PRIPOMBE: Kot bodo pokazale kasnejše študije, je DeltaFosB običajno molekularno stikalo tako za odvisnosti od drog kot za vedenje. Je transkripcijski faktor, kar pomeni, da vpliva na to, kateri geni so vklopljeni ali izklopljeni. Kot je navedeno drugje, mamila, ki povzročajo odvisnost, ugrabijo le običajne mehanizme. Zato je neumno trditi, da vedenjske odvisnosti ne morejo obstajati.
Proc Natl Acad Sci US A. 2001 September 25; 98 (20): 11042 – 11046.
doi: 10.1073 / pnas.191352698.
Eric J. Nestler *, Michel Barrot in David W. Self
Oddelek za psihiatrijo in Center za osnovno nevroznanost, University of Texas Southwestern Medical Center, 5323 Harry Hines Boulevard, Dallas, TX 75390-9070
Minimalizem
Dolga življenjska doba nekaterih vedenjskih nenormalnosti, ki so značilne za zasvojenost z drogami, je pokazala, da je regulacija izražanja nevralnih genov lahko vključena v proces, s katerim droge povzročajo zasvojenost. jazVečji dokazi kažejo, da transkripcijski faktor ΔFosB predstavlja en mehanizem, s katerim droge zlorabljajo relativno stabilne spremembe v možganih, ki prispevajo k fenotipu odvisnosti. ΔFosB, član družine Fos transkripcijskih faktorjev, se nabira v podskupini nevronov nukleusa accumbens in dorzalnega striatuma (področja možganov, ki so pomembna za zasvojenost) po večkratni uporabi mnogih vrst zlorab. Podobno kopičenje ΔFosB nastopi po kompulzivnem teku, kar kaže, da se lahko ΔFosB kopiči kot odziv na številne vrste kompulzivnega vedenja.. Pomembno je, da ΔFosB zaradi izjemne stabilnosti vztraja pri nevronih relativno dolgo časa. Zato ΔFosB predstavlja molekularni mehanizem, ki lahko sproži in nato vzdržuje spremembe v izražanju genov, ki trajajo dolgo po prenehanju izpostavljenosti zdravilu.. Študije na inducibilnih transgenskih miših, ki prekomerno izražajo ΔFosB ali dominantni negativni inhibitor proteina, neposredno dokazujejo, da ΔFosB povzroča povečano občutljivost za vedenjske učinke zlorabe drog in po možnosti povečano vedenje zaradi iskanja drog. To delo podpira stališče, da ΔFosB deluje kot vrsta trajnega "molekularnega stikala", ki postopoma pretvarja akutne odzive zdravil v relativno stabilne prilagoditve, ki prispevajo k dolgoročni nevralni in vedenjski plastičnosti, ki je osnova za zasvojenost.
Raziskovanje odvisnosti je osredotočeno na razumevanje kompleksnih načinov, s katerimi droge spremenijo možgane in povzročijo vedenjske nepravilnosti, ki so značilne za odvisnost. Eden od ključnih izzivov na tem področju je ugotoviti relativno stabilne spremembe v možganih, ki jih povzročajo droge, da bi upoštevali tiste vedenjske nepravilnosti, ki so še posebej dolgotrajne. Na primer, odvisnik od človeka je lahko izpostavljen povečanemu tveganju za ponovitev bolezni tudi po letih abstinence.
Stabilnost teh vedenjskih nenormalnosti je privedla do domneve, da jih lahko vsaj delno posredujejo spremembe v izražanju genov (1 – 3). V skladu s tem stališčem ponavljajoča se izpostavljenost drogam zlorabe večkrat povzroča motnje prenosa na določene sinapse v možganih, ki so občutljive na zdravilo. Takšne motnje sčasoma preko intracelularnih kurirskih sporočil posredujejo signal jedru, kjer najprej sprožijo in nato ohranijo spremembe v izražanju specifičnih genov. Primarni mehanizem, s katerim signalne transdukcijske poti vplivajo na gensko ekspresijo, je regulacija transkripcijskih faktorjev, proteinov, ki se vežejo na regulatorne regije genov in spreminjajo njihovo transkripcijo.
Eden od ciljev raziskav zasvojenosti je bil torej identificirati transkripcijske faktorje, ki se spreminjajo v regijah možganov, ki so vpleteni v odvisnost po kronični uporabi drog. V zadnjem desetletju je bilo ugotovljenih več takih transkripcijskih faktorjev (1 – 6). Ta pregled je osredotočen na določen transkripcijski faktor, imenovan ΔFosB.
Indukcija ΔFosB z zlorabo drog
ΔFosB, ki ga kodira gen fosB, je član Fosove družine transkripcijskih faktorjev, ki vključuje tudi c-Fos, FosB, Fra1 in Fra2 (7). Ti proteini iz družine Fos se heterodimerizirajo z beljakovinami iz družine Jun (c-Jun, JunB ali JunD), da tvorijo dejavnike transkripcije aktivnega AP-1 (aktivatorskega proteina-1), ki se vežejo na mesta AP-1 (konsenzna sekvenca: TGAC / GTCA) promotorje določenih genov za regulacijo njihove transkripcije.
Te beljakovine družine Fos se inducirajo hitro in prehodno v specifičnih regijah možganov po akutnem dajanju mnogih zdravil zlorabe (sl. 1) (8-11). Pomembne regije so nucleus accumbens in dorsal striatum, ki sta pomembna posrednika za vedenjske odzive na zdravila, zlasti na njihove nagrajevalne in lokomotorne učinke (12, 13). Te beljakovine se vrnejo na osnovne ravni v nekaj urah po dajanju zdravila.
