Predná. Behave. Neurosci., 23 March 2016 | http://dx.doi.org/10.3389/fnbeh.2016.00054

 Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2,
Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2, 

- 1Translačný výskumný ústav a Inštitút pre zdravie a biomedicínske inovácie, Queenslandská technologická univerzita, Brisbane, QLD, Austrália
- 2Škola biomedicínskych vied, Univerzita Queensland, Brisbane, QLD, Austrália
Moderná strava sa stala vysoko osladenou, čo viedlo k bezprecedentnej spotrebe cukru, najmä u dospievajúcich. Zatiaľ čo je známe, že chronický dlhodobý príjem cukru prispieva k rozvoju metabolických porúch vrátane obezity a diabetu typu II, je málo známe, čo sa týka priamych dôsledkov dlhodobej konzumácie cukru v mozgu. BKeďže sacharidy môžu spôsobiť uvoľňovanie dopamínu v nucleus accumbens (NAc) podobne ako drogy zneužívania, skúmali sme zmeny v morfológii neurónov v tejto oblasti mozgu po krátkom (4 týždňoch) a dlhodobom (12 týždňoch) ako je spotreba sacharózy s použitím intermitentnej paralelnej voľby dvoch fľašiek. Použili sme farbenie Golgiho-Cox na impregnáciu stredne štipľavých neurónov (MSN) z jadra a plášťa NAc krátkodobých a dlhodobých potkanov požívajúcich sacharózu a porovnali ich s kontrolou vody zodpovedajúcou veku. Ukazuje sa, že dlhodobá konzumácia sacharózy podobnej záchvatu výrazne znížila celkovú dendritickú dĺžku MSN škrupín NAc v porovnaní s kontrolnými potkanmi zodpovedajúcimi veku. Zistili sme tiež, že reštrukturalizácia týchto neurónov je výsledkom predovšetkým zhoršenej dendritickej zložitosti. Naopak, pozorovali sme zvýšené hustoty chrbtice v distálnych pobočkových poradiach krvných doštičiek NAc z dlhodobých potkanov konzumujúcich sacharózu. V kombinácii tieto výsledky poukazujú na neuronálne účinky dlhotrvajúceho príjmu sacharózy podobného záchvatu na morfológiu MSN škrupiny NAc.
úvod
Počas posledných rokov 40 došlo k zdokumentovanému nárastu spotreby nápojov sladených cukrom a potravín obsahujúcich pridaných cukrov (Nielsen a kol., 2002; Popkin, 2010; Ng a kol., 2012), pričom správy odhadujú, že až 75% všetkých potravín a nápojov obsahuje vysoké množstvá pridaných cukrov (Ford a Dietz, 2013; Bray a Popkin, 2014). Počas tohto obdobia sa tiež vyskytol súbežný nárast výskytu obezity a diabetu typu II, najmä u dospievajúcich (Arslanian, 2002; Reinehr, 2013; Dabelea a kol., 2014; Fryar a kol., 2014). Nedávne štúdie ukázali, že deti s nadváhou a obezitou často konzumujú vysoké množstvá pridaného cukru, avšak príspevok diét s vysokým obsahom cukru k zvýšenému výskytu detí s nadváhou a obezitou zostáva kontroverzný (Hu, 2013; Bray a Popkin, 2014; Bucher Della Torre a kol., 2015).
Zatiaľ čo rastúci počet dôkazov naznačuje, že konzumácia jedál s vysokým obsahom cukru môže čiastočne prispieť k zvýšeniu telesnej hmotnosti u detí a dospievajúcich (Malik a kol., 2010; Te Morenga a kol., 2013; Bray a Popkin, 2014) sa venovala menšia pozornosť nepriaznivým nemetabolickým dôsledkom vyplývajúcim z nadmerného príjmu cukru. Zaujímavé je, že niektoré bežné behaviorálne a psychologické vzorce sa často objavujú medzi podskupinou tých, ktorí nadmerne jedia a udržiavajú stravu s vysokým obsahom cukru. Najdôležitejšie sú vývoj porúch stravovania vrátane záchvatov, spojených so súčasným nástupom psychologických symptómov vrátane nedostatku motivácie a depresie Sheehan a Herman, 2015). Navyše, pretože jedinci, ktorí trpia záchvatmi, často vykazujú stratu kontroly a neschopnosť samo-obmedzovať príjem cukru, je pravdepodobné, že tieto správania vznikajú v dôsledku neurologických adaptácií v oblastiach mozgu, ktoré hodnotia hedonickú hodnotu veľmi chutného jedla (Saper a kol., 2002; Lutter a Nestler, 2009; Kenny, 2011). Toto zdôvodnenie podporuje aj dôkazy u ľudí, ktoré dokazujú, že cukor a sladkosť môžu spôsobiť craving, ktorý je podobný tomu, čo vyvolávajú návykové drogy, napríklad alkohol a nikotín (Volkow a kol., 2012).
Napriek tomu, že návykové vlastnosti cukru sú stále špekulatívne, tieto pozorovania v kombinácii so štúdiami preukazujúcimi prínos nadmerného príjmu cukru k zmenám obvodov odmeňovania a vývoju návykových správania a emocionálnych stavov na zvieracích modeloch (Avena a kol., 2008; Benton, 2010; Ventura a kol., 2014), zaručuje potrebu ďalšieho vyšetrovania. Predchádzajúce štúdie u hlodavcov ukázali, že prerušovaný prístup k sacharóze mení niektoré neurotransmitery v rámci mezolimbického systému, vrátane dopamínu, opioidov a acetylcholínu (pozri Avena a kol., 2008). Ukázalo sa, že závislosť konzumácie sacharózy uľahčuje uvoľňovanie dopamínu v nucleus accumbens (NAc), podobne ako drogy zneužívania (Avena a kol., 2008). Okrem toho sme preukázali, že dlhodobá spotreba sacharózy pomocou paradigmy výberu dvoch lámp 24 h s intermitentným prístupom (Simms a spol., 2008) moduluje expresiu nikotínového acetylcholínového receptora (nAChR) v NAc (Shariff a kol., V tlači). Zaujímavé je tiež, že zlúčeniny nAChR, o ktorých je známe, že modulujú dopamínovú a acetylcholínovú aktivitu v NAc, majú rôzne účinky na spotrebu sacharózy po krátkodobom a dlhodobom príjmeShariff a kol., V tlači).
Zatiaľ čo tieto štúdie preukázali podobnosti v behaviorálnych a neurochemických zmenách spôsobených prerušovaným prístupom k cukru a liekom zneužívania, nie je známe, či tieto účinky uľahčujú zmeny v neuronálnej morfológii v NAc. Tjeho kontrastuje s látkami zneužívania vrátane kokaínu, amfetamínu a nikotínu, ktoré produkujú dobre charakterizované zmeny v morfológii stredne štipľavých neurónov (MSN) v NAc vrátane zvýšenej hustoty chrbtice a zmenenej dendritickej zložitosti (Robinson a Kolb, 1999, 2004; Li et al., 2003; Crombag a kol., 2005). Pretože sme už predtým ukázali, že dlhodobá expozícia (týždeň 12) alkoholu a sacharóze pri použití intermitentnej paradigmy voľby dvoch fľašiek prináša diferencovanú odpoveď na farmakoterapeutické zákroky v porovnaní s krátkodobým príjmom (4 týždne; Steensland a kol., 2007; Shariff a kol., V tlači) sme hodnotili účinky krátkodobej a dlhodobej spotreby sacharózy na morfológiu MSN v NAc. Dospievajúcim krysám sme dovolili konzumovať sacharózu podobným spôsobom pre 4 (krátkodobé) alebo 12 (dlhodobé) týždne a potom analyzovali morfológiu NAc MSN z krátkodobých a dlhodobých potkanov konzumujúcich sacharózu a porovnali to s ktorí dostali prístup iba k vode. Naše výsledky ukazujú, že MSN z plášťa NAc sa menia po dlhodobej, ale nie krátkodobej spotrebe sacharózy, ktorá má zníženú dendritickú dĺžku, ale zvyšuje hustotu dendritického chrbtice. Navyše sme zistili, že morfológia MSN z jadra NAc zostala relatívne neporušená po krátkodobej a dlhodobej spotrebe sacharózy. Tieto výsledky poukazujú na priamy neurologický dôsledok dlhodobej spotreby sacharózy v podobnom záchvate. Okrem toho tieto údaje dokazujú potrebu ďalších štúdií zameraných na objasnenie molekulárnych a neurochemických zmien, ktoré sprevádzajú morfologickú reštrukturalizáciu MSN škrupín NAc indukovaných predĺženým, príliš sacharóznym príjmom.
Materiály a metódy
Vyhlásenie o etike
Všetky experimentálne postupy boli vykonané v súlade s austrálskym kódexom pre starostlivosť a používanie zvierat na vedecké účely, 8th Edition (Národná rada pre zdravotný a lekársky výskum, 2013). Protokoly boli schválené Výborom pre etiku zvierat na univerzite v Queenslande a Výborom pre etiku zvierat v Queenslandskej univerzite.
