Neurosci Biobehav Rev. 2011 Januar; 35 (3): 939-55. doi: 10.1016 / j.neubiorev.2010.10.014. Epub 2010 Nov 2.
- 1Bernard B. Brodie Oddelek za nevroznanost, Univerza v Cagliariju, Cittadella Universitaria, 09042 Monserrato, Kalifornija, Italija. [e-pošta zaščitena]
IZVLEČEK
Oksitocin je močan induktor erekcije penisa, če ga injiciramo v centralni živčni sistem. Pri samskih podganah je najbolj občutljivo območje možganov za proerektilni učinek oksitocina paraventrikularno jedro hipotalamusa. To jedro in okoliška območja vsebujejo celična telesa vseh oksitocinergičnih nevronov, ki štrlijo v ekstrahipotalamična področja možganov in hrbtenjače. Ta pregled kaže, da oksitocin povzroči erekcijo penisa tudi, če ga injiciramo na nekatere od teh področij (npr. Ventralno tegmentalno območje, ventralni subiculum hipokampusa, posteromedialno kortikalno jedro amigdale in torako-ledveno hrbtenjačo). Študije mikroinjekcije v kombinaciji z intra-cerebralno mikrodializo in dvojnimi imunofluorescentnimi študijami kažejo, da se oksitocin na teh območjih neposredno ali posredno (predvsem prek glutaminske kisline) aktivira mezolimbične dopaminergične nevrone. Dopamin, sproščen v jedru, se nato aktivira v nevronskih poteh, kar vodi do aktivacije incerto-hipotalamičnih dopaminergičnih nevronov v paraventrikularnem jedru. To aktivira ne samo oksitocinergične nevrone, ki štrlijo v hrbtenjačo in posredujejo erekcijo penisa, ampak tudi tiste, ki štrlijo na zgornja ekstrahipotalamična območja, neposredno ali posredno (preko glutaminske kisline) modulirajo aktivnost mezolimbičnih dopaminergičnih nevronov, ki nadzorujejo motivacijo in nagrado. Skupaj lahko te nevronske poti tvorijo kompleksen hipotetični sklop, ki igra vlogo ne le v fazi porabe spolne aktivnosti (erektilna funkcija in kopulacija), temveč tudi pri motivacijskih in nagrajujočih vidikih anticipirajoče faze spolnega vedenja.
1.Uvod
Erekcija penisa je moški spolni odziv, ki ima ključno vlogo pri razmnoževanju sesalcev, vključno s človekom, in ga lahko opazimo tudi v kontekstih, ki se razlikujejo od tistih, ki so strogo povezani z reprodukcijo. Glede na kontekst, v katerem pride do erekcije penisa, pri njegovi regulaciji sodelujejo različni centralni in periferni nevronski in / ali humoralni mehanizmi (glej Meisel in Sachs, 1994; Argiolas in Melis, 1995, 2004, 2005; Sachs, 2000, 2007; McKenna, 2000; Giuliano in Rampin, 2000, 2004; Andersson, 2001; Melis in Argiolas, 1995a, 2003; Hull in sod., 2002). Med osrednjimi nevrotransmiterji in nevropeptidi, ki nadzorujejo erekcijo penisa, so najbolj znani dopamin, serotonin, ekscitatorne aminokisline, dušikov oksid, adrenokortikotropin, oksitocin in opioidni peptidi. Olajšajo ali zavirajo erekcijo penisa z delovanjem na več možganskih področjih, tj. medialno preoptično območje, paraventrikularno jedro hipotalamusa, ventralno tegmentalno območje, hipokampus, amigdala, posteljno jedro stria terminalis, jedro accumbens, podolgata medule in hrbtenjača (Tabela 1) (glej Meisel in Sachs, 1994; Witt in Insel, 1994; Stancampiano idr., 1994; Argiolas in Melis, 1995, 2005; Argiolas, 1999; Bancila idr., 2002; Giuliano in Rampin, 2000, McKenna, 2000; Andersson, 2001; Hull in sod., 2002; Coolen in sod., 2004).
Oksitocin, nevrohipofizni peptid, ki je dobro znan po hormonski vlogi pri dojenju in porodu, je prisoten pri ženskah in samcih, ne le v nevronih s celičnimi telesi, ki se nahajajo v paraventrikularnem in supraoptičnem jedru hipotalamusa, ki štrli v nevrohipofizo, ampak tudi v nevronih štrlečih od paraventrikularnega jedra in okoliških struktur do ekstrahipotalamičnih možganskih področij (tj. septuma, ventralnega tegmentalnega območja, hipokampusa, amigdale, podolgate medule in hrbtenjače). Menijo, da so ti nevroni vključeni v številne osrednje funkcije, kot so spomin, učenje, pripadnost in družbeno-spolno vedenje, vključno z erekcijo penisa in kopulacijskim vedenjem (glej Buijs, 1978; Sofroniew, 1983; Argiolas in Gessa, 1991; Pedersen idr. ., 1992; Carter, 1992; Wagner in Clemens, 1993; Ivell in Russel, 1995; Carter idr., 1997; Tang idr., 1998; Veronneau-Longueville idr., 1999). Dejansko oksitocin olajša erektilno funkcijo in moško spolno vedenje pri miših, podganah, zajcih in opicah (glej Argiolas in Gessa, 1991; Carter, 1992; Pedersen idr., 1992; Argiolas in Melis, 1995, 2004; Argiolas, 1999). To se lahko pojavi tudi pri ljudeh, saj se oksitocin v plazmi poveča s spolnimi dražljaji, zlasti pri ejakulaciji (Carmichael in sod., 1987; Murphy in sod., 1987) ter z manipulacijo dojk in spolovil, ki se običajno pojavljajo med spolnim seksa (Tindall, 1974).
Olajševalni učinek oksitocina na moško spolno vedenje se je najprej pokazal s sposobnostjo intravenskega oksitocina, da zmanjša latenco do prve ejakulacije in zavira spolno izčrpanost samcev zajcev v paru z receptivnimi samicami (Melin in Kihlstrom, 1963). Vendar so bili spolni učinki oksitocina dokončno priznani šele v osemdesetih letih. Nato je bilo ugotovljeno, da je oksitocin, ki ga dajemo centralno v količinah nanograma, lahko sprožil erekcijo penisa (Argiolas et al., 1985, 1986) in izboljšal kopulacijsko vedenje (Arletti idr., 1985) pri samcih podgan in povečal lordozo pri samicah podgan (Arletti in Bertolini, 1985; Caldwell et al., 1986), očitno z delovanjem na maternične oksitocinergične receptorje (glej Argiolas in Melis, 1995, 2004; Argiolas, 1999; Melis in Argiolas, 2003; in reference v njih). Oksitocin izboljšuje spolno vedenje ne samo pri spolno močnih podganah (Arletti et al., 1985), temveč tudi pri starih samcih podgan (Arletti et al., 1990) in pri prevladujočih, ne pa tudi v podrejenih opicah moških veveric (Winslow in Insel , 1991).
Proerektilni učinek oksitocina je odvisen od testosterona, saj ga odpravi s hipofizektomijo in kastracijo ter obnovi z dopolnjevanjem testosterona ali njegovih presnovkov, estradiola in 5_-dihidro-testosterona, danih skupaj (Melis in sod., 1994a). Najbolj občutljivo možgansko območje za indukcijo erekcije penisa z oksitocinom je paraventrikularno jedro hipotalamusa (Melis et al., 1986), iz katerega izvirajo vse ekstrahipotalamične oksitocinergične projekcije (glejte zgoraj). Tu je bilo ugotovljeno, da oksitocin lahko povzroči erekcijo penisa (in zehanje), če ga injiciramo v odmerkih, ki so nižji od 3 pmol (glejte poglavje 2.1 spodaj). Erekcija penisa, ki jo povzroča oksitocin, se vnese tudi dvostransko v CA1 polje hipokampusa, ne pa v dorzalni subkuulum (glej oddelek 2.3 spodaj), bočni septum, kaudatno jedro, medialno preoptično območje, ventromedialno jedro hipotalamusa in supraoptično jedro (Melis in sod., 1986). Številne raziskave kažejo na mehanizem, s katerim oksitocin deluje v paraventrikularnem jedru, da sproži ta spolni odziv, da oksitocin aktivira lastne nevrone. V skladu s to hipotezo spolna interakcija poveča FOS, genski produkt neposrednega zgodnjega genskega c-fos v paraventrikularnih oksitocinergičnih nevronih, ki štrlijo v hrbtenjačo, ki sodelujejo pri nadzoru erekcije penisa (glejte Witt in Insel, 1994 in reference v njem) in spolna nemoč (npr. nezmožnost odraslega moškega, da se kopulira z ovariektomiziranim estrogenom-progesteronenim receptivnim samcem), je bila povezana pri samcu podgane z nizko stopnjo mRNA oksitocina v paraventrikularnem jedru hipotalamusa (Arletti et al., 1997).
Ali oksitocin vpliva na anticipirajočo fazo ali fazo porabe spolnega vedenja, za zdaj ni jasno. Ker oksitocin sproži erekcijo penisa in je glavni učinek oksitocina na kopulacijsko vedenje zmanjšanje intervala po ejakulaciji pri samcih podgan (Arletti et al., 1985), je smiselno domnevati, da peptid izboljšuje spolno sposobnost. Ker pa oksitocin povečuje tudi družbeno-spolno interakcijo (glejte Pedersen in sod., 1992; Carter in sod., 1997; Ivell in Russel, 1995), antagonisti receptorjev oksitocina preprečujejo nekontaktno erekcijo (Melis et al., 1999a), ki veljajo za indeks spolnega vzburjenja (glej Sachs, 1997, 2000, 2007; Melis et al., 1998, 1999b in njihove reference), ni mogoče izključiti možne vloge oksitocina pri spolnem vzburjenju in spolne motivacije.
Ta pregled povzema objavljene in neobjavljene rezultate zadnjih študij, ki kažejo, da oksitocin povzroči erekcijo penisa ne le, če ga injiciramo v paraventrikularno jedro hipotalamusa, temveč tudi v druga zunaj hipotalamična področja možganov, kot je ventralno tegmentalno območje (Melis et al., 2007, 2009a; Succu in sod., 2008), tventralni subiculum hipokampusa in posteriorno jedro amigdale (Melis et al., 2009b, 2010), ki so pomembne sestavine limbičnega sistema in so mišljene, da igrajo ključno vlogo v procesih motivacije in nagrajevanja. Te študije razkrivajo, da oksitocin sodeluje v nevronskih krogotokih, ki vključujejo druge nevrotransmiterje, kot sta dopamin in glutaminska kislina, in druga možganska območja, razen paraventrikularnega jedra, npr. Ventralno tegmentalno območje, okroglo jedro, hipokampus in območja, ki jih še ni. identificirali. Ta vezja verjetno vplivajo na interakcijo med mezolimbičnim in incerto-hipotalamičnim dopaminergičnim sistemom in igrajo vlogo ne le v fazi porabe moškega spolnega vedenja (npr. Erekcija in kopulacija penisa), temveč tudi pri spolni motivaciji in spolnem vzburjenje, s čimer je zagotovljena nevronska podlaga za razlago motivacijskih in koristnih lastnosti spolne aktivnosti.
2. Oksitocin vpliva na erekcijo penisa, saj deluje na različna področja možganov
2.1. Paraventrikularno jedro hipotalamusa
Kot smo spomnili zgoraj, je bilo kmalu opredeljeno paraventrikularno jedro hipotalamusa kot možgansko območje, ki je najbolj občutljivo za proerektilni učinek oksitocina. Ko so enostransko injicirali v to jedro, je bilo ugotovljeno, da je oksitocin aktiven v odmerkih, ki so nižji od 3 ng (3 pmol) (Melis et al., 1986). Študije razmerja med strukturo in aktivnostjo so pokazale, da je nastajanje oksitocina povzročilo erekcijo penisa, ki je bila posredovana z receptorji materinotipa oksitocinom, skupaj s prilivom Ca2 + v celična telesa oksitocinergičnih nevronov, ki so projecirali na ekstrapotirne možganske predele in na aktiviranje sinteze dušikovega oksida. Dušikov oksid pa deluje kot znotrajcelični glasnik z še neznanim mehanizmom (ne vključuje gvanilat ciklaze), kar povzroči aktiviranje oksitocinergičnih nevronov, ki štrlijo na hrbtenjačo in ekstrahipotalamična območja možganov, kar povzroči erekcijo penisa (slika 1) (glej spodaj ter Argiolas in Melis, 1995, 2004, 2005 in njihove reference). Sposobnost oksitocina za aktiviranje lastnih nevronov
Slika 1. (MISANJE) Shematski prikaz oksitocinergičnih nevronov, ki izvirajo v paraventrikularnem jedru hipotalamusa in štrlijo v ekstrahipotalamična možganska območja, kot so hrbtenjača, VTA, hipokampus, amigdala itd. Aktiviranje teh nevronov s dopamin, ekscitatorne aminokisline, oksitocin, heksarelinski analogni peptidi in peptidi, ki izhajajo iz VGF, vodijo do erekcije penisa, ki jo je mogoče zmanjšati in / ali odpraviti s stimulacijo GABAergičnih, opioidnih in kanabinoidnih receptorjev CB1. Aktivacija oksitocinergičnih nevronov je sekundarna od aktivacije dušikovega oksid-sintaze, ki je prisotna v teh nevronih. Dejansko endogeni dušikov oksid, ki nastane s stimulacijo dopaminskih, ekscitatornih receptorjev za aminokislino ali oksitocin ali eksogenega dušikovega oksida, saj dobljeni iz dajalcev dušikovega oksida, dani neposredno v paraventrikularno jedro, aktivirajo oksitocinergične nevrone s še neidentificiranim mehanizmom. stimulacija gvanilat ciklaze. To posledično povzroča sproščanje oksitocina v hrbtenjači in na možganov, ki niso hipotalamični. Nekaj podrobnosti o mehanizmih, s katerimi oksitocin povzroči erekcijo penisa, ko se sprosti na teh območjih, npr. VTA, ventralni subiculum in amigdala so opisani v ustreznih oddelkih možganskega območja. Tu oksitocin deluje na lastne receptorje in poveča proizvodnjo NO, kar vodi v erekcijo penisa, kot jo najdemo v PVN. Vendar v odstopanju od PVN v kaudalni VTA NO aktivira gvanilat ciklazo. To povzroči povečanje koncentracije cGMP, kar vodi do aktiviranja mezolimbičnih dopaminergičnih nevronov in do erekcije penisa. V VS NO aktivira glutamatergične nevrone, ki štrlijo na območja zunaj hipokampala, vključno z VTA. Glutaminska kislina v VTA aktivira mezolimbične dopaminergične nevrone, kot jih najdemo z oksitocinom. Mehanizmi, podobni zgoraj opisanim, bodo verjetno delovali tudi, ko se erekcija penisa pojavi v fizioloških okoliščinah, in sicer, ko so samci podgane nameščeni v primeru nedostopne dovzetne samice (npr. Brezkontaktne erekcije) ali med kopulacijo.
v paraventrikularnem jedru so podprle študije, ki kažejo, da: (1) oksitocinski receptorji so prisotni v tem hipotalamičnem jedru (Freund-Mercier in sod., 1987; Freund-Mercier in Stoeckel, 1995); (2) oksitocin olajša njegovo sproščanje in vitro in in vivo (Freund-Mercier in Richard, 1981, 1984; Moos in sod., 1984); in (3) oksitocin vzbudi lastne nevrone z delovanjem v paraventrikularnem jedru (Yamashita in sod., 1987). Poleg tega so bile ugotovljene tudi oksitocinergične sinapse, ki vplivajo na celična telesa magnoceličnih oksitocinergičnih nevronov v paraventrikularnem in supraoptičnem jedru hipotalamusa (Teodoza, 1985). Končno uničenje osrednjih oksitocinergičnih nevronov z elektrolitičnimi ali kemičnimi ekscitotoksičnimi lezijami paraventrikularnega jedra, ki v celoti izčrpa vsebnost oksitocina v osrednjem živčnem sistemu in hrbtenjači, odpravi ne le proerektilni učinek oksitocina, ampak tudi poslabša povzročene z zdravili erekcijo penisa in brezkontaktne erekcije (glej spodaj in Argiolas et al., 1987a, b; Liu in sod., 1997 in reference v njih). Rezultati, podobni tistim, ki so jih našli pri lezijah paraventrikularnega jedra, najdemo z močnimi in selektivnimi antagonisti receptorjev oksitocina. Dejansko so te spojine, vbrizgane v paraventrikularno jedro v nanogramskih količinah, preprečile erekcijo penisa, ki je bila povzročena z oksitocinom, medtem ko pri dajanju v lateralne ventrikle preprečijo ne samo erekcije penisa, ki jo povzroči sam oksitocin, temveč tudi erekcijo penisa, povzročeno z zdravili (glejte poglavje 3 spodaj ter Argiolas in Melis, 1995, 2004, 2005 in njihove reference) in brezkontaktne erekcije (Melis et al., 1999a), poleg tega pa so bili izjemno učinkoviti pri zmanjšanju kopulacijskega vedenja spolno močnih podgan (Argiolas et al., 1988). Poleg tega spolna interakcija poveča FOS, genski produkt neposrednega zgodnjega genskega c-fos v paraventrikularnih oksitocinergičnih nevronih, ki štrlijo v hrbtenjačo in sodelujejo pri nadzoru erekcije penisa (glejte Witt in Insel, 1994 in reference nanje). Končno spolna nemoč (npr. nezmožnost odraslega moškega podgane, da se kopulira z dojemljivo samico estrogena, progesterone) hkot so bili povezani tudi pri samcu podgane z nizko koncentracijo mRNA oksitocina in sinteze dušikovega oksida v paraventrikularnem jedru hipotalamusa (Benelli in sod., 1995; Arletti in sod., 1997) (za obširen pregled teh študij glej Argiolas, 1999; Argiolas in Melis, 2004, 2005).