Slika 1
Shema prikazuje postopno kopičenje ΔFosB v primerjavi s hitro in prehodno indukcijo drugih beljakovin družine Fos kot odziv na zlorabo zdravil. (A) Avtoradiogram prikazuje diferencialno indukcijo teh različnih beljakovin z akutno stimulacijo (1 – 2 h po enkratni izpostavljenosti zdravilu) v primerjavi s kronično stimulacijo (1 dan po ponavljajoči se izpostavljenosti zdravilu). (B) Več valov Fos podobnih beljakovin [ki jih sestavljajo c-Fos (52-do 58-kDa izooblike), FosB (46-do 50-kDa izoform), ΔFosB (33-kDa izoforma) in Fra1 ali Fra2 ( 40 kDa)] se sprožijo v nukleus akumbens in dorzalni striatni nevroni z akutnim dajanjem zlorabe drog. Prav tako so inducirane biokemično modificirane izoforme ΔFosB (35-37 kDa); tudi po akutnem dajanju zdravila so inducirani (čeprav na nizki ravni), vendar zaradi svoje stabilnosti vztrajajo v možganih dalj časa. (C) Pri ponavljajočem (npr. Dvakrat na dan) dajanju zdravila vsak akutni stimulus inducira nizko raven stabilnih ΔFosB izoform, kar je označeno z nižjim nizom prekrivajočih se linij, ki kažejo ΔFosB, inducirano z vsakim akutnim dražljajem. Rezultat je postopno povečanje skupne vrednosti ΔFosB s ponavljajočimi dražljaji med potekom kroničnega zdravljenja, kar je prikazano z naraščajočo stopničasto linijo v grafu.
Zelo različni odzivi so opaženi po kroničnem dajanju zdravil za zlorabo (slika 1).. Biokemično modificirane izoforme ΔFosB (molekulska masa 35-37 kDa) se kopičijo v istih možganskih regijah po večkratni izpostavljenosti zdravilu, medtem ko vsi drugi člani družine Fos izkazujejo toleranco (to je zmanjšano indukcijo v primerjavi z začetno izpostavljenostjo zdravilu). Tako kopičenje ΔFosB so opazili pri kokainu, morfinu, amfetaminu, alkoholu, nikotinu in fenciklidinu.e (11, 14-18). Obstajajo nekateri dokazi, da je ta indukcija selektivna za podskupino srednjih trnov, ki vsebujejo dynorphin / substance P, v teh regijah možganov (15, 17), čeprav je za zagotovitev tega potrebno več dela. Izoforme 35-37-kDa ΔFosB dimeriziramo pretežno z JunD, da tvorimo aktiven in dolgotrajen AP-1 kompleks v teh regijah možganov (19, 20). Te izoforme ΔFosB se kopičijo s kronično izpostavljenostjo zdravilu zaradi izredno dolge razpolovne dobe (21) in zato vztrajajo pri nevronih vsaj nekaj tednov po prenehanju dajanja zdravila. Zanimivo je, da so te izoforme ΔFosB zelo stabilni produkti takojšnjega zgodnjega gena (fosB). Stabilnost izooblik ΔFosB zagotavlja nov molekularni mehanizem, s katerim lahko povzročene spremembe v izražanju genov vztrajajo kljub relativno dolgim obdobjem odvzema zdravil.
Čeprav ima nucleus accumbens ključno vlogo pri nagrajevanju učinkov zlorabe drog, se domneva, da normalno deluje tako, da ureja odzive na naravne ojačevalce, kot so hrana, pijača, spol in socialne interakcije (12, 13). Posledica tega je, da obstaja veliko zanimanje za možno vlogo te možganske regije pri drugih kompulzivnih vedenjih (npr. Patološko prenajedanje, igranje na srečo, vadbo itd.). Zato smo preverili, ali je ΔFosB reguliran v živalskem modelu kompulzivnega teka. Dejansko se stabilne izoforme ΔFosB iz 35 do 37-kDa selektivno inducirajo znotraj nucleus accumbens pri podganah, ki kažejo kompulzivno obnašanje v teku. †
Biokemijska identiteta stabilnih ΔFosB Isoform
Kot je navedeno zgoraj, izooblike ΔFosB, ki se kopičijo po kroničnem dajanju zdravila ali zlorabe, kažejo molekulsko maso 35-37 kDa. Razlikujemo jih lahko iz izooblike 33-kDa ΔFosB, ki se inducira hitro, a prehodno po enkratni izpostavljenosti zdravilu (slika 1) (14, 19, 22). Trenutni dokazi kažejo, da je izoforma 33-kDa naravna oblika proteina, ki je spremenjena, da tvori bolj stabilne produkte 35-37-kDa (19, 21). Vendar je narava biokemijske modifikacije, ki pretvarja nestabilno izoformo 33-kDa v stabilne izoforme 35-37-kDa, še vedno nejasna. Špekuliralo se je, da je lahko odgovorna fosforilacija (11). Na primer, indukcija ΔFosB je oslabljena pri miših brez DARPP-32, striatno obogatenega proteina (23, 24). Ker DARPP-32 uravnava katalitično aktivnost protein fosfataze-1 in proteinske kinaze A (25, 26), zahteva za ta protein za normalno kopičenje stabilnih izooblik ΔFosB nakazuje možno vlogo fosforilacije pri tvorbi teh stabilnih produktov.