Zvieratá a bývanie
Päťtýždňové (dospievajúce) samce potkanov Wistar (kontrola: 176.4 ± 4.8 g, sacharóza: 178.3 ± 5.0 g) (ARC, WA, Austrália) boli individuálne umiestnené v odvetranom plexiskle s dvojitou úrovňou® klietky. Krysy sa aklimatizovali na jednotlivé podmienky umiestnenia, manipuláciu a cyklu 5 s obráteným osvetlením pred začiatkom pokusov. Všetky potkany boli umiestnené v klimatizovanom priestore 12-hr s obráteným svetlom / tmavým svetlom (zhasínajú pri izbe 9 am) so štandardným potkaním krmiva a vodou podľa chuti.
Intermitentný prístup Two-Bottle Choice Drinking Paradigm
Intermitentný prístup 5% sacharózy s dvoma fľaškami (Simms a spol., 2008) Múdry (1973), Všetky kvapaliny boli prezentované v plastových fľašiach s rozmermi 300 ml, ktoré obsahovali nápoje z nehrdzavejúcej ocele vložené cez dva priechodky v prednej časti klietky po začiatku cyklu tmavého osvetlenia. Hmotnosti každej fľaše boli zaznamenané pred prezentáciou fľaše. Súčasne boli predložené dve fľaše: jedna fľaša obsahujúca vodu; druhá fľaša obsahujúca 5% (hmotnosť / objem) sacharózy. Umiestnenie fľaše s obsahom sacharózy 5% (hmotn./objem) sa striedalo s každou expozíciou voči kontrole pre bočné preferencie. Fľaše boli odvážené 24 h potom, čo boli tekutiny prezentované a merania boli vykonané na najbližší 0.1 g. Hmotnosť každej krysy bola tiež meraná na výpočet gramov príjmu sacharózy na kilogram telesnej hmotnosti. V deň 1 doby pitia sa potkany (n = 6-9) dostali prístup do jednej fľaše 5% (hmotn./obj.) Sacharózy a jednej fľaše s vodou. Po 24 h bola fľaša na sacharózu nahradená druhou fľašou s vodou, ktorá bola k dispozícii pre nasledujúci 24 h. Tento vzor sa opakoval v stredu a piatok. Potkany mali neobmedzený prístup k vode vo všetkých ostatných dňoch. Závažná konzumácia sacharózy viedla k zvýšeniu celkového príjmu sacharózy (ml) v priebehu času (doplnkový obrázok 1) a boli sprevádzané stabilnými základnými hladinami pitia na základe telesnej hmotnosti [20 ± 5 g / kg 5% (w / v)] počas krátkodobých [4 týždňov (13 drinking session)] a dlhodobých [ ~ 12 týždňov (37 pitie)). Samostatná skupina kontrolných potkanov (n = 6-9) dostali prístup do vody v obidvoch fľašiach (tj bez sacharózy) za rovnakých podmienok opísaných vyššie. Priemerná telesná hmotnosť kontrolných potkanov a potkanov spotrebúvajúcich sacharózu na konci krátkodobej expozície bola 405.7 ± 40.8 g a 426.4 ± 31.2 g. Na konci dlhodobej expozície bola priemerná telesná hmotnosť kontrolných a sacharózových skupín 578.8 ± 53.4 g a 600.2 ± 45.2 g.
Golgi-Cox farbenie
Po poslednej konzumácii alkoholu sa potkany preniesli zo zariadenia na zvieranie, aby umožnili spracovanie vzoriek mozgu v histologickom zariadení na Fakulte biomedicínskych vied, University of Queensland (St Lucia, Austrália). Prijali sa všetky schválené opatrenia na zníženie stresu počas transportu, po ktorých sa krysy nechali cez noc späť. Nasledujúci deň boli potkany usmrtené predávkovaním pentobarbitalom sodným (60-80 mg / kg, ip Vetcare, Brisbane, Austrália) a intracardiálne perfundované s umelou mozgovou miechou 300 ml, ktorá obsahovala (v mM): 130 NaCl, 3 KCI, 26 NaHCO3, 1.25 NaH2PO4, 5 MgCl2, 1 CaCl2, a 10 D-glukóza. Každé zviera sa potom dekapitovalo a mozog sa odstránil a inkuboval v tme v roztoku Golgi-Cox, ktorý obsahoval 5% dvojchróman draselný, 5% chlorid draselný a 5% chlorid ortutnatý (všetky chemikálie od spoločnosti Sigma-Aldrich) pred usmrtením, ako bolo opísané skôr (Rutledge a kol., 1969). Golgi-Cox inkubácia škvŕn a metódy dodatočného spracovania boli modifikované z Ranjan a Mallick (2010), Mozgy z krátkodobých zvierat, ktoré konzumujú sacharózu, boli inkubované 6 dni pri 37 ° C, zatiaľ čo mozgy z dlhodobých zvierat náročných na sacharózu boli inkubované za 10 dni s jednou zmenou na čerstvý roztok Golgiho-Cox po 4 dňoch inkubácie.
Po inkubácii boli rezy 300 μm koronálne rezané pomocou vibračnej mikrotómy Zeiss Hyrax V50 (Carl Zeiss, Nemecko). Plátky boli potom umiestnené postupne do platní 24-well plnených 30% (hmotn./obj.) Sacharózy v fyziologickom roztoku pufrovanom fosforečnanom 0.1M a spracované tak, ako je uvedené v (Ranjan a Mallick, 2010). Stručne, sekcie boli dehydratované v 50% etanole pre 5 min, potom boli umiestnené do 0.1 M NH4Roztok OH pre 30 min, opláchnutý dvakrát destilovanou vodou na 5 min a umiestnený v Fujihunt film fixeri (Fujifilm, Singapur) pre 30 min v tme. Plátky boli potom opláchnuté dvakrát v destilovanej vode za min. 2 min a dehydratované v 70, 90, 95 a 100% etanole dvakrát pre každý 5 min. Rezy boli potom vyčistené v roztoku CXA (1: 1: 1 chloroform: xylén: alkohol) pre 10 min a namontované v DPX (Sigma-Aldrich) na sklíčkach Superfrost Plus (Menzel-Glaser, Lomb Scientific, Austrália) (Menzel-Glaser, Nemecko). Sklíčka boli ponechané v tme, aby sa usušila pri laboratórnej teplote cez noc.
Výber a sledovanie neurónov v rámci Nucleus Accumbens
Koronálne plátky medzi bregma + 2.8 a + 1.7 sa skúmali pre MSN v jadre a plášti NAc s využitím bočnej komory a prednej komisie ako orientačných bodov pomocou atlasu mozgu potkanaPaxinos a Watson, 2007) (Obrázok 1). Funkcia obrysu v systéme Neurolucida 7 (MBF Bioscience, VT, USA) bola použitá na vymedzenie jadra NAc a povlaku NAc v každom plátku (obrázok 2). Medzi neurónmi 2 a 9 na región na zviera bola sledovaná parametre dendritických dĺžok s použitím 63x cieľa alebo hustoty chrbtice (hlásené ako hroty na 100 μm) pomocou objektívu 100x na Zeiss Axioskop II (Carl Zeiss, Nemecko) s použitím automatizovaného xyz štádiu poháňaného Neurolucidou® Softvér 7 (MBF Biosciences, VT, USA). Všetky sledovanie sa uskutočnilo slepým spôsobom s ohľadom na liečbu. Morfologické parametre impregnovaných neurónov Golgi-Cox boli analyzované podobným spôsobom ako predchádzajúce správy (Klenowski a kol., 2015).
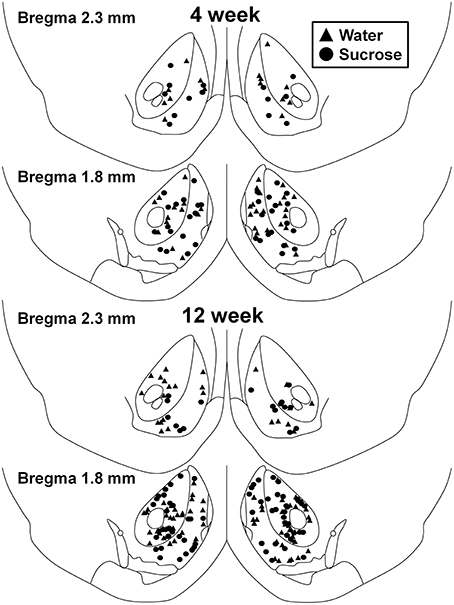
Obrázok 1. Mapa znázorňujúca umiestnenie stredne štipľavých neurónov odobratých z jadra nucleus accumbens jadra a škrupiny 4 a 12 týždeň konzumujúcich sacharózu potkanov a kontrolných vekových skupín, V horných dvoch paneloch sú zobrazené polohy neurónov odobratých z nucleus accumbens jadra a škrupiny týždennej kontroly 4 (trojuholníky) a zo sacharózy (kruhy) zvierat. Spodné dva panely ukazujú pozície neurónov, ktoré boli odobraté z týždennej kontroly 12 (trojuholníky) a zo sacharózy (kruhy).
