 Martin Zack
Martin Zack- 1ห้องปฏิบัติการองค์ความรู้จิตวิทยา, แผนกประสาท, ศูนย์การติดและสุขภาพจิต, โตรอนโต, ON, แคนาดา
- 2ภาควิชาจิตเวชศาสตร์คณะแพทยศาสตร์มหาวิทยาลัยเพนซิลเวเนียฟิลาเดลเฟียสหรัฐอเมริกา
- 3แผนกชีวจิต, แผนกประสาท, ศูนย์การติดและสุขภาพจิต, โตรอนโต, ON, แคนาดา
ติดยาเสพติดถือว่าเป็นโรคทางสมองที่เกิดจากการสัมผัสกับยาเสพติดเรื้อรัง อาการแพ้ของระบบสมองโดปามีน (DA) บางส่วนไกล่เกลี่ยผลกระทบนี้ การพนันทางพยาธิวิทยา (PG) ถือเป็นการติดพฤติกรรม ดังนั้น PG อาจเกิดจากการเล่นการพนันเรื้อรัง การระบุความไวที่เกิดจากการพนันของระบบ DA จะสนับสนุนความเป็นไปได้นี้ รางวัลการพนันทำให้เกิดการเปิดตัว DA ตอนหนึ่งของการเล่นสล็อตแมชชีนเปลี่ยนการตอบสนอง DA จากการส่งมอบรางวัลไปยังการโจมตีของคิว (วงล้อหมุน) เพื่อรับรางวัลตามหลักการเรียนรู้ที่แตกต่างกันชั่วคราว ดังนั้นสิ่งเร้าที่มีเงื่อนไข (CS) จึงมีบทบาทสำคัญในการตอบสนอง DA ต่อการพนัน ในไพรเมตการตอบสนอง DA ต่อ CS นั้นแข็งแกร่งที่สุดเมื่อความน่าจะเป็นรางวัลเป็น 50% ภายใต้กำหนดการนี้ CS นำความคาดหวังของรางวัลออกมาให้ แต่ไม่ได้ให้ข้อมูลว่าจะเกิดขึ้นในการทดลองที่กำหนดหรือไม่ ในระหว่างการเล่นการพนัน 50% กำหนดการควรลุ้น DA สูงสุด สิ่งนี้จับคู่ความถี่รางวัล (46%) บนเครื่องสล็อตเชิงพาณิชย์อย่างใกล้ชิด การปลดปล่อย DA สามารถก่อให้เกิดอาการแพ้ได้โดยเฉพาะอย่างยิ่งแอมเฟตามีน การได้รับ CS อย่างเรื้อรังที่ทำนายรางวัล 50% ของเวลาอาจเลียนแบบเอฟเฟกต์นี้ เราทดสอบสมมติฐานนี้ในสามการศึกษาด้วยหนู สัตว์ได้รับการเปิดเผย 15 × 45 นาทีให้กับ CS ซึ่งคาดการณ์รางวัลด้วยความน่าจะเป็น 0, 25, 50, 75 หรือ 100% CS เป็นแสงสว่าง รางวัลคือโซลูชันซูโครส 10% หลังการฝึกซ้อมหนูได้รับระบบการกระตุ้นอาการแพ้ในปริมาณที่แตกต่างกันห้าปริมาณ (1 mg / kg) ของ d-amphetamine ในที่สุดพวกเขาก็ได้รับการท้าทายยาบ้า 0.5 หรือ 1 mg / kg ก่อนที่จะทำการทดสอบฤทธิ์ของ 90-min locomotor ในการศึกษาทั้งสามกลุ่ม 50% แสดงกิจกรรมที่มากกว่ากลุ่มอื่นเพื่อตอบสนองต่อปริมาณความท้าทายทั้งสอง ขนาดของเอฟเฟกต์นั้นเรียบง่าย แต่สอดคล้องกันซึ่งสะท้อนโดยกลุ่มที่มีความสำคัญ×อันดับความสัมพันธ์ (ϕ = 0.986, p = 0.025) การเปิดรับสิ่งเร้าที่คาดเดายากเช่นการเล่นการพนันในระยะยาวสามารถกระตุ้นให้เกิดอาการแพ้ยาบ้าได้เช่นเดียวกับแอมเฟตามีน
บทนำ
ติดยาเสพติดมีลักษณะเป็นโรคสมองที่เกิดจากการสัมผัสเรื้อรังกับยาเสพติดของการละเมิด (Leshner, 1997) ความคิดแบบ Neuroplasticity เพื่อเป็นสื่อกลางในการรับรู้ถึงผลกระทบดังกล่าว (Nestler, 2001) การทำให้ไวของระบบสมองโดปามีน (DA) เป็นรูปแบบหนึ่งของการทำปฏิกิริยากับระบบประสาทซึ่งเกี่ยวข้องกับการกระตุ้นปฏิกิริยาต่อสิ่งกระตุ้น (CS) สำหรับยาเสพติดRobinson และ Berridge, 2001) การแพ้ถูกกำหนดโดยการเพิ่มการปลดปล่อย DA เพื่อตอบสนองต่อ CS สำหรับการให้รางวัลและโดยการเพิ่มการเคลื่อนไหวของการตอบสนองทางเภสัชวิทยาต่อความท้าทายทางเภสัชวิทยา (Robinson และ Berridge, 1993; เพียร์ซและคาลิวาส 1997; Vanderschuren และ Kalivas, 2000) แม้ว่าการแพ้จะเป็นเพียงหนึ่งในการเปลี่ยนแปลงของสมองที่เชื่อมโยงกับการเสพติด (cf. Robbins และ Everitt, 1999; Koob และ Le Moal, 2008) การเปลี่ยนแปลงในการปลดปล่อยโดพามีนได้รับการแนะนำให้เป็นตัวแทนของระบบประสาทที่เกี่ยวข้องกับการค้นหายาเสพติดที่ใช้ยาเสพติด (เช่นการกำเริบของโรค) ในยาเสพติดที่ชักนำให้เกิดความไวต่อการเคลื่อนไหว ยังทำให้เกิดการคืนสถานะของการตอบสนองของผู้ปฏิบัติการดับเฮโรอีนหรือโคเคนด้วยตนเอง - รูปแบบสัตว์ของการกำเริบของโรค (Vanderschuren และคณะ, 1999) หลักฐานที่แสดงว่าการกระตุ้นให้เกิดอาการแพ้ (มูลค่าที่เพิ่มขึ้นของการให้รางวัลยา) เป็นสิ่งที่เด่นชัดที่สุดหลังจากการสัมผัสกับยาเสพติดในระยะเริ่มต้นแสดงให้เห็นว่าอาจเกิดอาการแพ้ได้ในระยะแรกของการติดเช่นกันVanderschuren และ Pierce, 2010).
การพนันทางพยาธิวิทยา (PG) ได้รับการอธิบายว่าติดยาเสพติดพฤติกรรมและจัดประเภทใหม่เมื่อเร็ว ๆ นี้ในประเภทเดียวกับความผิดปกติของการพึ่งพาสารในรุ่น 5th ของคู่มือการวินิจฉัยและสถิติของความผิดปกติทางจิต (Frascella และคณะ, 2010; APA, 2013) นี่ก็หมายความว่า PG อาจเกิดจากการสัมผัสอย่างเรื้อรังต่อกิจกรรมการพนันซึ่งกลไกทั่วไปอาจไกล่เกลี่ยผลกระทบของการพนันและการสัมผัสยา (Zack และ Poulos, 2009; Leeman และ Potenza, 2012); และการทำให้ไวต่อการกระตุ้นเซลล์สมองในสมองอาจเป็นองค์ประกอบสำคัญอย่างหนึ่งของกระบวนการนี้
หลักฐานทางคลินิกสนับสนุนความเป็นไปได้ทางอ้อมนี้: การใช้เอกซเรย์ปล่อยโพซิตรอน (PET) Boileau และเพื่อนร่วมงานพบว่ากลุ่ม PG เพศชายแสดงการปลดปล่อย DA ที่มีนัยสำคัญมากขึ้นในการตอบสนองต่อแอมเฟตามีน (0.4 mg / kg)Boileau และคณะ, 2013) ความแตกต่างของกลุ่มโดยรวมมีความสำคัญในการเชื่อมโยงและ somatosensory striatum ใน strib limbic ซึ่งรวมถึงนิวเคลียส accumbens กลุ่มไม่แตกต่างกัน อย่างไรก็ตามในวิชาของ PG การปลดปล่อย DA ใน limbic striatum มีความสัมพันธ์โดยตรงกับความรุนแรงของอาการ PG การค้นพบเหล่านี้สอดคล้องกับการทำให้ไวต่อการกระตุ้นสมองในทางเดินหายใจใน PG แต่ยังแนะนำความแตกต่างที่สำคัญบางอย่างกับบุคคลที่ขึ้นอยู่กับสารของมนุษย์ ไม่เหมือนกับอาสาสมัคร PG และสัตว์ที่สัมผัสกับแอมเฟตามีนในปริมาณต่ำ (เปรียบเทียบ Robinson และคณะ, 1982) มนุษย์ที่อาศัยสารเสพติดมีการลดลงของการปลดปล่อย DA อย่างต่อเนื่องเพื่อความท้าทาย (Volkow et al., 1997; Martinez และคณะ, 2007) และหลักฐานจากสัตว์แสดงให้เห็นว่าสิ่งนี้อาจสะท้อนถึงการขาดดุลในการทำงานของ DA ในช่วงเริ่มต้นของการเลิกบุหรี่ตามรูปแบบของการใช้สารเสพติด (Mateo et al., 2005) ในการศึกษาที่แสดงให้เห็นถึงความไวต่อสารกระตุ้นในสัตว์มักจะพบการปลดปล่อย DA ที่ปรับปรุงแล้วใน limbic striatum แทนที่จะเป็นหลัง (เชื่อมโยง somatosensory) striatum (Vezina, 2004) อย่างไรก็ตามการกระตุ้นด้วยคิว (เช่นการปรับเงื่อนไข) การค้นหายาเสพติดในสัตว์ที่สัมผัสกับโคเคนซ้ำ ๆ นั้นมีการเชื่อมโยงกับการปลดปล่อย DA ที่เพิ่มขึ้นใน dorsal striatum ซึ่งเป็นผลมาจากความคิดที่บ่งบอกถึงพฤติกรรมที่เป็นแรงจูงใจIto et al., 2002) ดังนั้นความสูงโดยรวมในการเปิดตัว DA ในภูมิภาคหลังในอาสาสมัคร PG อาจเกี่ยวข้องกับการแสวงหารางวัลตามนิสัย (ไม่ยืดหยุ่น, เป็นกิจวัตร) ที่เกี่ยวข้องกับการค้นหาEveritt และ Robbins, 2005, p. 1481) ในขณะที่การปลดปล่อย DA ขึ้นอยู่กับความรุนแรงใน limbic striatum ในอาสาสมัครเหล่านี้อาจสอดคล้องกันอย่างใกล้ชิดกับการกระตุ้นให้เกิดแรงจูงใจตามแบบอย่างในสัตว์ การค้นพบ PET ไม่สามารถเปิดเผยได้ว่าปฏิกิริยาไฮเปอร์ไวเกินของ DA เป็นคุณสมบัติที่มีอยู่ก่อนของวิชา PG เหล่านี้ซึ่งเป็นผลมาจากการได้รับการพนันหรือเป็นผลมาจากกระบวนการอื่นทั้งหมด เพื่อตอบคำถามนี้มีความจำเป็นที่จะต้องแสดงให้เห็นถึงการเหนี่ยวนำให้เกิดอาการแพ้จากการพนันอย่างต่อเนื่องในวิชาที่เป็นเรื่องปกติก่อนที่จะได้รับสัมผัส นี่ทำให้เกิดคำถามว่าคุณลักษณะของการพนันมีแนวโน้มที่จะทำให้เกิดอาการแพ้ได้อย่างไร
สกินเนอร์ตั้งข้อสังเกตว่าตารางการเสริมกำลังที่ผันแปรเป็นพื้นฐานของเสน่ห์ของการพนัน (หรืออย่างน้อยก็คงอยู่) (สกินเนอร์ 1953) พฤติกรรมการเดิมพันในเกมสล็อตแมชชีนสอดคล้องกับหลักการพื้นฐานของการปรับสภาพอุปกรณ์ตามที่สะท้อนจากความสัมพันธ์ที่คาดหวังระหว่างการจ่ายเงินและขนาดการเดิมพันในการหมุนติดต่อกัน (Tremblay และคณะ 2011) ดังนั้นผู้ปฏิบัติงานอัตราส่วนตัวแปรตอบสนองที่ปรากฏขึ้นเพื่อให้รูปแบบการพนันสล็อตแมชชีนที่ถูกต้องจากภายนอก
การวิจัยเมื่อเร็ว ๆ นี้กับสัตว์ให้การสนับสนุนขั้นต้นที่แข็งแกร่งสำหรับผลกระทบเชิงสาเหตุของการพนันที่มีต่อการแพ้ นักร้องและเพื่อนร่วมงานตรวจสอบผลของ 55 1 – h เซสชันรายวันแบบคงที่ (FR20) หรือตัวแปร (VR20) saccharin เสริมแรงในกระบวนทัศน์คันกดแบบกดต่อการเคลื่อนไหวของแอมเฟตามีนในปริมาณต่ำ (0.5 mg / kg) Sprague Dawley) หนู (นักร้องและคณะ, 2012) พวกเขาตั้งสมมติฐานว่าหากการพนันนำไปสู่การแพ้หนูที่สัมผัสกับตารางตัวแปรซึ่งเลียนแบบการพนันควรแสดงการตอบสนองต่อแอมเฟตามีนมากกว่าหนูที่สัมผัสกับตารางที่แน่นอน ตามที่คาดการณ์ไว้กลุ่ม VR20 จะแสดงการตอบสนองของแอมเฟตามีนที่ดีกว่ากลุ่ม FR50 20% ในทางตรงกันข้ามกลุ่มแสดงการเคลื่อนไหวที่เท่าเทียมกันหลังจากฉีดน้ำเกลือ การค้นพบเหล่านี้ยืนยันว่าการได้รับการเสริมแรงแบบแปรผันนั้นเพียงพอที่จะกระตุ้นให้เกิดปฏิกิริยารุนแรงต่อความท้าทาย DA ในสัตว์ที่มีสุขภาพแบบสุ่มตามตารางเวลาที่เกี่ยวข้อง
มีคำถามจำนวนมากที่เกิดขึ้นจากผลลัพธ์นี้: ประการแรกการรับรู้ถึงสิ่งที่อาจเกิดขึ้นในระดับใดหรือขาดในระหว่างการตอบกลับของผู้ดำเนินการ ในแง่การเรียนรู้ผลกระทบนี้เกี่ยวข้องกับ“ ความคาดหวังผลการตอบสนอง” หรืออาจมีผลคล้ายกันในกรณีที่ไม่มีการตอบสนองของผู้ปฏิบัติงานคือ“ ความคาดหวังจากการกระตุ้นผล” ในกระบวนทัศน์ Pavlovian (cf. Bolles, 1972)? ประการที่สองระดับของความไม่แน่นอนระหว่างเหตุการณ์ก่อนเกิด (การตอบสนองหรือสิ่งเร้า) และผลลัพธ์ของมันมีอิทธิพลต่อระดับความรู้สึกไวหรือไม่?
