 Martin Zack
Martin Zack- 1Kognitívne psychofarmakologické laboratórium, oddelenie neurovedy, Centrum pre závislosť a duševné zdravie, Toronto, ON, Kanada
- 2Translačný neurovedecký program, Katedra psychiatrie, Lekárska fakulta, University of Pennsylvania, Philadelphia, PA, USA
- 3Sekcia biopsychológie, oddelenie neurovedy, Centrum pre závislosť a duševné zdravie, Toronto, ON, Kanada
Závislosť sa považuje za ochorenie mozgu spôsobené chronickou expozíciou drogám. Senzibilizácia systémov dopamínu mozgu (DA) čiastočne sprostredkováva tento účinok. Patologické hráčstvo (PG) sa považuje za behaviorálnu závislosť. PG môže byť preto spôsobený chronickou expozíciou hazardu. Identifikácia senzibilizácie systémov DA vyvolaných hazardom by túto možnosť podporila. Odmeny za hazardné hry evokujú vydanie DA. Jedna epizóda hry hracieho automatu posúva DA odozvu od odovzdania odmeny k nástupu cue (spinning kotúčov) za odmenu v súlade s princípmi časového rozlíšenia učenia. Podmienené stimuly (CS) teda hrajú kľúčovú úlohu v odpovediach DA na hazardné hry. V primátoch je odpoveď DA na CS najsilnejšia, keď pravdepodobnosť odmeny je 50%. Podľa tohto harmonogramu CS vyvoláva očakávanú odmenu, ale neposkytuje žiadne informácie o tom, či k nemu dôjde v danom konaní. Počas hrania hazardných hier by mal 50% program vyvolať maximálne uvoľňovanie DA. To úzko zodpovedá frekvencii odmeny (46%) na komerčnom automate. Uvoľňovanie DA môže prispieť k senzibilizácii, najmä pre amfetamín. Chronická expozícia CS, ktorá predpovedá odmenu 50% času, by mohla tento efekt napodobniť. Túto hypotézu sme testovali v troch štúdiách s potkanmi. Zvieratá dostali expozície 15 × 45-min do CS, ktoré predpovedali odmenu s pravdepodobnosťou 0, 25, 50, 75 alebo 100%. CS bolo svetlo; Odmenou bol roztok 10% sacharózy. Po tréningu dostávali potkany senzibilizačný režim piatich samostatných dávok (1 mg / kg) d-amfetamínu. Nakoniec dostávali 0.5 alebo 1 mg / kg amfetamínovú stimuláciu pred testom lokomotorickej aktivity 90-min. Vo všetkých troch štúdiách skupina 50% vykazovala vyššiu aktivitu ako ostatné skupiny v reakcii na obe stimulačné dávky. Veľkosti účinku boli skromné, ale konzistentné, čo sa odráža v významnej skupinovej asociácii (ϕ = 0.986, p = 0.025). Chronické vystavenie gamblingu podobného rozvrhu prediktívnych stimulov môže podporiť senzibilizáciu na amfetamín, podobne ako expozícia samotnému amfetamínu.
úvod
Závislosť bola charakterizovaná ako ochorenie mozgu spôsobené chronickým vystavením návykovým látkam (Leshner, 1997). Predpokladá sa, že neuroplasticita sprostredkováva účinky takejto expozície (\ tNestler, 2001). Senzibilizácia systémov dopamínu v mozgu (DA) je forma neuroplasticity, ktorá sa podieľa na hyperreaktivite na podmienené stimuly (CS) pre lieky a kompulzívne hľadanie liekov (Robinson a Berridge, 2001). Senzibilizácia bola operačne definovaná zvýšeným uvoľňovaním DA ako odozva na CS pre odmenu a zvýšenou lokomotorickou odpoveďou na farmakologickú DA výzvu (Robinson a Berridge, 1993; Pierce a Kalivas, 1997; Vanderschuren a Kalivas, 2000). Hoci senzibilizácia je len jednou z mnohých zmien mozgu spojených so závislosťou (porov. Robbins a Everitt, 1999; Koob a Le Moal, 2008) sa predpokladá, že zmeny v presynaptickom uvoľňovaní dopamínu predstavujú spoločné neuroadaptácie, ktoré sa podieľajú na hľadaní drog závislých od závislosti (napr. relaps), v tom, že lieky, ktoré indukujú lokomotorickú senzibilizáciu na opiátovú (napr. morfínovú) alebo stimulačnú stimuláciu (napr. amfetamín) spôsobujú aj opätovné obnovenie zaniknutých operatívnych reakcií na samoliečbu heroínom alebo kokaínom - zvierací model relapsu (Vanderschuren a kol., 1999). Dôkazy o tom, že senzitizácia podnetov (zvýšená hodnota odmien za drogy) je najvýraznejšia po počiatočnom vystavení návykovým látkam, ďalej svedčí o tom, že senzibilizácia môže byť zahrnutá aj v raných štádiách závislosti.Vanderschuren a Pierce, 2010).
Patologické hráčstvo (PG) bolo popísané ako behaviorálna závislosť a nedávno bolo zaradené do rovnakej kategórie ako poruchy látkovej závislosti v 5th vydanie Diagnostického a štatistického manuálu duševných porúch (Frascella a kol., 2010; APA, 2013). To znamená, že PG môže byť spôsobené chronickým vystavením účinkom podobných hazardných hier, že spoločné mechanizmy môžu sprostredkovať účinky hazardných hier a vystavenia drogám (Zack a Poulos, 2009; Leeman a Potenza, 2012); a že senzibilizácia dráh mozgu môže byť jedným z dôležitých prvkov tohto procesu.
Klinické dôkazy nepriamo podporujú túto možnosť: Použitie pozitrónovej emisnej tomografie (PET) Boileau a jeho kolegovia zistili, že mužskí PG jedinci vykazujú významne vyššie uvoľnenie striatálneho DA ako odpoveď na amfetamín (0.4 mg / kg) ako u zdravých mužských kontrol (Boileau a kol., 2013). Celkové skupinové rozdiely boli významné v asociatívnom a somatosenzorickom striate. V limbickom striate, ktoré obsahuje nucleus accumbens, sa skupiny nelíšili. U jedincov s PG však DA uvoľňovanie v limbickom striatu korelovalo priamo so závažnosťou symptómov PG. Tieto nálezy sú v súlade so senzibilizáciou dráh mozgu DA v PG, ale tiež naznačujú niektoré dôležité rozdiely medzi jedincami závislými od ľudskej látky a klasickým zvieracím modelom senzibilizácie amfetamínu. Na rozdiel od subjektov PG a zvierat vystavených nízkym dávkam amfetamínu (porovnaj Robinson a kol., 1982), ľudia so závislosťou od látky konzistentne vykazujú znížené uvoľňovanie DA do stimulačného stimulu (Volkow a kol., 1997; Martinez a kol., 2007), a dôkazy zo zvierat naznačujú, že to môže odrážať deficity funkcie DA počas počiatočných štádií abstinencie po vzplanutí návykových látok (Mateo a spol., 2005). V štúdiách, kde sa senzibilizácia stimulancií preukázala na zvieratách, sa zvýšené uvoľňovanie DA zvyčajne pozoruje skôr v limbickom striate, ako v dorzálnom (asociatívnom, somatosenzorickom) striatu (Vezina, 2004). Avšak cielené (tj podmienené) hľadanie liekov u zvierat opakovane vystavených kokaínu bolo spojené so zvýšeným uvoľňovaním DA v dorzálnom striate, čo je výsledok, ktorý naznačoval viac obvyklú formu motivovaného správania (Ito a kol., 2002). Celkové zvýšenie uvoľňovania DA v dorzálnych oblastiach u pacientov s PG môže byť teda spojené s vyhľadávaním na základe návykov (neflexibilné, rutinizované), ktoré zahŕňajú „progres od ventrálnej k viac dorzálnej doméne striatum“ (Everitt a Robbins, 2005, s. 1481), zatiaľ čo uvoľňovanie DA závislé od závažnosti v limbickom striate u týchto jedincov môže zodpovedať skôr stimulačnej senzibilizácii, ako je typicky modelované u zvierat. PET nálezy nedokážu odhaliť, či DA hyper-reaktivita bola už existujúcim znakom týchto PG jedincov, čo je dôsledok vystavenia hráčom, alebo výsledok nejakého iného procesu. Na riešenie tejto otázky je potrebné preukázať indukciu senzibilizácie chronickou expozíciou hazardných hier u subjektov, ktoré sú pred expozíciou normálne. To vyvoláva otázky, aké vlastnosti hazardných hier s najväčšou pravdepodobnosťou vyvolávajú senzibilizáciu.
Skinner poznamenal, že variabilný harmonogram posilňovania bol základom pre lákavosť hazardných hier (alebo aspoň pre ich pretrvávanie) (Skinner, 1953). Stávkové správanie v hre hracieho automatu je v súlade so základnými princípmi inštrumentálneho kondicionovania, čo sa odráža v budúcej korelácii medzi peňažnou návratnosťou a veľkosťou stávky na po sebe nasledujúcich spinoch (Tremblay a kol., 2011). Zdá sa teda, že operatívna reakcia variabilného pomeru poskytuje externe platný model hazardných hier na hracích automatoch.
Nedávny výskum so zvieratami poskytuje silnú počiatočnú podporu pre kauzálny účinok expozície hazardnými hrami na senzibilizáciu. Singer a jeho kolegovia skúmali účinky 55 1-h denných relácií fixného (FR20) alebo variabilného (VR20) posilnenia sacharínu v operatívnej pákovej tlakovej paradigme pri následnej lokomotorickej odpovedi na nízku dávku (0.5 mg / kg) amfetamínu u zdravých mužov ( Sprague Dawley) potkany (Singer a kol., 2012). Predpokladali, že ak by hazardné hry viedli k senzibilizácii, potkany vystavené variabilnému rozvrhu, ktoré napodobňujú hazardné hry, by mali vykazovať väčšiu odozvu na amfetamín ako potkany vystavené stanovenému rozvrhu. Ako bolo predpovedané, skupina VR20 vykazovala 50% väčšiu lokomotorickú odozvu na amfetamín ako skupina FR20. Naproti tomu skupiny vykazovali ekvivalentnú lokomóciu po injekcii fyziologického roztoku. Tieto zistenia potvrdzujú, že chronická expozícia premenlivému zosilneniu je dostatočná na vyvolanie hyperreaktivity na DA stimuláciu u zdravých zvierat randomizovaných do príslušných schém.
Z tohto výsledku vyplynulo niekoľko otázok: Po prvé, do akej miery sprostredkúva vnímaná podmienenosť - alebo jej nedostatok - medzi operatívnou odpoveďou a jej výsledkom tieto účinky? V termínoch učenia zahŕňa tento účinok „očakávanú dĺžku odozvy - výsledok“, alebo by mohol byť podobný účinok pozorovaný v neprítomnosti operantnej odpovede, tj „očakávaná dĺžka stimulu“ v Pavlovovskej paradigme (porovnaj Bolles, 1972)? Po druhé, stupeň závislosti medzi predchádzajúcou udalosťou (odpoveď alebo podnet) a jej výsledkom ovplyvňuje stupeň senzibilizácie?
Druhá otázka sa týka úlohy neistoty pri senzibilizácii. Napríklad, majú hry, ktorých výsledok je skutočne náhodný - úplne nepredvídateľný - majú väčší potenciál vyvolať senzibilizáciu ako hry, kde sú šance na výhru jasne definované, ale nie náhodné, aj keď absolútna miera odmeny je nízka? Tento výskum riešil tieto otázky.
Experimentálny návrh bol informovaný prostredníctvom kľúčovej štúdie o očakávanej odmene a odozve DA neurónov u opíc (Fiorillo a kol., 2003). Zvieratá v tejto štúdii dostali odmenu za šťavu (US) pod schémami 0, 25, 50, 75 alebo 100% variabilného pomeru. Plány boli určené 1 4 rôznych CS (ikon). Rozvrh 0% priniesol odmenu tak často ako plán 100%, ale vynechal CS. Rýchlosť streľby DA neurónov počas intervalu medzi nástupom CS a vynechaním alebo vynechaním v USA bola kľúčovo závislým meradlom. Štúdia zistila, že odpoveď DA sa zvýšila ako funkcia neistoty pri poskytovaní odmeny. Takže pod rozpisom 100% CS vyvolala malú aktivitu, pod schémami 25 a 75%, CS evokovala strednú a podobnú úroveň aktivity a pod plánom 50% CS vyvolala maximálnu aktivitu. V každom prípade sa rýchlosť streľby v priebehu intervalu CS-US zvyšovala, tzn.
Tieto zistenia naznačujú, že DA aktivita sa líši nielen s tým, či je alebo nie je odmena určitá (fixný pomer) alebo neistá (variabilný pomer), ale tiež sa mení v opačnom pomere k množstvu informácií o dodaní odmeny, ktoré sprostredkuje CS. V podmienkach 100%, CS evokuje očakávanú odmenu a tiež dokonale predpovedá jej doručenie. V podmienkach 25 a 75%, CS evokuje očakávania a predpovedá dodanie odmeny tri zo štyroch krát. V podmienkach 50, CS evokuje očakávania, ale neposkytuje žiadne informácie o samotnom odmeňovaní. Na základe ich zistení Fiorillo et al. dospel k záveru: „Toto zvýšenie neistoty vyvolané dopamínom by mohlo prispieť k odmeňovaniu vlastností hazardných hier“ (s. 1901).
Účinky 50% variabilnej odmeny v jednej relácii by sa nemali meniť v priebehu viacerých relácií, pretože pravdepodobnosť odmeny zostáva pri každom skúšaní úplne nepredvídateľná. Pri zvažovaní podmienok, ktoré by maximalizovali chronickú aktiváciu DA neurónov počas opakovaných epizód hazardu, by teda 50% schéma mala priniesť najtrvalejší aj najsilnejší účinok. To je pozoruhodné vzhľadom na to, že dlhodobá miera odmeny (výplata> 0) pozorovaná pri tisícoch roztočení na komerčnom hracom automate bola 45.8% (Tremblay a kol., 2011). Zdá sa teda, že variabilná odmena 50% presne odráža plán výplaty, ktorý spravujú skutočné herné zariadenia.