2.2. Ventralno tegmentalno območje
Ventralno tegmentalno območje je bilo odkrito šele pred kratkim kot mesto možganov, kjer oksitocin sproži erekcijo penisa. To območje vsebuje oksitocinergične živčne končiče, ki izvirajo iz paraventrikularnega jedra in oksitocinske receptorje (Freund-Mercier et al., 1987; Vaccari in sod., 1998). Natančneje, našli so oksitocin, ki je sposoben induciranje erekcije penisa, če ga enostransko injiciramo v kaudal, ne pa v testmentno območje rostralnega ventrala na način, ki je odvisen od odmerka (Melis in sod., 2007). Aktivni odmerki so bili višji od tistih, ki so bili potrebni pri injiciranju v paraventrikularno jedro, in podobni tistim, ki povzročajo erekcijo penisa, ko so bili injicirani v ventralni subiculum hipokampusa ali v posteromedialno kortikalno jedro amigdale (glejte spodaj). Očitno je proerektilni učinek posredovan z aktivacijo mezolimbičnih dopaminergičnih nevronov, ki štrlijo v lupino jedra akkule, kar posledično aktivira še neznane nevronske poti, ki štrlijo v incerto-hipotalamične dopaminergične nevrone, ki vplivajo na paraventrikularno oksitocinergično nevrono, ki posreduje erekcijo penisa (Melis in sod., 2007, 2009a).
Glede mehanizma, s katerim oksitocin aktivira dopaminergično nevrotransmisijo v ventralnem tegmentalnem območju, razpoložljivi podatki kažejo, da oksitocin stimulira oksitocinergične receptorje, ki se nahajajo v celičnih telesih mezolimbičnih dopaminergičnih nevronov. To poveča priliv Ca2 + v celična telesa dopaminergičnih nevronov in s tem aktivira dušikov oksid-sintazo (Succu et al., 2008). Za razliko od paraventrikularnega jedra (glej poglavje 3 spodaj) dušikov oksid nato aktivira gvanilat ciklazo in tako poveča koncentracijo cikličnega GMP. V skladu s tem mehanizmom se bodisi d (CH2) 5Tyr (Me) 2-Orn8-vazotocin, močan antagonist oksitocina, bodisi S-metil-tio-l-citrulin, močan zaviralec sinteze nevronskega dušikovega oksida. kaudalno ventralno tegmentalno območje pred oksitocinom, odpravljeno erekcijo penisa in povečanje koncentracije zunajceličnega dopamina v lupini jedrnih jezgrov, ki jih povzroča oksitocin. Poleg tega 8-bromo-ciklični GMP, aktivni ciklični analogni GMP, odporen na fosfodiesterazo, povzroči erekcijo penisa, kadar ga injiciramo v tegmentalno območje kaudalnega ventralnega tkiva in poveča koncentracijo zunajceličnega dopamina v lupini jedra jedra, kot je ugotovljeno z oksitocinom, injiciranim v kaudalno ventralno tegmentalno območje (Succu in sod., 2008; Melis in sod., 2009a) (glej tudi sliko 2).
Vedno v skladu s tem mehanizmom haloperidol, močan antagonist receptorja D2 dopamina, ki ga injiciramo v lupino nukleusnega tkiva, zmanjša erekcijo penisa, ki jo inducira oksitocin, vbrizgan v ventralno tegmentalno območje (Melis et al., 2007). Zgornji mehanizem podpirajo tudi dvojne imuno-fluorescentne študije, ki kažejo, da oksitocinska vlakna posegajo po celičnih telesih dopaminergičnih nevronov v kaudalnem ventralnem tegmentacijskem območju, ki so bili predhodno označeni z retrogradnim sledilcem Fluorogold, ki ga vbrizgajo v lupino nukleusnega jedra ( Melis in sod., 2007; Succu in sod., 2008). Aktivacija teh dopaminergičnih nevronov in dopaminskih receptorjev v akumulaciji jedra vodi v aktiviranje nevronskih poti, ki jih je treba še določiti, kar spodbudi incertno-hipotalamične dopaminergične nevrone, da sprostijo dopamin v paraventrikularnem jedru, s tem pa aktivirajo oksitocinergične nevrone. hrbtenjača in posredovana erekcija penisa (glej zgoraj in Melis in sod., 2007; Succu in sod., 2007, 2008). Dejansko je oksitocin vbrizgal v kaudalno ventralno tegmentalno območje v odmerku, ki je povzročil erekcijo penisa, povečal koncentracijo zunajceličnega dopamina v dializatu, pridobljenem ne le iz jedra jedra, temveč tudi iz paraventrikularnega jedra (Succu et al., 2007).
2.3. Hipokampus
Polje hipokampusa CA1 je bilo drugo možgansko območje, bogato z oksitocinergičnimi vlakni in receptorje, ugotovljene s prejšnjimi študijami, v katerih je injekcija oksitocina povzročila erekcijo penisa (glejte Bujis, 1978; Sofroniew, 1983). Vendar je bilo tu, v nasprotju s paraventrikularnim jedrom, tu ugotovljeno, da lahko oksitocin povzroči erekcijo penisa le, če ga injiciramo dvostransko in v višjih odmerkih, kot tisti, ki so bili aktivni v paraventrikularnem jedru (Melis in sod., 1986; Chen et al., 1992). V teh prejšnjih raziskavah so bile injekcije oksitocina v subiculum neaktivne. Vendar pa nedavne in previdnejše raziskave mikroinjekcije vodijo k identifikaciji a regiji ventralnega subiculuma pri čemer je injekcija oksitocina lahko povzročila erekcijo penisa na način, odvisen od odmerka (Melis et al., 2009b). Proerektilni učinek oksitocina, ki ga vbrizgajo v to možgansko območje, so opazili pri odmerkih, podobnih tistim, ki so bili po enostranskem injiciranju v ventralnem tegmentalnem območju (Melis in sod., 2007) aktivni v jedru paraventrikularne celice. Očitno oksitocin, ki se vbrizga v ventralni subiculum, povzroči erekcijo penisa z aktiviranjem oksitocinergičnih receptorjev v nevronih, ki vsebujejo dušikov oksid-sintazo, kar povzroči povečanje proizvodnje dušikovega oksida. Dušikov oksid pa deluje kot medcelični glasnik, aktivira nevrotransmisijo glutaminske kisline, kar vodi v erekcijo penisa, morda z nevronskimi (glutamatergičnimi) eferentnimi projekcijami iz ventralnega subiculuma na možganska območja zunaj hipokampusa, ki modulirajo aktivnost mezolimbičnih dopaminergičnih nevronov (npr. ventralno tegmentalno območje, prefrontalna skorja, paraventrikularno jedro) (glejte spodaj in Melis, 2007, 2009b; Succu idr., 2008).
Ta mehanizem delovanja podpirajo poskusi intra-cerebralne mikrodijalize, ki kažejo, da oksitocin, ki se vbrizga v ventralni subkulum v odmerkih, ki inducirajo erekcijo penisa, poveča proizvodnjo dušikovega oksida in koncentracijo zunajcelične glutaminske kisline v dializi iz ventralnega subiculuma (Melis et al. , 2010) in zunajceličnega dopamina v jedru jedra (Melis et al., 2007). Te odzive je antagoniziral ne le antagonist oksitocinskih receptorjev d (CH2) 5Tyr (Me) 2-Orn8-vazotocin, temveč tudi zaviralec nevronskega dušikovega oksida-sintaze S-metil-tio-lcitrulin in sredstvo za odstranjevanje dušikovega oksida hemoglob v ventralni subkulum nekaj minut pred oksitocinom (Melis in sod., 2010).
Poleg tega v skladu s tem mehanizmom delovanja aktivacija glutamatergičnega nevrotransmisije z NMDA, ki se vbrizga v ventralni subiculum, povzroči erekcijo penisa (Melis in sod., 2010). Fenotip eferentnih projekcij iz ventralnega subiculuma, ki povzročajo aktivacijo mezolimbičnih dopaminergičnih nevronov in povečanje zunajceličnega dopamina v jedru jedra, za zdaj ni znan. Ker pa pride do erekcije penisa, ki jo inducira oksitocin, vbrizgan v ventralni subiculum, sočasno do povečanja zunajcelične glutaminske kisline v dializatu iz ventralnega tegmentalnega območja, ne pa iz jedra, ki se nabira, in ga antagonizira (+) MK-801, močan nekonkurenčni antagonist ekscitacijskih receptorjev aminokislin podtipa NMDA (Woodruff et al., 1987), injiciran v ventralno tegmentalno območje, vendar ne v jedro jedra (glej slike 2 in Melis idr., 2009b) , verjetno te projekcije vodijo do aktiviranja glutamatergične nevrotransmisije v ventralnem tegmentalnem območju, kar posledično aktivira mezolimbične dopaminergične nevrone, ki štrlijo v akumulacijsko jedro. Ali se povečana koncentracija glutaminske kisline, ki jo najdemo v ventralnem tegmentalnem območju po vbrizganju oksitocina v ventralni subiculum, sprosti iz nevronov, ki izvirajo iz subiculuma ali iz drugih možganskih področij (npr. Predfrontalne skorje), za zdaj ni znano. Kljub temu to povzroči aktivacijo mezolimbičnih dopaminergičnih nevronov in povečano sproščanje dopamina v jedrih. Tu aktiviranje dopaminskih receptorjev vodi do aktiviranja incerto-hipotalamičnih dopaminergičnih nevronov, sprošča dopamin v paraventrikularnem jedru in s tem aktivira oksitocinergične nevrone, ki štrlijo v hrbtenjačo in posredujejo erekcijo penisa (glej zgoraj in Melis in sod., 2007, 2009a; Succu in sod., 2008).
2.4. Amigdala
Amigdala je drugo možgansko območje, bogato z oksitocinskimi vlakni in receptorji (glej Freund-Mercier in sod., 1987; Vaccari in sod., 1998; Uhl-Bronner idr., 2005). Mislimo, da je tu oksitocin vključen v različne funkcije, od anksiolize, socialnega spomina in kognicije, socialno okrepljenega učenja, čustvene empatije, čustvene obdelave obraza in strahu pri ljudeh do erektilne funkcije in spolnega vedenja (glej Kondo in sod., 1998; Dominguez idr. al., 2001; Ebner et al., 2005; Huber in sod., 2005; Domes in sod., 2007; Petrovic et al., 2008; Lee in sod., 2009; Donaldson in Young, 2009; Hurlemann idr. , 2010). Vendar pa je bila sposobnost oksitocina, da inducira erekcijo penisa pri samcih podgan pri indukciji v posteromedialnem kortikalnem jedru amigdale, odkrila šele pred kratkim (Melis in sod., 2009b). Ta odziv se je pojavil sočasno s povečanjem koncentracije zunajceličnega dopamina v dializatu, dobljenem iz lupine jedrnih jezgrov, kar smo ugotovili po vbrizganju oksitocina v ventralni subkulum (Melis in sod., 2009b). Mehanizem, s katerim se oksitocin injicira v posteromedialno kortikalno jedro amigdale, sproži erekcijo penisa, trenutno ni znan. Razpoložljivi podatki kažejo, da tako erekcija penisa kot povečanje koncentracije zunajceličnega dopamina v dializatu, dobljenem iz nukleusnih akumulacij, posreduje aktivacija oksitocinergičnih receptorjev, saj sta oba odgovora odpravila antagonist oksitocinskih receptorjev d (CH2) 5Tyr ( Me) 2-Orn8-vazotocin injiciran v jedro amigdale nekaj minut pred oksitocinom (Melis et al., 2009b).
Ne glede na to, kakšen mehanizem aktivira oksitocin v posteromedialnem kortikalnem jedru amigdale, se spolni odziv, ki ga povzroči peptid, odpravi z blokado vseh dopaminergičnih receptorjev s cis-flupentiksolom, ki se vbrizga v lupino jedra jedra in z blokado NMDA receptorjev z ( +) MK-801 se injicira v ventralno tegmentalno območje, vendar ne v jedro jedra, kot je bilo ugotovljeno za erekcijo penisa, ki jo inducira oksitocin, vbrizgan v ventralni subkulum (Melis et al., 2009b). To kaže, da oksitocin, injiciran v posteromedialno jedro amigdale, aktivira nevrotransmisijo glutaminske kisline v ventralnem tegmentalnem območju. To posledično povzroči aktiviranje mezolimbičnih dopaminergičnih nevronov, kar vodi v erekcijo penisa. Glede na študije, ki prikazujejo nevronske poti, ki povezujejo to jedro amigdale z ventralnim subiculumom (Canteras et al., 1995; French in Totterdell, 2003), te ugotovitve povečujejo možnost, da med tema dvema možganskima območjema obstaja interakcija, čeprav neposredna opisani so poti od amigdale do jedrskega jedra ali do ventralnega tegmentalnega območja (Kelley in Domesick, 1982; Witter, 2006).
2.5. Hrbtenjača
Hrbtenjača je še eno področje centralnega živčnega sistema, ki vsebuje oksitocinergična vlakna in receptorje (Freund-Mercier et al., 1987; Uhl-Bronner et al., 2005), kjer oksitocin inducira erekcijo penisa (Tang in sod., 1998 ; Veronneau-Longueville in sod., 1999; Giuliano in Rampin, 2000; Giuliano in sod., 2001). Kot je navedeno zgoraj, ta oksitocinergična vlakna izvirajo iz paraventrikularnega jedra hipotalamusa in prispevajo k padajočim poti, ki nadzorujejo spinalne avtonomne nevrone, ki posredujejo erekcijo penisa. Dejansko ta vlakna naredijo sinaptične stike v hrbtnem rogu preganglionskih simpatičnih in parasimpatičnih celičnih stolpcev v torako-ledvenem in ledveno-križnem traktu s hrbteničnimi nevroni, ki preplavijo kavernozno korpus penisa (Marson in McKenna, 1996; Giuliano in Rampin, 2000; Xulium et al. , 2001). Ti sinaptični stiki so bili dokazani z označevanjem hrbtenjačnih nevronov, ki izvirajo iz penisa in dosežejo hrbtenjačo s posebnimi retrogradnimi sledilci, ki jih injicirajo v kavernozno korpus, v kombinaciji z dvojno imuno-fluorescenco in konfokalnimi študijami laserske mikroskopije (Tang et al., 1998; Veronneau-Longueville et al., 1999). V skladu s temi študijami je pri anesteziranih samcih podganah intratekalno vbrizgavanje kumulativnih odmerkov oksitocina na ledveno-križnični stopnji, vendar ne na torako-ledvenem nivoju, povzročil intrakavernozni tlak, ki narašča od odmerka. Ti učinki so bili odpravljeni z blokado oksitocinergičnega receptorja z d (CH2) 5Tyr (Me) 2-Orn8-vazotocin in z odsekom medeničnih živcev (Giuliano in Rampin, 2000; Giuliano idr., 2001). Ti rezultati kažejo, da oksitocin, ki deluje na ledveno-križnični hrbtenjači, poveča intrakavernozni tlak in kažejo, da je oksitocin, ki se sprosti med fiziološko aktivacijo paraventrikularnega jedra, močan aktivator spinalnih proerektilnih nevronov, ki štrlijo v kavernozno korpus. Zanimivo je, da ti proerektilni hrbtenični nevroni, na katere oksitocin deluje, da izvajajo proerektilni učinek, prejemajo tudi sinaptične stike iz serotoninergičnih nevronov, ki izvirajo iz jedra paragigantocellularis retikularne tvorbe podolgata medule (Marson in McKenna, 1992; Tang et al. ., 1998). Uničenje teh serotoninergičnih nevronov olajša ejakulacijo in refleks penisa pri samcih podgan (Marson in McKenna, 1992; Yells et al., 1992). Ker zdravila, ki stimulirajo 5HT2C receptorje olajša erekcijo penisa pri dajanju intracerebroventricularly, vendar ne v paraventricular jedra, in zdravila, ki da blok 5HT2C receptorji zmanjšujejo tudi dopamin agonistand oksitocin-inducirano erekcijo penisa, medtem antagonisti dopamina ne zmanjšujejo 5HT2C-agonistom inducirane erekcijo penisa ( glej Stancampiano in sod., 1994 in njihove reference), je bilo tudi predlagano, da oksitocin olajša delovanje proerektilnih receptorjev 5HT2C na nivoju ledveno-sakralne hrbtenjače (Stancampiano et al., 1994). Lahko pa oksitocin vpliva na aktivnost serotoninergičnih nevronov hrbtenice, tako da deluje neposredno v jedru paragigantocellularis, od koder ti nevroni izvirajo (glej Stancampiano in sod., 1994).
3. Interakcije med oksitocinom, dopaminom in glutaminsko kislino v centralnem živčnem sistemu in erekcijo penisa
Kot je bilo opozorjeno v oddelku 1, vsi oksitocinergični nevroni, ki so prisotni v centralnem živčnem sistemu, izvirajo iz paraventrikularnega jedra in okoliških struktur. Aktivnost teh nevronov je pod nadzorom različnih nevrotransmiterjev in / ali nevropeptidov. Med najbolj raziskanimi na paraventrikularni ravni so dopamin, glutaminska kislina, gama-amino-maslačna kislina (GABA), dušikov oksid, endokanabinoidi, opioidni peptidi, peptidi, ki sproščajo rastni hormon, peptidi, povezani z VGF, in sam oksitocin. Dopamin, glutaminska kislina, peptidi, ki sproščajo rastni hormon, peptidi, ki izhajajo iz VGF, in oksitocin stimulirajo, npr. Te spojine in njihovi agonisti olajšajo erekcijo penisa, če se injicirajo v paraventrikularno jedro, medtem ko GABA, opioidni peptidi in endokanabinoidi zavirajo, npr. Te spojine ali njihovi agonisti zavirajo erekcijo penisa (glej Meisel in Sachs, 1994; Witt in Insel, 1994; Argiolas in Melis, 1995, 2004, 2005; Giuliano in Rampin, 2000, 2004; McKenna, 2000; Andersson, 2001; Hull idr., 2002).
Več vrstic eksperimentalnih dokazov kaže, da so ti oksitocinergični nevroni in zgoraj navedeni nevrotransmiterji in nevropeptidi vključeni v nadzor nad erektilno funkcijo in spolnim vedenjem v različnih fizioloških okoliščinah. Še več, oksitocin, ki se sprošča v možganih, ki niso hipotalamični v možganih, na primer v ventralnem tegmentalnem območju, hipokampusu in njegovih regijah, amigdali in hrbtenjači, lahko vpliva na aktivnost tistih nevronov, na katere posegajo oksitocinergične sinapse. Trenutno so edini nevroni, pomembni za erekcijo penisa, na katere vstopajo oksitocinergične sinapse, ki jih z gotovostjo identificirajo, celična telesa mezolimbičnih dopaminergičnih nevronov kaudalnega ventralnega tegmentalnega območja, ki štrlijo v lupino jedra jedra (Melis et al., 2007 ; Succu in sod., 2008) in proerektilnih hrbteničnih nevronov, ki štrlijo iz ledveno-križnega trakta v kavernozno korpus (glej Giuliano in Rampin, 2000; Giuliano idr., 2001) (glej tudi poglavji 2.2 in 2.5) . Čeprav so bili v ventrikalnem subkulumu, amigdali in hrbtenjači odkriti tudi oksitocinergični sinapse in receptorji, področja, ki so pomembna za erekcijo penisa (glejte zgoraj), je na teh območjih vrsta nevrotransmiterjev / prisoten v nevronih na kateri oksitocinergični živčni končiči prihajajo, še vedno niso znani.