คำถามที่สองเกี่ยวข้องกับบทบาทของความไม่แน่นอนในการทำให้แพ้ ตัวอย่างเช่นเกมที่มีผลการสุ่มอย่างแท้จริง - คาดเดาไม่ได้อย่างสมบูรณ์ - มีศักยภาพมากกว่าที่จะชักจูงให้เกิดความรู้สึกไวกว่าเกมที่มีการกำหนดอัตราต่อรองในการชนะอย่างชัดเจน แต่ไม่ใช่การสุ่ม การวิจัยในปัจจุบันตอบคำถามเหล่านี้
การออกแบบการทดลองได้รับแจ้งจากการศึกษาน้ำเชื้อเกี่ยวกับความคาดหวังของรางวัลและการตอบสนองของเซลล์ประสาท DA ในลิง (Fiorillo et al., 2003) สัตว์ในการศึกษานั้นได้รับรางวัลน้ำผลไม้ (US) ภายใต้ 0, 25, 50, 75 หรือ 100% ตารางอัตราตัวแปร กำหนดการถูกกำหนดโดย 1 ของ 4 CS ที่แตกต่างกัน (ไอคอน) กำหนดการ 0% ให้รางวัลบ่อยเท่าที่กำหนดการ 100% แต่ไม่รวม CS อัตราการยิงของเซลล์ประสาท DA ในช่วงเวลาระหว่างการโจมตี CS และการส่งหรือการละเลยของสหรัฐฯเป็นมาตรการที่สำคัญ การศึกษาพบว่าการตอบสนอง DA เพิ่มขึ้นเนื่องจากความไม่แน่นอนของการส่งมอบรางวัล ดังนั้นภายใต้กำหนดการ 100% CS ทำให้เกิดกิจกรรมเล็ก ๆ น้อย ๆ ภายใต้ตารางเวลา 25 และ 75% CS ทำให้เกิดกิจกรรมระดับปานกลางและคล้ายคลึงกันและภายใต้ตาราง 50% CS ทำให้เกิดกิจกรรมสูงสุด ในแต่ละกรณีอัตราการยิงเพิ่มขึ้นตลอดช่วงเวลาของ CS-US กล่าวคือเมื่อการคาดการณ์เริ่มขึ้น
การค้นพบเหล่านี้บ่งชี้ว่ากิจกรรม DA ไม่เพียง แต่จะแตกต่างกันไปตามว่ารางวัลนั้นแน่นอนหรือไม่ (อัตราส่วนคงที่) หรือไม่แน่นอน (Variable Ratio) แต่ยังแปรผันตามสัดส่วนผกผันกับปริมาณข้อมูลเกี่ยวกับการส่งมอบรางวัลโดย CS ในสภาพ 100% CS กระตุ้นความคาดหวังของรางวัลและคาดการณ์การส่งมอบของมันอย่างสมบูรณ์แบบ ในเงื่อนไข 25 และ 75% CS กระตุ้นความคาดหวังและคาดการณ์การส่งมอบรางวัลสามในสี่ครั้ง ในเงื่อนไข 50% CS ทำให้เกิดความคาดหวัง แต่ไม่ได้ให้ข้อมูลเกี่ยวกับการส่งมอบรางวัลเกินโอกาสเพียงอย่างเดียว จากการค้นพบของพวกเขา Fiorillo และคณะ สรุป:“ การเพิ่มโดปามีนในความไม่แน่นอนนี้อาจส่งผลต่อคุณสมบัติที่มีคุณค่าของการพนัน” (p. 1901)
ผลของรางวัลผันแปร 50% ในเซสชันเดียวไม่ควรเปลี่ยนแปลงในช่วงหลายเซสชันเนื่องจากความเป็นไปได้ที่จะได้รับรางวัลยังคงไม่สามารถคาดเดาได้อย่างสิ้นเชิงในทุกการทดลองใช้ ดังนั้นเมื่อพิจารณาเงื่อนไขที่จะเพิ่มการเปิดใช้งานเซลล์ประสาท DA อย่างเรื้อรังในช่วงการพนันซ้ำ ๆ ตารางเวลา 50% ควรทำให้เกิดผลที่ยั่งยืนและมีประสิทธิภาพมากที่สุด สิ่งนี้เป็นที่น่าสังเกตเนื่องจากอัตราผลตอบแทนระยะยาว (ผลตอบแทน> 0) ที่สังเกตได้จากการหมุนหลายพันครั้งในเครื่องสล็อตเชิงพาณิชย์คือ 45.8% (Tremblay และคณะ 2011) ดังนั้นรางวัลตัวแปร 50% จะปรากฏขึ้นตามตารางการจ่ายผลตอบแทนที่ถูกต้องโดยอุปกรณ์การพนันที่เกิดขึ้นจริง
การศึกษาในปัจจุบันใช้ตารางการปรับสภาพเดียวกันกับ Fiorillo et al ในการสัมผัสเรื้อรังการออกแบบระหว่างกลุ่มกับหนู สัตว์ได้รับการปรับสภาพประจำวันประมาณ 3 สัปดาห์โดยที่ CS (แสง) จับคู่กับสหรัฐอเมริกา (ซูโครสจำนวนเล็กน้อย) หลังจากขั้นตอนการฝึกสัตว์จะพักก่อนการประเมินความไวต่อการตอบสนองของขมิ้นอ้อยต่อแอมเฟตามีน จากวรรณกรรมพบว่าหนูที่ได้รับรางวัลตารางการให้รางวัลที่แตกต่างกันจะไม่แตกต่างกันในพฤติกรรมของขมิ้นอ้อยที่ปราศจากยา แต่จะแสดงระดับการเคลื่อนที่ที่แตกต่างกันอย่างมีนัยสำคัญหลังจากแอมเฟตามีนโดยกลุ่ม 50% แสดงการตอบสนองของขมิ้นอ้อยต่อยามากกว่าเมื่อเทียบกับ กลุ่มอื่น ๆ ในช่วงของปริมาณซึ่งเป็นรูปแบบที่คาดว่าจะเกิดขึ้นหากสัตว์ 50% เคยสัมผัสกับแอมเฟตามีนในปริมาณที่เพิ่มขึ้นมาก่อน (เช่นการแพ้ข้าม)
1 ทดลอง
วัสดุและวิธีการ
Subjects
สี่กลุ่ม (n = 8 / กลุ่ม) ของผู้ใหญ่ (300 – 350 g) หนู Sprague-Dawley (Charles River, St. Constant, Quebec, Canada) ตั้งอยู่แยกกันในกล่องโพลีคาร์บอเนต (20 × 43 × 22 cm) ภายใต้ reverse 12: 12 รอบแสงมืด พวกเขาได้รับ โฆษณาฟรี การเข้าถึงอาหารและน้ำและการจัดการรายวันโดยผู้ทดลองเป็นเวลา 2 สัปดาห์ก่อนการศึกษา แต่ละกลุ่มมีเงื่อนไขภายใต้กำหนดการรางวัลหนึ่งในสี่ตัวแปร: 0, 25, 50 หรือ 100% กลุ่ม 75% ถูกละเว้นในการศึกษาเริ่มต้นนี้เช่น Fiorillo และคณะ (2003) พบว่ามีการวางจำหน่าย post-CS DA ที่เทียบเท่าภายใต้กำหนดการรางวัล 25 และ 75% ซึ่งทั้งสองเงื่อนไขนำไปสู่การเปิดตัว DA มากขึ้นกว่าเงื่อนไข 100% CS-US แต่น้อยกว่าเงื่อนไข 50%
อุปกรณ์
การเข้าถึงงานนำเสนอซูโครสและ CS มีให้เป็นรายบุคคลในกล่องปรับสภาพโอเปอร์เรนเชียล (33 × 31 × 29 ซม.) แต่ละกล่องมีนิตยสาร reinforcer ตั้งอยู่ที่ผนังด้านหน้า แสงที่อยู่ด้านบนของนิตยสารทำหน้าที่เป็น CS ของเหลวกระบวยเหลวแบบใช้มอเตอร์ควบคุมสามารถยกขึ้นไปกองกับพื้นของนิตยสารได้ กิจกรรมในกล่องถูกควบคุมโดยอุปกรณ์และซอฟต์แวร์ของ Med Associates โดยใช้โปรแกรมภายในที่เขียนใน MED-PC การทดสอบหัวรถจักรได้ดำเนินการเป็นรายบุคคลในกรง Plexiglas (27 × 48 × 20 cm) กรงแต่ละหลังมีระบบตรวจสอบซึ่งประกอบด้วยเซลล์โฟโต้ลำแสงหกตัวเพื่อตรวจจับการเคลื่อนไหวในแนวนอน
การรักษาอื่นๆ
การฝึกอบรม. การศึกษาได้ดำเนินการตามแนวทางจริยธรรมที่กำหนดโดยสภาแคนาดาในการดูแลสัตว์ หนูถูก จำกัด อาหาร 90% ของน้ำหนักตัวของพวกเขาในช่วงระยะเวลาของการศึกษาและตั้งอยู่เป็นรายบุคคล หนูแต่ละคนได้รับการฝึกอบรมการให้รางวัลซูโครส 15 วัน (การแก้ปัญหาน้ำ 10% ที่ 0.06 มล. ต่อรางวัล): 5 วันติดต่อกัน× 3 สัปดาห์โดยหยุดสุดสัปดาห์ สัตว์ได้รับการบำรุงรักษาตามมาตรฐานอาหารก่อนและหลังการฝึกอบรม; การได้รับซูโครสนั้น จำกัด เพียงสิบห้า ~ 40 นาทีการฝึกซ้อม แต่ละเซสชั่นในแต่ละวันประกอบด้วยงานนำเสนอการกระตุ้น 15 (แสง; CS) แต่ละคนคั่นด้วยช่วงเวลาระหว่างการทดลองของ 120 s แสงนั้นตั้งอยู่ในแผงด้านบนของนิตยสารและยังคงเปิดอยู่สำหรับ 25 s โดยมีซูโครสให้บริการในช่วง 5 s สุดท้าย ในกรณีของกลุ่ม 0 กระบวยซูโครสจะถูกยกขึ้นทุก 140 s (สำหรับ 5 s) แต่แสงกระตุ้นไม่ได้ส่องสว่าง สิ่งนี้บรรจุช่วงเวลาระหว่างการนำเสนอของกระบวยในกลุ่ม 0 และกลุ่มอื่น ๆ (120 + 25 s) การรักษาแต่ละครั้งใช้เวลา ~ 40 ขั้นต่ำ โดยเฉลี่ยกลุ่ม 25 ได้รับซูโครสหนึ่งครั้งสำหรับทุก ๆ สี่งานนำเสนอ CS กลุ่ม 50 ได้รับซูโครสหนึ่งครั้งสำหรับทุก ๆ สองงานนำเสนอ CS และกลุ่ม 100 ได้รับซูโครสหลังจากนำเสนอ CS ทุกครั้ง
การทดสอบ. สองสัปดาห์หลังจากเซสชันการเข้าถึงซูโครสครั้งสุดท้าย (หรือ“ การปรับสภาพ”) การตอบสนองของหัวรถจักรต่อ d-amphetamine (AMPH; ip) ได้รับการประเมิน หนูได้รับ 2-h สามครั้งเพื่อทำให้เกิดความเคยชินกับกล่องหัวรถจักรตามด้วยการทดสอบ AMPH หกครั้ง วันทดสอบ AMPH เกิดขึ้นที่ช่วงเวลา 1-wk ในวันทดสอบหนูได้รับ 30 นาทีเพื่อทำให้เกิดความเคยชินกับกล่องจากนั้นได้รับ 0.5 mg / kg ปริมาณของ AMPH ที่ตามมาแยกเป็นรายสัปดาห์โดยแบ่งออกเป็นห้า 1.0 mg / kg (หนึ่งครั้งต่อวัน) ในวันทดสอบ 1 ผ่าน 5 . โพสต์ AMPH locomotion ถูกประเมิน 90 ขั้นต่ำในแต่ละเซสชั่น
วิธีการวิเคราะห์ข้อมูล
การวิเคราะห์ทางสถิติได้ดำเนินการกับ SPSS (v. 16 และ v. 21; SPSS Inc. , Chicago IL) การตอบสนองพฤติกรรมทันทีต่อ CS ได้รับการประเมินในแง่ของการเจาะจมูกเข้าไปในรูรับแสงที่มีการจ่ายซูโครส จำนวนเฉลี่ยของการเจาะจมูกในช่วงเวลานี้ (5 s ต่อการทดลอง) ถูกเปรียบเทียบกับจำนวนเฉลี่ยของการสะกิดจมูกในระยะเวลาเดียวกัน (5 s) เฉลี่ยในช่วงที่ CS ขาดไป Group × Session ANOVAs ของการเจาะจมูกด้วย CS ปัจจุบันและไม่ติดตามการได้มาของการเลือกปฏิบัติที่ตอบสนองต่อการตอบสนองต่อการชี้นำและการพิจารณาคัดจมูกภายใต้ตารางที่แตกต่างกันในระหว่างการฝึกซ้อม 15 ซูโครส
ผลของการรักษาต่อการตอบสนองของหัวรถจักรถูกประเมินด้วย Group × Session ANOVAs สำหรับเฟสการทำให้ไร้ความรู้สึกของยาเสพติด (สามครั้ง), การทำให้ไวต่อยาก่อนกำหนด 0.5 mg / kg AMPH ความท้าทาย (หนึ่งเซสชัน) และระหว่างห้า 1 mg / kg AMPH อาการแพ้เมื่อกลุ่มที่คาดว่าจะแตกต่างกันในการตอบสนองต่อปริมาณที่ซ้ำกันของ AMPH ANOVAs กลุ่ม× Session ยังประเมินการตอบสนองของหัวรถจักรโดยไม่ใช้ยาระหว่างขั้นตอนการทำให้เกิดก่อนการฉีด 30 นาทีขั้นต่ำจากการทดสอบ AMPH แต่ละครั้ง การเปรียบเทียบตามแผนประเมินความแตกต่างของประสิทธิภาพเฉลี่ยสำหรับกลุ่ม 50 กับกลุ่ม 0 (ไม่มีการควบคุมความคาดหวัง) และกลุ่ม 100 (ไม่มีการควบคุมความไม่แน่นอน) โดยใช้วิธีการ t- การทดสอบ (ธรรมด๊าธรรมดา, 1992) ใช้ข้อผิดพลาด MS และข้อกำหนดข้อผิดพลาด df สำหรับเอฟเฟกต์ที่เกี่ยวข้อง (เช่นปฏิสัมพันธ์ระหว่างกลุ่มหรือกลุ่ม×เซสชัน) จาก ANOVA (Winer, 1971) การวิเคราะห์แนวโน้มพหุนามทดสอบโปรไฟล์ของการเปลี่ยนแปลงตลอดระยะเวลาการประชุม
เพื่อตรวจสอบว่าการตอบสนองของวิธีการในการปรากฏตัวและขาดของ CS ในช่วงการฝึกอบรมซูโครส 15 มีส่วนทำให้เกิดการเปลี่ยนแปลงในการตอบสนองของหัวรถจักรเพื่อ AMPH หรือความแตกต่างของกลุ่มสื่อกลางในการตอบสนอง AMPH การวิเคราะห์ติดตามความแปรปรวนร่วม (ANCOVAs) ข้อมูลหัวรถจักรรวมถึงจมูกรวมทั้งหมด (ผลรวมสำหรับเซสชัน 15) เมื่อ CS ไม่อยู่ในรูปของโควาเรียต ผลกระทบที่สำคัญของโควาเรียตจะบ่งบอกว่าการตอบสนองต่อการใช้ยาเสพติดนั้นไม่มีการควบคุม (มีอิทธิพลต่อความแข็งแรงของ) ผลกระทบของกลุ่มหรือเซสชัน การลดลงอย่างมีนัยสำคัญของผลกระทบของกลุ่มหรือเซสชั่นในการปรากฏตัวของ covariate อย่างมีนัยสำคัญจะระบุว่าการตอบสนองวิธีการไกล่เกลี่ย (คิดเป็น) ผลกระทบของกลุ่มหรือเซสชั่น การลดลงของนัยสำคัญของผลกระทบของกลุ่มหรือเซสชั่นในกรณีที่ไม่มีผล covariate อย่างมีนัยสำคัญก็จะสะท้อนให้เห็นถึงการสูญเสียพลังงานทางสถิติเนื่องจากการจัดสรรใหม่ของ df จากคำผิดพลาดไปยัง covariate และจะไม่มีผลต่อการตีความ ผลกระทบของกลุ่มหรือเซสชั่น
ผลสอบ
จมูกทิ่มในระหว่างการปรับสภาพซูโครส
CS ปัจจุบัน. รูป 1A แสดงค่าเฉลี่ยของจมูกโผล่สำหรับกลุ่ม 25, 50, และ 100 ในขณะที่ CS อยู่ในช่วงการปรับสภาพซูโครส 15 (ไม่ได้รับการเข้ารหัสสำหรับกลุ่ม 0 ซึ่งไม่ได้รับ CS) กลุ่ม 3 ×เซสชัน 15 ANOVA ให้ผลหลักที่สำคัญของกลุ่ม F(2, 21) = 5.63, p = 0.011 และเซสชัน F(14, 294) = 14.00, p <0.001 พร้อมกับการโต้ตอบกลุ่ม×เซสชันที่สำคัญ F(28, 294) = 2.93, p <0.001. รูป 1A บ่งชี้ว่าผลกระทบหลักของเซสชันสะท้อนให้เห็นการเพิ่มขึ้นของการเจาะจมูกในเซสชันทั้งสามกลุ่มและผลหลักของกลุ่มสะท้อนคะแนนโดยรวมที่สูงขึ้นโดยทั่วไปในกลุ่ม 100 เทียบกับกลุ่ม 25 ด้วยคะแนนระดับกลางในกลุ่ม 50 การโต้ตอบของกลุ่ม×เซสชันที่สำคัญสำหรับแนวโน้มลูกบาศก์ F(2, 21) = 4.42, p = 0.030 บ่งชี้การเพิ่มขึ้นอย่างรวดเร็วการจุ่มและการปรับระดับในจมูกโผล่มาที่เซสชันในกลุ่ม 100 เมื่อเทียบกับการเพิ่มขึ้นเชิงเส้นในเซสชัน 50 กลุ่มและการเพิ่มขึ้นเชิงเส้นแบบตื้นกว่าเซสชันในกลุ่ม 25
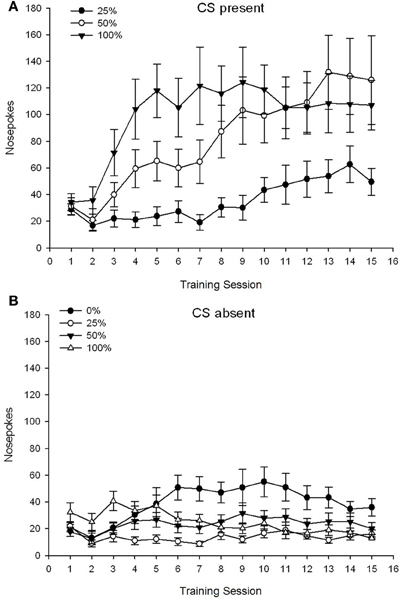
รูปที่ 1 Mean (SE) การตอบสนองวิธีการ (จมูกโผล่) ในการฝึก 15 ซูโครสในกลุ่มของหนู Sprague Dawley (n = 8 / กลุ่ม) ได้รับรางวัลน้ำตาลซูโครส (สารละลาย 10%) ส่งมอบภายใต้ตารางเวลาตัวแปร 0, 25, 50 หรือ 100%. มาตรการกระตุ้นคือแสง (120 s) กลุ่ม 0 ได้รับรางวัลจำนวนเท่ากันกับกลุ่ม 100 ในกรณีที่ไม่มีสิ่งเร้า (A) คะแนนเมื่อ CS มีอยู่ (การทดลอง 5 s × 15) (B) คะแนนเมื่อ CS ขาดงาน (ค่าเฉลี่ยสำหรับ 5 × 15 s ขณะที่แสงดับ)
CS ขาดไป. รูป 1B แสดงค่าเฉลี่ยของจมูกทั้งสี่กลุ่มในช่วงเวลาที่เท่าเทียมกัน (การทดลอง 5 s × 15) เฉลี่ยตลอดช่วงเวลาที่ CS ขาดงาน กลุ่ม 4 ×เซสชัน 15 ANOVA ให้ผลหลักที่สำคัญของกลุ่ม F(3, 28) = 7.06, p = 0.001 และเซสชัน F(14, 392) = 2.84, p <0.001 พร้อมกับการโต้ตอบกลุ่ม×เซสชันที่สำคัญ F(42, 392) = 3.93, p <0.001. ปฏิสัมพันธ์กลุ่ม×เซสชันที่สำคัญสำหรับแนวโน้มกำลังสอง F(3, 28) = 3.91, p = 0.019 พร้อมกับไม่มีการโต้ตอบสำหรับแนวโน้มลูกบาศก์ F(3, 28) <0.93, p > 0.44 ซึ่งสะท้อนให้เห็นโปรไฟล์ของจมูกแบบ "กลับหัว" โผล่เหนือเซสชันในกลุ่ม 0 เมื่อเทียบกับโปรไฟล์ที่มีเสถียรภาพโดยทั่วไปมากกว่าเซสชันในกลุ่มอื่น ๆ
ทำให้คุ้นเคยกับตู้รถไฟ
กลุ่ม 4 × 3 เซสชัน ANOVA ให้ผลหลักของเซสชัน F(2, 56) = 5.67, p = 0.006 และไม่มีผลกระทบที่สำคัญอื่น ๆ F(3, 28) <1.60, p > 0.21 ค่าเฉลี่ยของลำแสงแตกต่อ 2 ชั่วโมงในกล่องหัวรถจักรคือ 1681 (123) ในเซสชั่นที่ 1, 1525 (140) ในเซสชั่นที่ 2 และ 1269 (96) ในเซสชั่นที่ 3 การเปรียบเทียบตามแผนไม่พบความแตกต่างอย่างมีนัยสำคัญระหว่างกลุ่ม 50 และกลุ่ม 0 หรือกลุ่ม 100 ในเซสชันการทำให้คุ้นเคยครั้งแรกหรือครั้งสุดท้าย t(84) <1.69, p > 0.05 ดังนั้นในกรณีที่ไม่มี AMPH การได้รับกล่องทดสอบซ้ำ ๆ จึงสัมพันธ์กับการลดลงอย่างสม่ำเสมอของกิจกรรมขมิ้นอ้อยที่เกิดขึ้นเองใน XNUMX กลุ่ม (เช่นผลของเซสชัน) และไม่มีการตอบสนองที่แตกต่างกันตามตารางการฝึกของซูโครส (ไม่มีการโต้ตอบ) .
ช่วงทดสอบ
ผลกระทบของการแพ้ก่อนกำหนด 0.5 mg / kg AMPH ท้าทาย.
การเคลื่อนที่ก่อนฉีด. กลุ่มการวิเคราะห์ความแปรปรวนทางเดียวของกลุ่ม 4 กลุ่มของการตอบสนองของหัวรถจักรระหว่างขั้นตอนการทำให้เกิดความเคยก่อนฉีด 30-นาทีให้ผลอย่างไม่มีนัยสำคัญ F(3, 28) <1.05, p > 0.38. การเปรียบเทียบตามแผนไม่พบความแตกต่างอย่างมีนัยสำคัญระหว่างกลุ่ม 50 และกลุ่ม 0 หรือกลุ่ม 100 t(32) <0.87, p > 0.40. ดังนั้นความแตกต่างพื้นฐานของการเคลื่อนที่ก่อนการฉีดไม่ได้อธิบายถึงความแตกต่างของกลุ่มในการตอบสนองของหัวรถจักรต่อ AMPH คานเฉลี่ย (SE) แตกสำหรับตัวอย่างคือ 559 (77)
การเคลื่อนที่หลังฉีดกับการพักตัวครั้งสุดท้ายโดยไม่ต้องใช้ยา. การวิเคราะห์ความแปรปรวนของเซสชัน 4 กลุ่ม× 2 เปรียบเทียบการตอบสนองของหัวรถจักรในเซสชันการทำให้เกิดความเคยชินขั้นสุดท้ายและทันทีหลังจากการทดสอบ AMPH ก่อนการทำให้เกิดอาการแพ้ 0.5 มก. / กก. คะแนนสำหรับเซสชันความเคยชิน (120 นาที) ได้รับการปรับขนาดให้สอดคล้องกับระยะเวลาของเซสชันการทดสอบ AMPH (90 นาที) (คะแนนความเคยชินดิบ× 90/120) การวิเคราะห์ทำให้เกิดผลกระทบหลักที่สำคัญของเซสชัน F(1, 28) = 34.16, p <0.001 และไม่มีผลกระทบที่สำคัญอื่น ๆ F(3, 28) <2.26, p > 0.10. ผลของเซสชันสะท้อนให้เห็นการเพิ่มขึ้นของการแตกของลำแสงเฉลี่ย (SE) ในการตอบสนองต่อปริมาณจาก 952 (72) เป็น 1859 (151) การเปรียบเทียบตามแผนไม่พบความแตกต่างอย่างมีนัยสำคัญระหว่างกลุ่ม 50 และกลุ่ม 0 หรือกลุ่ม 100 ในการตอบสนองต่อขนาดยา t(56) <1.72, p > 0.10. อย่างไรก็ตามลำดับอันดับของคะแนนการแตกของลำแสง (M; SE) สอดคล้องกับสมมติฐาน: กลุ่ม 50 (2205; 264)> กลุ่ม 0 (2025; 203)> กลุ่ม 100 (1909; 407)> กลุ่ม 25 (1296; 299) .
