 Fabrizio Sanna1*,
Fabrizio Sanna1*,  杰西卡布拉祖1,
杰西卡布拉祖1,  Maria A. Piludu2,
Maria A. Piludu2,  Maria G. Corda2,
Maria G. Corda2,  Maria R. Melis1,
Maria R. Melis1,  奥斯瓦尔多乔治2 和
奥斯瓦尔多乔治2 和  Antonio Argiolas1,3
Antonio Argiolas1,3- 1意大利卡利亚里卡利亚里大学生物医学科学系,神经科学和临床药理学科,以及成瘾神经生物学卓越中心
- 2意大利卡利亚里卡利亚里大学生命与环境科学系,制药,药理和营养科学科
- 3意大利卡利亚里Cittadella Universitaria卡利亚里科国家研究委员会神经科学研究所
罗马高 - (RHA)和低回避(RLA)远交大鼠,在快车箱中的主动回避反应的快速与差的获得方面不同,在性生活中存在性活动的差异接受雌性大鼠。 事实上,RHA大鼠表现出比RLA大鼠更高水平的性动机和交配表现,RLA大鼠在反复性活动后也持续存在。 这些差异与RHA大鼠与RLA大鼠的中脑边缘多巴胺能系统的较高基调相关,通过在性活动期间从RHA大鼠获得的透析液中发现的多巴胺的增加高于RLA大鼠。 这项工作表明,细胞外多巴胺和去甲肾上腺素(NA)也从男性RHA和RLA大鼠的内侧前额叶皮层(mPFC)的透析液中增加,在存在不可接近的雌性大鼠的情况下,并且在直接性交互过程中更明显。 多巴胺(及其主要代谢产物3,4-二羟基苯乙酸,DOPAC)和NA的这种增加在性初始和有经验的动物中均有发现,但它们更高:(i)在RHA中比在RLA大鼠中; (ii)在性经验丰富的RHA和RLA大鼠中,而不是在他们的天真对应物中。 最后,mPFC中多巴胺和NA的差异与性活动中的差异同时发生,因为在性初始和经历的情况下,RHA大鼠表现出比RLA大鼠更高的性动机和交配性能水平。 这些结果表明,在mPFC中也发生更高的多巴胺能基调,同时增加的去甲肾上腺素能基调,其可能参与RHA和RLA大鼠中发现的不同交配模式,如中脑边缘多巴胺能系统所示。
介绍
罗马高 - (RHA)和低回避(RLA)远交大鼠系,最初选择用于快速与不良的获取飞梭箱中的主动回避反应(Bignami,1965; Broadhurst和Bignami,1965; Driscoll和Bättig,1982; Fernández-Teruel等,2002; Giorgi等,2007)当存在性接受性雌性大鼠时,表现出性活动的显着差异。 事实上,RHA大鼠表现出比RLA大鼠更高的性动机水平,正如这些大鼠在无法接近的接受女性存在时显示的非接触性阴茎勃起的更高数量所表明的那样,并且比RLA大鼠具有更好的交配性能,如所揭示的通过主要在两个大鼠系之间的第一次交配试验中发现的许多交配参数的显着差异,以及在五次交配试验后,尽管减毒(Sanna等,2014a)。 在RHA和RLA大鼠之间发现的最明显的差异是在第一次交配试验中射出的安装和插入的大鼠的百分比,80%的RHA大鼠分别对抗40%的RLA大鼠(Sanna等,2014a)。 大量实验证据表明,功能不同的多巴胺能基调参与RHA和RLA大鼠之间的交配差异。 事实上,已经发现RHA和RLA大鼠的不同交配模式被阿扑吗啡(一种混合的D1 / D2多巴胺受体激动剂,以促进性行为的剂量给药)和氟哌啶醇(一种D2多巴胺受体拮抗剂)的不同修饰。低剂量抑制性行为(Sanna等,2014b)。 因此,已经发现RLA大鼠分别对阿扑吗啡和氟哌啶醇诱导的性行为的促进和抑制更敏感,这由两种药物诱导的更大修饰证明,主要是在较低剂量下以不同交配参数施用时。 RLA大鼠与RHA大鼠(Sanna等,2014b)。 对于这项研究可能更重要的是,RHA和RLA大鼠的不同交配模式也与中脑边缘多巴胺能神经元的活性差异有关,众所周知,其活性在性行为期间会增加(Pfaus等,1990; Pleim等,1990; Pfaus和Phillips,1991; Damsma等,1992; Wenkstern等,1993; Balfour等人,2004; Pitchers等,2010, 2013; Beloate等,2016)。 事实上,尽管有性生活和性经验的RHA和RLA大鼠在伏隔核的透析液中具有相似的细胞外多巴胺基础值,但细胞外多巴胺和3,4-二羟基苯乙酸(DOPAC,其主要代谢物之一)的浓度也是如此。已经发现,在性行为的预期和完成阶段,从幼稚和性经历的RHA和RLA大鼠伏隔核获得的透析液中差异性增加。 性感天真的RHA和RLA大鼠之间的上述差异更为显着,但在经历过性行为的RHA和RLA大鼠之间存在差异,尽管这些大鼠的性行为差异趋于减弱(Sanna等,2015).
在性活动期间(在性行为的食欲和完成阶段)看到伏隔核中多巴胺能活性增加的功能性作用仍然是争论的问题以及在喂养期间看到的,如性活动,具有强烈的性能动机效价。 因此,尽管伏隔核中的多巴胺参与动机,并且中脑边缘多巴胺能神经元通常被称为有益神经元,但在最近的15年中,最近的研究支持了中脑边缘多巴胺不参与动机或奖励行为的主要表达的假设,而是学习和记忆刺激 - 奖励协会(Agmo等人,1995; Berridge和Robinson,1998; Ikemoto和Panksepp,1999; Pitchers等,2013, 2014; Beloate等,2016; Salamone等,2016)。 根据这一假设,最近发现对伏隔核中的多巴胺受体的阻断或腹侧被盖区域内多巴胺能神经元的失活不能改变雄性大鼠交配行为的食欲和完善方面的表达(Pitchers等,2013, 2014; Beloate等,2016).
然而,不能排除在RHA和RLA大鼠之间的性行为的上述差异中含有多巴胺的其他脑区域的作用。 因此,众所周知,多巴胺对实验动物的性行为的预期和完成阶段以及不仅在伏隔核中的人类也发挥促进作用(Everitt,1990; Pfaus等,1990; Hull等,1991; Pfaus和Everitt,1995; Melis和Argiolas,2011),但也在其他大脑区域,如内侧视前区,下丘脑及其核(即室旁核(PVN); Pfaus和Phillips,1991; Argiolas和Melis,1995, 2005, 2013; Hull等,1995, 1999; Melis和Argiolas,1995; Melis等,2003; Succu等,2007; Pfaus,2010)。 另一个含有多巴胺并且可能在性行为中发挥作用的区域是内侧前额皮质(mPFC; Fernández-Guasti等人,1994; Agmo和Villalpando,1995; Agmo等人,1995; Hernández-Gonzáles等,1998, 2007; Kakeyama等,2003; Balfour等人,2006; Afonso等人,2007; 戴维斯等人,2010; Febo,2011)。 事实上,在这个大脑区域发现了中皮质多巴胺神经元的神经末梢,它们的细胞体位于腹侧被盖区作为中脑边缘多巴胺神经元。 至于伏隔核,这个大脑区域在性行为中的确切作用还远未明确。 实际上,通常发现mPFC的病变不能改变性接受雌性的雄性大鼠的性行为(Fernández-Guasti等人,1994; Agmo和Villalpando,1995; Agmo等人,1995; Hernández-Gonzáles等,1998, 2007; Kakeyama等,2003; Balfour等人,2006; Afonso等人,2007),也不表达对性奖励的条件性地方偏好(戴维斯等人,2010)。 然而,mPFC病变并没有改变条件性地方对性奖励的偏好,在同一动物中消除了与厌恶刺激配对时形成对性活动的条件性厌恶的能力(戴维斯等人,2010并且在雄性大鼠的mPFC中测量了雄性大鼠接近无法接触的性接受雌性的行为期间的选择性细胞发射(Febo,2011)。 这些研究结果表明,性行为期间的mPFC激活在外部和内部信息的整合中起作用,用于执行和控制目标导向行为,而不是表达对天然强化物的先天反应(见 Goto和Grace,2005)。 因此,与伏隔核一起,mPFC是参与调节动机行为(目标导向行为)的复杂神经系统的一部分,其需要整合来自mPFC的认知信息,来自杏仁核的情绪信息和背景 - 相关信息来自海马,伏隔核(Goto和Grace,2005).
