúsených potkanov RHA a RLA v porovnaní s ich sexuálne naivnými náprotivkami. Sexuálna skúsenosť teda zjavne vyvoláva ďalšie zosilnenie noradrenergickej aktivity v mPFC RHA potkanov a v menšej miere RLA potkanov, ako už bolo diskutované pre dopamín. Sprievodné vyššie noradrenergické a dopaminergné tóny v mPFC RHA potkanov v porovnaní s RLA potkanmi si zaslúžia nejaký komentár. V skutočnosti sa uvádza, že noradrenergická aktivita v mPFC môže ovplyvňovať uvoľňovanie dopamínu rôznymi mechanizmami (napríklad pozri Carboni a kol., 1990, 2006; Gresch a kol., 1995; Westernik a kol., 1998 a odkazy na ne). Jeden z nich súvisí s aktivitou NET, o ktorej bolo dokázané, že viaže dopamín s ešte vyššou afinitou k NA pre synaptozómy získané z homogenátov PFC (Horn, 1973). Keby k tomu došlo v mPFC in vivo, dopamín uvoľnený v mPFC mohol byť zachytený nielen dopaminergnými, ale tiež noradrenergickými nervovými zakončeniami rýchlosťou ešte vyššou ako NA. To by mohlo zase spôsobiť zvýšenie uvoľňovania dopamínu z dopaminergných terminálov (napr. Zvýšený dopaminergný tón), aby sa kompenzovalo množstvo dopamínu odstráneného zo synaptickej štrbiny; teda vyšší noradrenergický tón nájdený u RHA potkanov by mohol prispievať, aspoň čiastočne, k vyššiemu dopaminergnému tónu zistenému u mPFC RHA potkanov v porovnaní s RLA potkanmi. Na overenie tejto možnosti sú potrebné ďalšie štúdie. Najmä rozdiel v noradrenergickej aktivite zistený v tejto štúdii v mPFC medzi potkanmi RHA a RLA môže tiež hrať úlohu v ich odlišnej reakcii na liečbu antidepresívami. V skutočnosti sú potkany RHA považované za model odolnosti a potkany RLA za model náchylnosti k depresii. Preto akútna alebo chronická liečba antidepresívami, ako je desipramín, fluoxetín a chlórimipramín, všetky významne zlepšili behaviorálne reakcie pri teste vynúteného plávania u potkanov RLA bez ovplyvnenia odpovedí potkanov RHA (Piras a kol., 2010, 2014).
Táto štúdia napokon potvrdzuje a rozširuje predchádzajúce zistenia, ktoré ukazujú, že mPFC sa podieľa na kontrole sexuálneho správania, hoci presná úloha tejto oblasti mozgu v sexuálnom správaní nie je ani zďaleka jasná (Fernández-Guasti a kol., 1994; Agmo a Villalpando, 1995; Agmo et al., 1995; Hernández-Gonzáles a kol., 1998, 2007; Kakeyama a kol., 2003; Balfour a spol., 2006; Afonso a kol., 2007; Davis a kol., 2010; Febo, 2011). Nedávno sa ukázalo selektívne vypaľovanie buniek v mPFC počas priblíženia správania sa samca potkana smerom k neprístupnej sexuálne vnímavej žene s použitím paradigmy záznamu jednej paľby (Febo, 2011). Je zaujímavé, že neuróny, ktoré neodpovedali počas prvej expozície samca neprístupnej vnímavej žene, sa počas druhej expozície aktivujú, akoby predchádzajúca skúsenosť dokázala vyvolať plastické zmeny, ktoré viedli k zmenám v reakciách na správanie. Zistilo sa však, že lézie s kyselinou iboténovou mPFC nie sú schopné zmeniť sexuálne správanie samcov potkanov so sexuálne vnímavou ženou, ani vyjadrenie podmieneného preferencie miesta pre sexuálnu odmenu. Schopnosť tvoriť podmienenú averziu voči sexuálnej aktivite, keď bola spárovaná s averzívnymi stimulmi, sa však u týchto poškodených zvierat úplne zrušila (Davis a kol., 2010). To naznačuje, že aktivácia mPFC počas sexuálneho správania zohráva úlohu pri integrácii vonkajších a vnútorných informácií na vykonávanie a kontrolu správania zameraného na cieľ, a nie na vyjadrovanie vrodených reakcií na prírodné zosilňovače (pozri pozri časť 4.4). Goto a Grace, 2005). Dôležitejšie pre túto prácu bolo niekoľko štúdií, ktoré ukázali, že oblasti mozgu zapojené do sexuálnej motivácie a sexuálneho správania, ako sú napríklad nucleus accumbens, ventrálna tegmentálna oblasť, stredná predoptická oblasť, lôžkové jadro stria terminis, basolaterálne amygdaly a parvocelulárne subparafaskulárne thalamické jadro, dostávajú vstupy z mPFC počas sexuálnej aktivity (pozri Balfour a spol., 2006). Mnoho z týchto oblastí mozgu sa podieľa na komplexnom nervovom okruhu zapojenom do kontroly sexuálneho správania, od sexuálnej motivácie a odmeňovania po sexuálny výkon. Tento obvod zahŕňa oxytocinergné neuróny, ktoré pochádzajú z PVN hypotalamu (PVN) a vyčnievajú do ventrálnej oblasti tegmentálu, nucleus accumbens, hippocampus, amygdala, PFC, lôžko jadra stria terminalis, medulla oblongata a miecha, mezolimbické / mezokortikálne dopaminergné neuróny vyčnievajúce z ventrálnej tegmentálnej oblasti do nucleus accumbens a PFC, incertohypotalamické dopaminergné neuróny a glutamatergické neuróny, ktoré sa zúčastňujú na miestnej a systémovej úrovni v niekoľkých oblastiach obvodu (pozri pozri časť 4.4). Melis a Argiolas, 1995, 2011; Melis a kol., 2003, 2007, 2009, 2010; Succu a kol., 2007, 2008, 2011). Zvýšenie extracelulárneho dopamínu (ale tiež NA) v dialyzátoch z mPFC pozorované počas sexuálnej aktivity by teda mohlo naznačovať, že mPFC je aktívna súčasť tohto okruhu, to znamená, že mezokortikálne dopaminergné neuróny sa spoločne zúčastňujú na kontrole sexuálnej aktivity. s noradrenergickými, mezolimbickými dopaminergnými, oxytocinergnými a glutamatergickými neurónmi, aj keď sa na vyjadrenie sexuálneho správania nejaví nevyhnutne (Davis a kol., 2010). V súlade s touto hypotézou táto štúdia ukazuje, že rozdiely v uvoľňovaní dopamínu a NA v mPFC počas sexuálnej aktivity sa vyskytli súbežne s rozdielmi v počte bezkontaktných erekcií penisu a zmenami kopulačných parametrov, s vyšším uvoľňovaním dopamínu a NA v priebehu chutné a konzumné fázy sa týkali vyššej sexuálnej motivácie a lepších kopulačných výkonov v oboch líniách rímskych potkanov, ako aj v sexuálne naivných a skúsených podmienkach.
