Sprednji Behav Neurosci. 2015 maj 19; 9: 128. doi: 10.3389 / fnbeh.2015.00128. eCollection 2015.
Andreatta M1, Pauli P1.
Minimalizem
V klasičnem pogojevanju se sprva nevtralni dražljaj (pogojeni dražljaj, CS) poveže z biološko pomembnim dogodkom (brezpogojni dražljaj, ZDA), ki je lahko bolečina (averzivno kondicioniranje) ali hrana (apetitivno kondicioniranje). Po nekaj asociacijah lahko CS sproži obrambni oziroma končni odgovor. V nasprotju z averzivnim pogojevanjem se pri ljudeh apetitivno pogojenost redko preiskuje, čeprav je njegov pomen za normalno in patološko vedenje (npr. Debelost, odvisnost) nesporen. Namen te študije je prevesti ugotovitve živali o kondicioniranju apetita ljudem, ki uporabljajo hrano kot ZDA. Triintrideset udeležencev so med 8. in 10. uro zjutraj preiskali brez zajtrka, da bi zagotovili, da se počutijo lačne. V dveh fazah pridobivanja je ena geometrijska oblika (avCS +) napovedovala nenavaden US (boleč električni udar), druga oblika (appCS +) pa apetitni US (čokolada ali slana pereca po želji udeležencev) in tretja oblika (CS– ) napovedal niti ZDA. V fazi izumrtja so bile te tri oblike in nova oblika (NOVO) ponovno predstavljene brez dostave v ZDA. Ocene valence in vzburjenosti ter odzivi na prestrašenost in prevodnost kože (SCR) so bili zbrani kot indeksi učenja. Ugotovili smo uspešno averzivno in apetitivno pogojevanje. Po eni strani je bil avCS + ocenjen kot bolj negativen in bolj vznemirljiv kot CS– in je povzročil presenetljivo okrepitev in izboljšan SCR. Po drugi strani pa je bil appCS + ocenjen bolj pozitivno kot CS– in povzročil dušenje in večji SCR. Če povzamemo, smo pri (lačnih) ljudeh uspešno potrdili ugotovitve živali z dokazovanjem apetitivnega učenja in običajnega averzivnega učenja.
Predstavitev
Napovedovanje nevarnosti in hrane je ključnega pomena za preživetje katerega koli organizma. V klasičnih pogojih (Pavlov, 1927), iztočnica pred averzivnim dogodkom, kot je blag boleči električni šok (averzivni brezpogojni dražljaj, ZDA) ali privlačen dogodek, kot je kroglica s hrano (apetitna ZDA), večkrat. Potem lahko ta spodbuda sama (zdaj označena kot CS, CS +) sproži bodisi obrambne bodisi potrošniške odzive. Prva vrsta asociativnega učenja se imenuje averzivno kondicioniranje, druga pa apetitna kondicija.
Čeprav je napovedovanje apetitnih dogodkov enako pomembno za preživetje kot napovedovanje averzivnih dogodkov, je apetitno kondicijo pri živalih še manj raziskano (Bouton in Peck, 1989; Koch et al., 1996; McDannald in sod., 2011, za pregled glej Martin-Soelch et al., 2007) kot tudi pri ljudeh (Klucken et al., 2009, 2013; Austin in Duka, 2010; Delgado et al., 2011; Levy in Glimcher, 2011). To pomanjkanje raziskav je morda posledica zapletenosti paradigme apetita v primerjavi z averzivno. Na primer, hrano kot primarni ojačevalec je treba oddati, ko je organizem lačen, da se lahko obrestuje (za nedavni pregled glej Dickinson in Balleine, 1994; Clark et al., 2012). V raziskavah na ljudeh smo to težavo premagali z uporabo denarja (Austin in Duka, 2010; Delgado et al., 2011; Levy in Glimcher, 2011) ali erotične slike (Klucken et al., 2009, 2013). Vendar pa nevro-slikovne študije kažejo, da primarni (npr. Prigrizki ali pijače) in sekundarni (tj. Denar) ojačevalci aktivirajo nekatere skupne možganske regije (npr. Striatum), vendar vključujejo tudi različne vzorce aktivacije (Delgado et al., 2011; Levy in Glimcher, 2011).
Kolikor vemo, je le nekaj študij o kondicioniranju ljudi preučilo učinke primarnih apetitnih ojačevalcev, kot je vonjGottfried et al., 2002), voda (Kumar et al., 2008) ali hrano (Prévost et al., 2012). V prvi študiji je dr. Gottfried in sod. (2002) pridruženi nevtralni obrazi (CS) z neprijetnim, prijetnim ali nevtralnim vonjem. Zanimivo je, da so ugotovili večjo aktivacijo v orbitofrontalni skorji (OFC) in v ventralnem striatumu kot odgovor na apetitni CS + proti averzivni CS +. Ugotovili so, da OFC obdeluje vrednost vonja in sodeluje pri prenosu afektivne vrednosti iz vonjave (ZDA) v vidni (CS) sistem. (Ventro) striatalna aktivacija je bila interpretirana kot odraz apetitnega CR, ki ga povzroča apetit CS +. V drugi študiji je dr. Kumar in sod. (2008) povabili svoje udeležence (zdravi nadzorniki in bolniki z veliko depresijo) zgodaj zjutraj v laboratorij in jih pozvali, naj se vzdržijo pitja ponoči, da bi zagotovili, da so žejni pred pregledom. Fraktalne slike so bile CS-ji in 0.1 ml vode v ZDA. Zanimivo je, da so med drugimi aktivacijami zdravi udeleženci (vendar ne bolniki z depresijo) pokazali večjo aktivacijo v ventralnem striatumu na apetitni CS +, kar kaže na to, da je ta spodbuda obravnavana kot koristna. V tretji študiji je dr. Prévost et al. (2012) predstavil tudi fraktalne slike kot CS in sladke ali slane prigrizke kot ZDA glede na želje udeležencev. CS je bil predstavljen 6 s, v zadnji sekundi pa je bila dodatno predstavljena tudi slika hrane. Vsakič, ko se je prikazala slika s hrano, je preizkuševalec položil kos hrane udeležencem v roke, ki so jim prigrizek takoj dovolili. Na žalost avtorji v fazi klasične kondicije niso poročali o aktivaciji možganov apetitivnemu CS +; vendar so opazili manjši srčni pojemek na nagrajeni CS + v primerjavi z nenagrajenim CS–, kar kaže na diferenčne učinke kondicioniranja.