Predložená štúdia používala rovnaké kondičné plány ako Fiorillo et al. pri chronickej expozícii medzi skupinami u potkanov. Zvieratá podstúpili ~ 3 týždne denných kondičných sedení, kde sa CS (svetlo) spárovalo s USA (malé množstvo sacharózy). Po fáze tréningu zvieratá odpočívali pred hodnotením senzibilizácie indexovanej pohybovou odpoveďou na amfetamín. Na základe literatúry sa predpokladalo, že potkany vystavené rôznym harmonogramom odmien sa nebudú líšiť v lokomotorickom správaní bez liečiva, ale budú vykazovať významne odlišné úrovne lokomócie po amfetamíne, pričom skupina s 50% prejaví vyššiu pohybovú odozvu na liečivo v porovnaní s u ostatných skupín v priebehu dávok, čo by sa dalo očakávať, keby bolo 50% zvierat predtým vystavených ďalším dávkam samotného amfetamínu (tj krížovej senzibilizácii).
experiment 1
Materiály a metódy
Predmety
Štyri skupiny (n = 8 / skupina) dospelých (300 – 350 g) samcov potkanov Sprague-Dawley (Charles River, St. Constant, Quebec, Kanada) sa umiestnili jednotlivo v priehľadných polykarbonátových boxoch (20 × 43 × 22 cm) pod reverzným 12: 12 cyklus svetlo-tma. Dostali podľa chuti prístup k potravinám a vode a denná manipulácia experimentátorom počas 2 týždňov pred štúdiou. Každá skupina bola podmienená jednou zo štyroch schém variabilného odmeňovania: 0, 25, 50 alebo 100%. Skupina 75% bola v tejto počiatočnej štúdii vynechaná, ako je uvedené vyššie Fiorillo a kol. (2003) našiel ekvivalentné post-CS DA vydanie v rámci 25 a 75% odmeny plány, takže obe podmienky viedli k väčšiemu vydaniu DA, ako tomu bolo v prípade 100% CS-US, ale menej ako podmienka 50%.
Zariadenie
Prístup k prezentáciám sacharózy a CS bol poskytovaný individuálne v operačných boxoch (33 × 31 × 29 cm). Každá krabica bola vybavená zosilňovačom, ktorý sa nachádza na prednej stene. Svetlo v hornej časti časopisu slúžilo ako CS. Motorizovaná, solenoidom riadená kvapalinová nádržka by mohla byť vyvýšená na podlahu zásobníka. Udalosti v boxe boli kontrolované zariadením a softvérom spoločnosti Med Associates, pričom sa použil interný program napísaný v programe MED-PC. Lokomotorické testovanie sa uskutočňovalo individuálne v klietkach Plexiglas (27 × 48 × 20 cm). Každá klietka bola vybavená monitorovacím systémom pozostávajúcim zo šiestich fotočiarnych buniek na detekciu horizontálneho pohybu.
Postup
Vzdelávanie, Štúdia bola vykonaná v súlade s etickými smernicami stanovenými Kanadskou radou pre starostlivosť o zvieratá. Potkany boli počas trvania štúdie obmedzené na 90% ich telesnej hmotnosti a boli umiestnené individuálne. Každý potkan dostal 15 dní tréningu sacharózovej odmeny (10% vodný roztok pri 0.06 ml za odmenu): 5 po sebe idúce dni × 3 týždňov, s víkendmi preč. Zvieratá sa udržiavali na štandardnom krmive pred a po tréningovej fáze; Expozícia sacharózy bola obmedzená na pätnásť školení 40-min. Každá denná relácia pozostávala z 15 stimulačných prezentácií (svetlo; CS), z ktorých každý bol oddelený inter-trial intervalom 120 s. Svetlo bolo umiestnené v hornom paneli časopisu a zostalo na 25 s, pričom sacharóza bola k dispozícii počas posledných 5 s. V prípade skupiny 0 sa zvýši obsah sacharózy každý 140 s (pre 5 s), ale stimulačné svetlo nebolo osvetlené. Toto sa rovnalo intervalu medzi prezentáciami ponorky v skupine 0 a ostatnými skupinami (120 + 25 s). Každá liečebná relácia trvala ~ 40 min. Skupina 25 dostala v priemere raz za štyri prezentácie CS raz sacharózu; skupina 50 dostala sacharózu raz za každé dve prezentácie CS a skupina 100 dostala sacharózu po každej prezentácii CS.
testovanie, Dva týždne po poslednom prístupe k sacharóze (alebo „kondicionácii“) sa hodnotila lokomotorická odpoveď na d-amfetamín (AMPH; ip). Potkanom sa podávali tri 2-h sedenia, aby sa zvykli na lokomotorické boxy, po ktorých nasledovalo šesť AMPH testovacích zasadnutí. Testovacie dni AMPH sa uskutočnili v intervaloch 1-wk. V testovacích dňoch boli potkanom podávané 30 min na navyknutie do boxov a potom bola podaná jedna dávka 0.5 mg / kg AMPH nasledovaná na oddelených týždenných reláciách piatimi dávkami 1.0 mg / kg (jedna dávka denne) v testovacích dňoch 1 cez 5 , Lokomotíva po AMPH bola hodnotená na 90 min na každej relácii.
Dátový analytický prístup
Štatistické analýzy sa uskutočnili pomocou SPSS (v. 16 a v. 21; SPSS Inc., Chicago IL). Okamžitá behaviorálna odozva na CS bola hodnotená z hľadiska výčnelkov nosa do otvoru, kde bola sacharóza vydávaná. Priemerný počet nosov sa počas tohto intervalu vystrelil (5 s na skúšku) a potom sa porovnal s priemerným počtom nosov v rovnakom trvaní (5 s) spriemerovaným v čase, keď CS chýbala. Skupina × Session ANOVA nosových výhonkov s prítomným CS a neprítomným sledovali získanie diskriminačných odpovedí na cue a nevyberané reakcie na nosný hluk podľa rôznych rozpisov počas tréningov 15 sacharózy.
Účinky liečby na lokomotorické odozvy sa hodnotili pomocou skupinových analýz ANOVA pre fázu návyku bez liečiva (tri sedenia), pred senzibilizáciou 0.5 mg / kg AMPH provokačnej dávky (jedna relácia) a počas piatich sedení 1 mg / kg AMPH ak sa očakáva, že sa skupiny budú líšiť v reakcii na opakované dávky AMPH. Skupina x Session ANOVA tiež hodnotili lokomotorické odpovede bez liečiva počas fázy navyknutia 30-min pred injekciou z každej testovacej relácie AMPH. Plánované porovnania hodnotili rozdiel v priemernom výkone pre skupinu 50 v porovnaní so skupinou 0 (žiadna kontrola očakávanej doby) a skupinou 100 (bez kontroly neistoty) pomocou t- testy (Howell, 1992) s použitím chybových výrazov MS a df pre relevantný efekt (tj interakcia skupiny alebo skupiny × relácie) z ANOVA (Víťaz, 1971). Polynomiálne analýzy trendov testovali profil zmien v priebehu relácií.
Aby sa určilo, či prístupové odpovede v prítomnosti a neprítomnosti CS počas tréningov 15 sacharózy prispeli k variácii lokomotorickej odpovede na AMPH, alebo k sprostredkovaným skupinovým rozdielom v odpovedi AMPH, na AMPH sa uskutočnili následné analýzy kovariancie (ANCOVA). lokomotorické dáta, vrátane totálneho nosu, sa strkajú (súčet pre 15 relácie), keď CS chýba ako kovariancia. Významný účinok kovariancie by naznačoval, že reakcie bezdrogového prístupu sa zmiernili (ovplyvnili silu) účinkov skupiny alebo relácie. Zníženie významu účinkov skupiny alebo relácie v prítomnosti významnej kovariancie by naznačovalo, že prístupy sprostredkované (započítané) k účinkom skupiny alebo relácie. Zníženie významu skupinových alebo session efektov v neprítomnosti významného kovariantného efektu by jednoducho odrážalo stratu štatistickej sily v dôsledku prerozdelenia df z chybového výrazu do kovariantu a nemalo by vplyv na interpretáciu efektov skupiny alebo relácie.
výsledky
Nos sa v priebehu sacharózovej kondicionácie stretáva
CS prítomný, figúra 1A ukazuje stredný nos pokes pre skupiny 25, 50 a 100, zatiaľ čo CS bol prítomný na 15 sacharózovej kondicionovacej relácii (nos pokes nebol kódovaný pre skupinu 0, ktorá nedostala CS). Skupina 3 × 15 Session ANOVA priniesla významné hlavné účinky skupiny, F(2, 21) = 5.63, p = 0.011 a relácia, F(14, 294) = 14.00, p <0.001 spolu s významnou interakciou skupina × relácia, F(28, 294) = 2.93, p <0.001. Obrázok 1A poukazuje na to, že hlavný efekt Session odráža zvýšenie počtu nosov v sedení vo všetkých troch skupinách a hlavný efekt skupiny odráža všeobecne vyššie celkové skóre v skupine 100 vs. skupina 25 s priebežným skóre v skupine 50. Významná skupina × Interakcia relácie pre kubický trend, F(2, 21) = 4.42, p = 0.030, indikoval rýchly vzostup, pokles a vyrovnanie v nose sa striekali cez sedenia v skupine 100, ako proti lineárnemu nárastu počas relácií v skupine 50 a plytší lineárny nárast v reláciách v skupine 25.
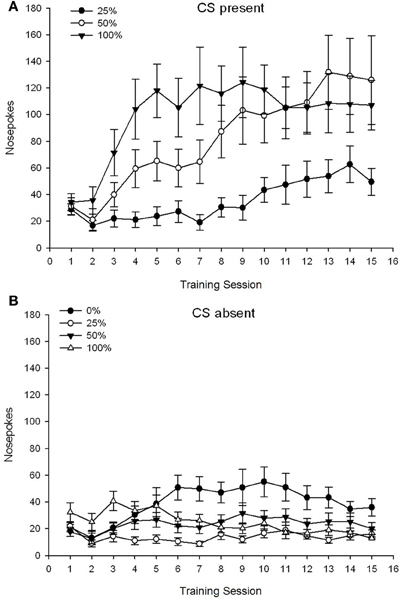
Obrázok 1. Stredné (SE) prístupové reakcie (nos pokes) na 15 tréningu sacharózy v skupinách potkanov Sprague Dawley (n = 8 / group) vystavená sacharózovej odmene (riešenie 10%) dodanej v premenných 0, 25, 50 alebo 100%, Podmieneným stimulom bolo svetlo (120 s). Skupina 0 dostala rovnaký počet odmien ako skupina 100 v neprítomnosti podmienených podnetov. (A) Skóre, keď bol CS prítomný (5 s × 15 skúšky). (B) Skóre, keď CS chýbala (priemer pre 5 × 15, zatiaľ čo svetlo bolo vypnuté).
CS chýba, figúra 1B ukazuje stredný nos pokes pre všetky štyri skupiny po ekvivalentné trvanie (5 s × 15 štúdie) spriemerované v čase, keď CS chýbala. Skupina 4 × 15 Session ANOVA priniesla významné hlavné účinky skupiny, F(3, 28) = 7.06, p = 0.001 a relácia F(14, 392) = 2.84, p <0.001 spolu s významnou interakciou skupina × relácia, F(42, 392) = 3.93, p <0.001. Významná interakcia skupiny × relácie pre kvadratický trend, F(3, 28) = 3.91, p = 0.019, bez interakcie pre kubický trend, F(3, 28) <0.93, p > 0.44, odráža profil „obráteného U“ nosa, ktorý trčí nad reláciami v skupine 0, oproti všeobecne stabilnému profilu počas relácií v ostatných skupinách.
Návyky na pohybové komory
Skupina 4 × 3 Session ANOVA priniesla hlavný efekt relácie, F(2, 56) = 5.67, p = 0.006 a žiadne iné významné účinky, F(3, 28) <1.60, p > 0.21. Priemerný (SE) zlom lúča za 2 h v lokomotorických boxoch bol 1681 (123) v relácii 1, 1525 (140) v relácii 2 a 1269 (96) v relácii 3. Plánované porovnania nezistili významné rozdiely medzi skupinou 50 a skupinou 0 alebo skupina 100 na prvom alebo poslednom zvykovom sedení, t(84) <1.69, p > 0.05. Takže pri absencii AMPH bolo opakované vystavenie testovacím boxom spojené s konzistentným poklesom spontánnej pohybovej aktivity v štyroch skupinách (tj. Efekt relácie) a bez rozdielovej odpovede v závislosti od tréningového harmonogramu sacharózy (bez interakcie) .
Testovacie stretnutia
Účinky predcitlivenia 0.5 mg / kg AMPH provokačná dávka.
Pohyb pred vstrekovaním, Jednosmerná ANOVA lokomotorickej odozvy skupiny 4 v priebehu fázy pred podaním injekcie 30-min nepriniesla žiadne významné účinky, F(3, 28) <1.05, p > 0.38. Plánované porovnania nezistili žiadny významný rozdiel medzi skupinou 50 a skupinou 0 alebo skupinou 100, t(32) <0.87, p > 0.40. Preto základné rozdiely v pohyblivosti pred injekciou nezohľadňovali skupinové rozdiely v pohybovej odpovedi na AMPH. Stredné (SE) zlomy lúča pre vzorku boli 559 (77).
Pooperačná lokomócia vs. finálna návyková návyk bez liečiva. Relácia ANOVA pri skupine 4 × 2 relácie porovnávala lokomočné reakcie skupín pri poslednej relácii návyku a bezprostredne po provizibilizácii 0.5 mg / kg AMPH. Skóre pre návykovú reláciu (120 minút) sa upravili tak, aby zodpovedali dĺžke testovacej relácie AMPH (90 minút) (hrubé návykové skóre × 90/120). Analýza priniesla významný hlavný efekt relácie, F(1, 28) = 34.16, p <0.001 a žiadne ďalšie významné účinky, F(3, 28) <2.26, p > 0.10. Efekt Session odrážal zvýšenie priemerných (SE) zlomov lúča v reakcii na dávku, z 952 (72) na 1859 (151). Plánované porovnania nezistili žiadne významné rozdiely medzi skupinou 50 a skupinou 0 alebo skupinou 100 v reakcii na dávku, t(56) <1.72, p > 0.10. Poradie skóre skóre zlomenia lúča (M; SE) však bolo v súlade s hypotézou: skupina 50 (2205; 264)> skupina 0 (2025; 203)> skupina 100 (1909; 407)> skupina 25 (1296; 299) .
