Dev Dev 2008; 28(1):-62 77 ดอย: 10.1016 / j.dr.2007.08.003
นามธรรม
วัยรุ่นเป็นช่วงเวลาการพัฒนาที่โดดเด่นด้วยการตัดสินใจและการกระทำที่ไม่เหมาะสมซึ่งก่อให้เกิดอุบัติการณ์ที่เพิ่มขึ้นของการบาดเจ็บและความรุนแรงโดยไม่ได้ตั้งใจการดื่มแอลกอฮอล์และยาเสพติดการตั้งครรภ์โดยไม่ตั้งใจและโรคติดต่อทางเพศสัมพันธ์ คำอธิบายระบบประสาทแบบดั้งเดิมและการรับรู้เกี่ยวกับพฤติกรรมของวัยรุ่นไม่สามารถอธิบายถึงการเปลี่ยนแปลงแบบไม่เชิงเส้นในพฤติกรรมที่สังเกตได้ในช่วงวัยรุ่นที่สัมพันธ์กับวัยเด็กและวัยผู้ใหญ่ การทบทวนนี้ให้แนวคิดที่เป็นไปได้ทางชีวภาพของกลไกประสาทที่มีพื้นฐานการเปลี่ยนแปลงแบบไม่เชิงเส้นในพฤติกรรมดังกล่าวเนื่องจากการตอบสนองต่อแรงจูงใจที่เพิ่มขึ้นในขณะที่การควบคุมแรงกระตุ้นยังคงไม่สมบูรณ์ในช่วงนี้ การถ่ายภาพมนุษย์และการศึกษาสัตว์เมื่อเร็ว ๆ นี้ได้ให้พื้นฐานทางชีววิทยาสำหรับมุมมองนี้แสดงให้เห็นการพัฒนาที่แตกต่างกันของระบบการให้รางวัลแบบลิมบิกเมื่อเทียบกับระบบควบคุมจากบนลงล่างในช่วงวัยรุ่นที่สัมพันธ์กับวัยเด็กและผู้ใหญ่ รูปแบบการพัฒนานี้อาจรุนแรงขึ้นในวัยรุ่นที่มีใจโอนเอียงไปสู่การรับความเสี่ยงเพิ่มความเสี่ยงสำหรับผลลัพธ์ที่ไม่ดี
จากศูนย์สถิติสุขภาพแห่งชาติระบุว่ามีการเสียชีวิตของวัยรุ่น 13,000 ในสหรัฐอเมริกาทุกปี ประมาณ 70% ของการเสียชีวิตเหล่านี้เป็นผลมาจากการชนของยานยนต์การบาดเจ็บโดยไม่เจตนาการฆาตกรรมและการฆ่าตัวตาย (Eaton และคณะ, 2006) ผลจากการสำรวจพฤติกรรมเสี่ยงต่อเยาวชนแห่งชาติ 2005 (YRBS) แสดงให้เห็นว่าวัยรุ่นมีส่วนร่วมในพฤติกรรมที่เพิ่มโอกาสในการเสียชีวิตหรือเจ็บป่วยโดยการขับรถยนต์หลังจากดื่มหรือไม่ใส่เข็มขัดนิรภัยพกพาอาวุธใช้สารที่ผิดกฎหมายและมีเพศสัมพันธ์ที่ไม่มีการป้องกัน ส่งผลให้เกิดการตั้งครรภ์ที่ไม่ตั้งใจและโรคติดต่อทางเพศสัมพันธ์รวมถึงการติดเชื้อเอชไอวี (Eaton และคณะ, 2006) สถิติเหล่านี้เน้นย้ำถึงความสำคัญของการทำความเข้าใจตัวเลือกและการกระทำที่มีความเสี่ยงในวัยรุ่น
จำนวนของสมมติฐานเกี่ยวกับความรู้ความเข้าใจและ neurobiological ได้รับการตั้งสมมติฐานว่าทำไมวัยรุ่นมีส่วนร่วมในพฤติกรรมทางเลือกที่ไม่ดี ในการทบทวนวรรณกรรมเกี่ยวกับการพัฒนาสมองของเด็กวัยรุ่น Yurgelun-Todd (2007) แสดงให้เห็นว่าการพัฒนาความรู้ความเข้าใจในช่วงวัยรุ่นมีความสัมพันธ์กับความสามารถในการควบคุมความรู้ความเข้าใจที่ก้าวหน้ามากขึ้น ประสิทธิภาพนี้ได้รับการอธิบายว่าขึ้นอยู่กับการเจริญเติบโตของเยื่อหุ้มสมอง prefrontal ดังที่เห็นได้จากกิจกรรมที่เพิ่มขึ้นภายในบริเวณ prefrontal โฟกัส (Rubia และคณะ 2000; Tamm, Menon และ Reiss, 2002) และกิจกรรมที่ลดลงในพื้นที่สมองที่ไม่เกี่ยวข้อง (Brown และคณะ, 2005; Durston และคณะ, 2006).
รูปแบบทั่วไปของการปรับปรุงการควบคุมความรู้ความเข้าใจกับการเจริญเติบโตของเยื่อหุ้มสมอง prefrontal แสดงให้เห็นการเพิ่มขึ้นเชิงเส้นในการพัฒนาตั้งแต่วัยเด็กจนถึงวัยผู้ใหญ่ แต่ตัวเลือกและการกระทำที่ไม่ได้ผลที่สังเกตได้ในช่วงวัยรุ่นแสดงให้เห็นถึงการเปลี่ยนแปลงที่ไม่เชิงเส้นในพฤติกรรมที่สามารถแยกความแตกต่างจากวัยเด็กและผู้ใหญ่ตามหลักฐานจากศูนย์สถิติสุขภาพแห่งชาติเกี่ยวกับพฤติกรรมวัยรุ่นและการเสียชีวิต หากการควบคุมความรู้ความเข้าใจและเยื่อหุ้มสมอง prefrontal ที่ยังไม่บรรลุนิติภาวะเป็นพื้นฐานสำหรับพฤติกรรมการเลือกที่ไม่ดีนั้นเด็ก ๆ ควรดูคล้ายกันอย่างน่าทึ่งหรือแย่กว่าวัยรุ่นที่ได้รับเยื่อหุ้มสมองส่วนหน้าและความสามารถในการคิด ดังนั้นฟังก์ชั่น prefrontal ที่ยังไม่บรรลุนิติภาวะเพียงอย่างเดียวไม่สามารถอธิบายพฤติกรรมของวัยรุ่นได้
แนวความคิดที่ถูกต้องของการเปลี่ยนแปลงทางปัญญาและ neurobiological ในช่วงวัยรุ่นจะต้องถือว่าวัยรุ่นเป็นระยะเวลาการพัฒนาในช่วงเปลี่ยนผ่าน (หอก 2000) แทนที่จะเป็นภาพรวมครั้งเดียว (Casey, Tottenham, Liston, & Durston, 2005) กล่าวอีกนัยหนึ่งเพื่อให้เข้าใจช่วงเวลาการพัฒนาการเปลี่ยนเข้าและออกจากวัยรุ่นเป็นสิ่งจำเป็นสำหรับการแยกแยะคุณลักษณะที่แตกต่างของขั้นตอนการพัฒนานี้ การสร้างวิถีการพัฒนาสำหรับกระบวนการทางปัญญาและระบบประสาทเป็นสิ่งจำเป็นในการอธิบายลักษณะการเปลี่ยนผ่านเหล่านี้และการตีความที่ จำกัด เกี่ยวกับการเปลี่ยนแปลงพฤติกรรมในช่วงเวลานี้ ในระดับความรู้ความเข้าใจหรือพฤติกรรมวัยรุ่นมีลักษณะที่หุนหันพลันแล่น (เช่นขาดการควบคุมความรู้ความเข้าใจ) และความเสี่ยงกับการสร้างสิ่งเหล่านี้ใช้ synonymously และไม่ซาบซึ้งสำหรับวิถีการพัฒนาที่แตกต่างกันของแต่ละคน ในระดับ neurobiological การถ่ายภาพมนุษย์และสัตว์ศึกษาแนะนำฐาน neurobiological ที่แตกต่างกันและวิถีการพัฒนาสำหรับระบบประสาทที่รองรับโครงสร้างที่แยกต่างหากเหล่านี้ของการควบคุมแรงกระตุ้นและการตัดสินใจที่มีความเสี่ยง
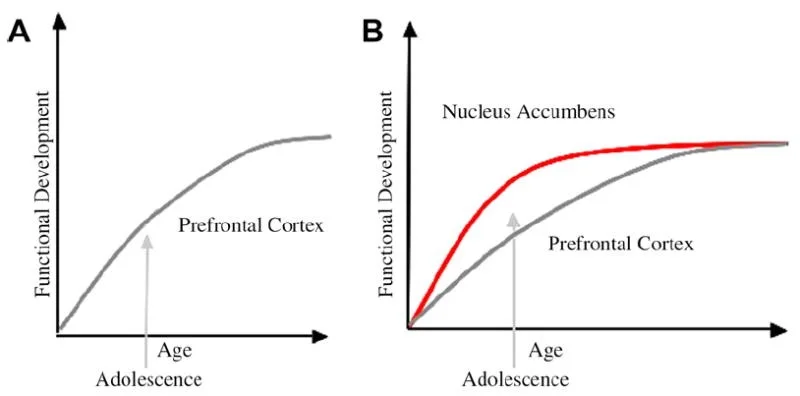
เราได้พัฒนาแบบจำลอง neurobiological ของการพัฒนาวัยรุ่นภายในกรอบนี้ที่สร้างแบบจำลองหนู (Laviola, Adriani, Terranova และ Gerra, 1999; หอก 2000) และการศึกษาการถ่ายภาพล่าสุดของวัยรุ่น (เอินส์ทและอัล, 2005; Galvan, Hare, Voss, Glover และ Casey, 2007; Galvan et al., 2006). มะเดื่อ. 1 ด้านล่างแสดงโมเดลนี้ ทางด้านซ้ายเป็นลักษณะดั้งเดิมของวัยรุ่นที่เกี่ยวข้องกับความไม่บรรลุนิติภาวะของเยื่อหุ้มสมอง prefrontal ด้านขวาเป็นแบบจำลองทางประสาทวิทยาที่เรานำเสนอซึ่งแสดงให้เห็นว่าขอบเขตการควบคุมจากบนลงล่างและแขนขาด้านหน้าและล่างจะต้องพิจารณาร่วมกันอย่างไร การ์ตูนแสดงวิถีการพัฒนาที่แตกต่างกันสำหรับระบบเหล่านี้ด้วยระบบลิมบิกที่พัฒนาเร็วกว่าส่วนควบคุมล่วงหน้า ตามแบบจำลองนี้บุคคลมีความเอนเอียงมากขึ้นจากภูมิภาคลิมบิกที่เป็นผู้ใหญ่ในช่วงวัยรุ่น (กล่าวคือความไม่สมดุลของลิมบิคสัมพันธ์กับการควบคุม prefrontal) เปรียบเทียบกับเด็กซึ่งระบบเหล่านี้ (เช่นลิมบิกและ prefrontal) ยังคงพัฒนา และเปรียบเทียบกับผู้ใหญ่ซึ่งระบบเหล่านี้มีความสมบูรณ์เต็มที่ มุมมองนี้ให้ข้อมูลพื้นฐานสำหรับการเปลี่ยนแปลงแบบไม่เชิงเส้นในพฤติกรรมข้ามการพัฒนาเนื่องจากการเจริญเติบโตก่อนหน้าของ limbic นี้เมื่อเทียบกับภูมิภาคการควบคุม prefrontal จากบนลงล่างที่ไม่เต็มที่ ด้วยการพัฒนาและประสบการณ์การเชื่อมต่อการทำงานระหว่างภูมิภาคเหล่านี้มีกลไกสำหรับการควบคุมจากบนลงล่างของภูมิภาคเหล่านี้ (Hare, Voss, Glover, & Casey, 2007a) นอกจากนี้รูปแบบการปรับความขัดแย้งของสถิติสุขภาพของพฤติกรรมเสี่ยงในช่วงวัยรุ่นด้วยการสังเกตที่ชาญฉลาดโดย Reyna และ Farley (2006) วัยรุ่นที่สามารถให้เหตุผลและเข้าใจความเสี่ยงของพฤติกรรมที่พวกเขามีส่วนร่วม ตามแบบจำลองของเราในสถานการณ์ที่สำคัญทางอารมณ์ระบบลิมบิกจะได้รับชัยชนะเหนือระบบควบคุมเนื่องจากความสมบูรณ์เมื่อเทียบกับระบบควบคุมพรีฟรอน หลักฐานจากการศึกษาเกี่ยวกับพฤติกรรมและการถ่ายภาพของมนุษย์เพื่อสนับสนุนโมเดลนี้มีให้ในบริบทของการกระทำในบริบทของการให้รางวัลและอารมณ์ (Galvan et al., 2006, 2007; Hare, Voss, Glover, & Casey, 2007b; Hare et al., 2007a) นอกจากนี้เรายังคาดการณ์ว่าเหตุใดสมองจึงอาจพัฒนาในลักษณะนี้และทำไมวัยรุ่นบางคนอาจมีความเสี่ยงมากขึ้นสำหรับการตัดสินใจที่ไม่เหมาะสมซึ่งนำไปสู่ผลลัพธ์ระยะยาวที่ไม่ดี (Galvan et al., 2007; Hare et al., 2007b).