实验证据表明,mPFC中多巴胺的释放参与了动机行为的适应性调节,并且这些机制的放松管制被认为在病理或适应不良的情况下发挥作用,例如精神分裂症,注意力缺陷和过度活跃的精神疾病(多动症),抑郁症(Dunlop和Nemeroff,2007; Masana等,2011),或滥用药物和赌博行为(Everitt和Robbins,2005)。 有趣的是,mPFC中释放的多巴胺的活性可能受去甲肾上腺素(NA)的影响,去甲肾上腺素在mPFC中的含量高于多巴胺,尤其是NA转运蛋白(NET),它不仅比NA转运蛋白更丰富。 mPFC中的多巴胺转运蛋白(DAT)Carboni等,1990, 2006; Gresch等,1995; Westernik等,1998),但也显示出对多巴胺的亲和力甚至高于NA(Horn,1973)。 由于多巴胺和NA通常在许多mPFC功能中起作用,从工作记忆和注意力集的形成和转变为逆转学习,反应抑制和对压力的反应(见 Robbins和Arnsten,2009),这提高了mPFC中单独或与NA一起使用的多巴胺也可能在RHA和RLA大鼠(包括性行为中发现的那些)之间的行为差异中发挥作用的可能性。
为了检验该假设,通过脑内微透析在两条RHA和RLA大鼠系中研究了中皮质多巴胺能系统和去甲肾上腺素能系统在mPFC中的活性。 简而言之,多巴胺(及其主要代谢产物DOPAC)和NA在透析液中测量,所述透析液得自性行为(例如,从未暴露于接受性女性)和性经历的RHA的mPFC的前肢(PrL)和下肢(IL)区室。 RLA大鼠(例如,其经历五次初步交配测试并且显示出恒定的交配活性水平)当存在难以接近的接受性雌性时和通过高压液相色谱与电化学检测(HPLC-ECD)的直接性相互作用期间。
材料和方法
动物
远交RHA和RLA雄性大鼠(N =每行的30,在实验开始时称重≈300g)均来自意大利卡利亚里大学1998成立的殖民地(Giorgi等,2007)。 用于选择性繁殖撒丁岛殖民地的程序已经详细描述(Giorgi等,2005).
在所有实验中使用的去卵巢刺激SD雌性大鼠(在实验开始时的250-300 g)获自Envigo(San Pietro al Natisone,Italy)。 在实验开始前至少38天,在60°C,湿度下,将每笼4只(20 cm×10 cm×24 cm)的动物适应卡利亚里大学生物医学科学系的住房设施。 60%,逆转12 h亮/暗循环(从08点亮:00 h到20:00 h),水和标准实验室食品 随意。 在整个适应期间每天处理动物的1-2 min,以限制实验期间的操作应力; 此外,与动物屋维修人员的接触仅限于一名服务员,家庭笼中的床上用品在前一天或实验当天从未改变过。 所有实验均在10:00 h和18:00 h之间进行。 该研究是根据欧洲共同体指南,9月22指令,2010(2010 / 63 / EU)和意大利立法(DL March 4,2014,n.26)的建议进行的。 该方案经卡利亚里大学动物实验伦理委员会批准(授权号361 / 2016-PR,4月08,2016至FS)。
实验组
使用性初始和性经历的雄性RHA和RLA大鼠。 性幼稚的大鼠是从未接触过性接受性卵巢切除和雌二醇+黄体酮引发的雌性大鼠; 有性生活的大鼠是已经接受了60 min连续5次交配测试的大鼠,每天3天接受雌性(Sanna等,2014a,b)。 通过分别在交配试验前用皮下雌二醇苯甲酸盐(花生油中的200μg/大鼠)和黄体酮(花生油中的0.5 mg /大鼠),48 h和6 h处理雌性动物。 在实验前,通过May-Grunwald-Giemsa染色和阴道涂片1 h的显微镜检查验证了发情。 与先前的研究一致(Sanna等,2014a,b, 2015)发现五个初步交配试验足以使两条线的雄性罗马大鼠显示出恒定的交配活性水平:例如,性经验的RHA和RLA大鼠满足在最后两个试验中的每一个中实现的至少一次射精的标准。 (一个RHA大鼠和两个不满足该标准的RLA大鼠在该阶段被丢弃)。 在这些初步交配试验两天后,性经验丰富的罗马大鼠进行立体定向手术,在mPFC中植入微透析探针,如下所述(Sanna等,2015).
性行为期间mPFC的微透析
在微透析前一天,将性初始或经历过的RHA和RLA大鼠置于异氟烷麻醉下的立体定位装置(Stoelting Co.,Wood Dale,IL,USA)中(1.5%-2%; Harvard Apparatus,Holliston,MA,USA)。并植入垂直的自制微透析探针(透析膜≈3mm的自由表面; Melis等,2003),单侧指导mPFC,PrL和IL区室(坐标:3.0 mm前部和前囟侧部0.7 mm,5.5 mm腹侧至硬脑膜; Paxinos和Watson,2004)。 在实验当天,将动物在周期的黑暗阶段转移到交配笼(45 cm×30 cm×24 cm),该笼子位于隔音红灯照亮的隔音室内并且包含在另一个内部。小型有机玻璃保持架(15 cm×15 cm×15 cm),垂直壁上有25孔(Ø2mm),可实现视觉,嗅觉和声学,但不能直接相互作用。 在2 h的适应期后,将微透析探针与聚乙烯管连接至CMA / 100微量输注泵(Harvard Apparatus,Holliston,MA,USA),并用林格氏溶液(147 mM NaCl,3 mM KCl和1.2 mM)灌注。氯化钙2,pH 6.5),流速为2.5μl/ min。 在灌注培养基的2小时与细胞外液的平衡期后,在实验期间在冰冷的聚乙烯管中每37.5 min收集透析液等分的15μL,用于测量多巴胺,DOPAC和NA的浓度,如所述下面。 在收集至少四个透析液等分试样后,将接受雌性大鼠引入位于交配笼内部的小笼中30 min。 在这些30 min期间,收集其他两个透析液等分试样。 在这些情况下,雄性大鼠不能直接与雌性相互作用,但表现出非接触性勃起(见下文)。 在此期间后,取出小笼子,允许交配75 min,并收集其他五个透析液等分试样。 在此期间结束时,将雌性从交配笼中取出,并收集另外的透析液等分试样(Pfaus和Everitt,1995; Melis等,2003; Sanna等,2015)。 在整个实验中记录与性行为的预期和完成阶段相关的性参数(见下文)。
性行为
在整个实验过程中,观察者记录了与性行为的预期和完成阶段相关的性动机和交配性能的几个参数,这些参数不了解具体的实验条件,例如,谁不知道线条和性经验的水平在该实验中使用的动物(见下文)。 简而言之,第一次非接触性勃起的延迟(NCPEL,从引入内部小笼子中的接受性雌性的时间)和它们的频率(NCPEF,在女性期间发生的非接触性阴茎勃起的数量)记录了存在于内笼中的情况。 这些信息素介导的阴茎勃起,发生在性能强大的雄性大鼠中,在难以接近的接受性雌性的情况下,被认为是性唤起的重要指标(Sachs等,1994; 萨克斯,2000; Melis等,2003)。 当允许性交互时,例如,在交配期间,安装和插入的潜伏期(ML和IL,分别从移除内部小笼到第一次安装或第一次插入的时间); 安装和插入的频率(MF和IF,分别在第一系列交配活动和整个交配期间的安装和插入次数); 射精潜伏期(EL,从第一次射入第一次射精到射精的时间); 记录射精频率(EF,交配测试期间的射精总数)和射精后间隔(PEI,从第一次射精到下一次插入的时间)。 此外,交配效率(CE;给定系列的插入次数除以同一系列中的坐位和插入次数的总和)和插入间隔(III;给定系列的射精潜伏期之间的比率)并计算了第一系列交配活动(该系列中的插入数量)(Sachs和Barfield,1976; Meisel和Sachs,1994; Melis等,2003; Sanna等,2014a,b, 2015).