Účinky 1 mg / kg AMPH.
Pohyb pred vstrekovaním, Skupina 4 × 5 Session ANOVA lokomotorickej odozvy počas fázy navyknutia 30-min pred podaním injekcie pri testovacích reláciách 1 mg / kg AMPH priniesla hlavný účinok Session, F(4, 112) = 43.64, p <0.0001 a žiadne ďalšie významné účinky, F(3, 28) <0.97, p > 0.42. Plánované porovnania nezistili žiadny významný rozdiel medzi skupinou 50 a skupinou 0 alebo skupinou 100 na prvom alebo poslednom testovacom sedení, t(140) <0.84, p > 0.30. Preto základné rozdiely v lokomócii nezohľadňovali skupinové rozdiely v lokomočnej odpovedi na AMPH. Priemerné skóre prerušenia lúča (SE) pre fázu návyku pred dávkou v reláciách 1–5 bolo: 454 (30), 809 (53), 760 (36), 505 (35), 756 (39).
Pohyb po injekcii, figúra 2 ukazuje účinky piatich injekcií 1 mg / kg AMPH (jedenkrát týždenne) na skóre lokomotorickej aktivity v štyroch skupinách. Skupina 4 × 5 Session ANOVA priniesla hlavný efekt relácie, F(4, 112) = 8.21, p <0.001, okrajový hlavný efekt skupiny, F(2, 45) = 3.28, p = 0.085 a žiadna významná interakcia, F(12, 122) <0.77, p > 0.68.
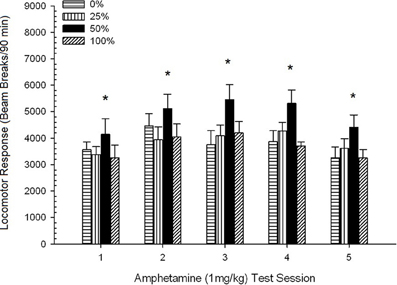
Obrázok 2. Priemerná (SE) lokomotorická odpoveď (počet zlomov lúča v elektronickom poli na 90 min) na 1 mg / kg d-amfetamínu (ip) na týždenných reláciách 5 v skupinách potkanov Sprague Dawley (n = 8 / group) predtým vystavené denným kondicionačným reláciám 15 s odmenou sacharózy (riešenie 10%) dodané pod premennými 0, 25, 50 alebo 100%, Podmieneným stimulom bolo svetlo (120 s). Skupina 0 dostala rovnaký počet odmien ako skupina 100 v neprítomnosti podmienených podnetov. *p <0.05 pre stredný rozdiel medzi skupinou 50 a skupinou 0, ako aj skupinou 100, na základe plánovaných porovnaní.
Plánované porovnania ukázali, že skupina 50 sa výrazne líšila od skupiny 0, t(14) = 2.19, p = 0.037 a skupina 100, t(14) = 2.36, p = 0.025 [a okrajovo sa líšil od skupiny 25, t(14) = 2.03, p = 0.051]. V skupine 50 teda lokomotorická odozva na 1 mg / kg AMPH spoľahlivo prevyšovala u ostatných troch skupín vo všetkých piatich testovacích reláciách. Analýza polynómneho trendu zistila výrazný kvadratický trend naprieč reláciami, F(1, 28) = 32.47, p <0.0001 a žiadne ďalšie významné trendy, F(1, 28) <1.78, p > 0.19. Obrázok 2 ukazuje, že tento výsledok odráža „invertovaný U“ vzor v reláciách.
Kontrola variácie v nose poke reagujúca počas tréningu sacharózy
Následná ANCOVA lokomotorických odpovedí na 1 mg / kg AMPH, s nosom pokes (CS prítomný) ako kovariát, v troch skupinách, ktoré dostali CS, priniesla marginálny hlavný účinok skupiny, F(2, 20) = 3.07, p = 0.069 a žiadne významné účinky súvisiace s kovarianciou, F(4, 80) <0.05, p > 0.85. Teda tlmený prístup reagujúci počas tréningu nevysvetlil významné variácie v pohybovej reakcii na 1 mg / kg AMPH v skupinách 25, 50 alebo 100.
Následná ANCOVA lokomotorických odpovedí na 1 mg / kg AMPH, s nosom pokes (CS chýba) ako kovarianciou, priniesla významný účinok kovariancie, F(1, 27) = 6.17, p = 0.020, významný hlavný efekt skupiny, F(3, 27) = 4.13, p = 0.016, okrajová relácia × interakcia s kovovarmi, p = 0.080 a žiadne iné významné účinky, F(4, 108) <1.48, p > 0.21. Takže nerozumný (nevyberavý) prístup reagujúci na tréning vysvetlil významné variácie v pohybovej reakcii na 1 mg / kg AMPH. Táto variácia sa však neprekrývala s rozptylom súvisiacim so skupinou, pretože zahrnutie kovariátu do analýzy skôr zvýšilo ako znížilo významnosť skupinového účinku.
Diskusia
Údaje o nosnom čreve počas prítomnosti CS ukazujú, že skupiny nadobudli asociáciu medzi dodaním CS a sacharózy, čo sa prejavilo zvýšením odpovedí v priebehu tréningu. Profil reakcie počas sedení, kým bol prítomný CS, naznačil, že plány 100 a 50% CS-US boli rovnako účinné pri vyvolaní prístupu, zatiaľ čo plán 25% vyvolal miernejší nárast prístupu vyvolaného cue. Údaje o nosnom nose, zatiaľ čo CS chýbala, naznačujú, že skupiny, ktoré dostali ktorýkoľvek z troch tréningových režimov CS-sacharózy (skupina 25, 50, 100), sa rýchlo naučili redukovať nos v dôsledku neprítomnosti CS, zatiaľ čo zvieratá v skupine 0 , ktorý nedostal CS, sa naučil znižovať svoje prístupové správanie v obmedzenom rozsahu po rozsiahlom tréningu.
Údaje o návyku ukazujú, že skupiny sa pred AMPH nelíšili a že opakovaná expozícia testovacím boxom bola spojená so zníženou lokomotorickou odpoveďou bez liečiva. Rozdiely medzi skupinami a zvýšená odpoveď na opakované dávky AMPH preto nemôžu byť pripisované už existujúcim rozdielom v lokomotorickom správaní.
Výsledky predsenzibilizačného testu s 0.5 mg / kg AMPH potvrdili, že liek zvyšuje lokomotorickú aktivitu v porovnaní s konečným dňom bez návyku na liečivo. V súlade s hypotézou sa skupina 50 umiestnila vyššie ako skupiny 0 alebo 100 (rovnako ako skupina 25), pokiaľ ide o priemernú odpoveď na dávku, hoci priemerné rozdiely medzi skupinami neboli významné.
Pokiaľ ide o senzibilizačné sedenia, plánované porovnania medzi skupinami ukázali, že predchádzajúca expozícia 50% podmienenej sacharózovej odmene viedla k významnému zvýšeniu pohybovej odpovede na dávku 1.0 mg / kg amfetamínu v porovnaní s ostatnými tromi režimami. Tento účinok bol zrejmý od prvej dávky a pri opakovaných dávkach sa výrazne nezmenil. Analýza trendu ukázala dvojfázovú odpoveď (pre celú vzorku) na opakované dávky AMPH, zvyšujúca sa až po tretiu dávku a potom klesajúca. Výsledky následného sledovania ANCOVA s vystrčenými nosmi (CS neprítomné) ako kovariátom potvrdili, že rozdiely v pohybových odpovediach štyroch skupín na 1 mg / kg AMPH neboli sprostredkované nekontrolovaným prístupom reagujúcim počas tréningov na sacharózu.
Skupinový účinok počas relácie senzibilizácie je v súlade s našou hypotézou. Bifázický efekt relácie nie je konzistentný s očakávanou pokračujúcou eskaláciou lokomotorických odpovedí s opakovanými dávkami AMPH. Môže to súvisieť s dávkovacím intervalom. Na riešenie tohto problému by sa mal použiť postup (alternatívne denné dávky), ktorý preukázal konzistentnú eskaláciu lokomotorickej odpovede na dávky 1.0 mg / kg AMPH (tj behaviorálna senzibilizácia). Vplyv senzibilizačného režimu AMPH na následnú reakciu na druhú výzvu 0.5 mg / kg by ďalej podporoval všeobecnosť tohto účinku. Začlenenie podania fyziologického roztoku pred AMPH by určilo úlohu očakávaných alebo injekčných (napr. Stresových) účinkov na lokomotorickú odpoveď na AMPH. Zahrnutie skupiny 75% kondicionovanej sacharózy by pomohlo objasniť úlohu neistoty v odmeňovaní v porovnaní s odmenou odmeny na modeli odpovedí pre skupiny 50 a skupinu 25. Okrem toho, aby sa umožnilo posúdenie (pomocou ANCOVA) príspevku odozvy bez prístupu k liekom na lokomóciu pod AMPH (s použitím nosa, ktorý sa vystríha s CS prítomným ako kovariát), boli nosové pokes kódované aj pre skupinu 0 počas intervalu, keď CS bol prítomný v ďalších štyroch skupinách (tj tak, že nos strká zo všetkých piatich skupín - vrátane skupiny 0, ktorá nedostala CS - by mohol byť zahrnutý do analýzy kovariancie s CS prítomným ako kovariát). Tieto vylepšenia boli začlenené do experimentu 2.
experiment 2
Materiály a metódy
Metodológia experimentu 2 bola podobná ako pri experimente 1, ale revidovaná tak, aby sa lepšie priblížila režimu, ktorý bol zistený ako spoľahlivo indukujúci senzibilizáciu AMPHFletcher a kol., 2005). Zmeny boli nasledovné: (a) Skupina 75% CS-sacharóza (n = 8); (b) Počas tréningu sacharózy dostávali potkany (okrem skupiny 0) 20 CS (svetlé) prezentácie (na rozdiel od 15 v experimente 1); (c) CS prezentácie boli oddelené priemerným inter-trial intervalom 90 s; rozsah: 30 – 180 s (vs. 120 s v experimente 1), ktorý kompenzoval nárast tréningových testov tak, aby sa dala porovnať dĺžka trvania každého tréningu s experimentom 1; (d) trvanie každej z troch habituačných sedení bolo znížené z 120 na 90 min, aby zodpovedalo dĺžke trvania testovania; (e) Pridal sa fyziologický roztok (ip, 1 ml / kg) (90 min) (deň po sacharóze 8), aby sa vyhodnotili lokomotorické účinky injekcie. sama o sebe (napr. očakávania, stres); (f) Senzibilizačné sedenia 1 mg / kg sa konali striedavo v pracovných dňoch (dni po tréningu 12 – 21) namiesto v týždenných intervaloch ako v experimente 1; (g) Spolu s predcitlivením 0.5 mg / kg AMPH provokačnou dávkou (po tréningu 9) bola pridaná druhá post-senzibilizačná dávka 0.5 mg / kg AMPH (po tréningovom dni po sacharóze 28), aby sa otestovala generálnosť. senzibilizačný účinok naprieč dávkami; (h) nos sa v priebehu prítomnosti CS kódoval pre všetky skupiny (vrátane skupiny 0); (i) nos sa strká, zatiaľ čo CS nie je prítomný, sa zaznamenáva špecificky z intervalu 5-u bezprostredne pred nástupom CS do indexu predčasného prístupu odpovedajúceho.
výsledky
Nos sa v priebehu sacharózovej kondicionácie stretáva
Skupina 5 × 15 Session × 2 Phase (prítomná CS, CS chýba) ANOVA nosa pokes priniesla významné hlavné účinky skupiny, F(4, 19) = 2.89, p = 0.050, relácia F(14, 266) = 2.28, p = 0.006 a Fáza, F(1, 19) = 14.72, p = 0.001, ako aj významnú trojstrannú interakciu, F(56, 266) = 1.38, p = 0.050. panely (A, B) obr 3 vynesie sa stredné skóre skokov v nose pre skupiny pre prítomnú CS a pre neprítomnú fázu. Porovnanie týchto dvoch panelov odhalilo, že hlavný účinok fázy odrážal viac celkových odpovedí na nose poke, keď bola prítomná CS, alebo neprítomná. Preto sa hromadné odpovede vyskytovali významne častejšie ako predčasné neodrážané odpovede. Hlavné účinky skupín a relácií neboli ľahko interpretované kvôli interakcii vyššieho rádu. Tento posledný výsledok odrážal konvergenciu skóre pre päť skupín na relatívne stabilnej nízkej úrovni naprieč reláciami, keď chýbala CS (obrázok) 3B), spolu s divergenciou skóre do vysokých (skupina 75, skupina 100), medziproduktov (skupina 50) a nízkych (skupina 0, skupina 25) hladín nosa poke odpovedajúcich počas sedení, keď bol prítomný CS (obrázok 3A). Z trendov polynómu nižšieho rádu (lineárny, kvadratický, kubický) sa priblížil iba trojsmerný interakcie pre lineárny trend. F(4, 19) = 2.32, p = 0.094, odrážajúc všeobecne monotónny nárast nosa, ktorý sa prevyšuje počas sedení v skupine 75 a relatívne rýchlejšiu stabilizáciu na vysokých, stredných a nízkych úrovniach odpovedí v ostatných skupinách, keď bol CS prítomný.
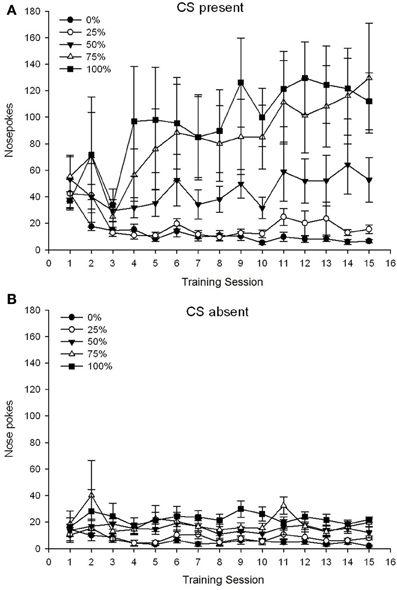
Obrázok 3. Stredné (SE) prístupové reakcie (nos pokes) na 15 tréningu sacharózy v skupinách potkanov Sprague Dawley (n = 8 / skupina) vystavená sacharózovej odmene (riešenie 10%) dodanej v premenných 0, 25, 50, 75 alebo 100%, Podmieneným stimulom bolo svetlo (120 s). Skupina 0 dostala rovnaký počet odmien ako skupina 100 v neprítomnosti podmienených podnetov. (A) Skóre, keď bol CS prítomný (5 s × 20 skúšky). (B) Skóre, keď CS chýbala (priemer pre 5 × 20, zatiaľ čo svetlo bolo vypnuté).
