 Martin Zack
Martin Zack- 1Laboratorij za kognitivno psihofarmakologijo, Oddelek za nevroznanost, Center za odvisnost in duševno zdravje, Toronto, ON, Kanada
- 2Program Translational Neuroscience, Oddelek za psihiatrijo, Medicinska šola, University of Pennsylvania, Philadelphia, PA, ZDA
- 3Oddelek za biopsihologijo, Oddelek za nevroznanost, Center za odvisnost in duševno zdravje, Toronto, ON, Kanada
Zasvojenost je možganska bolezen, ki jo povzroča kronična izpostavljenost drogam. Senzibilizacija možganskih dopaminskih (DA) sistemov delno posreduje ta učinek. Patološko igranje (PG) velja za vedenjsko odvisnost. Zato lahko PG povzroči kronična izpostavljenost igranju na srečo. Prepoznavanje senzibilizacije sistemov javnega naročanja, ki je posledica iger na srečo, bi podprlo to možnost. Nagrade za igre na srečo izzovejo izdajo DA. Ena epizoda igralnega avtomata premakne odziv DA od nagrajevanja do nastopa znakov (vrteči se koluti) za nagrado, v skladu s časovnimi načeli učenja razlike. Tako pogojeni dražljaji (CS) igrajo ključno vlogo pri odzivih DA na igre na srečo. Pri primatih je odgovor DA na CS najmočnejši, če je verjetnost nagrajevanja 50%. Po tem razporedu CS pričakuje pričakovano nagrado, vendar ne daje nobenih informacij o tem, ali se bo to zgodilo na določenem preskušanju. Med igranjem na srečo mora načrt 50% sprožiti maksimalno izdajo DA. To se zelo ujema s frekvenco nagrajevanja (46%) na komercialnem igralnem avtomatu. Sproščanje DA lahko prispeva k preobčutljivosti, zlasti za amfetamine. Kronična izpostavljenost CS-ju, ki napoveduje nagrado 50% časa, lahko posnema ta učinek. To hipotezo smo testirali v treh študijah s podganami. Živali so prejele 15 × 45-min izpostavljenosti v CS, ki je predvidel nagrado z verjetnostjo 0, 25, 50, 75 ali 100%. CS je bil lahek; nagrada je bila raztopina saharoze 10%. Po treningu so podgane prejele senzibilizirni režim petih ločenih odmerkov (1 mg / kg) d-amfetamina. Nazadnje so prejeli izziv 0.5 ali 1 mg / kg amfetamina pred testom lokomotorne aktivnosti 90-min. V vseh treh študijah je skupina 50% pokazala večjo aktivnost kot druge skupine kot odziv na obe izzivni odmerki. Velikosti efektov so bile skromne, vendar dosledne, kar se odraža v pomembni skupini x povezava (ϕ = 0.986, p = 0.025). Kronična izpostavljenost nagrajenega napovednega draženja, podobnega igralništvu, lahko spodbudi preobčutljivost na amfetamin, podobno izpostavljenosti samemu amfetaminu.
Predstavitev
Odvisnost je bila označena kot možganska bolezen, ki jo povzroča kronična izpostavljenost zlorabam drog (Leshner, 1997). Nevroplastičnost naj bi posredovala učinke takšne izpostavljenosti (Nestler, 2001). Preobčutljivost možganskih dopaminskih (DA) sistemov je oblika nevroplastičnosti, povezana s hiperreaktivnostjo na pogojene dražljaje (CS) za zdravila in kompulzivno iskanje drog (Robinson in Berridge, 2001). Preobčutljivost je operativno definirana z večjim sproščanjem DA kot odzivom na CS za nagrado in s povečanim lokomotornim odzivom na farmakološki izziv DA (Robinson in Berridge, 1993; Pierce in Kalivas, 1997; Vanderschuren in Kalivas, 2000). Čeprav je senzibilizacija le ena od mnogih možganskih sprememb, povezanih z odvisnostjo (prim. Robbins in Everitt, 1999; Koob in Le Moal, 2008) so predlagali spremembe v sproščanju presinaptičnega dopamina, ki predstavljajo običajne nevroadaptacije, ki so vključene v iskanje drog na osnovi odvisnosti (npr. recidiv), pri tistih zdravilih, ki povzročajo lokomotorno senzibilizacijo na opiate (npr. morfij) ali izziv stimulansa (npr. amfetamin) , povzroči tudi ponovno vzpostavitev ugaslih operantnih odzivov za heroin ali samo-dajanje kokaina - živalski model relapsa (Vanderschuren et al., 1999). Dokazi, da je spodbujevalna senzibilizacija (povečana vrednost nagrad za droge) najbolj izrazita po začetni izpostavljenosti odvisnim drogam, še kaže, da je lahko senzibilizacija vključena tudi v zgodnjih fazah zasvojenosti (Vanderschuren in Pierce, 2010).
Patološko igranje na srečo (PG) je bilo opisano kot vedenjska zasvojenost in v zadnjem času prerazvrščeno v isto kategorijo kot motnje odvisnosti od snovi v izdaji 5th, Diagnostičnega in statističnega priročnika duševnih motenj (Frascella et al., 2010; APA, 2013). To pomeni, da lahko PG povzroča kronična izpostavljenost podobnim dejavnostim, ki so podobne igranju, da lahko skupni mehanizmi posredujejo učinke iger na srečo in izpostavljenosti drogam (Zack in Poulos, 2009; Leeman in Potenza, 2012); in da je lahko senzibilizacija možganskih poti DA pomemben element tega procesa.
Klinični dokazi posredno podpirajo to možnost: Z uporabo pozitronske emisijske tomografije (PET) Boileau in sodelavci so ugotovili, da moški PG kažejo bistveno večje sproščanje striatnih DA kot odziv na amfetamin (0.4 mg / kg) kot zdravi moški (Boileau et al., 2013). Skupne razlike v skupini so bile pomembne v asociativnem in somatosenzoričnem striatumu. V limbičnem striatumu, ki vključuje jedro nucleus accumbens, se skupine niso razlikovale. Toda pri PG subjektih je sproščanje DA v limbičnem striatumu neposredno povezano z resnostjo simptomov PG. Te ugotovitve so skladne s preobčutljivostjo možganskih DA poti v PG, vendar kažejo tudi na nekatere pomembne razlike s posamezniki, odvisnimi od človeške snovi, in s klasičnim živalskim modelom senzibilizacije amfetamina. Za razliko od subjektov PG in živali, izpostavljenih nizkim odmerkom amfetamina (prim. Robinson et al., 1982), ljudje z odvisnostjo od snovi dosledno kažejo zmanjšano sproščanje DA na izziv stimulansa (Volkow et al., 1997; Martinez et al., 2007), in dokazi iz živali kažejo, da to lahko odraža pomanjkanje funkcije DA v začetnih fazah abstinence po vzorcih zlorabe drog (Mateo et al., 2005). V študijah, pri katerih je pri živalih dokazana senzibilizacija stimulansov, je povečano sproščanje DA običajno opazno v limbičnem striatumu in ne v dorzalnem (asociativnem, somatosenzoričnem) striatumu (Vezina, 2004). Vendar pa je bilo iskanje živali pri ponavljajočih se kokainu povezano z okrepljenim sproščanjem DA v hrbtnem striatumu, kar kaže na bolj običajno obliko motiviranega obnašanja (Ito et al., 2002). Tako je lahko celotno povišanje sproščanja DA v dorzalnih regijah v subjektih PG povezano z navadnimi (nefleksibilnimi, rutiniziranimi) nagradami, ki zahtevajo "napredovanje od ventralne do dorzalne domene striatuma" (Everitt in Robbins, 2005, str. 1481), medtem ko lahko resnost-odvisno sproščanje DA v limbičnem striatumu pri teh osebah bolj ustreza spodbujevalni senzibilizaciji, ki je tipično modelirana pri živalih. Ugotovitve PET ne morejo razkriti, ali je bila hiperreaktivnost DA že obstoječa značilnost teh subjektov PG, posledica izpostavljenosti iger na srečo, ali rezultat nekega drugega procesa v celoti. Za obravnavo tega vprašanja je treba pokazati indukcijo preobčutljivosti pri kronični izpostavljenosti na srečo pri osebah, ki so pred izpostavljenostjo normalne. To sproža vprašanja o tem, katere značilnosti iger na srečo najverjetneje povzročajo preobčutljivost.
Skinner je ugotovil, da je spremenljiv urnik okrepitve bistvenega pomena za privlačnost iger na srečo (ali vsaj njeno vztrajnost) (Skinner, 1953). Stilno vedenje v igri z igralnimi avtomati ustreza osnovnim načelom instrumentalne kondicije, kar se odraža v perspektivni korelaciji med denarnim izplačilom in velikostjo stav na zaporednih vrtljajih (Tremblay et al., 2011). Tako se zdi, da se odziv na operativno razmerje z variabilnim razmerjem zagotavlja eksterno veljaven model iger na srečo na igralnih avtomatih.
Nedavne raziskave z živalmi zagotavljajo močno začetno podporo za vzročni učinek izpostavljenosti iger na srečo občutljivosti. Singer in sodelavci so preučevali učinke dnevnih sej 55 1 na fiksno (FR20) ali spremenljivo (VR20) saharinovo ojačitev v operantski vzvodni skupini na poznejši lokomotorni odziv na nizke odmerke (0.5 mg / kg) amfetamina pri zdravih moških ( Sprague Dawley) podgane (Singer et al., 2012). Predvidevali so, da bi morale podgane, ki so izpostavljene variabilnemu urniku, ki posnemajo igre na srečo, pokazati večji odziv na amfetamin kot podgane, ki so bile izpostavljene fiksnemu urniku. Kot je bilo napovedano, je skupina VR20 pokazala 50% večji lokomotorni odziv na amfetamin kot skupina FR20. Nasprotno, skupine so pokazale enako lokomocijo po injiciranju slanice. Te ugotovitve potrjujejo, da je kronična izpostavljenost variabilni ojačitvi zadostna, da povzroči hiperreaktivnost na izziv DA pri zdravih živalih, ki so randomizirane na ustrezne sheme.
Iz tega izhaja več vprašanj: Prvič, v kolikšni meri zaznano nepredvidljivost - ali njeno pomanjkanje - med operantnim odzivom in njegovim izidom posreduje te učinke? Ali ta učinek v učnih izrazih vključuje »pričakovanje odziva na izid«, ali pa se podoben učinek vidi v odsotnosti operantnega odziva, tj. »Pričakovano spodbudo-rezultat« v Pavlovski paradigmi (prim. Bolles, 1972)? Drugič, ali stopnja kontingence med predhodnim dogodkom (odzivom ali dražljajem) in njegovim izidom vpliva na stopnjo senzibilizacije?
Drugo vprašanje zadeva vlogo negotovosti pri preobčutljivosti. Na primer, ali imajo igre, katerih rezultat je resnično naključen - popolnoma nepredvidljiv - večji potencial za spodbujanje senzibilizacije kot igre, kjer so možnosti za zmago jasno določene, vendar ne naključne, tudi če je absolutna stopnja nagrajevanja nizka? Ta raziskava je obravnavala ta vprašanja.
Eksperimentalna zasnova je bila utemeljena s semensko študijo o pričakovanem nagrajevanju in odzivu nevrona DA pri opicah (Fiorillo et al., 2003). Živali v tej študiji so prejele nagrado za sok (ZDA) na podlagi spremenljivih razporedov 0, 25, 50, 75 ali 100%. Razporede je določil 1 iz različnih CS (4) ikon. Razpored 0% je prinesel nagrado tako pogosto kot urnik 100%, vendar je izpustil CS. Ključni odvisni ukrep je bila hitrost žganja nevronov DA v intervalu med nastopom CS in ameriško dostavo ali opustitvijo. Študija je ugotovila, da se je odziv DA povečal v odvisnosti od negotovosti pri nagrajevanju. Tako je pod urnikom 100% CS evociral majhno aktivnost, pod urniki 25 in 75%, je CS sprožil zmerno in podobno raven aktivnosti, pod urnikom 50% pa je CS sprožil maksimalno aktivnost. V vsakem primeru se je stopnja streljanja povečala v času CS-US intervala, tj.
Te ugotovitve kažejo, da se aktivnost DA ne spreminja le z ali je nagrada določena (Fiksni Ratio) ali negotova (Variable Ratio), ampak se tudi spreminja v obratnem sorazmerju s količino informacij o dostavi nagrad, ki jih posreduje CS. V pogoju 100% CS prikliče pričakovanje nagrajevanja in tudi popolnoma napove njegovo dostavo. V pogojih 25 in 75% CS prikliče pričakovano dobo in napove dobavo nagrad trikrat od štirikrat. V stanju 50% CS prikliče pričakovano pričakovanje, vendar ne zagotavlja nobenih informacij o dostavi nagrade, ki bi presegla samo naključje. Na podlagi svojih ugotovitev so Fiorillo et al. Sklenil je: »To povečanje dopamina, povzročeno z negotovostjo, lahko prispeva k nagrajevanju lastnosti iger na srečo« (str. 1901).
Učinki 50-odstotne variabilne nagrade v eni seji se med več sejami ne bi smeli spreminjati, ker je verjetnost nagrade v celoti nepredvidljiva pri vsakem preizkusu. Tako bi moral pri razmisleku o pogojih, ki bi maksimizirali kronično aktivacijo DA nevronov v ponavljajočih se epizodah iger na srečo, 50-odstotni urnik povzročiti najbolj trajen in najmočnejši učinek. To je treba omeniti glede na to, da je bila dolgoročna stopnja nagrajevanja (izplačilo> 0), opažena pri tisočih vrtljajih na komercialnih igralnih avtomatih, 45.8% (Tremblay et al., 2011). Zdi se, da 50% variabilna nagrada natančno odraža časovni razpored izplačil, ki ga upravljajo dejanske naprave za igre na srečo.