Vyššie uvedená diskusia je založená hlavne na dobre známom zvýšení extracelulárneho dopamínu, ku ktorému dochádza v jadrovom mozgu samcov potkanov počas sexuálnej aktivity (Pfaus a kol., 1990; Pleim a kol., 1990; Pfaus a Phillips, 1991; Damsma a kol., 1992; Wenkstern a kol., 1993). Dopamín a iné neurotransmitery sa však môžu uvoľňovať v mPFC (a v nucleus accumbens) za iných experimentálnych podmienok, napríklad počas stresu alebo zvládania (Thierry a kol., 1976; Fadda a kol., 1978), ktoré nesúvisia výlučne so sexuálnym správaním. Možno teda tvrdiť, že zvýšenie dopamínu zistené v extracelulárnom dopamíne v mPFC (táto štúdia) alebo v nucleus accumbens (Sanna a kol., 2015) súvisí skôr so sekundárnym stresom alebo s vyrovnaním sa s prítomnosťou a / alebo interakciou so sexuálne vnímajúcou ženou než so sexuálnou aktivitou. To by mohlo mať určitý význam u RHA a RLA potkanov, ktoré vykazujú výrazne odlišné štýly zvládania záťaže a rôzne aktivácie osi hypothalamicko-hypofýza-nadobličky v reakcii na averzívne podmienky (potkany RLA vykazujú aktiváciu tejto osi oveľa vyššiu ako aktivácia RHA krýs; pozri časť Carrasco a kol., 2008; Díaz-Morán a kol., 2012). To tiež zvyšuje pravdepodobnosť, že stresové hormóny (tj kortikosteroidy) môžu ovplyvňovať dopaminergnú aktivitu v mPFC a ďalších mozgových oblastiach. Potkany RLA sú skutočne reaktívne kopolyméry a vykazujú hyperemotionálne správanie charakterizované hypomotilitou a mrazom, zatiaľ čo potkany RHA vykazujú proaktívne zvládanie správania zamerané na získanie kontroly nad stresorom (Driscoll a Bättig, 1982; Willig a kol., 1991; Escorihuela a kol., 1999; Steimer a Driscoll, 2003; Giorgi a kol., 2007). Aj keď nie je možné úplne vylúčiť, že k tomu môže dôjsť u sexuálne naivných potkanov (ktoré nikdy predtým neinteragovali s vnímavou samičkou), a najmä u sexuálne naivných RLA potkanov (pozri vyššie), je to nepravdepodobné. V našich experimentálnych podmienkach je urobené všetko pre to, aby sa zabránilo všetkým druhom stresu (pozri časť „Materiály a metódy“) a experimenty sú organizované tak, aby muži so ženou neinteragovali okamžite, ale až po 30 minútach expozície jej. , počas ktorých je nameraný nárast dopamínu oveľa nižší ako nárast, ku ktorému dochádza počas sexuálnej interakcie. Ak zvýšenie dopamínu zistené v mPFC (a v nucleus accumbens) rímskych potkanov (sexuálne naivné potkany RHA a RLA mali bazálne hladiny extracelulárneho dopamínu v dialyzáte z mPFC a nucleus accumbens, veľmi podobné), boli skutočne sekundárne k stres v dôsledku alebo interakcia so ženou: (i) zvýšenie dopamínu by sa zistilo oveľa vyššie v prvom období, keď sú muži v prítomnosti ženy, a nie neskôr, ako sa zistilo počas kopulácie; a ii) dalo by sa očakávať, že zvýšenie dopamínu zmizne alebo sa výrazne zníži po dosiahnutí stabilnej sexuálnej aktivity, to znamená po dokončení procesov učenia (vrátane zvládania nových vecí), ktoré budú hrať pri vykonávaní sexuálnych aktivít iba malú úlohu. naopak, stále sa vyskytuje, keď sa sexuálna aktivita opakuje. Inými slovami, extracelulárny dopamín v mPFC (a v nucleus accumbens) sa počas kopulácie vždy zvyšuje, dokonca aj po naučení sa sexuálneho správania. Na objasnenie funkčnej úlohy tohto zvýšenia aktivity dopamínu v mPFC (a v nucleus accumbens; pozri časť „Úvod a odkazy“) sú potrebné ďalšie štúdie.