Návyky na lokomotorické boxy
Skupina 5 × 3 Session ANOVA lokomotorických reakcií bez liečiva priniesla významný hlavný účinok Session, F(2, 70) = 60.01, p <0.0001 a žiadne ďalšie významné účinky, F(4, 35) <0.70, p > 0.60. Plánované porovnanie skupiny 50 so skupinou 0 a so skupinou 100 na prvom a poslednom sedení neprinieslo žiadne významné účinky, tje <0.84, p > 0.40. Preto sa priemerná pohybová reakcia bez liečiva v kľúčových skupinách pred testovaním nelíšila. Priemerný (SE) počet zlomov lúčov za 90 minút bol 2162 118 (1) v relácii 1470, 116 (2) v relácii 1250 a 98 (3) v relácii XNUMX.
Testovacie stretnutia
soľný, Skupina 5 × 2 Session ANOVA porovnávala lokomotorickú odozvu na konečnom sedení na návyku a relácii vyvolanej fyziologickým roztokom. ANOVA priniesla hlavný účinok Session, F(1, 35) = 62.46, p <0.0001 a žiadne ďalšie významné účinky, F(4, 35) <0.65, p > 0.64. Obrázok 4 Znázorňuje, že Session efekt odráža celkový pokles lokomotorickej odozvy z posledného návyku bez návyku na liečivo na fyziologický roztok, ktorý sa nelíši podľa skupiny. Pokles lokomotorickej odozvy pozorovaný počas troch sedení na návykoch teda pokračoval štvrtou expozíciou testovacích boxov bez liečiva.
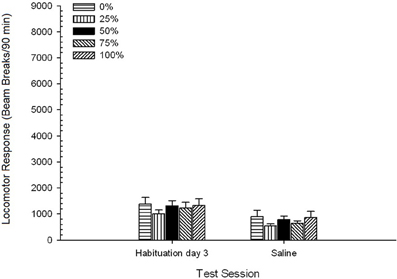
Obrázok 4. Priemerná (SE) lokomotorická odozva (počet lámaných lúčov v elektronickom poli na 90 min.) Na poslednom z 3 návykov bez návykových látok a na nasledujúcom sedení po injekcii fyziologického roztoku (ip, 1 ml / kg) v skupinách Sprague Dawley potkany (n = 8 / group) predtým vystavené denným kondicionačným reláciám 15 s odmenou sacharózy (riešenie 10%) dodané pod premennými 0, 25, 50, 75 alebo 100%, Podmieneným stimulom bolo svetlo (120 s). Skupina 0 dostala rovnaký počet odmien ako skupina 100 v neprítomnosti podmienených podnetov.
Účinky 0.5 mg / kg AMPH.
Pohyb pred vstrekovaním, Skupina 5 × 2 Session ANOVA pre-injekcie lokomócie (30-min) na testovacie dni pred a po senzibilizácii 0.5 mg / kg AMPH testovaných dní priniesla významný hlavný účinok Session, F(1, 35) = 13.39, p = 0.001 a žiadne iné významné účinky, F(4, 35) <1.79, p > 0.15. Plánované porovnania nezistili pri prvej relácii významné rozdiely medzi skupinou 50 a skupinou 0 alebo skupinou 100, t(70) <1.00, p > 0.30. Avšak v druhej (post-senzibilizačnej) relácii skupina 50 (1203; 121) vykazovala významne viac zlomov lúčov pred injekciou (M; SE) ako v skupine 100 (756; 103), t(70) = 5.11, p <0.001, ale nelíšili sa od skupiny 0 (1126; 211), t(7) <0.88, p > 0.40. Preto základné rozdiely v lokomócii nezohľadňovali skupinové rozdiely v pohybovej odpovedi na prvú dávku 0.5 mg / kg AMPH, ale mohli prispieť k rozdielom medzi skupinou 50 a skupinou 100 v pohybovej odpovedi na druhú dávku 0.5 mg / kg AMPH . Priemerný (SE) zlom lúča pre predinjekčnú fázu na prvom a druhom testovacom sedení 0.5 mg / kg AMPH bol 757 (41) a 974 (59).
Pohyb po injekcii, Skupina 5 × 2 Session ANOVA lokomotorickej odpovede na 0.5 mg / kg AMPH pred a po režime senzibilizácie na dávku 5 priniesol hlavný účinok Session, F(1, 35) = 76.05, p <0.0001 a žiadne ďalšie významné účinky, F(4, 35) <1.10, p > 0.37. Obrázok 5 ukazuje priemerné skóre pre každú skupinu a reláciu.
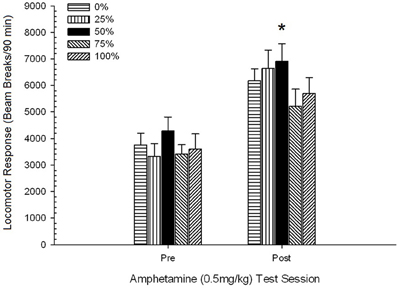
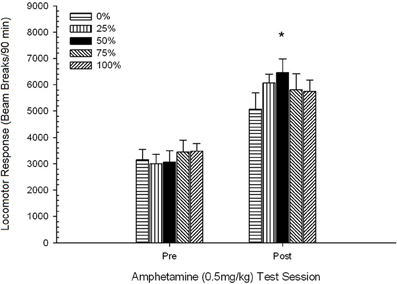
Obrázok 5. Priemerná (SE) lokomotorická odpoveď (počet lámaných lúčov v elektronickom poli na 90 min) na 0.5 mg / kg d-amfetamínu na oddelených sedeniach pred a po 5-senzibilizačnom režime d-amfetamínu (1.0 mg / kg; ip v skupine potkanov Sprague Dawley (n = 8 / group) predtým vystavené denným kondicionačným reláciám 15 s odmenou sacharózy (riešenie 10%) dodané pod premennými 0, 25, 50, 75 alebo 100%, Podmieneným stimulom bolo svetlo (120 s). Skupina 0 dostala rovnaký počet odmien ako skupina 100 v neprítomnosti podmienených podnetov. *p <0.05 pre stredný rozdiel medzi skupinou 50 a skupinou 0, ako aj skupinou 100, na základe plánovaných porovnaní.
Obrázok ukazuje, že Session efekt zahŕňal výrazné zvýšenie celkového priemerného (SE) lúča prerušenia na 90 min od dávky 0.5 mg / kg 1, 3674 (216) na 0.5 mg / kg dávky 2, 6123 (275). Nedostatok interakcie alebo skupinového efektu naznačil, že senzibilizácia na AMPH sa v skupinách spoľahlivo nemenila. Napriek tomu, že v štúdii ANOVA chýbajú významné účinky na skupinu, kontrola na obrázku ukazuje, že skupina 50 vykazovala najväčšiu odpoveď na prvú aj druhú dávku 0.5 mg / kg. Plánované porovnania odpovede na prvú dávku 0.5 mg / kg neodhalili signifikantný rozdiel medzi skupinou 50 a skupinou 0 alebo skupinou 100, t, s(35) <0.48, p > 0.50. Avšak v reakcii na druhú (post-senzibilizáciu) dávku 0.5 mg / kg vykazovala skupina 50 významne väčšiu lokomóciu ako skupina 0, t(35) = 2.00, p <0.05, rovnako ako skupina 100, t(35) = 3.29, p <0.01.
Vo svetle významného skupinového rozdielu v lokomócii pred podaním injekcie v druhej relácii 0.5 mg / kg AMPH uvedenej vyššie bola vykonaná následná kontrola 5 Group × 2 Session ANCOVA lokomotorickej odozvy na 0.5 mg / kg AMPH. injekcie na druhom zasadnutí. Táto analýza priniesla významný vplyv kovariancie, F(1, 34) = 8.65, p = 0.006, hlavný efekt relácie F(1, 34) = 10.83, p = 0.002 a žiadne iné významné účinky, F(4, 34) <0.85, p > 0.50. Dôležité je, že plánované porovnania založené na chybe MS a chybe df z ANCOVA potvrdili, že stredná pohybová odpoveď na druhú dávku 0.5 mg / kg AMPH zostala významne vyššia v skupine 50 ako v skupine 100, t(34) = 3.09, p <0.01 a skupina 0, t(34) = 1.88, p <0.05 (jednostranný), keď bola kontrolovaná variácia pred injekciou od relácie 2. Skupina 50 teda vykazovala významne vyššiu post-senzibilizačnú lokomočnú odpoveď na 0.5 mg / kg AMPH ako skupina 100 alebo skupina 0 a tieto rozdiely v skupine neboli sprostredkované predinjekčnou lokomóciou v testovacie dni.
Účinky 1.0 mg / kg AMPH.
Pohyb pred vstrekovaním, Skupina 5 × 5 Session ANOVA skóre pred injekciou 30-min pre relácie senzibilizácie 1 mg / kg AMPH priniesla hlavný účinok Session, F(4, 140) = 16.70, p <0.0001 a žiadne ďalšie významné účinky, F(4, 35) <0.94, p > 0.45. Plánované porovnania nezistili pri prvej relácii signifikantný rozdiel v lokomócii pred injekciou medzi skupinou 50 a skupinou 0 alebo skupinou 100, t(175) <1.66, p > 0.10. Na poslednom zasadnutí však skupina 50 (1167; 140) vykazovala významne viac zlomov lúčov (M; SE) ako skupina 100 (1000; 99), t(175) = 2.35, p <0.05, ale nelíšili sa od skupiny 0 (1085, 120), t(175) <1.16, p > 0.20. Preto rozdiely v predinjekčnej lokomócii prispeli k rozdielom medzi skupinami 50 a 100 v lokomočnej odpovedi na konečnú dávku 1 mg / kg AMPH. Priemerné (SE) celkové prerušenia lúča pre vzorku počas fázy pred injekciou pre relácie 1 až 5 boli: 810 (46), 784 (52), 760 (53), 726 (46), 1009 (51).
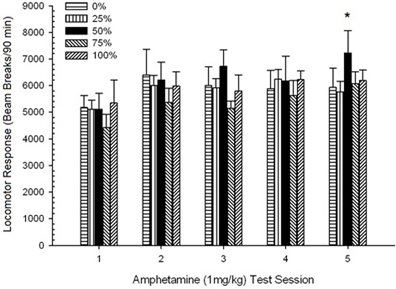
Pohyb po injekcii, Skupina 5 × 5 Session ANOVA odpovedí na 1 mg / kg AMPH priniesla významný hlavný účinok Session, F(4, 140) = 6.72, p <0.001, okrajová interakcia skupina × relácia, F(16, 140) = 1.57, p = 0.085 a žiadny hlavný efekt skupiny, F(4, 35) <0.44, p > 0.77. Analýzy polynomiálnych trendov odhalili významný lineárny trend, F(1, 35) = 9.19, p = 0.005 a kubický trend, F(1, 35) = 21.63, p <0.001, v priebehu relácií 1 až 5. Obrázok 6 ukazuje priemerné lokomotorické skóre pre každú skupinu a reláciu.
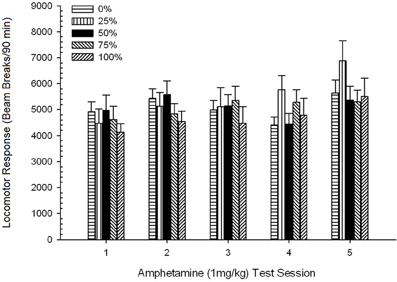
Obrázok 6. Priemerná (SE) lokomotorická odpoveď (počet zlomov lúča v elektronickom poli na 90 min) na 1 mg / kg d-amfetamínu (ip) na týždenných reláciách 5 v skupinách potkanov Sprague Dawley (n = 8 / group) predtým vystavené denným kondicionačným reláciám 15 s odmenou sacharózy (riešenie 10%) dodané pod premennými 0, 25, 50, 75 alebo 100%, Podmieneným stimulom bolo svetlo (120 s). Skupina 0 dostala rovnaký počet odmien ako skupina 100 v neprítomnosti podmienených podnetov.
Obrázok ukazuje, že Session efekt odrážal významný nárast celkových priemerných (SE) lámacích prerušení pre celú vzorku z relácie 1, 4624 (213) na reláciu 5, 5736 (272), čo potvrdzuje vznik senzibilizácie na AMPH. Kubický trend označoval relatívne maximá na reláciách 1, 3 a 5, s poklesmi na reláciách 2 a 4, najmä pre skupiny 0 a 50. Obrázok tiež ukazuje, že napriek nedostatku významnej interakcie, skupina 25 vykazovala postupne väčšie lokomotorické reakcie počas relácií a značne sa líšila od ostatných skupín na reláciách 4 a 5 (9 a 22% väčšie, než nasledujúca najvyššia skupina). Plánované porovnania zistili, že skupina 50 sa významne nelíšila od skupín 0 alebo 100, t(175) <0.89, p > 0.40 na prvom alebo poslednom testovacom stretnutí AMPH 1 mg / kg.
Kontrola variácie v nose poke reagujúca počas tréningu sacharózy
Dve 5 Group × 2 Session ANCOVA lokomotorickej odozvy na 0.5 mg / kg AMPH pred a po senzibilizačnom režime, vrátane totálneho nosa, sa počas tréningu sacharózy s CS prítomným a CS neprítomným ako separátnym kovariantom nezistili žiadne signifikantné účinky ani na jednu kovarianciu, F(1, 18) <1.03, p > 0.31. Preto prístup reagujúci počas tréningu nezmierňoval skupinové rozdiely v odpovedi na 0.5 mg / kg AMPH.
Dve skupiny 5 × 5 Sekcie ANCOVA lokomotorickej odpovede na 1 mg / kg počas senzibilizačných relácií s celkovým nosom sa strkajú (prítomnosť CS, CS neprítomná), pretože oddelené kovariáty nepriniesli žiadne významné účinky kovariancie, kým CS bola prítomná, F(4, 104) <1.04, p > 0.38 a okrajový hlavný účinok kovariátu, zatiaľ čo CS chýbala, F(1, 18) = 3.32, p = 0.085.