Ta študija je uporabila enake urnike kondicioniranja kot Fiorillo et al. pri kronični izpostavljenosti, zasnova skupine med podganami. Živali so prejemale ~ 3 tedne vsakodnevnega kondicioniranja, kjer je bil CS (svetloba) seznanjen z ZDA (majhna količina saharoze). Po fazi usposabljanja so se živali počivale pred oceno preobčutljivosti, indeksirane z gibalnim odzivom na amfetamin. Na podlagi literature je bilo napovedano, da se podgane, izpostavljene različnim urnikom nagrajevanja, ne bodo razlikovale v vedenju lokomotornega vedenja brez drog, ampak bodo pokazale bistveno različne ravni gibanja po amfetaminu, pri čemer je 50-odstotna skupina imela večji lokomotorni odziv na zdravilo glede na druge skupine med odmerki, vzorec, ki bi ga pričakovali, če bi bilo 50% živali predhodno izpostavljeno dodatnim odmerkom samega amfetamina (tj. navzkrižna preobčutljivost).
poskus 1
Materiali in metode
Predmeti
Štiri skupine (n = 8 / skupina) odraslih (300-350 g) samcev podgan Sprague-Dawley (Charles River, St. Constant, Quebec, Kanada) so bile nameščene posamično v jasnih polikarbonatnih škatlah (20 × 43 × 22 cm) pod vzvratno 12: 12 cikel svetlobe in teme. Prejeli so ad libitum dostop do hrane in vode ter vsakodnevno ravnanje s preskuševalcem za 2 tednov pred študijo. Vsaka skupina je bila pogojena z eno od štirih spremenljivih razporedov nagrajevanja: 0, 25, 50 ali 100%. Skupina 75% je bila izpuščena v tej začetni študiji, kot Fiorillo et al. (2003) ugotovili enakovredno objavo DA po CS-ju pod 25 in 75% razporedi nagrajevanja, tako da sta oba pogoja privedla do večje izdaje DA kot je bil 100% CS-US pogoj, vendar manj kot pogoj 50%.
Aparati
Dostop do predstavitev saharoze in CS je bil zagotovljen posamično v škatlah za operativno kondicioniranje (33 × 31 × 29 cm). Vsaka škatla je bila opremljena z ojačevalno revije, ki se nahaja na sprednji steni. Svetloba na vrhu revije je služila kot CS. Motorizirana, solenoidno nadzorovana tekočina za vodo se lahko dvigne na tla revije. Dogodke v škatli je nadzorovala oprema in programska oprema podjetja Med Associates z uporabo lastnega programa, napisanega v MED-PC. Lokomotorno testiranje je bilo izvedeno posamično v kletkah iz pleksi stekla (27 × 48 × 20 cm). Vsaka kletka je bila opremljena s sistemom za spremljanje, ki je sestavljen iz šestih celic foto-žarkov za zaznavanje vodoravnega gibanja.
Postopek
usposabljanje. Študija je bila izvedena v skladu z etičnimi smernicami, ki jih je določil Kanadski svet za nego živali. Podgane so bile v času študije omejene na 90% njihove telesne teže in so bile nameščene posamično. Vsaka podgana je prejela 15 dni saharoznega nagrajevanja (10% vodna raztopina pri 0.06 ml na nagrado): 5 zaporednih dni × 3 tednov, z izključenimi vikendi. Živali so vzdrževali na standardni mizi pred in po fazi usposabljanja; izpostavljenost saharozi je bila omejena na petnajst ur usposabljanja 40-min. Vsako dnevno sejo so sestavljale predstavitve stimulansov 15 (svetloba; CS), vsaka pa je bila ločena z medsebojnim intervalom 120 s. Svetloba se je nahajala v zgornji ploskvi revije in ostala vključena za 25 s, pri čemer je bila v zadnjih 5 s na voljo saharoza. V primeru skupine 0 je bila kapljica saharoze dvignjena na vsak 140 s (za 5 s), svetloba dražljaja pa ni bila osvetljena. To je izenačilo interval med predstavitvami potopa v skupini 0 in drugih skupinah (120 + 25 s). Vsako zdravljenje je trajalo ~ 40 min. V povprečju je skupina 25 prejela saharozo enkrat na vsake štiri predstavitve CS; skupina 50 je dobila saharozo enkrat na vsaki dve predstavitvi CS in skupina 100 je po vsaki predstavitvi CS dobivala saharozo.
Testiranje. Dva tedna po zadnjem dostopu (ali "kondicioniranju") saharoze so ocenili lokomotorni odziv na d-amfetamin (AMPH; ip). Podganam so dajali tri seje 2-h, da bi se privadili na lokomotorna polja, čemur je sledilo šest testov AMPH. Dnevi testiranja AMPH so se zgodili v intervalih 1-teden. V testnih dneh so podgane dobile 30 min, da se privadijo v škatle, nato pa so prejele en odmerek zdravila 0.5 mg / kg, ki je sledil, na ločenih tedenskih sejah, s petimi odmerki 1.0 mg / kg (en odmerek na dan) na testnih dneh 1 skozi 5 . Za vsako sejo smo ocenili X-NUMX min.
Podatkovni analitični pristop
Statistične analize so bile izvedene s SPSS (v. 16 in v. 21; SPSS Inc., Chicago IL). Takojšnji vedenjski odziv na CS je bil ocenjen v smislu, da se nos vtakne v odprtino, kjer je bila razpršena saharoza. Povprečno število pokesov v tem intervalu (5 s na poskus) je bilo nato primerjano s povprečnim številom nosnih pokesov za enako trajanje (5 s), povprečeno za čas, ko je bil CS odsoten. Skupina × seje ANOVAs nos-pokes s CS prisoten in odsoten sledenje pridobivanje diskriminatorne odziva na cue in neselektivno nos poke odzive pod različnimi urniki v času 15 saharoze usposabljanja.
Učinki zdravljenja na lokomotorne odzive so bili ocenjeni z ANOVA za skupino × sejo za fazo navajanja brez zdravil (tri seje), izzivi pred senzitizacijo 0.5 mg / kg AMPH (eno sejo) in med petimi sejami 1 mg / kg AMPH preobčutljivost, ko se je pričakovalo, da se bodo skupine razlikovale glede na ponavljajoče se odmerke AMPH. Skupina × seja ANOVAs je ocenila tudi lokomotorne odzive brez zdravil med fazo privzema pred injiciranjem 30-min iz vsake AMPH testne seje. Načrtovane primerjave so ocenile razliko v povprečni uspešnosti za skupino 50 v primerjavi s skupino 0 (brez nadzora nad pričakovanji) in skupino 100 (brez nadzora negotovosti) s pomočjo t-preskusi (Howell, 1992), z uporabo napak MS in napak df za ustrezen učinek (tj. interakcija skupine ali skupine × sejo) iz ANOVA (Winer, 1971). Polinomske analize trendov so testirale profil sprememb v času sej.
Da bi ugotovili, ali so pristopni odzivi v prisotnosti ali odsotnosti CS med treningi saharoze 15 prispevali k variaciji lokomotornega odziva na AMPH ali posredovanih razlikam v skupini AMPH, so bile na AMPH opravljene nadaljnje analize kovariance (ANCOVA). lokomotorni podatki, vključno s celotnim pokesom nosu (vsota za seje 15), ko CS ni bil prisoten kot kovarija. Pomemben učinek kovarija bi pokazal, da so odzivi pristopa brez drog omilili (vplivali na moč) učinkov skupine ali seje. Zmanjšanje pomena učinkov skupine ali seje v prisotnosti pomembne kovarija bi nakazalo, da so pristopni odzivi posredovani (upoštevani) učinki skupine ali seje. Zmanjšanje pomena učinkov skupine ali seje v odsotnosti pomembnega kovarijabilnega učinka bi preprosto odražalo izgubo statistične moči zaradi prerazporeditve df iz obdobja napake v kovarija in ne bi imelo vpliva na razlago učinki skupine ali seje.
Rezultati
Nos pokes med saharozno kondicioniranje
CS. Slika 1A kaže srednji nos za skupine 25, 50 in 100, medtem ko je bila CS prisotna na sejah s saharozo (nosne pokes niso bile kodirane za skupino 15, ki ni prejela CS). ANOVA skupine 0 × 3 je pokazala pomembne glavne učinke skupine, F(2, 21) = 5.63, p = 0.011 in seja, F(14, 294) = 14.00, p <0.001, skupaj s pomembno interakcijo Group × Session, F(28, 294) = 2.93, p <0.001. Slika 1A kaže, da je glavni učinek seje odražal povečanje nosa v vseh treh skupinah, glavni učinek skupine pa je pokazal na splošno višje skupne rezultate v skupini 100 v primerjavi s skupino 25 z vmesnimi rezultati v skupini 50. Pomembna interakcija skupine × seja za kubični trend, F(2, 21) = 4.42, p = 0.030, je pokazal hitro naraščanje, pomikanje in izravnavanje v nosnih pokesih nad sejami v skupini 100, v primerjavi z linearnim povečanjem nad sejami v skupini 50 in s plitkim linearnim povečanjem nad sejami v skupini 25.
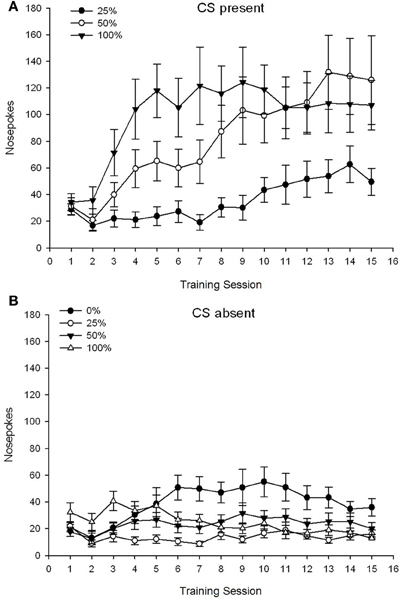
Slika 1. Srednji (SE) pristopni odzivi (nosni pokes) na treningih saharoze 15 v skupinah podgan Sprague Dawley (n = 8 / skupina) izpostavljena nagradi saharoze (rešitev 10%) pod 0, 25, 50 ali 100% spremenljivimi razporedi. Pogojni dražljaj je bila svetloba (120 s). Skupina 0 je prejela enako število nagrad kot skupina 100 v odsotnosti pogojenih dražljajev. () Rezultati, ko je bila CS prisotna (preskušanja 5 s × 15). (B) Rezultati, ko je bila CS odsotna (povprečje za 5 × 15 s, medtem ko je svetloba izklopljena).
CS odsoten. Slika 1B kaže povprečno pokanje nosu za vse štiri skupine za enakovredno trajanje (preskušanja 5 s × 15), povprečeno za čas, ko je bila CS odsotna. ANOVA skupine 4 × 15 je pokazala pomembne glavne učinke skupine, F(3, 28) = 7.06, p = 0.001 in seja F(14, 392) = 2.84, p <0.001, skupaj s pomembno interakcijo Group × Session, F(42, 392) = 3.93, p <0.001. Pomembna interakcija med skupino in sejo za kvadratni trend, F(3, 28) = 3.91, p = 0.019, brez interakcije za kubični trend, F(3, 28) <0.93, p > 0.44, je odražal "obrnjen-U" profil zbadanja v nosu med sejami v skupini 0, v primerjavi s splošno stabilnim profilom med sejami v drugih skupinah.
Prilagajanje lokomotornim komoram
ANOVA skupine 4 × 3 je dala glavni učinek seje, F(2, 56) = 5.67, p = 0.006 in nobenih drugih pomembnih učinkov, F(3, 28) <1.60, p > 0.21. Povprečni (SE) prelomi žarka na 2 uri v lokomotornih omaricah so bili 1681, 123 (1), 1525. in 140 (2), 1269. seji pa 96 (3). Načrtovane primerjave niso pokazale pomembnih razlik med skupino 50 in skupino 0 ali skupina 100 na prvi ali zadnji seji navajanja, t(84) <1.69, p > 0.05. Tako je bila v odsotnosti AMPH ponavljajoča izpostavljenost testnim poljem povezana s stalnim upadanjem spontane gibalne aktivnosti v štirih skupinah (tj. Učinek seje) in brez diferencialnega odziva kot funkcije urnika treninga saharoze (brez interakcije) .
Testne seje
Učinki pred senzitizacije izziva 0.5 mg / kg AMPH.
Premikanje pred injiciranjem. Enosmerna ANOVA skupine 4 za lokomotorni odziv med fazo privzema pred injiciranjem 30-min ni prinesla pomembnih učinkov, F(3, 28) <1.05, p > 0.38. Načrtovane primerjave niso odkrile pomembne razlike med skupino 50 in skupino 0 ali skupino 100, t(32) <0.87, p > 0.40. Zato izhodiščne razlike v gibu pred injiciranjem niso upoštevale skupinskih razlik v gibalnem odzivu na AMPH. Povprečni (SE) prelomi žarka za vzorec so bili 559 (77).
Post-injekcijska gibanja v primerjavi s končno terapijo brez navzočnosti zdravil. Na seji ANOVA s 4 skupinami × 2 so primerjali gibalne odzive skupin na zadnji seji navajanja in takoj po izzivu AMPH pred senzibilizacijo 0.5 mg / kg. Rezultati za navadno sejo (120 min) so bili prilagojeni tako, da ustrezajo trajanju preizkusne seje AMPH (90 min) (ocena surovega navajanja × 90/120). Analiza je dala pomemben glavni učinek seje, F(1, 28) = 34.16, p <0.001 in brez drugih pomembnih učinkov, F(3, 28) <2.26, p > 0.10. Učinek seje je odražal povečanje povprečnih (SE) prelomov žarka kot odziv na odmerek s 952 (72) na 1859 (151). Načrtovane primerjave niso pokazale pomembnih razlik med skupino 50 in skupino 0 ali skupino 100 kot odziv na odmerek, t(56) <1.72, p > 0.10. Vendar je vrstni red ocen loma žarka (M; SE) usklajen s hipotezo: skupina 50 (2205; 264)> skupina 0 (2025; 203)> skupina 100 (1909; 407)> skupina 25 (1296; 299) .
Učinki zdravila 1 mg / kg AMPH.