从mPFC中测定透析液中的多巴胺,DOPAC和去甲肾上腺素浓度
多巴胺,DOPAC和NA浓度在来自mPFC的相同透析液等分试样的20μL中通过高压液相色谱(HPLC)测量,使用4011双电池(Coulochem II,ESA,Cambridge,MA,USA)进行电化学检测。已经描述过(Melis等,2003)。 以还原模式在+ 350和-180 mV下进行检测。 HPLC配备有Supelcosil C18柱(7.5 cm×3.0 mm id,3μm粒径; Supelco,Supelchem,Milan,Italy),用0.06 M柠檬酸盐/乙酸盐pH 4.2洗脱,含有甲醇20%v / v,0.1 mM EDTA,1μM三乙胺和0.03 mM十二烷基硫酸钠作为流动相,流速为0.6 mL / min和室温。 该测定的灵敏度为多巴胺的0.125 pg,DOPAC的0.1 pg和NA的0.2 pg。
组织学
在实验结束时,通过断头处死大鼠,立即将脑从颅骨中取出并浸入4%甲醛水溶液中以进行12-15天。 在此期间后,用冷冻切片机制备40μm冠状脑切片,用中性红染色并在相差显微镜上检查。 然后通过一系列脑切片跟踪探头管道,将探头尖端的位置定位在mPFC中(见图) 1)。 只有发现透析膜的活性部分正确定位在mPFC的PrL和IL区室中的大鼠才被认为用于结果的统计学评估(来自每个实验组的一只大鼠在该阶段被丢弃)。
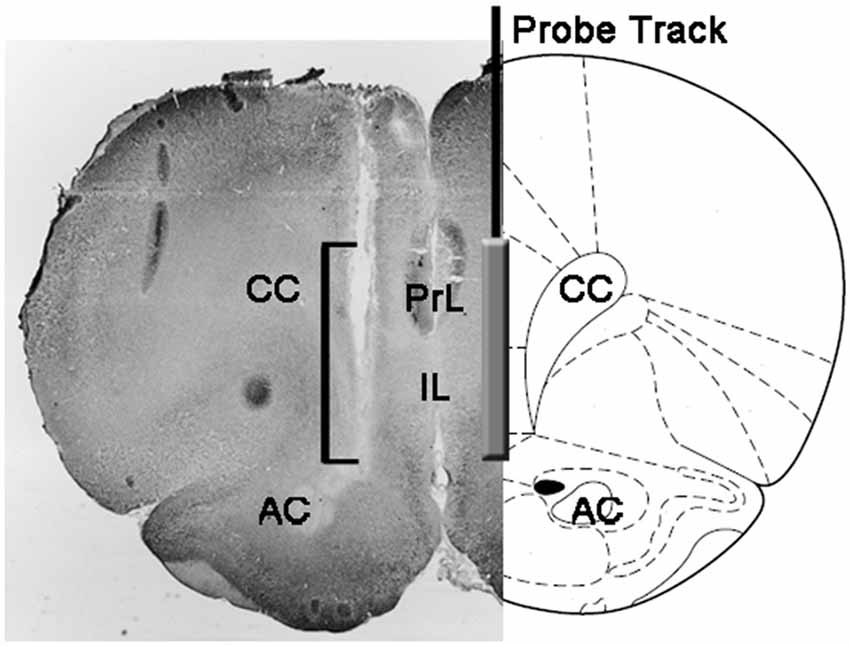 图1。 大鼠脑冠状切面的示意图,显示微透析探针在内侧前额叶皮质的PrL和IL部分的轨迹(mPFC; Paxinos和Watson,2004)。 微照片中的方括号表示中性红染色部分的部分,显示微透析探针的透析膜的活性部分。 缩写:PrL,prelimbic area; IL,infralimbic area; AC,前连合; CC,胼call体。
图1。 大鼠脑冠状切面的示意图,显示微透析探针在内侧前额叶皮质的PrL和IL部分的轨迹(mPFC; Paxinos和Watson,2004)。 微照片中的方括号表示中性红染色部分的部分,显示微透析探针的透析膜的活性部分。 缩写:PrL,prelimbic area; IL,infralimbic area; AC,前连合; CC,胼call体。
统计报表
进行了生化(多巴胺,DOPAC和NA)和行为(NCPEL和NCPEF,ML,IL和EL,MF,IF和EF,以及PEI)数据的统计分析,包括所有实验对象(传统上在雄性大鼠的研究中进行)交配行为)或排除那些在微透析实验期间不与射精交配的受试者。 在第一种情况下,当所有动物都被包括在分析中时,在女性无法接近或没有安装或插入或射精与可用女性的时间内未显示NCPE的动物被分配相应的全范围分数:1800 s if男性没有在女性无法进入的30分钟中显示NCPE; 900用于ML和IL,如果男性没有安装或插入可用的女性; 1800用于EL,如果男性没有达到射精,600用于PEI,如果男性在第一次射精后没有插入。 在第二种情况下,那些不与射精交配的动物被排除在分析之外。 这样做是为了评估性行为的差异是否与性活动的两个阶段中罗马鼠系mPFC中细胞外多巴胺,DOPAC和NA浓度的差异相关,排除了由于包含的可能造成的混乱干扰通过为分析的性参数指定固定值,未显示性行为的受试者。
使用ANOVA进行若干统计分析,以便检测并更好地表征所用实验条件之间可能的差异。 在进行ANOVA之前,检查每个不同实验变量的数据集的四个实验组之间的差异的均匀性(对所有动物或仅与射精交配的大鼠),根据情况进行Bartlett's或Levine's检验。 当发现方差的显着差异时,将数据集进行对数变换(即,将实验Y值改变为Log Y值),重新检查方差的均匀性,然后通过ANOVA分析。 简而言之,在幼稚和性经历的雄性RHA和RLA大鼠之前(例如,与难以接近的雌性)和第一系列交配活动(从第一次射入/插入到第一次射精后的第一次进入/插入)期间获得的行为参数在微透析期间,通过双向ANOVA分析,通过使用大鼠系和性经验水平作为受试者因素之间(行为参数的平均值在表中报告) 1 和 F 表中的ANOVA分析的值和显着性水平 2).