Vloga ΔFosB v vedenjski plastičnosti proti zlorabi drog
Vpogled v vlogo ΔFosB pri zasvojenosti z drogami je v veliki meri posledica študij transgenskih miši, v katerih se lahko ΔFosB selektivno inducira znotraj nucleus accumbens in drugih striatnih regij odraslih živali (27, 28). Pomembno je, da te miši prekomerno izražajo ΔFosB selektivno v srednjih živčnih nevronih, ki vsebujejo dinorfin / snov P, kjer se domneva, da zdravila inducirajo protein. Obnašanje fenotipa ΔFosB prekomerno izraženih miši, ki v mnogih pogledih spominja na živali po kronični izpostavljenosti zdravilu, je povzeto v tabeli 1. Po akutni in kronični uporabi (28) so miši pokazali povečan lokomotorni odziv na kokain. Prav tako kažejo večjo občutljivost na koristne učinke kokaina in morfina v testih za pripravo na mesto (11, 28) in bodo samostojno dajali manjše odmerke kokaina kot legla, ki ne prekomerno izražajo ΔFosB. senzibilizacija za kokain in normalno prostorsko učenje v vodnem labirintu Morris (28). Tti podatki kažejo, da ΔFosB poveča občutljivost živali na kokain in morda druge droge, ki zlorabljajo, in lahko predstavlja mehanizem za sorazmerno dolgotrajno preobčutljivost na zdravila.
 striatum
striatum
| Povečana aktivacija lokomotorja kot odziv na akutno in ponavljajočo uporabo kokaina. |
| Povečani nagrajeni odzivi na kokain in morfin v testih za pripravo na mesto. |
| Povečano samoinjiciranje nizkih odmerkov kokaina. |
| Povečana motivacija za kokain v postopnih analizah razmerja. |
| Povečan anksiolitični odziv na alkohol. |
| Povečano kompulzivno delovanje. |
Na podlagi podatkov v ref. 28 in 29.† ‡ §
Vedenjska plastičnost, ki jo posreduje ΔFosB v nucleus accumbens-dorsal striatum
IPoleg tega obstajajo predhodni dokazi, da se lahko učinki ΔFosB razširijo precej preko regulacije občutljivosti na droge per se na bolj zapleteno vedenje, povezano s procesom odvisnosti. Miševi, ki izražajo ΔFosB, si prizadevajo za samostojno uporabo kokaina v testih samoinjiciranja s progresivnim razmerjem, \ tugotavlja, da lahko ΔFosB zaznava živali na spodbujevalne motivacijske lastnosti kokaina in s tem povzroči nagnjenost k ponovitvi bolezni po prekinitvi zdravljenjal. ‡ ΔFosB-izražene miši kažejo tudi povečane anksiolitične učinke alkohola, fenotip, ki je bil povezan s povečanim vnosom alkohola pri ljudeh. Skupaj te zgodnje ugotovitve kažejo, da ΔFosB poleg povečane občutljivosti na zlorabe drog proizvaja kvalitativne spremembe v vedenju, ki spodbujajo vedenje, ki išče droge. Tako lahko ΔFosB deluje kot trajno "molekularno stikalo", ki pomaga inicirati in vzdrževati ključne vidike odvisnega stanja. V sedanji preiskavi je pomembno vprašanje, ali kopičenje ΔFosB med izpostavljenostjo drog spodbuja iskanje drog po daljšem karenci, tudi po normalizaciji ravni ΔFosB (glej spodaj).
Izobraževanje odraslih miši, ki prekomerno izražajo ΔFosB selektivno znotraj nucleus accumbens in dorzalnega striatuma, kažejo tudi večje kompulzivno delovanje v primerjavi s kontrolnimi legami. † Ta opazovanja vzbujajo zanimivo možnost, da akumulacija ΔFosB znotraj teh nevronov služi bolj splošni vlogi pri oblikovanju in vzdrževanju navadnih spominov in kompulzivnosti obnašanja, morda z okrepitvijo učinkovitosti nevronskih vezij, v katerih delujejo ti nevroni.
ΔFosB se po kronični izpostavljenosti kokainu kopiči v določenih regijah možganov zunaj nucleus accumbens in dorzalnega striatuma. Izstopajo med njimi regije so amigdala in medialni prefrontalni korteks (15). Glavni cilj trenutnih raziskav je razumeti prispevke indukcije ΔFosB v teh regijah k fenotipu odvisnosti.
Prejšnje delo na miših, ki so izločale fosB, je pokazalo, da te živali ne razvijejo preobčutljivosti na gibalne učinke kokaina, kar je v skladu z ugotovitvami zgoraj omenjenih miši s prekomernim izražanjem ΔFosB (22). Vendar so mutanti fosB pokazali večjo občutljivost na akutne učinke kokaina, kar ni v skladu s temi drugimi ugotovitvami. Interpretacija ugotovitev z mutanti fosB pa je zapletena zaradi dejstva, da tem živalim ne manjka le ΔFosB, temveč tudi FosB v celotni dolžini. Poleg tega mutantom v možganih in v najzgodnejših fazah razvoja primanjkuje beljakovin. Novejše delo dejansko podpira zaključke miši s prekomerno izražanjem ΔFosB: inducibilna prekomerna ekspresija okrnjenega mutanta c-Juna, ki deluje kot prevladujoči negativni antagonist ΔFosB, selektivno v nucleus accumbens in hrbtni striatum kaže zmanjšano občutljivost na koristne učinke kokaina .¶ Te ugotovitve poudarjajo previdnost, ki jo je treba uporabiti pri razlagi rezultatov miši z konstitutivnimi mutacijami, in ponazarjajo pomen miši z inducibilnimi in celično specifičnimi mutacijami pri študijah plastičnosti v možganih odraslih.
Ciljni geni za ΔFosB
Ker je ΔFosB transkripcijski faktor, verjetno protein povzroča vedenjsko plastičnost s spremembami v izražanju drugih genov. ΔFosB nastane z alternativnim spajanjem fosB gena in nima dela C-terminalne transaktivacijske domene, prisotne v FosB polni dolžini. Zato je bilo prvotno predlagano, da ΔFosB deluje kot transkripcijski represor (29). Vendar pa je delo v celični kulturi jasno pokazalo, da je ΔFosB lahko povzroči ali zatre Transkripcija, ki jo posreduje AP-1, odvisno od posameznega uporabljenega mesta AP-1 (21, 29-31). FosB s polno dolžino ima enake učinke kot ΔFosB na določene fragmente promotorjev, vendar ima različne učinke na druge. Nadaljnje delo je potrebno za razumevanje mehanizmov, na katerih temeljijo ti različni ukrepi ΔFosB in FosB.