Premikanje pred injiciranjem. ANOVA skupine 4 × 5 seje ANOV lokomotornega odziva med fazo predhodnega vbrizgavanja 30-min v fazi 1 mg / kg AMPH je pokazala glavni učinek seje, F(4, 112) = 43.64, p <0.0001 in brez drugih pomembnih učinkov, F(3, 28) <0.97, p > 0.42. Načrtovane primerjave niso pokazale pomembne razlike med skupino 50 in skupino 0 ali skupino 100 na prvem ali zadnjem preizkusu, t(140) <0.84, p > 0.30. Zato izhodiščne razlike v gibanju niso upoštevale skupinskih razlik v gibalnem odzivu na AMPH. Povprečne ocene preloma (SE) za fazo navajanja pred odmerkom na sejah 1–5 so bile: 454 (30), 809 (53), 760 (36), 505 (35), 756 (39).
Pospeševanje po injiciranju. Slika 2 prikazuje učinke petih injekcij 1 mg / kg AMPH (enkrat na teden) na rezultate lokomotorne aktivnosti v štirih skupinah. ANOVA skupine 4 × 5 je dala glavni učinek seje, F(4, 112) = 8.21, p <0.001, obrobni glavni učinek skupine, F(2, 45) = 3.28, p = 0.085 in brez pomembne interakcije, F(12, 122) <0.77, p > 0.68.
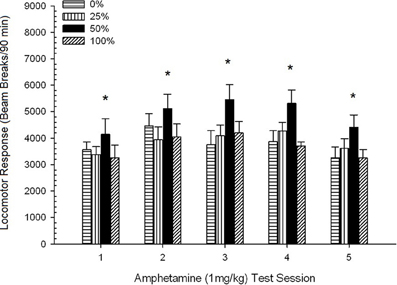
Slika 2. Povprečni (SE) lokomotorni odziv (število prelomov žarka v elektronskem polju na 90 min) na 1 mg / kg d-amfetamina (ip) na tedenskih sejah 5 v skupinah podgan Sprague Dawley (n = 8 / skupina), ki so bile predhodno izpostavljene 15 dnevnim kondicioniranjem z nagrado saharoze (rešitev 10%) pod 0, 25, 50 ali 100% spremenljivimi razporedi. Pogojni dražljaj je bila svetloba (120 s). Skupina 0 je prejela enako število nagrad kot skupina 100 v odsotnosti pogojenih dražljajev. *p <0.05 za povprečno razliko med skupino 50 in skupino 0 ter skupino 100 na podlagi načrtovanih primerjav.
Načrtovane primerjave so pokazale, da se rezultati skupine 50 bistveno razlikujejo od skupine 0, t(14) = 2.19, p = 0.037 in skupina 100, t(14) = 2.36, p = 0.025 [in se razlikuje od skupine 25, t(14) = 2.03, p = 0.051]. Tako je v skupini 50 zanesljivo presežen lokomotorni odziv na 1 mg / kg AMPH v primerjavi z drugimi tremi skupinami v vseh petih testnih obdobjih. Analiza polinomskih trendov je odkrila pomemben kvadratni trend na vseh sejah, F(1, 28) = 32.47, p <0.0001 in nobenih drugih pomembnih trendov, F(1, 28) <1.78, p > 0.19. Slika 2 kaže, da je ta rezultat na sejah odražal vzorec U.
Kontrola za spremembe v nosu, ki se odziva med treningom saharoze
Nadaljnje spremljanje ANCOVA o lokomotornih odzivih na 1 mg / kg AMPH z nosom na nos (CS prisotno) kot kovarijazo, v treh skupinah, ki so prejele CS, je imelo marginalni glavni učinek skupine, F(2, 20) = 3.07, p = 0.069 in ni pomembnih učinkov, povezanih z F(4, 80) <0.05, p > 0.85. Tako prisoten pristop, ki se je odzval med treningom, ni razložil pomembnih sprememb v gibalnem odzivu na 1 mg / kg AMPH v skupinah 25, 50 ali 100.
Nadaljnje spremljanje ANCOVA o lokomotornih odzivih na 1 mg / kg AMPH, s pokesom nosu (CS odsotno) kot kovarialo, je dalo pomemben učinek kovarija, F(1, 27) = 6.17, p = 0.020, pomemben glavni učinek skupine, F(3, 27) = 4.13, p = 0.016, obrobna interakcija x Covariate interakcija, p = 0.080 in nobenih drugih pomembnih učinkov, F(4, 108) <1.48, p > 0.21. Tako je neusoden (nediskriminatoren) pristop, ki se je odzval med treningom, razložil pomembne razlike v gibalnem odzivu na 1 mg / kg AMPH. Vendar se ta variacija ni prekrivala z varianco, povezano s skupino, ker je vključitev kovariata v analizo povečala in ne zmanjšala pomembnost učinka skupine.
Razprava
Podatki o nosu, medtem ko je bil CS prisoten, kažejo, da so skupine pridobile povezavo med dostavo CS in saharozo, kar se odraža v povečanju odgovorov na treningih. Profil odziva na seje, medtem ko je bil CS prisoten, je pokazal, da so bili načrti 100 in 50% CS-ZDA enako učinkoviti pri iskanju pristopa, medtem ko je urnik 25% sprožil bolj skromno povečanje pristopa, ki je bil sprožen. Podatki o nosu, medtem ko je bila CS odsotna, kažejo, da so se skupine, ki so prejele katerega od treh ur CS-saharoze za usposabljanje (skupina 25, 50, 100), hitro naučile zmanjšati nosne pokes v odsotnosti CS, medtem ko so živali v skupini 0 , ki ni prejela nobenega CS, se je po obsežnem usposabljanju le naučila zmanjšati svoje obnašanje pri pristopu v omejenem obsegu.
Podatki o habituaciji kažejo, da se skupine pred AMPH niso razlikovale in da je bila ponavljajoča se izpostavljenost testnim poljem povezana z zmanjšanim lokomotornim odzivom brez zdravil. Razlik med skupinami in povečan odziv na ponavljajoče se odmerke AMPH zato ni mogoče pripisati že obstoječim razlikam v obnašanju lokomotorja.
Rezultati pre-senzibilizacijskega izziva z 0.5 mg / kg AMPH so potrdili, da je zdravilo povečalo lokomotorno aktivnost glede na končni dan brez zdravljenja. V skladu s hipotezo se je skupina 50 glede na srednji odziv na odmerek uvrstila višje kot skupine 0 ali 100 (kot tudi skupina 25), čeprav povprečne razlike med skupinami niso bile pomembne.
Za preobčutljivostne seje so načrtovane primerjave med skupinami pokazale, da je predhodna izpostavljenost 50-odstotni nagrajeni saharozi privedla do znatnega povečanja lokomotornega odziva na 1.0 mg / kg odmerka amfetamina v primerjavi z ostalimi tremi programi. Ta učinek je bil očiten že pri prvem odmerku in se pri ponavljajočih se odmerkih ni bistveno spremenil. Analiza trenda je pokazala dvofazni odziv (za celoten vzorec) na ponavljajoče se odmerke AMPH, ki se je povečal do tretjega odmerka, nato pa zmanjšal. Rezultati nadaljnjega spremljanja ANCOVA z lupljenjem v nosu (CS odsoten) so potrdili, da razlike v gibalnih odzivih štirih skupin na 1 mg / kg AMPH niso bile posredovane z neučinkovitim pristopom med vadbo saharoze.
Učinek skupine med sejami preobčutljivosti je skladen z našo hipotezo. Učinek dvofaznega sevanja ni skladen s pričakovanim nadaljevanjem stopnjevanja lokomotornih odzivov s ponovljenimi odmerki AMPH. To je lahko povezano z intervalom odmerjanja. Za obravnavo tega vprašanja je treba uporabiti postopek (nadomestni dnevni odmerki), ki povzroča dosledno naraščanje lokomotornega odziva na odmerke AMPH (npr. Vedenjska senzibilizacija) na odmerke 1.0 mg / kg. Učinek preobčutljivega režima AMPH na poznejši odziv na drugi izziv 0.5 mg / kg bi še dodatno podprl splošnost tega učinka. Vključitev izločanja slanice pred AMPH bi določila vlogo pričakovanega trajanja ali učinkov, povezanih z injiciranjem (npr. Stres) na lokomotorni odziv na AMPH. Vključitev 75% kondicionirane skupine saharoze bi pripomogla k pojasnitvi vloge nagradne negotovosti v primerjavi z redko nagrajevanjem na vzorcih odgovorov za skupine 50 in skupino 25. Poleg tega, da bi omogočili oceno (z ANCOVA) o prispevku odgovorov, ki ne vključujejo drog, na lokomocijo pod AMPH (z uporabo pokesov z nosom s CS, ki je prisotna kot kovarija), so bili nosni pokes kodirani tudi za skupino 0 v intervalu, ko CS je bil prisoten v ostalih štirih skupinah (tj. tako, da bi se nosne pokes od vseh petih skupin - vključno s skupino 0, ki ni prejel CS-a - lahko vključil v analizo kovarianc s CS, ki je predstavljena kot kovarijata). Te izboljšave so bile vključene v eksperiment 2.
poskus 2
Materiali in metode
Metodologija eksperimenta 2 je bila podobna metodi eksperimenta 1, vendar je bila revidirana, da bi bila bolj približana shema, za katero je bilo ugotovljeno, da zanesljivo inducira senzibilizacijo AMPH (Fletcher et al., 2005). Spremembe so bile naslednje: (a) Skupina 75% CS-saharoza (n = 8); (b) Med treningom saharoze so podgane (razen skupine 0) prejele 20 CS (lahke) predstavitve (v nasprotju z 15 v poskusu 1); (c) vsaka predstavitev CS je bila ločena s povprečnim intervalom med preizkusi 90 s; območje: 30 – 180 s (v primerjavi z 120 s v eksperimentu 1), ki kompenzira povečanje preskusov usposabljanja, da se enačenje trajanja vsakega vadbe enači s poskusom 1; (d) trajanje vsakega od treh sej navajanja je bilo zmanjšano z 120 na 90 min, da bi ustrezalo trajanju testnih sej; (e) Dodana je bila slanica (ip, 1 ml / kg) (90 min) (dan po treningu saharoze 8) za oceno lokomotornih učinkov injekcije po sebi (npr. pričakovanje, stres); (f) seje senzibilizacije zdravila 1 mg / kg so potekale drugače po tednu (dnevi 12 – 21 po treningu) in ne v tedenskih intervalih kot v poskusu 1; (g) Poleg predhodne senzibilizacije 0.5 mg / kg AMPH izziva (9 po dnevu treninga) je bil dodan drugi post-senzibilizacijski 0.5 mg / kg AMPH izziv (dan po treningu saharoze 28), da se preveri splošnost učinek preobčutljivosti med odmerki; (h) nosni pokes med CS je bil kodiran za vse skupine (vključno s skupino 0); (i) nosni pokes, medtem ko CS ni bil prisoten, so bili zabeleženi posebej iz intervala 5-a, tik pred nastopom CS, da bi indeksirali prezgodnji odziv.
Rezultati
Nos pokes med saharozno kondicioniranje
Skupina 5 × 15 Session × 2 faza (CS prisotna, CS odsotna) ANOVA nosnih pokesov je prinesla pomembne glavne učinke skupine, F(4, 19) = 2.89, p = 0.050, seja F(14, 266) = 2.28, p = 0.006 in faza, F(1, 19) = 14.72, p = 0.001, kot tudi pomembna tristranska interakcija, F(56, 266) = 1.38, p = 0.050. Plošče (A, B) na sliki 3 narišite povprečne ocene skupin v nosu za trenutno CS in CS odsotno fazo. Primerjava obeh plošč razkrije, da je glavni učinek Phase odražal bolj splošne odzive na nos, ko je bil CS prisoten v primerjavi z odsotnostjo. Zato so se odzivi z znakom pojavili bistveno pogosteje kot prezgodnji odzivi brez odzivov. Glavni učinki skupine in seje niso bili razloženi zaradi interakcije višjega reda. Ta zadnji rezultat je odražal konvergenco rezultatov za pet skupin na razmeroma stabilni nizki ravni med sejami, ko ni bilo CS (slika 3B), skupaj z razhajanjem rezultatov v visokih (skupina 75, skupina 100), vmesne (skupina 50) in nizke (skupina 0, skupina 25) ravni nosnega sunka, ki se odzovejo na seje, ko je bil CS prisoten (slika 3A). Od trendov polinoma nižjega reda (linearnega, kvadratnega, kubičnega) se je približal le pomen trikotne interakcije za linearni trend, F(4, 19) = 2.32, p = 0.094, ki odraža na splošno monotono povečanje v nosu, ki sega po sejah v skupini 75, in relativno hitrejšo stabilizacijo pri visokih, srednjih in nizkih ravneh odzivanja v drugih skupinah, ko je bil CS prisoten.
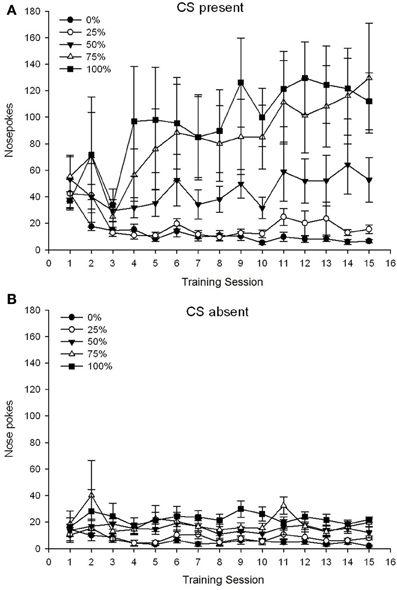
Slika 3. Srednji (SE) pristopni odzivi (nosni pokes) na treningih saharoze 15 v skupinah podgan Sprague Dawley (n = 8 / skupina) izpostavljena nagradi saharoze (rešitev 10%) pod 0, 25, 50, 75 ali 100% spremenljivimi razporedi. Pogojni dražljaj je bila svetloba (120 s). Skupina 0 je prejela enako število nagrad kot skupina 100 v odsotnosti pogojenih dražljajev. () Rezultati, ko je bila CS prisotna (preskušanja 5 s × 20). (B) Rezultati, ko je bila CS odsotna (povprečje za 5 × 20 s, medtem ko je svetloba izklopljena).