Pomanjkanje študij, ki bi uporabile zaznavni odziv kot indeks za apetitno kondicijo, je presenetljivo, zlasti če upoštevamo njegovo široko uporabo v averzivnem kondicioniranju. Odziven zagon je odziv prednikov in samodejen obrambni odziv na nenadne, nepričakovane in močne averzivne dogodke (Koch, 1999). Ta obrambni odziv posreduje razmeroma enostavna pot nevronov, ki vključuje kohlearne koreninske nevrone, kaudalno pontinsko jedro retikularne formacije (PnC) in hrbtenične motonevrone (Fendt in Fanselow, 1999; Koch, 1999). Študije na živalih so razkrile, da je potenciranje zagona odvisno od projekcij od amigdale do PnC (Fendt in Fanselow, 1999; Koch, 1999), ker je slabljenje zagona odvisno od nepoškodovanega jedra jedra (NAcc, Koch, 1999). Takšna modulacija zagonskega odziva je koristen implicitni ukrep za valenco predstavljenih prednjih dražljajev. Tako potenciranje kaže na negativno valenco, medtem ko slabljenje kaže na pozitivno valenco, oboje, ne da bi močno vplivali na kognitivne procese (Hamm in Weike, 2005; Andreatta in sod., 2010). Kolikor vemo, je le ena študija na živalih preučila apetitno kondicijo, tako da je merila odzivne rezultate kot odvisen ukrep. Zanimivo je, da so preučene podgane pokazale slabljenje zagona na CS + (tj. Svetlobo), povezano z izdajo raztopine saharoze. Zlasti je bilo takšno slabljenje oslabljeno pri živalih z lezijami NAcc, ne pa pri tistih z amigdalanimi lezijami, kar kaže na to, da ima NAcc posebno vlogo pri pridobivanju apetitnih CR in pri slabljenju odzivnosti zagona (Koch et al., 1996).
Trenutno je v naši raziskavi prva prevajalska paradigma tega pogovora na človeka prevedena s primarnimi ojačevalci kot ameriška, torej sladka (čokolada Smarties®) ali slana hrana (majhna slana perečka) in modulacija začudenja kot merilo CR. Pričakovali smo, da bo appCS + sprožil močne privlačne CR, kar se odraža v slabljenju zagona, povečanem SCR in pozitivni valenčni oceni v primerjavi z drugimi dražljaji, tj. AvCS + in CS–.
Materiali in metode
udeleženci
Dvainštirideset prostovoljcev je sprejelo sodelovanje v študiji in prejelo tečaj. Iz analize je bilo izključenih devet udeležencev, dva zaradi tehničnih težav, trije, ker so bili kodirani kot neodzivni (povprečna amplituda vznemirjenja <5 μV), in štirje, ker niso pokazali dovolj odzivnih odzivov na pogoj (najmanj = 2; za podrobnosti, glej Materiali in metode). Na koncu je bilo v analizo vključenih 33 udeležencev (16 moških; povprečna starost: 22.09 let, SD: 2.84; območje: 18 – 29 let). Štirje udeleženci niso bili domači Nemci, šest pa levičarjev. En udeleženec v celotnem poskusu ni vedel združenja CS-USs (glej Postopek), vendar smo se odločili, da tega ne izključimo, ker so bili njegovi odzivi normalni in niso vplivali na rezultate.
materiali
Brezpogojni Stimuli (ZDA)
Uporabljeni sta bili dve vrsti ZDA. Kot averzivni ZDA smo na ne dominantno podlaket udeležencev nanesli rahlo boleč električni udar. Električni udar je bil izveden s pomočjo dveh elektrod s premerom 9 mm in razmikom 30 cm. Električni dražljaj je bil sestavljen iz impulznega impulza s frekvenco 50 Hz in trajanjem 200 ms, ki ga ustvari trenutni stimulator (Digitimer DS7A, Digitimer Ltd, Welwyn Garden City, Velika Britanija, 400 V, največ 9.99 mA). Intenzivnost električnega udara je bila določena posamično s predhodno opisanim pragom (Andreatta in sod., 2010). Na kratko so udeleženci doživeli dve seriji naraščajoče in padajoče intenzitete v intervalih 0.5 mA. Vsak dražljaj so morali oceniti na vizualni lestvici, od 0 (sploh ne čutijo ničesar) do 10 (res intenzivna bolečina) z 4 kot sidrom za prag (samo opazna bolečina). Srednja intenzivnost električnega dražljaja je bila 2.12 mA (SD = 0.56) in je bil ocenjen kot boleč (M = 6.45, SD = 1.73). Apetitna ZDA je bila sestavljena bodisi iz čokolade (Smarties®) ali majhna slana pereca. Izbira apetitivnega ZDA je bila odvisna od individualnih preferenc udeleženca, kot so poročali med predhodnim intervjujem. Udeleženci so namreč morali sporočiti, ali med zajtrkom običajno jedo slano ali sladko hrano. Prav tako so lahko svobodno izbirali, ali imajo med samim poskusom raje čokolado ali slano pereco. Na koncu je 22 udeležencev izbralo čokolado in 11 majhnih slanih perecev.
Pogojeni Stimuli (CS)
Geometrijske oblike (modri kvadrat, rumen krog, zeleni trikotnik, rdeči šesterokotnik) z diagonalo 8 cm so bile predstavljene kot CS. Oblike so bile predstavljene na sredini črnega računalniškega zaslona za 8 s. Ena oblika (avCS +) je bila vedno povezana z averzivnimi ZDA (boleč električni šok), ena oblika (appCS +) je bila vedno povezana z apetitnimi ZDA (čokolada ali slana perečka), ena oblika (CS–) pa ni bila nikoli povezana niti z averzivno ali apetitne US, četrta oblika (NOVA) je bila predstavljena v fazi izumrtja, vendar ne v fazi pridobivanja, da se zagotovi njena nevtralnost.
Sonda za zagon
Kot sondo smo uporabili beli šum 103 dB s trajanjem 50 ms. Akustični dražljaji so bili predstavljeni binauralno prek slušalk in so se naključno pojavili 4–6 s po nastopu oblike.
Vprašalniki
Pred poskusom in po njem so morali udeleženci izpolniti nemške različice popisa anksioznosti države (STAI, Laux in sod., 1981) in razpored pozitivnih negativnih vplivov (PANAS, Krohne in sod., 1996). STAI je inventar za oceno lastnosti udeležencev in / ali stanja tesnobe, ki ga sestavlja 20 predmetov tako za lastnost kot za različico države. Anksioznost udeležencev pred (M = 37.06, SD = 7.80) in po (M = 39.33, SD = 9.16) se poskus ni bistveno spremenil [t(32) = 1.61, p = 0.117]. Rezultat tesnobe pri lastnostih v trenutnem vzorcu se je gibal med 20 in 58 (M = 36.6, SD = 8.98), kar je primerljivo z objavljenim normalnim razponom odraslih (Laux in sod., 1981). PANAS (Krohne in sod., 1996) je indeks pozitivnega in negativnega razpoloženja. Posamezniki z visokimi ocenami na lestvici pozitivnih vplivov (PAS) so nagnjeni k čustvom, kot je navdušenje, medtem ko so posamezniki z visokimi ocenami na lestvici negativnih vplivov (NAS) nagnjeni k čustvom, kot je stiska. Vsak predmet je sestavljen iz pridevnika, udeleženci pa na lestvici navedejo od 1 (zelo rahlo) do 5 (zelo), v kolikšni meri pridevnik odraža njihova občutja v določenem trenutku. V negativnem vplivu udeležencev kot rezultat eksperimenta niso bile ugotovljene nobene pomembne razlike [začetek: M = 11.67, SD = 2.29; konec: M = 12.88, SD = 4.69; t(32) = 1.55, p = 0.130]. Nekako so udeleženci že od začetka bistveno spremenili svoje pozitivno razpoloženje (M = 26.72, SD = 4.70) do konca (M = 23.66, SD = 7.02) preizkusa [t(31) = 3.11, p = 0.004]. To zmanjšanje pozitivnega razpoloženja udeležencev je bilo lahko povezano z neprijetnostjo paradigme (predstavljeni so bili boleči električni šoki in averzivni beli šum).