Naša skupina je uporabila dva pristopa za identifikacijo ciljnih genov za ΔFosB. Eden je pristop genskega kandidata. Sprva smo upoštevali glutamatne receptorje za α-amino-3-hidroksi-5-metil-4-izoksazolpropionsko kislino (AMPA) kot domnevne cilje glede na pomembno vlogo glutamatergičnega prenosa v nakopičenih jedrih. Dosedanje delo je pokazalo, da je ena določena podenota glutamatnega receptorja AMPA, GluR2, lahko resnična tarča za ΔFosB (slika 2). Ekspresija GluR2, ne pa tudi ekspresija drugih podenot receptorjev AMPA, se poveča v nucleus accumbens (ne pa tudi v hrbtnem striatumu) zaradi prekomerne ekspresije ΔFosB (28), ekspresija dominantnega negativnega mutanta pa oslabi sposobnost kokaina, da inducira protein. Poleg tega promotor gena GluR2 vsebuje soglasno mesto AP-1, ki veže ΔFosB (28). Prekomerna ekspresija GluR2 v nucleus accumbens z uporabo gensko posredovanega virusa poveča občutljivost živali na koristne učinke kokaina, s čimer posnema del fenotipa, opaženega pri miših, ki izražajo ΔFosB (28). Indukcija GluR2 bi lahko bila razlog za zmanjšano elektrofiziološko občutljivost nevronov nucleus accumbens na agoniste receptorjev AMPA po kroničnem dajanju kokaina (32), ker AMPA receptorji, ki vsebujejo GluR2, kažejo zmanjšano skupno prevodnost in zmanjšano prepustnost Ca2 +. Zmanjšana odzivnost teh nevronov na vzbujevalne vložke lahko nato poveča odziv na zlorabo drog. Vendar načini, kako dopaminergični in glutamatergični signali v nucleus accumbens uravnavajo zasvojenost, ostajajo neznani; to bo zahtevalo raven razumevanja nevronskega vezja, ki še ni na voljo.
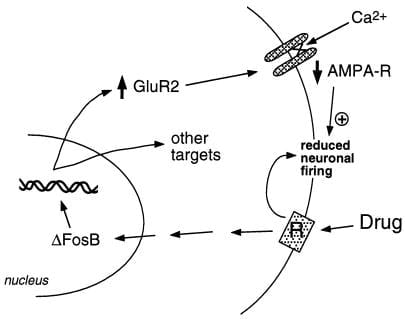
Slika 2
Podenota glutamatnega receptorja AMPA, GluR2, je domnevna tarča ΔFosB. Prikazano je, kako lahko indukcija GluR2, ki jo posreduje ΔFosB, spremeni fiziološko odzivnost nevronov nucleus accumbens in povzroči senzibiliziran odziv na zlorabo drog. V skladu s to shemo zlorabe drog povzročajo akutne okrepitvene učinke z zaviranjem nevronov nucleus accumbens. Z večkratno izpostavljenostjo zdravila inducirajo ΔFosB, ki uravnava številne ciljne gene, vključno z GluR2. To poveča delež receptorjev AMPA (AMPA-R) na nevronih nucleus accumbens, ki vsebujejo podenoto GluR2, kar povzroči zmanjšanje celotnega toka AMPA in zmanjšan tok Ca2 +. Ta zmanjšana razdražljivost bi lahko nevrone naredila bolj občutljive na akutne zaviralne učinke zdravil in s tem na okrepitvene učinke zdravil.
Drug domnevni cilj za ΔFosB je gen, ki kodira dynorphin. Kot je bilo že navedeno, se dynorphin izraža v podmnožici srednjih živčnih nevronov nucleus accumbens, ki kažejo indukcijo ΔFosB. Zdi se, da dynorphin deluje v medcelični povratni zanki: njegovo sproščanje zavira dopaminergične nevrone, ki inervirajo srednja živčna nevrona, preko κ opioidnih receptorjev, ki so prisotni na terminalih dopaminergičnih živcev v nucleus accumbens in tudi na celičnih telesih in dendritih v ventralnem tegmentalnem področju (Sl. 3) (33 – 35). Ta zamisel je skladna s sposobnostjo agonista receptorja κ po dajanju v katero od teh dveh možganskih regij, da zmanjša ponovno zdravljenje z zdravilom.d (35).
RDelo je pokazalo, da ΔFosB zmanjšuje izražanje dynorphina, ki bi lahko prispeval k izboljšanju nagrajevalnih mehanizmov, ki jih vidimo z indukcijo ΔFosB. Zanimivo je, da drugi faktor transkripcije, ki ga regulira zdravilo, CREB (cAMP odgovor element, ki veže beljakovino) (2, 3), deluje nasprotno: inducira ekspresijo dynorphina v nucleus accumbens in zmanjšuje nagrajene lastnosti kokaina in morfina. (4). **
Bzato, ker se zdravljenje, ki ga povzroča zdravilo CREB, hitro razširi po dajanju zdravila, lahko taka vzajemna regulacija dynorphina s strani CREB in ΔFosB pojasni vzajemne vedenjske spremembe, ki se pojavijo v zgodnjih in poznih fazah odtegnitve, z negativnimi čustvenimi simptomi in manjšo občutljivostjo zdravil v zgodnjih fazah umik in senzibilizacija za nagrajevanje in spodbujevalne motivacijske učinke zdravil, ki prevladujejo v poznejših časovnih točkah.