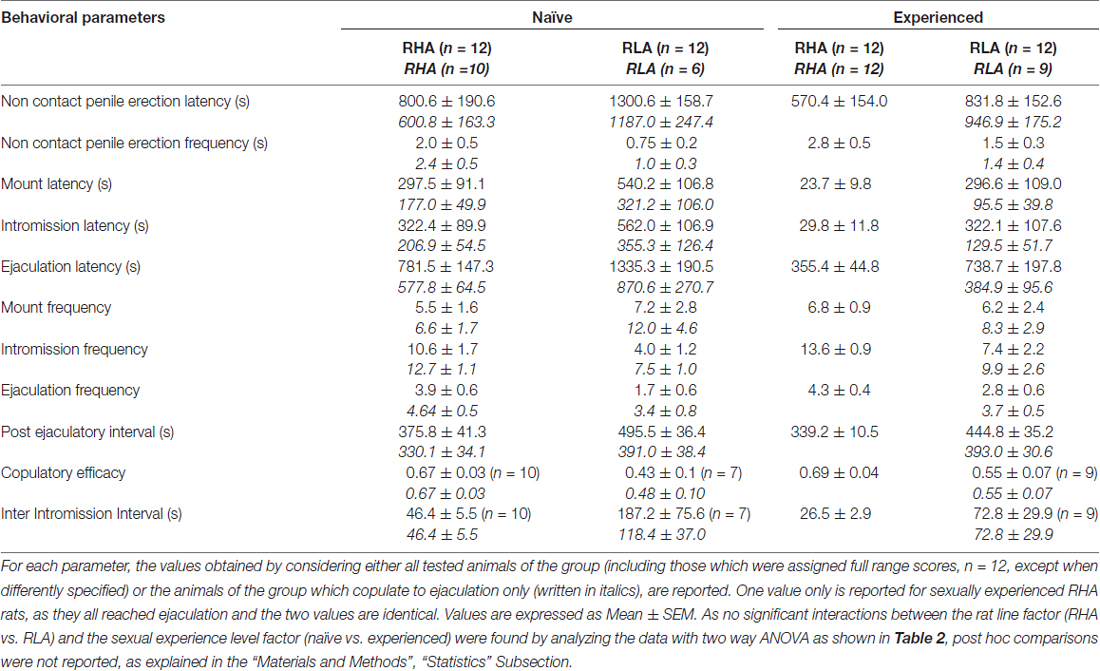 表1。 在第一系列交配活动中测量的交配参数(从女性可接近男性开始的第一次安装/插入开始,到第二次系列的第一次安装/介入后的射精后间隔结束后结束)和非接触式阴茎性生活和经验丰富的RHA和RLA大鼠的勃起。
表1。 在第一系列交配活动中测量的交配参数(从女性可接近男性开始的第一次安装/插入开始,到第二次系列的第一次安装/介入后的射精后间隔结束后结束)和非接触式阴茎性生活和经验丰富的RHA和RLA大鼠的勃起。
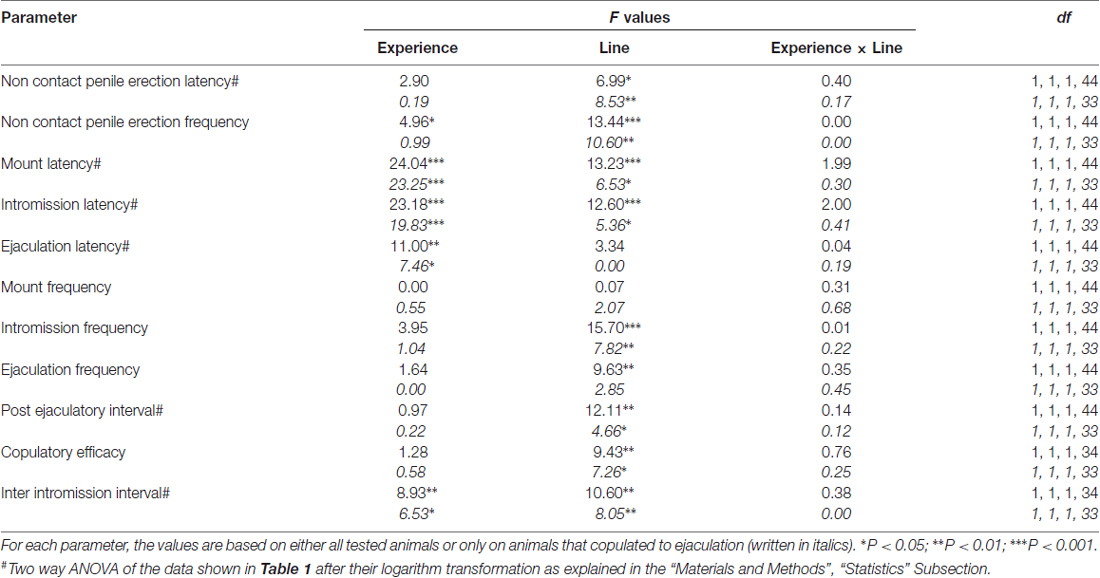 表2. F 对表中报告的数据进行双向ANOVA的值和显着性水平 1 通过使用大鼠系(RHA vs.RLA)和性经验水平(幼稚与经验)作为受试者因素之间。
表2. F 对表中报告的数据进行双向ANOVA的值和显着性水平 1 通过使用大鼠系(RHA vs.RLA)和性经验水平(幼稚与经验)作为受试者因素之间。
此外,通过首先计算通过绘制多巴胺,DOPAC或NA的浓度值或非接触性勃起,坐骑,插入和射精的数量获得的AUC,进行微透析期间从每只大鼠获得的数据的总体分析。时间(在交配笼中引入雄性大鼠至实验结束后开始1 h -180 min除以神经化学值的15 min分数 - 或在雌性被引入交配笼时的时间对于行为参数),然后通过使用大鼠线和性经验水平作为受试者因素之间的双向ANOVA比较计算值(神经化学和行为参数的AUC的平均值在表中报告) 4 和 F 表中的ANOVA分析的值和显着性水平 5)。 最后,通过因子分析对每组数据进行更详细的点对点分析(即多巴胺浓度,DOPAC,NA和非接触性勃起,坐骑,插入和射精与时间的关系)。重复测量的方差分析,通过使用大鼠线和主体因素和时间之间的性经验水平(即透析液分数)作为受试者因素( F 表中报告了这些ANOVA分析的值和显着性水平 6)。 进行类似的分析也只考虑神经化学参数的基础值(在接受雌性的交配笼中引入之前收集的最后四个透析液等分试样),其平均值在表中报告 3。 首先,当执行重复测量的一般因子ANOVA时,发现不是二阶相互作用,特别是神经化学参数的线×时间和经验×时间相互作用(见表 6)通过直接比较性初始或经验丰富的RHA与RLA大鼠,或性经验对比,对这些数据集进行双向ANOVA,其中线性或性经验水平在受试者因素和时间之间作为主体因素。幼稚的RHA大鼠或性经验与幼稚的RLA大鼠。 结果 事后 通过使用Tukey的HSD测试对由这些双向ANOVA揭示的显着相互作用进行的成对比较在图中报告 2。 在所有其他情况下, 事后 没有报告比较,因为方差分析未能揭示大鼠系,性经验水平和时间之间的任何显着相互作用。 统计分析均使用Graph Pad 5(PRISM,San Diego,CA,USA)和STATISTICA 12(Statsoft,Tulsa,OK,USA)进行,显着性水平设定为 P <0.05。
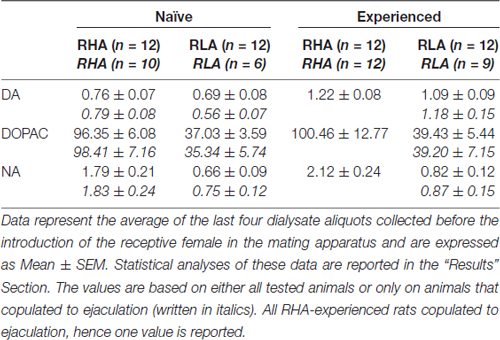 表3。 有性幼稚和经验丰富的RHA和RLA大鼠的内侧前额叶皮质(mPFC)的透析液中的基础多巴胺(DA),3,4-二羟基苯乙酸(DOPAC)和去甲肾上腺素(NA)浓度(nM)。
表3。 有性幼稚和经验丰富的RHA和RLA大鼠的内侧前额叶皮质(mPFC)的透析液中的基础多巴胺(DA),3,4-二羟基苯乙酸(DOPAC)和去甲肾上腺素(NA)浓度(nM)。
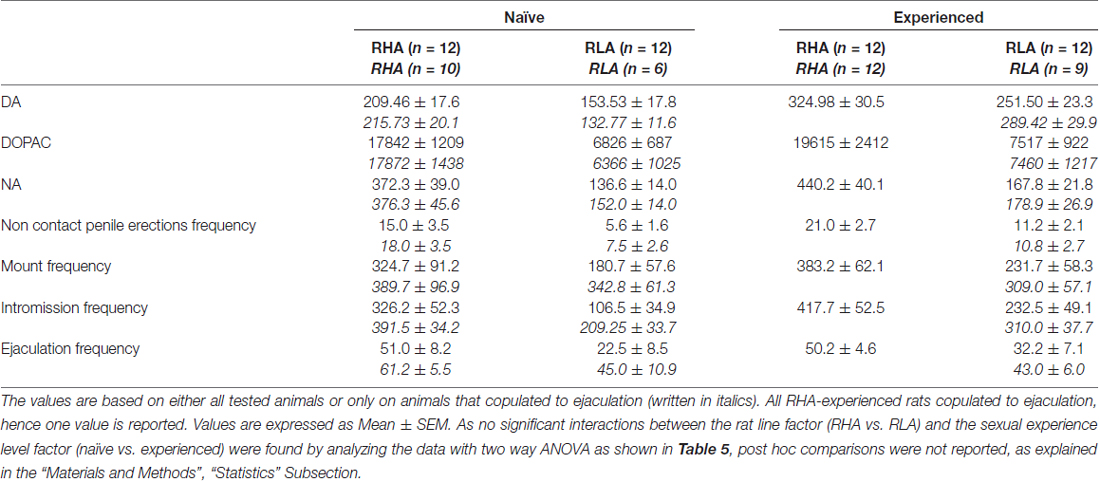
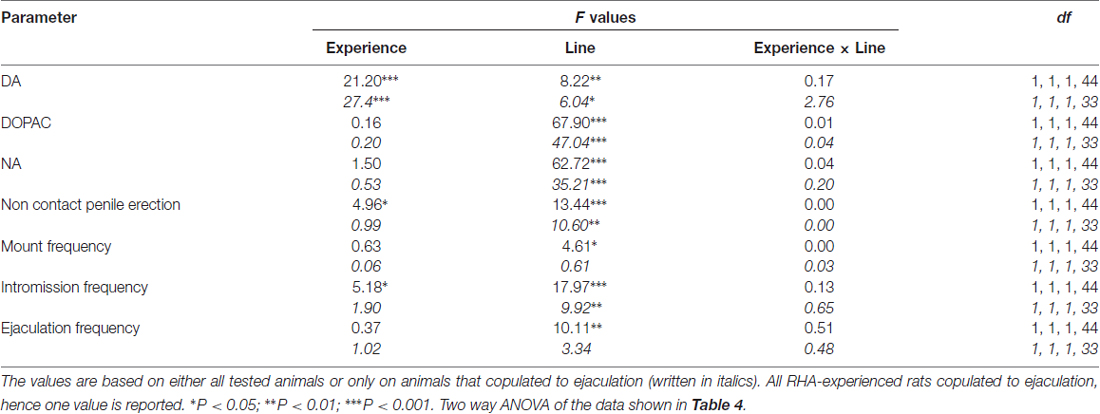 表5. F 对表中报告的数据进行双向ANOVA的值和显着性水平 4 通过使用大鼠系(RHA vs.RLA)和性经验水平(幼稚与经验)作为受试者因素之间。
表5. F 对表中报告的数据进行双向ANOVA的值和显着性水平 4 通过使用大鼠系(RHA vs.RLA)和性经验水平(幼稚与经验)作为受试者因素之间。
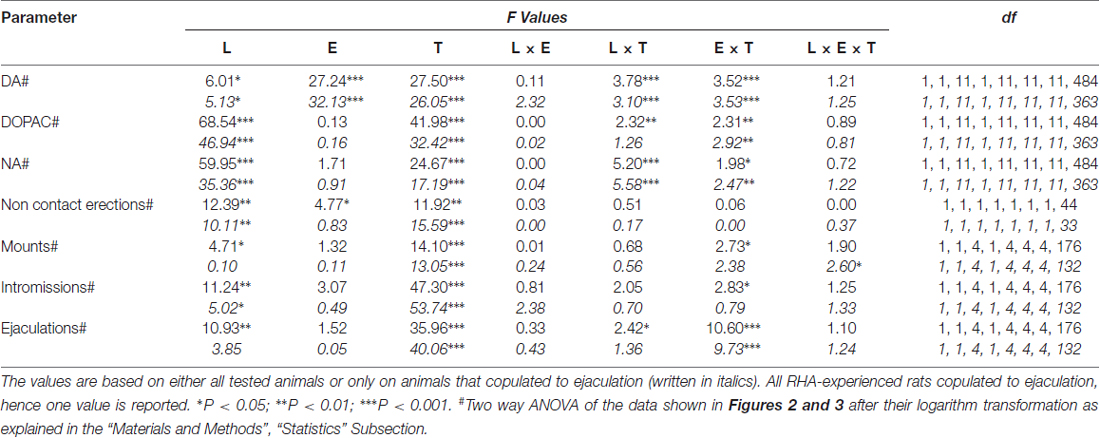 表6. F 对图中所示结果进行重复测量的一般因子ANOV的值和显着性水平 2 和 3 通过使用大鼠系(L; RHA vs.RLA)和性经验水平(E)(幼稚与经历)作为受试者因子和时间(T)(透析液分数)之间的受试者因子。
表6. F 对图中所示结果进行重复测量的一般因子ANOV的值和显着性水平 2 和 3 通过使用大鼠系(L; RHA vs.RLA)和性经验水平(E)(幼稚与经历)作为受试者因子和时间(T)(透析液分数)之间的受试者因子。