Postopek
Po prihodu v laboratorij so udeleženci prebrali in podpisali obrazec s informiranim soglasjem, ki ga je odobril etični odbor Oddelka za psihologijo Univerze v Würzburgu. Niso bili obveščeni o nepredvidenih dogodkih med CS-ji in ZDA. Po izpolnitvi vprašalnika smo elektrodi pritrdili in postopek praga bolečine izvedli, kot je opisano zgoraj.
Med faza privajanja, štiri geometrijske oblike so bile predstavljene dvakrat z inter-preskusnim intervalom (ITI), ki se spreminja med 18 in 25 s (povprečje: 21.5 s). V tej fazi niso bile dobavljene nobene ameriške ali zagonske sonde.
Pred fazo pridobivanja je bilo vsakih 7 – 15 s oddanih sedem udarcev belega hrupa, da se zmanjša začetna reaktivnost zagona.
Naslednja dva faze pridobivanja sta bila identična (slika 1). Vsaka faza pridobivanja je bila sestavljena iz preskusov 24: preskušanja 8 CS–, preskušanja 8 avCS + in preskušanja 8 appCS +. CS-zaporedje je bilo psevdo naključno, edina omejitev je bila, da istega dražljaja ni mogoče predstaviti več kot dvakrat zapored. Zlasti je bil avCS + predstavljen skupaj s strelo kot simbolom električnega udara in boleča ZDA je bila dostavljena ob izravnavi. Aplikacija appCS + je bila predstavljena v obliki slike Smarties ali slanega pereca in udeleženec je lahko iz kozarca izbral Smarties ali perece. CS– je bil predstavljen skupaj s simbolom prepovedi in ZDA niso bile dostavljene. Med tremi predstavitvami 8 CS vsake vrste je bila sprožena sonda za zagon med 4 in 6 s po začetku spodbude. Med ITI-ji so bile predstavljene tri dodatne sonde za zagon, da bi zagotovili njihovo nepredvidljivost in zmanjšali navadne navade. ITI, sestavljen iz črnega zaslona, se je spreminjal med 18 in 25 s s povprečjem 21.5 s.
SLIKA 1
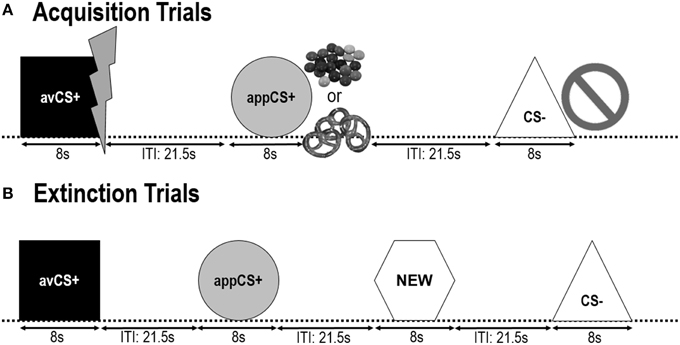
Slika 1. Preskusi med dvema fazama pridobivanja (A) in fazo izumiranja (B). Udeleženci so izvedeli, da ena oblika (avCS +) napoveduje blag boleč električni udar, ena oblika (appCS +) je napovedovala bodisi košček čokolade ali majhen slani peterelj (po njihovi želji), tretja oblika (CS–) pa ni predvidela nobenega biološko viden dogodek. Vsaka oblika je bila predstavljena skupaj s sliko, ki prikazuje električni udar, smarties / slano pereco ali nič, odvisno od povezanosti z ZDA. V fazi izumrtja so bile tri geometrijske oblike še enkrat predstavljene, vendar ZDA niso bile dostavljene. Poleg tega je bila kot nevtralna kontrola predstavljena še četrta geometrijska oblika (NOVO).
Med faza izumiranja, so udeleženci videli tri geometrijske oblike (tj. avCS +, appCS + in CS–) znova in novo nevtralno obliko (NOVO). ZDA niso bile dobavljene in oblike niso bile predstavljene skupaj s slikami bliskavic, čokoladnih / slanih perelov ali prepovedi. Vsak dražljaj je bil predstavljen osemkrat v psevdonamernem vrstnem redu (tj. Isti dražljaj ni bil predstavljen več kot dvakrat zapored), kar je ustvarilo preskuse 32. Med 4 iz predstavitev 8 spodbujevalcev vsake vrste CS so bili predstavljeni spodbujevalni stimuli. Tako kot v fazi pridobivanja in privajanja, so se tudi ITI spreminjale med 18 in 25 s, tudi dodatne sonde za zagon 4 so bile med ITI nepredvidljivo dobavljene.
Po vsaki fazi so udeleženci ocenili valenco (prijetnost) in vzburjenost (vznemirjenost) CS-jev z vizualno analognimi lestvicami (VAS), ki segajo od 1 do 9. Lestvica odstopanj se je gibala od "1", kar pomeni "zelo neprijetno" do "9", kar kaže na "zelo prijetno;" lestvica vzburjenja se je gibala od "1", kar kaže na "umirjeno" do "9", kar kaže na "vznemirljivo". Poleg tega so bile ocene nesreč ocenjeno po dveh fazah pridobivanja in fazi izumrtja. Udeleženci so videli 1 s geometrijsko obliko, nato pa so morali navesti, ali je ta oblika povezana z električnim udarom, s čokoladno / slano pereco, brez ničesar, ali se niso mogli povezati. Zlasti so se vsi udeleženci (razen enega) zavedali nepredvidljivih dogodkov po pridobitvi 2.
Zmanjšanje podatkov
Fiziološke odzive smo zabeležili z ojačevalnikom V-Amp 16 in programsko opremo Vis-Recorder V-Amp Edition (različica 1.03.0004, BrainProducts Inc., München, Nemčija). Uporabljena sta bila frekvenca vzorčenja 1000 Hz in zarezni filter 50 Hz. Analize brez povezave so bile izvedene z Brain Vision Analyzer (različica 2.0; BrainProducts Inc., München, Nemčija).