ผลของ 1 mg / kg AMPH.
การเคลื่อนที่ก่อนฉีด. 4 Group × 5 Session ANOVA ของการตอบสนองของ locomotor ในช่วงก่อนการฉีดพ่น 30-min ขั้นตอนก่อนหน้านี้ในการทดสอบ 1 mg / kg AMPH ให้ผลหลักของเซสชัน F(4, 112) = 43.64, p <0.0001 และไม่มีผลกระทบที่สำคัญอื่น ๆ F(3, 28) <0.97, p > 0.42 การเปรียบเทียบตามแผนไม่พบความแตกต่างอย่างมีนัยสำคัญระหว่างกลุ่ม 50 และกลุ่ม 0 หรือกลุ่ม 100 ในเซสชันการทดสอบครั้งแรกหรือครั้งสุดท้าย t(140) <0.84, p > 0.30. ดังนั้นความแตกต่างพื้นฐานของการเคลื่อนที่ไม่ได้อธิบายถึงความแตกต่างของกลุ่มในการตอบสนองของหัวรถจักรต่อ AMPH คะแนนการแบ่งลำแสงเฉลี่ย (SE) สำหรับระยะความเคยชินก่อนให้ยาในช่วงที่ 1–5 ได้แก่ 454 (30), 809 (53), 760 (36), 505 (35), 756 (39)
การเคลื่อนที่หลังฉีด. รูป 2 แสดงผลของการฉีดยา 1 มก. / กก. AMPH (ห้าครั้งต่อสัปดาห์) ห้าครั้งต่อคะแนนการเคลื่อนไหวของหัวรถจักรในสี่กลุ่ม กลุ่ม 4 × 5 เซสชัน ANOVA ให้ผลหลักของเซสชัน F(4, 112) = 8.21, p <0.001 ผลกระทบหลักเล็กน้อยของกลุ่ม F(2, 45) = 3.28, p = 0.085 และไม่มีการโต้ตอบที่สำคัญ F(12, 122) <0.77, p > 0.68
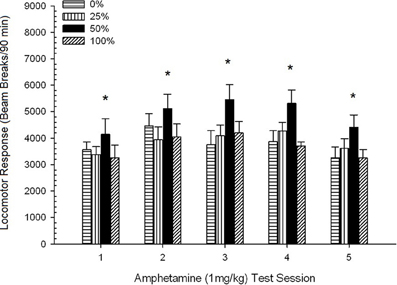
รูปที่ 2 การตอบสนองของหัวรถจักรเฉลี่ย (SE) (จำนวนการหยุดลำแสงในอาร์เรย์อิเล็กทรอนิกส์ต่อ 90 ขั้นต่ำ) ถึง 1 mg / kg d-amphetamine (ip) ในการประชุม 5 รายสัปดาห์ในกลุ่ม Sprague Dawley rats (n = 8 / กลุ่ม) เปิดเผยก่อนหน้านี้ในเซสชันการปรับสภาพ 15 รายวันด้วยการให้รางวัลซูโครส (โซลูชัน 10%) ส่งมอบภายใต้ 0, 25, 50, หรือ 100% ตารางเวลาตัวแปร. มาตรการกระตุ้นคือแสง (120 s) กลุ่ม 0 ได้รับรางวัลจำนวนเท่ากันกับกลุ่ม 100 ในกรณีที่ไม่มีสิ่งเร้า * * * *p <0.05 สำหรับค่าเฉลี่ยความแตกต่างระหว่างกลุ่ม 50 และกลุ่ม 0 และกลุ่ม 100 ตามการเปรียบเทียบที่วางแผนไว้
การเปรียบเทียบตามแผนเปิดเผยว่าคะแนนกลุ่ม 50 แตกต่างจากกลุ่ม 0 อย่างมาก t(14) = 2.19, p = 0.037 และกลุ่ม 100 t(14) = 2.36, p = 0.025 [และแตกต่างเล็กน้อยจากกลุ่ม 25 t(14) = 2.03, p = 0.051] ดังนั้นในกลุ่ม 50 locomotor ตอบสนองต่อ 1 mg / kg AMPH เกินความน่าเชื่อถือของอีกสามกลุ่มในช่วงการทดสอบทั้งห้าครั้ง การวิเคราะห์แนวโน้มพหุนามตรวจพบแนวโน้มกำลังสองที่สำคัญตลอดช่วง F(1, 28) = 32.47, p <0.0001 และไม่มีแนวโน้มสำคัญอื่น ๆ F(1, 28) <1.78, p > 0.19 รูป 2 แสดงให้เห็นว่าผลลัพธ์นี้สะท้อนให้เห็นถึงรูปแบบ "ฤvertedษี U" ในช่วงการประชุม
การควบคุมความแปรปรวนของการกระตุ้นจมูกตอบสนองระหว่างการฝึกซูโครส
ANCOVA ติดตามผลของการเคลื่อนไหวของการตอบสนองต่อ 1 mg / kg AMPH กับจมูกโผล่ (CS ปัจจุบัน) เป็น covariate ในสามกลุ่มที่ได้รับ CS ให้ผลกระทบหลักของกลุ่ม F(2, 20) = 3.07, p = 0.069 และไม่มีผลกระทบที่เกี่ยวข้องกับความแปรปรวนร่วมอย่างมีนัยสำคัญ F(4, 80) <0.05, p > 0.85 ดังนั้นวิธีการตอบสนองแบบ cued ในระหว่างการฝึกอบรมไม่ได้อธิบายถึงการเปลี่ยนแปลงที่มีนัยสำคัญในการตอบสนองของหัวรถจักรต่อ 1 มก. / กก. AMPH ในกลุ่ม 25, 50 หรือ 100
ANCOVA ติดตามผลของการเคลื่อนไหวของการตอบสนองต่อ 1 mg / kg AMPH กับจมูกโผล่ (CS ขาด) เป็น covariate ให้ผลสำคัญของ covariate F(1, 27) = 6.17, p = 0.020 ซึ่งเป็นผลหลักที่สำคัญของกลุ่ม F(3, 27) = 4.13, p = 0.016, เซสชั่นชายขอบ×ปฏิสัมพันธ์แบบ Covariate, p = 0.080 และไม่มีผลกระทบที่สำคัญอื่น ๆ F(4, 108) <1.48, p > 0.21 ดังนั้นวิธีการตอบสนองที่ไม่ได้รับการดูแล (ไม่เลือกปฏิบัติ) ในระหว่างการฝึกอบรมอธิบายถึงการเปลี่ยนแปลงที่สำคัญในการตอบสนองของหัวรถจักรต่อ 1 มก. / กก. AMPH อย่างไรก็ตามรูปแบบนี้ไม่ทับซ้อนกับความแปรปรวนที่เกี่ยวข้องกับกลุ่มเนื่องจากการรวมความแปรปรวนร่วมในการวิเคราะห์เพิ่มขึ้นแทนที่จะลดความสำคัญของผลกระทบของกลุ่ม
การสนทนา
ข้อมูลกระตุ้นจมูกในขณะที่ CS อยู่ในปัจจุบันแสดงให้เห็นว่ากลุ่มที่ได้รับความสัมพันธ์ระหว่าง CS และการส่งมอบซูโครสสะท้อนให้เห็นจากการตอบสนองที่เพิ่มขึ้นในช่วงการฝึกอบรม โพรไฟล์ของการตอบสนองผ่านเซสชันในขณะที่ CS แนะนำอยู่ว่ากำหนดการ 100 และ 50% CS-US มีประสิทธิภาพเท่ากันในแนวทางที่น่าทึ่งในขณะที่ตาราง 25% นำเสนอวิธีที่เพิ่มขึ้นเล็กน้อย ข้อมูลกระตุ้นจมูกในขณะที่ CS ไม่อยู่แนะนำว่ากลุ่มที่ได้รับตารางการฝึกอบรม CS-sucrose ใด ๆ (กลุ่ม 25, 50, 100) ได้เรียนรู้อย่างรวดเร็วเพื่อลดการเจาะจมูกในกรณีที่ไม่มี CS ในขณะที่สัตว์ในกลุ่ม 0 ซึ่งไม่ได้รับ CS เพียงเรียนรู้ที่จะลดพฤติกรรมวิธีการของพวกเขาในระดับที่ จำกัด หลังจากการฝึกอบรมที่กว้างขวาง
ข้อมูลความเคยชินแสดงให้เห็นว่ากลุ่มไม่แตกต่างกันก่อน AMPH และการเปิดรับซ้ำที่กล่องทดสอบนั้นมีความสัมพันธ์กับการตอบสนองของหัวรถจักรที่ปราศจากยาลดลง ดังนั้นความแตกต่างระหว่างกลุ่มและการตอบสนองที่เพิ่มขึ้นมากกว่าปริมาณที่ซ้ำกันของ AMPH ไม่สามารถนำมาประกอบกับความแตกต่างที่มีอยู่ก่อนในพฤติกรรมของหัวรถจักร
ผลลัพธ์ของความท้าทายก่อนการกระตุ้นด้วย 0.5 mg / kg AMPH ยืนยันว่ายานั้นเพิ่มการเคลื่อนไหวของหัวรถจักรเมื่อเทียบกับวันที่ปราศจากยาขั้นสุดท้าย สอดคล้องกับสมมติฐานกลุ่ม 50 อยู่ในอันดับที่สูงกว่ากลุ่ม 0 หรือ 100 (เช่นเดียวกับกลุ่ม 25) ในแง่ของการตอบสนองต่อปริมาณเฉลี่ยแม้ว่าความแตกต่างเฉลี่ยระหว่างกลุ่มไม่ได้มีนัยสำคัญ
สำหรับเซสชันการแพ้การเปรียบเทียบระหว่างกลุ่มที่วางแผนไว้แสดงให้เห็นว่าก่อนที่จะได้รับรางวัลซูโครสปรับอากาศ 50% ทำให้การตอบสนองของขมิ้นอ้อยเพิ่มขึ้นอย่างมีนัยสำคัญต่อปริมาณแอมเฟตามีน 1.0 มก. / กก. เมื่อเทียบกับตารางอื่น ๆ อีกสามตาราง ผลกระทบนี้เห็นได้ชัดจากการให้ยาครั้งแรกและไม่เปลี่ยนแปลงอย่างเห็นได้ชัดเมื่อรับประทานซ้ำ การวิเคราะห์แนวโน้มบ่งชี้ถึงการตอบสนองแบบสองเฟส (สำหรับตัวอย่างเต็มรูปแบบ) ต่อปริมาณ AMPH ซ้ำโดยเพิ่มขึ้นเป็นขนาดที่สามและลดลงหลังจากนั้น ผลของการตรวจ ANCOVA ที่ติดตามด้วยการกระตุ้นทางจมูก (ไม่มี CS) เนื่องจากความแปรปรวนร่วมยืนยันว่าความแตกต่างในการตอบสนองของหัวรถจักรทั้งสี่กลุ่มที่มีต่อ AMPH 1 มก. / กก. ไม่ได้เป็นสื่อกลางโดยวิธีที่ไม่ได้รับการดูแลในระหว่างการฝึกอบรมซูโครส
ผลของกลุ่มในระหว่างช่วงการกระตุ้นให้แพ้นั้นสอดคล้องกับสมมติฐานของเรา ผลเซสชั่น bi-phasic ไม่สอดคล้องกับการเพิ่มอย่างต่อเนื่องในการตอบสนองของหัวรถจักรที่มีปริมาณ AMPH ซ้ำ นี่อาจเกี่ยวข้องกับช่วงเวลาการเติม เพื่อแก้ไขปัญหานี้ควรมีการใช้ขั้นตอน (ปริมาณรายวันแบบอื่น) เพื่อกระตุ้นการเพิ่มระดับอย่างสม่ำเสมอในการตอบสนองต่อการเคลื่อนที่ของ 1.0 mg / kg ขนาดของ AMPH (เช่นการกระตุ้นให้เกิดพฤติกรรม) ผลกระทบของระบบการปกครองที่ไวของ AMPH ต่อการตอบสนองต่อความท้าทาย 0.5 mg / kg ครั้งที่สองจะช่วยสนับสนุนลักษณะทั่วไปของผลกระทบนี้ต่อไป การรวมความท้าทายของน้ำเกลือก่อนที่ AMPH จะกำหนดบทบาทของความคาดหวังหรือผลกระทบที่เกี่ยวข้องกับการฉีด (เช่นความเครียด) ที่มีต่อการตอบสนองของหัวรถจักรต่อ AMPH การรวมกลุ่มซูโครสที่มี 75% ปรับอากาศจะช่วยให้ชัดเจนในบทบาทของรางวัลความไม่แน่นอนเทียบกับความไม่บ่อยของรางวัลในรูปแบบของการตอบสนองสำหรับกลุ่ม 50 และกลุ่ม 25 นอกจากนี้เพื่ออนุญาตให้มีการประเมิน (โดย ANCOVA) ของการมีส่วนร่วมของการตอบสนองต่อการเคลื่อนที่ของยาฟรีภายใต้ AMPH (ใช้จมูกโผล่ด้วย CS ปัจจุบันเป็น covariate), จมูก pokes ถูกเข้ารหัสสำหรับกลุ่ม 0 ในช่วงเวลาที่ CS มีอยู่ในอีกสี่กลุ่ม (กล่าวคือจมูกที่โผล่ออกมาจากทั้งห้ากลุ่ม - รวมถึงกลุ่ม 0 ที่ไม่ได้รับ CS - อาจรวมอยู่ในการวิเคราะห์ความแปรปรวนร่วมกับ CS ปัจจุบันเป็น covariate) การปรับแต่งเหล่านี้รวมอยู่ในการทดลอง 2
2 ทดลอง
วัสดุและวิธีการ
วิธีการของการทดลอง 2 นั้นคล้ายคลึงกับการทดลอง 1 แต่ได้รับการแก้ไขให้ใกล้เคียงกับระบบการปกครองที่พบว่าทำให้เกิดอาการแพ้ AMPH ได้อย่างน่าเชื่อถือ (Fletcher และคณะ 2005) การเปลี่ยนแปลงมีดังนี้: (a) กลุ่ม 75% CS-sucrose (n = 8) รวมอยู่; (b) ระหว่างการฝึกซูโครสหนู (ยกเว้นกลุ่ม 0) ได้รับการนำเสนอ 20 CS (แสง) (ตรงข้ามกับ 15 ในการทดลอง 1); (c) งานนำเสนอ CS แต่ละครั้งถูกคั่นด้วยช่วงเวลาการทดลองใช้โดยเฉลี่ยที่ 90 s ช่วง: 30 – 180 s (เทียบกับ 120 s ในการทดลอง 1) ซึ่งชดเชยการเพิ่มขึ้นของการทดลองในการฝึกอบรมเพื่อเปรียบเทียบระยะเวลาของการฝึกซ้อมแต่ละครั้งกับการทดลอง 1; (d) ระยะเวลาของแต่ละช่วงเวลาการทำให้คุ้นเคยสามครั้งลดลงจาก 120 เป็น 90 นาทีเพื่อให้สอดคล้องกับระยะเวลาของเซสชันการทดสอบ (e) เพิ่มความท้าทายน้ำเกลือ (ip, 1 ml / kg) (90 ขั้นต่ำ) (วันฝึกอบรมหลังซูโครส 8) เพื่อประเมินผลของหัวรถจักรของการฉีด ต่อ se (เช่นความคาดหวังความเครียด); (f) ช่วงการแพ้ 1 mg / kg ถูกจัดขึ้นในวันธรรมดาอื่น (วันหลังการฝึกอบรม 12 – 21) มากกว่าช่วงเวลารายสัปดาห์เช่นเดียวกับในการทดลอง 1; (g) พร้อมกับการแพ้ก่อนกำหนด 0.5 mg / kg AMPH Challenge (วันหลังการฝึกอบรม 9) การเพิ่มความไวต่อการแพ้ครั้งที่สอง 0.5 mg / kg AMPH ท้าท้าย (วันฝึกอบรมหลังซูโครส 28) เพื่อทดสอบทั่วไป ผลกระทบที่ทำให้ไวต่อยา (h) จมูกโผล่ในขณะที่มี CS อยู่ทุกกลุ่ม (รวมถึงกลุ่ม 0) (i) การเจาะจมูกในขณะที่ CS ขาดหายไปนั้นถูกบันทึกไว้โดยเฉพาะจากช่วง 5-s ทันทีก่อนที่การโจมตีของ CS จะเข้าสู่ดัชนีก่อนวัยอันควร
ผลสอบ
จมูกทิ่มในระหว่างการปรับสภาพซูโครส
กลุ่ม 5 ×เซสชัน 15 × 2 เฟส (CS ปัจจุบัน, ขาด CS) ANOVA ของ pokes จมูกให้ผลกระทบอย่างมีนัยสำคัญที่สำคัญของกลุ่ม, F(4, 19) = 2.89, p = 0.050 เซสชัน F(14, 266) = 2.28, p = 0.006 และเฟส F(1, 19) = 14.72, p = 0.001 เช่นเดียวกับการโต้ตอบสามทางที่สำคัญ F(56, 266) = 1.38, p = 0.050 แผง (A, B) ของรูป 3 พล็อตคะแนนการโผล่จมูกเฉลี่ยของกลุ่มสำหรับ CS ปัจจุบันและขั้นตอนที่ขาด CS ตามลำดับ การเปรียบเทียบทั้งสองแผงแสดงให้เห็นว่าเอฟเฟกต์หลักของ Phase สะท้อนให้เห็นถึงการตอบสนองของจมูกโดยรวมมากขึ้นเมื่อ CS มีอยู่กับขาด ดังนั้นการตอบสนองแบบ cued จึงเกิดขึ้นบ่อยกว่าการตอบสนองก่อนกำหนด ผลกระทบหลักของกลุ่มและเซสชันไม่สามารถตีความได้อย่างง่ายดายเนื่องจากการโต้ตอบคำสั่งซื้อที่สูงขึ้น ผลลัพธ์หลังนี้สะท้อนให้เห็นถึงการรวมกันของคะแนนของทั้ง XNUMX กลุ่มในระดับต่ำที่ค่อนข้างคงที่ตลอดเซสชันเมื่อ CS ขาด (รูปที่ 3B) พร้อมกับความแตกต่างของคะแนนในระดับสูง (กลุ่ม 75, กลุ่ม 100), ระดับกลาง (กลุ่ม 50), และระดับต่ำ (กลุ่ม 0, กลุ่ม 25) ระดับจมูกโผล่ขึ้นมาตอบสนองต่อเซสชันเมื่อ CS ปรากฏอยู่ 3A) ของแนวโน้มพหุนามลำดับที่ต่ำกว่า (เชิงเส้น, กำลังสอง, ลูกบาศก์) เฉพาะปฏิสัมพันธ์สามทางสำหรับแนวโน้มเชิงเส้นที่มีนัยสำคัญเข้าหา F(4, 19) = 2.32, p = 0.094 ซึ่งสะท้อนถึงการเพิ่มขึ้นของเสียงโมโนในจมูกในการประชุมกลุ่ม 75 และมีความเสถียรที่ค่อนข้างรวดเร็วที่ระดับสูงกลางและระดับต่ำของการตอบสนองในกลุ่มอื่นเมื่อมี CS อยู่
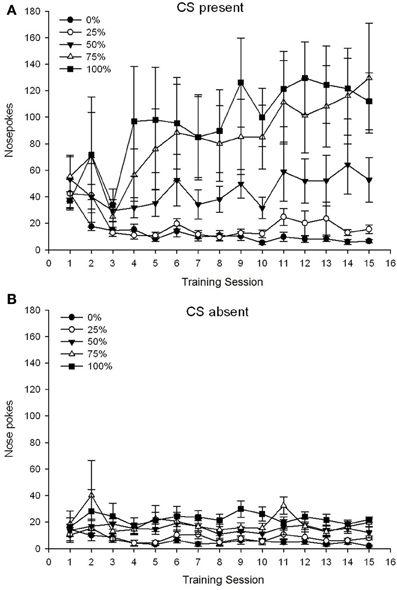
รูปที่ 3 Mean (SE) การตอบสนองวิธีการ (จมูกโผล่) ในการฝึก 15 ซูโครสในกลุ่มของหนู Sprague Dawley (n = 8 / กลุ่ม) สัมผัสกับรางวัลซูโครส (โซลูชัน 10%) ส่งมอบภายใต้ 0, 25, 50, 75, 100 หรือตารางเวลาตัวแปร XNUMX%. มาตรการกระตุ้นคือแสง (120 s) กลุ่ม 0 ได้รับรางวัลจำนวนเท่ากันกับกลุ่ม 100 ในกรณีที่ไม่มีสิ่งเร้า (A) คะแนนเมื่อ CS มีอยู่ (การทดลอง 5 s × 20) (B) คะแนนเมื่อ CS ขาดงาน (ค่าเฉลี่ยสำหรับ 5 × 20 s ขณะที่แสงดับ)
ทำให้คุ้นเคยกับตู้รถไฟ
กลุ่ม 5 × 3 เซสชัน ANOVA ของการตอบสนองของหัวรถจักรที่ปลอดยาเสพติดให้ผลสำคัญหลักของเซสชัน F(2, 70) = 60.01, p <0.0001 และไม่มีผลกระทบที่สำคัญอื่น ๆ F(4, 35) <0.70, p > 0.60. การเปรียบเทียบตามแผนของกลุ่ม 50 กับกลุ่ม 0 และกับกลุ่ม 100 ในการสร้างความเคยชินครั้งแรกและครั้งสุดท้ายไม่มีผลกระทบที่มีนัยสำคัญ t's <0.84, p > 0.40. ดังนั้นการตอบสนองของขมิ้นอ้อยโดยเฉลี่ยในกลุ่มสำคัญจึงไม่แตกต่างกันก่อนการทดสอบ จำนวนลำแสงเฉลี่ยต่อ 90 นาทีคือ 2162 (118) ในเซสชั่นที่ 1, 1470 (116) ในเซสชั่นที่ 2 และ 1250 (98) ในเซสชัน 3
ช่วงทดสอบ
น้ำเกลือ. 5 กลุ่ม× 2 เซสชัน ANOVA เปรียบเทียบการตอบสนองของหัวรถจักรในช่วงการทำให้เกิดความเคยชินและการท้าทายของน้ำเกลือ ANOVA ให้ผลหลักของเซสชัน F(1, 35) = 62.46, p <0.0001 และไม่มีผลกระทบที่สำคัญอื่น ๆ F(4, 35) <0.65, p > 0.64 รูป 4 พล็อตกลุ่มหมายถึงและแสดงให้เห็นว่าผลเซสชันสะท้อนให้เห็นถึงการลดลงโดยรวมในการตอบสนองของหัวรถจักรจากเซสชั่นทำให้เกิดความเคยชินปราศจากยาขั้นสุดท้ายไปยังเซสชันน้ำเกลือซึ่งไม่แตกต่างกันตามกลุ่ม ดังนั้นการลดลงของการตอบสนองของหัวรถจักรที่เห็นในช่วงที่สามทำให้เกิดความเคยชินอย่างต่อเนื่องในการสัมผัสที่ปราศจากยาเสพติดที่สี่ไปยังกล่องทดสอบ
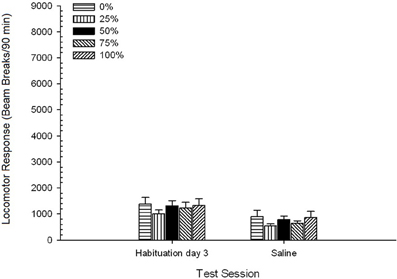
รูปที่ 4 การตอบสนองของหัวรถจักรเฉลี่ย (SE) (จำนวนการหยุดลำแสงในอาร์เรย์อิเล็กทรอนิกส์ต่อ 90 ขั้นต่ำ) ในช่วงสุดท้ายของการทำให้เกิดความคุ้นเคยกับยาเสพติด 3 และในช่วงต่อมาหลังจากการฉีดน้ำเกลือ (ip, 1 ml / kg) ในกลุ่ม Sprague Dawley หนู (n = 8 / กลุ่ม) เปิดเผยก่อนหน้านี้ในเซสชันการปรับสภาพ 15 รายวันด้วยการให้รางวัลซูโครส (โซลูชัน 10%) ส่งมอบภายใต้ 0, 25, 50, 75, 100 หรือ XNUMX% ตารางเวลา. มาตรการกระตุ้นคือแสง (120 s) กลุ่ม 0 ได้รับรางวัลจำนวนเท่ากันกับกลุ่ม 100 ในกรณีที่ไม่มีสิ่งเร้า
ผลของ 0.5 mg / kg AMPH.