คำอธิบายแบบดั้งเดิมของพฤติกรรมวัยรุ่นได้รับการแนะนำให้เป็นเพราะการพัฒนายืดเยื้อของเยื่อหุ้มสมอง prefrontal (A) แบบจำลองของเราคำนึงถึงการพัฒนาของ prefrontal cortex ร่วมกับ subbortical limbic region (เช่นนิวเคลียส accumbens) ซึ่งมีส่วนเกี่ยวข้องในการเลือกและการกระทำที่มีความเสี่ยง (B)
การพัฒนาพฤติกรรมตามเป้าหมาย
รากฐานที่สำคัญของการพัฒนาความรู้ความเข้าใจคือความสามารถในการปราบปรามความคิดและการกระทำที่ไม่เหมาะสมเพื่อสนับสนุนเป้าหมายที่กำหนดโดยเฉพาะอย่างยิ่งเมื่อมีแรงจูงใจที่น่าสนใจ (Casey, Galvan, & Hare, 2005; Casey et al., 2000b; Casey, Thomas, David-son, Kunz และ Franzen, 2002a; เคซีย์ท็อตแนมและฟอสเซลลา 2002b) การศึกษาเชิงพัฒนาการจำนวนมากแสดงให้เห็นว่าความสามารถนี้พัฒนาตลอดวัยเด็กและวัยรุ่นตัวพิมพ์ 1972; Flavell, Feach และ Chinsky, 1966; Keating & Bobbitt, 1978; Pascual-Leone, 1970) นักทฤษฎีหลายคนแย้งว่าการพัฒนาทางปัญญาเกิดจากการเพิ่มความเร็วและประสิทธิภาพในการประมวลผลและไม่ได้เกิดจากความสามารถทางจิตที่เพิ่มขึ้น (เช่น Bjorkland, 1985; Bjorkland, 1987; ตัวพิมพ์ 1985) นักทฤษฎีอื่น ๆ ได้รวมกระบวนการสร้าง“ การยับยั้ง” ไว้ในบัญชีของพวกเขาเกี่ยวกับการพัฒนาความรู้ความเข้าใจ (Harnishfeger & Bjorkland, 1993) ตามบัญชีนี้การรับรู้ที่ยังไม่บรรลุนิติภาวะเป็นลักษณะความไวต่อการรบกวนจากแหล่งแข่งขันที่จะต้องระงับ (เช่น Brainerd & Reyna, 1993; Casey, Thomas, Davidson, Kunz และ Franzen, 2002a; Dempster, 1993; ไดมอนด์ 1985; Munakata & Yerys, 2001) ดังนั้นพฤติกรรมที่มุ่งไปยังเป้าหมายนั้นต้องการการควบคุมแรงกระตุ้นหรือความล่าช้าของความพึงพอใจเพื่อให้ได้ผลลัพธ์ที่ดีที่สุดและความสามารถนี้ดูเหมือนจะเติบโตเต็มที่ในวัยเด็กและวัยรุ่น
พฤติกรรมวัยรุ่นได้รับการอธิบายว่าถูกหุนหันพลันแล่นและมีความเสี่ยงเกือบจะเหมือนกัน แต่สิ่งก่อสร้างเหล่านี้ขึ้นอยู่กับกระบวนการทางความคิดและระบบประสาทที่แตกต่างกันซึ่งแนะนำการสร้างที่แตกต่างด้วยวิถีการพัฒนาที่แตกต่างกัน โดยเฉพาะการทบทวนวรรณกรรมชี้ให้เห็นว่าแรงกระตุ้นลดลงตามอายุในวัยเด็กและวัยรุ่น (เคซี่ย์ และคณะ, 2002ก; Casey, Galvan และคณะ, 2005; Galvan et al., 2007) และมีความเกี่ยวข้องกับการพัฒนาที่ยืดเยื้อของเยื่อหุ้มสมอง prefrontal (Casey, Galvan และคณะ, 2005) แม้ว่าจะมีความแตกต่างในระดับที่บุคคลนั้นได้รับการหุนหันพลันแล่นหรือไม่ก็ตามโดยไม่คำนึงถึงอายุ
ในทางตรงกันข้ามการควบคุมแรงกระตุ้น / ความรู้ความเข้าใจความเสี่ยงปรากฏเพิ่มขึ้นในช่วงวัยรุ่นเมื่อเทียบกับวัยเด็กและวัยผู้ใหญ่และมีความสัมพันธ์กับระบบย่อยที่รู้จักกันว่ามีส่วนร่วมในการประเมินผลรางวัล การศึกษาการถ่ายภาพของมนุษย์ที่จะได้รับการตรวจสอบแนะนำการเพิ่มขึ้นของการเปิดใช้งาน subcortical (เช่น accumbens) เมื่อทำการเลือกที่มีความเสี่ยง (Kuhnen & Knutson, 2005; Matthews & et al., 2004; Montague & Berns, 2002) ที่พูดเกินจริงในวัยรุ่นสัมพันธ์กับเด็กและผู้ใหญ่ (เอินส์ทและอัล, 2005; Galvan et al., 2006) การค้นพบเหล่านี้แนะนำวิถีการทำงานที่แตกต่างกันสำหรับพฤติกรรมที่ได้รับรางวัลหรือเป็นแรงจูงใจโดยมีการพัฒนาระบบเหล่านี้ก่อนหน้านี้เมื่อเทียบกับระบบควบคุมที่แสดงหลักสูตรพัฒนาการที่ยืดเยื้อและเชิงเส้นในแง่ของการเอาชนะทางเลือกและการกระทำที่ไม่เหมาะสม
หลักฐานจากการศึกษา neuroimaging ของการพัฒนาสมองของมนุษย์
การตรวจสอบล่าสุดของการพัฒนาสมองของวัยรุ่นมีพื้นฐานมาจากความก้าวหน้าในวิธีการ neuroimaging ที่สามารถใช้งานได้ง่ายกับการพัฒนาประชากรมนุษย์ วิธีการเหล่านี้ขึ้นอยู่กับวิธีถ่ายภาพด้วยคลื่นสนามแม่เหล็ก (MRI) (ดู มะเดื่อ. 2) และรวมถึง: MRI โครงสร้างซึ่งใช้ในการวัดขนาดและรูปร่างของโครงสร้าง; functional MRI ซึ่งใช้วัดรูปแบบของการทำงานของสมอง และ diffusion tensor imaging (DTI) ซึ่งใช้ในการสร้างดัชนีการเชื่อมต่อของเส้นใยสสารสีขาว หลักฐานสำหรับรูปแบบการพัฒนาของเราในการแข่งขันระหว่างภูมิภาคเยื่อหุ้มสมองและภูมิภาค subcortical ได้รับการสนับสนุนโดยการเชื่อมต่อโครงสร้างที่ยังไม่บรรลุนิติภาวะและการทำงานที่วัดได้โดย DTI และ fMRI ตามลำดับ
MRI ศึกษาการพัฒนาสมองของมนุษย์
การศึกษาจำนวนมากได้ใช้โครงสร้าง MRI เพื่อทำแผนที่กายวิภาคของการพัฒนาสมองปกติ (ดูความคิดเห็นโดย Durston และคณะ, 2001) แม้ว่าขนาดของสมองโดยรวมจะอยู่ที่ประมาณ 90% ของขนาดผู้ใหญ่เมื่ออายุหกขวบ แต่ส่วนประกอบย่อยของสสารสีเทาและสีขาวของสมองยังคงได้รับการเปลี่ยนแปลงแบบไดนามิกตลอดช่วงวัยรุ่น ข้อมูลจากการศึกษา MRI ระยะยาวเมื่อเร็ว ๆ นี้ระบุว่าปริมาณสสารสีเทามีรูปแบบ U-shape คว่ำโดยมีการแปรผันของภูมิภาคมากกว่าสสารขาว (Giedd, 2004; Gogtay และคณะ, 2004; Sowell และคณะ 2003; Sowell, Thompson, & Toga, 2004) โดยทั่วไปภูมิภาคจะลดการทำงานของฟังก์ชั่นหลักเช่นระบบมอเตอร์และระบบประสาท พื้นที่การเชื่อมโยงลำดับสูงขึ้นซึ่งรวมฟังก์ชั่นหลักเหล่านี้ทำให้ครบกำหนดในภายหลัง (Gogtay และคณะ, 2004; Sowell, Thompson, & Toga, 2004) ตัวอย่างเช่นการศึกษาโดยใช้มาตรการที่ใช้ MRI แสดงให้เห็นว่าการสูญเสียสสารเยื่อหุ้มสมองสีเทาเกิดขึ้นเร็วที่สุดในพื้นที่เซ็นเซอร์หลักและล่าสุดใน dorsolateral prefrontal และ cortal ขมับด้านข้าง (Gogtay และคณะ, 2004) รูปแบบนี้สอดคล้องกับการศึกษาเจ้าคณะที่ไม่ใช่มนุษย์และการชันสูตรศพของมนุษย์แสดงให้เห็นว่าเยื่อหุ้มสมอง prefrontal เป็นหนึ่งในพื้นที่สมองสุดท้ายที่จะเติบโต (Bourgeois, Goldman-Rakic, & Rakic, 1994; Huttenlocher, 1979) ตรงกันข้ามกับวัตถุสีเทาปริมาณสสารสีขาวเพิ่มขึ้นในรูปแบบเชิงเส้นคร่าวๆเพิ่มขึ้นตลอดการพัฒนาสู่ความเป็นผู้ใหญ่ (Gogtay และคณะ, 2004) การเปลี่ยนแปลงเหล่านี้น่าจะสะท้อนให้เห็นถึง myelination อย่างต่อเนื่องของซอนโดย oligodendrocytes เสริมสร้างการนำและการสื่อสารของเซลล์ประสาท