Ta del pregleda na kratko povzema nedavno literaturo o mehanizmih, ki temeljijo na erektilnem učinku oksitocina, ki se vbrizga v kaudalno ventralno tegmentalno območje, v ventralni subiculum hipokampusa in v hrbtenjačo. Posebna pozornost je na teh območjih posvečena interakciji peptida z dopaminom in glutaminsko kislino, na vlogo te interakcije pa lahko pri osrednjem nadzoru erektilne funkcije. Najprej je podan kratek povzetek učinkov dopamina in glutaminske kisline na oksitocinergične nevrone v paraventrikularnem jedru, ki imajo tudi ključno vlogo pri erektilni funkciji, da bi bralca seznanili z zgodnjim stanjem raziskav na tem področju, saj te študije so bile že obsežno pregledane (glejte Argiolas in Melis, 1995, 2004, 2005; Melis in Argiolas, 2003). Tudi v tem primeru je posebna pozornost namenjena najnovejšim rezultatom, ki kažejo na pomembno vlogo tako povezave dopamin-oksitocin kot povezave glutaminske kisline-oksitocina ne le pri spolni uspešnosti (erekcija in kopulacija penisa), temveč tudi pri spolni uporabi. vzburjenje in spolna motivacija.
3.1. Interakcija dopamin-oksitocin v paraventrikularnem jedru
O sposobnost dopaminskih agonistov, da inducirajo erekcijo penisa z aktiviranjem centralnih oksitocinergičnih nevronov je bilo predlagano, da takoj po odkritju apomorfina povzroči erekcijo penisa, kadar ga injiciramo v paraventrikularno jedro (Melis et al., 1987), ko je antagonist oksitocinskih receptorjev d (CH2) 5Tyr (Me) 2-Orn8-vazotocin injiciran intracerebrovventralno). lahko skoraj popolnoma zmanjša erekcijo penisa, ki jo povzroča samo oksitocin, ki ga daje icv, ampak tudi apomorfin, ki se daje subkutano (Argiolas et al., 1987b). Tem rezultatom so sledili rezultati drugih raziskav, ki so pokazale podobne rezultate, ko je d (CH2) 5Tyr (Me) 2-Orn8-vazotocin prejel icv in apomorfin dal direktno v paraventrikularno jedro (Melis et al., 1989b), lizogibamo se domnevi, da dopaminski agonisti inducirajo erekcijo penisa z aktiviranjem paraventrikularnih oksitocinergičnih nevronov, ki štrlijo v možganske hipotalamične predele in zlasti na hrbtenjačo (glej Argiolas in Melis, 1995, 2004, 2005). V skladu s to hipotezo so pri anesteziranih podganah nedavno ugotovili blokado ledveno-sakralnih oksitocinergičnih receptorjev s strani nepeptidnega antagonista oksitocinskih receptorjev, ki so sposobni odpraviti povečanje, ki ga povzroča apomorfin v intrakavernoznem tlaku, ki ga povzroči agonist dopaminov agomorfin, kar zagotavlja dokaze za paraventrikulo-spinalna oksitocinergična pot, ki sodeluje pri erekciji penisa (Baskerville et al., 2009).
Študije, usmerjene v identifikacija receptorja za dopamin, ki je odgovoren za indukcijo erekcije penisa, je pokazala, da tudi agonisti dopaminskih receptorjev za paraventrikularno jedro inducirajo erekcijo penisa z delovanjem na dopaminske receptorje iz družine D2, kot so ugotovili pri agonistih receptorjev za dopamin (glejte Melis in sod., 1987; Eaton in sod., 1991; Melis in Argiolas, 1995a). V skladu s tem sta apomorfin, močan mešani agonist receptorjev D1 / D2, in kinpirol, močan selektivni agonist receptorjev D2, vendar ne SKF 38393, selektivni agonist D1 receptorjev, injiciran v to hipotalamično jedro, ki bi lahko povzročil nastanek odmerka. in spolni odziv, ki ga povzročajo ti agonisti receptorjev D2, so odpravili antagonisti receptorjev D2, kot sta haloperidol in l-sulfpirid, ne pa SCH 23390, selektivni antagonist D1 receptorjev (Melis et al., 1987). Sposobnost apomorfina, da pri injiciranju v paraventrikularno jedro inducira erekcijo penisa, je bila potrjena tudi s telemetrijskimi raziskavami, ki kažejo, da je agonist dopamina, ki ga dajemo v paraventrikularno jedro, sposoben zvišati intrakavernozni tlak pri budnih podganah, ne da bi pri tem spremenil sistemski krvni tlak (Chen et al. , 1999; Giuliano in Allard, 2001), kot so ugotovili po sistemski injekciji (Bernabè et al., 1999). Tudi te študije potrdili glavno vlogo receptorjev D2, saj je bilo ugotovljeno, da agonisti receptorjev D1 ob injiciranju v paraventrikularno jedro ne morejo zvišati intrakavernoznega tlaka (Chen in sod., 1999).
Nato je bilo na voljo več vrstic eksperimentalnih dokazov, ki kažejo, da so paraventrikularni receptorji D2 nameščeni na celičnih telesih oksitocinergičnih nevronov. Prvič, paraventrikularno jedro vsebuje dopaminergične živčne terminale, ki pripadajo tako imenovanim incertohipotalamičnim dopaminergičnim nevronom. Celična telesa teh nevronov se nahajajo v skupini A13 in A14 of Dahlstrom in Fuxe (1964), široko arborirajo in inervirajo več hipotalamičnih struktur, vključno s paraventrikularnimi oksitocinergičnimi nevroni, ki štrlijo na nevrohipofizo in / ali na ekstrahipotalamična možganska območja (Buijs et al., 1984; Lindvall et al., 1984).
Vključitev teh dopaminergičnih nevronov na paraventrikularni ravni pri nadzoru erekcije in kopulacije penisa je podprta z raziskavami mikrodialize, ki kažejo, da se koncentracije zunajceličnega dopamina in 3,4-dihidroksifeniloctene kisline (DOPAC), glavnega presnovka, povečajo v dializatu, dobljenem iz paraventrikularno jedro spolno močnih podgan samcev, ki kažejo brezkontaktne erekcije, kadar so jih dali v nedostopni samici z estrogenom, ki je bila napolnjena z estrogenom + progesteronom (Melis et al., 2003).
Povečanje koncentracij dopamina in DOPAC je bilo še večje, ko je bila dovoljena kopulacija z receptivno samico (Melis in sod., 2003), kot jo najdemo v območju medialnega predoptika (Hull in sod., 1995) in v jedru jedra (Pfaus in Everitt, 1995). Drugič, več raziskav kaže, da erekcijo penisa, ki jo povzroči stimulacija paraventrikularnih D2 receptorjev, posreduje oksitocin, ki se sprošča na teh območjih. V skladu s tem je bilo ugotovljeno, da je apomorfin v odmerkih, ki povzročajo erekcijo penisa, lahko povečal koncentracijo oksitocina, ne le v plazmi podgan in opic (Melis in sod., 1989a; Cameron in sod., 1992), temveč tudi v možgah, ki niso hipotalamični območja, kot je hipokampus (Melis et al., 1990). V skladu s temi rezultati je bil apomorfin, ki ga injicirajo v paraventrikularno jedro v odmerku, ki inducira erekcijo penisa, pred kratkim dokazano, da lahko poveča tudi koncentracijo zunajceličnega dopamina v jedru, kar vpliva na antagonist oksitocinskega receptorja d (CH2) 5Tyr (Me) 2-Orn8-vazotocin se injicira v ventralno tegmentalno območje (Succu in sod., 2007; Melis in sod., 2009a) (glej tudi poglavje 4). Tretjič, dvostranske elektrolitske lezije paraventrikularnega jedra, ki skoraj v celoti izločijo oksitocin iz ekstrahipotalamičnih možganskih področij (Hawthorn in sod., 1985), odpravijo erekcija penisa, povzročene z apomorfinom (Argiolas et al., 1987a), in selektivne antagoniste receptorjev oksitocina. dani v stranske preddrebe, vendar ne v paraventrikularno jedro, zmanjšajo erecijo penisa, ki je odvisna od odmerka, z vzporednostjo kot pri teh spojinah, ko blokirajo receptorje za oksitocin (Melis et al., 1989b). Antagonisti receptorjev za oksitocin so tudi izjemno močni pri zmanjšanju olajšanja moškega spolnega vedenja, ki ga povzroča ne le oksitocin, temveč tudi apomorfin (Argiolas et al., 1988, 1989).
Številni eksperimentalni podatki o mehanizmu, s katerim D2 receptorji, ki jih aktivirajo dopamin ali agonisti dopaminskih receptorjev, povečajo aktivnost oksitocinergičnih nevronov in tako sproščajo oksitocin v ekstrahipotalamičnih možganskih območjih in v hrbtenjači.podpirati hipotezo, da stimulacija receptorjev D2 poveča koncentracijo znotrajceličnih ionov Ca2 + v celičnih telesih oksitocinergičnih nevronov, kar vodi do aktiviranja dušikovega oksid-sintaze, encim, odvisen od Ca2 + -kalmodulina, ki je prisoten v teh celičnih telesih (Vincent in Kimura, 1992; Torres in sod., 1993; Sanchez et al., 1994; Sato-Suzuki idr., 1998). Povečana proizvodnja dušikovega oksida povzroči aktiviranje oksitocinergičnih nevronov. V skladu s tem so (1) apomorfinininizirano erekcijo penisa preprečili organski zaviralci kalcijevih kanalov in by-konotoksin GVIA, močan in selektiven blokator napetostno odvisnih kanalov Ca2 + N-tipa (McCleskey et al., 1987), dani v paraventrikularno jedro (glej Argiolas et al., 1990 in reference v njem); (2) apnoorfin, ki je povzročil erekcijo penisa, so preprečili zaviralci dušikovega oksida-sintaze, ki so bili dani v paraventrikularno jedro (Melis et al., 1994c); in (3) apomorfin in drugi agonisti receptorjev D2, dani pri odmerkih, ki inducirajo erekcijo penisa, povečajo proizvodnjo dušikovega oksida v paraventrikularnem dializatu, dobljenem z intra-cerebralno mikrodializo, kar je povečalo zaviralce paraventrikularnega dušikovega oksid-sintaze, dane v odmerkih, ki zmanjšujejo D2 receptor agonistinducirana erekcija penisa (Melis et al., 1996). Mehanizem, s katerim dušikov oksid aktivira paraventrikularne oksitocinergične nevrone, še vedno ni znan, čeprav razpoložljivi podatki kažejo, da dušikov oksid deluje kot znotrajcelični sel in da gvanilat ciklaza ne sodeluje. Dejansko je bilo ugotovljeno, da aktivni fosfodiesteraserezistentni analog cikličnega GMP, 8-bromo-ciklični GMP ne more inducirati erekcije penisa, ko ga damo v paraventrikularno jedro (slika 2) (glej Melis in Argiolas, 1995b in reference v njem).
Zgornja razlaga se pogosto šteje za neprepričljivo, predvsem zato, ker je stimulacija dopaminskih receptorjev D2 običajno povezana z inhibicijo in ne z vzbujanjem celičnih teles nevronov, ki vsebujejo te receptorje, z različnimi mehanizmi, povezanimi z G proteini (glej Sokoloff in Schwartz, 1995). Vendar pa je možno razlago tega neskladja, ki je v skladu z neposredno stimulacijo paraventrikularnih oksitocinergičnih nevronov z dopaminom, nedavno predlagalo odkritje Dopamin D4, povezan z beljakovinami receptor, ki je član družine receptorjev D2 (D2, D3 in D4), katerega stimulacija povečuje priliv Ca2 + v celične pripravke, ki vsebujejo klonirano različico tega podtipa receptorjev (Moreland in sod., 2004). Še pomembneje je, da je bil selektivni agonist receptorjev D4 (npr. ABT 724) (N-metil-4- (2-cianofenil) piperazinil-3metilbenzamid maleat) ugotovljen, da je sposoben inducirati erekcijo penisa pri samcih podgan, če jih dajemo sistemsko (Brioni et al., 2004). Ta učinek ni bil ugotovljen pri selektivnem agonističnem receptorju D2 receptorja PNU-95666E (R-5,6-dihidro-N, N-dimetil-4Himidazo [4,5,1-i] kinolin-5-amin) (Hsieh et al., 2004), ki prav tako ni mogel povečati priliva Ca2 + v celične pripravke, ki vsebujejo klonirano različico podtipa receptorjev D4 (Brioni in sod., 2004; Moreland in sod., 2004). V skladu z zgornjo hipotezo in ugotovitvami je PD 168,077 (N-metil-4- (2-cianofenil) piperazil-3metilbenzamid maleat), PIP-3EA (2- [4- (2-metoksifenil) piperazin-1-imzozo-1,2ida imzozo-4ida [1997-a] piridin) in drugi selektivni agonisti receptorjev D2006 (Heier in sod., 2009; Melis et al., 4b; Löber idr., 745,870) so lahko sprožili erekcijo penisa, kadar ga injiciramo sistemsko, icv in v paraventrikularno jedro, čeprav manj učinkovito kot apomorfin. Pro-erektilni učinek teh agonistov receptorjev D3 je preprečil L-4 (1- (1- [klorofenil] piperazin-2,3-il) -metil-4H-pirolo [1997-B] piridin trihidroklorid), selektivni DHNXX, selektivni DH, antagonist (Patel in sod., 2005; Melis in sod., 2006, 2009b; Löber in sod., XNUMX).
Nazadnje je proerektilni učinek zgoraj omenjenih agonistov D4 receptorjev zmanjšal tudi zaviralci dušikovega oksida-sintaze, dani v paraventrikularno jedro, in d (CH2) 5Tyr (Me) 2-Orn8-vazotocin, selektivni antagonist receptorjev oksitocina dani icv, vendar ne v paraventrikularnem jedru. Ti rezultati so v skladu s hipotezo, da agonisti receptorjev D4 prav tako stimulirajo oksitocinergične nevrone z aktiviranjem dušikovega oksid-sintaze in sproščanjem oksitocina v ekstrahipotalamičnih območjih možganov, kar posledično olajša erekcijo penisa, kot je razvidno za apomorphine in klasične agoniste D2 (Melis et al ., 2005, 2006b; Löber in sod., 2009).
Zgornja dognanja podpirajo tudi hipotezo, da dopamin sproži erekcijo penisa z delovanjem na D4 receptorje, ki se nahajajo na celičnih telesih paraventrikularnih oksitocinergičnih nevronov in povzročajo povečan priliv Ca2 + v celična telesa oksitocinergičnih nevronov, kar vodi do povečane proizvodnje dušikovega oksida. Dušikov oksid pa aktivira oksitocinergične nevrone, da sprostijo oksitocin v ekstrahipotalamičnih predelih možganov in v hrbtenjači, kot je že bilo razpravljano. V zvezi s tem, omembe vredno je, da so bili v celičnih telesih oksitocinergičnih nevronov v paraventrikularnem jedru prepoznani dopaminski receptorji šele pred kratkim z dvojnimi imuno-fluorescentnimi študijami z visoko selektivnimi protitelesi D2, D3 in D4 ter z oksitocinskimi protitelesi. Te študije so pokazale izražanje vseh treh podtipov D2receptorjev (D2, D3 in D4), ki se lokalizirajo ločeno v celičnih telesih oksitocinergičnih nevronov v paraventrikularnem jedru (in tudi v supraoptičnem jedru in medialnem predoptičnem območju) (Baskerville in Douglas, 2008; Baskerville in sod., 2009).
To zagotavlja močno nevroanatomsko podporo možnosti, da dopamin in agonisti dopaminskih receptorjev tipa D2 inducirajo erekcijo penisa z aktiviranjem neposrednih oksitocinergičnih nevronov, ki štrlijo v ekstrahipotalamična območja možganov, odpoklicana zgoraj, npr. hrbtenjača, ventralno tegmentalno območje, hipokampus in amigdala. Vendar te ugotovitve ne pomagajo pri prepoznavanju podtipov / receptorjev D2, katerih stimulacija povzroča erektilni odziv. Na žalost ni mogoče dobiti pomoči niti iz raziskav, katerih namen je prepoznati oksitocinergične nevrone, ki jih aktivirajo agonisti dopaminskih receptorjev v paraventrikularnem jedru. Dejansko so kljub različni aktivnosti različnih podtipov receptorjev dopamina bodisi mešani agonisti dopaminskih receptorjev (npr. Apomorfin) ali selektivni agonisti D2 receptorjev (npr. Kvinpirol, ki deluje na vse podtipe receptorjev D2) ali selektivni agonisti receptorjev D4 aktivacijo oksitocinergičnih nevronov, merjeno s povečanjem proteina FOS v parvoceličnih oksitocinergičnih nevronih paraventrikularnega jedra (Bitner et al., 2006). Vendar pa je bila ta ugotovitev nedavno vprašljiva, saj je bilo povečanje FOS beljakovin paraventrikularnih oksitocinergičnih nevronov ugotovljeno šele, ko je erekcijo penisa povzročil kinerolan, ki deluje predvsem na podtipe receptorjev D2 in D3, ne pa na PD 168077, agonist receptorja D4, kljub zmožnosti obeh spojin, da inducirata spolni odziv (Baskerville et al., 2009).
Nadaljnji poskusi s selektivnimi agonisti drugih podtipov receptorjev D2 (predvsem D2 in D3) so nato potrebni, da se ugotovi natančna vloga vsakega podtipa receptorjev dopamina pri nadzoru erektilne funkcije na paraventrikularni ravni. V zvezi s tem je, kot smo že omenili, apomorfin, ki deluje močno na vse podtipe receptorjev dopamina (glejte Brioni in sod., 2004 in reference v njem), veliko bolj učinkovit kot agonisti receptorjev D4 pri indukciji erekcije penisa, ko ga injiciramo v paraventrikularno jedro. To je mogoče razložiti z višjo afiniteto apomorfina na D4 receptorje v primerjavi z testiranimi agonisti D4 receptorjev, ali pa lahko agonisti D4 receptorjev delujejo kot delni agonisti receptorjev D4 ali pa sočasno aktiviranje različnih podtipov receptorjev dopamina s pomočjo apomorphina proizvajajo večjo aktivacijo oksitocinergičnih nevronov, ki posredujejo erekcijo penisa, kot pa aktiviranje samo agonistov receptorja D2 receptorja podtipa D4.