 图2。 多巴胺(DA),DOPAC和去甲肾上腺素(NA)浓度在mPFC透析液中从性天真 (高手) 和经验丰富的RHA和RLA雄性大鼠 (B,d,F) 在与性接触的女性进行性活动时。 对于使用立体定位植入mPFC的微透析探针的两个品系的性行为(即,从未暴露于性接受性女性之前)和性经历(其经历五次交配试验)被单独放入交配笼中。 如“材料和方法”部分所述,用透析缓冲液灌注微透析探针。 在收集四个透析液等分试样以确定基础值之后,然后将难以接近的接受雌性放置在配合装置的小笼内(时间= 0,不能使用)。 在30分钟后,移除小笼并允许交配75分钟(交配),之后从交配笼中取出雌性(fe.remov)。 在实验期间,测量非接触勃起和交配参数,并且每15 min收集透析液等分试样并如“材料和方法”部分中所述分析多巴胺,DOPAC和NA。 值是每组所有12大鼠获得的值的平均值±SEM(RHA_all =实线绿线,RLA_all =实心棕色线)或仅与射精交配的值(RHA_cop =红色虚线; RLA_cop =蓝色虚线)。 在经验丰富的RHA大鼠中,所有动物均达到交配并且值相同。 *P 相对于该组的基准值(无女性)<0.05(RHA_all为绿色; RHA_cop为红色,RLA_all为棕色,RLA_cop为蓝色); #P 相对于RLA组的相应值(红色,RHA_all与RLA_all;绿色,RHA_cop与RLA_cop),<0.05; §P 相对于有性行为的大鼠的时间匹配值<0.05(绿色,有经验的RHA_all vs.天真RHA_all;红色,有经验的RHA_cop与天真RHA_cop;棕色,有经验的RLA_all与天真RLA_all;蓝色,有经验的RLA_cop与天真RLA_all; RLA_cop)(对数转换后,对显示的数据进行双向方差分析,如“材料和方法”,“统计”小节中所述,然后进行Tukey的HSD测试)。
图2。 多巴胺(DA),DOPAC和去甲肾上腺素(NA)浓度在mPFC透析液中从性天真 (高手) 和经验丰富的RHA和RLA雄性大鼠 (B,d,F) 在与性接触的女性进行性活动时。 对于使用立体定位植入mPFC的微透析探针的两个品系的性行为(即,从未暴露于性接受性女性之前)和性经历(其经历五次交配试验)被单独放入交配笼中。 如“材料和方法”部分所述,用透析缓冲液灌注微透析探针。 在收集四个透析液等分试样以确定基础值之后,然后将难以接近的接受雌性放置在配合装置的小笼内(时间= 0,不能使用)。 在30分钟后,移除小笼并允许交配75分钟(交配),之后从交配笼中取出雌性(fe.remov)。 在实验期间,测量非接触勃起和交配参数,并且每15 min收集透析液等分试样并如“材料和方法”部分中所述分析多巴胺,DOPAC和NA。 值是每组所有12大鼠获得的值的平均值±SEM(RHA_all =实线绿线,RLA_all =实心棕色线)或仅与射精交配的值(RHA_cop =红色虚线; RLA_cop =蓝色虚线)。 在经验丰富的RHA大鼠中,所有动物均达到交配并且值相同。 *P 相对于该组的基准值(无女性)<0.05(RHA_all为绿色; RHA_cop为红色,RLA_all为棕色,RLA_cop为蓝色); #P 相对于RLA组的相应值(红色,RHA_all与RLA_all;绿色,RHA_cop与RLA_cop),<0.05; §P 相对于有性行为的大鼠的时间匹配值<0.05(绿色,有经验的RHA_all vs.天真RHA_all;红色,有经验的RHA_cop与天真RHA_cop;棕色,有经验的RLA_all与天真RLA_all;蓝色,有经验的RLA_cop与天真RLA_all; RLA_cop)(对数转换后,对显示的数据进行双向方差分析,如“材料和方法”,“统计”小节中所述,然后进行Tukey的HSD测试)。
成果
RHA和RLA大鼠显示不同数量的非接触性勃起和不同的交配行为模式
与早期研究一致(Sanna等,2014a,b, 2015),在通过脑内微透析从mPFC收集透析液等分试样期间,当与接受性雌性放在一起时,不同数量的性初始雄性RHA和RLA大鼠开始参与性活动。 简而言之,在该研究中,10中的12性初始雄性RHA大鼠(83%)在微透析实验期间在第一次交配试验中与6幼稚RLA大鼠中的12(50%)相互作用。 在性经历的雄性RHA和RLA大鼠中也发现了这种差异(在五次交配试验后),尽管是减毒的,所有12 RHA大鼠在微透析期间从9 RLA大鼠中射出针对12的射精。 始终与先前的研究一致,同样在该研究中,当暴露于难以接近的接受性雌性以及性交互过程中的不同交配行为模式时,两个罗马大鼠品系表现出不同的潜伏期和非接触性阴茎勃起的频率。 这些差异是在考虑给定组的所有实验动物的数据时发现的,无论它们是否与射精有关(例如,通过为那些没有表现出行为的动物分配完整分数,即没有交配的大鼠)射精)或考虑确实只显示给定组行为的动物数据(例如,与射精相关的动物;表 1)。 因此,通过双向ANOVA对第一系列交配活性中测量的非接触性勃起(NCPE)值和交配参数的统计分析显示,在考虑所有实验大鼠或仅交配大鼠时,两种大鼠系之间存在显着差异。在四个实验组中(见表 2)。 事实上,当考虑从所有实验动物获得的数据时,与RLA大鼠相比,RHA大鼠中非接触勃起的数量更高并且NCPEL,ML,IL和PEI显着更短。 此外,与RLA大鼠相比,在RHA大鼠中EF和CE更高,而III显着更短。 两次罗马鼠系之间的一些上述差异倾向于通过反复交配试验减少或消失。 尽管如此,在通过反复的性经验稳定性行为之后,仍然存在一些这些差异。 当仅考虑与射精相关的四个实验组的动物的值时,获得了类似的结果,除了EF(参见表格) 1, 2).