Začuda odziv
Odziv za zagon smo merili z elektromiografijo (EMG) na levi strani orbicularis oculi mišice z dvema 5 mm Ag / AgCl elektrodama. V skladu s smernicami (Blumenthal et al., 2005), ena elektroda je nameščena pod zenico, druga pa 1 cm bočno. Ozemljitvene in referenčne elektrode so bile postavljene na desni in levi mastoid. Pred pritrditvijo elektrod smo kožo rahlo podrgnili in očistili z alkoholom, da ohranimo impedanco pod 10 kΩ. Elektromiografski signal je bil filtriran brez povezave s filtrom z nizkim izklopom 28 Hz in filtrom z visokim izrezom 500 Hz. Nato je bil signal EMG odpravljen in dano je drsno povprečje 50 ms. Pred začetkom sonde za zagon kot osnovno črto smo uporabili ms 50 (Grillon in sod., 2006). Odzive na sonde za zagon so bili ocenjeni ročno, poskusi s prevelikimi izhodiščnimi premiki (± 5 μV) ali artefakti gibanja pa so bili izključeni iz nadaljnje analize. Odzivi na zagon nižji od 5 μV so bili kodirani kot nič in upoštevani za izračun velikosti zagona (Blumenthal et al., 2005). Skupno je bilo zavrnjenih 10.4% preskušanj, za fazo izumrtja pa je bilo potrebno najmanj 2 od zagonskih odzivov 3 v fazah pridobivanja in 4 iz zagonskih odzivov 8 v fazi izumrtja, da bi udeleženca ohranili v zbirki analiz. Zaradi tega so bili izključeni štirje udeleženci. Amplituda vrha je bila določena kot največji vrh glede na izhodišče v časovnem oknu 20 – 120 ms po zagonu sonde. Surovi podatki so bili potem normalizirani znotraj subjektov z uporabo z- ocene in nato T-ocene, da bi zmanjšali vpliv spremenljivosti posameznika in bolje zaznali psihološke procese. T-ocene so povprečili za vsak pogoj (avCS +, appCS +, CS–, NEW in ITI). Za raziskavo potenciranja zagona ali slabljenja zagona so bile ocene za odzive zagona ITI odštete od odzivov zagona za vsak pogoj.
Odziv kožne prevodnosti (SCR)
Odziv na prevodnost kože (SCR) smo zabeležili z uporabo dveh 5 mm Ag / AgCl elektrod, nameščenih na dlani nedominantne roke. Galvanski odziv je bil filtriran brez povezave s 1 Hz visokoreznim filtrom. SCR je bil opredeljen kot razlika (v μS) med začetkom odziva (1 – 3 s po pojavu dražljaja) in vrhom odziva (Tranel in Damasio, 1994; Delgado et al., 2011). Preskušanja, ki vsebujejo startne sonde, niso bila upoštevana v analizi SCR. Odzivi pod 0.02 μS so kodirani kot nič. Pet nadaljnjih udeležencev je bilo izključenih iz analize SCR, ker je imel povprečni SCR nižji od 0.02 μS. Podatki prevodnosti surove kože so bili spremenjeni s kvadratnim korenom, da se je normalizirala porazdelitev, rezultati pa so se povprečno izračunali za vsako stanje posebej za dve fazi pridobivanja (avCS +, appCS +, CS–) in fazo izumiranja (avCS +, appCS +, CS– in NOV ).
Statistična analiza
Vsi podatki so bili analizirani s SPSS za Windows (različica 20.0, SPSS Inc.). Za fiziološke odzive smo izračunali ločene multivariatne analize variance (ANOVA) za dve fazi pridobivanja in fazo izumiranja. ANOVA za faze prevzema je imela spodbujevalne dejavnike (avCS +, appCS +, CS–) in fazo (nakup 1, nakup 2) kot dejavnike znotraj subjektov. ANOVA za fazo izumrtja je imela kot dejavnik znotraj subjektov le dražljaj (avCS +, appCS +, CS–, NOV). Ocene valence, vzburjenja in nepredvidljivih dogodkov so bile analizirane z ločenimi ANOVA, ki vsebujejo spodbujevalni dejavnik znotraj subjektov (avCS +, appCS +, CS– in NOV) in fazo. Ta faktor je imel štiri stopnje ocenjevanja valenc in vzburjenja (T1: po habituacijski fazi, T2: po prvi fazi pridobivanja, T3: po drugi fazi pridobivanja, T4: po fazi izumrtja), vendar tri stopnje za oceno ob nepredvidljivih dogodkih (T1: po prva faza pridobivanja, T2: po drugi fazi pridobivanja, T3: po fazi izumrtja).
Za vse analize je bila vrednost alfa (α) nastavljena na 0.05. Velikost učinka je navedena kot delna η2.
Rezultati
Ocene valencnosti in vzburjenja za vsako fazo so prikazane na sliki 2; odzivi zagona in SCR so prikazani na sliki 3.
SLIKA 2
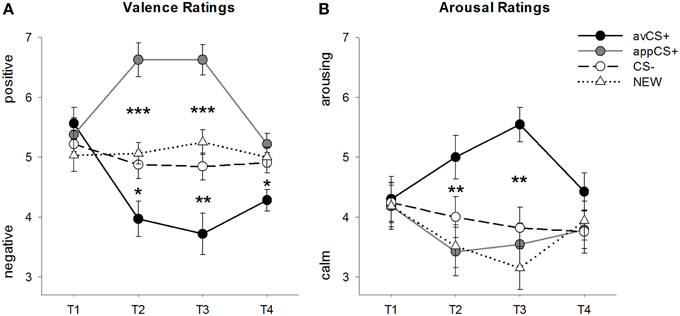
Slika 2. Ocene za valenco (A) in vzburjenje (B). Vrstice (s standardnimi napakami) prikazujejo ocene po fazi habituacije (T1), pridobitev 1 (T2), pridobitev 2 (T3) in fazo izumiranja (T4). Averzivni CS + (črna trdna črta) je po obeh fazah pridobivanja pridobil negativno valenco in močno vzburjenje v primerjavi s CS– (črna črtkana črta) in NOVO (črna črtkana črta). Pomembno je, da je privlačen CS + (siva trdna črta) pridobil pozitivno valenco v primerjavi s CS in NEW. *p <0.05, **p > 0.01, ***p <0.001.
SLIKA 3
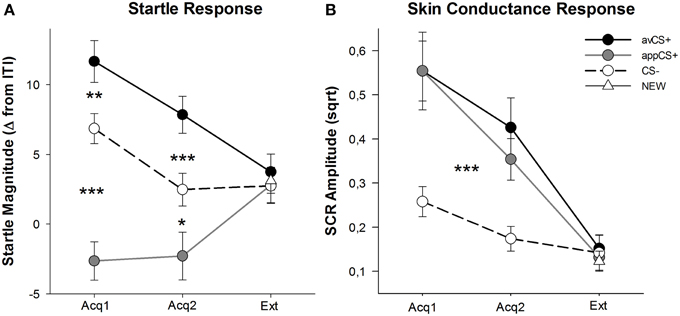
Slika 3. Začetni odzivi (A) in kožna prevodnost (B) (s standardnimi napakami) med prvo fazo pridobivanja (Acq1), drugo fazo pridobivanja (Acq2) in fazo izumiranja (Ext). Ustrezni odzivi so bili v potencialnem potencialu na averzivni CS + (črna trdna črta) in znatno oslabljeni na apetitni CS + (siva trdna črta) v primerjavi s CS - (črna črtkana črta) v fazah pridobivanja. SCR je bil bistveno večji od avCS + in appCS + v primerjavi s CS–. V fazi izumrtja niso bile ugotovljene razlike. *p <0.05, **p > 0.01, ***p <0.001.