Záverom je možné konštatovať, že táto štúdia prvýkrát ukazuje, že prítomnosť a ešte viac interakcia so sexuálne vnímavou ženou vedie k zvýšeniu extracelulárneho dopamínu a NA v dialyzáte z mPFC sexuálne naivných (nikdy nevystavených sexuálnym stimulom). ) a sexuálne skúsenosti (ktoré podstúpili päť predbežných kopulačných testov a vykazujú stabilné sexuálne výkony), potkany RHA a RLA, ktoré vykazujú zreteľne odlišné štýly zvládania reakcie v reakcii na averzívne podmienky a rôzne vzorce kopulačného správania so sexuálne vnímavou ženou. V súlade s tým sa zistilo, že koncentrácie dopamínu a NA sú vyššie v dialyzáte od sexuálne naivných a skúsených RHA potkanov v porovnaní s ich RLA náprotivkami v priebehu predikčnej aj konzumnej fázy sexuálnej aktivity. Tieto výsledky môžu byť čiastočne spôsobené dopaminergným a noradrenergickým tonom v mPFC u RHA potkanov vyšším ako u RLA potkanov, ako sa uvádza pre dopamín v nucleus accumbens (Sanna a kol., 2015). Okrem toho sa zistilo, že extracelulárny mPFC dopamín a NA sú tiež vyššie v sexuálne skúsených RHA a v menšej miere RLA potkany v porovnaní so svojimi sexuálne naivnými náprotivkami v bazálnych podmienkach, ako aj počas chutných a konzumných fáz sexuálneho správania. To naznačuje, že sexuálna skúsenosť indukuje plastické procesy, ktoré ďalej zosilňujú neurotransmisiu dopamínu a NA v mPFC, ako sa zistilo pre dopamín v jadre accumbens oboch línií rímskych potkanov (Sanna a kol., 2015). Vyššie uvedené rozdiely v dopaminergnej a noradrenergickej neurotransmisii v mPFC dvoch rímskych línií môžu hrať úlohu nielen v rôznych vzorcoch sexuálneho správania, ale aj v rôznych a často opačných behaviorálnych znakoch, ktoré charakterizujú tieto dve línie krýs.
Príspevky od autorov
Projekt navrhli FS, AA, MRM, OG a MGC. FS a JB navrhli, vykonali a analyzovali údaje z experimentov sexuálneho správania a mikrodialýzy. MAP, OG a MGC vybrané a chované rímske potkany. Štúdiu viedli FS, AA, MRM, OG a MGC. Rukopis napísali FS, AA, MRM, OG a MGC. Všetci autori diskutovali o výsledkoch a komentovali rukopis.
Financovanie
Táto práca bola čiastočne podporená grantmi talianskeho ministerstva univerzity a výskumu (MIUR) (vedecká nezávislosť mladých výskumných pracovníkov, SIR 2014, kód č. RBSI14IUX7) pre FS, z autonómnej oblasti Sardínia (ARS) (LR 7 / 2007). „Podpora vedeckého výskumu a technologických inovácií na Sardínii“, Európsky sociálny fond, 2007 – 2013, kód projektu č. CRP-59842) pre OG a z University of Cagliari na AA, MRM, FS, OG a MGC.
Vyhlásenie o konflikte záujmov
Autori vyhlasujú, že výskum bol vykonaný bez obchodných alebo finančných vzťahov, ktoré by mohli byť interpretované ako potenciálny konflikt záujmov.
Poďakovanie
Barbara Tuveri, Univerzita v Cagliari, pre ustajnenie a starostlivosť o zvieratá.
Referencie
Afonso, VM, Sison, M., Lovic, V. a Fleming, AS (2007). Mediálne prefrontálne kortexové lézie u samíc potkanov ovplyvňujú sexuálne a materské správanie a ich sekvenčnú organizáciu. Behave. Neurosci. 121, 515-526. dva: 10.1037 / 0735-7044.121.3.515
Agmo, A. a Villalpando, A. (1995). Stimulanty centrálneho nervového systému uľahčujú sexuálne správanie u samcov potkanov so strednými léziami prefrontálnej kortexu. Brain Res. 696, 187–193. doi: 10.1016/0006-8993(95)00853-i
Agmo, A., Villalpando, A., Picker, Z., a Fernández, H. (1995). Lézie stredného prefrontálneho kortexu a sexuálne správanie u samcov potkana. Brain Res. 696, 177–186. doi: 10.1016/0006-8993(95)00852-h
Argiolas, A., a Melis, MR (1995). Neuromodulácia erekcie penisu: prehľad úlohy neurotransmiterov a neuropeptidov. Prog. Neurobiol. 47, 235–255. doi: 10.1016/0301-0082(95)00020-8
Argiolas, A., a Melis, MR (2005). Centrálna kontrola erekcie penisu: úloha paraventrikulárneho jadra hypotalamu. Prog. Neurobiol. 76, 1-21. dva: 10.1016 / j.pneurobio.2005.06.002
Argiolas, A., a Melis, MR (2013). Neuropeptidy a centrálna kontrola sexuálneho správania od minulosti do súčasnosti: prehľad. Prog. Neurobiol. 108, 80-107. dva: 10.1016 / j.pneurobio.2013.06.006
Balfour, ME, Brown, JL, Yu, L. a Coolen, LM (2006). Potenciálny prínos efferentov zo stredného prefrontálneho kortexu k nervovej aktivácii po sexuálnom správaní u samcov potkana. Neurovedy 137, 1259-1276. dva: 10.1016 / j.neuroscience.2005.11.013
Balfour, ME, Yu, L. a Coolen, LM (2004). Prostredie sexuálneho správania a sexuálne prostredie aktivujú mezolimbický systém u samcov potkanov. neuropsychofarmakologie 29, 718-730. dva: 10.1038 / sj.npp.1300350
Beloate, LN, Omrani, A., Adan, RA, Webb, IC a Coolen, LM (2016). Aktivácia dopamínových buniek vo ventrálnej oblasti tegmentálu počas sexuálneho správania samcov potkanov reguluje neuroplasticitu a krížovú senzibilizáciu d-amfetamínu po abstinencii pohlavia. J. Neurosci. 36, 9949-9961. dva: 10.1523 / JNEUROSCI.0937-16.2016
Berridge, K., a Robinson, T. (1998). Aká je úloha dopamínu v odmeňovaní: hedonický dopad, učenie sa odmeňovania alebo stimulačný dôraz? Brain Res. Rev. 28, 309–369. doi: 10.1016/s0165-0173(98)00019-8
Bignami, G. (1965). Selekcia pre vysoké dávky a nízke dávky kondicionovania u potkanov. Anim. Behave. 13, 221–227. doi: 10.1016/0003-3472(65)90038-2
Broadhurst, PL a Bignami, G. (1965). Korelujúce účinky psychogénneho výberu: štúdia rímskych kmeňov krýs s vysokým a nízkym únikom. Behave. Res. Ther. 3, 273–280. doi: 10.1016/0005-7967(64)90033-6
Carboni, E., Silvagni, A., Vacca, C., a Di Chiara, G. (2006). Kumulatívny účinok blokády norepinefrínu a dopamínu na extracelulárny nárast dopamínu v jadre accumbens, v jadre lôžka stria terminalis a prefrontálnej kôre. J. Neurochem. 96, 473-481. dva: 10.1111 / j.1471-4159.2005.03556.x
Carboni, E., Tanda, GL, Frau, R., a Di Chiara, G. (1990). Blokáda noradrenalínového nosiča zvyšuje koncentrácie extracelulárneho dopamínu v prefrontálnej kôre: dôkaz, že dopamín je absorbovaný in vivo noradrenergickými terminálmi. J. Neurochem. 55, 1067–1070. doi: 10.1111/j.1471-4159.1990.tb04599.x
Carlsson, A. (1975). "Receptorom sprostredkovaná kontrola metabolizmu dopamínu," v Pre- a postsynaptické receptory, ed. E. Usdin a WE Bunney (New York, NY: Marcel Deker), 49 – 65.