Prilagajanje lokomotornim zabojem
ANOVA skupine sej 5 × 3 za lokomotorne odzive brez drog je povzročila pomemben glavni učinek seje, F(2, 70) = 60.01, p <0.0001 in brez drugih pomembnih učinkov, F(4, 35) <0.70, p > 0.60. Načrtovane primerjave skupine 50 s skupino 0 in skupino 100 na prvi in zadnji seji navajanja niso dale pomembnih učinkov, tje <0.84, p > 0.40. Zato se povprečni odziv gibal brez gibanja v ključnih skupinah pred testiranjem ni razlikoval. Povprečno (SE) število prekinitev žarka na 90 minut je bilo 2162 (118) na 1. sejo, 1470 (116) na 2. sejo in 1250 (98) na 3. sejo.
Testne seje
Slanica. ANOVA skupine 5 × 2 je primerjala lokomotorni odziv na končni seji navajanja in fiziološko izločanje. ANOVA je dala glavni učinek seje, F(1, 35) = 62.46, p <0.0001 in brez drugih pomembnih učinkov, F(4, 35) <0.65, p > 0.64. Slika 4 Skupina pomeni in kaže, da je učinek Session odražal splošno zmanjšanje lokomotornega odziva od končne seje navajanja brez droge na sejo slanice, ki se ni razlikovala glede na skupino. Tako se je zmanjšanje lokomotornega odziva, ki smo ga opazili med tremi seji navajanja, nadaljevalo s četrto izpostavljenostjo brez testa na testne škatle.
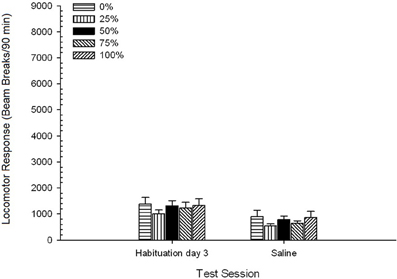
Slika 4. Srednje (SE) lokomotorni odziv (število prelomov žarka v elektronskem polju na 90 min) na zadnjem zasedanju brez navzkrižnega zdravljenja z zdravilom 3 in na naslednji seji po injiciranju fiziološke raztopine (ip, 1 ml / kg) v skupinah zdravila Sprague Dawley podgane (n = 8 / skupina), ki so bile predhodno izpostavljene 15 dnevnim kondicioniranjem z nagrado saharoze (rešitev 10%) pod 0, 25, 50, 75 ali 100% spremenljivimi razporedi. Pogojni dražljaj je bila svetloba (120 s). Skupina 0 je prejela enako število nagrad kot skupina 100 v odsotnosti pogojenih dražljajev.
Učinki zdravila 0.5 mg / kg AMPH.
Premikanje pred injiciranjem. Skupina 5 × 2 Sekcija ANOVA za lokomocijo pred injekcijo (30-min) na pre- in post-senzibilizacijske dni testiranja 0.5 mg / kg AMPH je povzročila pomemben glavni učinek zdravljenja, F(1, 35) = 13.39, p = 0.001 in nobenih drugih pomembnih učinkov, F(4, 35) <1.79, p > 0.15. Načrtovane primerjave niso pokazale pomembnih razlik med skupino 50 in skupino 0 ali skupino 100 na prvi seji, t(70) <1.00, p > 0.30. Vendar je na drugi seji (po preobčutljivosti) skupina 50 (1203; 121) pokazala bistveno več prekinitev žarka pred injiciranjem (M; SE) kot skupina 100 (756; 103), t(70) = 5.11, p <0.001, vendar se ni razlikoval od skupine 0 (1126; 211), t(7) <0.88, p > 0.40. Zato izhodiščne razlike v gibanju niso upoštevale skupinskih razlik v gibalnem odzivu na prvi odmerek AMPH 0.5 mg / kg, so pa morda prispevale k razlikam med odzivnostjo gibalnega odziva na drugi odmerek 50 mg / kg AMPH med skupino 100 in 0.5 . Povprečni (SE) prelomi žarka za fazo pred injiciranjem v prvem in drugem preskusu AMPH 0.5 mg / kg so bili 757 (41) in 974 (59).
Pospeševanje po injiciranju. Zelo pomemben učinek seje je pokazala ANOVA skupine 5 × 2 za sejo gibanja lokomotornega odziva na 0.5 mg / kg AMPH pred in po preobčutljivostnem režimu odmerka 5. F(1, 35) = 76.05, p <0.0001 in brez drugih pomembnih učinkov, F(4, 35) <1.10, p > 0.37. Slika 5 prikazuje povprečne ocene za vsako skupino in sejo.
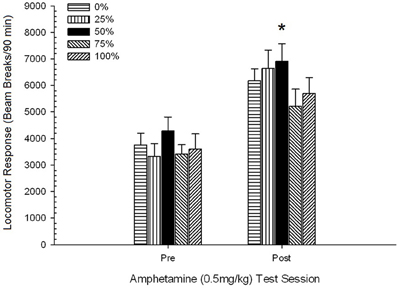
Slika 5. Povprečni (SE) lokomotorni odziv (število prelomov žarka v elektronskem polju na 90 min) na 0.5 mg / kg d-amfetamina na ločenih sejah pred in po preobčutljivosti d-amfetamina na sejo 5 (1.0 mg / kg; ip) na sejo) v skupinah podgan Sprague Dawley (n = 8 / skupina), ki so bile predhodno izpostavljene 15 dnevnim kondicioniranjem z nagrado saharoze (rešitev 10%) pod 0, 25, 50, 75 ali 100% spremenljivimi razporedi. Pogojni dražljaj je bila svetloba (120 s). Skupina 0 je prejela enako število nagrad kot skupina 100 v odsotnosti pogojenih dražljajev. *p <0.05 za povprečno razliko med skupino 50 in skupino 0 ter skupino 100 na podlagi načrtovanih primerjav.
Na sliki je razvidno, da je učinek seje vključeval pomembno povečanje povprečnih (SE) prelomov žarka na 90 min od odmerka 0.5 mg / kg 1, 3674 (216) na 0.5 mg / kg odmerek 2, 6123 (275). Pomanjkanje interakcije ali skupinski učinek je pokazalo, da se senzibilizacija za AMPH med skupinami ni zanesljivo razlikovala. Kljub pomanjkanju pomembnih učinkov, povezanih s skupino, v ANOVA, pregled slike kaže, da je skupina 50 pokazala največji odziv na prvi in drugi odmerek 0.5 mg / kg. Načrtovane primerjave odziva na prvi odmerek 0.5 mg / kg niso pokazale pomembne razlike med skupino 50 in skupino 0 ali skupino 100, tje(35) <0.48, p > 0.50. Kot odgovor na drugi odmerek (po preobčutljivosti) 0.5 mg / kg pa je skupina 50 pokazala znatno večje gibanje kot skupina 0, t(35) = 2.00, p <0.05, kot tudi skupina 100, t(35) = 3.29, p <0.01.
Glede na pomembno skupinsko razliko v gibanju pred injiciranjem na drugem zgoraj opisanem sejanju 0.5 mg / kg AMPH, je bila izvedena spremljanje 5 skupine 2 seje ANCOVA gibalnega odziva na 0.5 mg / kg AMPH. injekcije na drugi seji. Ta analiza je pokazala pomemben učinek kovarija, F(1, 34) = 8.65, p = 0.006, glavni učinek seje F(1, 34) = 10.83, p = 0.002 in nobenih drugih pomembnih učinkov, F(4, 34) <0.85, p > 0.50. Pomembno je, da so načrtovane primerjave na podlagi napake MS in napake df iz ANCOVA potrdile, da je povprečni gibalni odziv na drugi odmerek AMPH 0.5 mg / kg v skupini 50 ostal bistveno večji kot skupina 100, t(34) = 3.09, p <0.01 in skupina 0, t(34) = 1.88, p <0.05 (enostransko), ko je bilo nadzorovano odstopanje pred injiciranjem od 2. seje. Tako je skupina 50 pokazala občutno večji odziv gibalnega gibanja po preobčutljivosti na 0.5 mg / kg AMPH kot skupina 100 ali skupina 0, in te razlike v skupini niso bile posredovane s premikom pred injiciranjem v testnih dneh.
Učinki zdravila 1.0 mg / kg AMPH.
Premikanje pred injiciranjem. Skupina 5 × 5 seja ANOVA rezultatov pred injiciranjem 30-min za seje preobčutljivosti 1 mg / kg AMPH je povzročila glavni učinek seje, F(4, 140) = 16.70, p <0.0001 in brez drugih pomembnih učinkov, F(4, 35) <0.94, p > 0.45. Načrtovane primerjave niso pokazale pomembne razlike v gibanju pred injiciranjem med skupino 50 in skupino 0 ali skupino 100 na prvi seji, t(175) <1.66, p > 0.10. Na zadnji seji pa je skupina 50 (1167; 140) pokazala bistveno več prelomov žarka (M; SE) kot skupina 100 (1000; 99), t(175) = 2.35, p <0.05, vendar se ni razlikoval od skupine 0 (1085, 120), t(175) <1.16, p > 0.20. Zato so razlike v gibanju pred injiciranjem prispevale k razlikam med skupinami 50 in 100 v gibalnem odzivu na končni odmerek AMPH 1 mg / kg. Povprečni (SE) skupni prelomi žarka za vzorec med fazo pred injiciranjem za seje od 1 do 5 so bili: 810 (46), 784 (52), 760 (53), 726 (46), 1009 (51).
Pospeševanje po injiciranju. Skupina 5 × 5 Sekcija ANOVA odzivov na 1 mg / kg AMPH je dala pomemben glavni učinek seje, F(4, 140) = 6.72, p <0.001, obrobna interakcija med skupino in sejo, F(16, 140) = 1.57, p = 0.085 in brez glavnega učinka skupine, F(4, 35) <0.44, p > 0.77. Analize polinomskih trendov so pokazale pomemben linearni trend, F(1, 35) = 9.19, p = 0.005 in kubični trend, F(1, 35) = 21.63, p <0.001, v sejah od 1 do 5. Slika 6 prikazuje srednje vrednosti lokomotorjev za vsako skupino in sejo.
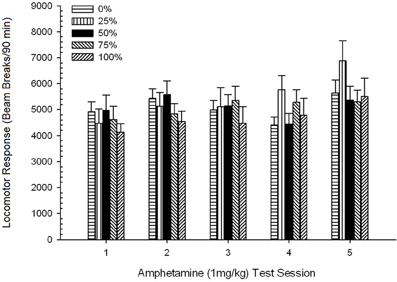
Slika 6. Povprečni (SE) lokomotorni odziv (število prelomov žarka v elektronskem polju na 90 min) na 1 mg / kg d-amfetamina (ip) na tedenskih sejah 5 v skupinah podgan Sprague Dawley (n = 8 / skupina), ki so bile predhodno izpostavljene 15 dnevnim kondicioniranjem z nagrado saharoze (rešitev 10%) pod 0, 25, 50, 75 ali 100% spremenljivimi razporedi. Pogojni dražljaj je bila svetloba (120 s). Skupina 0 je prejela enako število nagrad kot skupina 100 v odsotnosti pogojenih dražljajev.
Na sliki je razvidno, da je učinek Session odražal znatno povečanje povprečnih (SE) prelomov žarkov za celoten vzorec od seje 1, 4624 (213) do seje 5, 5736 (272), kar potrjuje nastanek senzibilizacije za AMPH. Kubični trend označuje relativne maksimume na sejah 1, 3 in 5, s padci na sejah 2 in 4, zlasti za skupine 0 in 50. Na sliki je tudi razvidno, da je skupina 25 kljub pomanjkanju pomembne interakcije postopoma kazala večji odziv lokomotorja na seje in se je bistveno razlikovala od ostalih skupin na sejah 4 in 5 (9 in 22% več kot naslednja najvišja skupina). Načrtovane primerjave so pokazale, da se skupina 50 ni bistveno razlikovala od skupin 0 ali 100, t(175) <0.89, p > 0.40 na prvem ali zadnjem preskusu AMPH 1 mg / kg.
Kontrola za spremembe v nosu, ki se odziva med treningom saharoze
Dve ANCOVA skupini 5 × 2 skupine za lokomotorni odziv na 0.5 mg / kg AMPH pred in po režimu preobčutljivosti, vključno s popolnim pokanjem nosu med treningom saharoze s prisotnostjo CS in CS odsotnimi kot ločenimi kovarijevami, ni našel nobenih pomembnih učinkov za kovarijazo, F(1, 18) <1.03, p > 0.31. Zato pristop, ki se je odzval med treningom, ni posredoval razlik v skupinah pri odzivu na 0.5 mg / kg AMPH.
Dve 5 skupini x 5 seji ANCOVA o gibanju lokomotornega odziva na 1 mg / kg med sejami preobčutljivosti s celotnim nosom (CS prisotni, CS odsotni) kot ločene kovarialnosti niso prinesli pomembnih učinkov kovarije med prisotnostjo CS \ t F(4, 104) <1.04, p > 0.38 in obrobni glavni učinek kovariate, medtem ko CS ni bilo, F(1, 18) = 3.32, p = 0.085.
Razprava
Rezultati te študije niso dosledno podpirali hipoteze, da bi skupina 50 pokazala višji lokomotorni odziv v primerjavi z drugimi skupinami. Podatki o 1 mg / kg AMPH so potrdili pojav preobčutljivosti z alternativnim režimom odmerjanja. Vzorec po skupinah je pokazal trend večje senzibilizacije med slednjimi sejami v skupini 25, brez takšnih dokazov za skupino 50. V nasprotju s tem pa so rezultati odmerjanja zdravila 0.5 mg / kg pokazali trend večje senzibilizacije v skupini 50, hkrati pa so potrdili pomembno splošno povečanje lokomotornega odziva v skupinah na drugi v primerjavi s prvim odmerkom zdravila 0.5 mg / kg AMPH. Nulti učinek injiciranja slanice je potrdil, da pričakovano trajanje ali stres, povezan z injiciranjem, ni prispeval k učinkom AMPH.