การเคลื่อนที่ก่อนฉีด. 5 Group × 2 Session ANOVA ของการเคลื่อนไหวก่อนฉีด (30-min) ในการทดสอบก่อนและหลังการกระตุ้น 0.5 mg / kg AMPH วันทดสอบให้ผลสำคัญอย่างมีนัยสำคัญของเซสชัน, F(1, 35) = 13.39, p = 0.001 และไม่มีผลกระทบที่สำคัญอื่น ๆ F(4, 35) <1.79, p > 0.15 การเปรียบเทียบตามแผนไม่พบความแตกต่างอย่างมีนัยสำคัญระหว่างกลุ่ม 50 และกลุ่ม 0 หรือกลุ่ม 100 ในเซสชันแรก t(70) <1.00, p > 0.30. อย่างไรก็ตามในกลุ่มเซสชันที่สอง (หลังการแพ้) 50 (1203; 121) แสดงการแตกลำแสงก่อนฉีด (M; SE) มากกว่ากลุ่ม 100 (756; 103) อย่างมีนัยสำคัญ t(70) = 5.11, p <0.001 แต่ไม่แตกต่างจากกลุ่ม 0 (1126; 211) t(7) <0.88, p > 0.40. ดังนั้นความแตกต่างพื้นฐานของการเคลื่อนที่ไม่ได้อธิบายถึงความแตกต่างของกลุ่มในการตอบสนองของหัวรถจักรต่อ AMPH 0.5 มก. / กก. แรก แต่อาจมีส่วนทำให้เกิดความแตกต่างระหว่างกลุ่ม 50 และกลุ่ม 100 ในการตอบสนองของขมิ้นอ้อยต่อ AMPH 0.5 มก. / กก. ที่สอง . ค่าเฉลี่ยของลำแสงแตกสำหรับระยะก่อนฉีดในการทดสอบ AMPH 0.5 มก. / กก. ครั้งแรกและครั้งที่สองเท่ากับ 757 (41) และ 974 (59)
การเคลื่อนที่หลังฉีด. 5 Group × 2 Session ANOVA ของ locomotor ตอบสนองต่อ 0.5 mg / kg AMPH ก่อนและหลัง 5-dose sensitizing regimen ส่งผลหลักของเซสชัน F(1, 35) = 76.05, p <0.0001 และไม่มีผลกระทบที่สำคัญอื่น ๆ F(4, 35) <1.10, p > 0.37 รูป 5 แสดงคะแนนเฉลี่ยสำหรับแต่ละกลุ่มและเซสชัน
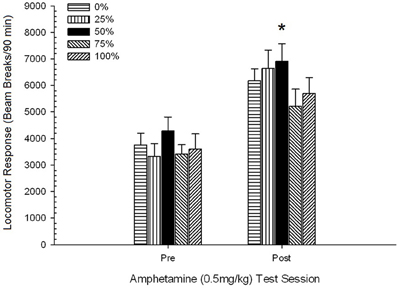
รูปที่ 5 การตอบสนองของหัวรถจักรเฉลี่ย (SE) (จำนวนการหยุดลำแสงในอาเรย์อิเล็กทรอนิกส์ต่อ 90 ขั้นต่ำ) เป็น 0.5 mg / kg d-amphetamine ในแต่ละรอบก่อนและหลัง 5-session sensitizing ระบบของ d-amphetamine (1.0 mg / kg; ต่อเซสชัน) ในกลุ่ม Sprague Dawley rats (n = 8 / กลุ่ม) เปิดเผยก่อนหน้านี้ในเซสชันการปรับสภาพ 15 รายวันด้วยการให้รางวัลซูโครส (โซลูชัน 10%) ส่งมอบภายใต้ 0, 25, 50, 75, 100 หรือ XNUMX% ตารางเวลา. มาตรการกระตุ้นคือแสง (120 s) กลุ่ม 0 ได้รับรางวัลจำนวนเท่ากันกับกลุ่ม 100 ในกรณีที่ไม่มีสิ่งเร้า * * * *p <0.05 สำหรับค่าเฉลี่ยความแตกต่างระหว่างกลุ่ม 50 และกลุ่ม 0 และกลุ่ม 100 ตามการเปรียบเทียบที่วางแผนไว้
รูปแสดงให้เห็นว่าผลเซสชันเกี่ยวข้องกับการเพิ่มขึ้นอย่างมีนัยสำคัญในการแบ่งลำแสงเฉลี่ย (SE) ต่อ 90 นาทีจากปริมาณ 0.5 mg / kg 1, 3674 (216) เป็น 0.5 mg / กิโลกรัมปริมาณ 2, 6123 (275) การขาดปฏิสัมพันธ์หรือผลของกลุ่มชี้ให้เห็นว่าการทำให้แพ้ต่อ AMPH นั้นไม่ได้มีความแตกต่างกันในแต่ละกลุ่ม แม้จะไม่มีผลกระทบที่เกี่ยวข้องกับกลุ่มอย่างมีนัยสำคัญใน ANOVA แต่การตรวจสอบภาพแสดงให้เห็นว่ากลุ่ม 50 แสดงการตอบสนองที่ยิ่งใหญ่ที่สุดต่อทั้งปริมาณ 0.5 mg / kg ที่หนึ่งและสอง การเปรียบเทียบการตอบสนองตามแผนกับปริมาณ 0.5 แรก / กิโลกรัมแสดงให้เห็นว่าไม่มีความแตกต่างอย่างมีนัยสำคัญระหว่างกลุ่ม 50 และกลุ่ม 0 หรือกลุ่ม 100 t's(35) <0.48, p > 0.50. อย่างไรก็ตามในการตอบสนองต่อครั้งที่สอง (หลังการแพ้) 0.5 มก. / กก. กลุ่ม 50 แสดงการเคลื่อนไหวมากกว่ากลุ่ม 0 อย่างมีนัยสำคัญ t(35) = 2.00, p <0.05 เช่นเดียวกับกลุ่ม 100 t(35) = 3.29, p <0.01.
ในแง่ของความแตกต่างของกลุ่มอย่างมีนัยสำคัญในการเคลื่อนที่ก่อนฉีดในช่วงที่สองของ 0.5 mg / kg AMPH ที่รายงานข้างต้น, การติดตาม 5 Group × 2 Session ANCOVA ของการเคลื่อนไหวของการตอบสนองของ locomotor ต่อ 0.5 mg / kg AMPH การเคลื่อนที่ของหัวฉีดในช่วงที่สอง การวิเคราะห์นี้ให้ผลที่สำคัญของ covariate F(1, 34) = 8.65, p = 0.006 ซึ่งเป็นเอฟเฟ็กต์หลักของเซสชัน F(1, 34) = 10.83, p = 0.002 และไม่มีผลกระทบที่สำคัญอื่น ๆ F(4, 34) <0.85, p > 0.50. ที่สำคัญการเปรียบเทียบตามแผนตามข้อผิดพลาด MS และข้อผิดพลาด df จาก ANCOVA ยืนยันว่าการตอบสนองของหัวรถจักรเฉลี่ยต่อ AMPH 0.5 มก. / กก. ที่สองยังคงสูงกว่ากลุ่ม 50 มากกว่ากลุ่ม 100 อย่างมีนัยสำคัญ t(34) = 3.09, p <0.01 และกลุ่ม 0 t(34) = 1.88, p <0.05 (ด้านเดียว) เมื่อมีการควบคุมรูปแบบก่อนการฉีดจากเซสชัน 2 ดังนั้นกลุ่มที่ 50 จึงแสดงการตอบสนองของหัวจักรโพสต์ไวไลเซชั่นที่มากกว่า 0.5 มก. / กก. AMPH มากกว่ากลุ่ม 100 หรือกลุ่ม 0 อย่างมีนัยสำคัญและความแตกต่างของกลุ่มเหล่านี้ไม่ได้ถูกสื่อกลางโดยการเคลื่อนที่ก่อนการฉีดในวันทดสอบ
ผลของ 1.0 mg / kg AMPH.
การเคลื่อนที่ก่อนฉีด. 5 Group × 5 Session ANOVA ของ 30-min คะแนนก่อนการฉีดสำหรับการประชุม 1 mg / kg AMPH ทำให้เกิดผลกระทบหลักของเซสชัน F(4, 140) = 16.70, p <0.0001 และไม่มีผลกระทบที่สำคัญอื่น ๆ F(4, 35) <0.94, p > 0.45 การเปรียบเทียบตามแผนไม่พบความแตกต่างอย่างมีนัยสำคัญในการเคลื่อนที่ก่อนการฉีดระหว่างกลุ่ม 50 และกลุ่ม 0 หรือกลุ่ม 100 ในเซสชั่นแรก t(175) <1.66, p > 0.10. อย่างไรก็ตามในเซสชันสุดท้ายกลุ่ม 50 (1167; 140) แสดงลำแสงแตก (M; SE) มากกว่ากลุ่ม 100 (1000; 99) อย่างมีนัยสำคัญ t(175) = 2.35, p <0.05 แต่ไม่แตกต่างจากกลุ่ม 0 (1085, 120) t(175) <1.16, p > 0.20. ดังนั้นความแตกต่างของการเคลื่อนที่ก่อนฉีดจึงทำให้เกิดความแตกต่างระหว่างกลุ่มที่ 50 และ 100 ในการตอบสนองของหัวรถจักรต่อปริมาณ AMPH ขั้นสุดท้าย 1 มก. / กก. ค่าเฉลี่ยของลำแสงโดยรวมสำหรับตัวอย่างในช่วงก่อนการฉีดสำหรับเซสชันที่ 1 ถึง 5 คือ 810 (46), 784 (52), 760 (53), 726 (46), 1009 (51)
การเคลื่อนที่หลังฉีด. 5 Group × 5 Session ANOVA ของการตอบสนองต่อ 1 mg / kg AMPH ให้ผลหลักอย่างมีนัยสำคัญของ Session, F(4, 140) = 6.72, p <0.001 ปฏิสัมพันธ์กลุ่มชายขอบ×เซสชัน F(16, 140) = 1.57, p = 0.085 และไม่มีผลกระทบหลักของกลุ่ม F(4, 35) <0.44, p > 0.77 การวิเคราะห์แนวโน้มพหุนามพบว่ามีแนวโน้มเชิงเส้นที่สำคัญ F(1, 35) = 9.19, p = 0.005 และแนวโน้มลูกบาศก์ F(1, 35) = 21.63, p <0.001 ในช่วงที่ 1 ถึง 5 6 แสดงคะแนนเฉลี่ยของหัวรถจักรสำหรับแต่ละกลุ่มและเซสชั่น
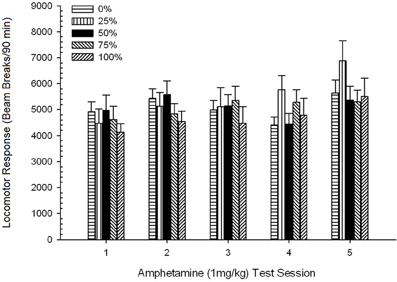
รูปที่ 6 การตอบสนองของหัวรถจักรเฉลี่ย (SE) (จำนวนการหยุดลำแสงในอาร์เรย์อิเล็กทรอนิกส์ต่อ 90 ขั้นต่ำ) ถึง 1 mg / kg d-amphetamine (ip) ในการประชุม 5 รายสัปดาห์ในกลุ่ม Sprague Dawley rats (n = 8 / กลุ่ม) เปิดเผยก่อนหน้านี้ในเซสชันการปรับสภาพ 15 รายวันด้วยการให้รางวัลซูโครส (โซลูชัน 10%) ส่งมอบภายใต้ 0, 25, 50, 75, 100 หรือ XNUMX% ตารางเวลา. มาตรการกระตุ้นคือแสง (120 s) กลุ่ม 0 ได้รับรางวัลจำนวนเท่ากันกับกลุ่ม 100 ในกรณีที่ไม่มีสิ่งเร้า
รูปแสดงให้เห็นว่าผลกระทบของเซสชั่นสะท้อนให้เห็นถึงการเพิ่มขึ้นอย่างมีนัยสำคัญในการแบ่งลำแสงเฉลี่ย (SE) สำหรับตัวอย่างเต็มรูปแบบจากเซสชั่น 1, 4624 (213) เป็นเซสชั่น 5, 5736 (272) ยืนยันการเกิดอาการแพ้ แนวโน้มลูกบาศก์แสดงถึงค่าสูงสุดที่สัมพันธ์กันในช่วง 1, 3 และ 5 โดยลดลงในช่วง 2 และ 4 โดยเฉพาะอย่างยิ่งสำหรับกลุ่ม 0 และ 50 รูปยังเผยให้เห็นว่าแม้จะขาดการปฏิสัมพันธ์ที่สำคัญกลุ่ม 25 แสดงการตอบสนองของหัวรถจักรที่เพิ่มขึ้นอย่างต่อเนื่องและแตกต่างอย่างมากจากกลุ่มอื่น ๆ ในเซสชัน 4 และ 5 (9 และ 22% ตามลำดับมากกว่ากลุ่มสูงสุดถัดไป) การเปรียบเทียบตามแผนพบว่ากลุ่ม 50 ไม่ได้แตกต่างอย่างมีนัยสำคัญจากกลุ่ม 0 หรือ 100 t(175) <0.89, p > 0.40 ในการทดสอบ AMPH 1 มก. / กก. ครั้งแรกหรือครั้งสุดท้าย
การควบคุมความแปรปรวนของการกระตุ้นจมูกตอบสนองระหว่างการฝึกซูโครส
5 สองกลุ่ม× 2 เซสชัน ANCOVAs ของหัวรถจักรตอบสนองต่อ 0.5 มก. / กก. AMPH ก่อนและหลังการควบคุมอาการแพ้รวมถึงจมูกรวมทั้งหมดในระหว่างการฝึกซูโครสกับ CS ปัจจุบันและ CS ขาดเป็นโควาเรียที่แยกจากกันพบว่าไม่มีผลกระทบอย่างมีนัยสำคัญ F(1, 18) <1.03, p > 0.31. ดังนั้นวิธีการตอบสนองในระหว่างการฝึกอบรมไม่ได้เป็นสื่อกลางในความแตกต่างของกลุ่มในการตอบสนองต่อ AMPH 0.5 มก. / กก.