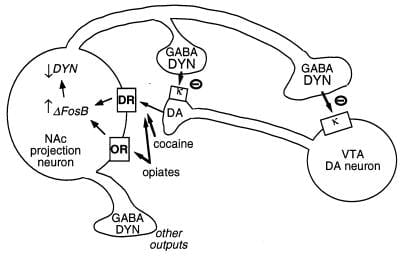
Slika 3
Dynorphin je domnevni cilj za ΔFosB. Prikazan je ventralni tegmentalni del (VTA) dopamin (DA) nevron, ki inervira razred nucleus accumbens (NAc) GABAergični projekcijski nevron, ki izraža dynorphin (DYN). Dynorphin služi povratni mehanizem v tem vezju: dynorphin, sproščen iz terminalov NAc nevronov, deluje na κ opioidne receptorje, ki se nahajajo na živčnih terminalih in celičnih telesih DA nevronov, da zavirajo njihovo delovanje. ΔFosBz zaviranjem ekspresije dynorphina lahko to povratno zanko regulira navzdol in izboljša nagrajevanje lastnosti zlorab. Ni prikazan vzajemni učinek CREB na ta sistem: CREB pospešuje ekspresijo dynorphina in s tem zmanjšuje nagrajene lastnosti zdravil zlorabe (4). GABA, y-aminomaslena kislina; DR, receptor za dopamin; OR, opioidni receptor.
Drugi pristop, uporabljen za identifikacijo ciljnih genov za ΔFosB, vključuje analizo mikromrež DNA. Inducibilna prekomerna ekspresija ΔFosB poveča ali zmanjša izražanje številnih genov v nucleus accumbens (36). Čeprav je zdaj treba potrditi vsakega od teh genov kot fiziološke tarče ΔFosB in razumeti njihov prispevek k fenotipu odvisnosti, se zdi, da je eden pomembnih ciljev Cdk5 (od ciklina odvisna kinaza-5). Tako je bil Cdk5 sprva identificiran kot ΔFosB-reguliran z uporabo mikromrež, kasneje pa se je pokazalo, da ga inducirajo v nucleus accumbens in dorsal striatum po kroničnem dajanju kokaina (37). ΔFosB aktivira gen cdk5 prek mesta AP-1, ki je prisotno v promotorju gena (36). Ti podatki skupaj podpirajo shemo, pri kateri kokain inducira ekspresijo Cdk5 v teh možganskih regijah prek ΔFosB. Zdi se, da indukcija Cdk5 vsaj delno spremeni dopaminergično signalizacijo s povečano fosforilacijo DARPP-32 (37), ki se iz fosforilacije s Cdk1 pretvori iz inhibitorja beljakovinske fosfataze-5 v inhibitor proteinske kinaze A (26).
Vloga ΔFosB pri posredovanju »trajne« plastičnosti z zlorabo drog
Čeprav je signal ΔFosB razmeroma dolgoživ, ni trajen. ΔFosB se postopoma razgradi in se v možganih ne more več odkriti po 1 – 2 mesecih odtegnitve drog, čeprav nekatere vedenjske anomalije trajajo dlje časa. Zato se zdi, da ΔFosB sam po sebi ne bi mogel posredovati teh polpovezanih vedenjskih nepravilnosti. Težava pri iskanju molekularnih prilagoditev, ki so osnova izjemno stabilnih vedenjskih sprememb, povezanih z zasvojenostjo, je podobna izzivom, ki se pojavljajo na področju učenja in spomina. Čeprav obstajajo elegantni celični in molekularni modeli učenja in spomina, doslej še ni bilo mogoče identificirati molekularnih in celičnih prilagoditev, ki so dovolj dolgožive, da bi lahko upoštevale zelo stabilne vedenjske spomine. Dejansko je ΔFosB najdaljša prilagoditev, za katero je znano, da se pojavi v možganih odraslih, ne samo kot odziv na zlorabo drog, temveč tudi na katero koli drugo motnjo (ki ne vključuje lezij). Razložila sta se dva predloga, tako na področju zasvojenosti kot na področju učenja in spomina, da bi razložili to neskladje.
Ena od možnosti je, da so bolj prehodne spremembe v izražanju genov, kot so tiste, ki so posredovane z ΔFosB ali drugimi transkripcijskimi faktorji (npr. CREB), lahko povzroči dolgotrajnejše spremembe nevronske morfologije in sinaptične strukture. Na primer, povečanje gostote dendritičnih bodic (zlasti povečanje dvoglavih bodic) spremlja povečana učinkovitost glutamatergičnih sinaps pri hipokampalnih piramidnih nevronih med dolgotrajnim potenciranjem (38-40), in vzporedno z okrepljeno vedenjsko občutljivostjo na kokain, posredovano na ravni srednjih živčnih nevronov iz nucleus accumbens (41). Ni znano, ali so takšne strukturne spremembe dovolj dolgo življenjske, da bi lahko upoštevale zelo stabilne spremembe v obnašanju, čeprav slednje trajajo vsaj mesec dni po umiku zdravila. Nedavni dokazi kažejo na možnost, da je ΔFosB in njegova indukcija Cdk1 eden od posrednikov sprememb v sinaptični strukturi nukleusa accumbens (slika 5), ki jo povzroča zdravilo. Tako je infuzija inhibitorja Cdk4 v nucleus accumbens preprečila sposobnost ponovne izpostavljenosti kokainu za povečanje gostote dendritičnih hrbtenic v tej regiji. To je skladno s stališčem, da Cdk5, ki je obogaten v možganih, uravnava živčno strukturo in rast (glej opombe 5 in 36). Možno je, čeprav nikakor ni dokazano, da takšne spremembe nevronske morfologije lahko presežejo sam signal ΔFosB.