Carrasco, J., Márquez, C., Nadal, R., Tobeña, A., Fernández-Teruel, A., a Armario, A. (2008). Charakterizácia centrálnych a periférnych zložiek osi hypotalamus-hypofýza-nadobličky v inbredných kmeňoch rímskych potkanov. Psychoneuroendocrinology 33, 437-445. dva: 10.1016 / j.psyneuen.2008.01.001
Coppens, CM, de Boer, SF, Steimer, T., a Koolhaas, JM (2012). Impulzívnosť a agresívne správanie u rímskych krýs s vysokým a nízkym únikom: základné rozdiely a zmeny vyvolané sociálnym stresom u dospievajúcich. Physiol. Behave. 105, 1156 – 1160. doi: 10.1016 / j.physbeh.2011.12.013
Corda, MG, Lecca, D., Piras, G., Di Chiara, G., a Giorgi, O. (1997). Biochemické parametre dopamínergnej a GABAergickej neurotransmisie v CNS rímskych potkanov s vysokým a minimálnym vyhýbaním sa. Behave. Genet. 27, 527-536. dva: 10.1023 / A: 1021452814574
Corda, MG, Piras, G., Piludu, MA, a Giorgi, O. (2014). Diferenciálne účinky dobrovoľnej konzumácie etanolu na produkciu dopamínu v jadre accumbens škrupiny rímskych potkanov s vysokou a nízkou vyhýbateľnosťou: štúdia behaviorálnej a mozgovej mikrodialýzy. World J. Neurosci. 4, 279 – 292. doi: 10.4236 / wjns.2014.43031
Dahlstrom, A. a Fuxe, K. (1964). Dôkaz o existencii neurónov obsahujúcich monoamíny v centrálnom nervovom systéme. I. Preukázanie monoamínov v bunkách tela neurónov mozgu. Acta Physiol. Scanda. Suppl. 62, 1-54.
Damsma, G., Pfaus, J., Wenkstern, D., Phillips, A., a Fibiger, H. (1992). Sexuálne správanie zvyšuje prenos dopamínu v nucleus accumbens a striatum samcov potkanov: porovnanie s novinkou a pohybom. Behave. Neurosci. 106, 181-191. dva: 10.1037 / 0735-7044.106.1.181
D'Angio, M., Serrano, A., Driscoll, P., a Scatton, B. (1988). Stresujúce environmentálne podnety zvyšujú hladiny extracelulárneho DOPAC v prefrontálnom kortexe hypoemotionických (rímske vysoké vyhýbanie sa), ale nie hyperemotionických (rímske nízke vyhýbanie sa). in vivo voltametrická štúdia. Brain Res. 451, 237–247. doi: 10.1016/0006-8993(88)90768-8
Davis, JF, Loos, M., Di Sebastiano, AR, Brown, JL, Lehman, MN a Coolen, LM (2010). Lézie stredného prefrontálneho kortexu spôsobujú maladaptívne sexuálne správanie u samcov potkanov. Biol. psychiatrie 67, 1199-1204. dva: 10.1016 / j.biopsych.2009.12.029
Díaz-Morán, S., Palècia, M., Mont-Cardona, C., Cañete, T., Blázquez, G., Martínez-Membrives, E., a kol. (2012). Reakcie na zvládanie štýlov a stresových hormónov u geneticky heterogénnych potkanov: porovnanie s kmeňmi rímskych potkanov. Behave. Brain Res. 228, 203 – 210. doi: 10.1016 / j.bbr.2011.12.002
Driscoll, P., a Bättig, K. (1982). „Behaviorálne, emocionálne a neurochemické profily potkanov vybraných pre extrémne rozdiely v aktívnom výkone obojsmerného vyhýbania sa,“ v Genetika mozgu, ed. I. Lieblich (Amsterdam: Elsevier), 95 – 123.