Štatistická analýza
Stredná a štandardná chyba priemeru (SEM) bola vypočítaná pre každú sadu údajov so zvieraťom ako n, pričom sa použili priemerné morfometrické údaje zo všetkých jadrových alebo shellových NAn MSN (n = 7 pre shell NAc a n = 6 pre týždeň NAn core 4, n = 9 pre 12-týždňové skupiny). Ak je to uvedené, nespárovaný dvojstranný študent ttesty alebo obojsmerné ANOVA s post-testmi Bonferroni boli vykonané pre všetky analýzy zahŕňajúce porovnanie skupinových prostriedkov pomocou GraphPad Prism verzie 6.02 (GraphPad Software, San Diego, CA). Štatistická významnosť bola prijatá na P <0.05. Všetky údaje v časti s výsledkami sú uvedené ako priemer ± SEM. Percentuálne zmeny sa počítajú ako relatívne k kontrolnej hodnote.
výsledky
Stredne štipľavé neuróny z jadra Nucleus Accumbens Shell majú zníženú dendritickú dĺžku, zníženú dendritickú komplexnosť, ale zvýšenú priemernú hustotu chrbtice v distálnych pobočkách po dlhšej ale nie krátkodobej spotrebe sacharózy
Po krátkodobej spotrebe sacharózy v týždni (4 týždňov) neboli žiadne významné rozdiely v morfometrických parametroch NAn shell MSN (tabuľka 1). Neboli ani významné rozdiely medzi krátkodobou spotrebou sacharózy a MSN v kontrolných vodách NAc shell v analýzach týkajúcich sa odstredivého poľa. Konkrétne dendritické segmenty na poradí pobočky (P = 0.4111), priemerná dendritická dĺžka na poradie odbočiek (P = 0.5581) a stredná hustota chrbtice podľa poradia odbočiek (P = 0.2977, obojsmerné ANOVA) sa medzi skupinami výrazne nelíšili. Mapová mapa s približnými pozíciami vybraných neurónov je uvedená na obrázku 1.
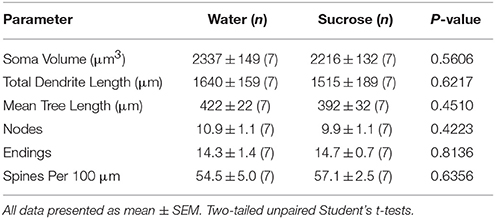
Tabuľka 1. Všeobecné morfologické parametre stredne štipľavých neurónov z jadrovo-jadrovej škrupiny krátkodobých potkanov požívajúcich sacharózu a kontroly v závislosti od veku.
Po dlhodobých (12 týždňoch) spotrebe sacharózy sa celková dĺžka dendritického kmeňa MSN na povrchu kmeňa NAc znížila o 21% v porovnaní s kontrolami spotrebujúcimi vodu (voda: 1827 ± 148 μm, n = 9; Sacharóza 1449 ± 78 μm, n = 9, *P = 0.0384, dvojstranný nepárový Student t-test, obrázok 2, Tabuľka 2). Porovnanie priemerného počtu dendritických bifurkácií (uzlov) a dendritických koncov medzi skupinou s vodou a so sacharózou odhalilo zníženú (hoci nie významnú) úroveň dendritickej zložitosti v MSN na plášti NAc (uzly: voda 12.9 ± 1.4 n = 9, sacharóza 10.1 ± 0.8 n = 9, P = 0.0879; koncovky: voda 17.9 ± 1.4 n = 9, sacharóza 14.8 ± 0.7 n = 9, P = 0.0657, dvojstranný nepárový Student t-test, tabuľka 2). V soma objemu nedošlo k žiadnej zmene (P = 0.9400), stredná dendritická dĺžka stromu (P = 0.1646) alebo celková hustota chrbtice (P = 0.3662) v MSN shell NAC z dlhodobých potkanov konzumujúcich sacharózu v porovnaní s kontrolou vody. Tieto morfometrické parametre sú uvedené v tabuľke 2.
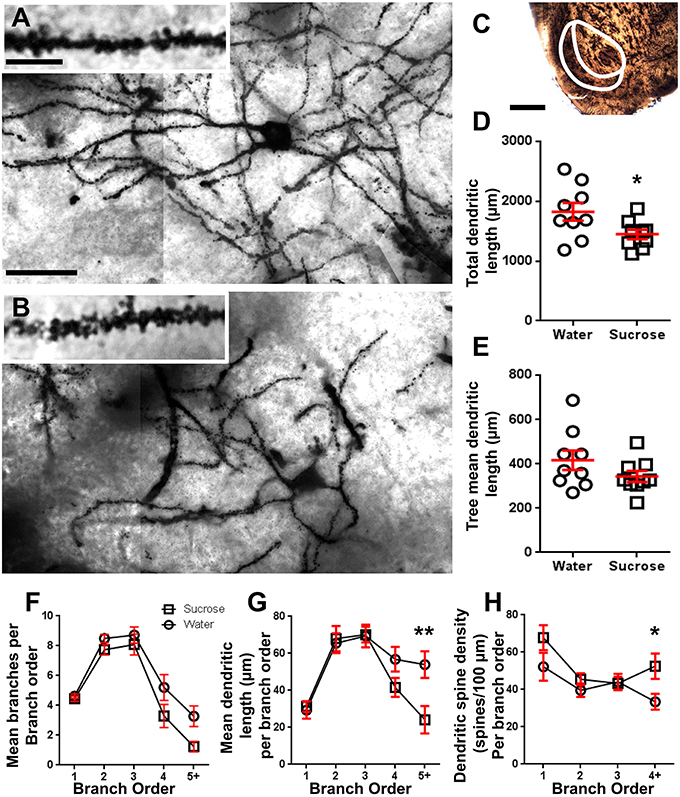 Obrázok 2. Znížená dendritická dĺžka klinčeka a zvýšená hustota dendritického chrbtice stredných štipcovitých neurónov (MSN) z jadra accumbens (NAc) dlhodobých potkanov ošetrených sacharózou v porovnaní s kontrolnými potkanmi. (A, B) ukazujú reprezentácie kontrolného (horného) a dlhodobého (12 týždeň) sacharózy (spodné) ošetreného jasného poľa z-stack mozaiky Golgi-impregnovaných MSN z plášťa NAc (zväčšenie 63x). Vložka (A, B) ukazuje kontrolné a dlhodobé obrazy jasného poľa ošetrené sacharózou z dendritov a dendritických trhlín impregnovaných Golgi z shell NAc (zväčšenie 100x). (C) ukazuje anatomické oblasti, z ktorých sa v tejto štúdii odobrali vzorky MSN. (D) znázorňuje rozptylový graf poklesu celkového dendritického arboru MSN (priemer ± SEM) z plášťa NAc u dlhodobých zvierat so sacharózou (štvorčeky) v porovnaní s kontrolami (kruhy), nespálených študentov t-test, *P <0.05, n = 9; ovládanie a n = 9; 12 týždeň sacharózy. (E) ukazuje priemer dendritického stromu MSN (priemer ± SEM) zo škrupiny NAc v dlhodobých zvieratách so sacharózou (štvorčeky) v porovnaní s kontrolami (kruhy), nezapojenými študentmi t-test, P > 0.05, n = 9; ovládanie a n = 9; 12 týždeň sacharózy. Analýza pobočkovej objednávky (priemer ± SEM) dendritického segmentu podľa pobočky (F), priemerná dendritická dĺžka na poradie odbočiek (G) a dendritickú hustotu chrbtice podľa poradia odbočiek (H), Dlhodobá spotreba sacharózy znížila dendritickú dĺžku pri vzdialených odbočkách (5 +) a zvýšila hustotu dendritického chrbtice v distálnych odbočkách (4 +) v porovnaní s kontrolami (G, H), obojsmerné ANOVA s post-testmi Bonferroni, *P <0.05, **P <0.01, n = 9; ovládanie a n = 9; dlhodobej sacharózy. Mierka: (A, B) = 20 μm; vložka (A, B) = 10 μm; (C) = 1 mm.
Obrázok 2. Znížená dendritická dĺžka klinčeka a zvýšená hustota dendritického chrbtice stredných štipcovitých neurónov (MSN) z jadra accumbens (NAc) dlhodobých potkanov ošetrených sacharózou v porovnaní s kontrolnými potkanmi. (A, B) ukazujú reprezentácie kontrolného (horného) a dlhodobého (12 týždeň) sacharózy (spodné) ošetreného jasného poľa z-stack mozaiky Golgi-impregnovaných MSN z plášťa NAc (zväčšenie 63x). Vložka (A, B) ukazuje kontrolné a dlhodobé obrazy jasného poľa ošetrené sacharózou z dendritov a dendritických trhlín impregnovaných Golgi z shell NAc (zväčšenie 100x). (C) ukazuje anatomické oblasti, z ktorých sa v tejto štúdii odobrali vzorky MSN. (D) znázorňuje rozptylový graf poklesu celkového dendritického arboru MSN (priemer ± SEM) z plášťa NAc u dlhodobých zvierat so sacharózou (štvorčeky) v porovnaní s kontrolami (kruhy), nespálených študentov t-test, *P <0.05, n = 9; ovládanie a n = 9; 12 týždeň sacharózy. (E) ukazuje priemer dendritického stromu MSN (priemer ± SEM) zo škrupiny NAc v dlhodobých zvieratách so sacharózou (štvorčeky) v porovnaní s kontrolami (kruhy), nezapojenými študentmi t-test, P > 0.05, n = 9; ovládanie a n = 9; 12 týždeň sacharózy. Analýza pobočkovej objednávky (priemer ± SEM) dendritického segmentu podľa pobočky (F), priemerná dendritická dĺžka na poradie odbočiek (G) a dendritickú hustotu chrbtice podľa poradia odbočiek (H), Dlhodobá spotreba sacharózy znížila dendritickú dĺžku pri vzdialených odbočkách (5 +) a zvýšila hustotu dendritického chrbtice v distálnych odbočkách (4 +) v porovnaní s kontrolami (G, H), obojsmerné ANOVA s post-testmi Bonferroni, *P <0.05, **P <0.01, n = 9; ovládanie a n = 9; dlhodobej sacharózy. Mierka: (A, B) = 20 μm; vložka (A, B) = 10 μm; (C) = 1 mm.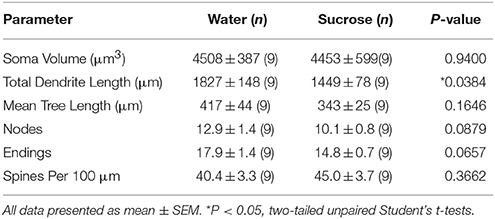
Tabuľka 2. Všeobecné morfologické parametre stredne štipľavých neurónov z jadrovacej škrupiny dlhodobých potkanov požívajúcich sacharózu a kontroly v závislosti od veku.