Interakcije med receptorji za dopamin D1 in D2 so bile že opisane pri nadzoru spolnega vedenja na ravni medialnega predoptičnega območja (glej Hull in sod., 1989). V primeru, da bo nezmožnost selektivnih agonistov receptorjev D2 povzročiti erekcijo penisa (Hsieh in sod., 2004) potrjena (vendar glejte Depoortère in sod., 2009), na primer tudi po vbrizganju teh spojin v paraventrikularno jedro. , je treba podrobno analizirati glavno vlogo samo D3 receptorjev ali skupaj z D4 receptorji pri aktivaciji oksitocinergičnih nevronov, ki posredujejo erekcijo penisa (glejte Baskerville in sod., 2009). Na žalost selektivnih agonistov receptorjev D2 in D3 (npr. Ki se razlikujejo po afiniteti do teh dveh podtipov receptorjev in vitro in vitro) ni na voljo. Zaradi tega je nedavna domneva, da receptorji D3 posredujejo erekcijo penisa, ki jo povzročajo klasični agonisti receptorjev D2, ki temelji predvsem na zmožnosti domnevnih antagonistov receptorjev D3, značilnih v poskusih in vitro, zmanjšati erekcijo penisa, ki jo povzročajo klasični agonisti D2, kot npr. apomorfin, kvinpirol in pramipeksol, ki močno aktivirajo vse podtipe receptorjev dopamina D2 (Collins et al., 2009), je treba zagotovo potrditi z drugimi poskusi. Ta validacija je potrebna tudi zato, ker v tej raziskavi niso ugotovili učinka agonistov receptorjev D4 na erekcijo penisa, kar je v nasprotju z rezultati zgoraj omenjenih študij, ki kažejo na erektilni učinek agonistov receptorjev D4. Dejansko celo sposobnost apomorfina povzroči erekcijo penisa (in zehanje) pri miših z izločanjem receptorja D4 s potenco, ki je enaka tisti, ki jo opazimo pri izločanju miši D4 receptorjev divjega tipa in zmožnostjo antagonistov receptorjev D3, da odpravi odziv apomorfina pri teh živalih ( Collins in sod., 2009) ne moremo obravnavati kot dokončen dokaz za selektivno vlogo podtipa receptorja D3 v erekciji penisa, ki jo povzroča agonist D2. Razlike med vrstami, študije z ablacijo genov na nevrotransmiter / nevropeptidni receptor (nevrotransmiter / nevropeptid in / ali živčni živčni receptor / nevrotransmiter / nevropeptidni receptor) so dodale dodatne zmede in zaplete pri potrditvi domnevne spolne vloge nevrotransmiterjev in / ali nevropeptidi in njihovi receptorji. Ablacija gena oksitocina namreč povzroči, da oksitocin izloči miši, ki se normalno parijo in kopulirajo, kot da bi bil oksitocin nepotreben za parjenje in kopulacijo. Tudi homozigotne miši oksitocina z izločanjem oksitocina kažejo normalno parjenje in paritev, čeprav z izrazito okvaro izpadanja mleka (Nishimori in sod., 1996; Young et al., 1996). Ablacija gena, ki kodira nevronski dušikov oksid sintazo, prav tako povzroči dušikov oksid sintazo, ki izloči miši, ki se normalno parijo in kopulirajo (Huang in sod., 1993). Vendar te ugotovitve verjetno kažejo na pomembno značilnost reproduktivne fiziologije, tj na odvečnost sistemov, ki sodelujejo pri njegovem nadzoru na centralni in periferni ravni.
Takšno odpuščanje ima zagotovo evolucijski izvor, saj zagotavlja prehod genov na naslednjo generacijo za preživetje vrste. Dejstvo, da ablacija gena receptorjev D4 ne spremeni proerektilnega učinka apomorfina, kaže na to, da so receptorji D4, kot oksitocin in dušikov oksid, le nekaj mediatorjev, ki delujejo v sistemih, ki nadzorujejo erektilno funkcijo, in ne kar kaže na to, da ti receptorji nimajo vloge pri nadzoru erekcije penisa in spolnega vedenja. V drugi raziskavi (Depoortère in sod., 4) so nedavno poročali o neuspehu agonistov D2009, da bi inducirali erekcijo penisov, ko jih sistemsko dajejo samcem podgan različnih sevov. Vendar v nasprotju z delom Collins in sod. (2009) in da bi slika o vlogi različnih podtipov receptorjev D2 pri nadzoru erekcije penisa postala še bolj zmedena, tudi ta raziskava kaže, da domnevni selektivni antagonisti receptorjev D3, dani sistemsko, niso mogli zmanjšati erekcije penisa, povzročene z apomorfinom. samci podgan seva, bolj občutljivi na proerektilni učinek apomorfina, medtem ko je spolni odziv (in zehanje) antagoniziral selektivni antagonist D2 L-741,626 (3 - [[4- (4-klorofenil) -4-hidroksipiperidin -1-il] metil-1H-indol), zaradi česar avtorji domnevajo, da so D2 receptorji in ne receptorji D3 in D4 tisti, ki igrajo glavno vlogo pri erekciji penisov, ki jo povzroča D2 (Depoortère et al., 2009). Nenazadnje je tudi možnost, da je vzbujalni učinek agonistov dopaminskih receptorjev na oksitocinergične nevrone, ki posredujejo erekcijo penisa, vsaj deloma posreden in ne neposreden, npr. Posredovan ali pod vplivom sprememb v delovanju drugih nevropeptidov nevropeptidov, ki lahko modulirajo aktivnost oksitocinergičnih nevronov v paraventrikularnem jedru ni mogoče popolnoma izključiti.
3.2. Interakcija glutaminske kisline in oksitocina v paraventrikularnem jedru
Paraventrikularno jedro hipotalamusa je zelo bogato s sinapsami, ki kot nevrotransmiter vsebujejo ekscitacijsko aminokislino (npr. Glutaminsko kislino in asparaginsko kislino) (Van Den Pol, 1991). Ekscitatorne aminokisline v tem jedru so vključene v številne funkcije, vključno z erekcijo penisa in spolnim vedenjem (Roeling in sod., 1991; Melis in sod., 1994b, 2000, 2004b). V skladu s tem je N-metil-d-asparaginska kislina (NMDA), selektivni agonist podtipa receptorjev NMDA, vendar ne (±) -_-amino-3-hidroksi-5-metil-izoksazol-4-propionska kislina (AMPA) , selektivni agonist podtipa AMPA receptorjev ali (±) -trans (1) -amino-1,3-ciklopentan dikarboksilna kislina (ACPD), selektivni agonist podtipa metabotropnih receptorjev, je bil sposoben inducirati erekcijo penisa, ko ga injiciramo v paraventrikularno jedro prosto gibajočih se podgan (Melis et al., 1994b). Proerektilni učinek 948 MR Melis, A. Argiolas / Neuroscience in Biobehavioral Reviews 35 (2011) 939-955 NMDA je preprečil (+) MK-801, nekonkurenčni antagonist receptorjev NMDA (Woodruff et al., 1987 ), injicirano v paraventrikularno jedro (Melis in sod., 1994b). V skladu s temi rezultati je bilo v študijah telemetrije, katerih cilj je spremljanje intrakavernoznega tlaka, NMDA veliko bolj aktivna kot agonisti drugih podtipov vzbujalnih aminokislinskih receptorjev, ko so jih injicirali v paraventrikularno jedro pri povečanju intrakavernoznega tlaka pri budnih ali anesteziranih samcih podgan (Zahran et al., 2000; Chen in Chang, 2003).
Kot je predlagano zgoraj za oksitocin in dopamin, je verjetno, da se receptorji NMDA, ki posredujejo erekcijo penisa, nahajajo v celičnih telesih oksitocinergičnih nevronov, saj živčni končiči ekscitacijskih aminokislin posegajo v celice oksitocinergičnih celic v paraventrikularnem jedru (Van Den Pol, 1991). Po analogiji s tistimi, ki so jih našli pri agonistih dopaminskih receptorjev, je proerektilni učinek NMDA očitno posredovan z aktiviranjem oksitocinergičnega nevrotransmisije, ki ga odpravi oksitocin antagonist d (CH2) 5Tyr (Me) 2-Orn8-vazotocin, ki ga daje icv, vendar ne v paraventrikularno jedro (glej Argiolas in Melis, 1995, 2004, 2005 in reference v njem). Prav tako je aktiviranje oksitocinergične nevrotransmisije, ki ga povzroča NMDA, tudi sekundarno pri aktivaciji dušikovega oksid-sintaze, saj erekcijo penisa, ki jo povzroča NMDA, preprečujejo zaviralci dušikovega oksid-sintaze (N-nitro-N-metil-arginin-metil ester in N- metil-tio-l-citrulin), ki se daje v paraventrikularno jedro, in NMDA, ki se vbrizga v paraventrikularno jedro v odmerkih, ki inducirajo erekcijo penisa, poveča proizvodnjo dušikovega oksida v hipotalamičnem jedru (glejte Argiolas in Melis, 1995, 2004, 2005 in reference nanje) ). Kar zadeva agoniste receptorjev za dopamin, je lahko aktiviranje dušikovega oksidintaze, ki ga povzroča NMDA, tudi sekundarno povečan priliv Ca2 + v oksitocinergična celična telesa prek receptorjev NMDA, vezanih na kanal Ca2 +, kot je prikazano v več nevronskih pripravkih (za pregled glejte Snyder, 1992; Southam in Garthwaite, 1993; Schuman in Madison, 1994 in njihove reference). Dušikov oksid pa aktivira oksitocinergični nevrotransmisijo (glejte zgoraj). Izvor glutamatergičnih projekcij, ki aktivirajo paraventrikularne oksitocinergične nevrone, ki posredujejo erekcijo penisa, ni znan, čeprav nekateri nevroanatomski in elektrofiziološki dokazi kažejo, da lahko vsaj deloma izvirajo iz hipokampusa (Saphier in Feldman, 1987; Chen et al., 1992) . Čeprav je potrebno nadaljnje delo, da bi bolje opredelili izvor glutamatergičnih projekcij v paraventrikularno jedro, pa v raziskave mikrodijalize očitno podpirajo vključitev glutamske kisline v paraventrikularno jedro v nadzor nad erekcijo penisa in spolno vedenje. Skladno s tem sta se v dializatu, dobljenem iz paraventrikularnega jedra samcev podgan, pokažele nekokontaktne erekcije, kadar so bile v prisotnosti nedostopnih dovzetnih samih estrogena + progesterona, podprte samice podgan (Melis et al., 2004b), penisne erekcije, ki jih posreduje tudi aktiviranje centralnega oksitocinergičnega prenosa (Melis et al., 1999a, b). Takšna povečanja so bila še večja, ko je bila dovoljena kopulacija s sprejemljivo samico (Melis in sod., 2004a). V skladu s hipotezo, da se v paraventrikularnem jedru med erekcijo in kopulacijo penisa pojavi povečana aktivnost ekscitatornih aminokislin, tako brezkontaktna erekcija kot kopulacijsko vedenje (med katerim prihaja do erekcije kopula) se zmanjšata z blokado receptorjev NMDA v paraventrikularnem jedru in temu zmanjšanju sledi zmanjšanje povečanja proizvodnje dušikovega oksida, ki se pojavi v tem hipotalamičnem jedru v teh fizioloških okoliščinah (Melis in sod., 2000). Povečanje koncentracije zunajcelične glutaminske kisline, ki je sekundarno zmanjšano sproščanje GABA iz živčnih končičev GABAergic, ki vplivajo na ekscitacijske aminokidergične sinapse, ki se namestijo na oksitocinergična celična telesa, je bilo ugotovljeno tudi v paraventrikularnem jedru po blokadi kanabinoidnih receptorjev CB1 s strani antagonista CB1, antagonista SR141761X dane v stranske preddrege ali neposredno v paraventrikularno jedro v odmerkih, ki inducirajo erekcijo penisa (glej Succu in sod., 2006; Castelli in sod., 2007). Takšno povečanje je privedlo do aktiviranja dušikovega oksid-sintaze v celičnih telesih oksitocinergičnih nevronov, kar je povečalo proizvodnjo dušikovega oksida. Dušikov oksid posledično aktivira oksitocinergične nevrone, ki posredujejo erekcijo penisa, kot je opisano zgoraj. Skladno s takšnim mehanizmom je bila erekcija penisa, povzročena s SR 141761A, zmanjšana z blokado receptorjev NMDA in zaviralci dušikovega oksida-sintaze, ne pa z blokado receptorjev dopamina ali oksitocina v paraventrikularnem jedru, medtem ko ga je preprečila blokada centralni receptorji za oksitocin, ki jih antagonisti oksitocinskih receptorjev dajo icv
3.3. Interakcija oksitocin-dopamin v ventralnem tegmentalnem območju
Oksitocin povzroči erekcijo penisa, če ga injiciramo v kaudalni del ventralnega tegmentalnega območja na način, odvisen od odmerka. (Melis in sod., 2007). Najnižji aktivni odmerek, ki ga enostransko injiciramo, je bil 20 ng, najvišji testirani odmerek pa 100 ng. Učinek oksitocina se posreduje z aktiviranjem oksitocinergičnih receptorjev, saj se spolni odziv odpravi s predhodno injekcijo antagonista oksitocina d (CH2) 5Tyr (Me) 2-Orn8-vazotocin v kaudalno ventralno tegmentalno območje. Ti receptorji so lokalizirani v celičnih telesih dopaminergičnih nevronov, ki štrlijo predvsem v lupino jedra jedra. Skladno s tem (1) dvojne imuno-fluorescentne študije kažejo, da so oksitocinergična vlakna v kaudalnem ventralnem tegmentatu v tesnem stiku s celičnimi telesi dopaminergičnih nevronov, katerih večina je bila pozitivno označena za tirozin-hidroksilazo in vsebuje retrogradni sledilnik Fluorogold injiciranje v lupino nukleus acumbens (Melis et al., 2007) in (2) ventralno tegmentalno območje erekcija penisa, povzročena z oksitocinom, se pojavlja sočasno s povečanjem koncentracije zunajceličnega dopamina v dializatu, dobljenem iz lupine the nucleus accumbens (Melis in sod., 2007). Erekcija penisa, ki jo povzroča oksitocin, se pojavlja sočasno s povečanjem proizvodnje dušikovega oksida v ventralnem tegmentalnem območju, saj sta oba odgovora antagonizirana ne samo z d (CH2) 5Tyr (Me) 2-Orn8-vazotocin in zaviralcem sinteze dušikovega oksida S- metil-tio-l-citrulin, pa tudi _-konotoksin, od napetosti odvisen blokator kanalov Ca2 + in ODQ (1H- [1,2,4] oksadiazol [4,3-a] kinoksalin-1-on), močan zaviralec gvanilata ciklaza, vsa navedena v kaudalnem ventralnem tegmentalnem območju pred oksitocinom (Succu et al., 2008). Ker je bilo veliko dopaminergičnih celičnih teles, označenih s fluororogoldom, na katere so prišli stiki z oksitocinergičnimi vlakni, za katere je bilo ugotovljeno, da so pozitivne na tirozin hidroksilazo v kaudalnem ventralnem tegmentalnem območju, so bile pozitivno označene tudi za dušikov oksid-sintazo in gvanilat ciklazo (Succu et al., 2008), oksitocin. -povedano erekcijo penisa lahko posreduje naslednji mehanizem. Aktivacija oksitocinergičnih receptorjev v dopaminergičnih celičnih telesih s peptidom poveča Ca2 + priliv v celična telesa dopaminergičnih nevronov. To aktivira dušikov oksid-sintazo, od Ca2 + -kalmodulina odvisen encim in tako poveča proizvodnjo dušikovega oksida. Dušikov oksid nato aktivira gvanilat ciklazo, kar vodi do povečane koncentracije cikličnega GMP. Ciklični GMP aktivira dopaminergične nevrone, ki štrlijo v akumulacijsko jedro. Vloga cikličnega GMP pri erekciji penisa, ki jo povzroča oksitocin, ki se vbrizga v kaudalno ventralno tegmentalno območje, podpira tudi zmožnost 8-bromo-cikličnega GMP, aktivnega analoga cikličnega GMP, odpornega na fosfodiesterazo, da sproži erekcijo penisa, ko ga injiciramo v kaudalno ventralno tegmentalno območje in za povečanje zunajceličnega dopamina v dializi iz jedrnih jezgrov (Succu in sod., 2008; Melis in sod., 2009a). To je v nasprotju z mehanizmom, s katerim dušikov oksid aktivira oksitocinergične nevrone v paraventrikularnem jedru, pri čemer 8-bromo-ciklični GMP ne more inducirati erekcije penisa, kadar ga injiciramo v to jedro (Melis in Argiolas, 1995b) (sl. 2). Glede nevronskih poti, ki jih aktivira dopamin v jedru jedra, kar vodi v erekcijo penisa, ti še vedno niso znani. Vendar razpoložljivi podatki kažejo, da ti poti aktivirajo nevrotransmisijo dopamina v paraventrikularnem jedru hipotalamusa. Skladno s tem se erekcija penisa, ki jo povzroča oksitocin, pojavi hkrati s povečanjem zunajceličnega dopamina, ne samo v nucleus accumbens, vendar tudi v paraventrikularnem jedru, antagonizira ga antagonist receptorja dopamina haloperidol, ki ga vbrizgajo v paraventrikularno jedro (Melis in sod., 2007). ASkupaj bodo ti rezultati podprli idejo, da oksitocinergični nevroni, ki izvirajo iz paraventrikularnega jedra in štrlijo v kaudalno ventralno tegmentalno območje, ko v tem območju sprostijo oksitocin in s tem aktivirajo NO-ciklični signalni sistem GMP, kar posledično aktivira mezolimbične dopaminergične nevrone (Melis in sod., 2007, 2009a; Succu in sod., 2008). Dopamin, sproščen v jedru, se nato aktivira nevronske poti, ki vodijo do aktivacije incerto-hipotalamičnih dopaminergičnih nevronov, ki spodbujajo paraventrikularne oksitocinergične nevrone, ki štrlijo v erekcijo hrbtenjače, ki posreduje na penisu. Hkrati lahko dopamin, sproščen v paraventrikularnem jedru, aktivira tudi oksitocinergične nevrone, ki segajo v ekstrahipotalamična možganska območja, kot so ventralno tegmentalno območje, hipokampus, amigdala in morda druga možganska območja.