แม้ว่าจะได้รับความสนใจน้อยลงในภูมิภาค subcortical เมื่อตรวจสอบการเปลี่ยนแปลงโครงสร้างบางส่วนของการเปลี่ยนแปลงที่ใหญ่ที่สุดในสมองในการพัฒนาจะเห็นในภูมิภาคเหล่านี้โดยเฉพาะอย่างยิ่งในปมประสาทฐาน (Sowell และคณะ, 1999โปรดดูที่ มะเดื่อ. 3) และโดยเฉพาะอย่างยิ่งในเพศชาย (Giedd et al., 1996) การเปลี่ยนแปลงเชิงพัฒนาการของปริมาตรโครงสร้างภายในฐานปมประสาทและบริเวณด้านหน้ามีความน่าสนใจในแง่ของกระบวนการพัฒนาที่เป็นที่รู้จัก (เช่น dendritic arborization, การตายของเซลล์, การตัดแต่ง synaptic, myelination) ที่เกิดขึ้นในช่วงวัยเด็กและวัยรุ่น กระบวนการเหล่านี้ช่วยให้การปรับแต่งและเสริมสร้างความเข้มแข็งของการเชื่อมต่อระหว่างภูมิภาค prefrontal และ subcortical ด้วยการพัฒนาและการเรียนรู้ที่อาจสอดคล้องกับการควบคุมความรู้ความเข้าใจที่มากขึ้น การเปลี่ยนแปลงโครงสร้างเหล่านี้เกี่ยวข้องกับการเปลี่ยนแปลงทางปัญญาอย่างไร มีการศึกษาจำนวนหนึ่งที่เกี่ยวข้องกับการเจริญเติบโตของกลีบสมองส่วนหน้าและการทำงานของสมองโดยใช้มาตรการทางประสาทวิทยาและการคิด (เช่น Sowell และคณะ, 2003) โดยเฉพาะมีการรายงานการเชื่อมโยงระหว่าง MRI-based prefrontal cortical และ basal ganglia ในระดับภูมิภาคและมาตรการของการควบคุมการรับรู้ (เช่นความสามารถในการแทนที่การตอบสนองที่ไม่เหมาะสมในความโปรดปรานของผู้อื่นหรือเพื่อยับยั้งความสนใจต่อคุณลักษณะการกระตุ้นที่ไม่เกี่ยวข้องเคซี่ย์ เทรนเนอร์ และคณะ 1997) การค้นพบเหล่านี้ชี้ให้เห็นว่าการเปลี่ยนแปลงทางปัญญาสะท้อนในการเปลี่ยนแปลงของสมองโครงสร้างและเน้นความสำคัญของ subcortical (basal ganglia) เช่นเดียวกับ cortical (เช่น prefrontal cortex)

การศึกษา DTI เกี่ยวกับการพัฒนาสมองของมนุษย์
การศึกษา morphometry จาก MRI ที่ได้รับการทบทวนชี้ให้เห็นว่าการเชื่อมต่อของเยื่อหุ้มสมองนั้นได้รับการปรับแต่งอย่างดีพร้อมกับการกำจัดการทำงานร่วมกันของความผิดปกติที่มากเกินไปและการเสริมสร้างความเข้มแข็งของการเชื่อมต่อที่เกี่ยวข้อง ความก้าวหน้าล่าสุดของเทคโนโลยี MRI เช่นเดียวกับ DTI เป็นเครื่องมือที่มีศักยภาพในการตรวจสอบบทบาทของสสารสีขาวเฉพาะเพื่อการพัฒนาสมองและพฤติกรรมที่มีรายละเอียดมากขึ้น ที่เกี่ยวข้องกับบทความนี้คือการศึกษา neuroimaging ที่เชื่อมโยงการพัฒนาของระบบทางเดินเส้นใยกับการปรับปรุงความสามารถทางปัญญา โดยเฉพาะอย่างยิ่งความสัมพันธ์ระหว่างมาตรการที่ใช้ DTI ของการพัฒนาสารสีขาวล่วงหน้าและการควบคุมความรู้ความเข้าใจในเด็กได้รับการแสดง ในการศึกษาหนึ่งการพัฒนาความสามารถนี้มีความสัมพันธ์เชิงบวกกับผืนเส้นใย prefrontal-parietal (Nagy, Westerberg และ Klingberg, 2004) สอดคล้องกับการศึกษา neuroimaging หน้าที่แสดงการรับสมัครที่แตกต่างกันของภูมิภาคเหล่านี้ในเด็กเมื่อเทียบกับผู้ใหญ่
ใช้วิธีการที่คล้ายกัน Liston และคณะ (2005) ได้แสดงให้เห็นว่าสสารสีขาวระหว่าง prefrontal-basal ganglia และ -front fiber tracts ยังคงพัฒนาข้ามวัยเด็กสู่วัยผู้ใหญ่ แต่มีเพียงบริเวณทางเดินที่อยู่ระหว่างเยื่อหุ้มสมอง prefrontal และ basal ganglia นั้นสัมพันธ์กับการควบคุมแรงกระตุ้นซึ่งวัดจากประสิทธิภาพในการเดิน / nogo งาน. ทางเดินเส้นใย prefrontal ถูกกำหนดโดยภูมิภาคของผลประโยชน์ที่ระบุไว้ในการศึกษา fMRI โดยใช้งานเดียวกัน จากการศึกษา DTI ทั้งพัฒนาการพัฒนาการของเส้นใยมีความสัมพันธ์กับการพัฒนา แต่ความจำเพาะของเส้นใยเฉพาะที่มีประสิทธิภาพการรับรู้แสดงโดยแยกส่วนทางเดินเฉพาะ (Liston และคณะ, 2005) หรือความสามารถทางปัญญา (นากี้ และคณะ 2004) การค้นพบนี้เน้นย้ำถึงความสำคัญของการตรวจสอบไม่เพียง แต่ในระดับภูมิภาค แต่ยังเกี่ยวข้องกับการเปลี่ยนแปลงของวงจรเมื่อทำการเรียกร้องเกี่ยวกับการเปลี่ยนแปลงตามอายุในพื้นผิวประสาทของการพัฒนาความรู้ความเข้าใจ
การศึกษา MRI เชิงหน้าที่ของการพัฒนาพฤติกรรมและสมอง
แม้ว่าการเปลี่ยนแปลงเชิงโครงสร้างที่วัดโดย MRI และ DTI นั้นเกี่ยวข้องกับการเปลี่ยนแปลงพฤติกรรมในระหว่างการพัฒนา แต่แนวทางที่ตรงกว่าสำหรับการตรวจสอบความสัมพันธ์ของโครงสร้างและหน้าที่คือการวัดการเปลี่ยนแปลงในสมองและพฤติกรรมพร้อมกันเช่นเดียวกับ fMRI ความสามารถในการวัดการเปลี่ยนแปลงการทำงานในสมองที่กำลังพัฒนาด้วย MRI นั้นมีศักยภาพที่สำคัญสำหรับสาขาวิทยาศาสตร์เพื่อการพัฒนา ในบริบทของบทความปัจจุบัน fMRI มีวิธีการในการตีความการตีความพฤติกรรมของวัยรุ่น ตามที่ระบุไว้ก่อนหน้านี้การพัฒนาของเยื่อหุ้มสมอง prefrontal เชื่อว่าจะมีบทบาทสำคัญในการกำหนดความสามารถทางปัญญาที่สูงขึ้นเช่นการตัดสินใจและการควบคุมความรู้ความเข้าใจ (เคซี่ย์ท็อตแนมและฟอสเซลลา 2002b; เคซี่ย์ เทรนเนอร์ และคณะ 1997) กระบวนทัศน์จำนวนมากได้ถูกนำมาใช้ร่วมกับ fMRI เพื่อประเมินพื้นฐานทางชีววิทยาของความสามารถเหล่านี้รวมถึง Flanker, Stroop และ go / nogo งาน (Casey, Castellanos และคณะ, 1997; Casey, Giedd และ Thomas, 2000a; Durston และคณะ, 2003) โดยรวมแล้วการศึกษาเหล่านี้แสดงให้เห็นว่าเด็ก ๆ รับสมัครงานที่แตกต่างกัน แต่บ่อยครั้งที่มีขนาดใหญ่และแพร่กระจายมากขึ้นในช่วง prefrontal ภูมิภาคเมื่อทำงานเหล่านี้มากกว่าผู้ใหญ่ รูปแบบของกิจกรรมภายในพื้นที่สมองเป็นศูนย์กลางในการปฏิบัติงาน (เช่นที่สัมพันธ์กับประสิทธิภาพการรับรู้) มากขึ้นโฟกัสหรือปรับตามอายุในขณะที่ภูมิภาคไม่สัมพันธ์กับการทำงานลดลงในกิจกรรมตามอายุ รูปแบบนี้ได้รับการสังเกตข้ามทั้งหน้าตัดBrown และคณะ, 2005) และการศึกษาระยะยาว (Durston และคณะ, 2006) และในกระบวนทัศน์ที่หลากหลาย แม้ว่าการศึกษา neuroimaging ไม่สามารถกำหนดลักษณะของกลไกของการเปลี่ยนแปลงการพัฒนาดังกล่าว (เช่น dendritic arborization, synaptic pruning) การค้นพบนี้สะท้อนให้เห็นถึงการพัฒนาภายในและการปรับแต่งของการคาดการณ์ไปและกลับจากบริเวณสมองที่เปิดใช้งาน ช่วงเวลา (Brown และคณะ, 2005; Bunge, Dudukovic, Thomason, Vaidya และ Gabrieli, 2002; เคซี่ย์ เทรนเนอร์ และคณะ 1997; เคซี่ย์ และคณะ, 2002ก; Crone, Donohue, Honomichl, Wendelken และ Bunge, 2006; Luna และคณะ, 2001; โมเสสและคณะ, 2002; Schlaggar และคณะ, 2002; Tamm et al., 2002; โทมัส et al., 2004; Turkeltaub, Gareau, Flowers, Zeffiro และ Eden, 2003).