Po charakterizácii všeobecnej dendritickej morfológie dlhodobých sacharózou spotrebúvajúcich NAc shell MSN sme analyzovali dendritické arborizácie a hustoty chrbtice s ohľadom na ich charakteristiky pobočiek. Naše komplexné posúdenie dendritických stromov kvantifikovalo počet dendritických segmentov na poradie vetvenia, priemernú dĺžku dendritických segmentov podľa poradia odbočiek a priemernú hustotu chrbtice podľa poradia krvných skupín NAn škrupín MSN vodnej kontroly a dlhodobých potkanov konzumujúcich sacharózu. Súhrn údajov a analýz odvetvovej objednávky je uvedený v tabuľke 3.
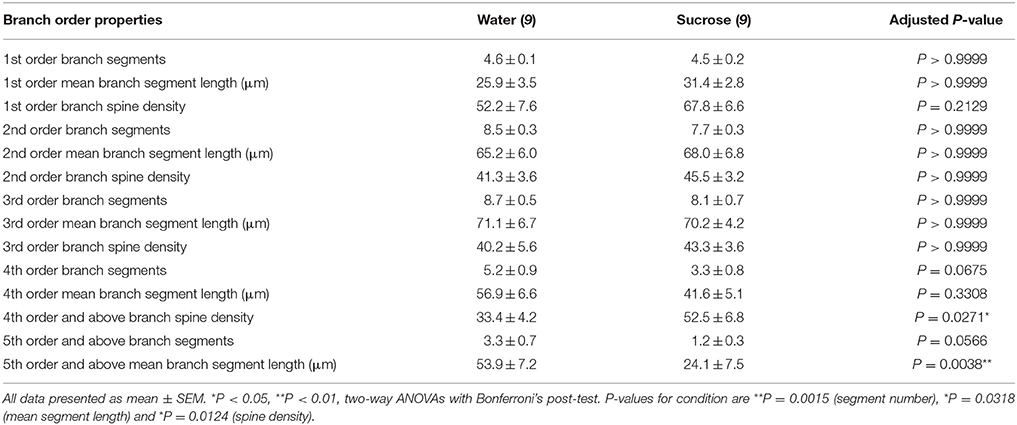 Tabuľka 3. Poruchové charakteristiky stredne štipľavých neurónov z dlhodobých potkanov s obsahom sacharózy a vody.
Tabuľka 3. Poruchové charakteristiky stredne štipľavých neurónov z dlhodobých potkanov s obsahom sacharózy a vody.Priemerný počet segmentov dendritických vetiev v poradí podľa MSNs v kmeňoch NAc bol významne znížený u potkanov s dlhodobou konzumáciou sacharózy v porovnaní s kontrolou vody (**)P = 0.0015, obojsmerná ANOVA). Post-testy Bonferroni odhalili trend smerujúci k zníženiu počtu segmentov v 4th (voda: 5.2 ± 0.9, n = 9; Sacharóza 3.3 ± 0.8, n = 9, P = 0.0675, obrázok 2F, Tabuľka 3) a príkazy 5 a vyššie uvedené objednávky (voda: 3.3 ± 0.7, n = 9; Sacharóza 1.2 ± 0.3, n = 9, P = 0.0566, obrázok 2F, Tabuľka 3). Stredná dendritická dĺžka segmentu podľa poradia krvných skupín NAn shell shellov bola tiež významne znížená u potkanov s dlhodobou konzumáciou sacharózy v porovnaní s kontrolou vody (*P = 0.0444, obojsmerná ANOVA). Post-testy Bonferroni preukázali zníženie 55% na 5th vetvy a ďalšie (Voda: 53.9 ± 7.2 μm, n = 9; Sacharóza 24.1 ± 7.5 μm, n = 9, **P = 0.0038, obrázok 2G, Tabuľka 3).
Analýza pobočiek ukázala významný nárast denzity dendritickej chrbtice nosníc MSN dlhodobých potkanov s obsahom sacharózy v porovnaní s kontrolnými zvieratami (*P = 0.0124, obojsmerná ANOVA). Post-testy Bonferroni preukázali zvýšenie hustoty chrbtice 57% v distálnych vedeniach 4th a ďalej (voda: 33.4 ± 4.2, n = 9; Sacharóza 52.5 ± 6.8, n = 9, P = 0.0271 *, vložka z obrázkov 2, B, H, Tabuľka 3). Reprezentatívne obrazy celkovej architektúry MSN a hustoty distálnej chrbtice (vložka) sú znázornené na obrázkoch 2, B.
Súhrnne tieto výsledky naznačujú, že krátkodobá spotreba sacharózy má malý vplyv na morfologické parametre MSN v plášti NAc. Avšak po dlhšej konzumácii dochádza k výraznému zníženiu dĺžky a zložitosti neurónového krovia, najmä v distálnych dendritických vetvách. Súčasné zvyšovanie hustoty distálnej chrbtice je tiež zrejmé z MSN s dlhými potkanmi, ktoré konzumujú sacharózu.
Stredne štipľavé neuróny z jadra Nucleus Accumbens majú zníženú zložitosť vetvenia po dlhej, ale nie krátkodobej spotrebe sacharózy
Po krátkodobej spotrebe sacharózy nedošlo k významným rozdielom v morfometrických parametroch NAc core (tab 4). Medzi analýzami týkajúcimi sa odstredivých odbočiek nebol ani významný rozdiel medzi konzumáciou sacharózy 4 v týždni a MSN v jadre kontroly vody. Konkrétne dendritické segmenty na poradí pobočky (P = 0.7717), priemerná dendritická dĺžka na poradie odbočiek (P = 0.2096) a stredná hustota chrbtice podľa poradia pobočiek (P = 0.3521, obojsmerné ANOVA) sa medzi skupinami nelíšili.
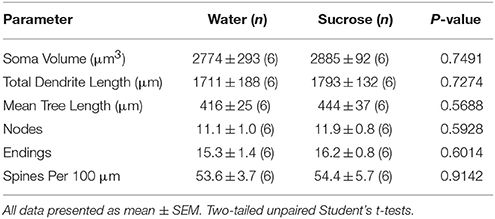 Tabuľka 4. Všeobecné morfologické parametre stredne štipľavých neurónov z nucleus accumbens jadra krátkodobých potkanov požívajúcich sacharózu a kontroly v závislosti od veku.
Tabuľka 4. Všeobecné morfologické parametre stredne štipľavých neurónov z nucleus accumbens jadra krátkodobých potkanov požívajúcich sacharózu a kontroly v závislosti od veku.Predĺžená spotreba sacharózy tiež nemala významné morfometrické parametre MSC na základe NAc (tab 5). Priemerný počet dendritických segmentov vetvenia podľa poradia MSN jadra NAc bol významne znížený u potkanov s dlhodobou konzumáciou sacharózy v porovnaní s kontrolou vody (*P = 0.0416, obojsmerná ANOVA), avšak žiadne významné rozdiely v priemernej dendritickej dĺžke na jedno usporiadanie vetvy (P = 0.0995) a stredná hustota chrbtice podľa poradia odbočiek (P = 0.4888, dvojcestné ANOVA) medzi MSN v jadre NAc dlhodobých potkanov konzumujúcich sacharózu v porovnaní s kontrolou vody. Spoločne, naše údaje ukazujú, že jadro NAc nie je tak citlivé na dlhodobú spotrebu sacharózy v porovnaní s MSN z regiónu NAc shell.
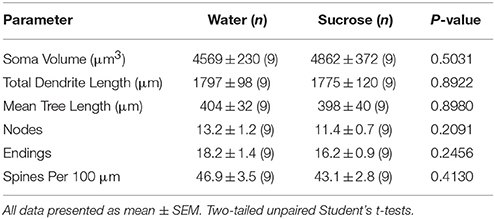 Tabuľka 5. Všeobecné morfologické parametre stredne štipľavých neurónov z nucleus accumbens jadra dlhodobých potkanov konzumujúcich sacharózu a kontroly veku zodpovedajúce veku.