Kot je že omenjeno zgoraj, v skladu s to hipotezo apomorfin, ki ga injiciramo v paraventrikularno jedro v odmerku, ki povzroči erekcijo penisa, poveča tudi koncentracijo zunajceličnega dopamina v jedru, kar vpliva na antagonist oksitocinskega receptorja d (CH2) 5Tyr (Me ) 2-Orn8-vazotocin injiciran v ventralno tegmentalno območje (Succu in sod., 2007; Melis in sod., 2009a). Zgornje nevronske poti lahko skupaj tvorijo hipotetično nevronsko vezje, ki vključuje dopamin, oksitocin in druge nevrotransmiterje (npr. Glutaminsko kislino, glej spodaj), ki vplivajo ne samo na spolno zmogljivost, temveč tudi na spolno motivacijo in spolno nagrajevanje (glej razdelek 4).
3.4. Medsebojno delovanje oksitocin-glutaminske kisline v ventralnem subkulumu hipokampusa
Oksitocin inducira erekcijo penisa, če ga injiciramo v ventral, vendar ne v dorzalni subkuulum, odvisno od odmerka (Melis in sod., 2009b). Spolni odziv je posredovan s stimulacijo oksitocinskih receptorjev, odpravljen pa je s predhodno injekcijo d (CH2) 5Tyr (Me) 2-Orn8-vazotocina, ki se daje na isto mesto oksitocina, kot ga najdemo na drugih možganskih območjih (glejte zgoraj) . Glede lokalizacije teh receptorjev razpoložljivi podatki kažejo, da so lokalizirani v celičnih telesih nevronov, bogatih z dušikovim oksid-sintazo.
Skladno s tem, raziskave mikrodialize kažejo, da se erekcija penisa, ki jo povzroča oksitocin, pojavi sočasno s povečanjem proizvodnje dušikovega oksida v ventralnem subkukulu in se ta porast ne odpravi samo s predhodnim vbrizgavanjem zaviralca dušikovega oksidintaze S-metil-tio-l-citrulina in s hemoglobinom, ki čistilec dušikovega oksida, pa tudi z d (CH2) 5Tyr (Me) 2-Orn8-vazotocin, vsi dani na isto mesto oksitocina v odmerkih, ki preprečujejo erekcijo penisa (Melis et al., 2010). Pomembnejše je, da erekcija penisa, ki jo povzroča oksitocin, pride tudi sočasno s povečanjem koncentracije zunajcelične glutaminske kisline v ventralnem subkukulu, ki ga le delno antagonizira nekonkurenčni antagonist receptorja NMDA (+) MK-801, dan v ventralni subiculum (Melis in sod., 2010). Ti rezultati skupaj kažejo, da novo nastali dušikov oksid, ki deluje kot medcelični glasnik, aktivira nevrotransmisijo glutaminske kisline, ki vodi v erekcijo penisa, morda z nevronskimi eferentnimi projekcijami iz ventralne celice subiculum na možganskih območjih zunaj hipokampa. V skladu s to hipotezo oz. NMDA, ki se vbrizga v ventralni subiculum, povzroči erekcijo penisa na način, odvisen od odmerka, in ta učinek se v celoti antagonizira s predhodno injekcijo na isto mesto (+) MK-801, ne pa s S-metil-tio-l-citrulinom , hemoglobin ali d (CH2) 5Tyr (Me) 2-Orn8-vazotocin (Melis et al., 2010). Kar zadeva nevronske eferentne poti, ki štrlijo v možganska območja zunaj hipokampa, aktivirana z ekscitacijskimi aminokislinami (tj. Glutaminsko kislino) v ventralnem subkukulu, je verjetno, da gre za glutamatergične učinke, kot je večina efektivnih projekcij hipokampa. Trenutno lahko le predlagamo, da te projekcije modulirajo aktivnost mezolimbičnih dopaminergičnih nevronov, ki nato modulirajo aktivnost incerto-hipotalamičnih dopaminergičnih nevronov v paraventrikularnem jedru, kar vodi do aktivacije oksitocinergičnih nevronov posredovanje erekcije penisa, kot je bilo že razpravljeno (glej zgoraj).
Skladno s tem se erekcija penisa, ki jo povzroči ventralni subiculum oksitocin, pojavi sočasno s povečanjem koncentracije zunajceličnega dopamina v lupini jedra jedra, in to povečanje, tako kot erekcija penisa, odpravi z d (CH2) 5Tyr (Me) 2- Orn8-vazotocin, ki ga dajemo v ventralni subkulum pred oksitocinom (Melis et al., 2009b). Ker se erekcijo penisa, ki jo povzroči ventralni subiculum oksitocin, zmanjša tudi (+) MK-801, ki se vbrizga v ventralno tegmentalno območje, ne pa v jedro jedra (Melis et al., 2009b) in se pojavi sočasno do povečanja zunajceličnih. glutaminska kislina v ventralnem tegmentalnem območju, vendar ne v jedru jedra, pri čemer sta oba odgovora odpravila d (CH2) 5Tyr (Me) -Orn8-vazotocin, injiciran v ventralni subkulum pred oksitocinom (glej sliko 3), verjetno je aktivacija mezolimbičnih dopaminergičnih nevronov sekundarna za povečano glutamatergično nevrotransmisijo v ventralnem tegmentalnem območju. To kaže, da na ventralnem tegmentalnem območju obstaja interakcija glutaminske kisline in dopamina, ki nadzira erekcijo penisa. Potrebne so nadaljnje študije, da se ugotovi, ali so proerektilne eferentne glutametergične poti od subiculuma do ventralnega tegmentalnega območja neposredne ali posredne, torej skozi predfrontalno skorjo ali druga možganska območja (glejte Melis et al., 2009b in njihove reference). Ker paraventrikularno jedro prejema tudi glutamatergične projekcije hipokampusa (glej zgoraj in Saphier in Feldman, 1987), in glutaminska kislina aktivira paraventrikularne oksitocinergične nevrone, vključno s tistimi, ki štrlijo v ventralno tegmentalno območje (glej Argiolas in Melis, 2005 in njihove reference) in oksitocin v ventralnem tegmentalnem območju povzroči erekcijo penisa in poveča aktivnost mezolimbičnega dopaminergičnega nevronas (glej zgoraj), skušnjava, da paraventrikularni oksitocinergični nevroni so lahko vsaj delno vključeni tudi v aktivacijo mezolimbičnih dopaminergičnih nevronov z oksitocinom, ki se vbrizga v ventralni subiculum (glej razdelek 4).
4. Zaključne opombe
Zgoraj opisane študije potrjujejo in razširjajo zgodnje ugotovitve, ki kažejo, da ima oksitocin pri samcih podgan ključno vlogo pri centralnem nadzoru erekcije penisa na ravni paraventrikularnega jedra hipotalamusa in hrbtenjače. Zlasti najbolj reŠtudije centov kažejo, da oksitocin vpliva na erekcijo penisa tudi z delovanjem na drugih možganskih območjih, tj. na ventralnem tegmentalnem območju, ventralnem subiculumu in posteromedialnem kortikalnem jedru amigdale.
Na paraventrikularni ravni je najpomembnejša nova ugotovitev morda odkritje izražanja vseh dopaminskih receptorjev iz družine D2 (D2, D3 in D4) v celičnih telesih oksitocinergičnih nevronov v paraventrikularnem jedru (in v supraoptičnem jedru in medialno preoptično območje) (Baskerville in Douglas, 2008; Baskerville et al., 2009). To zagotavlja močno nevroanatomsko podpora hipotezi, da lahko dopamin in agonisti dopaminskih receptorjev aktivirajo neposredno oksitocinergične nevrone, ki sodelujejo pri erektilni funkciji in štrlijo ne le na hrbtenjačo, temveč tudi na možgane, ki niso hipotalamični.. V zvezi s tem je pomembno tudi odkritje, da se povečanje intrakavernoznega tlaka, ki ga povzroča agonist dopaminskih receptorjev, zmanjša z blokado oksitocinergičnih receptorjev v ledveno-križnični hrbtenjači (Baskerville et al., 2009). Čeprav so bili takšni dokazi pridobljeni pri anesteziranih samcih podgan, ugotovitev potrjuje aktiviranje paraventrikulo-spinalne oksitocinergične poti padajoče erekcije, ki jo povzroča agonist dopaminskih receptorjev. Hvendar je treba še ugotoviti, ali je erekcija penisa, ki jo povzroča stimulacija receptorjev dopamina v telesih oksitocinergičnih celic, sekundarna za aktiviranje specifičnega podtipa receptorjev dopamina iz družine D2 (D2, D3 ali D4) ali če ti receptorski podtipi sodelujejo pri moduliranju erektilnega odziva, po možnosti na različne načine, odvisno od konteksta, v katerem se pojavi erekcija penisa (glej Moreland in sod., 2004; Enguehard-Gueiffier et al., 2006; Melis et al., 2006a, b; Löber in sod., 2009; Collins in sod., 2009; Depoortère in sod., 2009; Baskerville in sod., 2009).
Druga pomembna nova ugotovitev je, da oksitocin sproži erekcijo penisa, če ga injiciramo ne samo v paraventrikularno jedro ali v polje CA1 hipokampusa, temveč tudi v ventralno tegmentalno območje, ventralni subiculum in posteromedialno kortikalno jedro amigdale. Ta področja možganov niso bila testirana v prejšnjih študijah, ki kažejo, da je oksitocin povečal epizode spontane erekcije penisa pri samcih podgan, čeprav dobivajo podobno oksitocinergične projekcije ledvenega križa hrbtenjače iz paraventrikularnega jedra. Oksitocin je bil resnično sposoben povečati epizode spontane erekcije penisa, ki se pri odraslih moških podganah pojavijo brez kakršnih koli spolnih dražljajev, kot so tisti, ki izvirajo iz prisotnosti dostopne ali nedostopne receptivne (estrogen-progesteron nasičene) jajčne podgane ali manipulacija genitalij, ko se vbrizga v paraventrikularno jedro in v polje hipokampusa CA1, vendar ne v dorzalni subiculum, stranski septum, kaudatno jedro, medialno preoptično območje, ventromedialno jedro in supraoptično jedro (Melis et al. , 1986). V vseh teh študijah se je erekcija penisa običajno štela, ko je penis iz penine izšel iz opaznega opazovalca, ki se je med poskusom ali pozneje poznal na zdravljenje, ne da bi se seznanil z eksperimentom, posnetim na videokaseto z aparatom za video kamero. Vsaka epizoda erekcije penisa traja min. 0.5-1, običajno pa jo spremljata nego penilcev in / ali fleksija kolka. Na teh podganah običajno ne poteka nobenega eksperimenta, da bi ugotovili učinek spolnih izkušenj, starosti ali če jih lahko podgane razdelimo na nizke ali visoke odzive na proerektilni učinek oksitocina, ki ga vbrizgajo v različna možganska območja. To velja tudi za večino raziskav o erektilnem učinku drugih nevropeptidov in zdravil, ki povečajo epizode spontane erekcije penisa, vključno z agonisti dopamina, ekscitacijskimi aminokislinami, ACTH-MSH, heksarelinom in VGF peptidi. Vendar pa je proerektilni učinek teh spojin večkrat potrjen s telemetričnimi metodami, ki določajo pojav erekcije penisa s povečanjem intrakavernoznega tlaka, ki se pojavi spontano ali po dajanju teh spojin po različnih poteh, tj. Sistemsko, intracerebroventrikularno oz. neposredno po vsaditvi tlačnega mikrotransduktorja neposredno v kavernozno korpus (neposredno v Bernabè et al., 1999). V ventralnem tegmentalnem območju, ventralni subiculum in posteromedialno jedro amigdale tudi oksitocin povzroči erekcijo penisa z delovanjem na oksitocinergične receptorje. To vodi do aktiviranja mezolimbičnih dopaminergičnih nevronov, ki izvirajo iz ventralnega tegmentalnega območja in štrlijo v lupino jedra jedra, merjeno s povečanjem koncentracije zunajcelične dopamina v dializatu, dobljenem iz lupine jedrnih jezgrov, in zmanjšanjem erektilnega odziva, ki ga inducira peptid, vbrizgan v ta ekstrahipotalamična območja, ugotovljena po blokadi dopaminergičnih receptorjev v nucleus accumbens (glej spodaj). Kar zadeva mehanizme, ki jih aktivirajo oksitocinergični receptorji na teh možganskih območjih, ki vodijo do aktivacije mezolimbičnih dopaminergičnih nevronov in do erekcije penisa, so najbolje razjasnjeni tisti, ki se pojavljajo v kaudalnem ventralnem tegmentalnem območju. Dejansko tukaj farmakološki in imuno-fluorescentni rezultati kažejo, da živčni končiči oksitocina posegajo v celična telesa dopaminergičnih nevronov, ki štrlijo v lupino jedra jedra (Melis in sod., 2007, 2009a; Succu in sod., 2008). Mnogi od teh nevronov so bogati s sintezo dušikovega oksida in z gvanilat ciklazo. Stimulacija oksitocinergičnih receptorjev v celičnih telesih teh dopaminergičnih nevronov povzroči aktiviranje sintaze dušikovega oksida, kar vodi do povečane proizvodnje dušikovega oksida. Dušikov oksid nato aktivira gvanilat ciklazo in s tem poveča koncentracijo cikličnega GMP oz. kar privede do aktivacije mezolimbičnih dopaminergičnih nevronov in do sproščanja dopamina v nukleusu, kar merimo s povečanjem zunajceličnega dopamina v dializatu iz jedrnih jezgrov, pridobljenih z intra-cerebralno mikrodijalizo (Succu et al., 2008 ). Dopamin, sproščen v jedru, se nato aktivira nevronske poti, kar vodi do erekcije penisa. To podpira sposobnost antagonistov dopaminskih receptorjev haloperidola in / ali cis-flupentiksola, ki se vbrizga v jedro, da zmanjšajo erekcijo penisa, ki jo povzroči oksitocin v ventralni tegmentaciji (Succu et al., 2008). Zdi se, da eden od proerektilnih poti aktivira incerto-hipotalamične dopaminergične nevrone, zlasti tiste, ki štrlijo v celična telesa paraventrikularnih oksitocinergičnih nevronov. Dejansko oksitocin, ki se vbrizga v kaudalno ventralno tegmentalno območje, poveča zunajcelični dopamin ne le v jedru jedra, temveč tudi v paraventrikularnem jedru, blokada dopaminskih receptorjev v paraventrikularnem jedru pa znatno zmanjša ventralno tegmentalno območje, kar povzroča oksitocin, ki ga povzroča okončanje penisa (Succu et al. , 2007, 2008; Melis et al., 2007, 2009a). Obstoj teh jeder združuje dopaminsko-paraventrikularno dopaminsko-paraventrikularno oksitocin-ventralno tegmentalno območje oksitocin-dopaminske povezave predlaga tudi sposobnost proerektilnega odmerka apomorfina in agonista receptorja D4 PD 168077, ki se vbrizga v paraventrikularno jedro, da poveča zunajcelični dopamin v lupini jedrnega jedra (Succu et al., 2007), odziv ki se odpravi z d (CH2) 5Tyr (Me) -Orn8-vazotocin, ki ga damo v ventralno tegmentalno območje (Melis et al., 2009a, glej tudi spodaj). Vendar pa je potrebno nadaljnje delo za prepoznavanje nevronskih poti, ki povezujejo jedra jedra z incerto-hipotalamičnim dopaminergičnim sistemom.