Diskusia
Výsledky tejto štúdie konzistentne nepodporovali hypotézu, že skupina 50 by preukázala vyššiu lokomotorickú odozvu počas sedení v porovnaní s inými skupinami. Údaje 1 mg / kg AMPH potvrdili vznik senzibilizácie pri alternatívnom režime dávkovania. Vzor v skupinách naznačoval tendenciu k väčšej senzibilizácii počas posledných sedení v skupine 25, bez takéhoto dôkazu pre skupinu 50. Na rozdiel od toho výsledky dávky 0.5 mg / kg ukázali trend zvýšenej senzibilizácie v skupine 50, pričom súčasne potvrdili významné celkové zvýšenie lokomotorickej odpovede v skupinách na druhú oproti prvej dávke 0.5 mg / kg AMPH. Nulový účinok injekcie fyziologického roztoku potvrdil, že očakávaná dĺžka alebo stres spojený s injekciou neprispeli k účinkom AMPH.
Údaje o nosovom trakte opäť odhalili celkový nárast prístupu reagujúci v priebehu tréningu, keď bol CS prítomný, bez zodpovedajúceho nárastu, keď CS nebola prítomná. Preto sa zdá, že zvieratá získali vzťah medzi CS a vyhliadkou na získanie sacharózy. Skupinové rozdiely vo frekvencii nosa sa strčia, keď bol CS prítomný, zodpovedali zhruba frekvencii poskytovania odmeny podľa príslušných plánov, pričom skupiny 75 a 100 zobrazovali najviac nosov, skupina 50 zobrazovala stredné čísla nosov a skupiny 0 a skupiny 25 a 50 zobrazuje najmenší nos. Tieto výsledky naznačujú, že CS prišiel kontrolovať prístup, ktorý je v súlade s celkovou pravdepodobnosťou odmeny. Hoci špekulatívne, jedným z možných vysvetlení nižších frekvencií nosného poke s CS prítomným v skupine 2 v experimente 1 verzus experiment 1 môže byť skrátenie intervalu medzi skúškami, pretože dlhšie intervaly medzi pokusmi (experiment XNUMX) podporujú impulzívny impulz. súvisí so zvýšeným obratom DA pri prednom cinguláte, prelimbických a infralimbických kortikách (Dalley a kol., 2002). Zníženie intervalu medzi pokusmi v experimente 30 (a 2) v 3 môže mať preto zmenené hladiny kortikálneho DA a podporované selektívnejšie (tj riadené relatívnou frekvenciou odmeňovania) vs. v skupine 50 počas tréningových pokusov v experimente 2 v porovnaní s experimentom 1.
Absencia významných účinkov súvisiacich s kovarianciou pre nos sa vystríha v stave prítomného CS v ANCOVA, čo naznačuje, že prístup reagujúci počas tréningu sacharózy nesprostredkoval účinky rôznych schém CS-sacharózy na odpovede na AMPH. Okrajovo významný vplyv kovariancie na stav absencie CS v ANCOVA lokomotorických odpovedí na 1 mg / kg AMPH naznačuje, že tendencia k predčasnej reakcii bez liečiva vysvetlila niektoré z variability lokomotorických účinkov AMPH počas relácií senzibilizácie.
Dôkazy svedčia o tom, že účinky kondicionačnej anamnézy môžu byť viac rozoznateľné u 0.5 AMPH ako u 1 mg / kg AMPH a že protokol, ktorý vyvoláva senzibilizáciu v prípade absencie akejkoľvek inej manipulácie, môže zatemniť alebo spôsobiť redundanciu účinkov putatívnej liečby. behaviorálna manipulácia podporujúca senzibilizáciu (tj chronická premenlivá odmena).
Behaviorálna senzibilizácia na AMPH je robustný účinok v laboratóriu. Avšak mimo laboratória len menšina jedincov, ktorí hazardujú, chronicky eskalovala na patologické úrovne. Hoci riziko senzibilizácie súvisí s rizikom závislosti (alebo vyhľadávaním liekov), najmä pre psychostimulanciá (Vezina, 2004; Flagel a kol., 2008), mnoho faktorov, okrem rizika senzibilizácie, môže predisponovať k závislosti (napr. Verdejo-Garcia a kol., 2008; Conversano a kol., 2012; Volkow a kol., 2012). Faktory, ktoré spôsobujú zraniteľnosť voči senzibilizácii, však môžu interagovať s históriou kondicionovania, aby sa zvýraznili účinky nepredvídateľnej odmeny (napr. Plán 50% CS-US) na reaktivitu systému DA. Na preskúmanie tejto možnosti experiment 3 použil rovnaký postup ako experiment 2, ale namiesto potkanov kmeňa Sprague Dawley použil kmeň Lewis.
Potkany Sprague Dawley vykazujú stredné hladiny transportérov DA s nižšími hladinami ako potkany kmeňa Wistar (Zamudio a kol., 2005), ale vyššie hladiny ako potkany Wistar-Kyoto („depresívny“ kmeň podobný kmeňu) v nucleus accumbens, amygdale, ventrálnej tegmentálnej oblasti a substantia nigra (Jiao a kol., 2003). Tento profil môže spôsobiť, že potkany Sprague Dawley budú len mierne citlivé na environmentálne alebo farmakologické manipulácie funkcie DA. Na rozdiel od toho potkany Lewis vykazujú nízke hladiny DA transportérov, ako aj D2 a D3 DA receptorov v nucleus accumbens a dorzálnej striatum v porovnaní s inými kmeňmi (napr. F344) (Flores a kol., 1998). Tieto morfologické rozdiely môžu prispieť k diferenciálnej reakcii potkanov Lewis na manipulácie s DA. Potkany Lewis tiež vykazujú rad zvýraznených odpovedí na experimentálne manipulácie s liečivami v porovnaní s inými kmeňmi (napr. F344). Najdôležitejšie je, že potkany Lewis vykazujú väčšiu senzibilizáciu na metamfetamín, ktorá sa vyznačuje nízkou odpoveďou na počiatočné dávky, ale vyššou reakciou na neskoršie dávky (Camp et al., 1994). Lewisove potkany tiež vykazujú väčšiu lokomotorickú senzibilizáciu na celý rad dávok kokaínu (Kosten a kol., 1994; Haile a kol., 2001). Na základe tohto modelu účinkov sme usúdili, že Lewisove potkany nám umožnia skúmať, či citlivosť na senzibilizáciu zosilňuje účinky harmonogramu kondicionovania na následnú reakciu na AMPH.
experiment 3
Materiály a metódy
Metodológia bola rovnaká ako v experimente 2, okrem použitia Lewisových potkanov (200 – 225 g pri príchode, Charles River, Quebec, Kanada).
výsledky
Nos sa v priebehu sacharózovej kondicionácie stretáva
Skupina 5 × 15 Session × 2 Phase (prítomná CS, CS chýba) ANOVA nosa pokes priniesla významné hlavné účinky skupiny, F(4, 34) = 6.12, p = 0.001, relácia, F(14, 476) = 3.42, p <0.001 a fáza, F(1, 34) = 20.83, p <0.001, ako aj významná trojstranná interakcia, F(56, 476) = 1.56, p = 0.008. panely (A, B) obr 7 vynesie sa stredné skóre skokov v nose pre skupiny pre prítomnú CS a pre neprítomnú fázu. Porovnanie týchto dvoch panelov odhalilo, že hlavný účinok fázy odrážal viac celkových odpovedí na nose poke, keď bola prítomná CS, alebo neprítomná. Preto sa hromadné odpovede vyskytovali významne častejšie ako predzrelé odpovede. Hlavné účinky skupín a relácií neboli ľahko interpretované kvôli interakcii vyššieho rádu. Trojcestná interakcia odrážala konvergenciu skóre pre päť skupín na relatívne stabilnej nízkej úrovni naprieč reláciami, keď chýbala CS [Panel] (B)], spolu s divergenciou skóre, keď bol CS prítomný v relatívne diskrétnych profiloch pre každú skupinu, ktorá paralelne porovnávala ich poradie frekvencie odmien: od najvyššej (skupina 100) po najnižšiu (skupina 25) [Panel (A)]. Významný bol iba lineárny trend interakcie, F(4, 34) = 4.03, p = 0.009, odrážajúc všeobecne konzistentný vzrast nosa v priebehu sedenia v skupine 100, keď bol CS prítomný v porovnaní s relatívne nekonzistentným profilom nárastu nosa, ktorý sa v priebehu tejto fázy vyskytol v iných skupinách.
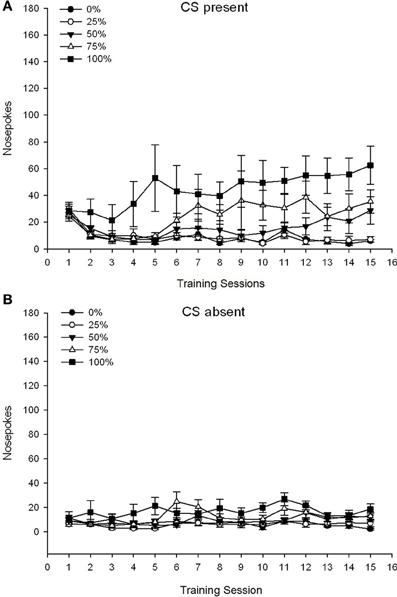
Obrázok 7. Stredné (SE) prístupové reakcie (nos pokes) na 15 sacharózových tréningoch v skupinách potkanov Lewis (n = 8 / skupina) vystavená sacharózovej odmene (riešenie 10%) dodanej v premenných 0, 25, 50, 75 alebo 100%, Podmieneným stimulom bolo svetlo (120 s). Skupina 0 dostala rovnaký počet odmien ako skupina 100 v neprítomnosti podmienených podnetov. (A) Skóre, keď bol CS prítomný (5 s × 20 skúšky). (B) Skóre, keď CS chýbala (priemer pre 5 × 20, zatiaľ čo svetlo bolo vypnuté).
Návyky na lokomotorické boxy
Skupina 5 × 3 Session ANOVA priniesla hlavný efekt relácie, F(2, 70) = 23.07, p <0.0001 a žiadne ďalšie významné účinky, F(8, 70) <1.47, p > 0.18. Krivočiary vzorec priemerných (SE) pohybových skóre sa objavil od relácie 1 1076 (74) cez reláciu 2 644 (48) až po reláciu 3 762 (59). Plánované porovnanie skupiny 50 so skupinou 0 a so skupinou 100 na prvých a posledných sedeniach bolo zistených významne menej zlomov lúčov v skupine 50 (M = 911; SE = 109) vs. skupina 0 (M = 1103; SE = 176) na návyku 1, t(105) = 2.02, p <0.05, ale žiadny rozdiel medzi skupinou 50 a skupinou 100 (M = 1066; SE = 150), t(105) <1.20, p > 0.20, počas tejto relácie. Skupina 50 sa významne nelíšila od skupiny 0 ani od skupiny 100 na poslednom návykovom stretnutí, t(105) <0.93, p > 0.30. Preto sa priemerná pohybová odpoveď bez liečiva v kľúčových skupinách pred testovaním konzistentne nelíšila.
Testovacie stretnutia
soľný, Skupina 5 × 2 Session ANOVA lokomotorických odpovedí na poslednom zasadnutí návyku a relácii testovania fyziologického roztoku priniesla významný hlavný účinok Session, F(1, 35) = 50.12, p <0.0001 a žiadne ďalšie významné účinky, F(4, 35) <0.57, p > 0.68. Obrázok 8 ukazuje priemerné skóre skupiny pre tieto dve sedenia a naznačuje, že účinok Session odrážal významný pokles od návyku na test fyziologického roztoku. Prijatie injekcie sama o sebe (napr. očakávaná dĺžka, stres) nezvyšovali lokomotorickú odpoveď.
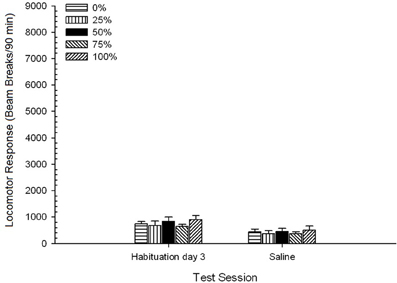
Obrázok 8. Priemerná (SE) lokomotorická odozva (počet lámaných lúčov v elektronickom poli na 90 min.) Na poslednom z 3 návykov bez návyku na liečivo a na nasledujúcom sedení po injekcii fyziologického roztoku (ip, 1 ml / kg) v skupinách potkanov Lewis (n = 8 / group) predtým vystavené denným kondicionačným reláciám 15 s odmenou sacharózy (riešenie 10%) dodané pod premennými 0, 25, 50, 75 alebo 100%, Podmieneným stimulom bolo svetlo (120 s). Skupina 0 dostala rovnaký počet odmien ako skupina 100 v neprítomnosti podmienených podnetov.
Účinky 0.5 mg / kg AMPH.
Pohyb pred vstrekovaním, Skupina 5 × 2 Session ANOVA pre-injekcie lokomócie priniesla významný hlavný účinok Session, F(1, 35) = 15.04, p <0.001 a žiadne ďalšie významné účinky, F(4, 35) <1.19, p > 0.33. Plánované porovnania nezistili žiadny signifikantný rozdiel medzi skupinou 50 a skupinou 0 alebo skupinou 100 ani pri jednom testovaní, t(70) <0.99, p > 0.30. Preto základné rozdiely v pohyblivosti pred injekciou nezohľadňovali skupinové rozdiely v pohybovej odpovedi na 0.5 mg / kg AMPH. Priemerný (SE) zlom lúča pre predinjekčnú fázu pre prvé a druhé (po senzibilizácii) 0.5 mg / kg sedenie bol 325 (25) a 473 (36).