Tabuľka 5. Všeobecné morfologické parametre stredne štipľavých neurónov z nucleus accumbens jadra dlhodobých potkanov konzumujúcich sacharózu a kontroly veku zodpovedajúce veku.Diskusia
Zvýšená dostupnosť vysoko osladených potravín v západnej strave nielenže prispela k zvýšenému výskytu a hospodárskej záťaži obezity a diabetu typu II, ale tiež viedla k nástupu porúch príjmu potravy, ako je prehnané jedenie (Swanson a kol., 2011; Kessler a kol., 2013; Davis, 2015). Hoci návykové vlastnosti cukrov vrátane fruktózy a sacharózy zostávajú špekulatívne, existuje výrazná podobnosť v behaviorálnych a neurálnych korelátoch, ktoré sa prejavujú v dôsledku nadmerného stravovania a dlhodobého užívania drog (Avena a kol., 2008, 2011). Cukor navyše aktivuje mozgové obvody odmien podobným spôsobom ako lieky proti zneužívaniu (Volkow a kol., 2012) a výsledky z výskumov na ľuďoch naznačujú, že cukor a sladkosť môžu vyvolať chute, ktoré sú porovnateľné v porovnaní s rozsahmi indukovanými návykovými látkami, ako je alkohol a nikotín (Volkow a kol., 2012). Preto sme použili model spotreby šarže-sacharózy u potkanov na určenie účinkov krátkodobých (4 týždňov) a dlhodobých (12 týždňov) spotreby sacharózy na neuronálnu morfológiu MSN v NAc, kľúčovej zložke prekrývajúcej sa odmeny obvodov ktorá je modulovaná cukrom a návykovými látkami. Ukazujeme, že MSN z pečene NAc chronických dlhodobých potkanov konzumujúcich sacharózu významne znížili dendritickú dĺžku a zložitosť, ale zvýšili hustotu dendritického chrbtice. Dlhodobá spotreba sacharózy nemala žiadny vplyv na morfológiu MSN z jadra NAc, zatiaľ čo krátkodobá spotreba sacharózy tiež nemala významný vplyv na morfológiu MSN z jadra alebo škrupiny NAc. Tieto výsledky ukazujú nielen priamy účinok predĺženého príjmu sacharózy podobného bláhovitosti na neuronálnu morfológiu MSN škrupín NAc, ale tiež poukazujú na potenciálne škodlivé dôsledky dlhotrvajúcej konzumácie diét s vysokým obsahom cukru.
NAc, ktorá tvorí súčasť ventrálneho striatu, pozostáva predovšetkým z MSN, ktoré sú morfologicky charakterizované ako stredne veľké neuróny s rozsiahlou dendritickou arborizáciou a vysokou hustotou chrbtice (Kemp a Powell, 1971; Graveland a DiFiglia, 1985; Rafols a kol., 1989; Kawaguchi a kol., 1990). Glutamatergické a dopaminergné neuróny sú dva primárne aferentné vstupy do NAc, primárne kontaktujúce dendritické šachty a trhliny MSN (Groves, 1980; Kaiya a Namba, 1981; Groves a kol., 1994). Konkrétne NAc shell a jadro dostanú glutamatergický vstup z funkčne odlišných kortikálnych oblastí (Brog a kol., 1993). NAc škrupina je tiež inervovaná excitačnými aferentmi z subkortikálnych oblastí, ako je hipokampus, talamus a bazolaterálna amygdala (Brog a kol., 1993; Wright a Groenewegen, 1995). Predchádzajúce štúdie preukázali, že tieto glutamatergické vstupy zohrávajú kľúčovú úlohu v motivácii a ciele orientovanom správaní, ako sú potraviny a odmeňovanie (Maldonado-Irizarry a kol., 1995; Kelley a Swanson, 1997; Reynolds a Berridge, 2003; Richard a Berridge, 2011). Druhý prevládajúci vstup na NAc MSNs je z dopaminergných aferentov, ktoré vychádzajú z ventrálnej tegmentálnej oblasti (Lindvall a Björklund, 1978; Veening a kol., 1980; Kalivas a Miller, 1984). Zaujímavé je, že predchádzajúce štúdie, ktoré používajú podobné modely prerušovaného prístupu k cukru, ukázali, že výsledná konzumácia podobná záchvatu vedie k zvýšeniu extracelulárneho dopamínu v NAc podobne (aj keď v menšej miere) (Rada et al., 2005; Avena a kol., 2006) a môže modulovať expresiu dopamínového receptora (Colantuoni a kol., 2001, 2002) v jadre a škrupine NAc. Zaujímavé je, že závislosť konzumácie sacharózy spôsobuje eskaláciu príjmu v priebehu času podobne ako samotná aplikácia zneužívajúcich liekov, ako je kokaín a hrdinovian (Ahmed a Koob, 1998; Ahmed a kol., 2000, 2003), ktorá je spojená s vývojom stavu "návykového typu".
Naša analýza morfometrie odbočiek ukazuje, že celkové zníženie dendritickej dĺžky MSN škrupín NAc spôsobené dlhodobým príjmom sacharózy je výsledkom predovšetkým znižovania zložitosti distálnych odbočiek. Zaznamenali sme znížené distálne rozvetvenie (4th a 5th poradie a vyššie rozdelenie) a výrazne znížili strednú dĺžku v poradí 5 a nad dendritmi, v kombinácii so zvýšenou hustotou chrbtice v týchto odbočkách. Bežný faktor, ktorý pravdepodobne ovplyvní tento typ dendritickej reštrukturalizácie, zahŕňa zmeny v synaptickej konektivite a / alebo funkcii (Russo a kol., 2010). Predchádzajúce štúdie ukázali, že glutamatergické synapsie na MSN sú tvorené primárne na tŕňoch, najmä pri distálnych dendritoch (Groenewegen a spol., 1999). Navyše ko-lokalizácia dopamínových a glutamatergických vstupov z prefrontálnej kôry (Sesack a Pickel, 1992), bájna morská príšera (Totterdell a Smith, 1989; Sesack a Pickel, 1990), a amygdala (Johnson a kol., 1994) boli pozorované na dendritických chvostoch MSN. Tieto pozorovania v kombinácii so zvýšenou hustotou chrbtice po dlhodobej spotrebe sacharózy v našej štúdii podporujú tvorbu zvýšených excitačných vstupov. Preto vzniká možnosť, že pretrvávajúce účinky spôsobené dlhotrvajúcim príjmom sacharózy podobného záchvatu môžu uľahčiť zvýšenú excitačnú synaptickú aktivitu u distálnych dendritov MSN v plášti NAc. V dôsledku toho môže dôjsť k redukcii a / alebo vytiahnutiu distálnych dendritov cez synaptický homeostatický mechanizmus (Reissner a Kalivas, 2010), zostáva sa to však určiť.
Je zaujímavé poznamenať, že Crombag a jeho kolegovia ukázali, že v NAc puzdre nedošlo k zvýšeniu hustoty chrbtice po konzumácii sacharózy v týždni 4 prostredníctvom paradigmy samoadministrácie nosa-poke napriek výraznejšej akvizícii a vyššej odpovedi na sacharózu v porovnaní s amfetamínom (Crombag a kol., 2005). Ich pozorovanie absencie zmeny v hustote chrbtice v týždňoch 4 odráža naše zistenia. Naproti tomu naša štúdia dokazuje, že po dlhodobom (12 týždennom) vystavení chronickej spotrebe sacharózy dochádza k významnému zvýšeniu hustoty distálnej chrbtice na MSN potkanov so skúsenosťami so sacharózou. Okrem toho naša laboratória predtým ukázala, že dlhodobá spotreba sacharózy v týždni 12 uľahčuje diferenciálnu farmakologickú odpoveď na farmakoterapeutiká, u ktorých sa ukázalo, že modulujú odpovede na dopamín a acetylcholín na úrovni NAc (Shariff a kol., V tlači). Súhrnne to naznačuje, že dlhodobé expozície sacharózy (12 týždňov a neskôr), ktoré presnejšie odráža scenáre v reálnom svete, vedie k morfologickým úpravám na úrovni NAc.
Pokiaľ ide o drogy zneužívania, opakované vystavenie rôznym liekom spôsobuje dlhotrvajúce zmeny v štruktúre dendritov a dendritických tŕňov. Napríklad amfetamíny a kokaín zvyšujú hustotu chrbtice v NAc v obale aj jadre (Robinson a Kolb, 2004). Zistilo sa tiež, že expozícia nikotínu zvyšuje hustotu chrbtice v kryte NAc. Naopak expozícia morfínu vedie k zníženiu hustoty chrbtice a zložitosti dendritických vetví (Robinson a Kolb, 2004). Pokiaľ ide o dlhodobú spotrebu sacharózy, pozorovali sme zvýšenie hustoty chrbtice podobne ako amfetamín, kokaín a nikotín a oproti účinku morfínu. Avšak na rozdiel od amfetamínu a kokaínu, ale podobne ako nikotín, je zvýšenie hustoty chrbtice pri dlhodobej expozícii sacharóze obmedzené na shell NAc. Je tiež zaujímavé, že zmeny v oboch dendritických vetvách (Robinson a Kolb, 1999) a hustota chrbtice (Li et al., 2003) produkovaných amfetamínom alebo kokaínom sú obmedzené na distálne dendrity MSN v NAc, čo odráža zistenia v našej štúdii, Okrem toho a potvrdzujúca vyššie opísané zmeny sa už predtým ukázalo, že spotreba sacharózy zvyšuje excitačnú synaptickú silu na akumbalových dopamínových neurónoch (Stuber a kol., 2008b), ako aj iné zložky mezolimbickej cesty odmeny (Stuber a kol., 2008a; Chen a kol., 2010). Spoločne to predstavuje sacharózu ako silný modulátor morfológie neurónov po dlhšom ťažkom užívaní, čo je podobné účinkom pozorovaným pri užívaní drog.