Ocene
ANOVA za valenčne ocene med pridobitvijo so pokazali pomembne glavne učinke dražljaja [F(3, 93) = 17.26, GG-ε = 0.801, p <0.001, delno η2 = 0.358] in faza [F(3, 93) = 3.30, GG-ε = 0.731, p = 0.039, delno η2 = 0.096] kot tudi pomembna interakcija med dražljajem in fazo [F(9, 279) = 9.54, GG-ε = 0.463, p <0.001, delno η2 = 0.235]. Spremljanje t-testi kažejo, da so bile valence štirih geometrijskih oblik na začetku eksperimenta enake (ps> 0.19), medtem ko je bil po pridobitvi 1 in pridobitvi 2 avCS + ocenjen kot posebej negativen, appCS + pa kot posebej pozitiven. Natančneje, avCS + je bil ocenjen kot bolj negativno valenciran v primerjavi s CS– [Acq1: t(31) = 2.34, p = 0.026; Acq2: t(31) = 3.07, p = 0.004], NOVO [Acq1: t(31) = 2.70, p = 0.011; Acq2: t(31) = 3.89, p <0.001] in appCS + [Acq1: t(31) = 5.41, p <0.001; Acq2: t(31) = 6.11, p <0.001]. AplikacijaCS + je bila ocenjena kot bistveno bolj pozitivna kot CS– [Acq1: t(31) = 4.99, p <0.001; Acq2: t(31) = 5.31, p <0.001] in NOVO [Acq1: t(31) = 4.92, p <0.001; Acq2: t(31) = 4.14, p <0.001]. Razlike med CS– in NEW nikoli niso bile pomembne (ps> 0.18).
Pomembno je, da nismo primerjali pomembnih razlik pri primerjavi ocen valencnosti za aplikacijo APCS +, povezano s čokolado in slano pereco [Acq1: t(30) = 0.03, p = 0.477; Acq2: t(30) = 0.29, p = 0.775].
Po fazi izumrtja je bil avCS + še vedno ocenjen kot bolj negativen v primerjavi s CS– [t(31) = 2.40, p = 0.023], NOVO [t(31) = 3.40, p = 0.002] in appCS + [t(31) = 3.35, p = 0.002]. Nasprotno, valenca appCS + se ni več razlikovala od CS– [t(31) = 1.77, p = 0.086] ali NOVO [t(31) = 1.07, p = 0.293].
ANOVA za ocene vzburjenja med pridobitvijo je pokazal pomemben glavni učinek spodbude [F(3, 96) = 7.07, GG-ε = 0.737, p = 0.001, delno η2 = 0.181], vendar ne iz faze [F(3, 96) = 1.27, GG-ε = 0.805, p = 0.289, delno η2 = 0.038] in pomembna interakcija med dražljajem in fazo [F(9, 288) = 4.53, GG-ε = 0.582, p = 0.001, delno η2 = 0.124]. Spremljanje t-testi niso pokazali bistvenih razlik med dražljaji glede njihovega začetnega vzburjenja (ps> 0.74). Po dveh fazah pridobivanja pa je bil avCS + ocenjen kot bolj vznemirljiv kot NOVO [Acq1: t(32) = 2.99, p = 0.005; Acq2: t(32) = 5.97, p <0.001] in appCS + [Acq1: t(32) = 2.62, p = 0.013; Acq2: t(32) = 4.42, p <0.001] in nekoliko bolj vznemirljivo kot CS - po prvi fazi pridobivanja [t(32) = 1.96, p = 0.058], vendar bistveno bolj vznemirljivo po drugi fazi pridobivanja [t(32) = 3.65, p = 0.001]. V nasprotju z bonitetno oceno se vzburjenje appCS + ni razlikovalo od vzburjenja CS - in NOV (ps > 0.13) po fazah prevzema.
Podobno kot ocene ranljivosti se tudi CS– in NEW pri vzburjenju nista razlikovala (ps> 0.07).
Kot je veljalo za valenco, ni bilo razkritih razlik za appCS +, ki je povezan s čokolado, in appCS +, povezan s slano pereco [Acq1: t(31) = 0.26, p = 0.797; Acq2: t(31) = 0.33, p = 0.724].
Po fazi izumrtja niso bile ugotovljene pomembne razlike v ocenah vzburjenja (ps> 0.08).
Za dodatno post-hoc t-testov, ki primerjajo ocene po habituacijski fazi, prvi in drugi fazi pridobivanja ter fazi izumrtja, glej Dodatna gradiva.
Začuda odziv
ANOVA za faze pridobivanja je vrnila glavni učinek spodbude [F(2, 64) = 49.92, GG-ε = 0.964, p <0.001, delno η2 = 0.609], ne pa faza [F(1, 32) = 3.16, p = 0.085, delno η2 = 0.090] in pomemben stik Stimulus × Faza [F(2, 64) = 3.37, GG-ε = 0.875, p = 0.048, delno η2 = 0.095]. Spremljanje t-testi so pokazali pomemben potencial za zagon avCS + v primerjavi s CS– med obema prvoma [t(32) = 3.27, p = 0.003] in drugo [t(32) = 4.00, p <0.001] faze pridobivanja. Presenetljivi odzivi na avCS + so bili tudi znatno okrepljeni v primerjavi z odgovori na appCS +, spet po obeh pridobitvah 1 [t(32) = 8.20, p <0.001] in pridobitev 2 [t(32) = 5.74, p <0.001]. Pomembno je, da je bila presenetljiva velikost appCS + znatno oslabljena v primerjavi s CS– med obema prevzemoma 1 [t(32) = 6.34, p <0.001] in pridobitev 2 [t(32) = 2.91, p = 0.007]. Ponovno in v skladu z ocenami ni bilo odkritih diferencialnih odzivov za zagon za appCS +, če so povezani s čokolado ali s slano pereco [Acq1: t(31) = 1.04, p = 0.309; Acq2: t(31) = 0.07, p = 0.947]. V fazi izumrtja ni bilo ugotovljenega pomembnega učinka [F(3, 96) = 0.26, GG-ε = 0.906, p = 0.833, delno η2 = 0.008].