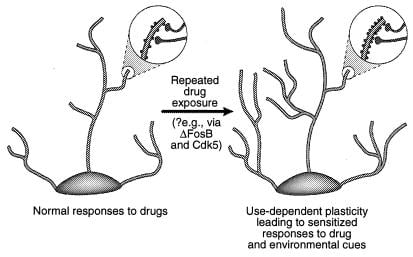
Slika 4
Uravnavanje dendritične strukture z zlorabo drog. Prikazana je ekspanzija dendritičnega drevesa nevrona po kronični izpostavljenosti zlorabi drog, kot je bilo opaženo pri kokainu v nucleus accumbens in prefrontalni skorji (41). Področja povečave kažejo na povečanje števila dendritičnih bodic, kar naj bi se zgodilo v povezavi z aktiviranimi živčnimi konci. To povečanje dendritične gostote hrbtenice se lahko posreduje preko ΔFosB in posledične indukcije Cdk5 (glej besedilo). Takšne spremembe v dendritični strukturi, ki so podobne tistim, ki jih opažamo v nekaterih učnih modelih (npr. Dolgoročno potenciranje), bi lahko posredovale dolgotrajne občutljive odzive na zlorabo drog ali okoljske namige. [Ponovljeno z dovoljenjem ref. 3 (Copyright 2001, Macmillian Magazines Ltd.)].
Druga možnost je, da je prehodna indukcija transkripcijskega faktorja (npr. ΔFosB, CREB) vodi do trajnejših sprememb v izražanju genov s spremembo kromatin. Ti in številni drugi transkripcijski faktorji verjamejo, da aktivirajo ali zavirajo transkripcijo tarčnega gena s pospeševanjem acetiliranja oziroma deacetilacije histonov v bližini gena (42). Čeprav se lahko taka acetilacija in deacetilacija histonov pojavi zelo hitro, je možno, da ΔFosB ali CREB povzročita dolgotrajnejše prilagoditve v encimskem stroju, ki nadzoruje acetilacijo histona. ΔFosB ali CREB lahko tudi spodbujata dolgotrajne spremembe v izražanju genov z uravnavanjem drugih modifikacij kromatina (npr. Metilacije DNA ali histona), ki so bile vpletene v trajne spremembe v transkripciji genov, ki se pojavijo med razvojem (glej opombe 42 in 43) . Čeprav te možnosti ostajajo špekulativne, lahko zagotovijo mehanizem, s katerim prehodne prilagoditve drogam zlorabe (ali kakšno drugo motnjo) vodijo do življenjskih vedenjskih posledic.
Reference
- ↵
- Nestler EJ,
- Hope BT,
- Widnell KL
(1993) Nevron 11: 995 – 1006.
- ↵
- Berke JD,
- Hyman SE
(2000) Nevron 25: 515 – 532.
- ↵
- Nestler EJ
(2001) Nat Rev Neurosci 2: 119 – 128.
- ↵
- Carlezon WA Jr,
- Thome J,
- Olson VG,
- Lane-Ladd SB,
- Brodkin ES,
- Hiroi N,
- Duman RS,
- Neve RL,
- Nestler EJ
(1998) Znanost 282: 2272 – 2275.
- O'Donovan KJ,
- WG Tourtellotte,
- Millbrandt J,
- Baraban JM
(1999) Trendi Neurosci 22: 167 – 173.
CrossRefMedlineSpletna znanost
- ↵
- Mackler SA,
- Korutla L,
- Cha XY,
- Koebbe MJ,
- Fournier KM,
- Bowers MS,
- Kalivas PW
(2000) J Neurosci 20: 6210 – 6217.
- ↵
- Morgan JI,
- Curran T
(1995) Trendi Neurosci 18: 66 – 67.
CrossRefMedlineSpletna znanost
- ↵
- Young ST,
- Porrino LJ,
- Iadarola MJ
(1991) Proc Natl Acad Sci ZDA 88: 1291 – 1295.
- Graybiel AM,
- Moratalla R,
- Robertson HA
(1990) Proc Natl Acad Sci ZDA 87: 6912 – 6916.
- Upam B,
- Kosofsky B,
- Hyman SE,
- Nestler EJ
(1992) Proc Natl Acad Sci ZDA 89: 5764 – 5768.
- ↵
- Kelz MB,
- Nestler EJ
(2000) Curr Opin Neurol 13: 715 – 720.
- ↵
- Koob GF,
- Sanna PP,
- Bloom FE
(1998) Nevron 21: 467 – 476.
- ↵
- Wise RA
(1998) Odvisnost od alkohola v zdravilih 51: 13 – 22.
- ↵
- Hope BT,
- Nye HE,
- Kelz MB,
- Self DW,
- Iadarola MJ,
- Nakabeppu Y,
- Duman RS,
- Nestler EJ
(1994) Nevron 13: 1235 – 1244.
CrossRefMedlineSpletna znanost
- ↵
- Nye H,
- Hope BT,
- Kelz M,
- Iadarola M,
- Nestler EJ
(1995) J Pharmacol Exp Ther 275: 1671 – 1680.
- Nye HE,
- Nestler EJ
(1996) Mol Pharmacol 49: 636-645.
- ↵
- Moratalla R,
- Elibol B,
- Vallejo M,
- Graybiel AM
(1996) Nevron 17: 147 – 156.
- ↵
- Pich EM,
- Pagliusi SR,
- Tessari M,
- Talabot-Ayer D,
- Hooft van Huijsduijnen R,
- Chiamulera C
(1997) Znanost 275: 83 – 86.