Hoci sú potrebné ďalšie výskumy na odhalenie bunkových a synaptických mechanizmov prispievajúcich k morfologickým zmenám pozorovaným v tejto štúdii, naše výsledky ukazujú na významné neuronálne účinky vyvolané dlhodobou konzumáciou sacharózy. Konkrétne sa v našej štúdii uvažovalo o tom, či možno pozorovať morfologické účinky sacharózy aj s nekalorickými sladidlami, ako je sacharín. V tejto súvislosti je dôležité poznamenať, že Lenoir a jeho kolegovia ukázali, že intenzívna sladkosť prevyšuje odmenu za kokaín, či už sacharín alebo sacharóza (Lenoir a kol., 2007). Okrem toho nedávna štúdia publikovaná našou laboratóriou (Shariff a kol., V tlači) dokazuje, že vareniklín, čiastočný agonista nikotínového acetylcholínového receptora, znížil príjem sacharózy a sacharínu u hlodavcov po rovnakom dlhodobom intermitentnom prístupovom režime použitom v tejto štúdii. Zaujímavé je, že predchádzajúce štúdie ukázali podobnosti akútnych účinkov nekalorických sladidiel, ako je sacharín a sacharóza, na úrovni NAc (Scheggi a kol., 2013; Tukey a kol., 2013; Carelli a West, 2014). Ďalšie štúdie však musia určiť, či nekalorické sladidlá môžu vyvolať dlhodobé účinky podobné zmenám v morfológii MSN škrupín NAc v dôsledku dlhodobej spotreby sacharózy tu uvádzanej.
Nedostatok vplyvu na morfológiu NAc MSN po krátkodobej spotrebe sacharózy poukazuje na dôležitosť uskutočnenia dlhodobých štúdií na posúdenie vplyvu dlhodobého zneužívania drog alebo prírodných odmien ako je sacharóza. Z hľadiska závislosti sú nielen opakované cykly záchvatu príjmu a abstinenčných kľúčových zložiek cyklu závislostí, rastúci počet dôkazov ukázal, že prechod na závislosť je progresívny proces, ktorý sa často vyskytuje počas dlhého časového obdobia. Napriek tomu, že návykové vlastnosti cukrov zostávajú neisté, čoraz častejšie sa skúma hodnovernosť závislosti na iných odmenách, ako sú pohlavie, hazardné hry a potraviny. Výsledky tejto štúdie pripisujú zásluhu na hypotéze, že cukry, ako napríklad sacharóza, majú potenciálne návykové vlastnosti po dlhodobej, bravčovej konzumácii. Naše výsledky majú aj dôsledky pre rastúci počet detí a dospievajúcich, ktorí udržujú nezdravé stravovacie návyky (vysoká spotreba cukru a konzumácia alkoholu) až do dospelosti. V súlade so zvýšeným rizikom vývoja metabolických účinkov je tiež možné, že z tohto správania môžu vyplynúť aj neurologické a psychické následky ovplyvňujúce náladu a motiváciu.
Príspevky od autorov
Podieľal sa na výskume: PK, SB. Vykonané experimenty: PK, MS, AB, MF, EM. Analýza údajov: PK, MF, MS. Interpretovali údaje a prispeli k písaniu rukopisu: PK, MS, MF, EM, MB, SB. Všetci autori prečítali a schválili konečný rukopis na podanie.
Vyhlásenie o konflikte záujmov
Autori vyhlasujú, že výskum bol vykonaný bez obchodných alebo finančných vzťahov, ktoré by mohli byť interpretované ako potenciálny konflikt záujmov.
Hodnotitelia SC, SA a spracovateľský redaktor vyhlásili svoju zdieľanú príslušnosť a editor spracovania hovorí, že tento proces napriek tomu spĺňal normy spravodlivého a objektívneho preskúmania.
Poďakovanie
Táto práca bola podporená z grantov z austrálskej rady pre výskum (FT1110884) do SB a Národnej rady pre zdravotný a lekársky výskum (1061979) pre SB a MB.
Doplnkový materiál
Doplnkový materiál tohto článku sa nachádza na adrese: http://journal.frontiersin.org/article/10.3389/fnbeh.2016.00054
Doplnkový obrázok 1. Príjem a uprednostňovanie sacharózy u potkanov konzumujúcich sacharózu 4 a 12 týždeň. (A, B) ukazujú eskaláciu celkového príjmu sacharózy (ml) počas 4 a 12 týždňov expozície. (C, D) vykazujú vysokú preferenciu pre sacharózu nad vodou počas obdobia prezentácie sacharózy.
Referencie
Ahmed, SH a Koob, GF (1998). Prechod z mierneho na nadmerný príjem liekov: zmena hedonickej požadovanej hodnoty. veda 282, 298-300. dva: 10.1126 / science.282.5387.298
Ahmed, SH, Lin, D., Koob, GF a Parsons, LH (2003). Eskalácia samotného podávania kokaínu nezávisí od zmenených hladín dopamínu indukovaných kokaínom indukovaným jadrom. J. Neurochem. 86, 102-113. dva: 10.1046 / j.1471-4159.2003.01833.x
Ahmed, SH, Walker, JR a Koob, GF (2000). Trvalé zvyšovanie motivácie užívať heroín u potkanov s eskaláciou drog v minulosti. neuropsychofarmakologie 22, 413–421. doi: 10.1016/S0893-133X(99)00133-5
Arslanian, S. (2002). Diabetes typu 2 u detí: klinické aspekty a rizikové faktory. Horm Res 57 (doplnok 1), 19-28. dva: 10.1159 / 000053308
Avena, NM, Bocarsly, ME, Hoebel, BG a Gold, MS (2011). Prekrývanie sa v nosológii zneužívania návykových látok a prejedania: translačné dôsledky "závislosti od stravy". Akt. Zneužívanie drog Rev. 4, 133-139. dva: 10.2174 / 1874473711104030133
Avena, NM, Rada, P. a Hoebel, BG (2008). Dôkaz o závislosti na cukre: behaviorálne a neurochemické účinky prerušovaného, nadmerného príjmu cukru. Neurosci. Biobehav. Rev. 32, 20-39. dva: 10.1016 / j.neubiorev.2007.04.019
Avena, NM, Rada, P., Moise, N. a Hoebel, BG (2006). Sacharóza, ktorá sa podieľa na schémach záchvatov, opakovane uvoľňuje adumbín dopamínu a eliminuje odpoveď na pocity sýtosti acetylcholínom. Neurovedy 139, 813-820. dva: 10.1016 / j.neuroscience.2005.12.037
Benton, D. (2010). Plausibilita závislostí od cukru a jej úloha v obezite a poruchách príjmu potravy. Clin. Nutr. 29, 288-303. dva: 10.1016 / j.clnu.2009.12.001
Bray, GA a Popkin, BM (2014). Dietný cukor a telesná hmotnosť: dostali sme sa v kríze epidémie obezity a cukrovky ?: zdravie zatraceně! Nalejte cukor. Diabetes Care 37, 950-956. dva: 10.2337 / dc13-2085
Brog, JS, Salypongse, A., Deutch, AY a Zahm, DS (1993). Vzory aferentnej inervácie jadra a škrupiny v "accumbens" časti potkanieho ventrálneho striatu: imunohistochemická detekcia retrográdne transportovaného fluoro-zlata. J. Comp. Neurol. 338, 255-278. dva: 10.1002 / cne.903380209
Bucher Della Torre, S., Keller, A., Laure Depeyre, J. a Kruseman, M. (2015). Riziko cukrových nápojov a obezity u detí a dospievajúcich: systematická analýza toho, ako môže metodická kvalita ovplyvniť závery. J. Acad. Nutr. Strave. [Epub pred tlačou]. dva: 10.1016 / j.jand.2015.05.020
Carelli, RM a západ, EA (2014). Keď sa dobrý chuť zvrtne: nervové mechanizmy, ktoré sú základom vzniku negatívneho vplyvu a súvisiacej devalvácie prirodzenej odmeny kokaínom. Neuropharmacology 76 (Pt B), 360-369. dva: 10.1016 / j.neuropharm.2013.04.025
Chen, BT, Hopf, FW a Bonci, A. (2010). Synaptická plasticita v mezolimbickom systéme: terapeutické dôsledky na zneužívanie návykových látok. Ann. NY Acad. Sci. 1187, 129-139. dva: 10.1111 / j.1749-6632.2009.05154.x
Colantuoni, C., Rada, P., McCarthy, J., Patten, C., Avena, NM, Chadeayne, A. a kol. (2002). Dôkaz, že prerušovaný, nadmerný príjem cukru spôsobuje endogénnu závislosť od opiátov. Obesí. Res. 10, 478-488. dva: 10.1038 / oby.2002.66
Colantuoni, C., Schwenker, J., McCarthy, J., Rada, P., Ladenheim, B., Cadet, JL a kol. (2001). Nadmerný príjem cukru mení väzbu na dopamínové a mu-opioidné receptory v mozgu. Neuroreport 12, 3549–3552. doi: 10.1097/00001756-200111160-00035
Crombag, HS, Gorny, G., Li, Y., Kolb, B. a Robinson, TE (2005). Opačné účinky skúsenosti s amfetamínovým samoadministráciou na dendritických chrbtoch v mediálnej a orbitálnej prefrontálnej kôre. Cereb. kôra 15, 341-348. dva: 10.1093 / cercor / bhh136
Dabelea, D., Mayer-Davis, EJ, Saydah, S., Imperatore, G., Linder, B., Divers, J. a kol. (2014). Prevalencia typu 1 a typu 2 diabetu u detí a adolescentov od 2001 po 2009. JAMA 311, 1778-1786. dva: 10.1001 / jama.2014.3201
Davis, C. (2015). Epidemiológia a genetika poruchy príjmu potravy (BED). CNS Spectr. 20, 522-529. dva: 10.1017 / s1092852915000462
Ford, ES a Dietz, WH (2013). Trendy v príjme energie medzi dospelými v Spojených štátoch: zistenia z NHANES. Am. J. Clin. Nutr. 97, 848-853. dva: 10.3945 / ajcn.112.052662
Fryar, CD, Carroll, MD a Ogden, CL (2014). Prevalencia nadváhy, obezity a extrémnej obezity u dospelých: Spojené štáty, 1960-1962 cez 2011-2012, Atlanta, GA: CDC.