Podatki o nosu so ponovno pokazali splošno povečanje pristopa, ki se je odzval v času treninga, ko je bil CS prisoten, brez ustreznega povečanja, ko CS ni bil prisoten. Zato so se zdele živali pridobile povezavo med CS in možnostjo nagrad saharoze. Skupinske razlike v frekvenci nosnih pokesov, ko je bil CS prisoten, je približno ustrezal pogostosti nagrajevanja v skladu z ustreznimi razporedi, pri čemer so skupine 75 in 100 prikazovale najbolj nosne pokes, skupina 50, ki prikazuje vmesne številke nosnih pokesov, in skupine 0 in 25 prikazuje najmanjše nosne pokes. Ti rezultati kažejo, da je CS prišel v nadzorni pristop, ki se je odzval na način, ki je skladen s splošno verjetnostjo nagrajevanja. Čeprav je špekulativno, je lahko ena od možnih razlag za stopnjo spodnjega nosu s CS, ki je prisotna v skupini 50 v eksperimentu 2 v primerjavi s poskusom 1, skrajšanje inter-sujalnega intervala, saj se zdi, da daljši inter-poskusni intervali (poskus 1) spodbujajo impulzivno To je povezano s povečanim prometom DA v sprednji cingularni, predhodni in infralimbi \ tDalley et al., 2002). Zato je lahko zmanjšanje 30% med poskusnim intervalom v eksperimentu 2 (in 3) spremenilo ravni kortikalnega DA in spodbudilo bolj selektivno (tj. Vodeno z relativno pogostostjo nagrajevanja) v primerjavi z impulzivnim (ne vodenim s nagrajevanjem) pristopom. odziv v skupini 50 med poskusnimi poskusi v eksperimentu 2 v primerjavi s poskusom 1.
Pomanjkanje pomembnih sankcijskih učinkov za pokes nosu v stanju CS v ANCOVA kaže, da pristop, ki se odziva med treningom saharoze, ni posredoval učinkov različnih urnikov CS-saharoze na odzive na AMPH. Marginalno pomemben učinek kovarijev za odsotnost stanja CS v ANCOVA lokomotornih odzivih na 1 mg / kg AMPH kaže, da je tendenca k prezgodnjemu odzivu brez zdravil pojasnila nekatere variabilnosti lokomotornih učinkov AMPH med sejami preobčutljivosti.
Skupaj dokazi kažejo, da so učinki anamneze kondicioniranja lahko opaznejši z 0.5 AMPH kot pri 1 mg / kg AMPH in da protokol, ki povzroča preobčutljivost v odsotnosti kakršnih koli drugih manipulacij, lahko zakrije ali povzroči odvečne učinke domnevnega vpliva vedenjska manipulacija, ki spodbuja preobčutljivost (tj. kronična variabilna nagrada).
Vedenjska preobčutljivost za AMPH je močan učinek v laboratoriju. Vendar pa je izven laboratorija le manjšina posameznikov, ki se igrajo kronično, prerasla v patološko raven. Čeprav je tveganje za preobčutljivost povezano s tveganjem za zasvojenost (ali iskanje drog), zlasti za psihostimulante (Vezina, 2004; Flagel et al., 2008), številni dejavniki, razen tveganja za preobčutljivost, lahko povzročijo eno odvisnost (npr. Verdejo-Garcia et al., 2008; Conversano et al., 2012; Volkow et al., 2012). Kljub temu lahko dejavniki lastnosti, ki omogočajo občutljivost za preobčutljivost, vplivajo na zgodovino pogojev, da bi poudarili učinke nepredvidljivega nagrajevanja (tj. 50% CS-US urnik) na reaktivnost sistema DA. Da bi raziskali to možnost, je eksperiment 3 uporabil isti postopek kot eksperiment 2, vendar je uporabil Lewisov sev namesto Sprague Dawley-jevih podgan.
Podgane Sprague Dawley prikazujejo vmesne nivoje DA transporterjev z nižjimi ravnmi kot Wistarjeve podgane (Zamudio in sod., 2005), toda višje ravni kot podgane Wistar-Kyoto (»depresivni« tip) v nucleus accumbens, amigdali, ventralnem tegmentalnem območju in substantia nigra (Jiao in sod., 2003). Ta profil lahko povzroči, da so podgane Sprague Dawley le zmerno občutljive na okoljske ali farmakološke manipulacije funkcije DA. Nasprotno pa imajo Lewis podgane nizke koncentracije DA transporterjev, kot tudi D2 in D3 DA receptorje v nucleus accumbens in dorzalni striatum v primerjavi z drugimi sevi (npr. F344) (Flores et al., 1998). Te morfološke razlike lahko prispevajo k diferencialnemu odzivu podgan Lewis na manipulacije z DA. Podgane Lewis kažejo tudi vrsto poudarjenih odzivov na eksperimentalne manipulacije z zdravili v primerjavi z drugimi sevi (npr. F344). Najpomembneje je, da podgane Lewis kažejo večjo preobčutljivost na metamfetamin, za katero je značilen majhen odziv na začetne odmerke, vendar večji odziv na poznejše odmerke (Camp in sod., 1994). Lewis podgane kažejo tudi večjo lokomotorno senzibilizacijo na številne odmerke kokaina (Kosten et al., 1994; Haile et al., 2001). Na podlagi tega vzorca učinkov smo domnevali, da bi nam Lewisovi podgani omogočili raziskati, ali dovzetnost za preobčutljivost poveča učinke urnika kondicije na poznejši odziv na AMPH.
poskus 3
Materiali in metode
Metodologija je bila enaka kot v eksperimentu 2, razen uporabe Lewisovih podgan (200-225 g ob prihodu, Charles River, Quebec, Kanada).
Rezultati
Nos pokes med saharozno kondicioniranje
Skupina 5 × 15 Session × 2 faza (CS prisotna, CS odsotna) ANOVA nosnih pokesov je prinesla pomembne glavne učinke skupine, F(4, 34) = 6.12, p = 0.001, seja, F(14, 476) = 3.42, p <0.001 in faza, F(1, 34) = 20.83, p <0.001, pa tudi pomembna trosmerna interakcija, F(56, 476) = 1.56, p = 0.008. Plošče (A, B) na sliki 7 narišite povprečne ocene skupin v nosu za trenutno CS in CS odsotno fazo. Primerjava obeh plošč razkrije, da je glavni učinek faze odražal bolj splošne odzive na nos, ko je bil CS prisoten v primerjavi z odsotnostjo. Zato so se znani odzivi pojavljali bistveno pogosteje kot predzreli odzivi. Glavni učinki skupine in seje niso bili zlahka interpretirani zaradi interakcije višjega reda. Trosmerna interakcija je odražala konvergenco rezultatov za pet skupin na razmeroma stabilni nizki ravni med sejami, ko CS ni bilo [Panel (B)], skupaj z razhajanjem rezultatov, ko je bil CS prisoten v relativno diskretnih profilih za vsako skupino, ki vzporedno uvršča njihov vrstni red frekvence nagrajevanja: od najvišje (skupina 100) do najnižje (skupina 25) [Panel ()]. Samo linearni trend interakcije je bil pomemben, F(4, 34) = 4.03, p = 0.009, ki odraža na splošno dosledno povečanje nosnih pokesov po sejah v skupini 100, ko je bila CS prisotna v primerjavi s sorazmerno neskladnim profilom povečanja nosnih pokesov po sejah v drugih skupinah v tej fazi.
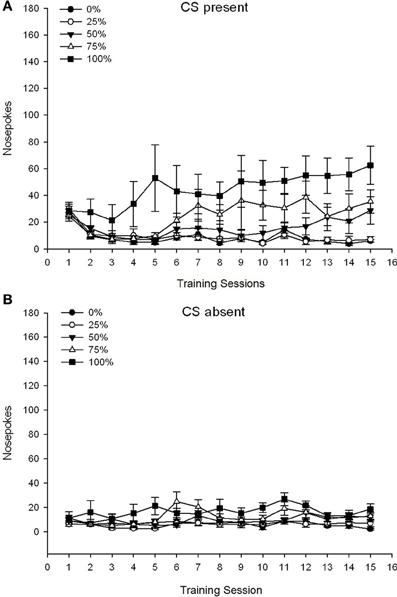
Slika 7. Srednji (SE) pristopni odzivi (nosni pokes) na treningih saharoze 15 v skupinah Lewisovih podgan (n = 8 / skupina) izpostavljena nagradi saharoze (rešitev 10%) pod 0, 25, 50, 75 ali 100% spremenljivimi razporedi. Pogojni dražljaj je bila svetloba (120 s). Skupina 0 je prejela enako število nagrad kot skupina 100 v odsotnosti pogojenih dražljajev. () Rezultati, ko je bila CS prisotna (preskušanja 5 s × 20). (B) Rezultati, ko je bila CS odsotna (povprečje za 5 × 20 s, medtem ko je svetloba izklopljena).
Prilagajanje lokomotornim zabojem
ANOVA skupine 5 × 3 je dala glavni učinek seje, F(2, 70) = 23.07, p <0.0001 in brez drugih pomembnih učinkov, F(8, 70) <1.47, p > 0.18. Ukrivljen vzorec povprečnih gibalnih rezultatov (SE) se je pojavil od seje 1, 1076 (74), do seje 2, 644 (48), do seje 3, 762 (59). Načrtovane primerjave skupine 50 s skupino 0 in skupino 100 na prvi in zadnji seji navajanja so pokazale bistveno manj prelomov žarka v skupini 50 (M = 911; SE = 109) v primerjavi s skupino 0 (M = 1103; SE = 176) na seji navajanja 1, t(105) = 2.02, p <0.05, vendar ni razlike med skupino 50 in skupino 100 (M = 1066; SE = 150), t(105) <1.20, p > 0.20, na tej seji. Skupina 50 se na zadnji seji privajanja ni bistveno razlikovala od skupine 0 ali skupine 100, t(105) <0.93, p > 0.30. Zato se povprečni odziv gibal brez gibanja v ključnih skupinah pred preskušanjem ni dosledno razlikoval.
Testne seje
Slanica. ANOVA skupine 5 × 2 za sejo gibanja lokomotornih odzivov na končni seji navajanja in preizkušanje slanice je pokazala pomemben glavni učinek seje, F(1, 35) = 50.12, p <0.0001 in brez drugih pomembnih učinkov, F(4, 35) <0.57, p > 0.68. Slika 8 prikazuje povprečne ocene skupin za dve seji in kaže, da je učinek Session odražal znatno zmanjšanje od navajanja na test fiziološke raztopine. Prejem injekcije po sebi (npr. pričakovano trajanje, stres) niso izboljšali lokomotornega odziva.
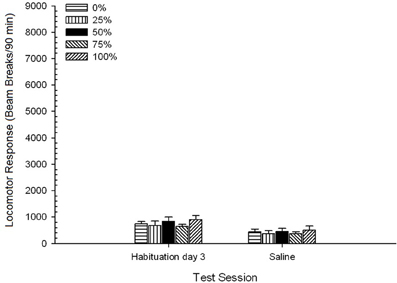
Slika 8. Povprečni (SE) lokomotorni odziv (število prelomov žarka v elektronskem polju na 90 min) na zadnjem zasedanju brez navzkrižnega zdravljenja z zdravilom 3 in na naslednji seji po injiciranju fiziološke raztopine (ip, 1 ml / kg) v skupinah Lewisovih podgan. (n = 8 / skupina), ki so bile predhodno izpostavljene 15 dnevnim kondicioniranjem z nagrado saharoze (rešitev 10%) pod 0, 25, 50, 75 ali 100% spremenljivimi razporedi. Pogojni dražljaj je bila svetloba (120 s). Skupina 0 je prejela enako število nagrad kot skupina 100 v odsotnosti pogojenih dražljajev.
Učinki zdravila 0.5 mg / kg AMPH.
Premikanje pred injiciranjem. ANOVA skupine 5 × 2 seje gibanja pred injiciranjem je povzročila pomemben glavni učinek seje, F(1, 35) = 15.04, p <0.001 in brez drugih pomembnih učinkov, F(4, 35) <1.19, p > 0.33. Načrtovane primerjave niso ugotovile nobene pomembne razlike med skupino 50 in skupino 0 ali skupino 100 na obeh testnih sejah, t(70) <0.99, p > 0.30. Zato osnovne razlike v gibanju pred injiciranjem niso upoštevale skupinskih razlik v gibalnem odzivu na 0.5 mg / kg AMPH. Povprečni (SE) prelomi žarka za fazo pred injiciranjem za prvo in drugo (po preobčutljivostni) 0.5 mg / kg seji so bili 325 (25) in 473 (36).
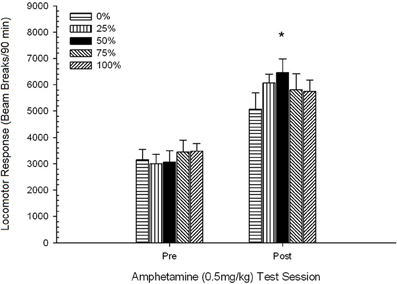
Pospeševanje po injiciranju. Skupina 5 × 2 Sekcija ANOVA za lokomotorni odziv na odmerke 0.5 mg / kg pred in po kroničnem 1 mg / kg AMPH je povzročila glavni učinek seje, F(1, 34) = 87.44, p <0.0001 in brez drugih pomembnih učinkov, F(4, 34) <0.94, p > 0.45. Slika 9 prikazuje povprečne vrednosti lokomotornih vrednosti za vsako skupino in sejo ter kaže, da je učinek seje odražal povečan splošni odziv na drugi odmerek 0.5 mg / kg, skladno s preobčutljivostjo. Slika kaže tudi, da so skupine izvajale zelo podobno na seji 1, toda ta skupina 50 je pokazala več lokomotorne aktivnosti kot druge skupine na seji 2. Načrtovane primerjave v odzivu na prvi odmerek 0.5 mg / kg niso pokazale pomembnih razlik med skupino 50 in skupino 0 ali skupino 100, t(35) <1.28, p > 0.20. Vendar je skupina 50 pokazala bistveno večji odziv gibalnega sistema na drugi odmerek 0.5 mg / kg kot skupina 0, t(35) = 4.32, p <0.001 ali skupina 100, t(35) = 2.24, p <0.05.