วิธีการนี้จะบอกเราได้อย่างไรว่าวัยรุ่นขาดการควบคุมการรับรู้อย่างเพียงพอ (หุนหันพลันแล่น) หรือมีความเสี่ยงในการเลือกและการกระทำของพวกเขา? การควบคุมแรงกระตุ้นตามที่วัดได้จากงานควบคุมการรับรู้เช่นงาน go / nogo แสดงรูปแบบเชิงเส้นของการพัฒนาในวัยเด็กและวัยรุ่นตามที่อธิบายไว้ข้างต้น อย่างไรก็ตามการศึกษา neuroimaging ล่าสุดได้เริ่มตรวจสอบการประมวลผลที่เกี่ยวข้องกับการให้รางวัลเฉพาะกับความเสี่ยงในวัยรุ่น (Bjork และคณะ, 2004; เอินส์ทและอัล, 2005; อาจ et al, 2004) การศึกษาเหล่านี้ได้มุ่งเน้นไปที่ภูมิภาคของ accumbens เป็นส่วนหนึ่งของฐานปมประสาทที่เกี่ยวข้องกับการทำนายรางวัลมากกว่าลักษณะของการพัฒนาของภูมิภาคนี้ร่วมกับภูมิภาคควบคุมจากบนลงล่าง (เยื่อหุ้มสมองด้านหน้า) แม้ว่ารายงานล่าสุดของกิจกรรม prefrontal หน้าท้องน้อยกว่าในวัยรุ่นที่เกี่ยวข้องกับผู้ใหญ่ในช่วงงานการตัดสินใจทางการเงินเกี่ยวกับพฤติกรรมความเสี่ยงได้รับการแสดง (Eshel, Nelson, Blair, Pine, & Ernst, 2007).
โดยรวมแล้วมีงานวิจัยจำนวนน้อยที่ตรวจสอบว่าการพัฒนาวงจรรางวัลในภูมิภาค subcortical (เช่น accumbens) เปลี่ยนแปลงร่วมกับการพัฒนาของภูมิภาค prefrontal เยื่อหุ้มสมอง ยิ่งไปกว่านั้นการเปลี่ยนแปลงของระบบประสาทเหล่านี้สอดคล้องกับพฤติกรรมการแสวงหารางวัลการกระตุ้นและพฤติกรรมเสี่ยง แบบจำลองทางประสาทวิทยาของเราเสนอว่าการรวมกันของการตอบสนองที่เพิ่มมากขึ้นกับการให้รางวัลและการไม่บรรลุนิติภาวะในพื้นที่ควบคุมพฤติกรรมอาจมีอคติกับวัยรุ่นในการแสวงหาผลประโยชน์ในระยะยาวมากกว่าการได้รับผลระยะยาวอาจอธิบายการเพิ่มขึ้นของพฤติกรรมเสี่ยง การติดตาม subcortical (เช่น accumbens) และเยื่อหุ้มสมอง (เช่น prefrontal) การพัฒนาของการตัดสินใจในวัยเด็กผ่านวัยผู้ใหญ่ให้ข้อ จำกัด เพิ่มเติมเกี่ยวกับการเปลี่ยนแปลงที่รายงานในวัยรุ่นเฉพาะในช่วงเวลาของการพัฒนานี้หรือสะท้อนให้เห็นถึงการสุกที่เกิดขึ้นใน รูปแบบเชิงเส้นค่อนข้างตั้งแต่วัยเด็กจนถึงวัยผู้ใหญ่
หลักฐานเชิงประจักษ์จากการศึกษา fMRI เมื่อเร็ว ๆ นี้ช่วยสนับสนุนโมเดล neurobiological ของเราและใช้วิธีการเปลี่ยนผ่านเพื่อทำความเข้าใจวัยรุ่นโดยการตรวจสอบการเปลี่ยนแปลงก่อนและหลังวัยรุ่น ในการศึกษานี้ (Galvan et al., 2006) เราตรวจสอบการตอบสนองเชิงพฤติกรรมและระบบประสาทเพื่อให้รางวัลแก่การพัฒนาข้ามการมุ่งเน้นไปที่บริเวณสมองที่เกี่ยวข้องกับการเรียนรู้และพฤติกรรมที่เกี่ยวข้องกับรางวัลในสัตว์ (ฮิโคซากะและวาตานาเบะ, 2000; Pecina, Cagniard, Berridge, Aldridge และ Zhuang, 2003; ชูลท์ซ 2006) และการศึกษาการถ่ายภาพผู้ใหญ่ (เช่น Knutson, Adams, Fong และ Hommer, 2001; O, Doherty, Kringelbach, Rolls, Hornak, Andrews, 2001; Zald et al., 2004) และในการศึกษาของการติดยาเสพติด (Hyman & Malenka, 2001; Volkow & Li, 2004) ขึ้นอยู่กับรุ่นหนูLaviola และคณะ, 1999; หอก 2000) และงานถ่ายภาพก่อนหน้า (เอินส์ทและอัล, 2005) เราตั้งสมมุติฐานว่าเมื่อเปรียบเทียบกับเด็กและผู้ใหญ่วัยรุ่นจะแสดงการเปิดใช้งานของผู้ถูกกล่าวหามากเกินไปในคอนเสิร์ตพร้อมกับการรับสมัครที่ไม่เป็นผู้ใหญ่ของเขตการควบคุมล่วงหน้าจากบนลงล่าง งานล่าสุดแสดงการเชื่อมต่อการทำงานล่าช้าระหว่างภูมิภาค subcortical prefrontal และ limbic เหล่านี้ในวัยรุ่นเมื่อเทียบกับผู้ใหญ่ให้กลไกสำหรับการขาดการควบคุมบนลงล่างของภูมิภาคเหล่านี้ (Hare et al., 2007a).
การค้นพบของเราสอดคล้องกับแบบจำลองหนู (Laviola, Macri, Morley-Fletcher และ Adriani, 2003) และการศึกษาการถ่ายภาพก่อนหน้า (เอินส์ทและอัล, 2005) แนะนำกิจกรรม accumbens ที่ปรับปรุงใหม่เพื่อให้รางวัลในช่วงวัยรุ่น แท้จริงแล้วเมื่อเปรียบเทียบกับเด็กและผู้ใหญ่แล้ววัยรุ่นแสดงให้เห็นถึงการตอบสนองที่เกินจริงในการรอรับรางวัล อย่างไรก็ตามเด็กและวัยรุ่นมีการตอบสนองที่เป็นผู้ใหญ่น้อยกว่าในเขตการควบคุมล่วงหน้ามากกว่าผู้ใหญ่ การค้นพบเหล่านี้แนะนำวิถีการพัฒนาที่แตกต่างกันสำหรับภูมิภาคเหล่านี้อาจรองรับการปรับปรุงในกิจกรรม accumbens เมื่อเทียบกับเด็กหรือผู้ใหญ่ซึ่งอาจเกี่ยวข้องกับพฤติกรรมหุนหันพลันแล่นและความเสี่ยงที่เพิ่มขึ้นในช่วงเวลาของการพัฒนานี้ (ดู มะเดื่อ. 4).

มีการรายงานการรับสมัครที่แตกต่างกันของ prefrontal และ subcortical ในการศึกษา fMRI เชิงพัฒนาการจำนวนหนึ่ง (Casey et al., 2002b; Monk et al., 2003; โทมัส et al., 2004) โดยทั่วไปแล้วการค้นพบนี้ได้รับการตีความในแง่ของภูมิภาค prefrontal ที่ยังไม่บรรลุนิติภาวะมากกว่าความไม่สมดุลระหว่างการพัฒนาภูมิภาค prefrontal และ subcortical ได้รับหลักฐานของภูมิภาค prefrontal ในแนวทางการดำเนินการที่เหมาะสมในบริบทที่แตกต่างกัน (มิลเลอร์แอนด์โคเฮน 2001) กิจกรรม prefrontal ที่ยังไม่บรรลุนิติภาวะอาจขัดขวางการประมาณค่าที่เหมาะสมของผลลัพธ์ในอนาคตและการประเมินทางเลือกที่มีความเสี่ยงและอาจมีอิทธิพลน้อยกว่าในการประเมินมูลค่ารางวัลกว่าผู้รับรางวัล รูปแบบนี้สอดคล้องกับการวิจัยก่อนหน้านี้แสดง subcortical สูงเมื่อเทียบกับกิจกรรมเยื่อหุ้มสมองเมื่อการตัดสินใจจะลำเอียงโดยทันทีมากกว่ากำไรระยะยาว (McClure, Laibson, Loewenstein และ Cohen, 2004) นอกจากนี้ยังมีการแสดงกิจกรรม accumbens กับ fMRI เพื่อสัมพันธ์เชิงบวกกับพฤติกรรมการเสี่ยงต่อไป (Kuhnen & Knutson, 2005) ในช่วงวัยรุ่นสัมพันธ์กับวัยเด็กหรือวัยผู้ใหญ่เยื่อหุ้มสมองส่วนหน้าท้องที่ยังไม่สมบูรณ์อาจไม่สามารถควบคุมการประมวลผลรางวัลที่มีแรงกระตุ้นจากภายนอกได้อย่างเพียงพอ (เช่น accumbens) ส่งผลให้ระบบ prefrontal น้อยลง (orbitofrontal cortex) การประเมินค่า
เหตุใดสมองจึงถูกโปรแกรมให้พัฒนาวิธีนี้
วัยรุ่นเป็นช่วงเปลี่ยนผ่านระหว่างวัยเด็กและผู้ใหญ่มักจะเกิดขึ้นกับวัยแรกรุ่น วัยแรกรุ่นนับเป็นจุดเริ่มต้นของการเติบโตทางเพศ (Graber & Brooks-Gunn, 1998) และสามารถกำหนดได้โดยเครื่องหมายทางชีวภาพ วัยรุ่นสามารถอธิบายได้ว่าเป็นการเปลี่ยนแปลงที่ก้าวหน้าไปสู่ความเป็นผู้ใหญ่ด้วยหลักสูตรเกี่ยวกับระยะเวลาของการก่อตัว (หอก 2000) การพูดเชิงวิวัฒนาการวัยรุ่นเป็นช่วงเวลาที่ได้รับทักษะความเป็นอิสระเพื่อเพิ่มความสำเร็จเมื่อแยกออกจากการคุ้มครองของครอบครัวแม้ว่าจะเพิ่มโอกาสสำหรับสถานการณ์ที่เป็นอันตราย (เช่นการบาดเจ็บการซึมเศร้าความวิตกกังวลการใช้ยาและการติดยาเสพติดKelley, Schochet และ Landry, 2004) พฤติกรรมการแสวงหาความเป็นอิสระนั้นแพร่หลายทั่วทั้งสปีชีส์เช่นการเพิ่มปฏิสัมพันธ์ทางสังคมกับเพื่อนและการเพิ่มความรุนแรงในพฤติกรรมการแสวงหาความแปลกใหม่และพฤติกรรมเสี่ยง ปัจจัยทางจิตสังคมส่งผลกระทบต่อแนวโน้มวัยรุ่นสำหรับพฤติกรรมเสี่ยง อย่างไรก็ตามพฤติกรรมที่มีความเสี่ยงเป็นผลมาจากความไม่สมดุลของแรงขับเคลื่อนทางชีวภาพระหว่างการเพิ่มความแปลกใหม่และการแสวงหาความรู้สึกร่วมกับ“ ความสามารถในการกำกับดูแลตนเองที่ยังไม่บรรลุนิติภาวะ” (Steinberg, 2004) ข้อมูลทางระบบประสาทของเราแนะนำว่าสิ่งนี้เกิดขึ้นผ่านการพัฒนาที่แตกต่างกันของทั้งสองระบบ (ลิมบิกและการควบคุม)