Graveland, GA a DiFiglia, M. (1985). Frekvencia a distribúcia stredne veľkých neurónov s oddelenými jadrami v primáte a hlodavcovi neostriatum. Brain Res. 327, 307–311. doi: 10.1016/0006-8993(85)91524-0
Groenewegen, HJ, Wright, CI, Beijer, AV a Voorn, P. (1999). Konvergencia a segregácia ventrálnych striatálnych vstupov a výstupov. Ann. NY Acad. Sci. 877, 49–63. doi: 10.1111/j.1749-6632.1999.tb09260.x
Groves, PM (1980). Synaptické koncovky a ich postsynaptické ciele v neostriatum: synaptické špecializácie odhalené z analýzy sériových sekcií. Proc. Natl. Acad. Sci. USA 77, 6926-6929. dva: 10.1073 / pnas.77.11.6926
Groves, PM, Linder, JC a Young, SJ (1994). Dopaminergné axóny označené 5-hydroxydopamínom: trojrozmerné rekonštrukcie axónov, synapsií a postsynaptických cieľov u krys neostriatum. Neurovedy 58, 593–604. doi: 10.1016/0306-4522(94)90084-1
Hu, FB (2013). Vyriešené: existujú dostatočné vedecké dôkazy, že zníženie spotreby nápojov osladeného cukrom zníži prevalenciu obezity a ochorení súvisiacich s obezitou. Obesí. Rev. 14, 606-619. dva: 10.1111 / obr.12040
Johnson, LR, Aylward, RL, Hussain, Z. a Totterdell, S. (1994). Vstup z amygdaly do krysieho nucleus accumbens: jeho vzťah s imunoreaktivitou tyrozínhydroxylázy a identifikovanými neurónmi. Neurovedy 61, 851–865. doi: 10.1016/0306-4522(94)90408-1
Kaiya, H. a Namba, M. (1981). Dva typy dopaminergných nervových zakončení v krysom neostriatum. Ultraštruktúrna štúdia. Neurosci. Letí. 25, 251–256. doi: 10.1016/0304-3940(81)90400-6
Kalivas, PW a Miller, JS (1984). Neurónové neuroturóny v oblasti ventrálneho tegmentálneho priestoru smerujú k strednému jadru accumbens. Brain Res. 300, 157–160. doi: 10.1016/0006-8993(84)91351-9
Kawaguchi, Y., Wilson, CJ a Emson, PC (1990). Projekčné podtypy buniek krysích neostriatálnych matríc odhalené intracelulárnou injekciou biocytínu. J. Neurosci. 10, 3421-3438.
Kelley, AE a Swanson, CJ (1997). Kŕmenie indukované blokádou AMPA a kainátových receptorov vo vnútri ventrálneho striatu: štúdia mikroinfúzneho mapovania. Behave. Brain Res. 89, 107–113. doi: 10.1016/S0166-4328(97)00054-5
Kemp, JM a Powell, TP (1971). Synaptická organizácia kazatej jadra. Philos. Trans. R. Soc. Londa. B Biol. Sci. 262, 403-412. dva: 10.1098 / rstb.1971.0103
Kenny, PJ (2011). Mechanizmy odmeňovania v obezite: nové poznatky a budúce smerovanie. Neurón 69, 664-679. dva: 10.1016 / j.neuron.2011.02.016
Kessler, RC, Berglund, PA, Chiu, WT, Deitz, AC, Hudson, JI, Shahly, V. a kol. (2013). Prevalencia a korelácia poruchy príjmu potravy z dôvodu nadmernej výživy v Svetových zdravotníckych organizáciách Svetové prieskumy duševného zdravia. Biol. psychiatrie 73, 904-914. dva: 10.1016 / j.biopsych.2012.11.020
Klenowski, PM, Fogarty, MJ, Belmer, A., Noakes, PG, Bellingham, MC a Bartlett, SE (2015). Štruktúrna a funkčná charakterizácia dendritických trávnikov a GABAergických synaptických vstupov na interneuronoch a hlavných bunkách v basolaterálnej amygade. J. Neurophysiol. 114, 942-957. dva: 10.1152 / jn.00824.2014
Lenoir, M., Serre, F., Cantin, L. a Ahmed, SH (2007). Intenzívna sladkosť prevyšuje odmenu za kokaín. PLoS ONE 2: e698. dva: 10.1371 / journal.pone.0000698
Li, Y., Kolb, B. a Robinson, TE (2003). Umiestnenie pretrvávajúcich amfetamínom indukovaných zmien v hustote dendritických tŕňov na stredne štipľavých neurónoch v nucleus accumbens a caudate-putamen. neuropsychofarmakologie 28, 1082-1085. dva: 10.1038 / sj.npp.1300115
Lindvall, O., a Björklund, A. (1978). Anatómia dopaminergných neurónových systémov v mozgu potkanov. Prísl. Biochem. Psychopharmacol. 19, 1-23.
Lutter, M. a Nestler, EJ (2009). Homeostatické a hedonické signály interagujú pri regulácii príjmu potravy. J. Nutr. 139, 629-632. dva: 10.3945 / jn.108.097618
Maldonado-Irizarry, CS, Swanson, CJ a Kelley, AE (1995). Glutamátové receptory v správaní kŕmenia jadra accumbens kontrolujú napájanie cez laterálny hypotalamus. J. Neurosci. 15, 6779-6788.
Malik, VS, Popkin, BM, Bray, GA, Després, JP a Hu, FB (2010). Cukry osladené nápoje, obezita, diabetes mellitus typu 2 a riziko kardiovaskulárnych ochorení. Obeh 121, 1356-1364. dva: 10.1161 / CIRCULATIONAHA.109.876185
Ng, SW, Slining, MM a Popkin, BM (2012). Používanie kalorických a nekalorických sladidiel v amerických spotrebiteľských balených potravinách, 2005-2009. J. Acad. Nutr. Strave. 112, 1828-1834 e1821-e1826. dva: 10.1016 / j.jand.2012.07.009
Nielsen, SJ, Siega-Riz, AM a Popkin, BM (2002). Trendy v príjme energie v USA medzi 1977 a 1996: podobné posuny vidieť vo všetkých vekových skupinách. Obesí. Res. 10, 370-378. dva: 10.1038 / oby.2002.51
Paxinos, G. a Watson, C. (2007). Krysa mozog v stereotaxických súradniciach, Amsterdam; Boston, MA: Academic Press / Elsevier.