5 Group × 5 สองเซสชัน ANCOVAs ของ locomotor ตอบสนองต่อ 1 mg / kg ในระหว่างช่วงการกระตุ้นอาการแพ้ที่มีจมูกรวมทั้งหมด (CS ปัจจุบัน, CS ขาดอยู่) เนื่องจาก covariates แยกไม่มีผลสำคัญของ covariate ในขณะที่ CS อยู่ F(4, 104) <1.04, p > 0.38 และผลกระทบหลักเล็กน้อยของความแปรปรวนร่วมในขณะที่ CS ไม่อยู่ F(1, 18) = 3.32, p = 0.085
การสนทนา
ผลของการศึกษาครั้งนี้ไม่ได้สนับสนุนสมมติฐานอย่างสม่ำเสมอว่ากลุ่ม 50 จะแสดงการตอบสนองของหัวรถจักรที่สูงขึ้นเมื่อเทียบกับกลุ่มอื่น ๆ ข้อมูล AMPH 1 mg / kg ยืนยันการเกิดอาการแพ้โดยใช้ระบบการให้ยาแบบวันต่อวัน รูปแบบในกลุ่มต่างๆบ่งชี้ถึงแนวโน้มของการแพ้ที่มากขึ้นในช่วงหลังในกลุ่ม 25 โดยไม่มีหลักฐานดังกล่าวสำหรับกลุ่ม 50 ในทางตรงกันข้ามผลลัพธ์ปริมาณรังสี 0.5 mg / kg บ่งชี้ถึงแนวโน้มของการแพ้ในกลุ่ม 50 ในขณะเดียวกันก็ยืนยันการเพิ่มขึ้นอย่างมีนัยสำคัญโดยรวมในการตอบสนองของหัวรถจักรข้ามกลุ่มเป็นวินาทีและปริมาณ 0.5 mg / kg AMPH แรก ผลของการฉีดน้ำเกลือยืนยันว่าความคาดหวังหรือความเครียดที่เกี่ยวข้องกับการฉีดไม่ได้ส่งผลต่อ AMPH
ข้อมูลจากจมูกโผล่ขึ้นมาอีกครั้งเผยให้เห็นถึงการเพิ่มขึ้นโดยรวมของวิธีการตอบสนองในระหว่างการฝึกซ้อมเมื่อมี CS อยู่โดยไม่เพิ่มขึ้นเมื่อ CS ขาดไป ดังนั้นสัตว์ดูเหมือนจะได้รับความสัมพันธ์ระหว่าง CS และโอกาสในการได้รับรางวัลซูโครส ความแตกต่างของกลุ่มในความถี่ของจมูกโผล่เมื่อ CS สอดคล้องกับความถี่ของการส่งมอบรางวัลภายใต้ตารางเวลาที่เกี่ยวข้องโดยมีกลุ่ม 75 และ 100 แสดงจมูกโผล่มากที่สุดกลุ่ม 50 แสดงตัวเลขโผล่กลางจมูกและกลุ่ม 0 และ 25 แสดงให้เห็นจมูกที่เล็กที่สุด ผลลัพธ์เหล่านี้ชี้ให้เห็นว่า CS มาเพื่อควบคุมวิธีการตอบสนองในลักษณะที่สอดคล้องกับความน่าจะเป็นโดยรวมของรางวัล แม้ว่าจะเป็นการเก็งกำไรคำอธิบายที่เป็นไปได้สำหรับอัตราการกระตุ้นจมูกที่ต่ำกว่าด้วย CS ที่มีอยู่ในกลุ่ม 50 ในการทดลอง 2 เทียบกับการทดลอง 1 อาจเป็นการลดระยะเวลาการทดลองใช้ระหว่างการทดลอง แนวโน้มและสิ่งนี้เกี่ยวข้องกับการหมุนเวียนที่เพิ่มขึ้นของ DA ใน cingulate ล่วงหน้า, prelimbic และ infralimbic cortices (Dalley และคณะ 2002) ดังนั้นการลด 30% ในช่วงเวลาระหว่างการทดลองในการทดลอง 2 (และ 3) อาจเปลี่ยนระดับเยื่อหุ้มสมอง DA และเลื่อนระดับการคัดเลือกมากขึ้น (เช่นได้รับคำแนะนำจากความถี่สัมพัทธ์ของรางวัล) เทียบกับวิธีการหุนหัน การตอบสนองในกลุ่ม 50 ระหว่างการทดสอบการฝึกซ้อมในการทดลอง 2 เมื่อเปรียบเทียบกับการทดลอง 1
การขาดผลกระทบที่เกี่ยวข้องกับโควาเรียอย่างมีนัยสำคัญสำหรับการเจาะจมูกในสภาพปัจจุบันของ CS ใน ANCOVAs บ่งชี้ว่าวิธีการตอบสนองในระหว่างการฝึกซูโครสไม่ได้เป็นสื่อกลางของตาราง CS-sucrose ที่แตกต่างกันต่อการตอบสนองต่อ AMPH ผลกระทบสำคัญของ covariate สำหรับอาการขาด CS ใน ANCOVA ของ locomotor ตอบสนองต่อ 1 mg / kg AMPH แสดงให้เห็นว่าแนวโน้มต่อการตอบสนองที่ปราศจากยาก่อนกำหนดอธิบายถึงความแปรปรวนของผลกระทบของ locomotor ระหว่าง AMPH
หลักฐานบ่งชี้ว่าผลกระทบของประวัติการปรับสภาพอาจจะมองเห็นได้ด้วย 0.5 AMPH มากกว่า 1 mg / kg AMPH และโปรโตคอลที่สร้างความรู้สึกไวหากไม่มีการจัดการอื่น ๆ อาจปิดบังหรือทำให้ผลกระทบซ้ำซ้อนของสมมุติฐาน การกระตุ้นให้เกิดการกระตุ้นพฤติกรรม (เช่นรางวัลตัวแปรเรื้อรัง)
การทำให้ไวต่อพฤติกรรมต่อ AMPH เป็นผลที่แข็งแกร่งในห้องปฏิบัติการ อย่างไรก็ตามนอกห้องปฏิบัติการมีเพียงส่วนน้อยของบุคคลที่เล่นการพนันเพิ่มระดับเรื้อรังถึงระดับพยาธิวิทยา แม้ว่าความเสี่ยงในการเกิดอาการแพ้นั้นสัมพันธ์กับความเสี่ยงในการติดยาเสพติด (หรือการแสวงหายาเสพติด) โดยเฉพาะอย่างยิ่งสำหรับยาจิตเวช (Vezina, 2004; Flagel et al., 2008) ปัจจัยหลายอย่างนอกเหนือจากความเสี่ยงต่อการเกิดอาการแพ้อาจจูงใจให้เกิดการเสพติด (เช่น Verdejo-Garcia และคณะ, 2008; Conversano et al., 2012; Volkow et al., 2012) อย่างไรก็ตามปัจจัยลักษณะที่ทำให้เกิดความเสี่ยงต่อการแพ้อาจมีผลกระทบกับประวัติการปรับสภาพเพื่อเน้นผลของรางวัลที่ไม่สามารถคาดการณ์ได้ (เช่นกำหนดการ 50% CS-US) ต่อปฏิกิริยาของระบบ DA เพื่อตรวจสอบความเป็นไปได้นี้การทดลอง 3 ใช้ขั้นตอนเดียวกับการทดลอง 2 แต่ใช้สายพันธุ์ Lewis แทนหนูสายพันธุ์ Sprague Dawley
Sprague Dawley rats แสดงระดับกลางของตัวส่งสัญญาณ DA โดยมีระดับต่ำกว่าหนู Wistar สายพันธุ์ (Zamudio และคณะ, 2005) แต่ระดับที่สูงกว่าหนูวิสตาร์ - เกียวโต (ความเครียดแบบ "ซึมเศร้า") ในนิวเคลียส accumbens, amygdala, บริเวณหน้าท้องและพื้นที่เทเชียลนิโกร (substantia nigra)เจียวและคณะ, 2003) โปรไฟล์นี้อาจแสดงให้เห็นว่า Sprague Dawley หนูมีความอ่อนไหวต่อสภาพแวดล้อมหรือเภสัชวิทยาของ DA ในทางตรงกันข้ามหนู Lewis แสดงระดับการขนย้าย DA ต่ำเช่นเดียวกับตัวรับ D2 และ D3 DA ในนิวเคลียส accumbens และ dorsal striatum เมื่อเทียบกับสายพันธุ์อื่น (เช่น F344) (Flores et al., 1998). ความแตกต่างทางสัณฐานวิทยาเหล่านี้อาจนำไปสู่การตอบสนองที่แตกต่างกันของหนู Lewis ต่อการปรับเปลี่ยน DA หนูลูอิสยังแสดงการตอบสนองที่เน้นการใช้ยาทดลองเมื่อเทียบกับสายพันธุ์อื่น ๆ (เช่น F344) ที่สำคัญที่สุดหนูลูอิสแสดงอาการแพ้ต่อเมทแอมเฟตามีนมากขึ้นโดยมีการตอบสนองต่อปริมาณเริ่มต้นต่ำ แต่ตอบสนองต่อปริมาณที่มากขึ้นในภายหลัง (Camp et al., 1994) ลูอิสหนูยังแสดงอาการแพ้หัวรถจักรมากขึ้นในช่วงของโคเคน (Kosten และคณะ, 1994; Haile et al., 2001) จากรูปแบบของเอฟเฟกต์นี้เราคาดการณ์ว่าหนูลูอิสจะช่วยให้เราตรวจสอบว่าความไวต่อการแพ้นั้นขยายผลของตารางการปรับสภาพต่อการตอบสนองต่อ AMPH หรือไม่
3 ทดลอง
วัสดุและวิธีการ
วิธีการนั้นเหมือนกับในการทดลอง 2 นอกเหนือจากการใช้หนูลูอิส (200 – 225 g เมื่อมาถึง, แม่น้ำชาร์ลส์, ควิเบก, แคนาดา)
ผลสอบ
จมูกทิ่มในระหว่างการปรับสภาพซูโครส
กลุ่ม 5 ×เซสชัน 15 × 2 เฟส (CS ปัจจุบัน, ขาด CS) ANOVA ของ pokes จมูกให้ผลกระทบอย่างมีนัยสำคัญที่สำคัญของกลุ่ม, F(4, 34) = 6.12, p = 0.001 เซสชัน F(14, 476) = 3.42, p <0.001 และเฟส F(1, 34) = 20.83, p <0.001 เช่นเดียวกับปฏิสัมพันธ์สามทางที่สำคัญ F(56, 476) = 1.56, p = 0.008 แผง (A, B) ของรูป 7 พล็อตคะแนนการโผล่จมูกเฉลี่ยของกลุ่มสำหรับ CS ปัจจุบันและขั้นตอนที่ขาด CS ตามลำดับ การเปรียบเทียบทั้งสองแผงแสดงให้เห็นว่าเอฟเฟกต์หลักของ Phase สะท้อนให้เห็นถึงการตอบสนองของจมูกโดยรวมมากขึ้นเมื่อ CS มีอยู่กับขาด ดังนั้นการตอบสนองที่ได้รับการดูแลจึงเกิดขึ้นบ่อยกว่าการตอบสนองก่อนวัยผู้ใหญ่อย่างมีนัยสำคัญ ผลกระทบหลักของกลุ่มและเซสชันไม่สามารถตีความได้อย่างง่ายดายเนื่องจากการโต้ตอบคำสั่งซื้อที่สูงขึ้น การโต้ตอบสามทางสะท้อนให้เห็นถึงการรวมกันของคะแนนของทั้งห้ากลุ่มในระดับที่ค่อนข้างคงที่ต่ำในช่วงที่ CS ขาด [Panel (B)] พร้อมกับความแตกต่างของคะแนนเมื่อ CS อยู่ในโปรไฟล์ที่แยกกันสำหรับแต่ละกลุ่มที่ขนานลำดับของความถี่รางวัล: จากสูงสุด (กลุ่ม 100) ถึงต่ำสุด (กลุ่ม 25) [แผง (A)] เฉพาะแนวโน้มเชิงเส้นสำหรับการโต้ตอบเท่านั้น F(4, 34) = 4.03, p = 0.009 ซึ่งสะท้อนถึงการเพิ่มขึ้นอย่างต่อเนื่องของการเจาะจมูกในการประชุมกลุ่ม 100 เมื่อมี CS เมื่อเทียบกับโปรไฟล์ที่ไม่สอดคล้องกันของการเพิ่มขึ้นของการเจาะจมูกในการประชุมในกลุ่มอื่น ๆ ในช่วงนี้
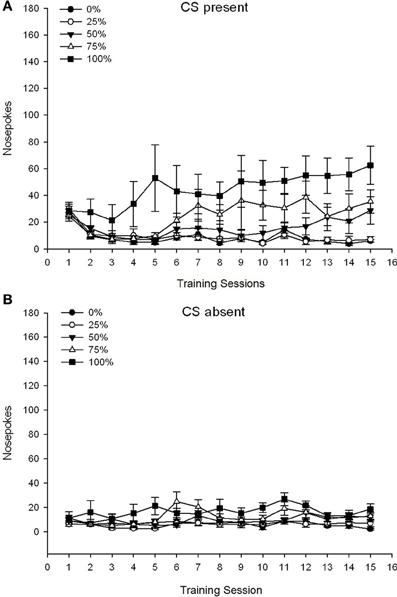
รูปที่ 7 Mean (SE) การตอบสนองวิธีการ (จมูกโผล่) ในการฝึก 15 ซูโครสในกลุ่มของหนู Lewis (n = 8 / กลุ่ม) สัมผัสกับรางวัลซูโครส (โซลูชัน 10%) ส่งมอบภายใต้ 0, 25, 50, 75, 100 หรือตารางเวลาตัวแปร XNUMX%. มาตรการกระตุ้นคือแสง (120 s) กลุ่ม 0 ได้รับรางวัลจำนวนเท่ากันกับกลุ่ม 100 ในกรณีที่ไม่มีสิ่งเร้า (A) คะแนนเมื่อ CS มีอยู่ (การทดลอง 5 s × 20) (B) คะแนนเมื่อ CS ขาดงาน (ค่าเฉลี่ยสำหรับ 5 × 20 s ขณะที่แสงดับ)
ทำให้คุ้นเคยกับตู้รถไฟ
กลุ่ม 5 × 3 เซสชัน ANOVA ให้ผลหลักของเซสชัน F(2, 70) = 23.07, p <0.0001 และไม่มีผลกระทบที่สำคัญอื่น ๆ F(8, 70) <1.47, p > 0.18 รูปแบบเส้นโค้งของคะแนนหัวรถจักรเฉลี่ย (SE) เกิดขึ้นจากเซสชัน 1, 1076 (74) ถึงเซสชัน 2, 644 (48) ถึงเซสชัน 3, 762 (59) การเปรียบเทียบตามแผนของกลุ่ม 50 กับกลุ่ม 0 และกับกลุ่ม 100 ในการสร้างนิสัยครั้งแรกและครั้งสุดท้ายพบว่ามีการแบ่งลำแสงน้อยลงอย่างมีนัยสำคัญในกลุ่ม 50 (M = 911; SE = 109) กับกลุ่ม 0 (M = 1103; SE = 176) ในเซสชันการทำให้คุ้นเคย 1 t(105) = 2.02, p <0.05 แต่ไม่มีความแตกต่างระหว่างกลุ่ม 50 และกลุ่ม 100 (M = 1066; SE = 150) t(105) <1.20, p > 0.20 ในเซสชันนี้ กลุ่ม 50 ไม่มีความแตกต่างอย่างมีนัยสำคัญจากกลุ่ม 0 หรือกลุ่ม 100 ในเซสชันความเคยชินขั้นสุดท้าย t(105) <0.93, p > 0.30. ดังนั้นการตอบสนองของขมิ้นอ้อยโดยเฉลี่ยในกลุ่มสำคัญจึงไม่แตกต่างกันอย่างสม่ำเสมอก่อนการทดสอบ
ช่วงทดสอบ
น้ำเกลือ. 5 Group × 2 Session ANOVA ของการตอบสนองของหัวรถจักรในช่วงสุดท้ายของการทำให้เกิดความเคยชินและการทดสอบน้ำเกลือให้ผลที่สำคัญอย่างมากของเซสชัน F(1, 35) = 50.12, p <0.0001 และไม่มีผลกระทบที่สำคัญอื่น ๆ F(4, 35) <0.57, p > 0.68 รูป 8 แสดงคะแนนเฉลี่ยกลุ่มสำหรับสองเซสชันและระบุว่าผลเซสชันสะท้อนการลดลงอย่างมีนัยสำคัญจากการทำให้คุ้นเคยกับการทดสอบน้ำเกลือ ดังนั้นการรับฉีด ต่อ se (เช่นความคาดหวังความเครียด) ไม่ได้เพิ่มการตอบสนองของหัวรถจักร
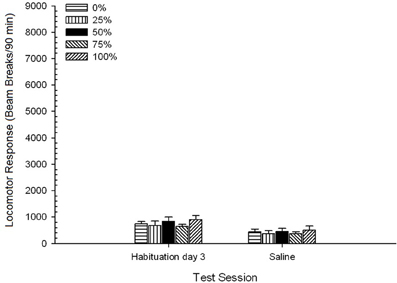
รูปที่ 8 การตอบสนองของหัวรถจักรเฉลี่ย (SE) (จำนวนการหยุดลำแสงในอาร์เรย์อิเล็กทรอนิกส์ต่อ 90 ขั้นต่ำ) ในช่วงสุดท้ายของการทำให้เกิดความคุ้นเคยกับยา 3 และในช่วงต่อมาหลังจากฉีดน้ำเกลือ (ip, 1 ml / kg) ในกลุ่มของหนูลูอิส (n = 8 / กลุ่ม) เปิดเผยก่อนหน้านี้ในเซสชันการปรับสภาพ 15 รายวันด้วยการให้รางวัลซูโครส (โซลูชัน 10%) ส่งมอบภายใต้ 0, 25, 50, 75, 100 หรือ XNUMX% ตารางเวลา. มาตรการกระตุ้นคือแสง (120 s) กลุ่ม 0 ได้รับรางวัลจำนวนเท่ากันกับกลุ่ม 100 ในกรณีที่ไม่มีสิ่งเร้า
ผลของ 0.5 mg / kg AMPH.
การเคลื่อนที่ก่อนฉีด. 5 Group × 2 Session ANOVA ของการเคลื่อนไหวก่อนฉีดให้ผลหลักที่สำคัญของเซสชัน F(1, 35) = 15.04, p <0.001 และไม่มีผลกระทบที่สำคัญอื่น ๆ F(4, 35) <1.19, p > 0.33 การเปรียบเทียบตามแผนไม่พบความแตกต่างอย่างมีนัยสำคัญระหว่างกลุ่ม 50 และกลุ่ม 0 หรือกลุ่ม 100 ในเซสชันการทดสอบใด ๆ t(70) <0.99, p > 0.30. ดังนั้นความแตกต่างพื้นฐานของการเคลื่อนที่ก่อนการฉีดจึงไม่ได้อธิบายถึงความแตกต่างของกลุ่มในการตอบสนองของขมิ้นอ้อยที่ 0.5 มก. / กก. AMPH ลำแสงเฉลี่ย (SE) แตกสำหรับระยะก่อนการฉีดสำหรับครั้งแรกและครั้งที่สอง (หลังการแพ้) ครั้งละ 0.5 มก. / กก. เท่ากับ 325 (25) และ 473 (36)
การเคลื่อนที่หลังฉีด. กลุ่ม 5 × 2 เซสชัน ANOVA ของการตอบสนองของหัวรถจักรต่อปริมาณ 0.5 mg / kg ที่ส่งก่อนและหลังเรื้อรัง 1 mg / kg AMPH ให้ผลหลักของเซสชัน F(1, 34) = 87.44, p <0.0001 และไม่มีผลกระทบที่สำคัญอื่น ๆ F(4, 34) <0.94, p > 0.45 รูป 9 วางแผนคะแนนเฉลี่ยของหัวรถจักรสำหรับแต่ละกลุ่มและเซสชันและแสดงให้เห็นว่าผลเซสชันสะท้อนให้เห็นถึงการตอบสนองโดยรวมที่เพิ่มขึ้นกับปริมาณ 0.5 mg / kg ที่สองซึ่งสอดคล้องกับการแพ้ รูปยังแสดงให้เห็นว่ากลุ่มดำเนินการคล้ายกันมากในเซสชัน 1 แต่กลุ่ม 50 นั้นแสดงกิจกรรมของหัวรถจักรมากกว่ากลุ่มอื่น ๆ ในเซสชัน 2 การเปรียบเทียบตามแผนเพื่อตอบสนองต่อปริมาณ 0.5 แรก / ปริมาณรังสีกิโลกรัมต่อกิโลกรัมพบว่าไม่มีความแตกต่างอย่างมีนัยสำคัญระหว่างกลุ่ม 50 และกลุ่ม 0 หรือกลุ่ม 100 t(35) <1.28, p > 0.20. อย่างไรก็ตามกลุ่มที่ 50 แสดงการตอบสนองของขมิ้นอ้อยต่อยา 0.5 มก. / กก. ที่สองมากกว่ากลุ่ม 0 อย่างมีนัยสำคัญ t(35) = 4.32, p <0.001 หรือกลุ่ม 100 t(35) = 2.24, p <0.05.