การเก็งกำไรจะแนะนำว่ารูปแบบการพัฒนานี้เป็นคุณสมบัติวิวัฒนาการ คุณต้องมีส่วนร่วมในพฤติกรรมที่มีความเสี่ยงสูงที่จะออกจากครอบครัวและหมู่บ้านของคุณเพื่อหาคู่ครองและการเสี่ยงในเวลาเดียวกันขณะที่ฮอร์โมนขับวัยรุ่นเพื่อหาคู่นอน ในสังคมปัจจุบันเมื่อวัยรุ่นอาจขยายไปเรื่อย ๆ โดยที่เด็กอาศัยอยู่กับพ่อแม่และมีการพึ่งพาทางการเงินและเลือกเพื่อนในภายหลังในชีวิตวิวัฒนาการนี้อาจถือว่าไม่เหมาะสม
มีหลักฐานข้ามสปีชีส์เพื่อค้นหาสิ่งแปลกใหม่และการเสี่ยงในช่วงวัยรุ่น ค้นหาเพื่อนในวัยเดียวกันและต่อสู้กับพ่อแม่ซึ่งช่วยให้วัยรุ่นออกไปจากดินแดนบ้านเพื่อการผสมพันธุ์จะเห็นในสายพันธุ์อื่น ๆ รวมถึงหนูสัตว์ที่ไม่ใช่มนุษย์และนกบางตัว (หอก 2000) เมื่อเปรียบเทียบกับผู้ใหญ่หนูในวัยเจริญพันธุ์แสดงพฤติกรรมการแสวงหาสิ่งแปลกใหม่เพิ่มขึ้นในกระบวนทัศน์ทางเลือกที่แปลกใหม่ฟรี (Laviola และคณะ, 1999) หลักฐานทางประสาทวิทยาระบุว่าความสมดุลในสมองวัยรุ่นระหว่างระบบโดปามีนเยื่อหุ้มสมองและกล้ามเนื้อสมองเริ่มเปลี่ยนไปสู่ระดับโดปามีนเยื่อหุ้มสมองส่วนใหญ่ในช่วงวัยรุ่น (หอก 2000) การกระตุ้นโดปามีนที่คล้ายกันยืดเยื้อผ่านวัยรุ่นเข้าสู่วัยผู้ใหญ่ได้รับการแสดงในเยื่อหุ้มสมอง prefrontal เจ้าคณะนอกมนุษย์เช่นกัน (Rosenberg & Lewis, 1995) ดังนั้นความเสี่ยงที่เด่นชัดนี้ดูเหมือนจะข้ามสายพันธุ์และมีวัตถุประสงค์ในการปรับตัวที่สำคัญ
ความบกพร่องทางชีวภาพการพัฒนาและความเสี่ยง
ความแตกต่างระหว่างบุคคลในการควบคุมแรงกระตุ้นและการรับความเสี่ยงได้รับการยอมรับในด้านจิตวิทยามาระยะหนึ่งแล้ว (Benthin, Slovic และ Severson, 1993) บางทีหนึ่งในตัวอย่างคลาสสิกของความแตกต่างระหว่างบุคคลที่รายงานในความสามารถเหล่านี้ในวรรณคดีจิตวิทยาสังคม, ความรู้ความเข้าใจและการพัฒนาเป็นความล่าช้าของความพึงพอใจ (Mischel, Shoda และ Rodriguez, 1989) ความล่าช้าของความพึงพอใจมักจะได้รับการประเมินในเด็กวัยหัดเดิน 3- ถึง 4 ปี เด็กวัยหัดเดินจะถูกถามว่าพวกเขาจะต้องการรางวัลเล็ก ๆ (หนึ่งคุกกี้) หรือรางวัลใหญ่ (สองคุกกี้) จากนั้นเด็กคนนั้นก็บอกว่าผู้ทดลองจะออกจากห้องเพื่อเตรียมสำหรับกิจกรรมที่จะเกิดขึ้นและอธิบายให้เด็กฟังว่าถ้าเธอยังคงนั่งและไม่กินคุกกี้เธอจะได้รับรางวัลใหญ่ หากเด็กทำไม่ได้หรือไม่สามารถรอได้เธอควรกดกริ่งเพื่อเรียกผู้ทดลองและได้รับรางวัลน้อยลง เมื่อเห็นได้ชัดว่าเด็กเข้าใจภาระงานเธอจะนั่งที่โต๊ะพร้อมรางวัลสองรางวัลและเสียงระฆัง สิ่งรบกวนในห้องลดน้อยลงโดยไม่มีของเล่นหนังสือหรือรูปภาพ ผู้ทดลองจะกลับมาหลังจาก 15 นาทีหรือหลังจากที่เด็กดังกริ่งระฆังกินรางวัลหรือแสดงอาการเจ็บปวดใด ๆ Mischel แสดงให้เห็นว่าเด็ก ๆ มักจะประพฤติหนึ่งในสองวิธี: (1) ไม่ว่าพวกเขาจะส่งเสียงกริ่งดังขึ้นเกือบจะในทันทีเพื่อให้มีคุกกี้ซึ่งหมายความว่าพวกเขาจะได้รับเพียงครั้งเดียว หรือ (2) พวกเขารอและเพิ่มประสิทธิภาพกำไรของพวกเขาและรับทั้งสองคุกกี้ การสังเกตนี้แสดงให้เห็นว่าบางคนดีกว่าคนอื่น ๆ ในความสามารถในการควบคุมแรงกระตุ้นในการเผชิญกับแรงจูงใจที่เด่นชัดและสามารถตรวจพบอคตินี้ในวัยเด็ก (Mischel et al., 1989) และดูเหมือนว่าจะยังคงอยู่ตลอดช่วงวัยรุ่นและวัยหนุ่มสาว (Eigsti et al., 2006).
อะไรอาจอธิบายความแตกต่างของแต่ละบุคคลในการตัดสินใจและพฤติกรรมที่ดีที่สุด? นักทฤษฎีบางคนได้ตั้งสมมติฐานว่าวงจร dopaminergic mesolimbic ที่เกี่ยวข้องในการประมวลผลรางวัลรองรับพฤติกรรมเสี่ยง ความแตกต่างของแต่ละคนในวงจรนี้เช่นอัลลีลสายพันธุ์ในยีนที่เกี่ยวข้องกับโดปามีนส่งผลให้โดพามีนน้อยหรือมากเกินไปในภูมิภาค subcortical อาจเกี่ยวข้องกับนิสัยชอบที่จะมีส่วนร่วมในพฤติกรรมเสี่ยง (O'Doherty, 2004) นิวเคลียส accumbens ได้รับการแสดงเพื่อเพิ่มกิจกรรมทันทีก่อนที่จะตัดสินใจเลือกที่มีความเสี่ยงในกระบวนทัศน์ทางการเงินที่มีความเสี่ยง (Kuhnen & Knutson, 2005; Matthews และคณะ 2004; Montague & Berns, 2002) และตามที่อธิบายไว้ก่อนหน้านี้วัยรุ่นแสดงกิจกรรมที่เกินจริงเพื่อให้รางวัลแก่ผลลัพธ์ที่สัมพันธ์กับเด็กหรือผู้ใหญ่ (เอินส์ทและอัล, 2005; Galvan et al., 2006) โดยรวมแล้วข้อมูลเหล่านี้ชี้ให้เห็นว่าวัยรุ่นอาจมีแนวโน้มที่จะมีทางเลือกที่มีความเสี่ยงมากกว่าในกลุ่มGardener & Steinberg, 2005) แต่วัยรุ่นบางคนจะมีแนวโน้มมากกว่าคนอื่น ๆ ที่จะมีส่วนร่วมในพฤติกรรมเสี่ยงทำให้พวกเขามีความเสี่ยงมากขึ้นสำหรับผลลัพธ์เชิงลบ ดังนั้นจึงเป็นสิ่งสำคัญที่จะต้องพิจารณาความแปรปรวนของแต่ละบุคคลเมื่อตรวจสอบความสัมพันธ์ของสมองและพฤติกรรมที่ซับซ้อนที่เกี่ยวข้องกับการเสี่ยงและการให้รางวัลในการพัฒนาประชากร
เพื่อสำรวจความแตกต่างของบุคคลในพฤติกรรมเสี่ยง Galvan และคณะ (2007) เมื่อเร็ว ๆ นี้ตรวจสอบความสัมพันธ์ระหว่างกิจกรรมในวงจรประสาทที่เกี่ยวข้องกับการให้รางวัลในการคาดหวังผลตอบแทนทางการเงินจำนวนมากด้วยมาตรการลักษณะบุคลิกภาพของการรับความเสี่ยงและความหุนหันพลันแล่นในวัยรุ่น การถ่ายภาพด้วยเรโซแนนซ์แม่เหล็กและระดับการรายงานด้วยตนเองแบบไม่ระบุชื่อของพฤติกรรมเสี่ยงการรับรู้ความเสี่ยงและการกระตุ้นได้รับในบุคคลที่มีอายุระหว่าง 7 และ 29 ปี มีความสัมพันธ์ในเชิงบวกระหว่างกิจกรรมของผู้ป่วยและโอกาสในการมีพฤติกรรมเสี่ยงในการพัฒนา กิจกรรมนี้แตกต่างกันไปตามฟังก์ชั่นของการจัดอันดับของบุคคลที่คาดว่าจะเกิดผลทางบวกหรือทางลบของพฤติกรรมดังกล่าว บุคคลเหล่านั้นที่รับรู้พฤติกรรมเสี่ยงที่นำไปสู่ผลกระทบที่เลวร้ายทำให้ผู้ที่ได้รับผลกระทบน้อยลงเพื่อให้รางวัล ความสัมพันธ์นี้ถูกขับเคลื่อนโดยเด็กส่วนใหญ่โดยผู้ใหญ่ให้คะแนนผลของพฤติกรรมดังกล่าวเท่าที่จะทำได้ การให้คะแนนความหุนหันพลันแล่นไม่เกี่ยวข้องกับกิจกรรม accumbens แต่ขึ้นกับอายุ การค้นพบเหล่านี้ชี้ให้เห็นว่าในช่วงวัยรุ่นบางคนอาจมีแนวโน้มที่จะมีส่วนร่วมในพฤติกรรมเสี่ยงเนื่องจากการเปลี่ยนแปลงการพัฒนาในคอนเสิร์ตพร้อมกับความแปรปรวนในความโน้มเอียงของบุคคลที่กำหนดให้มีส่วนร่วมในพฤติกรรมเสี่ยงมากกว่าการเปลี่ยนแปลงง่าย ๆ มะเดื่อ. 5).