Odziv kožne prevodnosti (SCR)
Od ANOVA za SCR med dvema fazama pridobivanja so bili glavni spodbujevalni učinki [F(2, 54) = 18.04, GG-ε = 0.908, p <0.001, delno η2 = 0.401] in faza [F(1, 27) = 20.91, p <0.001, delno η2 = 0.436] se je izkazalo za pomembno, vendar ni njihova interakcija [F(2, 54) = 0.68, GG-ε = 0.637, p = 0.451, delno η2 = 0.024]. Post-hoc t-testi kažejo bistveno večji SCR na avCS + [t(27) = 6.46, p <0.001] in v aplikacijoCS + [t(27) = 4.84, p <0.001] v primerjavi s CS–, medtem ko so udeleženci pokazali primerljiv SCR z avCS + in appCS + [t(27) = 0.64, p = 0.527]. Zlasti ni bilo ugotovljenih razlik v SCR-jih v aplikaciji čokoladeCC + in v aplikaciji slanega perecaCC + [Acq1: t(26) = 2.55, p = 0.120; Acq2: t(26) = 1.29, p = 0.210]. Kot je veljalo za ocene in odziv na zagon, za fazo izumrtja niso našli pomembnih učinkov [F(3, 81) = 0.28, GG-ε = 0.634, p = 0.743, delno η2 = 0.010].
Razprava
Cilj te študije je bil prevesti ugotovitve živali na človeka z uporabo klasične apetitne kondicijske paradigme s primarnim ojačevalcem kot brezpogojno spodbudo (ZDA). Da bi to storili, so udeleženci prišli v laboratorij zgodaj zjutraj brez zajtrka, da bi zagotovili, da so lačni, zato so po svojih željah kot čokolado uporabili koščke čokolade ali slane perece. Med fazo pridobivanja je ena geometrijska oblika (avCS +) postala povezana z rahlo bolečim električnim udarcem (averzivni ZDA), druga oblika (appCS +) z apetitnimi ZDA in tretja oblika (CS–), ne da bi imeli privlačne ZDA niti averzivni ZDA. Rezultati kažejo na uspešno averzivno in apetitno kondicijo na izrecni verbalni ravni (tj. Ocene), na implicitni vedenjski ravni (tj. Zaznavni odziv) in na fiziološki ravni (tj. SCR). Konkretno, avCS + je v primerjavi s CS-jem ustvaril več negativnih ocen valencnosti, višje ocene vzburjenja, potenciranje zagona in večji SCR. Najpomembneje je, da je appCS + v primerjavi s CS-jem sprožil več pozitivnih valenčnih ocen, slabljenje zagona in večji SCR. Naše ugotovitve o averzivnem kondicioniranju so bile v skladu s pričakovanji, saj so v prejšnjih raziskavah ugotovili, da je spodbuda, ki napoveduje grožnjo (avCS +), ocenjena kot averzivno, povzroča večje odzive na strah in povečuje fiziološko vzburjenje (Fendt in Fanselow, 1999; Hamm in Weike, 2005; Andreatta in sod., 2010, 2013). Naši rezultati glede apetitne kondicije so bili tudi v skladu s prejšnjimi raziskavami na ljudeh in živalih, ki kažejo, da je spodbuda, ki napoveduje nagrado (appCS +), ocenjena kot pozitivna, zavira odzive na strah in povečuje fiziološko vzburjenje (Koch et al., 1996; Gottfried et al., 2002; Kumar et al., 2008; Klucken et al., 2009, 2013; Austin in Duka, 2010; Prévost et al., 2012). Kolikor vemo, je to prva študija, ki je pokazala, da je pri ljudeh pogojeno slabljenje zagona na spodbudo, ki napoveduje osnovno nagrado. Pomembno je, da smo lahko prenesli in potrdili rezultate študije na živalih (Koch et al., 1996). Ta študija na živalih je pokazala, da je oslabitev zaleta pri podganah odvisna od projekcij od NAcc (del ventralnega striatuma) do PnC. Zato bi oslabitev zagona v naši študiji lahko pomenila aktivnost NAcc, kar je tudi v skladu z ugotovitvami fMRI (Gottfried et al., 2002; Kumar et al., 2008; Klucken et al., 2009, 2013; Delgado et al., 2011; Levy in Glimcher, 2011). Zato zaključujemo, da je bila naša pozitivna kondicijska paradigma uspešna, saj sta kazali tako eksplicitna (ocena) kot implicitna (slabljenje zagona) pozitivna valenca.
Poleg te nove, a dokaj predvidljive ugotovitve, velja omeniti še dva zanimiva rezultata. Prvič, verbalni in fiziološki odzivi na vzburjenje na appCS + disociirani. Drugič, ugotovili smo hitrejše izumrtje privlačnih CR v primerjavi z averzivnimi CR.
SCR označuje aktivacijo simpatikusa, ki se poveča tako na spodbudo, povezano z nagrajevanjem (appCS +), kot na spodbudo, povezano z grožnjo (avCS +). Ta rezultat je v skladu s prejšnjo kondicijsko študijo, v kateri so bile erotične slike uporabljene kot privlačne ZDA (Klucken et al., 2013), in študijo, ki razkriva primerljiv SCR na iztočnici, ki napoveduje denar, in na iztočnici, ki napoveduje odporni hrup (Austin in Duka, 2010). Zlasti je SCR orientacijski odziv, povezan z aktiviranjem simpatičnega sistema. Ta odziv je bil predlagan, da odraža pripravo vedenjske reakcije na motivacijsko vidne dogodke (Bradley, 2009). Na podlagi tega mislimo, da so spodbude, povezane z grožnjo in z nagradami, sprožile pripravljalni odziv za zaporedne vedenjske odzive. Z drugimi besedami, hrana in bolečina sta sprožila pripravo na pristop in izogibanje vedenju. V nasprotju z veliko fiziološko vzburjenostjo so verbalni odzivi kazali na nizko vzburjenost zaradi spodbude, povezane z nagrado. Mogoče bi lahko na verbalno vzburjenje bolj vplivala narava ZDA, ne pa fiziološka aktivacija po sebi. V resnici menimo, da so bili appCS + proti avCS + ocenjeni kot nizki in zelo vznemirljivi, ker so bili povezani z nizko in zelo vznemirjajočimi ZDA. Žal nismo zbirali ocene vzburjenja za ZDA, zato bodo prihodnje študije, da to hipotezo izrecno preizkusijo.
V fazi izumrtja ni bilo dobavljenih ZDA. To je morda povzročilo novo zaviralno učenje, imenovano izumrtje (za pregled glej Milad in Quirk, 2012), tako da se averzivni in apetitni CR posledično zmanjšujejo. Očitno na vedenjskih (tj. Odzivnem zagonu) in na fizioloških (tj. SCR) ravneh ni bilo več zaznati diferencialnih reakcij na avCS +, appCS + in CS–. Podobno je bilo izrecno vzburjenje pogojenih dražljajev, ocenjenih po fazi izumrtja, izenačeno na nizki ravni, kar kaže na uspešno učenje izumrtja. Vendar pa je avCS + še vedno ocenjen bistveno bolj negativno kot CS–, medtem ko se valenca appCS + ni več razlikovala od valencije CS. Počasnejše izumrtje averzivnega eksplicitnega odziva je lahko posledica evolucijskega konzervativizma, kar pomeni, da je signale o grožnji še posebej težko pozabiti, ker lahko neodziv na signal ogroženosti ogrozi življenje.