- ↵
- Chen JS,
- Nye HE,
- Kelz MB,
- Hiroi N,
- Nakabeppu Y,
- Hope BT,
- Nestler EJ
(1995) Mol Pharmacol 48: 880-889.
- ↵
- Hiroi N,
- Brown J,
- Ye H,
- Saudou F,
- Vaidya VA,
- Duman RS,
- Greenberg ME,
- Nestler EJ
(1998) J Neurosci 18: 6952 – 6962.
- ↵
- Chen J,
- Kelz MB,
- Hope BT,
- Nakabeppu Y,
- Nestler EJ
(1997) J Neurosci 17: 4933 – 4941.
- ↵
- Hiroi N,
- Brown J,
- Haile C,
- Ye H,
- Greenberg ME,
- Nestler EJ
(1997) Proc Natl Acad Sci ZDA 94: 10397 – 10402.
- ↵
- Fienberg AA,
- Hiroi N,
- Mermelstein P,
- Song WJ,
- Snyder GL,
- Nishi A,
- Cheramy A,
- O'Callaghan JP,
- Miller D,
- Cole DG,
- et al.
(1998) Znanost 281: 838 – 842.
- ↵
- Hiroi N,
- Feinberg A,
- Haile C,
- Greengard P,
- Nestler EJ
(1999) Eur J Neurosci 11: 1114 – 1118.
- ↵
- Greengard P,
- Allen PB,
- Nairn AC
(1999) Nevron 23: 435 – 447.
- ↵
- Bibb JA,
- Snyder GL,
- Nishi A,
- Yan Z,
- Meijer L,
- Fienberg AA,
- Tsai LH,
- Kwon YT,
- Girault JA,
- Czernik AJ,
- et al.
(1999) Narava (London) 402: 669 – 671.
- ↵
- Chen JS,
- Kelz MB,
- Zeng GQ,
- Sakai N,
- Steffen C,
- Shockett PE,
- Picciotto M,
- Duman RS,
- Nestler EJ
(1998) Mol Pharmacol 54: 495-503.
- ↵
- Kelz MB,
- Chen JS,
- Carlezon WA,
- Whisler K,
- Gilden L,
- Beckmann AM,
- Steffen C,
- Zhang YJ,
- Marotti L,
- Self SW,
- et al.
(1999) Narava (London) 401: 272 – 276.
- ↵
- Dobrazanski P,
- Noguchi T,
- Kovary K,
- Rizzo CA,
- Lazo PS,
- Bravo R
(1991) Mol Cell Biol 11: 5470-5478.
- Nakabeppu Y,
- Nathans D
(1991) Celica 64: 751 – 759.
- ↵
- Yen J,
- Modrost RM,
- Tratner I,
- Verma IM
(1991) Proc Natl Acad Sci ZDA 88: 5077 – 5081.
- ↵
- Beli FJ,
- Hu XT,
- Zhang XF,
- Wolf ME
(1995) J Pharmacol Exp Ther 273: 445 – 454.
- ↵
- Hyman SE
(1996) Nevron 16: 901 – 904.
CrossRefMedlineSpletna znanost
- Kreek MJ
(1997) Pharmacol Biochem Behav 57: 551 – 569.
- ↵
- Shippenberg TS,
- Rea W
(1997) Pharmacol Biochem Behav 57: 449 – 455.
- ↵
- Chen JS,
- Zhang YJ,
- Kelz MB,
- Steffen C,
- Ang ES,
- Zeng L,
- Nestler EJ
(2000) J Neurosci 20: 8965 – 8971.
- ↵
- Bibb JA,
- Chen JS,
- Taylor JR,
- Svenningsson P,
- Nishi A,
- Snyder GL,
- Yan Z,
- Sagawa ZK,
- Nairn AC,
- Nestler EJ,
- et al.
(2001) Narava (London) 410: 376 – 380.
- ↵
- Luscher C,
- Nicoll RA,
- Malenka RC,
- Muller D
(2000) Nat Neurosci 3: 545 – 550.
- Malinow R,
- Mainen ZF,
- Hayashi Y
(2000) Curr Opin Neurobiol 10: 352 – 357.
- ↵
- Scannevin RH,
- Huganir RL
(2000) Nat Rev Neurosci 1: 133 – 141.
Robinson, TE in Kolb, B. (1999) (1997) EUR. J. Neurosci.11, 1598-1604.
- ↵
- Carey M,
- Smale ST
(2000) Transkripcijska ureditev v evkariontih (laboratorij Cold Spring Harbour Lab. Press, Plainview, NY).
- ↵
- Spencer VA,
- Davie JR
(1999) Gene 240: 1 – 12.