Pohyb po injekcii, Skupina 5 × 2 Session ANOVA lokomotorickej odpovede na dávky 0.5 mg / kg podávané pred a po chronickej dávke 1 mg / kg AMPH priniesla hlavný účinok Session, F(1, 34) = 87.44, p <0.0001 a žiadne ďalšie významné účinky, F(4, 34) <0.94, p > 0.45. Obrázok 9 Znázorňuje priemerné lokomotorické skóre pre každú skupinu a sedenie a ukazuje, že účinok Session odrážal zvýšenú celkovú odpoveď na druhú dávku 0.5 mg / kg, čo je v súlade so senzibilizáciou. Obrázok tiež ukazuje, že skupiny na relácii 1 fungovali veľmi podobne, ale skupina 50 zobrazovala viac lokomotorickej aktivity ako ostatné skupiny na relácii 2. Plánované porovnania v reakcii na prvú dávku 0.5 mg / kg neodhalili významné rozdiely medzi skupinou 50 a skupinou 0 alebo skupinou 100, t(35) <1.28, p > 0.20. Skupina 50 však vykazovala signifikantne vyššiu pohybovú odpoveď na druhú dávku 0.5 mg / kg ako skupina 0, t(35) = 4.32, p <0.001 alebo skupina 100, t(35) = 2.24, p <0.05.
Obrázok 9. Priemerná (SE) lokomotorická odpoveď (počet lámaných lúčov v elektronickom poli na 90 min) na 0.5 mg / kg d-amfetamínu na oddelených sedeniach pred a po 5-senzibilizačnom režime d-amfetamínu (1.0 mg / kg; ip na jednu reláciu) v skupinách potkanov Lewis (n = 8 / group) predtým vystavené denným kondicionačným reláciám 15 s odmenou sacharózy (riešenie 10%) dodané pod premennými 0, 25, 50, 75 alebo 100%, Podmieneným stimulom bolo svetlo (120 s). Skupina 0 dostala rovnaký počet odmien ako skupina 100 v neprítomnosti podmienených podnetov. *p <0.05 pre stredný rozdiel medzi skupinou 50 a skupinou 0, ako aj skupinou 100, na základe plánovaných porovnaní.
Účinky 1 mg / kg AMPH.
Pohyb pred vstrekovaním, Skupina 5 × 5 Session ANOVA 30-min skóre pred injekciou pre relácie senzibilizácie priniesla hlavný účinok Session, F(4, 140) = 4.10, p = 0.004 a žiadne iné významné účinky, F(4, 35) = 1.25, p > 0.31. Plánované porovnania zistili, že zlomenie lúča počas fázy pred injekciou (M; SE) bolo významne nižšie v skupine 50 (395; 62) ako v skupine 100 (508; 62), t(175) = 2.58, p <0.01, ale nie skupina 0, t(175) <1.83, p > 0.10, v relácii 1 mg / kg AMPH 1. Na poslednej relácii 1 mg / kg AMPH sa v plánovaných porovnaniach tiež zistilo, že lokomócia pred injekciou v skupine 50 (378; 60) bola významne nižšia ako v skupine 100 (650; 75 ), t(175) = 6.17, p <0.001, ale nie v skupine 0, t(175) <1.84, p > 0.10. Pretože smer týchto skupinových rozdielov (kontrolná skupina = skupina 50) bol opačný k hypotézovanému vzoru, nemožno skupinové rozdiely v postinjekčnej lokomócii, ktoré sa zhodujú s hypotézou, pripísať základným rozdielom pred injekciou. Priemerné (SE) celkové zlomy lúča počas fázy pred injekciou pre relácie 1 až 5 boli: 442 (34), 452 (32), 542 (40), 411 (26), 504 (37).
Pohyb po injekcii, Skupina 5 × 5 Session ANOVA odpovedí na dávky 1 mg / kg priniesla významný hlavný účinok Session, F(4, 140) = 6.15, p <0.001 a žiadne ďalšie významné účinky, F(4, 35) <0.57, p > 0.68. Analýzy polynomiálnych trendov odhalili významný lineárny trend, F(1, 35) = 9.34, p = 0.004 a kubický trend, F(1, 35) = 5.08, p = 0.031, tento výsledok označuje relatívne maximá na reláciách 3 a 5. figúra 10 Vykazuje tieto skóre a ukazuje, že napriek nedostatku významnej interakcie v ANOVA skupina 50 vykazovala podstatne väčšiu lokomóciu ako ostatné štyri skupiny v reakcii na konečnú dávku 1 mg / kg. Preto plánované porovnania ukázali výrazne vyššie priemerné skóre na relácii 5 v skupine 50 ako vo všetkých ostatných skupinách, t(35) > 3.68, p <0.001.
Obrázok 10. Priemerná (SE) lokomotorická odpoveď (počet zlomov lúča v elektronickom poli na 90 min) na 1 mg / kg d-amfetamínu (ip) na týždenných sedeniach 5 u skupín potkanov Lewis (n = 8 / group) predtým vystavené denným kondicionačným reláciám 15 s odmenou sacharózy (riešenie 10%) dodané pod premennými 0, 25, 50, 75 alebo 100%, Podmieneným stimulom bolo svetlo (120 s). Skupina 0 dostala rovnaký počet odmien ako skupina 100 v neprítomnosti podmienených podnetov. *p <0.05 pre stredný rozdiel medzi skupinou 50 a skupinou 0, ako aj skupinou 100, na základe plánovaných porovnaní.
Kontrola variácie v nose poke reagujúca počas tréningu sacharózy
Dve 5 Group × 2 Session ANCOVA lokomotorickej odozvy na 0.5 mg / kg AMPH pred a po senzibilizačnom režime, vrátane totálneho nosa, sa počas tréningu sacharózy s CS prítomným a CS neprítomným ako separátnym kovariantom nezistili žiadne signifikantné účinky ani na jednu kovarianciu, F(1, 32) <0.44 p > 0.51. Dve 5 skupinové × 5 relácie ANCOVA lokomočnej odpovede na 1 mg / kg AMPH počas senzibilizačných sedení s celkovým vpichom do nosa (prítomný CS, neprítomný CS), pretože samostatné kovariáty nepriniesli žiadne významné účinky kovariátov, kým bol alebo nebol prítomný CS, F(1, 33) <0.14, p > 0.71. Bezdrogový prístup reagujúci na liečbu preto nezohľadňoval skupinové rozdiely v lokomotorických reakciách na ktorúkoľvek dávku AMPH.
Diskusia
Senzibilizácia sa vyvinula na účinky opakovaného 1.0 mg / kg amfetamínu. Údaje o návykoch a fyziologickom roztoku potvrdzujú, že tento účinok nebol spôsobený už existujúcimi rozdielmi, očakávanou dĺžkou alebo reakciami súvisiacimi so stresom. ANCOVA s vpichmi do nosa potvrdzujú, že tieto účinky neboli spôsobené prístupom bez liekov. Samotné dáta z nosa naznačovali, že skupiny získali súvislosť medzi CS a vyhliadkou na odmenu za sacharózu. Úroveň zaradenia skupín „nose-poke“ odpovedajúcich na konci tréningu zodpovedala celkovej frekvencii odmien podľa rôznych časových plánov od najvyššej (skupina 100) po najnižšiu (skupina 0), ako to bolo v experimente 2. Relatívne nižší celkový priemer Úrovne prepichnutia nosa v tomto experimente v porovnaní s experimentmi 1 a 2 môžu odrážať selektívnejší prístup reagujúci na podnety k odmene u potkanov Lewis (Kosten a kol., 2007).
Údaje o dávke 0.5 mg / kg ukázali, že počiatočná lokomotorická odpoveď na AMPH u potkanov Lewis (obrázok č 9) bol v porovnaní s potkanmi Sprague Dawley (experiment 2; obrázok č 5), ale zvýšenie reakcie v rámci skupiny na druhú dávku u potkanov kmeňa Lewis bolo značné (takmer dvojnásobok odpovede na prvú dávku 0.5 mg / kg) po režime liečby AMPH v režime 5. Najmä skupina 50 vykazovala väčšiu lokomotorickú odozvu ako všetky skupiny okrem skupiny 25 na druhú (tj post-senzibilizačnú) dávku 0.5 mg / kg AMPH a väčšiu lokomotorickú odpoveď ako všetky ostatné skupiny, vrátane skupiny 25, na konečnú dávku 1 mg / kg AMPH (konečná senzibilizácia). ,
Súhrnná analýza skupinových rebríčkov naprieč experimentmi
Aby sa určila spoľahlivosť skupinových rozdielov v senzibilizácii, neparametrická analýza vyhodnotila kontingenciu medzi skupinou a hodnosťou priemernej lokomotorickej odpovede na druhú dávku (post-chronickú AMPH) 0.5 mg / kg a konečnú dávku 1.0 mg / kg AMPH. z experimentov 3. Analýza priniesla významný účinok, φ = 0.986, p = 0.025, čo odráža skutočnosť, že skupina 50 sa umiestnila na prvom mieste v porovnaní s jedným z porovnaní. Vyššia hodnosť skupiny 50 v porovnaní so všetkými ostatnými skupinami v reakcii na druhú dávku (post-chronická AMPH) 0.5 mg / kg je znázornená na obrázku. 5 (experiment 2) a obrázok 9 (experiment 3). Vyššia hodnosť skupiny 50 v porovnaní s inými skupinami v reakcii na konečnú dávku 1.0 mg / kg je znázornená na obrázku. 2 (experiment 1) a obrázok 10 (experiment 3). Jedinou výnimkou z tohto vzoru bola odpoveď na konečnú dávku 1.0 mg / kg u potkanov Sprague-Dawley v experimente 2.
Všeobecná diskusia
Súčasná séria experimentov testovala hypotézu, že chronické vystavenie gamblingovému programu odmeňovania môže senzibilizovať mozgové dráhy DA, podobne ako chronické vystavenie drogám. Dôkazy pre takýto účinok by naznačovali, že neuroplasticita rovnakého druhu, o ktorej sa predpokladá, že prispieva k drogovej závislosti, môže byť vyvolaná chronickým vystavením nepredvídateľným plánom odmeňovania. V súlade s literatúrou o drogovej závislosti, lokomotorickej odozve na dávky 0.5 a 1.0 mg / kg dávky AMPH indexovanej reaktivity DA systému, s väčšou lokomóciou v reakcii na neskoršie dávky operatívne definujúce senzibilizáciu (porovnaj Robinson a Berridge, 1993; Pierce a Kalivas, 1997; Vanderschuren a Kalivas, 2000).
Celkovo sú výsledky v súlade s našou hypotézou. Naznačujú však tiež značnú variabilitu experimentálnych účinkov spôsobených procesnými faktormi. Účinky harmonogramu kondicionovania boli mierne, ale konzistentné, pričom skupina 50 preukázala väčšiu odozvu ako ostatné štyri skupiny na obe dávky po piatich dávkových režimoch. Hoci celkovo FHodnoty pre účinky súvisiace so skupinou v analýzach rozptylu boli často nevýznamné, rozdiely v kľúčových skupinách boli potvrdené párovo plánovanými porovnaniami. V tejto súvislosti treba poznamenať, že „súčasné myslenie je však tento celkový význam [pre F v ANOVA] nie je potrebné. Po prvé, hypotézy testované celkovým testom a viacnásobným porovnávacím testom sú dosť odlišné, s pomerne odlišnými úrovňami výkonu. Napríklad celkový F rozdeľuje rozdiely medzi skupinami naprieč počtom stupňov voľnosti pre skupiny. To má za následok zriedenie celkového F v situácii, keď sa viaceré skupinové prostriedky navzájom rovnajú, ale líšia sa od iných prostriedkov “(Howell, 1992, s. 338). To je presne tá situácia, ktorá sa aplikovala v súčasných experimentoch, kde sa očakávalo, že skupina 50 sa bude líšiť od skupiny 0 a skupiny 100 kontrol, ale žiadny rozdiel medzi týmito kontrolnými skupinami nebol predpovedaný pre skupinu 25 alebo skupinu 75.
Údaje z nosa poke potvrdili, že v každom experimente zvieratá získali spojenie medzi CS a vyhliadkou na sacharózovú odmenu. Zhoda medzi frekvenciou nosného poke pre rôzne skupiny a celkovým výskytom odmeňovania v rámci ich príslušných školiacich programov naznačuje, že priemerná miera odmeňovania závislá od odmeňovania sacharózy sa riadi. Nedostatočný sprostredkujúci účinok nosa sa však vyvíja na lokomotorické reakcie súvisiace s skupinou na AMPH v ANCOVA, ktoré naznačujú, že tieto dva spôsoby správania sú základom oddelených procesov.
V niektorých prípadoch bol účinok režimu kondicionovania evidentný v reakcii na prvú dávku AMPH; v iných prípadoch sa objavil až po opakovaných dávkach. Skupinové rozdiely v lokomotorickej odpovedi na prvú dávku AMPH naznačujú, že vystavenie účinkom hazardných hier podobných schémam odmeňovania je samo o sebe dostatočné na vyvolanie senzibilizácie. Skupinové rozdiely v lokomócii po viacnásobných dávkach AMPH poukazujú na jemnejší účinok, ktorý možno charakterizovať ako „citlivosť“, ktorá sa prejavuje len v kombinácii s pokračujúcou expozíciou primárnemu senzibilizačnému činidlu (tj amfetamínu).
Rozdiely v modeli odozvy naprieč experimentmi naznačujú, že dlhší interval medzi tréningom a počiatočnou expozíciou AMPH môže maximalizovať možnosť detegovať inherentný senzibilizačný účinok kondicionačnej liečby. To na druhej strane naznačuje, že účinky podmienenej expozície odmeňovania sa môžu časom inkubovať, čo je jav, ktorý sa pozoruje aj pri senzibilizácii stimulancií (Grimm a kol., 2006). Vzor odpovede na dve dávky amfetamínu naznačuje, že dávka 0.5 mg / kg môže byť účinnejšia pri odhaľovaní účinkov kondicionovanej histórie. To zase naznačuje, že účinky kondicionovania podľa súčasného školiaceho protokolu sú trochu subtílne a môžu byť maskované účinkami stropu v rámci dávok AMPH a podmienok, ktoré vytvárajú de novo senzibilizácie.
V experimente 3 je dvojfázový model odpovede na dávky 0.5 mg / kg a progresívny výskyt nadradenosti v skupine 50 zhodný s očakávaným profilom pre potkany Lewis v reakcii na metamfetamín (Camp et al., 1994). To podporuje opodstatnenosť súčasných zistení a navrhuje prekrývanie faktorov, ktoré zmierňujú zraniteľnosť voči psychostimulačnej senzibilizácii a gamblingovým programom odmien.