Mehanizem, s katerim oksitocin inducira erekcijo penisa in aktivira mezolimbične dopaminergične nevrone, kadar ga injiciramo v ventralni subiculum ali v posteromedialno jedro amigdale, je trenutno le delno razumljiv. Tudi na teh območjih oksitocin aktivira lastne receptorje, ki vodijo v aktivacijo dušikovega oksid-sintaze in s tem povečajo proizvodnjo dušikovega oksida. Dušikov oksid pa aktivira neznane eferentne projekcije, ki očitno povečujejo glutamatergično nevrotransmisijo v ventralnem tegmentalnem območju. Glutaminska kislina nato stimulira mezolimbične dopaminergične nevrone, kar vodi v erekcijo penisa. To hipotezo podpira predvsem sposobnost oksitocina, ki se vbrizga v ventralni subiculum, da poveča zunajcelično glutaminsko kislino na ventralnem tegmentalnem območju (sl. 3) in nekonkurenčnega antagonista receptorjev NMDA (+) MK-801 injicirajo v ventralno tegmentalno območje, vendar ne v jedru jedra, da bi zmanjšali erekcijo penisa, ki jo povzroči oksitocin, injiciran bodisi v ventralni subkulum ali v posteromedialno jedro amigdale (Melis in sod., 2009b). Trenutno je na voljo več podrobnosti za erekcijo penisa, ki jo povzroča oksitocin. Tu se zdi, da je erekcija penisov, povzročena z oksitocinom, sekundarna zaradi aktiviranja oksitocinergičnih receptorjev, ki se nahajajo v celičnih telesih nevronov, ki vsebujejo dušikov oksid-sintazo. To povzroči povečanje proizvodnje dušikovega oksida, ki aktivira glutamatergično nevrotransmisijo, tako da deluje kot medcelični sel z mehanizmom, podobnim kot je opisan za dolgotrajno potenciranje (glej Snyder, 1992; Southam in Garthwaite, 1993; Schuman in Madison, 2004) . V skladu s to hipotezo erekcijo penisa, ki jo povzroča oksitocin, se pojavlja sočasno s povečanjem zunajcelične glutaminske kisline v dializatu iz ventralnega subkukulata in stimulacijo ekscitacijskih receptorjev za aminokisline v ventralnem subicumu z NMDA, povzroči erekcijo penisa. Glutaminska kislina aktivira nevronske eferentne projekcije, ki vodijo do aktiviranja mezolimbičnih dopaminergičnih nevronov v ventralnem tegmentalnem območju, kot je navedeno zgoraj. Če ti mehanizmi delujejo tudi v posteromedialnem jedru amigdale, za zdaj ni znano. Poleg tega so potrebne nadaljnje študije, ki dokazujejo, da so (1) oksitocinergični živčni končiči in receptorji v ventralnem subkuumu in v posteromedialnem jedru amigdale lokalizirani v celičnih telesih nevronov, ki vsebujejo dušikov oksid-sintazo (2), če so ti nevroni sčasoma pod nadzorom ekscitacijskih aminokislinskih (glutamatergičnih) sinaps in (3) za identifikacijo signalizacijskega sistema, ki ga aktivira dušikov oksid. V zvezi s tem je treba opozoriti, da spominja sposobnost oksitocina, da poveča aktivnost dušikovega oksida-sintaze v celičnih telesih dopaminergičnih nevronov v kaudalnem ventralnem tegmentacijskem območju in še vedno neznanih nevronov v ventralnem subkukulu in posteromedialnem jedru amigdale. sposobnost oksitocina, da v celičnih telesih oksitocinergičnih nevronov v paraventrikularnem jedru aktivira dušikov oksid-sintazo (Melis et al., 1997). Toda medtem ko povečana proizvodnja dušikovega oksida v celičnih telesih dopaminergičnih nevronov vodi v aktivacijo teh nevronov z aktiviranjem gvanilat ciklaze in povečanjem ciklične GMP, se to v paraventrikularnem jedru ne zgodi. V skladu s tem 8-bromo-ciklični GMP, injiciran v paraventrikularno jedro, ne povzroči erekcije penisa, medtem ko to počne, če ga injiciramo v ventralno tegmentalno območje. Druga signalna pot, ki se razlikuje od dušikovega oksidocikličnega GMP sistema, je nato na paraventrikularni ravni vključena v aktivacijo oksitocinergičnih nevronov, ki posredujejo erekcijo penisa z endogenim in / ali eksogenim dušikovim oksidom (Melis in Argiolas, 1995b; Melis et al., 1997) (sl . 1). Po drugi strani se zdi, da ciklični GMP v ventralnem tegmentalnem območju igra ključno vlogo tudi pri aktivaciji mezolimbičnih dopaminergičnih nevronov in pri povečanju zunajceličnega dopamina, ki se pojavlja v dializatu, pridobljenem iz lupine jedra jedra samcev podgan izbrano za prikaz ali ne prikaz nekontaktne erekcije penisa, če ga dajemo v prisotnosti nedostopne jajčnike, podvržene jajčniku (z estrogenom + progesteronom) podgane. V teh eksperimentalnih razmerah pri samcih podgan, ki kažejo brezkontaktno penisno erekcijo, ugotovimo povečanje koncentracije zunajcelične dopamina, kot je bilo pričakovano, in še povečajo, čeprav le skromno, zaviralci fosfodiesteraze, ki jih dajo v tegmentalno območje kaudalnega ventrala (Sanna et al., 2009).
Sposobnost oksitocina, ki se vbrizga v ventralno tegmentalno območje, v ventralni subkulum in v posteromedialno kortikalno jedro amigdale, skupaj s tistimi dopaminskih agonistov, injiciranih v paraventrikularno jedro, da sproži erekcijo penisa in aktivira mezolimbične dopaminergične nevrone, zasluži nekaj komentarja . Prvič, mehanizmi, podobni zgoraj omenjenim, lahko delujejo, kadar pride do erekcije penisa v fizioloških okoliščinah, na primer med kopulacijo (ko pride do erekcije penisa kopule) ali med nekontaktno erekcijo penisa. Te erekcije so feromonske erekcije penisov, ki jih ni mogoče razlikovati od tistih, ki jih povzročajo droge ali oksitocin, in se pojavijo, ko se spolno močne podgane položijo v prisotnost nedostopne dovzetne (ovariektomizirane estrogene + progesteronske nasičene) ženske podgane in se štejejo za indeks spolnih vzburjenje (Sachs, 1997, 2007). Čeprav ti rezultati ne kažejo, da ima oksitocin na teh področjih vlogo pri erekciji penisa, ki se pojavlja v fizioloških okoliščinah ali po dajanju zdravil, dodajo dodatno moč zgodnjim ugotovitvam, ki kažejo, da ta področja možganov pripadajo tistim, kjer oksitocin centralno deluje na povečanje ne samo pri spontanih erekcijah penisa, ki so jih opazili po proerektilnih zdravilih, temveč tudi za izboljšanje moškega (in ženskega) spolnega vedenja (glejte Argiolas in Melis, 2004 in reference v njih). V skladu s tem se koncentracija oksitocina poveča v hipokampusu samcev podgan, ki se zdravijo s proerektilnim odmerkom apomorfina, klasičnega agonista dopamina (Melis et al., 1990) in d (CH2) 5Tyr (Me) -Orn8-vazotocin, ki blokira oksitocin receptorjev, je izjemno učinkovit ne le pri motenju kopulacijskega vedenja (Argiolas et al., 1987a), ampak tudi pri lajšanju učinka apomorfina na moško kopulacijsko vedenje (Argiolas et al., 1987b) pri spolno močnih podganah, med katerimi je v erekciji penula copula se pojavijo. d (CH2) 5Tyr (Me) -Orn8-vazotocin je tudi izjemno močan pri zmanjševanju nekontaktnih erekcij pri spolno močnih podganah, če ga dajemo v količini nanograma v stranske preddvore, ne pa v PVN (Melis et al., 1999a).
Drugič, mezolimbični dopaminergični nevroni igrajo ključno vlogo pri motivacijskih in nagrajujočih lastnostih naravnih ojačujočih dražljajev, kot so hrana, voda in spolna aktivnost (Fibiger in Phillips, 1988; Wise in Rompre, 1989; Everitt, 1990). Zlasti se domneva, da dopamin, sproščen iz teh nevronov, posreduje pri prenosu motivacijskih vidikov naravnih dražljajev v ciljno usmerjena vedenja, na primer v primeru spolne aktivnosti, iskanja spolnega partnerja in spolnega odnosa, da bi dosegli nagrado in zadovoljstvo (Goto in milost, 2005). V skladu s tem se koncentracija zunajceličnega dopamina v dializi iz nukleusnih akumulatorjev spolno močnih podgan med izpostavljenostjo nedostopnim jajčnim podmrežljivim estrogenom + progesteronom, ki so bile primirirane samice podgane (Pfaus in Everitt, 1995).
Tretjič, pričujoči rezultati podpirajo hipotezo, da nevronsko vezje povezuje paraventrikularno jedro z ventralnim tegmentalnim območjem neposredno ali posredno (skozi ventralni subiculum in / ali posteromedialno kortikalno jedro amigdale) in se jedro pripoji in od tu naprej neznano poti nazaj v paraventrikularno jedro za nadzor aktivnosti oksitocinergičnih nevronov, ki štrlijo na erekcijo hrbtenjače na hrbtenjači, in oksitocinergičnih nevronov, ki štrlijo v ventralno tegmentalno območje, ventralni subiculum in posteromedialno kortikalno jedro amigdale, ki na ta način modulirajo aktivnost mezolimbičnih dopaminergičnih nevronov (slika 4). To kompleksno nevronsko vezje ima lahko vlogo pri vključevanju nevronskih dejavnosti, ki sodelujejo pri nadzoru potrošnih (erektilno-ejakulatornih) in anticipirajočih (motivacijskih in nagrajujočih) vidikov moškega spolnega vedenja v fizioloških okoliščinah. Dejansko se zunajcelični dopamin poveča v jedru jedra (Pfaus in Everitt, 1995) in v paraventrikularnem jedru spolno močnih podgan med izpostavljenostjo nedostopni dovzetni samici podgane, ko pride do brezkontaktnih erekcij in še bolj, ko je dovoljeno kopičenje, na primer, ko pride do erekcije penisa v copuli (Melis in sod., 2003). Čeprav so potrebne nadaljnje študije za razjasnitev vloge endogenega oksitocina v ventralnem tegmentalnem območju, ventralnem subkulumu in amigdali med brezkontaktnimi erekcijami in spolnim vedenjem, je morda smiselno domnevati, da lahko ta hipotetični nevronski krog hkrati prispeva k potrošniški vidiki spolnega vedenja lahko istočasno aktivirajo tudi mezolimbični dopaminergični sistem, ki zagotavlja nevronski substrat za razlago koristnih lastnosti spolne aktivnosti (Everitt, 1990; Pfaus in Everitt, 1995). V zvezi s tem je treba opozoriti, da je mezolimbični dopaminergični sistem, aktiviran z oksitocinom, ki se vbrizga v ventralno tegmentalno območje, enako aktiviran z zlorabo drog, kot so opiati, kanabinoidi, amfetamin, kokain in alkohol (Tanda idr., 1997) in da je bilo ugotovljeno, da je oksitocin sposoben zmanjšati toleranco in odvisnost od kokaina, morfija, alkohola in kanabinoidov (Kovacs in sod., 1998; Cui in sod., 2001). Za zaključek se zdi, da oksitocin, ki se sprošča ne le v ventralnem tegmentalnem območju, temveč tudi v ventralnem subkukulu in posteromedialnem kortikalnem jedru amigdale, lahko aktivira mezolimbične dopaminergične nevrone, ki so lahko vključeni v apetitne in nagrajujoče učinke spolne aktivnosti . Aktivacija mezolimbičnih dopaminergičnih nevronov je lahko neposredna, prek oksitocinergičnih receptorjev v celičnih telesih mesolimbičnih dopaminergičnih nevronov ali posredna s pomočjo aktivacije nevrotransmisije glutaminske kisline v ventralnem tegmentalnem območju.
Dopamin, sproščen v jedru, zajema lupino, modulira aktivnost incerto-hipotalamičnih dopaminergičnih nevronov v paraventrikularnem jedru, ki povzročajo bodisi erekcijo penisa (z aktivacijo oksitocinergičnih nevronov, ki štrli v hrbtenjačo), bodisi spolno motivacijo in nagrajevanje (z aktivacijo oksitocinergike nevroni, ki štrlijo na ventralno tegmentalno območje, ventralni subiculum ali posteromedialno kortikalno jedro amigdale). Ker se dopamin sprošča tudi v lupini nukleus accumbens in v paraventrikularnem jedru, ko pride do erekcije penisa v fizioloških kontekstih (npr. Brezkontaktne erekcije in kopulacije) (Succu et al., 2007; Melis et al., 2003, 2007), je verjetno, da osrednji oksitocinergični nevroni sodelujejo v nevronskih vezjih, ki posredujejo medsebojno delovanje med mezolimbičnim in incerto-hipotalamičnim dopaminergičnim sistemom. Ti nevronski krogi lahko igrajo vlogo ne samo v fazi porabe spolnega vedenja (npr. Erekcija in kopulacija penisa), temveč tudi pri spolni motivaciji, spolnem vzburjenju in spolni nagradi.
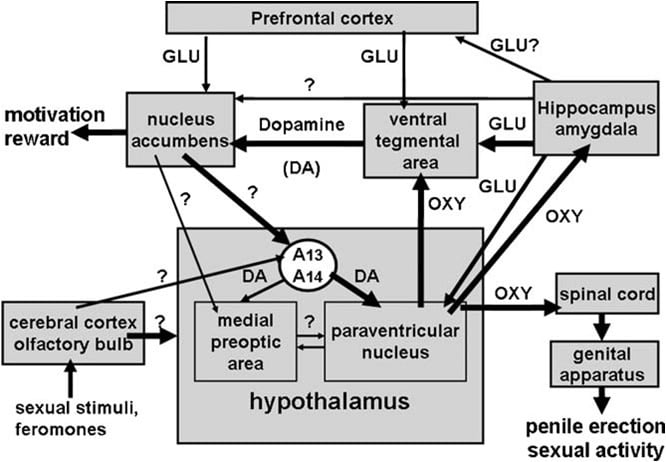
Shematski prikaz hipotetičnega nevronskega vezja, ki vključuje oksitocin, ki vpliva na spolno motivacijo, nagrajevanje in spolno uspešnost, kot predlagajo rezultati tega poglavja in prejšnja poročila. Oksitocinergični nevroni, ki izvirajo iz paraventrikularnega jedra in štrlijo v hrbtenjačo, ko jih aktivirajo na primer dopamin in glutaminska kislina (lahko pa tudi drugi nevrotransmiterji in / ali nevropeptidi), olajšajo erekcijo penisa in spolno delovanje z aktiviranjem oksitocinergičnih nevronov, ki štrlijo v hrbtenjačo. Dopamin in glutaminska kislina (pa tudi nevrotransmiterji in nevropeptidi) v paraventrikularnem jedru aktivirajo tudi oksitocinergične nevrone, ki štrlijo v ventralno tegmentalno območje, s čimer aktivirajo mezolimbične dopaminergične nevrone, ki štrlijo v jedro, prilagajajo spolno motivacijo in nagrado. Dopamin, ki se sprosti v jedru jedra (NA), aktivira še vedno neznane nevronske poti, ki povečajo aktivnost incerto-hipotalamičnih dopaminergičnih nevronov (ki izvirajo iz skupin A13-A14 iz Dahlstroma in Fuxeja), ki vsrkavajo oksitocinergične nevrone, vključno s tistimi, ki projektirajo hrbtenjača, kar vodi do erekcije penisa. To vezje se lahko aktivira tudi z oksitocinom, ki se injicira ne le v kaudalno ventralno tegmentalno območje, ampak tudi v ventralni subiculum in amigdalo, ki prav tako prejemajo oksitocinergično inervacijo iz paraventrikularnega jedra, po možnosti z neposrednim ali posrednim glutamatergičnim eferentom do ventral Tegmentalno področje, kar vodi k modulaciji tako spolne motivacije kot erekcije penisa. Nazadnje se zgornje vezje lahko aktivira tudi s spolnimi dražljaji in feromoni, saj se zunajcelični dopamin in glutaminska kislina povečujeta v paraventrikularnem jedru (in v območju medialnega predoptika) med nekontaktnimi erekcijami in kopulacijo, ki jih posreduje feromon (za primerne reference glejte Seznam referenc).