 Facebook
Facebook - Twitter
- Google+
- CiteULike
- Delicious
 Digg
Digg- Mendeley
Članki, ki so gostili članke HighWire Press, navajajo ta članek
- Zakon o naravnih in zdravniških nagradah o mehanizmih skupne nevralne plastičnosti z {Delta} FosB kot ključnim posrednikom J. Neurosci. 2013 33 (8) 3434-3442
- Droge, kriminal in epigenetika hedonistične alostaze Časopis za sodobno kazensko pravosodje 2012 28 (3) 314-328
- Minimalizem
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Minimalizem
- Celotno besedilo (HTML)
- Celotno besedilo (PDF)
- Morfij aktivira E-dvajset-šest-podoben transkripcijski faktor-1 / odzivni faktor serumskega odziva preko zunajceličnih signalov reguliranih kinaz J. Pharmacol. Exp. Ther. 2012 342 (1) 41-52
- Molekularni mehanizem za zdravilo Gateway: Epigenetske spremembe, ki jih je sprožila kokainska genska ekspresija nikotina Sci Transl Med 2011 3 (107) 107ra109
- Okrepljena saharoza in kokain, samonadzorna in sprožena z iskanjem zdravil po izgubi VGLUT2 v nevronih medijev možganskega dopamina v miših J. Neurosci. 2011 31 (35) 12593-12603
- Kronična intermitentna hipoksija zvišuje krvni tlak in izražanje FosB / {Delta} FosB v osrednjih avtonomnih regijah Am. J. Physiol. Regul. Integr. Comp. Fiziol. 2011 301 (1) R131-R139
- Odsotnost GPR37 / PAEL receptorja slabi striatno Akt in ERK2 fosforilacijo, ekspresijo {Delta} FosB in pogojeno prednost pred amfetaminom in kokainom FASEB J. 2011 25 (6)
- Povezava med trajanjem začetne izpostavljenosti alkoholu in obstojnostjo molekularne tolerance je izrazito nelinearna J. Neurosci. 2011 31 (7) 2436-2446
- In vivo slikanje z bioluminiscenco razkriva aktivacijo protein-1 z reguliranim redoksom pri paraventricularnem jedru miši z Renovaskularno hipertenzijo Hipertenzija 2011 57 (2) 289-297
- Striatalna prekomerna ekspresija {Delta} FosB reproducira prisilno gibanje, povzročeno s kronično levodopo J. Neurosci. 2010 30 (21) 7335-7343
- Epigenetska mediacija okoljskih vplivov v večjih psihotičnih motnjah Schizophr Bull 2009 35 (6)
- MRI s sondo na osnovi DNA za specifično odkrivanje kronične izpostavljenosti amfetaminu v živih možganih J. Neurosci. 2009 29 (34) 10663-10670
- Spremenjena plastičnost dendritične hrbtenice pri podganjih, ki so bili odvzeti kokainu J. Neurosci. 2009 29 (9) 2876-2884
- Zaslon s prekomerno ekspresijo v drosofili določa nevronske vloge GSK-3 {beta} / dlakavosti kot regulator AP-1-odvisne razvojne plastičnosti Genetika 2008 180 (4) 2057-2071
- MRI za transkripcijo: nov pogled na živi možgani Nevroznanstvenik 2008 14 (5) 503-520
- {Delta} FosB indukcija v orbitofrontalni skorji posreduje toleranco kognitivne disfunkcije, povzročene s kokainom J. Neurosci. 2007 27 (39) 10497-10507
- Dolgotrajna ranljivost za ponovno vzpostavitev obnašanja metamfetamina v nevrotrofnem faktorju mutantov, pridobljenih iz glijalnih celic FASEB J. 2007 21 (9)
- {Delta} FosB v jedrskem akumulatorju uravnava ojačano instrumentalno vedenje in motivacijo za hrano J. Neurosci. 2006 26 (36) 9196-9204
- Regulacija stabilnosti {Delta} FosB s fosforilacijo. J. Neurosci. 2006 26 (19) 5131-5142
- Ekspresija mutantnih NMDA receptorjev v celicah, ki vsebujejo dopamin D1, preprečuje senzibilizacijo kokaina in zmanjšuje prednost kokaina J. Neurosci. 2005 25 (28) 6651-6657
- D1 Dopaminski receptorji modulirajo {Delta} FosB indukcijo v Striatumu podgane po intermitentni uporabi morfina J. Pharmacol. Exp. Ther. 2005 314 (1) 148-154
- Nevrobiologija miši, izbrana za visoko prostovoljno delovanje na kolesih Integr. Comp. Biol. 2005 45 (3) 438-455
- Vpliv pomanjkanja vode in rehidracije na obarvanje c-Fos in FosB v supraoptičnem jedru podgane in regiji lamina terminalis \ t Am. J. Physiol. Regul. Integr. Comp. Fiziol. 2005 288 (1) R311-R321
- Transkripcijska indukcija gena FosB / {Delta} FosB z mehanskim stresom v osteoblastih J Biol Chem 2004 279 48-49795
- Indukcija {Delta} FosB v strukturah možganov, povezanih z nagrado, po kroničnem stresu J. Neurosci. 2004 24 (47) 10594-10602
- Odmerek gena Sim1 uravnava odziv homeostatskega krmljenja na povečano prehransko maščobo pri miših Am. J. Physiol. Endokrinol. Metab. 2004 287 (1) E105-E113
- Analiza DNA mikromrež z izražanjem genov v astrocitih glave človeškega optičnega živca kot odgovor na hidrostatski tlak Fiziol. Genomika 2004 17 (2) 157-169
- Superoksid je vključen v aktivacijo centralnega živčnega sistema in simpatično ekscitacijo srčnega popuščanja zaradi miokardnega infarkta Circ. Res. 2004 94 (3) 402-409
- Adenozinski receptorji A2A pri nevroadaptaciji na ponavljajočo se dopaminergično stimulacijo: posledice za zdravljenje diskinezij pri Parkinsonovi bolezni Nevrologija 2003 61 (90116) S74-81
- Citoplazmatska Versus Nuklearna lokalizacija Fos-sorodnih beljakovin v žabici, Rana esculenta, testis: In Vivo in neposredni in vitro učinek agonista, ki sprosti gonadotropin Biol. Reprod. 2003 68 (3) 954-960
- Periadolescent Mice kažejo povečano povečanje Delta FosB v odgovor na kokain in amfetamin \ t J. Neurosci. 2002 22 (21) 9155-9159
- Delta FosB uravnava tek kolesa J. Neurosci. 2002 22 (18) 8133-8138
- Aktivnost CREB v lupini nucleus accumbens nadzoruje preoblikovanje vedenjskih odzivov na čustvene dražljaje Proc. Natl. Acad. Sci. ZDA 2002 99 (17) 11435-11440
- Psihogenomika: priložnosti za razumevanje odvisnosti J. Neurosci. 2001 21 (21) 8324-8327