Pri experimentoch post-senzibilizačná lokomotorická odpoveď skupiny 50 vo všeobecnosti prevyšovala u ostatných skupín pod rôznymi dávkami amfetamínu a u rôznych kmeňov zvierat. Vysoká variabilita v rámci skupiny a skromné veľkosti efektu medzi skupinami však naznačujú úlohu ďalších faktorov v reaktivite DA systému na amfetamín po vystavení rôznym rozpisom podmienenej odmeny sacharózy. Hoci reakcie DA neurónov na odmeňovanie signálov môžu poskytnúť hrubý model hazardných hier (Fiorillo a kol., 2003), podobne ako všetky modely, dochádza k strate informácií kvôli parsimonii - tj k demonštrovaniu kľúčového procesu. V dôsledku toho vzor účinkov v podmienkach CS-US v pôvodnom Fiorillo et al. štúdia úplne nezovšeobecňuje lokomotorickú odpoveď na amfetamín. Ďalšie vylepšenia modelu sú potrebné na úplné zachytenie aspektov hazardných hier, ktoré majú vplyv na funkciu systému DA.
Celkovo vzaté, výsledky tejto série experimentov poskytujú predbežnú podporu pre hypotézu, že chronické vystavenie gambling-like rozvrhom odmeňovania zvyšuje reaktivitu mozgového DA systému na psychostimulačnú výzvu. Ako také rozširujú zistenia z Singer a kol. (2012) ktorí preukázali, že v porovnaní s pevne stanovenou schémou predchádzajúce vystavenie premenlivému harmonogramu posilňovania v operatívnej paradigme zvyšuje následnú lokomotorickú odpoveď na amfetamín. Presnejšie povedané, súčasné zistenia poukazujú na neistotu poskytovania odmeny ako rozhodujúceho faktoru, ktorý je základom účinkov variabilnej odmeny. Veľkosť účinkov v operatívnej paradigme bola podstatne väčšia ako účinky zistené v predložených experimentoch. To môže odrážať väčšie chronické vystavenie účinkom podobných hrám (55 vs. 15 dní); môže odrážať účinky vyžadujúce operatívnu odpoveď na získanie odmeny (tj úlohu agentúry) namiesto pasívnej expozície, ako je to v tejto štúdii. Zvýšenie trvania odbornej prípravy v súčasnej paradigme by pomohlo vyriešiť tieto otázky.
Z týchto experimentov nie je možné zozbierať platnosť variabilných schém odmeňovania a posilňovania ako modelov hazardných hier. Budúci výskum, ktorý skúma vplyv histórie kondicionovania na rizikové správanie pri hazardných hrách na hlodavcoch, by mohol tento problém riešiť. Podobne, korešpondencia medzi behaviorálnou senzibilizáciou zistenou tu a zvýšenou striatálnou DA reakciou na amfetamín, ktorý sa nedávno našiel u patologických hráčov, musí čakať na ďalšie vyšetrenie (Boileau a kol., 2013). Mikrodialýza by sa mohla zaoberať touto otázkou a predikcia založená na údajoch u ľudí by bola, že väčšie uvoľňovanie DA v skupine 50 „hazard fenotyp“ by bolo najzreteľnejšie pozorované skôr v dorzálnom (senzorimotorickom) striatu než v ventrálnom (limbickom) striate. , Validácia 50% variabilnej expozície CS + odmeny v týchto iných paradigmách by podporila jeho užitočnosť ako bona fide experimentálny model PG.
Zatiaľ čo niektoré formy hazardných hier jednoznačne znamenajú inštrumentálnu reakciu (napr. Hracie automaty), v iných formách hazardných hier (napr. Lotérie) existuje súvislosť medzi akciou (zakúpením tiketu, tj uzavretím stávky), dôvodmi pre odmenu (tj. , čísla lotérie) a samotná odmena (výherné číslo a peňažná výplata) je oveľa rozptýlenejšia. Napriek tomu sa môže pokojne vyskytnúť aktivácia DA počas intervalu CS-US. To môže vysvetľovať, prečo sa pri vyhlásení „výherného čísla“ pozornosť upriamuje na pozornosť, keď každá jednotlivá loterijná guľa klesá postupne a je možné zostaviť konkrétnu postupnosť číslic vo výhernom čísle. Aj keď je pravdepodobnosť výskytu konkrétnej číslice matematicky definovaná, výsledok pre každú jednotlivú loterijnú loptičku je binárny - zásah (zhoduje sa s číslom hráča) alebo miss (nezhoduje sa s číslom hráča) - a výsledok ľubovoľného daného pokusu nie je známy. Takýto scenár môže lepšie charakterizovať skúsenosť skupiny 50 v súčasných experimentoch, kde bola odmena poskytnutá náhodne, ale aj nepredvídateľne a CS iba naznačila potenciál odmeny bez toho, aby prezradila, či k nej v danom pokuse dôjde. Výherné automaty sú viac spojené s PG ako s tiketmi (Cox a kol., 2000; Bakken a kol., 2009), čo poukazuje na dôležitú úlohu inštrumentálnych faktorov (a bezprostrednosti) v odmeňovaní aspektov hazardných hier pre túto populáciu (Loba a kol., 2001). Pavlovovský proces modelovaný v súčasných experimentoch (CS + neistá odmena) sa však javí ako nevyhnutný, ak nie dostatočný prvok skúseností s hazardom.
Spolu s chýbajúcou jasnou inštrumentálnou požiadavkou mohlo k relatívne skromnému a variabilnému vzoru experimentálnych účinkov prispieť množstvo ďalších konštrukčných prvkov. Skupiny sa líšili celkovým vystavením sacharóze, ako aj kontingenciou medzi hodnotami CS a sacharózy. Hoci to mohlo prispieť k variabilite medzi skupinami, nemôže ľahko vysvetliť, prečo zvieratá s najväčšou expozíciou sacharózy (skupina 100) vykazovali menšiu senzibilizáciu ako skupina 50. Okrem toho skupina 0 nedostávala žiadny stimul pred vystavením sacharóze pri každom skúšaní. Aj keď to vylučovalo očakávanie odmeny vyvolané cue, nekontrolovalo prítomnosť stimulu pred dodaním odmeny, ktorá existovala vo všetkých ostatných skupinách. Aby sa tento problém riešil, budúci výskum by mal zahŕňať stav, v ktorom zvieratá dostávajú odmenu v každom skúšaní po náhodnom vystavení neutrálnemu stimulu (tj ktorého prítomnosť neznamená potenciál pre odmenu).
Ďalším obmedzením návrhu je potenciálny vznik prídavného správania, ktoré by mohlo ovplyvniť účinky tréningového plánu. Vzhľadom na neistotu sa u zvierat môžu vyvinúť poverčivé správanie určené na zvýšenie vnímanej kontroly a zníženie neistoty vyvolanej aktivácie DA (porov. Harris a kol., 2013). Je preto možné, že nekontrolované aspekty experimentálneho dizajnu umožnili zvieratám kompenzovať účinky harmonogramu kondicionovania. Takýto účinok by mohol prispieť k relatívne skromnej a variabilnej odpovedi na amfetamín v skupine 50 po tréningu CS + sacharózy. Budúci výskum by mal zaznamenávať spontánne správanie, okrem výkyvov nosa, počas tréningov, aby sa táto možnosť otestovala a štatisticky by sa mala kontrolovať, ak sa objaví. Pretože sa očakáva, že takéto správanie bude pôsobiť proti účinkom neistoty spôsobenej harmonogramom alebo tlmí účinky, lokomotorická odpoveď na amfetamín v skupine 50 by sa mala zvýšiť, ak je kontrolovaná (procesne alebo štatisticky). Súčasný (nekontrolovaný) dizajn teda poskytuje konzervatívny test účinkov 50% CS + na amfetamínovú senzibilizáciu.
Pokiaľ ide o vonkajšiu platnosť, použitie samcov potkanov tiež obmedzuje zovšeobecniteľnosť výsledkov. Chýbajúca jasná podmienka „trestu“ sa líši aj od hazardných hier, kde sú veľké peňažné straty bežné a majú významné motivačné účinky (Nieuwenhuis a kol., 2005; Singh a Khan, 2012). Schopnosť akumulovať odmenu chýba aj v súčasnej paradigme a kumulatívne výhry v hracom automate sa ukázali ako interakcie s DA manipuláciami u ľudí (Tremblay a kol., 2011; Smart a kol., 2013). Podobne, príležitosť na jackpot je dôležitým rozdielom medzi súčasným modelom a skutočným hazardom.
Napriek týmto obmedzeniam, súčasné výsledky naznačujú, že 50% variabilná CS + odmena môže zapojiť DA cesty zahrnuté v posilňujúcich účinkoch hazardných hier (Fiorillo a kol., 2003; Anselme, 2013). Krížová senzibilizácia odpovede na AMPH v súlade s týmto harmonogramom podobným hazardu je v súlade s kľúčovou úlohou DA pri hrách a psychostimulačných účinkoch liekov (Zack a Poulos, 2009) a rozširuje predchádzajúce štúdie o krížovom primingu motivácie k hazardu zo strany AMPH u patologických hráčov (Zack a Poulos, 2004). Súčasné výsledky tiež nepriamo naznačujú, že skromné dávky AMPH, ktoré nespôsobujú supra-fyziologické uvoľňovanie DA, môžu lepšie modelovať mozgovú aktivitu v reakcii na intermitentné signály odmien (tj počas hry), ako je vystavenie vysokým (napr. Nárazom). dávky stimulačných liekov (porovnaj Vanderschuren a Pierce, 2010). Priama podpora pre túto korešpondenciu by mohla byť odvodená z hodnotenia uvoľňovania DA ako odozvy na 50% variabilný CS-US plán a rôznych dávok AMPH pomocou mikrodialýzy.
Z experimentálneho hľadiska sa zdá, že súčasný Pavlovov model a predchádzajúci operatívny model variabilného vystuženia vyvolávajú fenotyp podobný ľudskému patologickému hráčovi. Ako také poskytujú hodnotný doplnok k úlohám v oblasti hazardných hier hlodavcov, ktoré modelujú správanie sa hazardných hier (ako závislé opatrenie), ale doteraz používali len zdravé zvieratá, čo je ekvivalent ľudských sociálnych hráčov. Na základe literatúry sa zvieratá chronicky vystavené variabilnej odmene môžu v týchto úlohách veľmi líšiť, najmä v reakcii na DA-ergické lieky. Kombinácia fenotypu potkaniaho hazardu s úlohami hazardných hier môže umožniť systematický rozvoj liekov na liečbu PG, ktoré sa nemusia úplne dosiahnuť len u zdravých zvierat. Ďalšie vylepšenia v experimentálnom dizajne a tréningovom režime, ako je opísané vyššie, by mali zlepšiť súlad medzi zvieratami vyškolenými v tejto paradigme a skutočnými patologickými hráčmi.
Z klinicko-sociologického hľadiska zistenie, že expozícia 50% premenná CS + odmena, ktorá úzko zodpovedá plánu odmien na komerčnom hracom automate (Tremblay a kol., 2011), zmeny v systéme DA mozgu spoľahlivými a trvalými spôsobmi naznačujú, že v niektorých prípadoch môže byť hazardná aktivita, podobne ako drogy zneužívania, „patogénom“ schopným spôsobiť závislosť. Avšak skromná veľkosť účinku a vysoká variabilita v reakcii na 50% CS + odmenu naznačujú, že podobne ako drogy zneužívania, tendencia hazardných hier, ako sú programy odmeňovania na podporu závislosti, bude vo veľkej miere závisieť od vopred existujúceho rizikového profilu hráča. Napriek tomu, aby sa ušetrili osoby s vysokým rizikom vystavenia potenciálnym nepriaznivým účinkom spojeným s hraním hazardných hier, zdá sa byť rozumné, aby sa politiky uplatňované na odradenie od používania a minimalizáciu škôd spôsobených zneužívaním drog mohli rozšíriť aj na hazardné hry.
Vyhlásenie o konflikte záujmov
Autori vyhlasujú, že výskum bol vykonaný bez obchodných alebo finančných vzťahov, ktoré by mohli byť interpretované ako potenciálny konflikt záujmov.
Poďakovanie
Tento výskum bol financovaný z grantov Kanady pre prírodné vedy a inžiniersky výskum Paula J. Fletchera. Úprimne ďakujeme pani Djurdjovi Djordjevičovej za prípravu údajov.
Referencie
Anselme, P. (2013). Dopamín, motivácia a evolučný význam správania podobného hazardu. Behave. Brain Res, 256, 1 – 4. doi: 10.1016 / j.bbr.2013.07.039
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
APA (2013). Diagnostický a štatistický manuál duševných porúch, publikácia 5th Edn, Arlington, VA: American Psychiatric Publishing.
Bakken, IJ, Gotestam, KG, Grawe, RW, Wenzel, HG a Oren, A. (2009). Chovanie v hazardných hrách a problémy s hazardnými hrami v Nórsku 2007. Scanda. J. Psychol, 50, 333-339. dva: 10.1111 / j.1467-9450.2009.00713.x
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Boileau, I., Payer, D., Chugani, B., Lobo, DS, Houle, S., Wilson, AA, et al. (2013). In vivo dôkaz o vyššom uvoľňovaní dopamínu indukovanom amfetamínom pri patologickom hazardnom hraní: štúdia pozitrónovej emisnej tomografie s [C] - (+) - PHNO. Mol. psychiatrie doi: 10.1038 / mp.2013.163. [Epub pred tlačou].
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Bolles, RC (1972). Posilnenie, očakávania a vzdelávanie. Psychol. otáčka, 79, 394 – 409. doi: 10.1037 / h0033120
Camp, DM, Browman, KE a Robinson, TE (1994). Účinky metamfetamínu a kokaínu na motorické správanie a extracelulárny dopamín vo ventrálnom striate Lewis verzus Fischer 344 potkanov. Brain Res. 668, 180–193. doi: 10.1016/0006-8993(94)90523-1
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Conversano, C., Marazziti, D., Carmassi, C., Baldini, S., Barnabei, G. a Dell'Osso, L. (2012). Patologické hráčstvo: systematické preskúmanie biochemických, neurovíznych a neuropsychologických nálezov. Harv. Psychiatria 20, 130-148. dva: 10.3109 / 10673229.2012.694318
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Cox, BJ, Kwong, J., Michaud, V. a Enns, MW (2000). Problém a pravdepodobný patologický hazard: úvahy z komunitného prieskumu. Môcť. J. Psychiatria 45, 548-553.