Zahvala To delo je delno podprla donacija italijanskega ministrstva za univerzo in raziskave za AA in MRM
Reference
Andersson, KE, 2001. Farmakologija erekcije penisa. Farmakol. Rev. 53, 417-450. Argiolas, A., 1994. Dušikov oksid je osrednji mediator erekcije penisa. Nevrofarmakologija 33, 1339-1344. Argiolas, A., 1999. Nevropeptidi in spolno vedenje. Neurosci. Biobehav. Rev. 23, 1127-1142. Argiolas, A., Gessa, GL, 1991. Osrednje funkcije oksitocina. Neurosci. Biobehav. Rev. 15, 217-231. Argiolas, A., Melis, MR, 1995. Nevromodulacija erekcije penisa: pregled vloge nevrotransmiterjev in nevropeptidov. Prog. Neurobiol. 47, 235-255. Argiolas, A., Melis, MR, 2004. Vloga oksitocina in paraventrikularnega jedra v spolnem vedenju moških sesalcev. Fiziol. Behav. 83, 309-317. Argiolas, A., Melis, MR, 2005. Centralna kontrola erekcije penisa: vloga paraventrikularnega jedra hipotalamusa. Prog. Neurobiol. 76, 1-21. Argiolas, A., Collu, M., Gessa, GL, Melis, MR, Serra, G., 1988. Antagonist oksitocina d (CH2) 5Tyr (Me) -Orn8-vazotocin antagonizira kopulacijsko vedenje samcev pri podganah. E. J. Farmakol. 149, 389-392. Argiolas, A., Collu, M., D'Aquila, P., Gessa, GL, Melis, MR, Serra, G., 1989. Pri podganah stimulira apomorfinsko stimulacijo moškega kopulacijskega vedenja antagonist oksitocina d (CH2) 5Tyr (Me) -Orn8-vazotocin. Farmakol. Biochem. Behav. 33, 81-83. Argiolas, A., Melis, MR, Gessa, GL, 1985. Intraventrikularni oksitocin pri podganah sproži zehanje in erekcijo penisa. E. J. Farmakol. 117, 395-396. Argiolas, A., Melis, MR, Gessa, GL, 1986. Oksitocin: izjemno močan povzročitelj erekcije penisa in zehanja pri samcih podgan. E. J. Farmakol. 130, 265-272. Argiolas, A., Melis, MR, Mauri, A., Gessa, GL, 1987a. Lezija paraventrikularnega jedra preprečuje zehanje in erekcijo penisa, ki jo pri podganah povzročata apomorfin in oksitocin, ne pa tudi ACTH. Brain Res. 421, 349-352. Argiolas, A., Melis, MR, Vargiu, L., Mauri, A., Gessa, GL, 1987b. d (CH2) 5Tyr (Me) -Orn8-vazotocin, močan oksitocinski antagonist, antagonizira erekcijo in zehanje penisa, ki ju povzročata oksitocin in apomorfin, ne pa ACTH 1-24. E. J. Farmakol. 134, 221-224. Argiolas, A., Melis, MR, Stancampiano, R., Gessa, GL, 1990. _-Konotoksin preprečuje epopenijo penisa in apnoorfina in oksitocina pri samcih podgan. Farmakol. Biochem. Behav. 37, 253-257. Arletti, R., Bertolini, A., 1985. Oksitocin spodbuja vedenje lordoze pri samicah podgan. Nevropeptidi 6, 247-255. Arletti, R., Bazzani, C., Castelli, M., Bertolini, A., 1985. Oksitocin izboljšuje moški kopulacijo pri podganah. Horm. Behav. 19, 14-20. Arletti, R., Benelli, A., Bertolini, A., 1990. Spolno vedenje starajočih se podgan spodbuja oksitocin. E. J. Farmakol. 179, 377-382. Arletti, R., Calzà, L., Giardino, L., Benelli, A., Cavazzutti, E., Bertolini, A., 1997. Spolna nemoč je povezana z zmanjšano proizvodnjo oksitocina in povečano proizvodnjo opioidnih peptidov v paraventrikularnem jedru hipotalamusa. Neurosci. Lett. 233, 65-68. Bancila, M., Giuliano, F., Rampin, O., Mailly, P., Brisorgueil, MJ, Clas, A., Verge, D., 2002. Dokazi za neposredno projekcijo iz paraventrikularnega jedra hipotalamusa v domnevne serotoninergične nevrone jedra paragigantocellularis, ki sodelujejo pri nadzoru erekcije pri podganah. E. J. Neurosci. 16, 1240-1249. Burnett, AL, Lowenstein, CJ, Bredt, DS, Chang, TSK, Snyder, SH, 1992. Dušikov oksid: fiziološki mediator erekcije penisa. Znanost 257, 401-403. Baskerville, TA, Douglas, AJ, 2008. Interakcija med dopaminom in oksitocinom pri nadzoru spolnega vedenja. Prog. Brain Res. 170, 277-289. Baskerville, TA, Allard, J., Wayman, C., Douglas, AJ, 2009. Interakcija dopamin-oksitocin pri erekciji penisa. E. J. Neurosci. 30, 2151-2164. Benelli, A., Bertolini, A., Poggioli, R., Cavazzutti, E., Calzà, L., Giardino, L., Arletti, R., 1995. Dušikov oksid je vključen v moško spolno vedenje podgan. E. J. Farmakol. 294, 505-510. Bernabè, J., Rampin, O., Sachs, BD, Giuliano, F., 1999. Intraverzalen tlak med erekcijo na podganah: integrativni pristop, ki temelji na telemetričnem snemanju. Am. J. Fiziol. 276, R441-R449. Bitner, RS, Nikkel, AL, Otte, S., Martino, B., Barlow, EH, Bhatia, P., Stewart, AO, Brioni, JD, Decker, MW, Moreland, RB, 2006. Signalizacija receptorja dopamina D4 v paraventrikularnem hipotalamičnem jedru pri podganah: dokazi naravnega spajanja, ki vključujejo takojšnjo zgodnjo indukcijo genov in fosforilacijo proteina kinaze, aktivirane z mitogenom. Nevrofarmakologija 50, 521-531. Brioni, JD, Moreland, RB, Cowart, M., Hsieh, GC, Stewart, AO, Hedlund, P., Donnelly-Roberts, DL, Nakane, M., Lynch 3rd., J., Kolasa, T., Polakowski , JS, Osinski, MA, Marsh, K., Andersson, KE, Sullivan, JP, 2004. Aktivacija receptorjev dopamina D4 s pomočjo ABT-724 povzroči erekcijo penisa pri podganah. Proc. Natl. Acad. Sci. ZDA 101, 6758-6763. Buijs, RM, 1978. Intra- in ekstra-hipotalamični poti vazopresina in oksitocina pri podganah. Cell Tissue Res. 192, 423-435. Buijs, RM, Geffard, M., Pool, CW, Hoorneman, EMD, 1984. Dopaminergična inervacija supraoptičnega in paraventrikularnega jedra. Lahka in elektronska mikroskopska študija. Brain Res. 323, 65-72. Caldwell, JD, Prange, AJ, Pedersen, CA, 1986. Oksitocin olajša spolno dovzetnost samic podgan, ki se zdravijo z estrogenom. Nevropeptidi 7, 175-189. Cameron, JL, Pomerantz, SM, Layden, LM, Amico, JA, 1992. Dopaminergična stimulacija koncentracij oksitocina v plazmi moških in samic opic z apomorfinom in agonistom D2 receptorjev. J. Clin. Endokrinol. Metab. 75, 855-860. Canteras, NS, Simerly, RB, Swanson, LW, 1995. Organizacija projekcij iz medialnega jedra amigdale: študija PHAL na podganah. J. Comp. Nevrol. 360, 213-245. Carmichael, MS, Humbert, R., Dixen, J., Palmisano, G., Greeleaf, W., Davidson, JM, 1987. Oksitocin v plazmi se poveča pri spolnem odzivu ljudi. J. Clin. Endokrinol. Metab. 64, 27-31. Carter, CS, 1992. Oksitocin in spolno vedenje. Neurosci. Biobehav. Rev. 16, 131-144. Carter, CS, Lederhendler, II, Kirkpatrick, B., 1997. Interaktivna nevrobiologija pripadnosti, Annals Newyorške akademije znanosti, vol. 807. Newyorška akademija znanosti, New York. Castelli, poslanec, Piras, AP, Melis, T., Succu, S., Sanna, F., Melis, MR, Collu, S., Ennas, MG, Diaz, G., Mackie, K., Argiolas, A. , 2007. Kanabinoidni CB1 receptorji v paraventrikularnem jedru in centralna kontrola erekcije penisa: imunocitokemija, avtoradiografija in vedenjske študije. Nevroznanost 147, 197-206. Chen, KK, Chang, LS, 2003. Vpliv agonistov ekscitacijskih aminokislin na erekcijo penisa po aplikaciji v paraventrikularno jedro hipotalamusa pri podganah. J. Urol. 62, 575-580. Chen, KK, Chan, JYH, Chang, LS, Chen, MT, Chang, SHH, 1992. Izločanje erekcije penisa po aktivaciji hipokampalne formacije pri podganah. Neurosci. Lett. 141, 218-222. Chen, KK, Chan, JYH, Chang, LS, 1999. Dopaminergična nevrotransmisija v paraventrikularnem jedru hipotalamusa pri centralni regulaciji erekcije penisa pri podganah. J. Urol. 162, 237-242. Collins, GT, Truccone, A., Haji-Abdi, F., Newman, AH, Grundt, P., Rice, KC, Husbands, SM, Greedy, BM, Enguehard-Gueiffer, C., Gueiffer, A., Chen , J., Wang, S., Katz, JL, Grandy, DK, Sunahara, RK, Woods, JH, 2009. Proerektilne učinke dopaminD2 podobnih agonistov posreduje receptor D3 pri podganah in miših. J. Farmakol. Exp. Ther. 329, 210-217. Coolen, LM, Allard, J., Truitt, WA, McKenna, KE, 2004. Centralna ureditev ejakulacije. Fiziol. Behav. 83, 203-215. Cui, SS, Bowen, RC, Gu, GB, Hannesson, BK, Yu, PH, Zhang, X., 2001. Preprečevanje sindroma odtegnitve kanabinoidov z litijem: vključitev oksitocinergične nevronske aktivacije. J. Neurosci. 21, 9867-9876. Dahlstrom, A., Fuxe, K., 1964. Dokazi za obstoj nevronov, ki vsebujejo monoamin, v centralnem živčnem sistemu. I. Dokaz monoaminov v celičnih telesih nevronov možganskega debla. Acta Physiol. Skand. 62 (Suppl. 232), 1-54. Depoortère, R., Bardin, L., Rodrigues, M., Abrial, E., Aliaga, M., Newman-Tancredi, A., 2009. Erekcija in zehanje penisa, ki jo pri podganah povzročajo dopaminski receptorji, podobni D2: vpliv seva in prispevek dopaminskih D2, ne pa receptorjev D3 in D4. Behav. Farmakol. 20, 303-311. Domes, G., Heinrichs, M., Buchel, C., Braus, DF, Herpertz, SC, 2007. Oksitocin zmanjšuje amigdalane odzive na čustvene obraze ne glede na valenco. Biol. Psihiatrija 62, 11871190. Dominguez, J., Riolo, JV, Xu, Z., Hull, ME, 2001. Uravnavanje medialne amigdale kopulacije in medialnega predoptičnega sproščanja dopamina. J. Neurosci. 21, 349-355. Donaldson, ZR, Young, LJ, 2009. Oksitocin, vazopresin in nevrogenetika socialnosti. Znanost 322, 900-904. Eaton, RG, Markowski, VF, Lumley, LA, Thompson, JT, Moses, J., Hull, EM, 1991. D2 receptorji v paraventrikularnem jedru uravnavajo genitalne odzive in kopulacijo pri samcih podgan. Farmakol. Biochem. Behav. 39, 177-181. Ebner, K., Bosch, UL, Krömer, SA, Singewald, N., Neumann, ID, 2005. Sprostitev oksitocina v osrednji amigdali pri podganah modulira obvladovanje stresa in sproščanje vzbujajočih aminokislin. Neuropsychopharmacology 30, 223-230. Enguehard-Gueiffier, C., Hübner, H., El Hakmaoui, A., Allouchi, H., Gmeiner, P., Argiolas, A., Melis, MR, Gueiffier, A., 2006. 2 - [(4-fenilpiperazin-1-il) metil] imidazo (di) azini kot selektivni D4-ligandi. Indukcija erekcije penisa z 2- [4- (2-metoksifenil) piperazin-1-ilmetil] imidazo [1,2-a] piridinom (PIP3EA), močnim in selektivnim agonistom D4. J. Med. Chem. 49, 3938-3947. Everitt, BJ, 1990. Spolna motivacija: nevronska in vedenjska analiza mehanizmov, na katerih temeljijo apetitni in kopulacijski odzivi samcev podgan. Neurosci. Biobehav. Rev. 14, 217-232. Fibiger, HC, Phillips, AG, 1988. Mezokortikolimimski dopaminski sistem in nagrada. Ann. N. Y. Acad. Sci. 5, 206-215. Francoščina, SJ, Totterdell, S., 2003. Pri podganah prejemajo posamezne nukleon-projekcijske nevrone bazolateralno amigdalo in ventralni subtikularni aferent. Nevroznanost 119, 19-31. 954 MR Melis, A. Argiolas / Neuroscience and Biobehavioral Reviews 35 (2011) 939-955 Freund-Mercier, MJ, Richard, P., 1981. Vzbujajoči učinki intraventrikularnih injekcij oksitocina na refleks izmetanja mleka pri podganah. Neurosci. Lett. 23, 193-198. Freund-Mercier, MJ, Richard, P., 1984. Elektrofiziološki dokazi za olajšanje nadzora oksitocinskih nevronov z oksitocinom med dojenjem pri podganah. J. Fiziol. (Lond.) 352, 447-466. Freund-Mercier, MJ, Stoeckel, ME, 1995. Somatodendritični avtoreceptorji na nevronih oksitocina. V: Ivell, R., Russel, JA (Ur.), Oksitocin, celični in molekularni pristopi v medicini in raziskavah. Adv. Exp. Med. Biol., 365. Plenum Press, New York in London, pp. 185-194. Freund-Mercier, MJ, Stoeckel, ME, Palacios, JM, Pazos, JM, Richard, PH, Porte, A., 1987. Farmakološke značilnosti in anatomska porazdelitev krajev vezave 3H oksitocina v možganih podgan Wistar, preučenih z avtoradiografijo. Nevroznanost 20, 599-614. Giuliano, F., Rampin, O., 2000. Centralna kontrola erekcije penisa. Neurosci. Biobehav. Rev. 24, 517-533. Giuliano, F., Allard, J., 2001. Dopamin in spolna funkcija. Int. J. Impotenca Res. 13 (Suppl. 3), 18-28. Giuliano, F., Rampin, O., 2004. Nevronska kontrola erekcije. Fiziol. Behav. 83, 189-201. Giuliano, F., Bernabè, J., McKenna, KE, Longueville, F., Rampin, O., 2001. Spinalni proerectilni učinek oksitocina pri anesteziranih podganah. Am J. Fiziol. Regul. Integ. Comp. Fiziol. 280, R1870-R1877. Goto, Y., Grace, AA, 2005. Dopaminergična modulacija limbičnega in kortikalnega nagona jedra v ciljno usmerjenem vedenju. Nat. Neurosci. 8, 805-812. Hawthorn, J., Ang, VT, Jenkins, JS, 1985. Učinki lezij v hipotalamičnih paraventrikularnih, supraoptičnih in suprahijazmatičnih jedrih na vazopresin in oksitocin v možganih in hrbtenjači podgane. Brain Res. 346, 51-57. Heier, RF, Dolak, LA, Duncan, JN, Hyslop, DK, Lipton, MF, Martin, LJ, Mauragis, MA, Piercey, MF, Nichols, NF, Schreur, PJ, Smith, MW, Moon, MW, 1997. Sinteza in biološke aktivnosti (R) -5,6-dihidro-N, N-dimetil-4H-imidazo [4,5,1-ij] kinolin-5-amina) in njegovih presnovkov. J. Med. Chem. 40, 639-646. Hsieh, GC, Hollingsworth, PR, Martino, B., Chang, R., Terranova, MA, O'Neill, AB, Lynch, JJ, Moreland, RB, Donnelly-Roberts, DL, Kolasa, T., Mikusa, JP , McVey, JM, Marsh, KC, Sullivan, JP, Brioni, JD, 2004. Osrednji mehanizmi za uravnavanje erekcije penisa pri zavestnih podganah: dopaminergični sistemi, povezani s proerectilnim učinkom apomorfina. J. Farmakol. Exp. Ther. 308, 330-338. Huang, PL, Dawson, TM, Bredt, DS, Snyder, SH, Fishman, MC, 1993. Ciljna motnja gena nevronskega dušikovega oksida sintaze. Celica 75, 1273-1286. Huber, D., Veinante, P., Stoop, R., 2005. Vasopresin in oksitocin vzbujata izrazito populacijo nevronov v osrednji amigdali. Znanost 308, 245-248. Hull, EM, Warner, RK, Bazzett, TJ, Eaton, RC, Thompson, JT, 1989. Razmerje D2 / D1 v območju medialnega predoptika vpliva na kopulacijo samcev podgan. J. Farmakol. Exp. Ther. 251, 422-427. Hull, EM, Du, J., Lorrain, DS, Matuszewich, L., 1995. Izvencelični dopamin v medialnem predoptičnem območju: posledice za spolno motivacijo in hormonsko kontrolo kopulacije. J. Neurosci. 15, 7465-7471. Hull, EM, Meisel, RL, Sachs, BD, 2002. Moško spolno vedenje. V: Pfaff, DW, Arnold, AP, Etgen, AM, Fahrbach, SE, Rubin, RT (Ur.), Hormoni, možgani in vedenje. Academic Press, New York, pp. 3-137. Hurlemann, R., Patin, A., Onur, OA, Cohen, MX, Baumgartner, T., Metzler, S., Dziobek, I., Gallinat, J., Wagner, M., Maier, W., Kendrick, KM, 2010. Oksitocin povečuje amigdala, ki je odvisen od socialne ojačitve učenja in čustvene empatije pri ljudeh. J. Neurosci. 30, 4999-5007. Ivell, R., Russel, JA, 1995. Oksitocin: celični in molekularni pristopi v medicini in raziskavah. Napredek eksperimentalne medicine in biologije, vol. 395. Plenum Press, New York. Kelley, AE, Domesick, VB, 1982. Razporeditev projekcije od tvorbe hipokampa do jedra v podganah: študija o anterogradni in retrogradni hrenovi peroksidazi. Nevroznanost 7, 2321-2335. Kimura, Y., Naitou, Y., Wanibuchi, F., Yamaguchi, T., 2008. Aktivacija receptorjev 5-HT (2C) je pogost mehanizem na proerectilne učinke apomorfina, oksitocina in melanotana-II pri podganah. E. J. Farmakol. 589, 157-162. Kondo, Y., Sachs, BD, Sakuma, Y., 1998. Pomen medialne amigdale pri erekciji penisa pri podganah, ki jo povzročajo oddaljeni dražljaji iz estrostičnih samic. Behav. Brain Res. 91, 215-222. Kovacs, GL, Sarnyai, Z., Szabo, G., 1998. Oksitocin in odvisnost: pregled. Psihoneuroendokrinologija 23, 945-962. Lee, HJ, Macbeth, AH, Pagani, JH, Scott Young 3rd, W., 2009. Oksitocin: velik olajševalec življenja. Prog. Neurobiol. 88, 127-151. Lindvall, O., Bjorklund, A., Skagerberg, G., 1984. Selektivna istokemična demonstracija terminalnih sistemov dopamina v di- in telencefalonu podgan: novi dokazi za dopaminergično inervacijo hipotalamičnih nevrosekretornih jeder. Brain Res. 306, 19-30. Liu, YC, Salamone, JD, Sachs, BD, 1997. Moten spolni odziv po poškodbah paraventrikularnega jedra hipotalamusa pri samcih podgan. Behav. Neurosci. 111, 1361-1367. Löber, S., Tschammer, N., Hübner, H., Melis, MR, Argiolas, A., Gmeiner, P., 2009. Okvir azulena kot nov bioizoster: zasnova močnih dopaminskih receptorjev D4 receptorjev, ki povzročajo erekcijo penisa. Chem. Med. Chem. 4, 325-328. McCleskey, EW, Fox, AP, Feldman, DH, Cruz, LJ, Olivera, BM, Tsien, RW, Yoshikami, D., 1987. _-Konotoksin: neposredna in vztrajna blokada posebnih vrst kalcijevih kanalov v nevronih, vendar ne v mišicah. Prot. Nat. Acad. Sci. ZDA 84, 4327-4331. McKenna, KE, 2000. Nekaj predlogov glede organizacije centralnega živčnega sistema nad erekcijo penisa. Neurosci. Biobehav. Rev. 24, 535-540. Marson, L., McKenna, KE, 1992. Vloga 5-hidroksitriptamina pri zmanjševanju spinalnih spolnih refleksov. Exp. Brain Res. 88, 313-318. Marson, L., McKenna, KE, 1996. Censke skupine CNS, vključene v nadzor nad mišicami ischiocavernosusa in bulbospongiosusa: transnevronalna sledilna študija z uporabo psevdorabies virusa. J. Comp. Nevrol. 