来自性初生和经验丰富的RHA和RLA大鼠的mPFC透析液中细胞外多巴胺,DOPAC和去甲肾上腺素的基础浓度
在目前的实验条件下,从mPFC获得的透析液中的多巴胺,DOPAC和NA的量(例如,无论它们是否与射精有关)性生幼稚的RHA和RLA大鼠,是多巴胺的2.32 pg和2.11 pg, 322.56 pg和124.42 pg用于DOPAC和6.05 pg和2.24 pg用于NA,分别在20μl透析液中。 在从有经验的RHA和RLA大鼠的mPFC获得的透析液中测量相似的量(多巴胺:3.73 pg和3.33 pg,DOPAC:353.62 pg和133.32 pg,NA:7.17 pg和2.78 pg分别在RHA和RLA大鼠中) 。 这些值分别表示细胞外多巴胺和DOPAC的≅0.8-1.2 nM和≅95-100 nM浓度,以及RHA大鼠mPFC中细胞外NA的≅1.8-2.2 nM浓度和浓度≅0.8-1.2 nM和≅35-40 nM分别用于细胞外多巴胺和DOPAC,而≅0.6-0.9 nM用于细胞外NA,用于RLA大鼠的mPFC(表 3)。 在透析缓冲液与mPFC细胞外液的2 h平衡期后获得上述值。 随着真实多巴胺的回收,透析探针的DOPAC和NA估计接近20%,在多巴胺,≅4的两个细胞系中,细胞外多巴胺,DOPAC和NA浓度可估计接近≅5-500 nM。并且≅200nM用于DOPAC和≅10和≅4nM分别用于RHA和RLA大鼠的mPFC中的NA。 在对数转换后对上述值进行重复测量的因子ANOVA揭示了性初始和性经历大鼠之间多巴胺基础水平的显着差异(F(1,44,132) = 26.05, P <0.001),以及在DOPAC的基础水平(F(1,44,132) = 63.36, P <0.001)和NA(F(1,44,132) = 42.14, P 在RHA和RLA大鼠之间(<0.001)(在将雌性引入小笼子之前收集了最后四个样品)。 当仅考虑在微透析实验中交配射精的大鼠时,发现相似的值(表 3)。 同样在这种情况下,对数变换后对数值进行重复测量的因子方差分析显示性性幼稚和性经验大鼠之间多巴胺的基础水平存在显着差异(F(1,33,99) = 26.97, P <0.001),以及在DOPAC的基础水平(F(1,33,99) = 42.95, P <0.001)和NA(F(1,33,99) = 27.63, P (<0.001)。
性活性和经验丰富的RHA和RLA大鼠mPFC透析液中细胞外多巴胺,DOPAC和去甲肾上腺素的浓度在性活动中发生差异
内部小笼中难以接近的接受性雌性的存在以及随后的直接性相互作用增加了从性初始和经历的雄性RHA和RLA大鼠获得的mPFC透析液中细胞外多巴胺,DOPAC和NA的浓度,尽管两者之间存在显着差异。两个大鼠系和经验条件水平,要么考虑所有实验动物,要么只考虑那些与射精相关的动物(图 2)。 事实上,多巴胺,DOPAC和NA浓度的AUC双向ANOVA显示,与RLA大鼠相比,RHA大鼠的整个试验中多巴胺,DOPAC和NA的总含量显着更高,并且在性方面与幼稚大鼠相比有经验(见表 4 对于AUCs值和表 5 F 价值观和意义水平)。 这些差异通过点对点因子ANOVA进一步证实,用于重复测量多巴胺,DOPAC和NA实验中两种性初始和经历过的大鼠行的NA量,其显示出线,经验,时间和显着线×时间的显着主要影响。和经验×时间一阶交互(见表 6 F 价值观和意义水平)。
从性生活的RHA和RLA大鼠获得的mPFC透析液中细胞外多巴胺,DOPAC和去甲肾上腺素的浓度在性活动期间发生差异变化
在性初始的RHA和RLA大鼠中,细胞外多巴胺(但不是DOPAC)在存在难以接近的雌性时首先增加,而NA仅在RHA大鼠中增加。 在幼稚的RHA大鼠中,细胞外多巴胺在DOPAC和NA发现的性交互作用的第一个15 min中进一步增加。 细胞外多巴胺,DOPAC和NA的增量持续所有交配时间,多巴胺,DOPAC和NA分别在60 min,75 min和75 min达到峰值,然后是细胞外多巴胺,并且在较小程度上达到DOPAC和NA ,去除雌性后,降至与基础类似的值。
另一方面,在交配的第一个15分钟期间,幼稚RLA大鼠中出现细胞外多巴胺,DOPAC和NA的第一次显着增加和峰值。 然而,与RHA大鼠不同,在RLA大鼠中,细胞外多巴胺,DOPAC和NA倾向于恢复到与女性直接相互作用的第一个30 min内与基础相似的值(参见图 2A,C,E).