Dalley, JW, Theobald, DE, Eagle, DM, Passetti, F. a Robbins, TW (2002). Deficity v impulznej kontrole spojené s tonicky zvýšenou serotonergnou funkciou v prefrontálnom kortexe potkanov. neuropsychofarmakologie 26, 716–728. doi: 10.1016/S0893-133X(01)00412-2
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Everitt, BJ a Robbins, TW (2005). Neurónové systémy posilňovania drogovej závislosti: od činov k návykom k donucovaniu. Nat. Neurosci, 8, 1481 – 1489. doi: 10.1038 / nn1579
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Fiorillo, CD, Tobler, PN a Schultz, W. (2003). Diskrétne kódovanie pravdepodobnosti odmien a neistoty dopamínovými neurónmi. veda 299, 1898-1902. dva: 10.1126 / science.1077349
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Flagel, SB, Watson, SJ, Akil, H. a Robinson, TE (2008). Individuálne rozdiely v pripisovaní stimulačného významu k odmeňovaniu súvisiacemu s odmenou: vplyv na senzibilizáciu kokaínu. Behave. Brain Res, 186, 48 – 56. doi: 10.1016 / j.bbr.2007.07.022
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Fletcher, PJ, Tenn, CC, Rizos, Z., Lovic, V. a Kapur, S. (2005). Senzibilizácia na amfetamín, ale nie PCP, zhoršuje posun nastavenia pozornosti: reverziu pomocou agonistu receptora D1 vstreknutého do mediálneho prefrontálneho kortexu. Psychofarmakológia (Berl.) 183, 190–200. doi: 10.1007/s00213-005-0157-6
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Flores, G., Wood, GK, Barbeau, D., Quirion, R. a Srivastava, LK (1998). Lewis a Fischer potkany: porovnanie hladín dopamínového transportéra a receptorov. Brain Res. 814, 34–40. doi: 10.1016/S0006-8993(98)01011-7
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Frascella, J., Potenza, MN, Brown, LL a Childress, AR (2010). Zdieľané zraniteľnosti mozgu otvárajú cestu pre nezáväzné závislosti: rezbárska závislosť na novom spoločnom? Ann. NY Acad. sci, 1187, 294-315. dva: 10.1111 / j.1749-6632.2009.05420.x
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Grimm, JW, Buse, C., Manaois, M., Osincup, D., Fyall, A. a Wells, B. (2006). Časovo závislá disociácia účinkov kokaínovej dávky na chuť a lokomóciu sacharózy. Behave. Pharmacol, 17, 143 – 149. doi: 10.1097 / 01.fbp.0000190686.23103.f8
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Haile, CN, Hiroi, N., Nestler, EJ a Kosten, TA (2001). Diferenciálne behaviorálne reakcie na kokaín sú spojené s dynamikou mezolimbických dopamínových proteínov u potkanov Lewis a Fischer 344. Synapse 41, 179 – 190. doi: 10.1002 / syn.1073
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Harris, JA, Andrew, BJ a Kwok, DW (2013). Časopis prístup počas signálu pre potraviny závisí na Pavlovian, nie inštrumentálne, klimatizácia. J. Exp. Psychol. Anim. Behave. proces, 39, 107 – 116. doi: 10.1037 / a0031315
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Howell, DC (1992). Štatistické metódy pre psychológiu, Boston, MA: Duxbury.
Ito, R., Dalley, JW, Robbins, TW a Everitt, BJ (2002). Uvoľňovanie dopamínu v dorzálnom striate počas správania pri vyhľadávaní kokaínu pod kontrolou podania súvisiaceho s liekom. J. Neurosci, 22, 6247-6253.
Jiao, X., Pare, WP a Tejani-Butt, S. (2003). Rozdiely medzi kmeňmi v distribúcii miest dopamínového transportéra v mozgu potkana. Prog. Neuropsychopharmacol. Biol. psychiatrie 27, 913–919. doi: 10.1016/S0278-5846(03)00150-7
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Koob, GF, a Le Moal, M. (2008). Preskúmanie. Neurobiologické mechanizmy pre oponentské motivačné procesy v závislosti. Philos. Trans. R. Soc. Londa. B Biol. sci, 363, 3113-3123. dva: 10.1098 / rstb.2008.0094
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Kosten, TA, Miserendino, MJ, Chi, S. a Nestler, EJ (1994). Fischerove a Lewisove potkanie kmene vykazujú rozdielne kokaínové účinky v podmienenom mieste preferencie a behaviorálnej senzibilizácie, ale nie v lokomotorickej aktivite alebo podmienenej averzii chuti. J. Pharmacol. Exp. Ther, 269, 137-144.
Kosten, TA, Zhang, XY a Haile, CN (2007). Rozdiely medzi kmeňmi v udržiavaní samosprávy kokaínu a ich vzťahu k reakciám na nové aktivity. Behave. Neurosci, 121, 380 – 388. doi: 10.1037 / 0735-7044.121.2.380
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Leeman, RF a Potenza, MN (2012). Podobnosti a rozdiely medzi patologickými hráčmi a poruchami užívania drog: zameranie sa na impulzívnosť a kompulzívnosť. Psychofarmakológia (Berl.) 219, 469–490. doi: 10.1007/s00213-011-2550-7
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Leshner, AI (1997). Závislosť je ochorenie mozgu a záleží na tom. veda 278, 45-47. dva: 10.1126 / science.278.5335.45
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Loba, P., Stewart, SH, Klein, RM a Blackburn, JR (2001). Manipulácie vlastností štandardných hier VLT (video lottery terminal): účinky v patologických a nepatologických hráčoch. J. Gambl. gombík, 17, 297 – 320. doi: 10.1023 / A: 1013639729908
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Martinez, D., Narendran, R., Foltin, RW, Slifstein, M., Hwang, DR, Broft, A., et al. (2007). Uvoľňovanie dopamínu indukované amfetamínom: značne otupené v závislosti od kokaínu a prediktívnosť voľby kokaínu. Am. J. Psychiatry 164, 622-629. dva: 10.1176 / appi.ajp.164.4.622
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Mateo, Y., Lack, CM, Morgan, D., Roberts, DC a Jones, SR (2005). Znížená terminálna funkcia dopamínu a necitlivosť na kokaín po kokaínovom samovoľnom podaní a deprivácii. neuropsychofarmakologie 30, 1455-1463. dva: 10.1038 / sj.npp.1300687
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Nestler, EJ (2001). Molekulárna podstata dlhodobej závislosti od závislosti. Nat. Neurosci, 2, 119 – 128. doi: 10.1038 / 35053570
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Nieuwenhuis, S., Heslenfeld, DJ, von Geusau, NJ, Mars, RB, Holroyd, CB a Yeung, N. (2005). Aktivita v oblastiach mozgu citlivých na ľudské odmeny je silne závislá od kontextu. Neuroimage 25, 1302-1309. dva: 10.1016 / j.neuroimage.2004.12.043
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Pierce, RC a Kalivas, PW (1997). Obvodový model vyjadrenia behaviorálnej senzibilizácie na amfetamínové psychostimulanty. Brain Res. Brain Res. otáčka, 25, 192. doi: 10.1016 / S0165-0173 (97) 00021-0
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Robbins, TW a Everitt, BJ (1999). Drogová závislosť: zvyšujú sa zlé návyky. príroda 398, 567-570. dva: 10.1038 / 19208
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Robinson, TE, Becker, JB a Presty, SK (1982). Dlhodobé uľahčenie amfetamínom indukovaného rotačného správania a uvoľňovanie striatálneho dopamínu vyvolaného jednorazovou expozíciou amfetamínu: rozdiely medzi pohlaviami. Brain Res. 253, 231–241. doi: 10.1016/0006-8993(82)90690-4
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Robinson, TE a Berridge, KC (1993). Neurálny základ túžby po drogách: teória motivácie a senzibilizácie závislosti. Brain Res. Brain Res. otáčka, 18, 247 – 291. doi: 10.1016 / 0165-0173 (93) 90013-P
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Robinson, TE a Berridge, KC (2001). Stimulačná senzibilizácia a závislosť. Závislosť 96, 103-114. dva: 10.1046 / j.1360-0443.2001.9611038.x
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Singer, BF, Scott-Railton, J. a Vezina, P. (2012). Nepredvídateľné posilňovanie sacharínu zlepšuje lokomotorickú reakciu na amfetamín. Behave. Brain Res, 226, 340 – 344. doi: 10.1016 / j.bbr.2011.09.003
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Singh, V. a Khan, A. (2012). Rozhodovanie v odmeňovaní a variantoch trestu hazardnej hry iowa: dôkaz o „predvídavosti“ alebo „rámovaní“? Predná. Neurosci, 6: 107. doi: 10.3389 / fnins.2012.00107
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Skinner, BF (1953). Veda a ľudské správanie, New York, NY: Free Press.
Smart, K., Desmond, RC, Poulos, CX a Zack, M. (2013). Modafinil zvyšuje výplatu odmeny v hracom automate v patologických hrách s nízkou a vysokou impulsivitou. Neuropharmacology 73, 66-74. dva: 10.1016 / j.neuropharm.2013.05.015
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Tremblay, AM, Desmond, RC, Poulos, CX a Zack, M. (2011). Haloperidol modifikuje inštrumentálne aspekty hazardných hier v patologických hrách a zdravých kontrolách. Narkoman. Biol, 16, 467-484. dva: 10.1111 / j.1369-1600.2010.00208.x
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Vanderschuren, LJ a Kalivas, PW (2000). Zmeny v dopaminergnom a glutamatergickom prenose v indukcii a expresii behaviorálnej senzibilizácie: kritický prehľad predklinických štúdií. Psychofarmakológia (Berl.) 151, 99-120. dva: 10.1007 / s002130000493
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Vanderschuren, LJ, a Pierce, RC (2010). Senzibilizačné procesy v drogovej závislosti. Akt. Top. Behave. Neurosci, 3, 179 – 195. doi: 10.1007 / 7854_2009_21
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Vanderschuren, LJ, Schoffelmeer, AN, Mulder, AH, a De Vries, TJ (1999). Dopaminergné mechanizmy sprostredkujúce dlhodobú expresiu lokomotorickej senzibilizácie po predexpozícii morfínu alebo amfetamínu. Psychofarmakológia (Berl.) 143, 244-253. dva: 10.1007 / s002130050943
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Verdejo-Garcia, A., Lawrence, AJ, a Clark, L. (2008). Impulzívnosť ako marker zraniteľnosti pre poruchy užívania látok: prehľad zistení z vysoko rizikového výskumu, problémových hráčov a štúdií genetickej asociácie. Neurosci. Biobehav. otáčka, 32, 777 – 810. doi: 10.1016 / j.neubiorev.2007.11.003
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Vezina, P. (2004). Senzibilizácia reaktivity dopamínového neurónu stredného mozgu a samopodávanie psychomotorických stimulantov. Neurosci. Biobehav. otáčka, 27, 827 – 839. doi: 10.1016 / j.neubiorev.2003.11.001
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Volkow, ND, Wang, GJ, Fowler, JS, Logan, J., Gatley, SJ, Hitzemann, R., et al. (1997). Znížená striatálna dopaminergná citlivosť u detoxikovaných subjektov závislých od kokaínu. príroda 386, 830 – 833. doi: 10.1038 / 386830a0
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Volkow, ND, Wang, GJ, Fowler, JS a Tomasi, D. (2012). Závislosť obvodov v ľudskom mozgu. Annu. Pharmacol. Toxicol, 52, 321 – 336. doi: 10.1146 / annurev-pharmtox-010611-134625
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Winer, B. (ed.). (1971). Štatistické princípy v experimentálnom dizajne, New York, NY: McGraw-Hill.
Zack, M., a Poulos, CX (2004). Amfetamín vyvoláva motiváciu k sémantickým sieťam týkajúcim sa hazardných hier a hazardných hier v problémových hráčoch. neuropsychofarmakologie 29, 195-207. dva: 10.1038 / sj.npp.1300333
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Zack, M., a Poulos, CX (2009). Paralelné úlohy dopamínu pri patologickom hráčstve a psychostimulačnej závislosti. Akt. Zneužívanie drog Rev, 2, 11 – 25. doi: 10.2174 / 1874473710902010011
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Zamudio, S., Fregoso, T., Miranda, A., De La Cruz, F. a Flores, G. (2005). Kmeňové rozdiely hladín dopamínových receptorov a správania súvisiaceho s dopamínom u potkanov. Brain Res. býk, 65, 339 – 347. doi: 10.1016 / j.brainresbull.2005.01.009
Pubmed Abstract | Publikovaný celý text | CrossRef Plný text
Kľúčové slová: patologické hráčstvo, senzibilizácia, amfetamín, dopamín, neistota
Citácia: Zack M, Featherstone RE, Mathewson S a Fletcher PJ (2014) Chronické vystavenie gamblingu podobného harmonogramu stimulačných stimulov môže podporiť senzibilizáciu na amfetamín u potkanov. Predná. Behave. Neurosci. 8: 36. dva: 10.3389 / fnbeh.2014.00036
Prijaté: 01 November 2013; Prijaté: 23 január 2014;
Publikované online: 11 Február 2014.
strih:
Bryan F. Singer, University of Michigan, USA
Hodnotené:
Louk Vanderschuren, University of Utrecht, Holandsko
Ruud Van Den Bos, Radboud University Nijmegen, Holandsko
Patrick Anselme, Univerzita v Liège, Belgicko
Copyright © 2014 Zack, Featherstone, Mathewson a Fletcher. Toto je článok s otvoreným prístupom distribuovaný podľa podmienok Creative Commons Attribution License (CC BY), Používanie, distribúcia alebo reprodukcia na iných fórach je povolené za predpokladu, že pôvodný autor (autorov) alebo poskytovateľ licencie je pripísaný a že pôvodná publikácia v tomto časopise je citovaná v súlade s prijatou akademickou praxou. Nepoužíva sa žiadna distribúcia alebo reprodukcia, ktorá nie je v súlade s týmito podmienkami.
* Korešpondencia: Martin Zack, Kognitívne psychofarmakologické laboratórium, Neuroscience Department, Centrum pre závislosť a duševné zdravie, 33 Russell Street, Toronto, ON M5S 2S1, Kanada e-mail: [chránené e-mailom]