374, 161-179. Meisel, RL, Sachs, BD, 1994. Fiziologija moškega spolnega vedenja. V: Knobil, E., Neil, J. (Ur.), Fiziologija reprodukcije, vol. 2, druga izdaja. Raven Press, New York, pp. 3-96. Melin, P., Kihlstrom, JE, 1963. Vpliv oksitocina na spolno vedenje pri samcih zajcev. Endokrinologija 73, 433-435. Melis, MR, Argiolas, A., 1995a. Dopamin in spolno vedenje. Neurosci. Biobehav. Rev. 19, 19-38. Melis, MR, Argiolas, A., 1995b. Darovalci dušikovega oksida povzročajo erekcijo in zehanje penisa, če ga injicirajo v centralni živčni sistem samcev podgan. E. J. Farmakol. 294, 1-9. Melis, MR, Argiolas, A., 2003. Osrednji oksitocinergični nevrotransmisija: tarča zdravil za zdravljenje psikogene erektilne disfunkcije. Curr Cilji glede drog 4, 55-66. Melis, MR, Argiolas, A., Gessa, GL, 1986. Oksitocin, ki ga povzročajo zehanje in erekcija penisa: mesto delovanja v možganih. Brain Res. 398, 259-265. Melis, MR, Argiolas, A., Gessa, GL, 1987. Apomorfin, ki ga povzročajo zehanje in erekcija penisa: mesto delovanja v možganih. Brain Res. 415, 98-104. Melis, MR, Argiolas, A., Gessa, GL, 1989a. Apomorfin poveča koncentracijo oksitocina v plazmi pri podganah. Neurosci. Lett. 98, 351-355. Melis, MR, Argiolas, A., Gessa, GL, 1989b. Dokazi, da apomorfin sproži erekcijo in zehanje penisa, tako da sprošča oksitocin v centralnem živčnem sistemu. E. J. Phamacol. 164, 565-570. Melis, MR, Argiolas, A., Stancampiano, R., Gessa, GL, 1990. Vpliv apomorfina na koncentracijo oksitocina v različnih možganskih območjih in plazmi samcev podgan. E. J. Farmakol. 182, 101-107. Melis, MR, Mauri, A., Argiolas, A., 1994a. Erekcija penisov, povzročena z apomorfinom in oksitocinom, in zehanje pri nepoškodovanih in kastriranih samcih podgan: učinek spolnih steroidov. Nevroendokrinologija 59, 349-354. Melis, MR, Stancampiano, R., Argiolas, A., 1994b. Erekcijo in zehanje penisa, ki jo povzročajo paraventrikularne injekcije NMDA, posreduje oksitocin. Farmakol. Biochem. Behav. 48, 203-207. Melis, MR, Stancampiano, R., Argiolas, A., 1994c. Preprečevanje z NG-nitro-l-arginin metilnim estrom erekcije in zehanja, ki jo povzroča apomorfin in oksitocin: mesto delovanja v možganih. Farmakol. Biochem. Behav. 48, 799-804. Melis, MR, Succu, S., Argiolas, A., 1996. Agonisti dopamina povečajo proizvodnjo dušikovega oksida v paraventrikularnem jedru hipotalamusa: korelacija z erekcijo penisa in zehanje. E. J. Neurosci. 8, 2056-2063. Melis, MR, Succu, S., Iannucci, U., Argiolas, A., 1997. Oksitocin poveča proizvodnjo dušikovega oksida v paraventrikularnem jedru hipotalamusa: korelacija z erekcijo penisa in zehanje. Reg. Peptidi 69, 105-112. Melis, MR, Succu, S., Mauri, A., Argiolas, A., 1998. Proizvodnja dušikovega oksida se poveča v paraventrikularnem jedru hipotalamusa samcev podgan med brezkontaktnimi penicami in kopulacijo. E. J. Neurosci. 10, 1968-1974. Melis, MR, Spano, MS, Succu, S., Argiolas, A., 1999a. Antagonist oksitocina d (CH2) 5Tyr (Me) 2-Orn8-vazotocin zmanjšuje brezkontaktne erekcije penisa pri samcih podgan. Neurosci. Lett. 265, 171-174. Melis, MR, Succu, S., Spano, MS, Argiolas, A., 1999b. Morfin, ki ga injiciramo v paraventrikularno jedro hipotalamusa, preprečuje brezkontaktno erekcijo in poslabša kopulacijo: vključevanje dušikovega oksida. E. J. Neurosci. 11, 1857-1864. Melis, MR, Spano, MS, Succu, S., Argiolas, A., 2000. Vpliv ekscitacijskih aminokislin, dopamina in antagonistov receptorjev za oksitocin na nekontaktno erekcijo penisa in proizvodnjo paraventrikularnega dušikovega oksida pri samcih podgan. Behav. Neurosci. 114, 849-857. Melis, MR, Succu, S., Mascia, MS, Cortis, L., Argiolas, A., 2003. Izvencelični dopamin med spolno aktivnostjo narašča v paraventrikularnem jedru samcev podgan. E. J. Neurosci. 17, 1266-1272. Melis, MR, Succu, S., Mascia, MS, Argiolas, A., 2004a. Antagonizem kanabinoidnih CB1receptorjev v paraventrikularnem jedru samcev podgan sproži erekcijo penisa. Neurosci. Lett. 359, 17-20. Melis, MR, Succu, S., Mascia, MS, Cortis, L., Argiolas, A., 2004b. Zunajcelične ekscitatorne aminokisline se povečajo v paraventrikularnem jedru samcev podgan med spolno aktivnostjo: glavna vloga receptorjev NMDA pri erektilni funkciji. E. J. Neurosci. 19, 2569-2575. Melis, MR, Succu, S., Mascia, MS, Argiolas, A., 2005. PD-168,077, selektivni agonist receptorjev D4 za dopamin, povzroči erekcijo penisa, kadar ga injiciramo v paraventrikularno jedro samcev podgan. Neurosci. Lett. 379, 59-62. Melis, MR, Succu, S., Mascia, MS, Sanna, F., Melis, T., Succu, S., Castelli, MP, Argiolas, A., 2006a. Erise penisov, povzročene s SR 141716A, pri samcih podgan: vključevanje paraventrikularne glutaminske kisline in dušikovega oksida. Nevrofarmakologija 50, 219-228. Melis, MR, Succu, S., Sanna, F., Mascia, MS, Melis, T., Enguehard-Gueiffier, C., Hubner, H., Gmenier, P., Gueiffier, A., Argiolas, A., 2006b. PIP3EA in PD168077, dva selektivna agonista receptorja D4 za dopamin, pri moških podganah sprožijo erekcijo penisa: mesto in mehanizem delovanja v možganih. E. J. Neurosci. 24, 2021-2030. Melis, MR, Melis, T., Cocco, C., Succu, S., Sanna, F., Pillolla, G., Boi, A., Ferri, GL, Argiolas, A., 2007. Oksitocin, ki ga injiciramo v ventralno tegmentalno območje, povzroči erekcijo penisa in poveča zunajcelični dopamin v jedru. Melis, A. Argiolas / Neuroscience and Biobehavioral Reviews 35 (2011) 939-955 955 in paraventrikularno jedro hipotalamusa samcev podgan. E. J. Neurosci. 26, 1026-1035. Melis, MR, Sanna, F., Succu, S., Zarone, P., Boi, A., Argiolas, A., 2009a. Vloga oksitocina v predhodni in potrošniški fazi spolnega vedenja samcev. V: Jastrow, H., Feuerbach, D. (Ur.), Priročnik za raziskave oksitocina: sinteza, shranjevanje in sproščanje, ukrepi in oblike zdravil. Nova Publishers Inc, New York, ZDA, pp. 109-125. Melis, MR, Succu, S., Sanna, F., Boi, A., Argiolas, A., 2009b. Oksitocin, ki se injicira v ventralni subkulum ali posteromedialno kortikalno jedro amigdale, povzroči erekcijo penisa in poveča zunajcelični dopamin v jedru podgan samcev podgan. E. J. Neurosci. 30, 1349-1357. Melis, MR, Succu, S., Cocco, C., Caboni, E., Sanna, F., Boi, A., Ferri, GL, Argiolas, A., 2010. Oksitocin povzroči erekcijo penisa, če ga injiciramo v ventralni subiculum: vloga dušikovega oksida in glutaminske kisline. Nevrofarmakologija 58, 1153-1160. Moreland, RB, Nakane, M., Donnelly-Roberts, DL, Miller, LN, Chang, R., Uchic, ME, Terranova, MA, Gubbins, EJ, Helfrich, RJ, Namovic, MT, El-Kouhen, OF, Masters, JN, Brioni, JD, 2004. Primerjalna farmakologija človeških dopaminskih D (2) podobnih celičnih linij, podobnih receptorjem, povezana s kalcijevim tokom skozi Galpha (qo5). Biochem. Farmakol. 68, 761-772. Moos, F., Freund-Mercier, MJ, Guerne, Y., Guerne, JM, Stoeckel, ME, Richard, P., 1984. Sprostitev oksitocina in vazopresina s pomočjo magnoceličnih jeder in vitro: poseben olajševalni učinek oksitocina na njegovo sproščanje. J. Endokrinol. 102, 63-72. Murphy, MR, Seckl, JR, Burton, S., Checkley, SA, Lightman, SL, 1987. Spremembe izločanja oksitocina in vazopresina med spolno aktivnostjo pri moških. J. Clin. Endokrinol. Metab. 65, 738-741. Nishimori, K., Young, LJ, Guo, Q., Wang, Z., Insel, TR, Matzuk, MM, 1996. Oksitocin je potreben za nego, vendar ni nujno potreben za porod ali reproduktivno vedenje. Proc. Natl. Acad. Sci. ZDA 93, 11699-11704. Patel, S., Freedman, S., Chapman, KL, Emms, F., Fletcher, AE, Knowles, M., Marwood, R., Mccallister, G., Myers, J., Curtis, J., Kulagowski, JJ, Leeson, PD, Ridgill, M., Graham, M., Matheson, S., Rathbone, D., Watt, AP, Bristow, LJ, Rupniak, NM, Baskin, E., Lynch, JJ, Ragan, CI , 1997. Biološki profil L 745,870, selektivnega antagonista z visoko afiniteto do receptorja D4 za dopamin. J. Farmakol. Exp. Ther. 283, 636-647. Pedersen, CA, Caldwell, JD, Jirikowski, GF, Insel, TR, 1992. Oksitocin v materinskem, spolnem in družbenem vedenju, Anali New York Academy of Sciences, vol. 652. Newyorška akademija znanosti, New York. Petrovic, P., Kalisch, R., Singer, T., Dolan, RJ, 2008. Oksitocin zmanjšuje afektivne ocene kondicijskih obrazov in aktivnost amigdale. J. Neurosci. 28, 6607-6615. Pfaus, JG, Everitt, BJ, 1995. Psihofarmakologija spolnega vedenja. V: Knobil, FE, Kupfer, DJ (Ur.), Psihoparmakologija: četrta generacija napredka. Raven Press, New York, pp. 742-758. Rajfer, J., Aronson, WJ, Bush, PA, Dorey, FJ, Ignarro, LJ, 1992. Dušikov oksid kot mediator sprostitve kavernozusa telesa kot odgovor na nenadrenergično, nehlinergično nevrotransmisijo. N. Engl. J. Med. 326, 90-94. Roeling, TAP, Van Erp, AMM, Meelis, W., Kruk, MR, Veening, JG, 1991. Vedenjski učinki NMDA, ki se injicirajo v hipotalamično paraventrikularno jedro podgane. Brain Res. 550, 220-224. Sachs, BD, 1997. Erekcijo pri moških podganah povzroča vonj po zraku od strašnih samic. Fiziol. Behav. 62, 921-924. Sachs, BD, 2000. Kontekstualni pristopi k fiziologiji in klasifikaciji erektilne funkcije, erektilne disfunkcije in spolnega vzburjenja. Neurosci. Biobehav. Rev. 24, 541-560. Sachs, BD, 2007. Kontekstualna definicija moškega spolnega vzburjenja. Horm. Behav. 51, 569-578. Sanchez, F., Alonso, JR, Arevalo, R., Blanco, E., Aijon, J., Vazquez, R., 1994. Soobstoj NADPH-diforaze z vazopresinom in oksitocinom v hipotalamičnih magnocelularnih nevrosekretornih jedrih podgane. Cell Tissue Res. 276, 31-34. Sanna, F., Succu, S., Boi, A., Melis, MR, Argiolas, A., 2009. Zaviralci 5 tipa fosfodiesteraze olajšajo brezkontaktno erekcijo pri samcih podgan: mesto delovanja v možganih in mehanizem delovanja. J. Seks. Med. 6, 2680-2689. Saphier, D., Feldman, S., 1987. Učinki septalnih in hipokampalnih dražljajev na nevrone paraventrikularnega jedra. Nevroznanost 20, 749-755. Sato-Suzuki, I., Kita, I., Oguri, M., Arita, H., 1998. Stereotipni zevajoči odzivi, ki jih povzročajo električna in kemična stimulacija paraventrikularnega jedra podgane. J. Nevrofiziol. 80, 2765-2775. Schuman, EM, Madison, DV, 1994. Dušikov oksid in sinaptična funkcija. Ann. Rev. Neurosci. 17, 153-183. Snyder, SH, 1992. Dušikov oksid: prvi v novem razredu nevrotransmiterjev? Znanost 254, 494-496. Sofroniew, MV, 1983. Vasopresin in oksitocin v možganih in hrbtenjači sesalcev. Trendi Neurosci. 6, 467-472. Sokoloff, P., Schwartz, JC, 1995. Pol desetletja pozneje so novi dopaminski receptorji. Trendi Pharmacol. Sci. 16, 270-275. Southam, E., Garthwaite, J., 1993. Cestna signalizacija GMP dušikovega oksida v možganih podgan. Nevrofarmakologija 32, 1267-1277. Stancampiano, R., Melis, MR, Argiolas, A., 1994. Erekcija in zevanje penisa, ki jo povzročajo agonisti 5-HT1c pri samcih podgan: odnos z dopaminergičnim in oksitocinergičnim prenosom. E. J. Farmakol. 261, 149-155. Succu, S., Mascia, MS, Sanna, F., Melis, T., Argiolas, A., Melis, MR, 2006. Kanabinoidni antagonist receptorjev CB1 SR 141716A povzroči erekcijo penisa s povečanjem zunajcelične glutaminske kisline v paraventrikularnem jedru samcev podgan. Behav. Brain Res. 169, 274-281. Succu, S., Sanna, F., Melis, T., Boi, A., Argiolas, A., Melis, MR, 2007. Stimulacija dopaminskih receptorjev v paraventrikularnem jedru hipotalamusa samcev podgan povzroči erekcijo penisa in poveča zunajcelični dopamin v okoliščinah jedra: vključitev osrednjega oksitocina. Nevrofarmakologija 52, 1034-1043. Succu, S., Sanna, F., Cocco, C., Melis, T., Boi, A., Ferri, GL, Argiolas, A., Melis, MR, 2008. Oksitocin povzroči erekcijo penisa, če ga injiciramo v ventralno tegmentalno območje samcev podgan: vloga dušikovega oksida in ciklični GMP. E. J. Neurosci. 28, 813-821. Tanda, G., Pontieri, FE, Di Chiara, G., 1997. Kanabinoidi in heroin aktiviranje mezolimbičnega prenosa dopamina s skupnim mehanizmom opioidnih receptorjev mu1. Znanost 276, 2048-2050. Tang, Y., Rampin, O., Calas, A., Facchinetti, P., Giuliano, F., 1998. Oksitocinergična in serotoninergična inervacija identificiranih lumbosakralnih jeder, ki nadzirajo erekcijo penisa pri samcu podgane. Nevroznanost 82, 241-254. Teodoza, DT, 1985. Oksitocin-imunoreaktivni terminali sinapsijo na nevronih oksitocina v supraoptičnih jedrih. Narava (London) 313, 682-684. Tindall, JS, 1974. Stimuli, ki povzročajo sproščanje oksitocina. V: Geiger, SR, Knobil, E., Sawyer, WH, Greef, R., Astwood, EB (Ur.), Priročnik za fiziologijo. Sekta. 7, Endokrinologija, vol. IV. Ameriško fiziološko društvo, Washington DC, pp. 257-267. Torres, G., Lee, S., Rivier, C., 1993. Ontogenija sinteze dušikovega oksida podgane in kolokalizacija z nevropeptidi. Mol. Celica. Neurosci. 4, 155-163. Uhl-Bronner, S., Waltisperger, E., Martinez-Lorenzana, G., Condes, LM, Freund-Mercier, MJ, 2005. Spolno dimorfno izražanje mest vezave oksitocina v sprednjem mozgu in hrbtenjači podgane. Nevroznanost 135, 147-154. Vaccari, C., Lolait, SJ, Ostrowski, NL, 1998. Primerjalna porazdelitev ribonukleinskih kislin vazopresina V1b in oksitocinskih receptorjev v možganih. Endokrinologija 139, 5015-5033. Van Den Pol, A., 1991. Imunoreaktivnost glutamata in aspartata v hipotalamičnih presinaptičnih aksonih. J. Neurosci. 11, 2087-2101. Veronneau-Longueville, F., Rampin, O., Freund-Mercier, MJ, Tang, Y., Calas, A., Marson, L., McKenna, KE, Stoeckel, ME, Benoit, G., Giuliano, F. , 1999. Oksitocinergična inervacija avtonomnih jeder, ki nadzorujejo erekcijo penisa pri podganah. Nevroznanost 93, 1437-1447. Vincent, SR, Kimura, H., 1992. Histokemično kartiranje sintaze dušikovega oksida v možganih podgane. Nevroznanost 46, 755-784. Wagner, CK, Clemens, LG, 1993. Pot, ki vsebuje nevrofizin, od paraventrikularnega jedra hipotalamusa do spolno dimorfnega motoričnega jedra v ledvenem delu hrbtenjače. J. Comp. Nevrol. 336, 106-116. Winslow, JT, Insel, TR, 1991. Socialni status pri parih opic moških veveric določa vedenjski odziv na centralno dajanje oksitocina. J. Neurosci. 11, 2032-2038. Wise, RA, Rompre, P.-P., 1989. Bazični dopamin in nagrada. Ann. Rev. Psihola. 40, 191-225. Witt, DM, Insel, TR, 1994. Moško spolno vedenje aktivira c-fos podobno beljakovino v nevronih oksitocina v paraventrikularnem jedru hipotalamusa. J. Nevroendokrinol. 6, 13-18. Witter, MP, 2006. Povezave podprograma podgane: topografija glede na kolonsko in laminarno organizacijo. Behav Brain Res. 174, 251-264. Woodruff, GN, Foster, AC, Gill, R., Kemp, JA, Wong, EH, Iversen, LL, 1987. Medsebojno delovanje med MK-801 in receptorji za N-metil-d-aspartat: funkcionalne posledice. Nevrofarmakologija 26, 903-909. Yamashita, H., Shigeru, O., Inenaga, K., Kasai, M., Uesugi, S., Kannan, H., Kaneko, T., 1987. Oksitocin v in vitro pretežno vzbuja domnevne oksitocinske nevrone v supraoptičnem jedru podgane. Brain Res. 416, 364-368. Yells, DP, Hendricks, SE, Prendergast, MA, 1992. Lezije jedra paragigantocellularis: učinki na parjenje pri samcih podgan. Brain Res. 596, 73-79. Young, WS, Shepard, E., Amico, J., Hennighausen, L., LaMarca, ME, McKinney, C., Ginns, EI, 1996. Pomanjkanje mišjega oksitocina preprečuje izmet mleka, ne pa tudi plodnosti ali poglobitve. J. Nevroendokrinol. 8, 847-854. Zahran, AR, Vachon, P., Courtois, F., Carrier, S., 2000. Povišanje intrakavernoznega tlaka na penisu po injiciranju agonistov ekscitacijskih aminokislin v hipotalamično paraventrikularno jedro anesteziranih podgan. J. Urol.