Dunlop, BW a Nemeroff, CB (2007). Úloha dopamínu v patofyziológii depresie. Arch. Gen. Psychiatry 64, 327-337. dva: 10.1001 / archpsyc.64.3.327
Escorihuela, RM, Fernández-Teruel, A., Gil, L., Aguilar, R., Tobeña, A., a Driscoll, P. (1999). Inbredné rímske potkany s vysokým a nízkym vyhýbaním sa: rozdiely v správaní sa úzkosti, v hľadaní novosti a v raketopláne. Physiol. Behave. 67, 19–26. doi: 10.1016/S0031-9384(99)00064-5
Everitt, BJ (1990). Sexuálna motivácia: nervová a behaviorálna analýza mechanizmov, ktoré sú základom chuti do jedla a kopulácie u samcov potkanov. Neurosci. Biobehav. Rev. 14, 217–232. doi: 10.1016/s0149-7634(05)80222-2
Everitt, BJ a Robbins, TW (2005). Neurónové systémy posilňovania drogovej závislosti: od činov k návykom k donucovaniu. Nat. Neurosci. 8, 1481 – 1489. doi: 10.1038 / nn1579
Fadda, F., Argiolas, A., Melis, MR, Tissari, AH, Onali, PL, a Gessa, GL (1978). Stresom vyvolané zvýšenie hladín kyseliny 3,4-dihydroxyfenyloctovej (DOPAC) v mozgovej kôre a n.akcumbénoch: zvrat diazepamom. Life Sci. 23, 2219–2224. doi: 10.1016/b978-0-08-023768-8.51934-4
Fattore, L., Piras, G., Corda, MG, a Giorgi, O. (2009). Rímske línie krýs s vysokým a nízkym vyhýbaním sa líšia v získavaní, udržiavaní, vyhynutí a obnovení intravenózneho podania kokaínu. neuropsychofarmakologie 34, 1091-1101. dva: 10.1038 / npp.2008.43
Febo, M. (2011). Prefrontálne vypaľovanie buniek u samcov potkanov počas prístupu k sexuálne vnímavej žene: interakcie s kokaínom. Synapse 65, 271 – 277. doi: 10.1002 / syn.20843
Fernández-Guasti, A., Omaña-Zapata, I., Luján, M., a Condés-Lara, M. (1994). Účinky ligácie sedacieho nervu na sexuálne správanie sexuálne skúsených a neskúsených samcov potkanov: účinky dekortikácie predného pólu. Physiol. Behave. 55, 577–581. doi: 10.1016/0031-9384(94)90119-8
Fernández-Teruel, A., Driscoll, P., Gil, L., Aguilar, R., Tobeña, A., a Escorihuela, RM (2002). Pretrvávajúce účinky obohacovania životného prostredia na hľadanie nových výrobkov, príjem sacharínu a etanolu v dvoch líniách potkanov (RHA / Verh a RLA / Verh) sa líšia v motívoch pri hľadaní správania. Pharmacol. Biochem. Behave. 73, 225–231. doi: 10.1016/s0091-3057(02)00784-0
Giménez-Llort, L., Cañete, T., Guitart-Masip, M., Fernández-Teruel, A., a Tobeña, A. (2005). Dva charakteristické fenotypy indukované apomorfínom u rímskych krýs s vysokým a nízkym vyhýbaním sa. Physiol. Behave. 86, 458 – 466. doi: 10.1016 / j.physbeh.2005.07.021
Giorgi, O., Lecca, D., Piras, G., a Corda, MG (2005). Diferenciálna aktivácia uvoľňovania dopamínu v jadre accumbens po akútnych alebo opakovaných injekciách amfetamínu: porovnávacia štúdia v rímskych líniách krýs s vysokým a nízkym vylučovaním. Neurovedy 135, 987-998. dva: 10.1016 / j.neuroscience.2005.06.075
Giorgi, O., Lecca, D., Piras, G., Driscoll, P., a Corda, MG (2003a). Disociácia medzi mezokortikálnym uvoľňovaním dopamínu a chovaním spojeným so strachom v dvoch psychogeneticky vybraných líniách krýs, ktoré sa líšia stratégiou zvládania záhy averzívnych stavov. Eur. J. Neurosci. 17, 2716-2726. dva: 10.1046 / j.1460-9568.2003.02689.x
Giorgi, O., Piras, G., Lecca, D., Hansson, S., Driscoll, P., a Corda, MG (2003b). Diferenciálne neurochemické vlastnosti centrálneho serotonergného prenosu u rímskych potkanov s vysokým a nízkym vyhýbaním sa. J. Neurochem. 86, 422-431. dva: 10.1046 / j.1471-4159.2003.01845.x
Giorgi, O., Orlandi, M., Escorihuela, RM, Driscoll, P., Lecca, D., a Corda, MG (1994). GABAergický a dopaminergný prenos v mozgu rímskych potkanov s vysokou vyhýbavosťou a rímskych potkanov s nízkym vyhýbaním sa. Brain Res. 638, 133–138. doi: 10.1016/0006-8993(94)90642-4
Giorgi, O., Piras, G., a Corda, MG (2007). Psychogeneticky vybrané rímske línie krýs s vysokým a nízkym únikom: model na štúdium individuálnej zraniteľnosti jednotlivcov voči drogovej závislosti. Neurosci. Biobehav. Rev. 31, 148-163. dva: 10.1016 / j.neubiorev.2006.07.008
Goto, Y. a Grace, AA (2005). Dopaminergná modulácia limbického a kortikálneho pohonu jadra pripadá na cieľové správanie. Nat. Neurosci. 8, 805 – 812. doi: 10.1038 / nn1471
Gresch, PJ, Sved, AF, Zigmond, MJ a Finlay, JM (1995). Lokálny vplyv endogénneho norepinefrínu na extracelulárny dopamín v mediálnej prefrontálnej kôre potkanov. J. Neurochem. 65, 111-116. dva: 10.1046 / j.1471-4159.1995.65010111.x
Hernández-Gonzáles, M., Guevara, MA, Cervantes, M., Morali, G., a Corsi-Cabrera, M. (1998). Charakteristické frekvenčné pásma kortiko-frontálneho EEG počas sexuálnej interakcie samcov potkanov ako výsledok faktoriálnej analýzy. J. Physiology 92, 43–50. doi: 10.1016/s0928-4257(98)80022-3
Hernández-Gonzáles, M., Prieto-Beracoechea, Kalifornia, Arteaga-Silva, M. a Guevara, MA (2007). Rôzna funkčnosť mediálnej a orbitálnej prefrontálnej kôry počas sexuálne motivovanej úlohy u potkanov. Physiol. Behave. 90, 450 – 458. doi: 10.1016 / j.physbeh.2006.10.006
Horn, AS (1973). Vzťahy štruktúry a aktivity na inhibíciu absorpcie katecholamínu do synaptozómov z noradrenergných a dopaminergných neurónov v mozgových homogenátoch potkana. Br. J. Pharmacol. 47, 332–338. doi: 10.1111/j.1476-5381.1973.tb08331.x
Hull, EM, Du, J., Lorrain, DS a Matuszewich, L. (1995). Extracelulárny dopamín v strednej predoptickej oblasti: dôsledky pre sexuálnu motiváciu a hormonálnu kontrolu kopulácie. J. Neurosci. 15, 7465-7471.