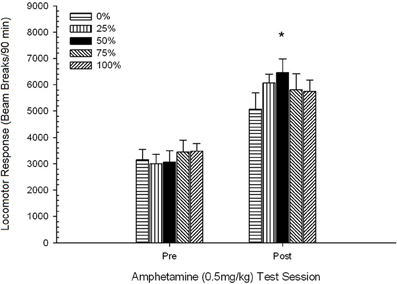
รูปที่ 9 การตอบสนองของหัวรถจักรเฉลี่ย (SE) (จำนวนการหยุดลำแสงในอาเรย์อิเล็กทรอนิกส์ต่อ 90 ขั้นต่ำ) เป็น 0.5 mg / kg d-amphetamine ในแต่ละรอบก่อนและหลัง 5-session sensitizing ระบบของ d-amphetamine (1.0 mg / kg; ต่อเซสชัน) ในกลุ่มของ Lewis rats (n = 8 / กลุ่ม) เปิดเผยก่อนหน้านี้ในเซสชันการปรับสภาพ 15 รายวันด้วยการให้รางวัลซูโครส (โซลูชัน 10%) ส่งมอบภายใต้ 0, 25, 50, 75, 100 หรือ XNUMX% ตารางเวลา. มาตรการกระตุ้นคือแสง (120 s) กลุ่ม 0 ได้รับรางวัลจำนวนเท่ากันกับกลุ่ม 100 ในกรณีที่ไม่มีสิ่งเร้า * * * *p <0.05 สำหรับค่าเฉลี่ยความแตกต่างระหว่างกลุ่ม 50 และกลุ่ม 0 และกลุ่ม 100 ตามการเปรียบเทียบที่วางแผนไว้
ผลของ 1 mg / kg AMPH.
การเคลื่อนที่ก่อนฉีด. 5 Group × 5 Session ANOVA ของ 30-min คะแนนก่อนการฉีดสำหรับช่วงการแพ้ให้ผลหลักของเซสชัน F(4, 140) = 4.10, p = 0.004 และไม่มีผลกระทบที่สำคัญอื่น ๆ F(4, 35) = 1.25, p > 0.31. การเปรียบเทียบตามแผนพบว่าคานแตกในช่วงก่อนการฉีด (M; SE) ต่ำกว่ากลุ่ม 50 (395; 62) อย่างมีนัยสำคัญกว่าในกลุ่ม 100 (508; 62) t(175) = 2.58, p <0.01 แต่ไม่ใช่กลุ่ม 0 t(175) <1.83, p > 0.10 ในช่วง AMPH 1 มก. / กก. 1 ในช่วง AMPH 1 มก. / กก. สุดท้ายการเปรียบเทียบตามแผนยังพบว่าการเคลื่อนที่ก่อนฉีดในกลุ่ม 50 (378; 60) ต่ำกว่ากลุ่ม 100 อย่างมีนัยสำคัญ (650; 75 ), t(175) = 6.17, p <0.001 แต่ไม่อยู่ในกลุ่ม 0 t(175) <1.84, p > 0.10. เนื่องจากทิศทางของความแตกต่างของกลุ่มเหล่านี้ (กลุ่มควบคุม = กลุ่ม 50) ตรงข้ามกับรูปแบบที่ตั้งสมมติฐานความแตกต่างของกลุ่มในการเคลื่อนที่หลังการฉีดที่สอดคล้องกับสมมติฐานจึงไม่สามารถนำมาประกอบกับความแตกต่างพื้นฐานก่อนการฉีดได้ ค่าเฉลี่ยของลำแสงโดยรวมจะแตกระหว่างระยะก่อนฉีดสำหรับเซสชันที่ 1 ถึง 5 ได้แก่ 442 (34), 452 (32), 542 (40), 411 (26), 504 (37)
การเคลื่อนที่หลังฉีด. กลุ่ม 5 × 5 ประชุม ANOVA ของการตอบสนองต่อปริมาณ 1 mg / kg ให้ผลหลักที่สำคัญของเซสชัน F(4, 140) = 6.15, p <0.001 และไม่มีผลกระทบที่สำคัญอื่น ๆ F(4, 35) <0.57, p > 0.68 การวิเคราะห์แนวโน้มพหุนามพบว่ามีแนวโน้มเชิงเส้นที่สำคัญ F(1, 35) = 9.34, p = 0.004 และแนวโน้มลูกบาศก์ F(1, 35) = 5.08, p = 0.031 ซึ่งเป็นผลลัพธ์หลังที่แสดงถึงค่าสูงสุดที่สัมพันธ์กันในเซสชัน 3 และ 5 รูป 10 วางแผนคะแนนเหล่านี้และแสดงให้เห็นว่าแม้จะขาดการมีส่วนร่วมอย่างมีนัยสำคัญใน ANOVA กลุ่ม 50 แสดงการเคลื่อนไหวที่สำคัญกว่ากลุ่มอื่น ๆ อีกสี่กลุ่มเพื่อตอบสนองต่อปริมาณ 1 mg / kg สุดท้าย ดังนั้นการเปรียบเทียบตามแผนเปิดเผยคะแนนเฉลี่ยที่มากขึ้นในเซสชัน 5 ในกลุ่ม 50 กว่าในกลุ่มอื่น ๆ ทั้งหมด t(35) > 3.68, p <0.001.
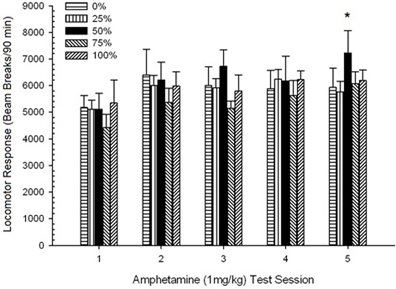
รูปที่ 10 การตอบสนองของหัวรถจักรเฉลี่ย (SE) (จำนวนการหยุดลำแสงในอาเรย์อิเล็กทรอนิกส์ต่อ 90 ขั้นต่ำ) เป็น 1 mg / kg d-amphetamine (ip) ในการประชุม 5 รายสัปดาห์ในกลุ่มของหนูลูอิส (n = 8 / กลุ่ม) เปิดเผยก่อนหน้านี้ในเซสชันการปรับสภาพ 15 รายวันด้วยการให้รางวัลซูโครส (โซลูชัน 10%) ส่งมอบภายใต้ 0, 25, 50, 75, 100 หรือ XNUMX% ตารางเวลา. มาตรการกระตุ้นคือแสง (120 s) กลุ่ม 0 ได้รับรางวัลจำนวนเท่ากันกับกลุ่ม 100 ในกรณีที่ไม่มีสิ่งเร้า * * * *p <0.05 สำหรับค่าเฉลี่ยความแตกต่างระหว่างกลุ่ม 50 และกลุ่ม 0 และกลุ่ม 100 ตามการเปรียบเทียบที่วางแผนไว้
การควบคุมความแปรปรวนของการกระตุ้นจมูกตอบสนองระหว่างการฝึกซูโครส
5 สองกลุ่ม× 2 เซสชัน ANCOVAs ของหัวรถจักรตอบสนองต่อ 0.5 มก. / กก. AMPH ก่อนและหลังการควบคุมอาการแพ้รวมถึงจมูกรวมทั้งหมดในระหว่างการฝึกซูโครสกับ CS ปัจจุบันและ CS ขาดเป็นโควาเรียที่แยกจากกันพบว่าไม่มีผลกระทบอย่างมีนัยสำคัญ F(1, 32) <0.44 p > 0.51 ANCOVA 5 กลุ่ม× 5 เซสชันสองครั้งของการตอบสนองของขมิ้นอ้อยต่อ 1 มก. / กก. AMPH ในระหว่างการกระตุ้นอาการแพ้โดยมีการกระตุ้นทางจมูกทั้งหมด (มี CS ปัจจุบัน, CS ไม่อยู่) เนื่องจากสารแปรปรวนที่แยกจากกันไม่ได้ให้ผลกระทบอย่างมีนัยสำคัญของความแปรปรวนร่วมในขณะที่ CS มีอยู่หรือไม่อยู่ F(1, 33) <0.14, p > 0.71 ดังนั้นวิธีการตอบสนองโดยไม่ใช้ยาจึงไม่ได้อธิบายถึงความแตกต่างของกลุ่มในการตอบสนองของหัวรถจักรต่อ AMPH ทั้งสองขนาด
การสนทนา
การแพ้ที่พัฒนาขึ้นเพื่อผลของแอมเฟตามีน 1.0 มก. / กก. ข้อมูลความเคยชินและน้ำเกลือยืนยันว่าผลกระทบนี้ไม่ได้เกิดจากความแตกต่างที่มีอยู่ก่อนแล้วความคาดหวังหรือการตอบสนองที่เกี่ยวข้องกับความเครียดต่อการฉีดยา ANCOVAs ที่มีจมูกโผล่ยืนยันว่าผลกระทบเหล่านี้ไม่ได้เกิดจากพฤติกรรมการใช้ยาฟรี ข้อมูลการโผล่จมูกบ่งชี้ว่ากลุ่มต่างๆได้รับความสัมพันธ์ระหว่าง CS และโอกาสที่จะได้รับรางวัลซูโครส ระดับอันดับของการตอบสนองของจมูกชี้เมื่อสิ้นสุดการฝึกอบรมตรงกับความถี่โดยรวมของรางวัลภายใต้ตารางเวลาที่แตกต่างกันจากสูงสุด (กลุ่ม 100) ไปจนถึงต่ำสุด (กลุ่ม 0) เช่นเดียวกับในการทดลอง 2 ค่าเฉลี่ยโดยรวมที่ค่อนข้างต่ำกว่า ระดับการโผล่จมูกในการทดลองนี้เมื่อเทียบกับการทดลองที่ 1 และ 2 อาจสะท้อนถึงวิธีการเลือกที่ตอบสนองต่อสัญญาณเพื่อให้รางวัลในหนูลูอิส (Kosten และคณะ, 2007).
ข้อมูลปริมาณ 0.5 mg / kg แสดงให้เห็นว่า locomotor เริ่มต้นตอบสนองต่อ AMPH ในหนู Lewis (รูปที่ 9) ถูกระงับค่อนข้างเมื่อเปรียบเทียบกับหนู Sprague Dawley (ทดลอง 2; รูปที่ 5) แต่การเพิ่มขึ้นของกลุ่มภายในในการตอบสนองต่อปริมาณที่สองในหนูลูอิสเป็นจำนวนมาก (เกือบสองเท่าของการตอบสนองต่อปริมาณ 0.5 mg / kg แรก) หลังจากตามระบบการปกครอง AMPH 5 เซสชันที่สะดุดตาที่สุดกลุ่ม 50 แสดงการตอบสนองที่ดีกว่า กว่าทุกกลุ่มยกเว้นกลุ่ม 25 ไปยังกลุ่มที่สอง (เช่นโพสต์อาการแพ้) ปริมาณ 0.5 mg / kg AMPH และการตอบสนองที่ดีกว่ากลุ่มอื่น ๆ รวมถึงกลุ่ม 25 ไปยังขนาด AMP 1 mg / kg สุดท้าย (การแพ้สุดท้าย) .
การวิเคราะห์สรุปการจัดอันดับกลุ่มในการทดลอง
เพื่อตรวจสอบความน่าเชื่อถือของความแตกต่างของกลุ่มในการแพ้การวิเคราะห์ที่ไม่ใช่พารามิเตอร์ประเมินความบังเอิญระหว่างกลุ่มและอันดับของการตอบสนองของหัวรถจักรเฉลี่ยต่อ AMPH ที่สอง (โพสต์เรื้อรังเรื้อรัง) ปริมาณ 0.5 mg / kg สุดท้ายและปริมาณ 1.0 mg / kg สุดท้ายของ AMPH จากการทดสอบ 3 การวิเคราะห์ให้ผลที่สำคัญ, φ = 0.986, p = 0.025 ซึ่งสะท้อนถึงความจริงที่ว่ากลุ่ม 50 ได้รับการจัดอันดับเป็นอันดับแรกในทุกกลุ่มยกเว้นการเปรียบเทียบอย่างใดอย่างหนึ่ง อันดับที่เหนือกว่าของกลุ่ม 50 เมื่อเทียบกับกลุ่มอื่น ๆ ในการตอบสนองต่อ AMPH ที่สอง (โพสต์เรื้อรังเรื้อรัง) ปริมาณ 0.5 mg / kg อยู่ในรูป 5 (ทดลอง 2) และรูปที่ 9 (ทดลอง 3) อันดับที่เหนือกว่าของกลุ่ม 50 ที่สัมพันธ์กับกลุ่มอื่น ๆ ในการตอบสนองต่อปริมาณ 1.0 สุดท้าย / ปริมาณ mg อยู่ในรูป 2 (ทดลอง 1) และรูปที่ 10 (ทดลอง 3) ข้อยกเว้นเพียงอย่างเดียวสำหรับรูปแบบนี้คือการตอบสนองต่อปริมาณ 1.0 mg / kg สุดท้ายในหนู Sprague-Dawley ในการทดลอง 2
พูดคุยเรื่องทั่วไป
การทดลองในปัจจุบันได้ทดสอบสมมติฐานที่ว่าการได้รับผลตอบแทนตามกำหนดเวลาเหมือนการพนันอย่างเรื้อรังสามารถทำให้สมองเกิดความเสียหายได้อย่างรวดเร็วเช่นเดียวกับการใช้ยาในทางที่ผิด หลักฐานที่แสดงถึงผลกระทบดังกล่าวจะแนะนำว่า neuroplasticity ซึ่งเป็นความคิดแบบเดียวกันที่นำไปสู่การติดยาสามารถเกิดขึ้นได้จากการสัมผัสเรื้อรังกับตารางรางวัลที่คาดเดาไม่ได้ สอดคล้องกับวรรณกรรมที่เกี่ยวกับการติดยาเสพติดการตอบสนองของหัวรถจักรต่อปริมาณ 0.5 และ 1.0 mg / kg ของ AMPH ที่ทำดัชนีการตอบสนองของระบบ DA ด้วยการเคลื่อนที่ที่มากขึ้นในการตอบสนองต่อปริมาณที่กำหนดในภายหลัง Robinson และ Berridge, 1993; เพียร์ซและคาลิวาส 1997; Vanderschuren และ Kalivas, 2000).