Slika 9. Povprečni (SE) lokomotorni odziv (število prelomov žarka v elektronskem polju na 90 min) na 0.5 mg / kg d-amfetamina na ločenih sejah pred in po preobčutljivosti d-amfetamina na sejo 5 (1.0 mg / kg; ip) na sejo) v skupinah podgan Lewis (n = 8 / skupina), ki so bile predhodno izpostavljene 15 dnevnim kondicioniranjem z nagrado saharoze (rešitev 10%) pod 0, 25, 50, 75 ali 100% spremenljivimi razporedi. Pogojni dražljaj je bila svetloba (120 s). Skupina 0 je prejela enako število nagrad kot skupina 100 v odsotnosti pogojenih dražljajev. *p <0.05 za povprečno razliko med skupino 50 in skupino 0 ter skupino 100 na podlagi načrtovanih primerjav.
Učinki zdravila 1 mg / kg AMPH.
Premikanje pred injiciranjem. Skupina 5 × 5 Sekcija ANOVA rezultatov pred injiciranjem 30-min za seje preobčutljivosti je povzročila glavni učinek seje, F(4, 140) = 4.10, p = 0.004 in nobenih drugih pomembnih učinkov, F(4, 35) = 1.25, p > 0.31. Načrtovane primerjave so pokazale, da so bili prelomi žarka med fazo pred vbrizgom (M; SE) v skupini 50 (395; 62) bistveno nižji kot v skupini 100 (508; 62), t(175) = 2.58, p <0.01, ne pa skupine 0, t(175) <1.83, p > 0.10 na seji AMPH 1 mg / kg 1. Na zadnji seji AMPH 1 mg / kg so načrtovane primerjave pokazale tudi, da je bilo gibanje pred injiciranjem v skupini 50 (378; 60) bistveno nižje kot v skupini 100 (650; 75 ), t(175) = 6.17, p <0.001, vendar ne v skupini 0, t(175) <1.84, p > 0.10. Ker je bila smer teh skupinskih razlik (kontrolna skupina = skupina 50) nasprotna predpostavljenemu vzorcu, skupinskih razlik v gibanjih po injiciranju, ki se ujemajo s hipotezo, ni mogoče pripisati razlikam v izhodišču pred injiciranjem. Povprečni (SE) skupni prelomi žarka med fazo pred injiciranjem za seje od 1 do 5 so bili: 442 (34), 452 (32), 542 (40), 411 (26), 504 (37).
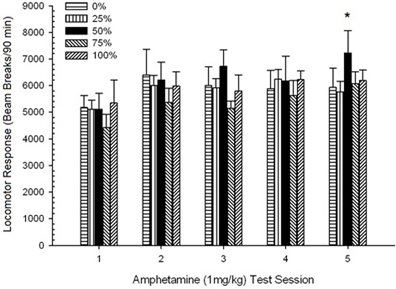
Pospeševanje po injiciranju. Zasedanja skupine 5 × 5 ANOVA odzivov na odmerke 1 mg / kg so pokazala pomemben glavni učinek seje, F(4, 140) = 6.15, p <0.001 in brez drugih pomembnih učinkov, F(4, 35) <0.57, p > 0.68. Analize polinomskih trendov so pokazale pomemben linearni trend, F(1, 35) = 9.34, p = 0.004 in kubični trend, F(1, 35) = 5.08, p = 0.031, slednji rezultat označuje relativne maksimume na sejah 3 in 5. Slika 10 prikazuje rezultate in kaže, da je skupina 50 kljub pomanjkanju pomembne interakcije v ANOVA pokazala bistveno večjo lokomocijo kot druge štiri skupine kot odziv na končni odmerek 1 mg / kg. V skladu s tem so načrtovane primerjave pokazale bistveno večje povprečne rezultate na seji 5 v skupini 50 kot v vseh drugih skupinah, t(35) > 3.68, p <0.001.
Slika 10. Povprečni (SE) lokomotorni odziv (število prelomov žarka v elektronskem polju na 90 min) na 1 mg / kg d-amfetamina (ip) na tedenskih sejah 5 v skupinah podgan Lewis (n = 8 / skupina), ki so bile predhodno izpostavljene 15 dnevnim kondicioniranjem z nagrado saharoze (rešitev 10%) pod 0, 25, 50, 75 ali 100% spremenljivimi razporedi. Pogojni dražljaj je bila svetloba (120 s). Skupina 0 je prejela enako število nagrad kot skupina 100 v odsotnosti pogojenih dražljajev. *p <0.05 za povprečno razliko med skupino 50 in skupino 0 ter skupino 100 na podlagi načrtovanih primerjav.
Kontrola za spremembe v nosu, ki se odziva med treningom saharoze
Dve ANCOVA skupini 5 × 2 skupine za lokomotorni odziv na 0.5 mg / kg AMPH pred in po režimu preobčutljivosti, vključno s popolnim pokanjem nosu med treningom saharoze s prisotnostjo CS in CS odsotnimi kot ločenimi kovarijevami, ni našel nobenih pomembnih učinkov za kovarijazo, F(1, 32) <0.44 p > 0.51. Dve ANCOVA seji skupine 5 × 5 lokomotornega odziva na 1 mg / kg AMPH med sejami preobčutljivosti s skupnimi vdori v nosu (prisoten CS, odsoten CS) kot ločeni kovariati nista povzročila pomembnih učinkov kovariate, medtem ko je bil CS prisoten ali odsoten, F(1, 33) <0.14, p > 0.71. Zato odziv na pristop brez zdravil ni upošteval skupinskih razlik v gibalnih odzivih na noben odmerek AMPH.
Razprava
Preobčutljivost se je razvila do učinkov ponavljajočih se 1.0 mg / kg amfetamina. Podatki o navajanju in fiziološki raztopini potrjujejo, da ta učinek ni bil posledica že obstoječih razlik, pričakovane dobe ali odzivov na injekcijo, povezanih s stresom. ANCOVA-i z zamaški v nosu potrjujejo, da ti učinki niso posledica vedenja brez zdravil. Podatki o poke z nosom kažejo, da so skupine dobile povezavo med CS in možnostjo nagrajevanja saharoze. Stopnja ranga skupin, ki so se odzvale na koncu treninga, se je ujemala s splošno pogostnostjo nagrajevanja v različnih urnikih od najvišje (skupina 100) do najnižje (skupina 0), kot je to storila v poskusu 2. Relativno nižja skupna povprečna vrednost ravni poke v nosu v tem poskusu v primerjavi s poskusoma 1 in 2 lahko odražajo bolj selektiven pristop pri odzivu na nagrade za podgane Lewis (Kosten et al., 2007).
Podatki o odmerkih 0.5 mg / kg so pokazali, da je začetni lokomotorni odziv na AMPH pri podganah Lewisa (slika 3) 9je bil nekoliko zatreti v primerjavi s podganami Sprague Dawley (poskus 2; slika 5), vendar je bilo povečanje odziva na drugi odmerek pri Lewisovih podganah znotraj skupine večje (skoraj dvakrat večji od odziva na prvi odmerek 0.5 mg / kg), potem ko je 5-session AMPH najbolj opazno, skupina 50 je pokazala večji lokomotorni odziv od vseh skupin, razen skupine 25, do druge (tj. post-senzibilizacijske) doze 0.5 mg / kg AMPH in večjega lokomotornega odziva kot vse druge skupine, vključno s skupino 25, do končnega odmerka 1 mg / kg AMPH (končno sejo senzibilizacije). .
Povzetek analize razvrstitev skupin med poskusi
Za določitev zanesljivosti skupinskih razlik v preobčutljivosti je neparametrična analiza ocenila kontingenco med skupino in rangom srednjega lokomotornega odziva na drugi (post-kronični AMPH) odmerek 0.5 mg / kg in končni odmerek 1.0 mg / kg AMPH. iz poskusov 3. Analiza je dala pomemben učinek, φ = 0.986, p = 0.025, ki odraža dejstvo, da je skupina 50 na prvem mestu med vsemi primerjavami. Višji rang skupine 50 v primerjavi z vsemi drugimi skupinami kot odziv na drugi odmerek 0.5 mg / kg na drugi (po kronični AMPH) \ t 5 (eksperiment 2) in slika 9 (poskus 3). Višji rang skupine 50 v primerjavi z drugimi skupinami kot odziv na končni odmerek 1.0 mg / kg je prikazan na sliki 2 (eksperiment 1) in slika 10 (poskus 3). Edina izjema pri tem vzorcu je bil odziv na končni odmerek 1.0 mg / kg pri podganah Sprague-Dawley v poskusu 2.
Splošna razprava
Ta serija eksperimentov je preizkusila hipotezo, da lahko kronična izpostavljenost nagrajenemu podobnemu nagrajevanju senzibilizira možganske poti DA, podobno kot kronična izpostavljenost drogam. Dokazi za takšen učinek kažejo, da lahko nevroplastičnost, enake vrste, za katero misli, da prispeva k zasvojenosti z drogami, povzroči kronična izpostavljenost nepredvidljivim načrtom nagrajevanja. V skladu z literaturo o zasvojenosti z drogami, lokomotorni odziv na 0.5 in 1.0 mg / kg odmerki AMPH indeksirane reaktivnosti DA sistema, z večjo lokomocijo kot odziv na poznejše odmerke, ki operativno določajo preobčutljivost (prim. Robinson in Berridge, 1993; Pierce in Kalivas, 1997; Vanderschuren in Kalivas, 2000).
Na splošno so rezultati v skladu z našo hipotezo. Vendar pa kažejo tudi na znatno variabilnost eksperimentalnih učinkov zaradi postopkovnih dejavnikov. Učinki urnika kondicioniranja so bili skromni, vendar dosledni, pri čemer je skupina 50 pokazala večji odziv kot druge štiri skupine na oba odmerka po petem režimu odmerjanja. Čeprav na splošno F-vrednosti za skupinske učinke v analizah varianc so bile pogosto brez pomena, razlike v ključnih skupinah pa so bile potrjene s parnimi načrtovanimi primerjavami. V zvezi s tem je treba opozoriti, da je »sedanje razmišljanje tisti splošni pomen [za F v ANOVA] ni potrebna. Prvič, hipoteze, ki jih je preskusil splošni test in test večkratne primerjave, so precej drugačne, s precej različnimi stopnjami moči. Na primer, na splošno F dejansko razdeli razlike med skupinami po številu stopenj svobode za skupine. To ima za posledico redčenje celotnega proizvoda F v primeru, ko je več skupinskih sredstev enakovrednih, vendar drugačnih od drugih srednjih vrednosti (Howell, 1992, str. 338). To je ravno tista situacija, ki je bila uporabljena v teh poskusih, kjer se je pričakovalo, da se skupina 50 razlikuje od kontrolnih skupin 0 in skupine 100, vendar za skupino 25 ali skupino 75 ni bila predvidena nobena razlika.
Podatki o nosu so potrdili, da so živali v vsakem poskusu pridobile povezavo med CS in možnostjo nagrad saharoze. Ujemanje med frekvenco nosa za različne skupine in splošno pogostost nagrajevanja v skladu s svojimi urniki usposabljanja kaže, da se odziva na povprečno stopnjo nagrajenega saharoze. Vendar pa je pomanjkanje posredovalnega učinka nosnega pokrova na skupine, povezane z lokomotornimi odzivi na AMPH v ANCOVA, pokazalo, da ločeni procesi temeljijo na teh dveh vedenjih.
V nekaterih primerih je bil učinek urnika kondicije viden kot odziv na prvi odmerek AMPH; v drugih primerih se je pojavil le po večkratnih odmerkih. Razlike v skupini lokomotornega odziva na prvi odmerek AMPH kažejo, da je izpostavljenost igralniškemu nagrajevanju dovolj sama po sebi, da povzroči preobčutljivost. Skupinske razlike v gibanju po večkratnih odmerkih AMPH kažejo na bolj subtilen učinek, ki bi ga lahko označili kot „občutljivost“, ki se kaže le v kombinaciji s stalno izpostavljenostjo primarnemu senzibilizatorju (tj. Amfetaminu).
Razlike v vzorcu odziva med poskusi kažejo, da lahko daljši interval med usposabljanjem in začetnim izzivom AMPH poveča možnost za odkrivanje inherentnega senzibilizacijskega učinka kondicioniranja. To pa kaže, da se lahko učinki pogojene izpostavljenosti nagrajevanja sčasoma inkubirajo, kar se pojavi tudi pri senzibilizaciji s stimulansi (Grimm et al., 2006). Vzorec odziva na dva odmerka amfetamina kaže, da je lahko odmerek zdravila 0.5 mg / kg učinkovitejši pri razkrivanju učinkov zgodovine kondicioniranja. To pa kaže, da so učinki kondicioniranja pod trenutnim protokolom o usposabljanju nekoliko subtilni in jih lahko prikrivajo učinki stropa pod odmerki AMPH in pogoji, ki ustvarjajo de novo preobčutljivost.
V eksperimentu 3 je dvofazni vzorec odziva na odmerke 0.5 mg / kg in progresivni pojav superiornosti v skupini 50 skladen s pričakovanim profilom za podgane Lewis kot odgovor na metamfetamin (Camp in sod., 1994). To podpira veljavnost sedanjih ugotovitev in kaže na prekrivanje med dejavniki, ki zmerno vplivajo na senzibilizacijo psihostimulantov in na igre na srečo podobne načrte nagrajevanja.
Po poskusih je post-senzibilizacijski lokomotorni odziv skupine 50 na splošno presegel odziv drugih skupin pri različnih odmerkih amfetamina in različnih sevih živali. Vendar pa visoka variabilnost znotraj skupine in skromne velikosti medskupinskega učinka kažejo na vlogo drugih dejavnikov v reaktivnosti DA sistema na amfetamin po izpostavljenosti različnim časovnim razporedom kondicionirane nagrade saharoze. Čeprav odzivi DA nevronov na nagrajevanje signalov lahko zagotovijo grob model igranja (Fiorillo et al., 2003), tako kot vsi modeli, obstaja izguba informacij zaradi varčevanja, tj. prikazati ključni proces. Posledično je vzorec učinkov v pogojih CS-US v originalnih Fiorillo et al. Študija ni v celoti posplošena na lokomotorni odziv na amfetamin. Potrebne so nadaljnje izboljšave modela, da bi v celoti zajeli vidike iger na srečo, ki vplivajo na delovanje sistema DA.