Hull, EM, Lorrain, DS, Du, J., Matuszewich, L., Lumley, LA, Putnam, SK, a kol. (1999). Interakcie hormónov s neurotransmitermi pri kontrole sexuálneho správania. Behave. Brain Res. 105, 105–116. doi: 10.1016/s0166-4328(99)00086-8
Hull, EM, Weber, MS, Eaton, RC, Dua, R., Markowski, VP, Lumley, L., a kol. (1991). Dopamínové receptory vo ventrálnej oblasti tegmentálu ovplyvňujú motorické, ale nie motivujúce alebo reflexívne zložky kopulácie u samcov potkanov. Brain Res. 554, 72–76. doi: 10.1016/0006-8993(91)90173-S
Ikemoto, S., a Panksepp, J. (1999). Úloha jadra accumbens dopamínu v motivovanom správaní: zjednocujúca interpretácia s osobitným dôrazom na hľadanie odmeny. Brain Res. Rev. 31, 6–41. doi: 10.1016/S0165-0173(99)00023-5
Kakeyama, M., Sone, H., Miyabara, Y., a Tohyama, C. (2003). Perinatálna expozícia 2,3,7,8-tetrachlórdibenzo-p-dioxínu mení aktivitu závislú expresiu BDNF mRNA v neokortexe a sexuálne správanie samcov potkanov v dospelosti. Neurotoxicology 24, 207–217. doi: 10.1016/S0161-813x(02)00214-0
Lecca, D., Piras, G., Driscoll, P., Giorgi, O. a Corda, MG (2004). Diferenciálna aktivácia produkcie dopamínu v škrupinách a jadre nucleus accumbens je spojená s motorickými odpoveďami na návykové látky: štúdia na dialýze mozgu u rímskych potkanov s vysokým a nízkym únikom. Neuropharmacology 46, 688-699. dva: 10.1016 / j.neuropharm.2003.11.011
Manzo, L., Gómez, MJ, Callejas-Aguilera, JE, Donaire, R., Sabariego, M., Fernández-Teruel, A., a kol. (2014). Vzťah medzi preferenciou etanolu a hľadaním senzácie / novosti. Physiol. Behave. 133, 53 – 60. doi: 10.1016 / j.physbeh.2014.05.003
Manzo, L., Gómez, MJ, Callejas-Aguilera, JE, Fernández-Teruel, A., Papini, MR a Torres, C. (2014b). Samoliečba proti úzkosti vyvolaná motivačnou stratou u potkanov. Physiol. Behave. 123, 86 – 92. doi: 10.1016 / j.physbeh.2013.10.002
Masana, M., Bortolozzi, A., a Artigas, F. (2011). Selektívne zvýšenie mezokortikálneho dopaminergného prenosu noradrenergickými liekmi: terapeutické možnosti pri schizofrénii. Int. J. Neuropsychopharmacol. 14, 53-68. dva: 10.1017 / s1461145710000908
Meisel, RL a Sachs, BD (1994). "Fyziológia sexuálneho správania mužov", v Fyziológia reprodukcie, Zv. 2, 2 a Edn., Ed. E. Knobil a J. Neil (New York, NY: Raven Press), 3 – 96.
Melis, MR a Argiolas, A. (1995). Dopamín a sexuálne správanie. Neurosci. Biobehav. Rev. 19, 19–38. doi: 10.1016/0149-7634(94)00020-2
Melis, MR a Argiolas, A. (2011). Centrálna kontrola erekcie penisu: opätovná návšteva úlohy oxytocínu a jeho interakcie s dopamínom a kyselinou glutámovou u samcov potkanov. Neurosci. Biobehav. Rev. 35, 939-955. dva: 10.1016 / j.neubiorev.2010.10.014
Melis, MR, Melis, T., Cocco, C., Succu, S., Sanna, F., Pillolla, G., a kol. (2007). Oxytocín vstreknutý do ventrálnej tegmentálnej oblasti indukuje erekciu penisu a zvyšuje extracelulárny dopamín v nucleus accumbens a paraventrikulárnom jadre hypotalamu samcov potkanov. Eur. J. Neurosci. 26, 1026-1035. dva: 10.1111 / j.1460-9568.2007.05721.x
Melis, MR, Succu, S., Cocco, C., Caboni, E., Sanna, F., Boi, A., a kol. (2010). Oxytocín vyvoláva erekciu penisu, keď sa vstrekne do ventrálneho subikula: úloha oxidu dusnatého a kyseliny glutámovej. Neuropharmacology 58, 1153-1160. dva: 10.1016 / j.neuropharm.2010.02.008
Melis, MR, Succu, S., Mascia, MS, Cortis, L., a Argiolas, A. (2003). Extracelulárny dopamín zvyšuje paraventrikulárne jadro samcov potkanov počas sexuálnej aktivity. Eur. J. Neurosci. 17, 1266-1272. dva: 10.1046 / j.1460-9568.2003.02558.x
Melis, MR, Succu, S., Sanna, F., Boi, A., a Argiolas, A. (2009). Oxytocín vstreknutý do ventrálneho subikula alebo posteromediálneho kortikálneho jadra amygdaly indukuje erekciu penisu a zvyšuje hladiny extracelulárneho dopamínu v jadre accumbens samcov potkanov. Eur. J. Neurosci. 30, 1349-1357. dva: 10.1111 / j.1460-9568.2009.06912.x
Moreno, M., Cardona, D., Gómez, MJ, Sánchez-Santed, F., Tobeña, A., Fernández-Teruel, A., a kol. (2010). Charakterizácia impulzivity v rímskych kmeňoch potkanov s vysokým a nízkym únikom: rozdiely v správaní a neurochemické vlastnosti. neuropsychofarmakologie 35, 1198-1208. dva: 10.1038 / npp.2009.224
Oliveras, I., Río-Alamos, C., Cañete, T., Blázquez, G., Martínez-Membrives, E., Giorgi, O., a kol. (2015). Prepulzná inhibícia predpovedá výkon priestorovej pracovnej pamäte u inbredných rímskych potkanov s vysokým a nízkym vyhýbaním sa a u geneticky heterogénnych potkanov NIH-HS: relevantnosť pri štúdiu pre-pozorných a kognitívnych anomálií pri schizofrénii. Predná. Behave. Neurosci. 9: 213. dva: 10.3389 / fnbeh.2015.00213
Paxinos, G. a Watson, C. (2004). Krysa mozog v stereotaxických súradniciach, 4th Edn, New York: Academic Press.