พฤติกรรมของวัยรุ่นได้รับการโดดเด่นเป็นหุนหันพลันแล่นและมีความเสี่ยง (Steinberg, 2004, 2007) แต่จากการทบทวนวรรณกรรมการถ่ายภาพนี้แสดงให้เห็นพื้นผิว neurobiological ที่แตกต่างกันและวิถีการพัฒนาที่แตกต่างกันสำหรับพฤติกรรมเหล่านี้ โดยเฉพาะอย่างยิ่งแรงกระตุ้นที่เกี่ยวข้องกับการพัฒนา prefrontal หน้าท้องที่ยังไม่บรรลุนิติภาวะและค่อยๆลดลงตั้งแต่วัยเด็กจนถึงวัยผู้ใหญ่ (Casey, Galvan และคณะ, 2005) ความสัมพันธ์เชิงลบระหว่างการจัดอันดับแรงกระตุ้นและอายุในการศึกษาโดย Galvan และคณะ (2007) สนับสนุนแนวคิดนี้เพิ่มเติม ในทางตรงกันข้ามการรับความเสี่ยงนั้นสัมพันธ์กับการเพิ่มขึ้นของกิจกรรมKuhnen & Knutson, 2005; Matthews และคณะ 2004; Montague & Berns, 2002) ที่พูดเกินจริงในวัยรุ่นสัมพันธ์กับเด็กและผู้ใหญ่ (เอินส์ทและอัล, 2005; Galvan et al., 2006) ดังนั้นทางเลือกวัยรุ่นและพฤติกรรมไม่สามารถอธิบายได้ด้วยการกระตุ้นหรือการพัฒนาของเยื่อหุ้มสมองส่วนหน้าเพียงอย่างเดียวเนื่องจากเด็กจะได้รับการทำนายว่าเป็นผู้รับความเสี่ยงมากขึ้น การค้นพบนี้ให้พื้นฐานทางประสาทสำหรับสาเหตุที่วัยรุ่นบางคนมีความเสี่ยงมากกว่าคนอื่น ๆ แต่ยังเป็นพื้นฐานสำหรับพฤติกรรมของวัยรุ่นที่แตกต่างจากเด็กและผู้ใหญ่ในการเสี่ยง
โดยรวมแล้วข้อมูลเหล่านี้ชี้ให้เห็นว่าถึงแม้ว่าวัยรุ่นในฐานะกลุ่มจะได้รับการพิจารณาว่าเป็นผู้เสี่ยงGardener & Steinberg, 2005) วัยรุ่นบางคนจะมีแนวโน้มมากกว่าคนอื่น ๆ ที่จะมีส่วนร่วมในพฤติกรรมเสี่ยงทำให้พวกเขามีความเสี่ยงมากขึ้นสำหรับผลลัพธ์เชิงลบ การค้นพบนี้เน้นย้ำถึงความสำคัญของการพิจารณาความแปรปรวนของแต่ละบุคคลเมื่อตรวจสอบความสัมพันธ์ของสมองและพฤติกรรมที่ซับซ้อนที่เกี่ยวข้องกับการเสี่ยงและการให้รางวัลในการพัฒนาประชากร นอกจากนี้ความแตกต่างระหว่างบุคคลและการพัฒนาเหล่านี้อาจช่วยอธิบายถึงช่องโหว่ในบุคคลบางคนเพื่อรับความเสี่ยงจากการใช้สารเสพติด
สรุป
การศึกษาเกี่ยวกับการถ่ายภาพของมนุษย์แสดงให้เห็นถึงการเปลี่ยนแปลงเชิงโครงสร้างและหน้าที่ในภูมิภาคส่วนหน้า (Giedd et al., 1996, 1999; Jernigan และคณะ, 1991; Sowell และคณะ, 1999; เพื่อการตรวจสอบ Casey, Galvan และคณะ, 2005) ที่ดูเหมือนจะเพิ่มขึ้นแบบขนานในการควบคุมการรับรู้และการควบคุมตนเอง (เคซี่ย์ เทรนเนอร์ และคณะ 1997; Luna & Sweeney, 2004; Luna และคณะ, 2001; Rubia และคณะ 2000; Steinberg, 2004; ดูเพิ่มเติม Steinberg, 2008ปัญหานี้) การเปลี่ยนแปลงเหล่านี้ดูเหมือนจะแสดงให้เห็นถึงการเปลี่ยนแปลงในการเปิดใช้งานของภูมิภาค prefrontal จากการกระจายไปยังโฟกัสที่มากขึ้นเมื่อเวลาผ่านไป (Brown และคณะ, 2005; Bunge et al., 2002; เคซี่ย์ เทรนเนอร์ และคณะ 1997; Durston และคณะ, 2006; โมเสสและคณะ, 2002) และการรับสมัครที่เพิ่มขึ้นของภูมิภาคย่อยในช่วงวัยรุ่น (เคซี่ย์ และคณะ, 2002ก; Durston และคณะ, 2006; Luna และคณะ, 2001) แม้ว่าการศึกษา neuroimaging ไม่สามารถกำหนดลักษณะของกลไกของการเปลี่ยนแปลงการพัฒนาดังกล่าวได้อย่างชัดเจน แต่การเปลี่ยนแปลงของปริมาณและโครงสร้างเหล่านี้อาจสะท้อนถึงการพัฒนาภายในและการปรับการคาดการณ์ไปยังและออกจากบริเวณสมองเหล่านี้ในระหว่างการเจริญเติบโต
เมื่อนำมารวมกันการค้นพบที่สังเคราะห์ที่นี่บ่งชี้ว่าพฤติกรรมการรับความเสี่ยงที่เพิ่มขึ้นในวัยรุ่นนั้นมีความสัมพันธ์กับวิถีการพัฒนาที่แตกต่างกันของความสุข subcortical และภูมิภาคควบคุมเยื่อหุ้มสมอง การเปลี่ยนแปลงการพัฒนาเหล่านี้สามารถทำให้รุนแรงขึ้นด้วยความแตกต่างของแต่ละบุคคลในกิจกรรมของระบบการให้รางวัล แม้ว่าวัยรุ่นได้รับการโดดเด่นเป็นช่วงเวลาที่โดดเด่นด้วยการแสวงหารางวัลและพฤติกรรมการเสี่ยง (Gardener & Steinberg, 2005; หอก 2000) ความแตกต่างของแต่ละบุคคลในการตอบสนองต่อระบบประสาทเพื่อให้รางวัลจูงใจวัยรุ่นบางคนให้รับความเสี่ยงมากกว่าคนอื่น ๆ ทำให้พวกเขามีความเสี่ยงมากขึ้นสำหรับผลลัพธ์เชิงลบ การค้นพบเหล่านี้ให้รากฐานที่สำคัญโดยการสังเคราะห์การค้นพบต่าง ๆ ที่เกี่ยวข้องกับพฤติกรรมเสี่ยงในวัยรุ่นและในการทำความเข้าใจความแตกต่างของแต่ละบุคคลและเครื่องหมายการพัฒนาสำหรับ propensities ที่จะมีส่วนร่วมในพฤติกรรมเชิงลบ
กิตติกรรมประกาศ
งานนี้ได้รับการสนับสนุนบางส่วนโดยทุนจากสถาบันยาเสพติดแห่งชาติ R01 DA18879 และสถาบันสุขภาพจิตแห่งชาติ 1P50 MH62196
อ้างอิง
- Benthin A, Slovic P, Severson H. การศึกษาไซโครเมทริกของการรับรู้ความเสี่ยงของวัยรุ่น วารสารวัยรุ่น 1993;16: 153 168- [PubMed]
- Bjork JM, Knutson B, Fong GW, Caggiano DM, Bennett SM, Hommer DW การกระตุ้นสมองที่กระตุ้นให้เกิดแรงจูงใจในวัยรุ่น: ความเหมือนและความแตกต่างจากคนหนุ่มสาว วารสารประสาทวิทยา 2004;24: 1793 1802- [PubMed]
- Bjorkland DF บทบาทของความรู้ทางแนวคิดในการพัฒนาองค์กรในความทรงจำของเด็ก ใน: Brainerd CJ, Pressley M, บรรณาธิการ กระบวนการพื้นฐานในการพัฒนาหน่วยความจำ: ความก้าวหน้าในการวิจัยการพัฒนาองค์ความรู้ นิวยอร์ก: Springer-Verlag; 1985 pp. 103 – 142
- Bjorkland DF การเปลี่ยนแปลงอายุในฐานความรู้มีส่วนช่วยในการพัฒนาความจำของเด็กอย่างไร: การทบทวนเชิงตีความ รีวิวการพัฒนา 1987;7: 93 130-
- Bourgeois JP, Goldman-Rakic PS, Rakic P. Synaptogenesis ในเยื่อหุ้มสมองส่วนหน้าของลิงจำพวก Cortex สมอง 1994;4: 78 96- [PubMed]
- Brainerd CJ, Reyna VF ความเป็นอิสระของหน่วยความจำและการรบกวนหน่วยความจำในการพัฒนาความรู้ความเข้าใจ รีวิวจิตวิทยา 1993;100: 42 67- [PubMed]
- สีน้ำตาล TT, Lugar HM, Coalson RS, Miezin FM, Petersen SE, Schlaggar BL การเปลี่ยนแปลงเชิงพัฒนาการขององค์การการทำงานของสมองมนุษย์เพื่อการสร้างคำ Cortex สมอง 2005;15: 275 290- [PubMed]
- Bunge SA, Dudukovic NM, Thomason ME, Vaidya CJ, Gabrieli JD การมีส่วนร่วมของพูหน้าผากที่ยังไม่บรรลุนิติภาวะต่อการควบคุมความรู้ความเข้าใจในเด็ก: หลักฐานจาก fMRI เซลล์ประสาท 2002;33: 301 311- [PubMed]
- Case R. การตรวจสอบความถูกต้องของโครงสร้างกำลังการผลิตนีโอ - Piagetian วารสารจิตวิทยาเด็กทดลอง 1972;14: 287 302-
- เคสอาร์ การพัฒนาทางปัญญา: กำเนิดสู่ความเป็นผู้ใหญ่ นิวยอร์ก: นักวิชาการสื่อมวลชน; 1985
- Casey BJ, Castellanos FX, Giedd JN, มาร์ช WL, Hamburger SD, Schubert AB et al. ความหมายของวงจรด้านหน้าขวาในการยับยั้งการตอบสนองและความผิดปกติของสมาธิสั้น / สมาธิสั้น วารสารของสถาบันจิตเวชศาสตร์เด็กและวัยรุ่นอเมริกัน 1997;36: 374 383- [PubMed]
- Casey BJ, Galvan A, Hare TA การเปลี่ยนแปลงในการทำงานของสมองในระหว่างการพัฒนาความรู้ความเข้าใจ ความคิดเห็นปัจจุบันทางชีววิทยา 2005;15: 239 244- [PubMed]
- Casey BJ, Giedd JN, Thomas KM. การพัฒนาสมองเชิงโครงสร้างและหน้าที่และความสัมพันธ์กับการพัฒนาทางปัญญา จิตวิทยาชีวภาพ 2000a;54: 241 257- [PubMed]
- Casey BJ, Thomas KM, Davidson MC, Kunz K, Franzen PL การแยกฟังก์ชัน striatal และ hippocampal ได้รับการพัฒนาด้วยงานความเข้ากันได้กับสิ่งกระตุ้น วารสารประสาทวิทยา 2002a;22: 8647 8652- [PubMed]