最后,在交配期间,细胞外多巴胺的增量在性初始RHA中比RLA大鼠更大且更持久(见图 2A而在没有雌性的情况下,它们的浓度在两条大鼠品系中相似,尽管不相同(见上文和表格) 3)。 有趣的是,虽然在DOPAC增加百分比和两个罗马品系之间没有发现差异,但在接受雌性存在期间,NA浓度在较小程度上没有差异,DOPAC和NA浓度的绝对值差异超过两倍在基础条件下观察到两条大鼠系之间的差异(参见表 3并且在整个测试过程中接受女性(见图 2C,E).
从性经验的RHA和RLA大鼠获得的mPFC透析液中细胞外多巴胺,DOPAC和去甲肾上腺素的浓度在性活动期间发生差异变化
如在性初始的RHA和RLA大鼠中发现的,不可接近的雌性和随后的直接性相互作用的存在导致从性经历的雄性RHA和RLA大鼠获得的mPFC透析液中细胞外多巴胺和DOPAC的增加。 然而,在这种情况下,也发现了大鼠系相关的差异,或者在考虑所有实验动物时,或者在较小程度上,只考虑那些与射精相关的动物(图 2)。 在性经验丰富的RHA和RLA大鼠中,细胞外多巴胺的第一次增加发生在难以接近的女性身上。 此后,在两种大鼠的交配期间多巴胺浓度增加。 交配后45-60 min后多巴胺浓度达到峰值,并在交配试验结束时缓慢降低至基础值(见图) 2B)。 与性初始大鼠相似,性交经验的RHA大鼠的细胞外多巴胺浓度高于RLA大鼠,主要是在交配阶段的中心部分(交配的30-45分钟后)收集的透析液的等分试样中。 仅在RHA大鼠中存在难以接近的雌性期间DOPAC浓度增加,而在交配期间两种大鼠品系中DOPAC浓度增加至相似程度(75-90 min处的峰值),并且在结束时倾向于恢复至基础值。交配试验(见图 2D)。 在RHA大鼠的90分钟和RN大鼠的45分钟引入雌性达到峰值后,两种性经验大鼠的NA浓度也增加(参见图 2F)。 与多巴胺(参见上文)不同,当考虑所有动物与仅到达射精的动物时,在来自两个品系的性经历的大鼠中DOPAC和NA浓度的增加中发现了类似的差异。 然而,正如在性行为幼稚大鼠中发现的那样,虽然DOPAC和NA浓度的增加百分比相似,但在整个测试过程中(即,之前,期间和之后),在性经历的RHA和RLA大鼠之间发现DOPAC和NA绝对值的显着差异。接受雌性的存在,RHA大鼠显示出比RLA大鼠更高的值(见表 3 对于基础值和数字 2D,F 对于整个测试的值)。
性经验影响RHA和RLA大鼠性活动时mPFC透析液中多巴胺和去甲肾上腺素浓度的变化
性行为和经验丰富的RHA和RLA大鼠的细胞外多巴胺和NA浓度的比较表明,在考虑所有实验动物时,性经验改变了两种大鼠细胞外多巴胺的浓度,并且在较小程度上改变了NA的浓度。或仅与射精相关的那些(图 2A,B,E,F)。 因此,两种品系的性初始和经历过的大鼠之间多巴胺的基础值出现显着差异(见表1) 3与性生活的RHA和RLA大鼠相比,交配活动期间的多巴胺值通常高于其幼稚的同龄人(参见图 2B)。 此外,在经历过性行为的RHA大鼠中,尽管在难以接近的雌性存在期间发现的多巴胺增加遵循在性幼稚大鼠中看到的相同的时间模式,但在交配期间发现时间模式的差异。 实际上,在这种情况下,与性初始RHA大鼠(在60 min处的一个主峰)中观察到的不同,发现两个主要峰值,第一个在15分钟之后,第二个在45分钟交配之后。 在最后一次增加之后,多巴胺值倾向于恢复到基础值(见图 2B).
在性初始与经验的RLA大鼠中发现了相似的图片。 事实上,与在RHA大鼠中发现的相似,在经验丰富的RLA大鼠中也发现了以细胞外多巴胺的三个主要增加为特征的时间模式:第一个具有不可接近的雌性和另外两个在交配期间,即在15 min和60之后。交配的最小值(见图 2A,B)。 一般来说,性经验似乎会在从性初始到经验状态的两种大鼠行中多巴胺释放的时间模式中产生更持久的变化。
如图所示 2C-F在两种幼稚和有经验的大鼠之间,DOPAC和NA浓度仅检测到微小差异。 然而,在基础中观察到细胞外NA与未经治疗的大鼠相比在经历更高值时的显着趋势(在引入接受性雌性之前,参见表 3和AUCs值所揭示的总NA数量(见表 4计算从整个测试获得的细胞外浓度(在接受雌性存在之前和期间)。 最后,当将两只大鼠系的经验大鼠与其幼稚对应物进行比较时,NA浓度的时间模式非常相似,表明性经验可导致细胞外NA的普遍增加,而不是其增加模式的变化(见图 2E,F).