Pfaus, JG (2010). Dopamín: Pomáhajú mužom kopulovať sa najmenej 200 miliónov rokov: teoretická poznámka o Kleitz-Nelson et al. (2010). Behave. Neurosci. 124, 877-880. dva: 10.1037 / a0021823
Pfaus, JG, Damsma, G., Nomikos, GG, Wenkstern, DG, Blaha, CD, Phillips, AG, a kol. (1990). Sexuálne správanie zvyšuje prenos dopamínu u samcov potkanov. Brain Res. 530, 345–348. doi: 10.1016/0006-8993(90)91309-5
Pfaus, JG a Everitt, BJ (1995). "Psychofarmakológia sexuálneho správania", v Psychofarmakológia: štvrtá generácia pokroku, ed. FE Bloom a DJ Kupfer (New York, NY: Raven Press), 742 – 758.
Pfaus, JG a Phillips, AG (1991). Úloha dopamínu v predvídacích a konzumných aspektoch sexuálneho správania u samcov potkanov. Behave. Neurosci. 105, 727-747. dva: 10.1037 / 0735-7044.105.5.727
Piras, G., Giorgi, O., a Corda, MG (2010). Účinky antidepresív na výkon v teste núteného plávania dvoch psychogeneticky vybraných línií potkanov, ktoré sa líšia stratégiou zvládania záhy averzívnych stavov. Psychofarmakológiu 211, 403–414. doi: 10.1007/s00213-010-1904-x
Piras, G., Piludu, MA, Giorgi, O., a Corda, MG (2014). Účinky chronickej antidepresívnej liečby na domnelý genetický model zraniteľnosti (rímske potkany s nízkym vyhýbaním sa) a rezistencie (rímske potkany s vysokým vyhýbaním sa) na depresiu vyvolanú stresom. Psychofarmakológiu 231, 43–53. doi: 10.1007/s00213-013-3205-7
Pitchers, KK, Frohmader, KS, Vialou, V., Mouzon, K., Nestler, EJ, Lehman, MN, a kol. (2010). FOSB v nucleus accumbens je rozhodujúce pre zosilnenie účinkov sexuálnej odmeny. Gény Brain Behav. 9, 831–840. doi: 10.1111/j.1601-183X.2010.00621.x
Pitchers, KK, Coppens, CM, Beloate, LN, Fuller, J., Van, S., Frohmader, KS, a kol. (2014). Endogénna opioidmi indukovaná neuroplasticita dopaminergných neurónov vo ventrálnej tegmentálnej oblasti ovplyvňuje prirodzenú a opiátovú odmenu. J. Neurosci. 34, 8825-8836. dva: 10.1523 / JNEUROSCI.0133-14.2014
Pitchers, KK, Vialou, V., Nestler, EJ, Laviolette, SR, Lehman, MN a Coolen, LM (2013). Prírodné a liekové výhody pôsobia na bežné mechanizmy nervovej plasticity s AFOS ako kľúčovým mediátorom. J. Neurosci. 33, 3434-3442. dva: 10.1523 / JNEUROSCI.4881-12.2013
Pleim, E., Matochik, J., Barfield, R., a Auerbach, S. (1990). Korelácia uvoľňovania dopamínu v jadre s mužským sexuálnym správaním u potkanov. Brain Res. 524, 160–163. doi: 10.1016/0006-8993(90)90507-8
Robbins, TW a Arnsten, AFT (2009). Neuropsychofarmakológia frontu-výkonnej funkcie: monoaminergná modulácia. Ann. Neurosci. 32, 267 – 287. doi: 10.1146 / annurev.neuro.051508.135535
Sabariego, M., Morón, I., Gómez, MJ, Donaire, R., Tobeña, A., Fernández-Teruel, A., a kol. (2013). Stimulačná strata a expresia hippocampálnych génov u potkanov s invazívnym rímskym vysoko (RHA-I) a rímskym nízkym (RLA-I). Behave. Brain Res. 257, 62 – 70. doi: 10.1016 / j.bbr.2013.09.025
Sachs, BD (2000). Kontextové prístupy k fyziológii a klasifikácii erektilnej funkcie, erektilnej dysfunkcie a sexuálneho vzrušenia. Neurosci. Biobehav. Rev. 24, 541–560. doi: 10.1016/S0149-7634(00)00022-1
Sachs, BD, Akasofu, K., Citro, JK, Daniels, SB a Natoli, JH (1994). Bezkontaktná stimulácia z estrusových samíc vyvoláva erekciu penisu u potkanov. Physiol. Behave. 55, 1073–1079. doi: 10.1016/0031-9384(94)90390-5
Sachs, BD a Barfield, RJ (1976). Funkčná analýza mužského kopulačného správania u potkanov. Prísl. Stud. Behave. 7, 91–154. doi: 10.1016/s0065-3454(08)60166-7
Salamone, JD, Pardo, M., Yohn, SE, López-Cruz, L., San Miguel, N., a Correa, M. (2016). Mezolimbický dopamín a regulácia motivovaného správania. Akt. Top. Behave. Neurosci. 27, 231–257. doi: 10.1007/7854_2015_383