Če povzamemo, rezultati te serije eksperimentov zagotavljajo začasno podporo hipotezi, da kronična izpostavljenost nagrajevanja, podobnega igranju, povečuje reaktivnost sistema možganskega DA na psihostimulantni izziv. Kot take razširjajo ugotovitve Singer et al. (2012) ki so pokazali, da v primerjavi s fiksnim časovnim razporedom predhodna izpostavljenost variabilnemu urniku okrepitve v operantni paradigmi povečuje poznejši lokomotorni odziv na amfetamin. Natančneje, sedanje ugotovitve kažejo na negotovost pri zagotavljanju nagrajevanja kot kritični dejavnik, na katerem temeljijo učinki spremenljivega plačila. Obseg učinkov v operantni paradigmi je bil bistveno večji od učinkov, ugotovljenih v teh poskusih. To lahko odraža večjo kronično izpostavljenost dejavnosti, podobnih igranju na srečo (dnevi 55 v primerjavi z dnevi 15); lahko odraža učinke, ki zahtevajo operantni odziv, da se doseže nagrada (tj. vloga za agencijo), ne pa pasivna izpostavljenost, kot v tej študiji. Povečanje trajanja usposabljanja v sedanji paradigmi bi pomagalo rešiti ta vprašanja.
Veljavnosti variabilnega načrta nagrajevanja in okrepitve kot modelov iger na srečo ni mogoče pridobiti iz teh poskusov. To vprašanje bi lahko obravnavali prihodnje raziskave, ki preučujejo vpliv zgodovine pogojev na obnašanje tveganega tveganja pri igrah na srečo pri glodalcih. Podobno mora biti ustreznost med vedenjsko senzibilizacijo, ki jo najdemo tukaj, in povišanim odzivom striatnega DA na amfetamine, ki so jih pred kratkim odkrili patološki hazarderji, čakati na nadaljnje preiskave (Boileau et al., 2013). Mikro-dializa bi lahko rešila to vprašanje in napoved, ki temelji na podatkih za ljudi, bi bila, da bi bilo večje sproščanje DA v skupini 50 »fenotip kockanja« najbolj opazno v hrbtnem (senzimotornem) striatumu namesto v ventralnem (limbičnem) striatumu . Validacija 50% variabilne izpostavljenosti CS + nagrajevanja v teh drugih paradigmah bi podprla njeno uporabnost kot dobronamerni eksperimentalni model PG.
Medtem ko nekatere oblike iger na srečo jasno vključujejo instrumentalni odziv (npr. Igralni avtomati), pri drugih oblikah iger na srečo (npr. Loterija) povezava med akcijo (nakup vstopnice, tj. Stava), znaki za nagrado (tj. , številke loterije) in sama nagrada (dobitna številka in denarno izplačilo) je veliko bolj razpršena. Kljub temu lahko pride do aktivacije DA med intervalom CS-US. To lahko pojasni, zakaj je ob razglasitvi "dobitne številke" pritegnjena pozornost, saj vsaka posamezna kroglica loterije zaporedoma pade, da sestavi določeno zaporedje številk v dobitni številki. Čeprav je verjetnost, da se bo zgodila določena številka, matematično določena, je izid vsake posamezne loterijske žoge dvojiški - zadetek (se ujema z igralčevo številko) ali zgrešen (ne ujema z igralčevo številko) - in rezultat na katerem koli preizkusu ni znan. Tak scenarij lahko bolje označi izkušnjo skupine 50 v sedanjih poskusih, kjer je bila nagrada dodeljena nenapovedano, a tudi nepredvidljivo, CS pa je zgolj nakazal potencial za nagrado, ne da bi razkril, ali bi do nje prišlo v danem poskusu. Igralni avtomati so močneje povezani s PG kot loterijski vstopnice (Cox et al., 2000; Bakken in sod., 2009), kar kaže na pomembno vlogo instrumentalnih dejavnikov (in neposrednosti) pri nagrajevanju vidikov iger na srečo za to populacijo (Loba in sod., 2001). Kljub temu se zdi, da je Pavlovov proces, ki je bil modeliran v sedanjih poskusih (nagrada CS + negotova), nujen, če ne zadosten element izkušnje z igrami na srečo.
Poleg pomanjkanja jasne instrumentalne zahteve so lahko številne druge značilnosti oblikovanja prispevale k razmeroma skromnemu in spremenljivemu vzorcu eksperimentalnih učinkov. Skupine so se med seboj razlikovale glede skupne izpostavljenosti saharozi, pa tudi glede na primere med nagrado CS in saharozo. Čeprav je to lahko prispevalo k variabilnosti med skupinami, ni mogoče zlahka pojasniti, zakaj so bile živali z največjo izpostavljenostjo saharozi (skupina 100) manj občutljive kot skupina 50. Poleg tega skupina 0 ni prejela nobenega dražljaja pred izpostavljenostjo saharozi na vsakem preskušanju. Čeprav je to izključevalo pričakovano nagrajevanje, ki bi ga povzročilo iztočnico, ni nadzoroval prisotnosti dražljaja pred dostavo nagrad, ki je obstajal v vseh drugih skupinah. Za reševanje tega vprašanja bi morale prihodnje raziskave vključevati stanje, v katerem živali prejemajo nagrado pri vsakem preskušanju po naključni izpostavljenosti nevtralnemu stimulusu (tj., Čigar prisotnost ne signalizira možnosti za nagrado).
Druga omejitev zasnove je potencialni pojav dodatnega vedenja, ki bi lahko vplival na učinek urnika usposabljanja. Glede na negotovost lahko živali razvijejo vraževerno vedenje, namenjeno povečanju zaznanega nadzora in zmanjšanju aktivacije DA pri negotovosti (prim. Harris et al., 2013). Zato je mogoče, da so nenadzorovani vidiki zasnove eksperimenta omogočili živalim, da izravnajo učinke urnika načrtovanja. Tak učinek bi lahko prispeval k relativno skromnemu in spremenljivemu odzivu na amfetamin v skupini 50 po treningu CS + saharoze. Prihodnje raziskave bi morale zabeležiti spontano vedenje, poleg pokesov za nos, med treningi, da bi preizkusili to možnost, in nadzor za to statistično, če bi se pojavila. Ker se pričakuje, da bo takšno obnašanje preprečilo ali ublažilo učinke negotovosti zaradi urnika, je treba lokomotorni odziv na amfetamin v skupini 50 okrepiti, ko je nadzorovan (proceduralno ali statistično). Zato sedanja (nekontrolirana) zasnova zagotavlja konzervativen preskus učinkov nagrade 50% CS + na senzibilizacijo amfetamina.
Z vidika zunanje veljavnosti uporaba moških podgan omejuje posplošitev rezultatov. Pomanjkanje jasnega pogoja kaznovanja se razlikuje tudi od iger na srečo, kjer so velike denarne izgube pogoste in imajo pomembne motivacijske učinke (Nieuwenhuis et al., 2005; Singh in Khan, 2012). Možnost nagrajevanja je odsotna tudi iz sedanje paradigme in ugotovljeno je, da so kumulativni dobički v igri igralnih avtomatov v interakciji z manipulacijami DA pri ljudeh (Tremblay et al., 2011; Smart et al., 2013). Podobno je priložnost za jackpot pomembna razlika med trenutnim modelom in dejanskim igranjem na srečo.
Kljub tem omejitvam sedanji rezultati kažejo, da 50% variabilna nagrada CS + lahko vključi poti DA, ki so vpletene v okrepitev učinkov iger na srečo (Fiorillo et al., 2003; Anselme, 2013). Navzkrižna senzibilizacija odziva na AMPH po tem razporedu, podobnem igrah na srečo, je v skladu s ključno vlogo DA pri učinkih iger na srečo in psihostimulantih (Zack in Poulos, 2009) in razširja prejšnje študije o navzkrižnem motiviranju motivacije za igranje z AMPH pri patoloških hazarderjih (Zack in Poulos, 2004). Predstavljeni rezultati tudi posredno kažejo, da lahko skromni odmerki AMPH, ki ne povzročajo supra-fiziološkega sproščanja DA, bolje modelirajo možgansko aktivnost kot odziv na intermitentne signale nagrajevanja (npr. Med igranjem na srečo) kot izpostavljenost visokim (npr. Binge-like) odmerki stimulativnih zdravil (prim. Vanderschuren in Pierce, 2010). Neposredna podpora za to ujemanje se lahko izvede z ocenjevanjem sproščanja DA kot odzivom na 50-ov variabilni urnik CS-US in različnih odmerkov AMPH z uporabo mikrodialize.
Z eksperimentalnega stališča se zdi, da sedanji Pavlovski model in prejšnji operantni model variabilne ojačitve ustvarjata fenotip, ki spominja na človeškega patološkega kockarja. Kot taki zagotavljajo dragoceno dopolnilo pri igrah na srečo pri glodalcih, ki modelirajo obnašanje iger na srečo (kot odvisen ukrep), vendar so do sedaj uporabljale le zdrave živali, ki so enakovredne človeškim socialnim gamblerjem. Glede na literaturo se lahko živali, ki so kronično izpostavljene variabilni nagradi, v teh nalogah zelo razlikujejo, zlasti kot odgovor na DA-ergična zdravila. Kombinacija fenotipa iger na srečo s podganami pri igrah na srečo lahko dovoljuje sistematičen razvoj zdravil za zdravljenje PG, ki morda ne bodo v celoti doseženi samo pri zdravih živalih. Nadaljnje izpopolnitve pri načrtovanju eksperimenta in režimu usposabljanja, kot je opisano zgoraj, bi morale izboljšati korespondenco med živalmi, ki so bile usposobljene v tej paradigmi, in dejanskimi patološkimi hazarderji.
S klinično-sociološkega stališča je ugotovitev, da je izpostavljenost 50% variabilni nagradi CS +, ki se zelo ujema z urnikom nagrajevanja na komercialnem igralnem avtomatu (Tremblay et al., 2011), spremeni možganski sistem DA na zanesljive in trajne načine, kar nakazuje, da je v nekaterih primerih dejavnost iger na srečo, kot so zlorabe drog, lahko »patogen«, ki lahko povzroči zasvojenost. Vendar skromna velikost učinka in velika variabilnost kot odziv na nagrado 50% CS + kažejo, da je, tako kot zloraba drog, tendenca nagrajevanja, podobna nagradam za igre na srečo, v veliki meri odvisna od že obstoječega profila tveganja igralca. Kljub temu se zdi, da je za preprečevanje izpostavljenosti visoko tveganih posameznikov potencialnim škodljivim učinkom, povezanim z igrami na srečo, smiselno, da se politike, ki se uporabljajo za preprečevanje uporabe in zmanjševanje škode zaradi zlorabe drog, razširijo tudi na igre na srečo.
Izjava o konfliktu interesov
Avtorji izjavljajo, da je bila raziskava izvedena v odsotnosti komercialnih ali finančnih odnosov, ki bi se lahko razumeli kot potencialno navzkrižje interesov.
Priznanja
Ta raziskava je bila financirana s subvencijami Kanadskega sveta za naravoslovne in tehnične raziskave Paulu J. Fletcherju. Iskreno se zahvaljujemo gospe Đurđi Đorđević za pripravo številk.
Reference
Anselme, P. (2013). Dopamin, motivacija in evolucijski pomen vedenja, podobnega igranju na srečo. Behav. Brain Res. 256, 1 – 4. doi: 10.1016 / j.bbr.2013.07.039
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
APA (2013). Diagnostični in statistični priročnik o duševnih motnjah, 5th Edn. Arlington, VA: American Psychiatric Publishing.
Bakken, IJ, Gotestam, KG, Grawe, RW, Wenzel, HG in Oren, A. (2009). Obnašanje pri igrah na srečo in težave z igrami na Norveškem 2007. Scand. J. Psychol. 50, 333 – 339. doi: 10.1111 / j.1467-9450.2009.00713.x
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Boileau, I., Payer, D., Chugani, B., Lobo, DS, Houle, S., Wilson, AA, et al. (2013). In vivo dokazi za večjo sproščanje dopamina pri amfetaminu pri patološkem igranju: pozitronska emisijska tomografska študija z [C] - (+) - PHNO. Mol. Psihiatrija doi: 10.1038 / mp.2013.163. [Epub pred tiskanjem].
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Bolles, RC (1972). Okrepitev, pričakovano trajanje in učenje. Psihol. Rev. 79, 394 – 409. doi: 10.1037 / h0033120
Camp, DM, Browman, KE in Robinson, TE (1994). Učinki metamfetamina in kokaina na motorično vedenje in zunajcelični dopamin v ventralnem striatumu Lewisa v primerjavi s podganami Fischer 344. Brain Res. 668, 180–193. doi: 10.1016/0006-8993(94)90523-1
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Conversano, C., Marazziti, D., Carmassi, C., Baldini, S., Barnabei, G. in Dell'Osso, L. (2012). Patološka igra na srečo: sistematičen pregled biokemičnih, nevro-slikovnih in nevropsiholoških ugotovitev. Harv. Rev. Psihiatrija 20, 130 – 148. doi: 10.3109 / 10673229.2012.694318
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Cox, BJ, Kwong, J., Michaud, V., in Enns, MW (2000). Problem in verjetno patološko igranje na srečo: premisleki iz raziskovanja skupnosti. Can. J. Psihiatrija 45, 548-553.