Popkin, BM (2010). Čo je zlé na prístupe USA k obezite? Virtuálny mentor 12, 316–320. doi: 10.1001/virtualmentor.2010.12.4.pfor2-1004
Rada, P., Avena, NM a Hoebel, BG (2005). Každodenné záchvaty cukru opakovane uvoľňujú dopamín do shellu accumbens. Neurovedy 134, 737-744. dva: 10.1016 / j.neuroscience.2005.04.043
Rafols, JA, Cheng, HW a McNeill, TH (1989). Golgiho štúdia myšacieho striatu: vekové dendritické zmeny v rôznych populáciách neurónov. J. Comp. Neurol. 279, 212-227. dva: 10.1002 / cne.902790205
Ranjan, A. a Mallick, BN (2010). Modifikovaná metóda na konzistentné a spoľahlivé zafarbenie Golgiho koxu vo výrazne zníženom čase. Predná. Neurol. 1: 157. dva: 10.3389 / fneur.2010.00157
Reinehr, T. (2013). Diabetes mellitus typu 2 u detí a dospievajúcich. Svet J. Diabetes 4, 270-281. dva: 10.4239 / wjd.v4.i6.270
Reissner, KJ a Kalivas, PW (2010). Použitie homeostázy glutamátu ako cieľa liečby návykových porúch. Behave. Pharmacol. 21, 514–522. doi: 10.1097/FBP.0b013e32833d41b2
Reynolds, SM a Berridge, KC (2003). Glutamátové motivačné súbory v nucleus accumbens: rostrokaudálne škrupiny škrupiny strachu a kŕmenia. Eur. J. Neurosci. 17, 2187-2200. dva: 10.1046 / j.1460-9568.2003.02642.x
Richard, JM a Berridge, KC (2011). Nucleus accumbens dopamín / glutamátová interakcia prepína režimy na generovanie túžby verzus strach: D (1) samotný pre potrava apetitívne, ale D (1) a D (2) spoločne zo strachu. J. Neurosci. 31, 12866-12879. dva: 10.1523 / JNEUROSCI.1339-11.2011
Robinson, TE a Kolb, B. (1999). Zmeny v morfológii dendritov a dendritických tŕňov v nucleus accumbens a prefrontálnej kôre po opakovanej liečbe amfetamínom alebo kokaínom. Eur. J. Neurosci. 11, 1598-1604. dva: 10.1046 / j.1460-9568.1999.00576.x
Robinson, TE a Kolb, B. (2004). Štruktúrna plasticita spojená s vystavením zneužívaniu drog. Neuropharmacology 47 (doplnok 1), 33-46. dva: 10.1016 / j.neuropharm.2004.06.025
Russo, SJ, Dietz, DM, Dumitriu, D., Morrison, JH, Malenka, RC a Nestler, EJ (2010). Závislosť synapsa: mechanizmy synaptickej a štrukturálnej plasticity v nucleus accumbens. Trendy Neurosci. 33, 267-276. dva: 10.1016 / j.tins.2010.02.002
Rutledge, LT, Duncan, J. a Beatty, N. (1969). Štúdia pyramídových bunkových axonových kolaterálov v neporušenej a čiastočne izolovanej dospelosti mozgovej kôry. Brain Res. 16, 15–22. doi: 10.1016/0006-8993(69)90082-1
Saper, CB, Chou, TC a Elmquist, JK (2002). Potreba kŕmiť: homeostatická a hedonická kontrola stravovania. Neurón 36, 199–211. doi: 10.1016/S0896-6273(02)00969-8
Scheggi, S., Secci, ME, Marchese, G., De Montis, MG a Gambarana, C. (2013). Vplyv chutnosti na motiváciu pracovať pre kalorické a nekalorické potraviny u potracov, ktorí nie sú zbavení potraviny a zbavení potravy. Neurovedy 236, 320-331. dva: 10.1016 / j.neuroscience.2013.01.027
Sesack, SR a Pickel, VM (1990). V stredu mediálneho jadra accumbens, hippocampal a katecholaminergné terminály sa zbiehajú na špinavé neuróny a navzájom sa prispôsobujú. Brain Res. 527, 266–279. doi: 10.1016/0006-8993(90)91146-8
Sesack, SR a Pickel, VM (1992). Prefrontálne kortikálne eferenty v synapse potkanov na neoznačených neuronálnych cieľoch katecholamínových terminálov v nucleus accumbens septi a na dopamínových neurónoch v ventrálnej tegmentálnej oblasti. J. Comp. Neurol. 320, 145-160. dva: 10.1002 / cne.903200202
Shariff, M., Quik, M., Holgate, JY, Morgan, M., Patkar, OL, Tam, V. a kol. (v tlači). Modulátory neuronálneho nikotínového acetylcholínového receptora znižujú príjem cukru. PLoS ONE.
Sheehan, DV a Herman, BK (2015). Psychologické a medicínske faktory spojené s neliečenou poruchou príjmu potravy. Prim. Care companion CNS Disord. 17. dva: 10.4088 / PCC.14r01732
Simms, JA, Steensland, P., Medina, B., Abernathy, KE, Chandler, LJ, Wise, R. a kol. (2008). Prerušovaný prístup k etanolu 20% spôsobuje vysokú spotrebu etanolu u potkanov Long-Evans a Wistar. Alkohol. Clin. Exp. Res. 32, 1816-1823. dva: 10.1111 / j.1530-0277.2008.00753.x
Steensland, P., Simms, JA, Holgate, J., Richards, JK a Bartlett, SE (2007). Vareniklín, parciálny agonista nikotínového acetylcholínového receptora alpha4beta2, selektívne znižuje spotrebu a hľadanie etanolu. Proc. Natl. Acad. Sci. USA 104, 12518-12523. dva: 10.1073 / pnas.0705368104
Stuber, GD, Hopf, FW, Hahn, J., Cho, SL, Guillory, A. a Bonci, A. (2008a). Dobrovoľný príjem etanolu zvyšuje excitačnú synaptickú silu vo ventrálnej tegmentálnej oblasti. Alkohol. Clin. Exp. Res. 32, 1714-1720. dva: 10.1111 / j.1530-0277.2008.00749.x
Stuber, GD, Klanker, M., de Ridder, B., Bowers, MS, Joosten, RN, Feenstra, MG a kol. (2008b). Predvídateľné odozvy zvyšujú excitačnú synaptickú silu na dopamínové neuróny stredného mozgu. veda 321, 1690-1692. dva: 10.1126 / science.1160873
Swanson, SA, Crow, SJ, Le Grange, D., Swendsen, J. a Merikangas, KR (2011). Prevalencia a korelácia porúch stravovania u dospievajúcich. Výsledky z národného doplnku adolescentnej replikácie pri prieskume komorbidity. Arch. Gen. Psychiatry 68, 714-723. dva: 10.1001 / archgenpsychiatry.2011.22
Te Morenga, L., Mallard, S. a Mann, J. (2013). Dietetické cukry a telesná hmotnosť: systematické preskúmanie a metaanalýzy randomizovaných kontrolovaných štúdií a kohortových štúdií. BMJ 346: e7492. dva: 10.1136 / bmj.e7492
Totterdell, S. a Smith, AD (1989). Konvergencia hipokampálneho a dopaminergného vstupu na identifikované neuróny v nucleus accumbens potkana. J. Chem. Neuroanat. 2, 285-298.
Tukey, DS, Ferreira, JM, Antoine, SO, D'Amour, JA, Ninan, I., Cabeza de Vaca, S. a kol. (2013). Požitie sacharózy indukuje rýchle obchodovanie s AMPA receptorom. J. Neurosci. 33, 6123-6132. dva: 10.1523 / JNEUROSCI.4806-12.2013
Veening, JG, Cornelissen, FM a Lieven, PA (1980). Aktuálna organizácia príbuzných kauzotoputánov potkana. Štúdia chrenovej peroxidázy. Neurovedy 5, 1253–1268. doi: 10.1016/0306-4522(80)90198-0
Ventura, T., Santander, J., Torres, R. a Contreras, AM (2014). Neurobiologický základ túžby po sacharidoch. Výživa 30, 252-256. dva: 10.1016 / j.nut.2013.06.010
Volkow, ND, Wang, GJ, Fowler, JS, Tomasi, D. a Baler, R. (2012). Odmena za potraviny a drogy: prekrývajúce sa obvody v ľudskej obezite a závislosti. Akt. Top. Behave. Neurosci. 11, 1–24. doi: 10.1007/7854_2011_169
Múdre, RA (1973). Dobrovoľný príjem etanolu u potkanov po expozícii etanolu v rôznych plánoch. Psychopharmacologia 29, 203-210. dva: 10.1007 / BF00414034
Wright, CI a Groenewegen, HJ (1995). Vzory konvergencie a segregácie v mediálnom nucleus accumbens potkana: vzťahy prefrontálnej kortikálnej, strednej talamickej a bazálnych amygdaloidných aferentov. J. Comp. Neurol. 361, 383-403. dva: 10.1002 / cne.903610304
Kľúčové slová: spotreba podobná záchvatu, dlhodobý, stredne štipľavý neurón, nucleus accumbens, sacharóza
Citácia: Klenowski PM, Shariff MR, Belmer A, Fogarty MJ, Mu EWH, Bellingham MC a Bartlett SE (2016) Predĺžená spotreba sacharózy v podobe záchvatu, mení morfológiu stredne štipľavých neurónov v Nucleus Accumbens Shell. Predná. Behave. Neurosci, 10: 54. dva: 10.3389 / fnbeh.2016.00054
Prijaté: 03 December 2015; Prijaté: 07 March 2016;
Publikované: 23 March 2016.
strih:
Djoher Nora Abrous, Institut des Neurosciences de Bordeaux, Francúzsko
Hodnotené:
Serge H. Ahmed, Centrum National de la Recherche Scientifique, Francúzsko
Stéphanie Caille, Centrum National de la Recherche Scientifique, Francúzsko
Copyright © 2016 Klenowski, Shariff, Belmer, Fogarty, Mu, Bellingham a Bartlett. Toto je článok s otvoreným prístupom distribuovaný podľa podmienok Creative Commons Attribution License (CC BY), Používanie, distribúcia alebo reprodukcia na iných fórach je povolené za predpokladu, že pôvodný autor (autorov) alebo poskytovateľ licencie je pripísaný a že pôvodná publikácia v tomto časopise je citovaná v súlade s prijatou akademickou praxou. Nepoužíva sa žiadna distribúcia alebo reprodukcia, ktorá nie je v súlade s týmito podmienkami.
* Korešpondencia: Selena E. Bartlettová, [chránené e-mailom]