Sanna, F., Corda, MG, Melis, MR, Piludu, MA, Giorgi, O. a Argiolas, A. (2014a). Samce rímskych potkanov s vysokým a nízkym vyhýbaním sa vykazujú rôzne vzorce kopulačného správania: porovnanie s potkanmi sprague dawley. Physiol. Behave. 127, 27 – 36. doi: 10.1016 / j.physbeh.2014.01.002
Sanna, F., Piludu, MA, Corda, MG, Argiolas, A., Giorgi, O. a Melis, MR (2014b). Dopamín sa podieľa na rôznych vzorcoch kopulačného správania sa rímskych potkanov s vysokým a nízkym únikom: štúdie s apomorfínom a haloperidolom. Pharmacol. Biochem. Behave. 124, 211-219. dva: 10.1016 / j.pbb.2014.06.012
Sanna, F., Piludu, MA, Corda, MG, Melis, MR, Giorgi, O. a Argiolas, A. (2015). Účasť dopamínu na rozdieloch v sexuálnom správaní medzi rímskymi potkanmi s vysokým a nízkym únikom: štúdia intracerebrálnej mikrodialýzy. Behave. Brain Res. 281, 177 – 186. doi: 10.1016 / j.bbr.2014.12.009
Scatton, B., D'Angio, M., Driscoll, P., a Serrano, A. (1988). in vivo voltametrické štúdium odpovede mezokortikálnych a mezoaccumbens dopaminergných neurónov na environmentálne stimuly u kmeňov potkanov s rôznymi úrovňami emotivity. Ann. NY Acad. Sci. 537, 124–137. doi: 10.1111/j.1749-6632.1988.tb42101.x
Steimer, T., a Driscoll, P. (2003). Odlišné stresové reakcie a zvládanie štýlov zvládania psychogeneticky vybraných rímskych potkanov s vysokou (RHA) a nízkou (RLA) hladinou: behaviorálne, neuroendokrinné a vývojové aspekty. Stres 6, 87-100. dva: 10.1080 / 1025389031000111320
Succu, S., Sanna, F., Argiolas, A., a Melis, MR (2011). Oxytocín vstreknutý do hipokampálneho ventrálneho subikula indukuje erekciu penisu u samcov potkanov zvýšením glutamatergickej neurotransmisie vo ventrálnej tegmentálnej oblasti. Neuropharmacology 61, 181-188. dva: 10.1016 / j.neuropharm.2011.03.026
Succu, S., Sanna, F., Cocco, C., Melis, T., Boi, A., Ferri, GL, a kol. (2008). Oxytocín vyvoláva erekciu penisu, keď sa injikuje do ventrálnej tegmentálnej oblasti samcov potkanov: úloha oxidu dusnatého a cyklický GMP. Eur. J. Neurosci. 28, 813-821. dva: 10.1111 / j.1460-9568.2008.06385.x
Succu, S., Sanna, F., Melis, T., Boi, A., Argiolas, A. a Melis, MR (2007). Stimulácia dopamínových receptorov v paraventrikulárnom jadre hypotalamu samcov potkanov vyvoláva erekciu penisu a zvyšuje mimobunkový dopamín v jadre accumbens: zapojenie centrálneho oxytocínu. Neuropharmacology 52, 1034-1043. dva: 10.1016 / j.neuropharm.2006.10.019
Thierry, AM, Tassin, JP, Blanc, G. a Glowinski, J. (1976). Selektívna aktivácia mezokortikálneho DA systému stresom. príroda 263, 242 – 244. doi: 10.1038 / 263242a0
Wenkstern, D., Pfaus, J., a Fibiger, H. (1993). Prenos dopamínu sa zvyšuje v nucleus accumbens samcov potkanov počas ich prvého vystavenia pohlavne vnímavým samiciam potkanov. Brain Res. 618, 41–46. doi: 10.1016/0006-8993(93)90426-N
Westernik, BHC, De Boer, P., De Vries, JB, a Long, S. (1998). Antipsychotické lieky vyvolávajú podobné účinky na uvoľňovanie dopamínu a noradrenalínu v strednom prefrontálnom kortexe mozgu potkana. Eur. J. Pharmacol. 361, 27–33. doi: 10.1016/s0014-2999(98)00711-0
Willig, F., M'Harzi, M., Bardelay, C., Viet, D., a Delacour, J. (1991). Rímske kmene ako psychogénny model na štúdium pracovnej pamäte: behaviorálne a biochemické údaje. Pharmacol. Biochem. Behave. 40, 7–16. doi: 10.1016/0091-3057(91)90313-q
Kľúčové slová: sexuálne správanie, dopamín, noradrenalín, mPFC, mikrodialýza, potkany RHA a RLA
Citácia: Sanna F, Bratzu J, Piludu MA, Corda MG, Melis MR, Giorgi O a Argiolas A (2017) dopamín, noradrenalín a rozdiely v sexuálnom správaní medzi rímskymi vysokými a nízkymi únikmi u potkanov: štúdia mikrodialýzy v strednom prefrontálnom kortexe , Predná. Behave. Neurosci. 11: 108. doi: 10.3389 / fnbeh.2017.00108
Prijaté: 22 December 2016; Prijaté: 19 Môže 2017;
Pridané: 07 June 2017.