โดยรวมแล้วผลลัพธ์เป็นไปตามสมมติฐานของเรา อย่างไรก็ตามพวกเขายังระบุความแปรปรวนจำนวนมากในผลการทดลองเนื่องจากปัจจัยขั้นตอน ผลกระทบของตารางการปรับสภาพนั้นเรียบง่าย แต่มีความสอดคล้องกับกลุ่ม 50 แสดงให้เห็นถึงการตอบสนองที่ดีกว่ากลุ่มอื่น ๆ ทั้งสี่กลุ่มต่อปริมาณที่ได้รับทั้งห้าตามระบบการให้ยาทั้งห้า ถึงแม้ว่าโดยรวมแล้ว F- ค่าสำหรับผลกระทบที่เกี่ยวข้องกับกลุ่มในการวิเคราะห์ความแปรปรวนมักจะไม่สำคัญความแตกต่างของกลุ่มที่สำคัญได้รับการยืนยันด้วยการเปรียบเทียบตามแผนคู่ ในเรื่องนี้ควรสังเกตว่า“ ความคิดในปัจจุบันมีความสำคัญโดยรวม [สำหรับ F ใน ANOVA] ไม่จำเป็น ก่อนอื่นสมมติฐานที่ทดสอบโดยการทดสอบโดยรวมและการทดสอบแบบเปรียบเทียบหลายแบบนั้นแตกต่างกันมากโดยมีระดับพลังงานที่แตกต่างกันมาก ตัวอย่างเช่นภาพรวม F จริง ๆ แล้วกระจายความแตกต่างระหว่างกลุ่มตามจำนวนองศาอิสระสำหรับกลุ่ม นี่เป็นผลของการเจือจางโดยรวม F ในสถานการณ์ที่หลายกลุ่มมีค่าเท่ากัน แต่แตกต่างจากค่าเฉลี่ยอื่น ๆ ” (ธรรมด๊าธรรมดา, 1992, p. 338) นี่คือสถานการณ์ที่นำไปใช้อย่างแม่นยำในการทดลองปัจจุบันซึ่งคาดว่ากลุ่ม 50 จะแตกต่างจากกลุ่ม 0 และกลุ่ม 100 แต่ไม่มีความแตกต่างระหว่างกลุ่มควบคุมเหล่านี้สำหรับกลุ่ม 25 หรือกลุ่ม 75
ข้อมูลกระตุ้นจมูกยืนยันว่าในการทดลองทุกครั้งสัตว์จะได้รับความสัมพันธ์ระหว่าง CS และโอกาสในการได้รับซูโครส การติดต่อกันระหว่างความถี่ในการกระตุ้นจมูกสำหรับกลุ่มที่แตกต่างกันและความถี่โดยรวมของรางวัลภายใต้ตารางการฝึกอบรมของพวกเขานั้นแสดงให้เห็นว่าอัตราเฉลี่ยของซูโครสที่ได้รับคำแนะนำวิธีการปราศจากยาเสพติด อย่างไรก็ตามการขาดผลกระทบของการกระตุ้นจมูกต่อการตอบสนองของ locomotor ที่เกี่ยวข้องกับกลุ่มของ AMPH ใน ANCOVAs บ่งชี้ว่ากระบวนการแยกกันรองรับพฤติกรรมทั้งสอง
ในบางกรณีผลกระทบของตารางการปรับสภาพเห็นได้ชัดในการตอบสนองต่อปริมาณ AMPH แรก ในกรณีอื่นมันเกิดขึ้นหลังจากปริมาณซ้ำ ความแตกต่างของกลุ่มในการตอบสนองของหัวรถจักรต่อยา AMPH แรกแนะนำว่าการได้รับตารางการให้รางวัลเหมือนการพนันนั้นเพียงพอที่จะทำให้เกิดอาการแพ้ได้ ความแตกต่างของกลุ่มในการเคลื่อนที่ตามขนาดของ AMPH หลายครั้งบ่งบอกถึงผลกระทบที่ลึกซึ้งยิ่งขึ้นซึ่งอาจมีลักษณะเป็น“ ความอ่อนไหว” ซึ่งจะปรากฏเมื่อรวมกับการสัมผัสกับสารกระตุ้นอาการแพ้หลักอย่างต่อเนื่อง (เช่นแอมเฟตามีน)
ความแตกต่างในรูปแบบของการตอบสนองในการทดลองแสดงให้เห็นว่าช่วงเวลาที่ยาวนานขึ้นระหว่างการฝึกอบรมและการท้าทาย AMPH เริ่มต้นอาจเพิ่มโอกาสในการตรวจจับผลกระทบที่ไวต่อการรับสัมผัสของการปรับสภาพด้วยตนเอง สิ่งนี้สะท้อนให้เห็นว่าผลกระทบจากการได้รับผลตอบแทนตามเงื่อนไขอาจฟักตัวเมื่อเวลาผ่านไปปรากฏการณ์หนึ่งที่เห็นด้วยการกระตุ้นอาการแพ้Grimm et al., 2006) รูปแบบของการตอบสนองต่อแอมเฟตามีนในสองขนาดแสดงให้เห็นว่ายา 0.5 mg / kg อาจมีประสิทธิภาพมากกว่าในการเปิดเผยผลกระทบของประวัติปรับอากาศ ในทางกลับกันนี่แสดงให้เห็นว่าเอฟเฟกต์ปรับอากาศภายใต้โปรโตคอลการฝึกอบรมในปัจจุบันนั้นค่อนข้างบอบบางและอาจถูกพรางด้วยเอฟเฟกต์เพดานภายใต้ขนาดของ AMPH และเงื่อนไขที่สร้างขึ้น de novo แพ
ในการทดลอง 3 รูปแบบ biphasic ของการตอบสนองต่อปริมาณ 0.5 mg / kg และความก้าวหน้าที่เหนือกว่าในกลุ่ม 50 สอดคล้องกับโปรไฟล์ที่คาดไว้สำหรับหนู Lewis ในการตอบสนองต่อ methamphetamine (Camp et al., 1994) สิ่งนี้ให้การสนับสนุนความถูกต้องของการค้นพบในปัจจุบันและแนะนำการทับซ้อนระหว่างปัจจัยที่มีช่องโหว่ปานกลางถึงอาการแพ้ทางจิตและการให้รางวัลแบบกำหนดการพนัน
จากการทดลองพบว่าการตอบสนองของหัวรถจักรหลังการแพ้ของกลุ่ม 50 นั้นเกินกว่ากลุ่มอื่น ๆ ภายใต้ปริมาณยาบ้าที่แตกต่างกันและในสัตว์ที่ต่างกัน อย่างไรก็ตามความแปรปรวนภายในกลุ่มสูงและขนาดผลระหว่างกลุ่มเล็ก ๆ น้อย ๆ บ่งบอกถึงบทบาทสำหรับปัจจัยอื่น ๆ ในการทำปฏิกิริยาของระบบ DA ต่อแอมเฟตามีนหลังจากการสัมผัสกับตารางเวลาที่แตกต่างกันของรางวัลซูโครสปรับอากาศ แม้ว่าการตอบสนองของเซลล์ประสาท DA เพื่อรับรางวัลสัญญาณอาจให้รูปแบบการพนันที่หยาบ (Fiorillo et al., 2003) เช่นเดียวกับทุกรุ่นที่มีการสูญเสียข้อมูลเพื่อประโยชน์ของความประหยัด - เพื่อแสดงให้เห็นถึงกระบวนการที่สำคัญ เป็นผลให้รูปแบบของผลกระทบทั่วทั้งเงื่อนไข CS-US ใน Fiorillo และคณะ การศึกษาไม่ได้พูดคุยกันโดยทั่วไปเพื่อตอบสนองต่อแอมเฟตามีน การปรับแต่งเพิ่มเติมของโมเดลได้รับการเรียกร้องให้จับภาพด้านการพนันที่ส่งผลกระทบต่อการทำงานของระบบ DA อย่างเต็มที่
เมื่อนำมารวมกันผลลัพธ์ของการทดลองในชุดนี้ให้การสนับสนุนชั่วคราวสำหรับสมมติฐานที่ว่าการได้รับผลกระทบจากกำหนดการเล่นการพนันแบบเรื้อรังจะช่วยเพิ่มการตอบสนองของระบบสมอง DA ต่อความท้าทายทางจิต เช่นนี้พวกเขาขยายการค้นพบของ นักร้องและคณะ (2012) ที่แสดงให้เห็นว่าเมื่อเทียบกับกำหนดเวลาการสัมผัสก่อนที่จะมีตารางเวลาการเสริมตัวแปรในกระบวนทัศน์การผ่าตัดช่วยเพิ่มการตอบสนองของหัวรถจักรที่ตามมาเพื่อยาบ้า โดยเฉพาะอย่างยิ่งการค้นพบในปัจจุบันชี้ไปที่ความไม่แน่นอนของการให้รางวัลเนื่องจากปัจจัยสำคัญที่เป็นพื้นฐานของผลกระทบของรางวัลที่ผันแปร ขนาดของเอฟเฟกต์ในกระบวนทัศน์ผู้ดำเนินการมีค่ามากกว่าผลกระทบที่พบในการทดลองปัจจุบัน สิ่งนี้อาจสะท้อนถึงกิจกรรมการเล่นการพนันที่เรื้อรังมากขึ้น (55 กับ 15 วัน) มันอาจสะท้อนให้เห็นถึงผลกระทบของการต้องมีการตอบสนองการผ่าตัดเพื่อลุ้นรับรางวัล (เช่นบทบาทสำหรับหน่วยงาน) มากกว่าการสัมผัสแฝงเช่นในการศึกษาปัจจุบัน การเพิ่มระยะเวลาของการฝึกอบรมในกระบวนทัศน์ปัจจุบันจะช่วยแก้ไขคำถามเหล่านี้
ความถูกต้องของการให้รางวัลผันแปรและกำหนดการเสริมเนื่องจากรูปแบบการพนันไม่สามารถรวบรวมได้จากการทดลองเหล่านี้ การวิจัยในอนาคตที่ตรวจสอบผลกระทบของประวัติปรับอากาศต่อพฤติกรรมเสี่ยงในงานการพนันหนูสามารถแก้ไขปัญหานี้ ในทำนองเดียวกันการโต้ตอบระหว่างการตอบสนองพฤติกรรมที่พบได้ที่นี่และการตอบสนอง DA ระดับสูงในการตอบสนองต่อแอมเฟตามีนที่เพิ่งพบในนักพนันทางพยาธิวิทยาต้องรอการสอบสวนเพิ่มเติม (Boileau และคณะ, 2013) การล้างไตด้วยวิธีไมโครสามารถตอบคำถามนี้ได้และการคาดการณ์จากข้อมูลของมนุษย์ก็คือว่าการเปิดตัว DA มากขึ้นในกลุ่ม 50“ ฟีโนไทป์การพนัน” จะสังเกตได้ชัดเจนที่สุดใน striatum หลัง (sensorimotor) มากกว่า ventral (limbic) striatum . การตรวจสอบความถูกต้องของตัวแปร 50% CS + การได้รับรางวัลในกระบวนทัศน์อื่น ๆ เหล่านี้จะสนับสนุนยูทิลิตี้ในฐานะรุ่นทดลองที่แท้จริงโดยสิ้นเชิงของ PG
ในขณะที่การพนันบางรูปแบบมีการตอบสนองโดยใช้เครื่องมืออย่างชัดเจน (เช่นสล็อตแมชชีน) ในรูปแบบอื่น ๆ ของการพนัน (เช่นลอตเตอรี) การเชื่อมโยงระหว่างการกระทำ (การซื้อตั๋วเช่นการวางเดิมพัน) สัญญาณสำหรับรางวัล (เช่น , หมายเลขลอตเตอรี) และรางวัล (หมายเลขที่ชนะและผลตอบแทนที่เป็นตัวเงิน) จะกระจายมากขึ้น อย่างไรก็ตามการเปิดใช้งาน DA ในช่วง CS-US อาจเกิดขึ้นได้ดี สิ่งนี้อาจอธิบายได้ว่าเหตุใดเมื่อมีการประกาศ“ หมายเลขที่ชนะ” ความสนใจจึงถูกตรึงใจเมื่อลอตเตอรี่แต่ละลูกลดลงติดต่อกันเพื่อเขียนลำดับตัวเลขที่เฉพาะเจาะจงในหมายเลขที่ชนะ แม้ว่าความน่าจะเป็นของตัวเลขที่เฉพาะเจาะจงจะถูกกำหนดทางคณิตศาสตร์ แต่ผลลัพธ์ของลอตเตอรีแต่ละลูกจะเป็นแบบไบนารี - ตี (ตรงกับหมายเลขของผู้เล่น) หรือพลาด (ไม่ตรงกับหมายเลขของผู้เล่น) และไม่ทราบผลลัพธ์ของการทดลองใด ๆ สถานการณ์ดังกล่าวอาจบ่งบอกลักษณะของประสบการณ์ของกลุ่ม 50 ในการทดลองปัจจุบันได้ดีขึ้นโดยที่รางวัลนั้นให้โดยไม่เกิดขึ้นโดยบังเอิญ แต่ยังไม่สามารถคาดเดาได้และ CS เพียงระบุถึงศักยภาพในการให้รางวัลโดยไม่เปิดเผยว่าจะเกิดขึ้นในการทดลองหนึ่ง ๆ หรือไม่ เครื่องสล็อตเชื่อมโยงกับ PG มากกว่าตั๋วลอตเตอรี (Cox และคณะ, 2000; Bakken et al., 2009) แสดงถึงบทบาทที่สำคัญสำหรับปัจจัยเครื่องมือ (และความฉับไว) ในแง่มุมที่มีคุณค่าของการพนันสำหรับประชากรกลุ่มนี้ (Loba et al., 2001) อย่างไรก็ตามกระบวนการ Pavlovian ที่จำลองในการทดลองปัจจุบัน (CS + รางวัลที่ไม่แน่นอน) ดูเหมือนจะเป็นสิ่งจำเป็นหากองค์ประกอบของประสบการณ์การพนันไม่เพียงพอ
นอกเหนือไปจากการขาดข้อกำหนดที่ชัดเจนเกี่ยวกับอุปกรณ์จำนวนของคุณสมบัติการออกแบบอื่น ๆ อาจมีส่วนร่วมในรูปแบบที่ค่อนข้างเรียบง่ายและตัวแปรของเอฟเฟกต์การทดลอง กลุ่มที่แตกต่างกันในการสัมผัสซูโครสโดยรวมเช่นเดียวกับความบังเอิญระหว่าง CS และรางวัลซูโครส แม้ว่าสิ่งนี้อาจมีส่วนทำให้เกิดความแปรปรวนระหว่างกลุ่ม แต่ก็ไม่สามารถอธิบายได้อย่างง่ายดายว่าทำไมสัตว์ที่ได้รับซูโครสที่ยิ่งใหญ่ที่สุด (กลุ่ม 100) แสดงอาการแพ้น้อยกว่ากลุ่ม 50 นอกจากนี้กลุ่ม 0 ไม่ได้รับการกระตุ้นก่อนที่จะได้รับซูโครสในทุก ๆ การทดลอง แม้ว่าสิ่งนี้จะจำกัดความคาดหวังของรางวัลที่ชักนำ แต่ก็ไม่ได้ควบคุมสิ่งกระตุ้นก่อนที่จะส่งมอบรางวัลซึ่งมีอยู่ในกลุ่มอื่น ๆ ทั้งหมด เพื่อแก้ไขปัญหานี้การวิจัยในอนาคตควรมีเงื่อนไขที่สัตว์จะได้รับรางวัลในทุก ๆ การทดลองหลังจากได้รับการกระตุ้นแบบเป็นกลาง (เช่นการปรากฏตัวของพวกเขาไม่ได้บ่งบอกถึงศักยภาพของการให้รางวัล)
ข้อ จำกัด ในการออกแบบอีกประการหนึ่งคือการเกิดขึ้นของพฤติกรรมเสริมที่อาจมีผลต่อผลกระทบของตารางการฝึกอบรม ในหน้าของความไม่แน่นอนสัตว์อาจพัฒนาพฤติกรรมที่เชื่อโชคลางที่ออกแบบมาเพื่อปรับปรุงการควบคุมการรับรู้และลดการกระตุ้นการกระตุ้น DA ที่ไม่แน่นอน (เปรียบเทียบ แฮร์ริสและคณะ 2013) ดังนั้นจึงเป็นไปได้ว่าการควบคุมการออกแบบด้านการทดลองช่วยให้สัตว์สามารถชดเชยผลกระทบของตารางการปรับสภาพ ผลดังกล่าวอาจนำไปสู่การตอบสนองที่ค่อนข้างเรียบง่ายและตอบสนองต่อแอมเฟตามีนในกลุ่ม 50 หลังจากการฝึกอบรม CS + ซูโครส การวิจัยในอนาคตควรบันทึกพฤติกรรมที่เกิดขึ้นเองนอกเหนือจากการเจาะจมูกในระหว่างการฝึกซ้อมเพื่อทดสอบความเป็นไปได้นี้และการควบคุมมันควรจะเกิดขึ้นทางสถิติ เนื่องจากพฤติกรรมดังกล่าวคาดว่าจะตอบโต้หรือลดผลกระทบจากความไม่แน่นอนที่เกิดขึ้นตามตารางดังนั้นจึงควรเพิ่มการตอบสนองของแอมเฟตามีนในกลุ่ม 50 เมื่อมีการควบคุม (ขั้นตอนหรือสถิติ) ดังนั้นการออกแบบปัจจุบัน (ไม่มีการควบคุม) ให้การทดสอบแบบอนุรักษ์นิยมของผลกระทบของรางวัล 50% CS + ต่อการแพ้แอมเฟตามีน
ในแง่ของความถูกต้องภายนอกการใช้หนูตัวผู้ยัง จำกัด การใช้งานทั่วไปของผลลัพธ์ การขาดเงื่อนไข“ การลงโทษ” ที่ชัดเจนนั้นแตกต่างจากการพนันซึ่งการสูญเสียเงินจำนวนมากเป็นเรื่องปกติและออกแรงจูงใจสำคัญ (Nieuwenhuis และคณะ, 2005; ซิงห์และข่าน 2012) ความสามารถในการสะสมรางวัลนั้นขาดไปจากกระบวนทัศน์ปัจจุบันและการชนะสะสมในเกมสล็อตแมชชีนพบว่ามีการโต้ตอบกับกิจวัตร DA ในมนุษย์ (Tremblay และคณะ 2011; สมาร์ทและอัล 2013) ในทำนองเดียวกันโอกาสในการแจ็คพอตเป็นความแตกต่างที่สำคัญระหว่างรูปแบบปัจจุบันและการพนันจริง
แม้จะมีข้อ จำกัด เหล่านี้ แต่ผลลัพธ์ในปัจจุบันชี้ให้เห็นว่ารางวัลตัวแปร CSN + 50% สามารถมีส่วนร่วมกับวิถีทาง DA ที่เกี่ยวข้องในการเสริมผลกระทบของการพนัน (Fiorillo et al., 2003; Anselme, 2013) การตอบโต้ข้ามการตอบสนองต่อ AMPH ตามกำหนดการเล่นการพนันนี้สอดคล้องกับบทบาทสำคัญสำหรับ DA ในการพนันและผลกระทบของยาจิตZack และ Poulos, 2009) และขยายการศึกษาก่อนหน้านี้เกี่ยวกับ cross-priming ของแรงจูงใจในการเดิมพันโดย AMPH ในการพนันทางพยาธิวิทยา (Zack และ Poulos, 2004) ผลในปัจจุบันยังแนะนำโดยอ้อมว่าปริมาณเล็กน้อยของ AMPH ซึ่งไม่ก่อให้เกิดการปลดปล่อย DA ทางสรีรวิทยาอาจทำให้รูปแบบการทำงานของสมองดีขึ้นในการตอบสนองต่อสัญญาณรางวัลเป็นระยะ ๆ (เช่นระหว่างการเล่นพนัน) มากกว่าการสัมผัสสูง ปริมาณยากระตุ้น (cf. Vanderschuren และ Pierce, 2010) การสนับสนุนโดยตรงสำหรับการติดต่อนี้อาจเกิดขึ้นได้จากการประเมินการปล่อย DA เพื่อตอบสนองต่อกำหนดการตาราง CS-US ตัวแปร 50% และขนาดต่าง ๆ ของ AMPH โดยใช้ microdialysis
จากมุมมองของการทดลองแบบจำลอง Pavlovian ปัจจุบันและแบบจำลองการเสริมกำลังตัวแปรก่อนหน้านี้ทั้งคู่ดูเหมือนจะทำให้เกิดฟีโนไทป์ที่คล้ายกับนักพนันทางพยาธิวิทยาของมนุษย์ เช่นนี้พวกเขามีส่วนประกอบที่มีคุณค่าสำหรับงานการพนันหนูซึ่งรูปแบบพฤติกรรมการเล่นการพนัน (เป็นมาตรการขึ้นอยู่กับ) แต่มีจนถึงขณะนี้เพียงใช้สัตว์ที่มีสุขภาพเทียบเท่ากับการเล่นการพนันทางสังคมของมนุษย์ บนพื้นฐานของวรรณกรรมสัตว์ที่สัมผัสกับรางวัลที่ผันแปรอาจแตกต่างกันในงานเหล่านี้โดยเฉพาะอย่างยิ่งในการตอบสนองต่อยา DA-ergic การรวมฟีโนไทป์การพนันของหนูกับงานการพนันอาจอนุญาตให้มีการพัฒนายาสำหรับรักษา PG ซึ่งอาจไม่สามารถทำได้อย่างสมบูรณ์กับสัตว์ที่มีสุขภาพเพียงอย่างเดียว การปรับแต่งเพิ่มเติมในการออกแบบการทดลองและระบบการฝึกอบรมตามที่อธิบายไว้ข้างต้นควรปรับปรุงการติดต่อระหว่างสัตว์ที่ได้รับการฝึกฝนในกระบวนทัศน์นี้และนักพนันทางพยาธิวิทยาที่เกิดขึ้นจริง
จากมุมมองทางคลินิกสังคมวิทยาการค้นพบการสัมผัสกับรางวัล 50% ตัวแปร CS + ซึ่งใกล้เคียงกับตารางการให้รางวัลบนเครื่องสล็อตเชิงพาณิชย์ (Tremblay และคณะ 2011) การเปลี่ยนแปลงระบบ DA ของสมองด้วยวิธีที่เชื่อถือได้และยั่งยืนชี้ให้เห็นว่าในบางกรณีกิจกรรมการพนันเช่นยาเสพติดอาจเป็น "เชื้อโรค" ที่สามารถทำให้ติดยาได้ อย่างไรก็ตามขนาดผลกระทบเล็กน้อยและความแปรปรวนสูงในการตอบสนองต่อรางวัล 50% CS + แนะนำว่าเช่นเดียวกับยาเสพติดในทางที่ผิดแนวโน้มของตารางการให้รางวัลเหมือนการพนันเพื่อส่งเสริมการติดจะขึ้นอยู่กับความเสี่ยงที่มีอยู่ก่อนของนักพนัน อย่างไรก็ตามเพื่อสำรองบุคคลที่มีความเสี่ยงสูงที่อาจได้รับผลกระทบที่เกี่ยวข้องกับการเล่นการพนันดูเหมือนว่ามีเหตุผลที่นโยบายที่ใช้ในการยับยั้งการใช้และลดอันตรายจากยาเสพติดอาจถูกขยายไปสู่การพนันเช่นกัน
คำชี้แจงความขัดแย้งทางผลประโยชน์
ผู้เขียนประกาศว่าการวิจัยได้ดำเนินการในกรณีที่ไม่มีความสัมพันธ์ทางการค้าหรือทางการเงินใด ๆ ที่อาจตีความได้ว่าเป็นความขัดแย้งทางผลประโยชน์ที่อาจเกิดขึ้น
กิตติกรรมประกาศ
งานวิจัยนี้ได้รับทุนจากทุนจากสภาวิทยาศาสตร์ธรรมชาติและวิศวกรรมแห่งแคนาดาถึง Paul J. Fletcher เราขอขอบคุณคุณ Djurdja Djordjevic อย่างจริงใจสำหรับการเตรียมตัวเลข
อ้างอิง
Anselme, P. (2013) โดปามีนแรงจูงใจและความสำคัญเชิงวิวัฒนาการของพฤติกรรมการเล่นการพนัน Behav ความต้านทานของสมอง. 256, 1 – 4 ดอย: 10.1016 / j.bbr.2013.07.039
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
APA (2013) คู่มือการวินิจฉัยและสถิติของความผิดปกติทางจิต 5th Edn. อาร์ลิงตัน, เวอร์จิเนีย: สำนักพิมพ์จิตเวชอเมริกัน
Bakken, IJ, Gotestam, KG, Grawe, RW, Wenzel, HG และ Oren, A. (2009) พฤติกรรมการพนันและปัญหาการพนันในนอร์เวย์ 2007 Scand J. Psychol. 50, 333 – 339 doi: 10.1111 / j.1467-9450.2009.00713.x
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Boileau, I. , ผู้จ่าย, D. , Chugani, B. , Lobo, DS, Houle, S. , Wilson, AA, et al. (2013) ในร่างกาย หลักฐานการปล่อยโดปามีนจากแอมเฟตามีนที่มากขึ้นในการพนันทางพยาธิวิทยา: การศึกษาเอกซเรย์ปล่อยโพซิตรอนด้วย [C] - (+) - PHNO mol จิตเวช doi: 10.1038 / mp.2013.163 [Epub ก่อนพิมพ์]
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Bolles, RC (1972) การเสริมแรงความคาดหวังและการเรียนรู้ จิตวิทยา การหมุนรอบ. 79, 394 – 409 doi: 10.1037 / h0033120