- Casey BJ, Thomas KM, TF ชาวเวลส์, Badgaiyan RD, Eccard CH, Jennings JR, และคณะ การแตกแยกของความขัดแย้งในการตอบสนองการเลือกแบบตั้งใจและความคาดหวังกับการถ่ายภาพด้วยคลื่นสนามแม่เหล็ก การดำเนินการของ National Academy of Science 2000b;97: 8728 8733-
- Casey BJ, Tottenham N, Fossella J. คลินิก, การถ่ายภาพ, แผลและวิธีการทางพันธุกรรมไปสู่รูปแบบของการควบคุมความรู้ความเข้าใจ จิตวิทยาพัฒนาการ 2002b;40: 237 254- [PubMed]
- Casey BJ, Tottenham N, Liston C, Durston S. การถ่ายภาพสมองกำลังพัฒนา: เราเรียนรู้อะไรบ้างเกี่ยวกับการพัฒนาทางปัญญา? แนวโน้มในด้านวิทยาศาสตร์การรับรู้ 2005;9: 104 110-
- Casey BJ, Trainor RJ, Orendi JL, Schubert AB, Nystrom LE, Giedd JN, และคณะ การศึกษา MRI เชิงหน้าที่เชิงพัฒนาการของการเปิดใช้งานล่วงหน้าในระหว่างการทำงานของงานที่ไม่ต้องเดินทาง วารสารประสาทวิทยาศาสตร์ 1997;9: 835 847-
- Crone E, Donohue S, Honomichl R, Wendelken C, Bunge S. พื้นที่สมองเป็นสื่อกลางการใช้กฎที่ยืดหยุ่นในระหว่างการพัฒนา วารสารประสาทวิทยา 2006;26: 11239 11247- [PubMed]
- Dempster FN ความต้านทานต่อสัญญาณรบกวน: การเปลี่ยนแปลงการพัฒนาในกลไกการประมวลผลขั้นพื้นฐาน ใน: Howe ML, Pasnak R, บรรณาธิการ ธีมที่เกิดขึ้นใหม่ในการพัฒนาความรู้ความเข้าใจเล่ม 1: ฐานราก นิวยอร์ก: สปริงเกอร์; 1993 pp. 3 – 27
- Diamond A. การพัฒนาความสามารถในการใช้การเรียกคืนเพื่อเป็นแนวทางในการดำเนินการตามที่ระบุโดยประสิทธิภาพของทารกใน AB พัฒนาการของเด็ก 1985;56: 868 883- [PubMed]
- Durston S, Davidson MC, Thomas KM, Worden MS, Tottenham N, Martinez A, et al. การจัดการความขัดแย้งและการตอบสนองแบบ Parametric โดยใช้ fMRI ที่เกี่ยวข้องกับการทดลองผสมเหตุการณ์อย่างรวดเร็ว Neuroimage 2003;20: 2135 2141- [PubMed]
- Durston S, Davidson MC, Tottenham N, Galvan A, Spicer J, Fossella J, และคณะ การเปลี่ยนจากการกระจายไปสู่การทำกิจกรรมนอกสมองกับการพัฒนา วิทยาศาสตร์พัฒนาการ 2006;1: 18 20- [PubMed]
- Durston S, Hulshoff Pol HE, Casey BJ, Giedd JN, Buitelaar JK, Van Engeland H. กายวิภาค MRI ของสมองมนุษย์ที่กำลังพัฒนา: เราเรียนรู้อะไรบ้าง? วารสาร American Academy จิตเวชศาสตร์เด็กวัยรุ่น 2001;40: 1012 1020-
- Eaton LK, Kinchen S, Ross J, Hawkins J, Harris WA, Lowry R, et al. การเฝ้าระวังพฤติกรรมเสี่ยงของเยาวชน - สหรัฐอเมริกา, 2005, บทสรุปการเฝ้าระวัง รายงานการเสียชีวิตและความตายประจำสัปดาห์ 2006;55: 1 108- [PubMed]
- Eigsti IM, Zayas V, Mischel W, Shoda Y, Ayduk O, Dadlani MB และอื่น ๆ การทำนายการควบคุมความรู้ความเข้าใจตั้งแต่วัยเด็กจนถึงวัยปลายและวัยหนุ่มสาว วิทยาศาสตร์จิตวิทยา 2006;17: 478 484- [PubMed]
- เอินส์ทเอ็ม, เนลสัน EE, Jazbec S, McClure EB, พระภิกษุสงฆ์, Leibenluft E, และคณะ Amygdala และนิวเคลียส accumbens ในการตอบสนองต่อการรับและการละเว้นของกำไรในผู้ใหญ่และวัยรุ่น Neuroimage 2005;25: 1279 1291- [PubMed]
- Eshel N, เนลสัน EE, แบลร์ RJ, ไพน์เอส, เอิร์นส์เอ็ม. สารตั้งต้นของการเลือกทางเลือกในผู้ใหญ่และวัยรุ่น: การพัฒนาของเยื่อหุ้มปอดด้านหน้าและหน้าท้อง Neuropsychologia 2007;45: 1270 1279- [บทความฟรี PMC] [PubMed]
- Flavell JH, Feach DR, Chinsky JM การฝึกฝนด้วยวาจาโดยธรรมชาติในงานความทรงจำในฐานะหน้าที่ของอายุ พัฒนาการของเด็ก 1966;37: 283 299- [PubMed]
- Galvan A, Hare TA, Parra CE, Penn J, Voss H, Glover G, et al. การพัฒนา accumbens ก่อนหน้านี้เมื่อเทียบกับ orbitofrontal cortex อาจรองรับพฤติกรรมเสี่ยงในวัยรุ่น วารสารประสาทวิทยา 2006;26: 6885 6892- [PubMed]
- Galvan A, Hare T, Voss H, Glover G, Casey BJ ความเสี่ยงและสมองวัยรุ่น: ใครมีความเสี่ยง วิทยาศาสตร์พัฒนาการ 2007;10: F8-F14 [PubMed]
- Gardener M, Steinberg L. Peer มีอิทธิพลต่อการรับความเสี่ยงการตั้งค่าความเสี่ยงและการตัดสินใจที่มีความเสี่ยงในวัยรุ่นและวัยผู้ใหญ่: การศึกษาเชิงทดลอง จิตวิทยาพัฒนาการ. 2005;41: 625 635- [PubMed]
- Giedd JN การถ่ายภาพด้วยคลื่นสนามแม่เหล็กของสมองวัยรุ่น พงศาวดารของ New York Academy of Sciences 2004;1021: 77 85- [PubMed]
- Giedd JN, Blumenthal J, Jeffries NO, Castellanos FX, Liu H, Zijdenbos A, et al. การพัฒนาสมองในวัยเด็กและวัยรุ่น: การศึกษา MRI ระยะยาว ธรรมชาติประสาทวิทยา 1999;2: 861 863-
- Giedd JN, Snell JW, Lange N, Rajapakse JC, Casey BJ, Kozuch PL, และคณะ การถ่ายภาพด้วยคลื่นสนามแม่เหล็กเชิงปริมาณของการพัฒนาสมองของมนุษย์: อายุ 4 – 18 Cortex สมอง 1996;6: 551 560- [PubMed]
- Gogtay N, Giedd JN, รูสก์ L, ฮายาชิ KM, Greenstein D, Vaituzis AC, et al. การทำแผนที่แบบไดนามิกของการพัฒนาเยื่อหุ้มสมองของมนุษย์ในวัยเด็กผ่านวัยผู้ใหญ่ตอนต้น การดำเนินการของ National Academy of Sciences ของสหรัฐอเมริกา 2004;101: 8174 8179- [บทความฟรี PMC] [PubMed]
- Graber JA, Brooks-Gunn J. วัยแรกรุ่น ใน: Blechman EA, Brownell KD, editors คู่มือยาและเวชศาสตร์เชิงพฤติกรรมสำหรับผู้หญิง New York, NY: Guilford Press; 1998 pp. 51 – 58
- Hare TA, Voss HU, Glover GH, Casey BJ สมองของวัยรุ่นและความเสี่ยงที่อาจเกิดขึ้นกับความวิตกกังวลและภาวะซึมเศร้า 2007a ส่งเพื่อการเผยแพร่
- Hare TA, Voss HU, Glover GH, Casey BJ การแข่งขันระหว่างระบบลิมบิก prefrontal และ subcortical ภายใต้ปฏิกิริยาทางอารมณ์ในช่วงวัยรุ่น 2007b ส่งเพื่อการเผยแพร่
- Harnishfeger KK, Bjorkland F. กลไกการยับยั้งการทำงานของร่างกาย: แนวทางใหม่ในการพัฒนาความรู้ความเข้าใจ ใน: Howe ML, Pasnek R, บรรณาธิการ ธีมใหม่ ๆ ในการพัฒนาความรู้ความเข้าใจ ฉบับ 1 นิวยอร์ก: Springer-Verlag; 1993 pp. 28 – 49
- Hikosaka K, Watanabe M. กิจกรรมที่ล่าช้าของเซลล์ประสาทวงโคจรและด้านข้างด้านหน้าของลิงที่แตกต่างกันไปกับรางวัลที่แตกต่างกัน Cortex สมอง 2000;10: 263 271- [PubMed]
- Huttenlocher PR ความหนาแน่นของ Synaptic ในเยื่อหุ้มสมองส่วนหน้า - การเปลี่ยนแปลงและพัฒนาการของริ้วรอย การวิจัยสมอง 1979;163: 195 205- [PubMed]
- Hyman SE, Malenka RC ติดยาเสพติดและสมอง: ชีววิทยาของการบังคับและความเพียรของมัน รีวิวจากธรรมชาติ Neuroscience 2001;2: 695 703-
- Jernigan TL, Zisook S, Heaton RK, Moranville JT, Hesselink JR, Braff DL ความผิดปกติของการถ่ายภาพด้วยคลื่นสนามแม่เหล็กในนิวเคลียสแม่และลูกสมองในโรคจิตเภท จดหมายเหตุของจิตเวชทั่วไป 1991;48: 811 823-
- Keating DP, Bobbitt BL ความแตกต่างระหว่างบุคคลและพัฒนาการในองค์ประกอบการประมวลผลความรู้ความเข้าใจของความสามารถทางจิต พัฒนาการของเด็ก 1978;49: 155 167-
- ตวัด AE, Schochet T, Landry C. พงศาวดารของ New York Academy of Sciences 2004;1021: 27 32- [PubMed]
- Knutson B, Adams CM, Fong GW, Hommer D. ความคาดหวังของการเพิ่มรางวัลทางการเงินที่คัดเลือกเข้ามาคัดเลือกนิวเคลียส accumbens วารสารประสาทวิทยา 2001;21: RC159 [PubMed]
- Kuhnen CM, Knutson B. พื้นฐานทางประสาทของการรับความเสี่ยงทางการเงิน เซลล์ประสาท 2005;47: 763 770- [PubMed]