Dalley, JW, Theobald, DE, Eagle, DM, Passetti, F., in Robbins, TW (2002). Primanjkljaji v kontroli impulzov, povezani s tonično-zvišano serotonergično funkcijo v prefrontalnem korteksu podgan. Neuropsychopharmacology 26, 716–728. doi: 10.1016/S0893-133X(01)00412-2
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Everitt, BJ in Robbins, TW (2005). Nevronski sistemi okrepitve za odvisnost od drog: od dejanj do navad do prisile. Nat. Neurosci. 8, 1481 – 1489. doi: 10.1038 / nn1579
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Fiorillo, CD, Tobler, PN, in Schultz, W. (2003). Diskretno kodiranje verjetnosti nagrajevanja in negotovosti z dopaminskimi nevroni. Znanost 299, 1898 – 1902. doi: 10.1126 / science.1077349
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Flagel, SB, Watson, SJ, Akil, H., in Robinson, TE (2008). Posamezne razlike pri pripisovanju spodbujevalne pomembnosti za nagrado, povezano z namenom: vpliv na senzibilizacijo kokaina. Behav. Brain Res. 186, 48 – 56. doi: 10.1016 / j.bbr.2007.07.022
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Fletcher, PJ, Tenn, CC, Rizos, Z., Lovic, V., in Kapur, S. (2005). Preobčutljivost na amfetamin, ne pa tudi na PCP, poslabša spreminjanje nastavitev pozornosti: preobrat z agonistom D1 receptorja, vbrizganim v medialni prefrontalni korteks. Psihofarmakologija (Berl.) 183, 190–200. doi: 10.1007/s00213-005-0157-6
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Flores, G., Wood, GK, Barbeau, D., Quirion, R., in Srivastava, LK (1998). Lewis in Fischer podgane: primerjava ravni dopaminskega transporterja in receptorjev. Brain Res. 814, 34–40. doi: 10.1016/S0006-8993(98)01011-7
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Frascella, J., Potenza, MN, Brown, LL in Childress, AR (2010). Skupne možganske ranljivosti odprejo pot za nesamostojne odvisnosti: rezba zasvojenosti v novem sklepu? Ann. NY Acad. Sci. 1187, 294 – 315. doi: 10.1111 / j.1749-6632.2009.05420.x
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Grimm, JW, Buse, C., Manaois, M., Osincup, D., Fyall, A., in Wells, B. (2006). Časovno odvisna disociacija učinkov odziva na odmerek kokaina na željo po saharozi in gibanje. Behav. Pharmacol. 17, 143 – 149. doi: 10.1097 / 01.fbp.0000190686.23103.f8
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Haile, CN, Hiroi, N., Nestler, EJ, in Kosten, TA (2001). Diferencialni vedenjski odzivi na kokain so povezani z dinamiko mezolimbičnih dopaminskih proteinov v podganah Lewisa in Fischerja 344. Synapse 41, 179 – 190. doi: 10.1002 / syn.1073
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Harris, JA, Andrew, BJ in Kwok, DW (2013). Pristop revije v času signala za hrano je odvisen od pavlovskega, ne instrumentalnega, kondicioniranja. J. Exp. Psihol. Anim. Behav. Proces. 39, 107 – 116. doi: 10.1037 / a0031315
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Howell, DC (1992). Statistične metode za psihologijo. Boston, MA: Duxbury.
Ito, R., Dalley, JW, Robbins, TW in Everitt, BJ (2002). Sproščanje dopamina v dorzalnem striatumu med obnašanjem, ki išče kokain, pod nadzorom opozorila, povezanega z zdravilom. J. Neurosci. 22, 6247 – 6253.
Jiao, X., Pare, WP in Tejani-Butt, S. (2003). Razlike v sevih v porazdelitvi mest transporterja dopamina v možganih podgan. Prog. Nevropsihofarmakol. Biol. Psihiatrija 27, 913–919. doi: 10.1016/S0278-5846(03)00150-7
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Koob, GF in Le Moal, M. (2008). Pregled. Nevrobiološki mehanizmi za motivacijske procese nasprotnikov v odvisnosti. Philos. Trans. R. Soc. Lond. B Biol. Sci. 363, 3113 – 3123. doi: 10.1098 / rstb.2008.0094
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Kosten, TA, Miserendino, MJ, Chi, S., in Nestler, EJ (1994). Fischerjevi in Lewisovi sevi podgan kažejo diferencirane učinke kokaina v pogojenih preferencah na mestu in vedenjski senzibilizaciji, vendar ne pri lokomotorni aktivnosti ali pogojeni averziji okusa. J. Pharmacol. Exp. Ther. 269, 137 – 144.
Kosten, TA, Zhang, XY in Haile, CN (2007). Razlike v sevanju v vzdrževanju kokainske samouprave in njihova povezanost z novimi aktivnostmi. Behav. Neurosci. 121, 380 – 388. doi: 10.1037 / 0735-7044.121.2.380
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Leeman, RF, in Potenza, MN (2012). Podobnosti in razlike med patološkimi igrami in motnjami uporabe snovi: poudarek na impulzivnosti in kompulzivnosti. Psihofarmakologija (Berl.) 219, 469–490. doi: 10.1007/s00213-011-2550-7
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Leshner, AI (1997). Odvisnost je bolezen možganov in je pomembna. Znanost 278, 45 – 47. doi: 10.1126 / science.278.5335.45
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Loba, P., Stewart, SH, Klein, RM in Blackburn, JR (2001). Manipulacije s značilnostmi iger standardnih video loterijskih terminalov (VLT): učinki pri patoloških in nepatoloških igralcih iger na srečo. J. Gambl. Stud. 17, 297 – 320. doi: 10.1023 / A: 1013639729908
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Martinez, D., Narendran, R., Foltin, RW, Slifstein, M., Hwang, DR, Broft, A., et al. (2007). Sproščanje dopamina, povzročeno z amfetaminom: izrazito zavito v odvisnosti od kokaina in napovedovanje izbire za samo-dajanje kokaina. Am. J. Psihiatrija 164, 622 – 629. doi: 10.1176 / appi.ajp.164.4.622
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Mateo, Y., Lack, CM, Morgan, D., Roberts, DC, in Jones, SR (2005). Zmanjšana funkcija dopaminskega terminala in neobčutljivost na kokain po samoupravljanju in pomanjkanju kokaina. Neuropsychopharmacology 30, 1455 – 1463. doi: 10.1038 / sj.npp.1300687
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Nestler, EJ (2001). Molekularna osnova dolgoročne plastičnosti, ki je v osnovi zasvojenosti. Nat. Rev. Neurosci. 2, 119 – 128. doi: 10.1038 / 35053570
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Nieuwenhuis, S., Heslenfeld, DJ, von Geusau, NJ, Mars, RB, Holroyd, CB, in Yeung, N. (2005). Aktivnost v možganskih območjih, občutljivih na nagrajevanje, je močno odvisna od konteksta. Neuroimage 25, 1302 – 1309. doi: 10.1016 / j.neuroimage.2004.12.043
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Pierce, RC in Kalivas, PW (1997). Model vezja za izražanje vedenjske senzibilizacije do amfetaminskih podobnih psihostimulantov. Brain Res. Brain Res. Rev. 25, 192. doi: 10.1016 / S0165-0173 (97) 00021-0
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Robbins, TW, in Everitt, BJ (1999). Zasvojenost z drogami: slabe navade. Narava 398, 567 – 570. doi: 10.1038 / 19208
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Robinson, TE, Becker, JB in Presty, SK (1982). Dolgoročno olajšanje rotacijskega obnašanja, ki ga povzroča amfetamin, in sproščanje dopamina pri striatah, ki ga povzroči enkratna izpostavljenost amfetaminu: razlike med spoloma. Brain Res. 253, 231–241. doi: 10.1016/0006-8993(82)90690-4
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Robinson, TE, in Berridge, KC (1993). Nevronske osnove za hrepenenje po drogah: teorija o zasvojenosti, ki spodbuja občutljivost. Brain Res. Brain Res. Rev. 18, 247 – 291. doi: 10.1016 / 0165-0173 (93) 90013-P
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Robinson, TE, in Berridge, KC (2001). Spodbujevalna senzibilizacija in zasvojenost. Odvisnost 96, 103 – 114. doi: 10.1046 / j.1360-0443.2001.9611038.x
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Singer, BF, Scott-Railton, J. in Vezina, P. (2012). Nepredvidljiva ojačitev saharina poveča lokomotorni odziv na amfetamin. Behav. Brain Res. 226, 340 – 344. doi: 10.1016 / j.bbr.2011.09.003
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Singh, V., in Khan, A. (2012). Sprejemanje odločitev v nagradnih in kaznih variantah pri igralništvu na Iovu: dokaz "predvidevanja" ali "uokvirjanja"? Spredaj. Neurosci. 6: 107. doi: 10.3389 / fnins.2012.00107
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Skinner, BF (1953). Znanost in človeško vedenje. New York, NY: Free Press.
Smart, K., Desmond, RC, Poulos, CX in Zack, M. (2013). Modafinil poveča nagrajevanje nagrad v igri igralnih avtomatov pri patoloških škodljivih igralcih z nizko in visoko impulznostjo. Neurofarmakologija 73, 66 – 74. doi: 10.1016 / j.neuropharm.2013.05.015
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Tremblay, AM, Desmond, RC, Poulos, CX in Zack, M. (2011). Haloperidol spreminja instrumentalne vidike iger na srečo pri patoloških hazarderjih in zdravih kontrolah. Addict. Biol. 16, 467 – 484. doi: 10.1111 / j.1369-1600.2010.00208.x
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Vanderschuren, LJ in Kalivas, PW (2000). Spremembe dopaminergičnega in glutamatergičnega prenosa pri indukciji in izražanju vedenjske senzibilizacije: kritični pregled predkliničnih študij. Psihofarmakologija (Berl.) 151, 99 – 120. doi: 10.1007 / s002130000493
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Vanderschuren, LJ, in Pierce, RC (2010). Senzibilizacijski procesi v odvisnosti od drog. Curr. Na vrh. Behav. Neurosci. 3, 179 – 195. doi: 10.1007 / 7854_2009_21
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Vanderschuren, LJ, Schoffelmeer, AN, Mulder, AH in De Vries, TJ (1999). Dopaminergični mehanizmi, ki posredujejo pri dolgotrajnem izražanju lokomotorne senzibilizacije po predhodni izpostavljenosti morfinu ali amfetaminu. Psihofarmakologija (Berl.) 143, 244 – 253. doi: 10.1007 / s002130050943
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Verdejo-Garcia, A., Lawrence, AJ in Clark, L. (2008). Impulzivnost kot oznaka ranljivosti za motnje uporabe snovi: pregled ugotovitev raziskav z visokim tveganjem, problematičnih igralcev in študij genetskih združenj. Neurosci. Biobehav. Rev. 32, 777 – 810. doi: 10.1016 / j.neubiorev.2007.11.003
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Vezina, P. (2004). Preobčutljivost reaktivnosti dopaminskega nevrona na srednji možgani in samo-dajanje psihomotoričnih stimulansov. Neurosci. Biobehav. Rev. 27, 827 – 839. doi: 10.1016 / j.neubiorev.2003.11.001
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Volkow, ND, Wang, GJ, Fowler, JS, Logan, J., Gatley, SJ, Hitzemann, R., et al. (1997). Zmanjšan dopaminergični odziv pri striatah pri detoksificiranih osebah, odvisnih od kokaina. Narava 386, 830 – 833. doi: 10.1038 / 386830a0
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Volkow, ND, Wang, GJ, Fowler, JS, in Tomasi, D. (2012). Vezje odvisnosti v človeških možganih. Annu. Rev. Pharmacol. Toxicol. 52, 321 – 336. doi: 10.1146 / annurev-pharmtox-010611-134625
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Winer, B. (ur.). (1971). Statistična načela pri načrtovanju eksperimentov. New York, NY: McGraw-Hill.
Zack, M., in Poulos, CX (2004). Amfetamin namesti motivacijo za kockanje in semantične mreže, povezane z igrami na srečo, v problematičnih hazarderjih. Neuropsychopharmacology 29, 195 – 207. doi: 10.1038 / sj.npp.1300333
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Zack, M., in Poulos, CX (2009). Vzporedne vloge za dopamin pri patološkem hazardiranju in psihostimulantni zasvojenosti. Curr. Zloraba drog Rev. 2, 11 – 25. doi: 10.2174 / 1874473710902010011
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Zamudio, S., Fregoso, T., Miranda, A., De La Cruz, F., in Flores, G. (2005). Razlike v sevih odmerkov dopaminskih receptorjev in vedenja, povezanih z dopaminom pri podganah. Brain Res. Bull. 65, 339 – 347. doi: 10.1016 / j.brainresbull.2005.01.009
Pubmed Abstract | Objavljeno celotno besedilo | CrossRef Celotno besedilo
Ključne besede: patološko igranje, senzibilizacija, amfetamin, dopamin, negotovost
Navedba: Zack M, Featherstone RE, Mathewson S in Fletcher PJ (2014) Kronična izpostavljenost nagnjenega predvidevalnega dražljaja, podobnega kockanju, lahko spodbudi senzibilizacijo amfetamina pri podganah. Spredaj. Behav. Neurosci. 8: 36. doi: 10.3389 / fnbeh.2014.00036
Prejeto: 01 November 2013; Sprejeto: 23 januar 2014;
Objavljeno na spletu: 11 februar 2014.
Uredil:
Bryan F. Singer, Univerza v Michiganu, ZDA
Pregledal:
Louk VanderschurenUniverza v Utrechtu, Nizozemska
Ruud Van Den Bos, Univerza Radboud Nijmegen, Nizozemska
Patrick AnselmeUniverza v Liègu, Belgija
Avtorske pravice © 2014 Zack, Featherstone, Mathewson in Fletcher. To je članek z odprtim dostopom, ki je razdeljen pod pogoji iz. \ T Licenca za priznanje Creative Commons (CC BY). Uporaba, distribucija ali reprodukcija v drugih forumih je dovoljena pod pogojem, da so avtor (ji) ali dajalec licence priznani in da je navedena izvirna objava v tej reviji v skladu s sprejeto akademsko prakso. Uporaba, distribucija ali reprodukcija ni dovoljena, kar ni v skladu s temi pogoji.
* Korespondenca: Martin Zack, Kognitivni psihofarmakološki laboratorij, Oddelek za nevroznanost, Center za odvisnost in duševno zdravje, 33 Russell Street, Toronto, ON M5S 2S1, Kanada e-pošta: [e-pošta zaščitena]