ค่าย DM, Browman, KE และ Robinson, TE (1994) ผลของเมทแอมเฟตามีนและโคเคนต่อพฤติกรรมยนต์และโดปามีนนอกเซลล์ใน ventat striatum ของ Lewis กับ Fischer 344 หนู สมอง Res. 668, 180–193. doi: 10.1016/0006-8993(94)90523-1
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Conversano, C. , Marazziti, D. , Carmassi, C. , Baldini, S. , Barnabei, G. , และ Dell'Osso, L. (2012) การพนันทางพยาธิวิทยา: การทบทวนอย่างเป็นระบบเกี่ยวกับการค้นพบทางชีวเคมีการสร้างภาพทางระบบประสาทและระบบประสาทวิทยา Harv รายได้จิตเวช 20, 130 – 148 doi: 10.3109 / 10673229.2012.694318
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Cox, BJ, Kwong, J. , Michaud, V. , และ Enns, MW (2000) ปัญหาและการพนันทางพยาธิวิทยาที่น่าจะเป็น: การพิจารณาจากการสำรวจชุมชน สามารถ. เจจิตเวช 45, 548-553
Dalley, JW, Theobald, DE, Eagle, DM, Passetti, F. , และ Robbins, TW (2002) การขาดดุลในการควบคุมแรงกระตุ้นที่เกี่ยวข้องกับฟังก์ชั่น serotonergic สูงขึ้นในเยื่อหุ้มสมองหนู prefrontal Neuropsychopharmacology 26, 716–728. doi: 10.1016/S0893-133X(01)00412-2
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Everitt, BJ และ Robbins, TW (2005) ระบบประสาทของการเสริมแรงสำหรับการติดยาเสพติด: จากการกระทำไปจนถึงนิสัยการบังคับ ชัยนาท Neurosci. 8, 1481 – 1489 ดอย: 10.1038 / nn1579
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Fiorillo, CD, Tobler, PN และ Schultz, W. (2003) การเข้ารหัสแบบไม่ต่อเนื่องของรางวัลความน่าจะเป็นและความไม่แน่นอนโดยเซลล์ประสาทโดปามีน วิทยาศาสตร์ 299, 1898 – 1902 ดอย: 10.1126 / วิทยาศาสตร์. 1077349
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Flagel, SB, Watson, SJ, Akil, H. , และ Robinson, TE (2008) ความแตกต่างของแต่ละบุคคลในการระบุถึงสิ่งกระตุ้นที่มีผลต่อคิวที่เกี่ยวข้องกับการให้รางวัล: มีอิทธิพลต่อการกระตุ้นอาการแพ้โคเคน Behav ความต้านทานของสมอง. 186, 48 – 56 ดอย: 10.1016 / j.bbr.2007.07.022
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Fletcher, PJ, Tenn, CC, Rizos, Z. , Lovic, V. , และ Kapur, S. (2005) การทำให้เป็นแอมเฟตามีน แต่ไม่ใช่ PCP ทำให้การตั้งค่าเปลี่ยนไปโดยไม่ตั้งใจลดลง: กลับมาโดยตัวเอกผู้รับ D1 ซึ่งถูกฉีดเข้าไปในเยื่อหุ้มสมอง prefrontal ที่อยู่ตรงกลาง Psychopharmacology (Berl.) 183, 190–200. doi: 10.1007/s00213-005-0157-6
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Flores, G. , Wood, GK, Barbeau, D. , Quirion, R. , และ Srivastava, LK (1998) Lewis และ Fischer หนู: การเปรียบเทียบระดับการขนส่งโดปามีนและตัวรับ สมอง Res. 814, 34–40. doi: 10.1016/S0006-8993(98)01011-7
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Frascella, J. , Potenza, MN, Brown, LL และ Childress, AR (2010) ช่องโหว่ของสมองที่ใช้ร่วมกันเปิดทางให้ผู้เสพติดไม่ติดสาร: แกะติดที่ข้อต่อใหม่? แอน NY Acad วิทย์. 1187, 294 – 315 doi: 10.1111 / j.1749-6632.2009.05420.x
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Grimm, JW, Buse, C. , Manaois, M. , Osincup, D. , Fyall, A. และ Wells, B. (2006) การแยกตัวออกจากกันของเวลาขึ้นอยู่กับผลกระทบของการตอบสนองต่อยาโคเคนต่อความอยากซูโครสและการเคลื่อนที่ Behav Pharmacol. 17, 143 – 149 ดอย: 10.1097 / 01.fbp.0000190686.23103.f8
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Haile, CN, Hiroi, N. , Nestler, EJ และ Kosten, TA (2001) การตอบสนองเชิงพฤติกรรมที่แตกต่างของโคเคนนั้นสัมพันธ์กับการเปลี่ยนแปลงของโปรตีนโดปามีนใน mesolimbic ในหนู Lewis และ Fischer 344 ไซแนปส์ 41, 179 – 190 doi: 10.1002 / syn.1073
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Harris, JA, Andrew, BJ และ Kwok, DW (2013) วิธีการอ่านนิตยสารระหว่างสัญญาณอาหารขึ้นอยู่กับพาฟโลเวียน J. ประสบการณ์ จิตวิทยา Anim Behav กระบวนการ. 39, 107 – 116 doi: 10.1037 / a0031315
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
ธรรมด๊าธรรมดา, DC (1992) วิธีการทางสถิติสำหรับจิตวิทยา. บอสตันแมสซาชูเซตส์: Duxbury
Ito, R. , Dalley, JW, Robbins, TW และ Everitt, BJ (2002) โดปามีนปล่อยออกมาในหลัง striatum ระหว่างพฤติกรรมการค้นหาโคเคนภายใต้การควบคุมของคิวยาที่เกี่ยวข้อง J. Neurosci. 22, 6247 – 6253
Jiao, X. , Pare, WP และ Tejani-Butt, S. (2003) ความแตกต่างของความเครียดในการกระจายตัวของโดพามีนในสมองหนู Prog Neuropsychopharmacol Biol จิตเวช 27, 913–919. doi: 10.1016/S0278-5846(03)00150-7
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Koob, GF และ Le Moal, M. (2008) ทบทวน กลไกทางระบบประสาทสำหรับกระบวนการสร้างแรงบันดาลใจของคู่ต่อสู้ในการเสพติด Philos ทรานส์ ร. Lond B Biol วิทย์. 363, 3113 – 3123 doi: 10.1098 / rstb.2008.0094
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Kosten, TA, Miserendino, MJ, Chi, S. และ Nestler, EJ (1994) ฟิสเชอร์และลูอิสหนูสายพันธุ์แสดงผลโคเคนที่แตกต่างกันในการเลือกสถานที่ที่มีการปรับอากาศและการทำให้ไวต่อพฤติกรรม แต่ไม่ได้อยู่ในกิจกรรมของหัวรถจักร เจ Pharmacol ประสบการณ์ Ther. 269, 137 – 144
Kosten, TA, Zhang, XY และ Haile, CN (2007) ความแตกต่างของความเครียดในการดูแลโคเคนการดูแลตนเองและความสัมพันธ์กับการตอบสนองต่อกิจกรรมแปลกใหม่ Behav Neurosci. 121, 380 – 388 ดอย: 10.1037 / 0735-7044.121.2.380
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Leeman, RF และ Potenza, MN (2012) ความเหมือนและความแตกต่างระหว่างการพนันทางพยาธิวิทยากับความผิดปกติในการใช้สารเสพติด: มุ่งเน้นไปที่การกระตุ้นและการบังคับ Psychopharmacology (Berl.) 219, 469–490. doi: 10.1007/s00213-011-2550-7
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Leshner, AI (1997) ติดยาเสพติดเป็นโรคทางสมองและมันเป็นเรื่องสำคัญ วิทยาศาสตร์ 278, 45 – 47 ดอย: 10.1126 / วิทยาศาสตร์. 278.5335.45
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Loba, P. , Stewart, SH, Klein, RM, และ Blackburn, JR (2001) การควบคุมคุณสมบัติของเกมลอตเตอรีวิดีโอลอตเตอรี (VLT): ผลกระทบของการเล่นการพนันทางพยาธิวิทยาและที่ไม่ใช่ทางพยาธิวิทยา J. Gambl แกน. 17, 297 – 320 ดอย: 10.1023 / A: 1013639729908
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Martinez, D. , Narendran, R. , Foltin, RW, Slifstein, M. , Hwang, DR, Broft, A. , et al. (2007) การปลดปล่อยโดปามีนที่เกิดจากแอมเฟตามีน: ทื่ออย่างชัดเจนในการพึ่งพาโคเคนและการทำนายการเลือกโคเคนด้วยตนเอง am J. จิตเวชศาสตร์ 164, 622 – 629 doi: 10.1176 / appi.ajp.164.4.622
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Mateo, Y. , Lack, CM, Morgan, D. , Roberts, DC และ Jones, SR (2005) ลดการทำงานของโดปามีนขั้วและลดความรู้สึกไวต่อโคเคนหลังดื่มโคเคนด้วยตนเองและการกีดกัน Neuropsychopharmacology 30, 1455 – 1463 doi: 10.1038 / sj.npp.1300687
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Nestler, EJ (2001) พื้นฐานระดับโมเลกุลของการติดอยู่กับพลาสติกในระยะยาว ชัยนาท รายได้ Neurosci. 2, 119 – 128 doi: 10.1038 / 35053570
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Nieuwenhuis, S. , Heslenfeld, DJ, von Geusau, NJ, Mars, RB, Holroyd, CB, และ Yeung, N. (2005) กิจกรรมในพื้นที่สมองที่ไวต่อผลตอบแทนของมนุษย์นั้นขึ้นอยู่กับบริบทอย่างมาก Neuroimage 25, 1302 – 1309 doi: 10.1016 / j.neuroimage.2004.12.043
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Pierce, RC และ Kalivas, PW (1997) แบบจำลองวงจรของการแสดงออกของความไวต่อพฤติกรรมต่อแอมเฟตามีนที่มีลักษณะคล้ายจิต ความต้านทานของสมอง ความต้านทานของสมอง การหมุนรอบ. 25, 192 ดอย: 10.1016 / S0165-0173 (97) 00021-0
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Robbins, TW และ Everitt, BJ (1999) ติดยาเสพติด: นิสัยที่ไม่ดีเพิ่มขึ้น ธรรมชาติ 398, 567 – 570 doi: 10.1038 / 19208
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
โรบินสัน, TE, เบกเกอร์, JB, และ Presty, SK (1982) การอำนวยความสะดวกในระยะยาวของพฤติกรรมการหมุนของแอมเฟตามีนที่เกิดขึ้นและการปลดปล่อยโดปามีนแบบ striatal ที่เกิดจากการสัมผัสกับแอมเฟตามีนเพียงครั้งเดียว: ความแตกต่างทางเพศ สมอง Res. 253, 231–241. doi: 10.1016/0006-8993(82)90690-4
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
โรบินสัน, TE, และ Berridge, KC (1993) พื้นฐานทางประสาทของความอยากติดยา: ทฤษฎีการกระตุ้นให้ติดยาเสพติด ความต้านทานของสมอง ความต้านทานของสมอง การหมุนรอบ. 18, 247 – 291 ดอย: 10.1016 / 0165-0173 (93) 90013-P
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
โรบินสัน, TE, และ Berridge, KC (2001) การกระตุ้นและการเสพติด ติดยาเสพติด 96, 103 – 114 doi: 10.1046 / j.1360-0443.2001.9611038.x
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
นักร้อง, BF, Scott-Railton, J. และ Vezina, P. (2012) การเสริมแซคคารินในรูปแบบที่คาดเดาไม่ได้ช่วยเพิ่มการเคลื่อนไหวของแอมเฟตามีน Behav ความต้านทานของสมอง. 226, 340 – 344 ดอย: 10.1016 / j.bbr.2011.09.003
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
ซิงห์, V. , และ Khan, A. (2012) การตัดสินใจเกี่ยวกับการให้รางวัลและการลงโทษในภารกิจการพนันไอโอว่ามีหลักฐานของการมองอนาคตหรือการวางกรอบหรือไม่? ด้านหน้า Neurosci. 6: 107 doi: 10.3389 / fnins.2012.00107
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
สกินเนอร์, BF (1953) วิทยาศาสตร์และพฤติกรรมมนุษย์. New York, NY: กดฟรี
สมาร์ท, K. , Desmond, RC, Poulos, CX และ Zack, M. (2013) Modafinil เพิ่มความสดชื่นให้รางวัลในเกมสล็อตแมชชีนในนักพนันทางพยาธิวิทยาที่มีความดันต่ำและสูง Neuropharmacology 73, 66 – 74 ดอย: 10.1016 / j.neuropharm.2013.05.015
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Tremblay, AM, Desmond, RC, Poulos, CX และ Zack, M. (2011) Haloperidol ปรับเปลี่ยนแง่มุมที่เป็นประโยชน์ของการพนันสล็อตแมชชีนในนักพนันทางพยาธิวิทยาและการควบคุมสุขภาพ ผู้เสพติด Biol. 16, 467 – 484 doi: 10.1111 / j.1369-1600.2010.00208.x
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Vanderschuren, LJ และ Kalivas, PW (2000) การเปลี่ยนแปลงในการส่งโดปามินเนอร์จิคและกลูตามาเทอจิคในการเหนี่ยวนำและการแสดงออกของการกระตุ้นให้เกิดพฤติกรรม: การทบทวนที่สำคัญของการศึกษาพรีคลินิก Psychopharmacology (Berl.) 151, 99 – 120 ดอย: 10.1007 / s002130000493
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Vanderschuren, LJ และ Pierce, RC (2010) กระบวนการแพ้ในการติดยา ฟี้ ด้านบน Behav Neurosci. 3, 179 – 195 ดอย: 10.1007 / 7854_2009_21
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Vanderschuren, LJ, Schoffelmeer, AN, Mulder, AH, และ De Vries, TJ (1999) กลไก Dopaminergic เป็นสื่อกลางในการแสดงออกในระยะยาวของอาการแพ้หัวรถจักรหลังจากการสัมผัสกับมอร์ฟีนหรือแอมเฟตามีน Psychopharmacology (Berl.) 143, 244 – 253 ดอย: 10.1007 / s002130050943
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Verdejo-Garcia, A. , Lawrence, AJ และ Clark, L. (2008) Impulsivity เป็นช่องโหว่สำหรับความผิดปกติของการใช้สาร: ทบทวนผลการวิจัยที่มีความเสี่ยงสูง, นักพนันที่มีปัญหาและการศึกษาความสัมพันธ์ทางพันธุกรรม Neurosci Biobehav การหมุนรอบ. 32, 777 – 810 doi: 10.1016 / j.neubiorev.2007.11.003
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Vezina, P. (2004) อาการแพ้ของปฏิกิริยาโดปามีนเซลล์ประสาท midbrain และการบริหารตนเองของยากระตุ้นจิต Neurosci Biobehav การหมุนรอบ. 27, 827 – 839 doi: 10.1016 / j.neubiorev.2003.11.001
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Volkow, ND, Wang, GJ, Fowler, JS, Logan, J. , Gatley, SJ, Hitzemann, R. , et al. (1997) การลดลงของการตอบสนองแบบ dopaminergic ในทารกแรกเกิดในเรื่องขึ้นกับการล้างพิษโคเคน ธรรมชาติ 386, 830 – 833 doi: 10.1038 / 386830a0
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Volkow, ND, Wang, GJ, Fowler, JS และ Tomasi, D. (2012) วงจรติดยาเสพติดในสมองของมนุษย์ Annu รายได้ Pharmacol Toxicol. 52, 321 – 336 ดอย: 10.1146 / annurev-pharmtox-010611-134625
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Winer, B. (ed.) (1971) หลักการทางสถิติในการออกแบบการทดลอง. นิวยอร์กนิวยอร์ก: McGraw-Hill
Zack, M. , และ Poulos, CX (2004) แอมเฟตามีน primes แรงจูงใจในการเล่นการพนันและเครือข่ายความหมายที่เกี่ยวข้องกับการพนันในการเล่นการพนันปัญหา Neuropsychopharmacology 29, 195 – 207 doi: 10.1038 / sj.npp.1300333
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Zack, M. , และ Poulos, CX (2009) บทบาทแบบขนานสำหรับโดปามีนในการพนันทางพยาธิวิทยาและการเสพติดสารกระตุ้นจิต ฟี้ การใช้ยาในทางที่ผิด. 2, 11 – 25 doi: 10.2174 / 1874473710902010011
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
Zamudio, S. , Fregoso, T. , Miranda, A. , De La Cruz, F. , และ Flores, G. (2005) ความแตกต่างของระดับโดพามีนและพฤติกรรมที่เกี่ยวข้องกับโดพามีนในหนู ความต้านทานของสมอง วัว. 65, 339 – 347 doi: 10.1016 / j.brainresbull.2005.01.009
ตีพิมพ์บทคัดย่อ | เผยแพร่ข้อความแบบเต็ม | CrossRef ข้อความแบบเต็ม
คำสำคัญ: การพนันทางพยาธิวิทยาการทำให้แพ้แอมเฟตามีนโดปามีนความไม่แน่นอน
การอ้างอิง: Zack M, Featherstone RE, Mathewson S และ Fletcher PJ (2014) การสัมผัสเรื้อรังในตารางการเล่นที่เหมือนการพนันที่มีสิ่งเร้าการทำนายผลตอบแทนสามารถส่งเสริมการแพ้แอมเฟตามีนในหนู ด้านหน้า Behav Neurosci. 8: 36 doi: 10.3389 / fnbeh.2014.00036
ได้รับ: 01 พฤศจิกายน 2013; ยอมรับแล้ว: 23 มกราคม 2014;
เผยแพร่ออนไลน์: 11 กุมภาพันธ์ 2014
แก้ไขโดย:
ไบรอันเอฟซิงเกอร์มหาวิทยาลัยมิชิแกนสหรัฐอเมริกา
บทวิจารณ์โดย:
Louk Vanderschurenมหาวิทยาลัยอูเทรคต์เนเธอร์แลนด์
Ruud Van Den Bos, Radboud University Nijmegen, เนเธอร์แลนด์
Patrick Anselme, มหาวิทยาลัยLiège, เบลเยี่ยม
ลิขสิทธิ์© 2014 Zack, Featherstone, Mathewson และ Fletcher นี่เป็นบทความแบบเปิดที่เผยแพร่ภายใต้เงื่อนไขของ ใบอนุญาตแสดงที่มาของครีเอทีฟคอมมอนส์ (CC BY). อนุญาตให้ใช้งานแจกจ่ายหรือทำซ้ำในฟอรัมอื่นโดยผู้แต่งหรือผู้ออกใบอนุญาตจะได้รับเครดิตและสิ่งพิมพ์ต้นฉบับในวารสารนี้ได้รับการอ้างอิงตามแนวทางปฏิบัติทางวิชาการที่เป็นที่ยอมรับ ไม่อนุญาตให้ใช้แจกจ่ายหรือทำซ้ำซึ่งไม่เป็นไปตามข้อกำหนดเหล่านี้
* สารบรรณ: Martin Zack, ห้องปฏิบัติการจิตวิทยาความรู้, แผนกประสาทวิทยาศาสตร์, ศูนย์การติดและสุขภาพจิต, 33 Russell Street, โทรอนโต, ON M5S 2S1, แคนาดา e-mail: [ป้องกันอีเมล]