Še vedno ostaja eno vprašanje: zakaj je potem odzivnost zagona (tj. Implicitna valenca) v fazi izumrtja popolnoma ugasnila? Najprej so bili ti odzivi izračunani skozi celoten potek faze. Zato je možno, da bi bilo mogoče diskriminatorne CR še vedno zaznati med prvimi poskusi faze izumiranja. Na raziskovalni način smo sledili tej hipotezi in si ogledali tako zastrašujoče odzive kot tudi SCR v celotni fazi izumrtja (glej Dodatno gradivo). Čeprav nismo našli bistvenih razlik, smo za avCS + opazili nekoliko večjo velikost zagona v primerjavi s CS- in novimi kontrolnimi spodbudami. Opazili smo tudi nekoliko večjo slabljenje zagona na appCS + v primerjavi s CS - in novim spodbujevalnim spodbudom na samem začetku faze izumiranja, ki pa je v nekaj preskusih izginila. SCR na appCS + je padel že po preskusu iztrebljanja 2nd, medtem ko je SCR na avCS + ostal skoraj višji za skoraj vse izumrtje v primerjavi s SCR na NOVO. Čeprav bi morali te rezultate razlagati zelo previdno, se zdi, da so se odmevni odzivi vzporedno z valenci. Poleg tega tako osupljivi odzivi kot tudi SCR nadalje podpirajo idejo evolucijskega konzervativizma proti dražljajem groženj.
Nazadnje moramo priznati nekatere omejitve te študije. Prvič, zaradi tehnične težave nismo mogli poročati o oceni prijetnosti (in vznemirljivosti) za čokolado in slano pereco. Vendar apetitni CR kažejo, da so udeleženci resnično doživeli dve ZDA kot apetitne. Drugič, trajanje averzije in apetitivnih ZDA se je močno razlikovalo. Tako je bil boleč električni šok dostavljen ob izravnavi avCS + in je trajal natanko 200 ms, čokolada in slana pereca pa sta bila udeležencem predstavljena okrog 2 s po začetku appCS +, trajanje pa je bilo nedoločljivo, saj je bilo odvisno od tega, kako hitro posameznik jih je pojedel. Izbira, da bodo na ta način dostavili apetitne ZDA, je temeljila na predhodni študiji na ljudeh (Prévost et al., 2012). Vendar bi bilo metodološko bolj elegantno dostaviti privlačne ZDA, primerljive z averzivnimi ZDA, kot sta požirek soka ali ingverjev ale pri odmeri appCS +. Da bi nadomestili to veliko zaznavno razliko, smo predstavili geometrijske oblike v povezavi z vizualnim dražljajem, ki simbolizira ZDA. Tretjič, ne moremo dokončno izključiti možnosti, da so hitro ugasnjeni fiziološki odzivi v fazi izumrtja posledica metodološkega vidika. V resnici vizualni dražljaji v tej fazi niso bili predstavljeni v povezavi z ameriškim simbolom, kot je to veljalo v fazah pridobivanja. Možno je, da je samo predstavitev CS-jev vplivalo na CR in zato med prvimi poskusi izumiranja ni bilo opaziti bistvenih razlik. Vendar so naši hitro ugasnjeni odzivi v skladu z ugasnjenimi odzivi v prejšnji študiji, v kateri je bil CS + (obraz) predstavljen v povezavi z ZDA (krik) med pridobitvijo, ne pa med izumrtjem (Lissek in sod., 2008).
Za zaključek smo našli uspešne averzivne in apetitno pogojene odzive na dražljaj, povezan z grožnjo, in na dražljaj, povezan z nagrado. Zanimivo je, da sta eksplicitna (ocene) in implicitna (refleksni zagon) delovala sinergično, ker je bilo avCS + prijavljeno kot negativno in inducirano potenciranje zagona, appCS + pa kot pozitivno in inducirano slabljenje zagona. Poleg tega sta eksplicitna (ocene) in fiziološka (SCR) vzburjenja appCS + disociirana, kar odraža dva različna procesa.
Izjava o konfliktu interesov
Avtorji izjavljajo, da je bila raziskava izvedena v odsotnosti komercialnih ali finančnih odnosov, ki bi se lahko razumeli kot potencialno navzkrižje interesov.
Priznanja
To delo je podprlo Skupno raziskovalno središče "Strah, tesnoba in anksiozne motnje", SFB-TRR 58, projekt B1.
Dodatni material
Dodatno gradivo za ta članek lahko najdete na spletni strani: http://journal.frontiersin.org/article/10.3389/fnbeh.2015.00128/abstract
Reference
Andreatta, M., Mühlberger, A., Glotzbach-Schoon, E. in Pauli, P. (2013). Predvidljivost bolečine razveljavi oceno valencnosti olajšavnega dražljaja. Spredaj. Syst. Nevrosci. 7: 53. doi: 10.3389 / fnsys.2013.00053
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Andreatta, M., Mühlberger, A., Yarali, A., Gerber, B., in Pauli, P. (2010). Razkorak med implicitno in eksplicitno pogojeno valenco po učenju lajšanja bolečine pri ljudeh. Proc. Biol. Sci. 277, 2411 – 2416. doi: 10.1098 / rspb.2010.0103
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Austin, AJ in Duka, T. (2010). Mehanizmi pozornosti za apetitne in averzivne izide pri Pavlovianovi kondicioniranju. Behav. Brain Res. 213, 19 – 26. doi: 10.1016 / j.bbr.2010.04.019
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Blumenthal, TD, Cuthbert, BN, Filion, DL, Hackley, S., Lipp, OV in van Boxtel, A. (2005). Poročilo odbora: smernice za elektromiografske študije zrcala na človeku. Psihofiziologija 42, 1 – 15. doi: 10.1111 / j.1469-8986.2005.00271.x
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Bouton, ME in Peck, CA (1989). Kontekstni učinki na kondicioniranje, izumrtje in ponovno vzpostavitev apetitivnega pripravka. Žival. Naučite se. Behav. 17, 188 – 198. doi: 10.3758 / BF03207634
CrossRef Celotno besedilo | Google Scholar
Bradley, MM (2009). Naravna selektivna pozornost: orientacija in čustva. Psihofiziologija 46, 1 – 11. doi: 10.1111 / j.1469-8986.2008.00702.x
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Clark, JJ, Hollon, NG in Phillips, PEM (2012). Pavlovijski sistemi vrednotenja pri učenju in odločanju. Curr. Opin. Neurobiol. 22, 1054 – 1061. doi: 10.1016 / j.conb.2012.06.004
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Delgado, MR, Jou, RL in Phelps, EA (2011). Nevronski sistemi, ki temeljijo na averzivnem kondicioniranju pri ljudeh s primarnimi in sekundarnimi ojačevalci. Spredaj. Neurosci. 5: 71. doi: 10.3389 / fnins.2011.00071
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Dickinson, A. in Balleine, B. (1994). Motivacijski nadzor ciljno usmerjenega delovanja. Žival. Naučite se. Behav. 22, 1 – 18. doi: 10.3758 / BF03199951