- Laviola G, Adriani W, Terranova ML, Gerra G. ปัจจัยเสี่ยงทางจิตวิทยาสำหรับความอ่อนแอต่อยาจิตเวชในวัยรุ่นและสัตว์ทดลอง ประสาทวิทยาศาสตร์และชีวจิตรีวิว 1999;23: 993 1010- [PubMed]
- Laviola G, Macri S, Morley-Fletcher S, Adriani W. พฤติกรรมการรับความเสี่ยงในหนูวัยรุ่น: ปัจจัยทางจิตวิทยาและอิทธิพล epigenetic ในช่วงต้น ประสาทวิทยาศาสตร์และชีวจิตรีวิว 2003;27: 19 31- [PubMed]
- Liston C, Watts R, Tottenham N, Davidson MC, Niogi S, Ulug AM, et al. โครงสร้างจุลภาค Frontostriatal ปรับการสรรหาที่มีประสิทธิภาพของการควบคุมความรู้ความเข้าใจ Cortex สมอง 2005;16: 553 560- [PubMed]
- Luna B, Sweeney JA การเกิดขึ้นของการทำงานของสมองที่ทำงานร่วมกัน: การศึกษา FMRI ของการพัฒนาของการยับยั้งการตอบสนอง พงศาวดารของ New York Academy of Sciences 2004;1021: 296 309- [PubMed]
- Luna B, Thulborn KR, Munoz DP, Merriam EP, Garver KE, Minshew NJ, และคณะ การครบกำหนดของฟังก์ชั่นสมองกระจายอย่างกว้างขวางช่วยลดการพัฒนาความรู้ความเข้าใจ Neuroimage 2001;13: 786 793- [PubMed]
- Matthews SC และคณะ การเปิดใช้งานแบบเลือกของนิวเคลียสจะเกิดขึ้นระหว่างการตัดสินใจเสี่ยง Neuroreport 2004;15: 2123 2127- [PubMed]
- JC, Delgado MR, Dahl RE, Stenger VA, Ryan ND, Fiez JA, et al. เหตุการณ์เกี่ยวกับการถ่ายภาพด้วยคลื่นสนามแม่เหล็กที่เกี่ยวข้องกับการทำงานของวงจรสมองที่เกี่ยวข้องกับการให้รางวัลในเด็กและวัยรุ่น จิตเวชชีวภาพ 2004;55: 359 366- [PubMed]
- McClure SM, Laibson DI, Loewenstein G, Cohen JD ระบบประสาทที่แยกต่างหากให้ความสำคัญกับผลตอบแทนทางการเงินทันที วิทยาศาสตร์ 2004;306: 503 507- [PubMed]
- มิลเลอร์ EK โคเฮน JD ทฤษฎีเชิงบูรณาการของฟังก์ชันเยื่อหุ้มสมองส่วนหน้า การทบทวนประสาทวิทยาประจำปี 2001;24: 167 202-
- Mischel W, Shoda Y, Rodriguez MI ความล่าช้าของความพึงพอใจในเด็ก วิทยาศาสตร์ 1989;244: 933 938- [PubMed]
- พระ CS, McClure EB, เนลสัน EE, Zarahn E, Bilder RM, Leibenluft E, และอื่น ๆ วัยรุ่นที่ยังไม่บรรลุนิติภาวะในการมีส่วนร่วมของสมองที่เกี่ยวข้องกับความสนใจต่อการแสดงออกทางอารมณ์ Neuroimage 2003;20: 420 428- [PubMed]
- Montague PR, Berns GS เศรษฐศาสตร์ประสาทและสารตั้งต้นทางชีวภาพ เซลล์ประสาท 2002;36: 265 284- [PubMed]
- Moses P, Roe K, Buxton RB, Wong EC, Frank LR, Stiles J. MRI ที่ใช้งานได้ของการประมวลผลระดับโลกและระดับท้องถิ่นในเด็ก Neuroimage 2002;16: 415 424- [PubMed]
- Munakata Y, Yerys พ.ศ. ทั้งหมดเข้าด้วยกันตอนนี้: เมื่อความแตกต่างระหว่างความรู้และการกระทำหายไป วิทยาศาสตร์กายภาพ 2001;12: 335 337-
- Nagy Z, Westerberg H, Klingberg T. การเติบโตของสสารสีขาวนั้นสัมพันธ์กับพัฒนาการของการรับรู้ในช่วงวัยเด็ก วารสารประสาทวิทยาศาสตร์ 2004;16: 1227 1233- [PubMed]
- O'Doherty J, Kringelbach ML, Rolls ET, Hornak J, Andrews C. การให้รางวัลและการลงโทษเชิงนามธรรมในเยื่อหุ้มสมอง orbitofrontal ของมนุษย์ Neurosci ธรรมชาติ 2001;4: 95 102- [PubMed]
- JP โดเฮอร์ตี้ ให้รางวัลเป็นตัวแทนและการเรียนรู้ที่เกี่ยวข้องกับผลตอบแทนในสมองมนุษย์: ข้อมูลเชิงลึกจากระบบประสาท ความคิดเห็นปัจจุบันทางชีววิทยา 2004;14: 769 776-
- Pascual-Leone JA แบบจำลองทางคณิตศาสตร์สำหรับการเปลี่ยนแปลงในระยะการพัฒนาของเพียเจต์ Acta Psychologica 1970;32: 301 345-
- Pecina S, Cagniard B, Berridge KC, Aldridge JW, จ้วง X. หนูกลายพันธุ์ Hyperdopaminergic มีหนูสูงกว่า“ ต้องการ” แต่ไม่“ ชอบ” สำหรับรางวัลหวาน วารสารประสาทวิทยา 2003;23: 9395 9402- [PubMed]
- Reyna VF, Farley F. ความเสี่ยงและความมีเหตุผลในการตัดสินใจของวัยรุ่น: ความหมายสำหรับทฤษฎีการปฏิบัติและนโยบายสาธารณะ วิทยาศาสตร์ทางจิตวิทยาเพื่อสาธารณประโยชน์ 2006;7: 1 44-
- Rosenberg DR, Lewis DA การเจริญเติบโตหลังคลอดของการปกคลุมด้วยเส้น dopaminergic ของลิง prefrontal และ cortices ยนต์: การวิเคราะห์อิมมูโนฮิสโตเคมี hydroxylase ไทโรซีน วารสารประสาทวิทยาเปรียบเทียบ 1995;358: 383 400- [PubMed]
- Rubia K, Overmeyer S, Taylor E, Brammer M, Williams SC, Simmons A, et al. การทำงานของ frontalisation กับอายุ: การทำแผนที่เส้นทางการพัฒนาระบบประสาทกับ fMRI ประสาทวิทยาศาสตร์และชีวจิตรีวิว 2000;24: 13 19- [PubMed]
- Schlaggar BL, TT สีน้ำตาล, Lugar HM, Visscher KM, Miezin FM, Petersen SE ฟังก์ชั่นความแตกต่างทางประสาทวิทยาระหว่างผู้ใหญ่และเด็กวัยเรียนในการประมวลผลคำเดียว วิทยาศาสตร์ 2002;296: 1476 1479- [PubMed]
- Schultz W. ทฤษฎีพฤติกรรมและสรีรวิทยาของรางวัล ความคิดเห็นประจำปีของจิตวิทยา 2006;57: 87 115-
- Sowell ER, Peterson BS, Thompson PM, ยินดีต้อนรับ SE, Henkenius AL, Toga AW การทำแผนที่การเปลี่ยนแปลงของเยื่อหุ้มสมองในช่วงชีวิตมนุษย์ ธรรมชาติประสาทวิทยา 2003;6: 309 315-
- Sowell ER, Thompson PM, Holmes CJ, Jernigan TL, Toga AW ในหลักฐานการวิฟสำหรับการเจริญเติบโตของสมองหลังวัยรุ่นในภูมิภาคหน้าผากและ striatal ธรรมชาติประสาทวิทยา 1999;2: 859 861-
- Sowell ER, Thompson PM, Toga AW การทำแผนที่การเปลี่ยนแปลงในเยื่อหุ้มสมองของมนุษย์ตลอดช่วงชีวิต ประสาทวิทยา 2004;10: 372 392- [PubMed]
- หอก LP สมองวัยรุ่นและอาการทางพฤติกรรมที่เกี่ยวข้องกับอายุ ประสาทวิทยาศาสตร์และชีวจิตรีวิว 2000;24: 417 463- [PubMed]
- Steinberg L. รับความเสี่ยงในวัยรุ่น: การเปลี่ยนแปลงอะไรและทำไม? พงศาวดารของ New York Academy of Sciences 2004;1021: 51 58- [PubMed]
- Steinberg L. รับความเสี่ยงในวัยรุ่น: มุมมองใหม่จากสมองและวิทยาศาสตร์พฤติกรรม ทิศทางปัจจุบันทางวิทยาศาสตร์จิตวิทยา. 2007;16: 55 59-
- Steinberg L. มุมมองทางประสาทวิทยาศาสตร์ทางสังคมเกี่ยวกับการรับความเสี่ยงของวัยรุ่น รีวิวการพัฒนา 2008;28: 78 106- [บทความฟรี PMC] [PubMed]
- Tamm L, Menon V, Reiss AL ความสมบูรณ์ของการทำงานของสมองที่เกี่ยวข้องกับการยับยั้งการตอบสนอง วารสารของสถาบันจิตเวชศาสตร์เด็กและวัยรุ่นอเมริกัน 2002;41: 1231 1238- [PubMed]
- Thomas KM, Hunt RH, Vizueta N, Sommer T, Durston S, Yang Y, และคณะ หลักฐานของความแตกต่างของพัฒนาการในการเรียนรู้ลำดับโดยนัย: การศึกษา FMRI ของเด็กและผู้ใหญ่ วารสารประสาทวิทยาศาสตร์ 2004;16: 1339 1351- [PubMed]
- Turkeltaub PE, Gareau L, ดอกไม้ DL, Zeffiro TA, Eden GF การพัฒนากลไกทางประสาทสำหรับการอ่าน ธรรมชาติประสาทวิทยา 2003;6: 767 773-
- Volkow ND, Li TK การติดยาเสพติด: ชีววิทยาของพฤติกรรมผิดไป รีวิวจากธรรมชาติ Neuroscience 2004;5: 963 970-
- Yurgelun-Todd D. การเปลี่ยนแปลงทางอารมณ์และความรู้ความเข้าใจในช่วงวัยรุ่น ความคิดเห็นปัจจุบันทางชีววิทยา 2007;17: 251 257- [PubMed]
- Zald DH, Boileau I, El-Dearedy W, Gunn R, McGlone F, Dichter GS, และคณะ การส่งโดปามีนใน striatum ของมนุษย์ในระหว่างงานให้รางวัลทางการเงิน วารสารประสาทวิทยา 2004;24: 4105 4112- [PubMed]