CrossRef Celotno besedilo | Google Scholar
Fendt, M. in Fanselow, MS (1999). Nevroanatomska in nevrokemična osnova pogojenega strahu. Neurosci. Biobehav. Rev. 23, 743 – 760. doi: 10.1016 / j.ijpsycho.2012.09.006
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Gottfried, JA, O'Doherty, J., in Dolan, RJ (2002). Apetitvno in averzivno vohalno učenje pri ljudeh, ki so ga preučevali z uporabo funkcionalne magnetne resonance, povezane s dogodki. J. Neurosci. 22, 10829 – 10837. Na voljo na spletu: http://www.jneurosci.org/content/22/24/10829.full
Grillon, C., Baas, JM, Cornwell, B. in Johnson, L. (2006). Kondicioniranje kontekstov in izogibanje vedenju v okolju virtualne resničnosti: učinek predvidljivosti. Biol. Psihiatrija 60, 752 – 759. doi: 10.1016 / j.biopsych.2006.03.072
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Hamm, AO in Weike, AI (2005). Nevropsihologija učenja strahu in regulacija strahu. Int. J. Psihofiziol. 57, 5 – 14. doi: 10.1016 / j.ijpsycho.2005.01.006
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Klucken, T., Schweckendiek, J., Merz, CJ, Tabbert, K., Walter, B., Kagerer, S., et al. (2009). Nevronske aktivacije pridobivanja pogojene spolne vzburjenosti: učinki zavedanja ob nepredvidljivih dogodkih in spola. J. Spol. Med. 6, 3071 – 3085. doi: 10.1111 / j.1743-6109.2009.01405.x
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Klucken, T., Wehrum, S., Schweckendiek, J., Merz, CJ, Hennig, J., Vaitl, D., et al. (2013). Polimorfizem 5-HTTLPR je povezan s spremenjenimi hemodinamskimi odzivi med apetitivnim kondicioniranjem. Hum. Brain Mapp. 34, 2549 – 2560. doi: 10.1002 / hbm.22085
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Koch, M. (1999). Nevrobiologija zagorelosti. Prog. Neurobiol. 59, 107–128. doi: 10.1016/S0301-0082(98)00098-7
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Koch, M., Schmid, A. in Schnitzler, H.-U. (1996). Užitek slabljenja zagona moti lezija jedrnega jedra. Nevroport 7, 1442–1446. doi: 10.1097/00001756-199605310-00024
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Krohne, HW, Egloff, B., Kohmann, C.-W., in Tausch, A. (1996). Različna različica "Pozitivnega in negativnega vpliva" (PANAS). Diagnostika 42, 139-156.
Kumar, P., natakar, G., Ahearn, T., Milders, M., Reid, I., in Steele, JD (2008). Nenormalne razlike v časovni razliki navajajo na velike depresije. Brain 131, 2084 – 2093. doi: 10.1093 / možgani / awn136
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Laux, L., Glanzmann, P., Schaffner, P., in Spielberger, CD (1981). Das State-Trait Angstinventar. Weinheim: Beltz test.
Levy, DJ in Glimcher, PW (2011). Primerjava jabolk in pomaranč: z uporabo nagrajevanja in splošne subjektivne zastopanosti vrednosti v možganih. J. Neurosci. 31, 14693 – 14707. doi: 10.1523 / JNEUROSCI.2218-11.2011
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Lissek, S., Levenson, J., Biggs, AL, Johnson, LL, Ameli, R., Pine, DS, et al. (2008). Zvišan strah pogojuje socialno pomembne brezpogojne dražljaje pri socialni anksiozni motnji. Am. J. Psihiatrija 165, 124 – 132. doi: 10.1176 / appi.ajp.2007.06091513
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Martin-Soelch, C., Linthicum, J., in Ernst, M. (2007). Apetitno kondicioniranje: nevronske podlage in posledice za psihopatologijo. Neurosci. Biobehav. Rev. 31, 426 – 440. doi: 10.1016 / j.neubiorev.2006.11.002
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
McDannald, MA, Lucantonio, F., Burke, KA, Niv, Y. in Schoenbaum, G. (2011). Ventralni striatum in orbitofrontalna skorja sta potrebna za učenje okrepitve, ki temelji na modelu, ne pa tudi brez modela. J. Neurosci. 31, 2700 – 2705. doi: 10.1523 / JNEUROSCI.5499-10.2011
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Milad, MR in Quirk, G. (2012). Izumiranje strahu kot model translacijske nevroznanosti: deset let napredka. Annu. Rev. Psychol. 63, 129 – 111. doi: 10.1146 / annurev.psych.121208.131631
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Pavlov, IP (1927). Kondicioni refleksi: preiskava fiziološke aktivnosti možganske skorje. London: Oxford University Press.
Prévost, C., Liljeholm, M., Tyszka, JM in O'Doherty, JP (2012). Nevronski korelati specifičnega in splošnega prenosa pavlovija na instrumental v človeških amigdalarnih podregijah: študija fMRI z visoko ločljivostjo. J. Neurosci. 32, 8383 – 8390. doi: 10.1523 / jneurosci.6237-11.2012
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Tranel, D. in Damasio, H. (1994). Nevroanatomski korelati elektrodermalnih kožnih prevodnih odzivov. Psihofiziologija 31, 427–438. doi: 10.1111/j.1469-8986.1994.tb01046.x
PubMed Povzetek | CrossRef Celotno besedilo | Google Scholar
Ključne besede: klasično kondicioniranje, nagrada, kazen, začudni refleks, odzivnost prevodnosti kože
Citiranje: Andreatta M in Pauli P (2015) Apetitna proti averzivni kondiciji pri ljudeh. Spredaj. Behav. Neurosci. 9: 128. doi: 10.3389 / fnbeh.2015.00128
Prejeto: 11 februar 2015; Sprejeto: 05 maj 2015;
Objavljeno: 19 maj 2015.
Uredil:
Niels Birbaumer, Univerza v Tuebingenu, Nemčija
Pregledal:
Tim Klucken, Univerza Justus Liebig Giessen, Nemčija
Alessandro Angrilli, Univerza v Padovi, Italija
Copyright © 2015 Andreatta in Pauli. To je članek z odprtim dostopom, ki se distribuira pod pogoji Licenca za priznanje Creative Commons (CC BY). Uporaba, distribucija ali reprodukcija v drugih forumih je dovoljena pod pogojem, da so avtor (ji) ali dajalec licence priznani in da je navedena izvirna objava v tej reviji v skladu s sprejeto akademsko prakso. Uporaba, distribucija ali reprodukcija ni dovoljena, kar ni v skladu s temi pogoji.
* Korespondenca: Marta Andreatta, oddelek za psihologijo (biološka psihologija, klinična psihologija in psihoterapija), Univerza v Würzburgu, Marcusstraße 9-11, D-97070 Würzburg, Nemčija, [e-pošta zaščitena]