从性生活和经验丰富的RHA和RLA大鼠中获得的mPFC透析液中细胞外多巴胺和去甲肾上腺素浓度的变化与性行为的变化同时发生
在试验期间,在性生活和经历过的RHA和RLA大鼠中发现的mPFC透析液中细胞外多巴胺,DOPAC和NA浓度的差异(图 2)与实验的两个主要阶段(即女性难以接近且分别可用于男性)测量的不同性参数的修改一起发生,其中包括非接触性阴茎勃起(图中) 3A,B),坐骑(数字 3C,d),插入(图 3E,F)和射精(图 3G,H)。 在考虑所有实验动物或仅与射精相关的动物时,发现了这些差异。 事实上,通过双向ANOVA对这些参数的AUC值进行了初步分析(见表 4 对于AUCs值和表 5 F 值和显着性水平)随后通过对重复测量的因子ANOVA对实验中这些参数的值进行点对点分析,揭示了线,经验,时间,显着的一阶线×时间和经验×时间相互作用的显着主效应,和坐骑的重要二阶互动(见表 6 F 价值观和意义水平)。 因此,符合以前的研究(Sanna等,2015):( i)性行为幼稚的RHA大鼠表现出比其RLA对应物更多的非接触性阴茎勃起,坐位,插入和射精; (ii)经验丰富的RHA大鼠表现出比其RLA对应物更多的非接触性阴茎勃起,插入和射精; (iii)两只品系的性经历的大鼠,特别是RLA大鼠,与性交天真的对应物相比,显示出更多的非接触性阴茎勃起,插入和射精。 事实上,在RHA大鼠中,性经验产生的大部分变化发生在时间过程而不是行为参数的频率(例如,经验丰富的大鼠主要在测试的前半部分交配,而幼稚的大鼠表现出来在整个试验过程中,类似的交配趋势),在RLA大鼠中,通过性经验产生性行为的更显着的总体改善,主要表现在性行为上,正如经验丰富的RLA大鼠的交配参数值的变化所揭示的那样。天真的同行。
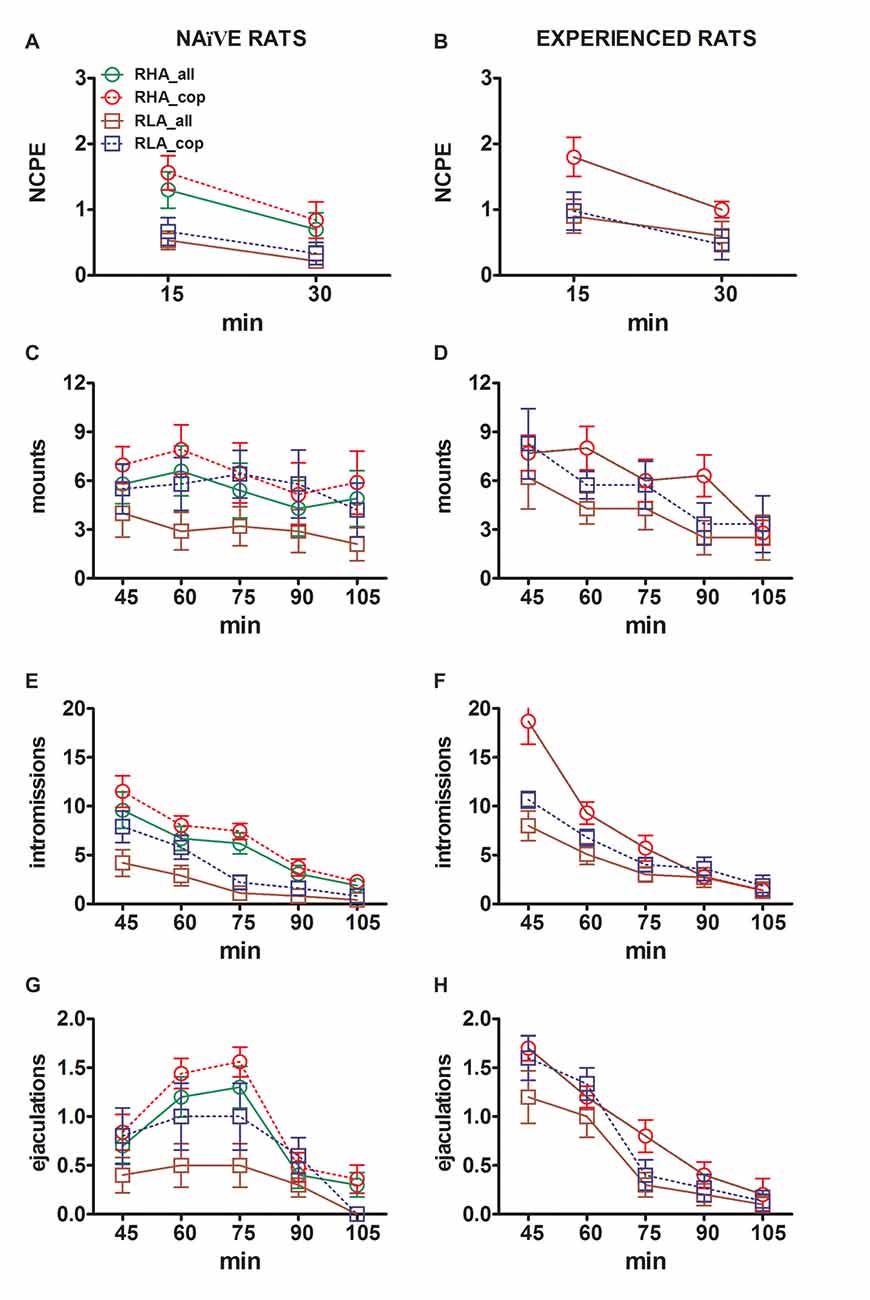 图3。 从性生活中记录的非接触性阴茎勃起(NCPE),坐骑,插入和射精次数的变化 (A,C,E,G) 和经验丰富的RHA和RLA大鼠 (B,d,F,H) 在微透析实验中报道了图 2。 所有实验条件与图中图例中描述的相同 2。 值是每组所有12大鼠获得的值的平均值±SEM(包括那些被指定为全时评分; RHA =实线绿线,RLA =实线棕色线)或仅与射精交配的那些(RHA =红色虚线) ; RLA =蓝色虚线)。 在经验丰富的RHA大鼠中,所有动物均达到交配并且值相同。 由于通过双向ANOVA分析数据,未发现大鼠线因子(RHA与RLA)之间的显着相互作用,性经验水平因素(天真与经历)和时间,如表所示 6, 事后 没有报告比较,如“材料和方法”,“统计”小节中所述。
图3。 从性生活中记录的非接触性阴茎勃起(NCPE),坐骑,插入和射精次数的变化 (A,C,E,G) 和经验丰富的RHA和RLA大鼠 (B,d,F,H) 在微透析实验中报道了图 2。 所有实验条件与图中图例中描述的相同 2。 值是每组所有12大鼠获得的值的平均值±SEM(包括那些被指定为全时评分; RHA =实线绿线,RLA =实线棕色线)或仅与射精交配的那些(RHA =红色虚线) ; RLA =蓝色虚线)。 在经验丰富的RHA大鼠中,所有动物均达到交配并且值相同。 由于通过双向ANOVA分析数据,未发现大鼠线因子(RHA与RLA)之间的显着相互作用,性经验水平因素(天真与经历)和时间,如表所示 6, 事后 没有报告比较,如“材料和方法”,“统计”小节中所述。
讨论
这项研究证实并扩展了先前研究的结果,显示RHA和RLA大鼠在厌恶条件下表现出明显不同的应对方式(Driscoll和Bättig,1982; Giorgi等人,2003a; Steimer和Driscoll,2003),在存在难以接近的接受女性和经典交配测试期间显示不同的行为模式(Sanna等,2014a,b, 2015)。 因此,这项研究也表明性生幼稚RHA大鼠(以前从未接触过接受过的女性)与性初始RLA大鼠相比表现出更高的性动机和更好的性行为,例如,RHA大鼠表现出更多的非接触性阴茎勃起(信息素 - 诱发阴茎勃起被认为是性唤起的指标; Sachs等,1994; 萨克斯,2000)以及之前研究中发现的几个交配参数的不同性别变化(Sanna等,2014a,b, 2015)。 这些差异在获得性经验之后仍然存在于罗马大鼠品系中,即在性行为稳定后通过五次接触和与接受性女性进行交配测试。 该研究还首次表明,在幼稚或经历的RHA和RLA大鼠系中,非接触性阴茎勃起和性交互作用同时伴随着细胞外多巴胺(及其主要代谢物DOPAC)浓度的增加。通过脑内微透析从两个罗马大鼠系的mPFC获得的透析液中的NA和NA。 有趣的是,mPFC中细胞外多巴胺和NA的这些增加似乎与大鼠表型或性经历水平的差异有关。 因此,如结果部分中广泛显示的,性行为和经历过的RHA大鼠在性行为的食欲和完成阶段与其RLA对应物相比,细胞外多巴胺和NA显示出更高和更持久的增加。 此外,与他们的幼稚对应物相比,两种罗马品系的性经验大鼠在性行为的两个阶段中显示出mPFC中细胞外多巴胺和NA的更高且持久的增加。 最后,mPFC中细胞外多巴胺和NA的所有上述差异显然与性行为的行为参数的差异有关,例如,与他们的天真对应物相比,两种罗马品系的性经验大鼠表现出更高的性动机和更好的交配性能。与性生稚和经验丰富的RLA对照相比,性初始和经验丰富的RHA大鼠表现出更高的性动机和更好的交配表现。
至于在性活动期间mPFC透析液中发现的细胞外多巴胺的增加,第一次增加发生在交配装置中性接受雌性引入后的第一个15分钟,当雌性不能接近雄性时,通常表现出来在这种情况下非接触性阴茎勃起。 在雌性可接近雄性交配后的第一个15分钟期间发生细胞外多巴胺的第二次和更大的增加。 有趣的是,在幼稚的情况下,细胞外多巴胺主要在测试的第一部分增加,而在第二部分,可接触的女性有回到基础值的趋势,在性经历的情况下,多巴胺在整个测试期间保持升高达到峰值在45-60之后,在两个罗马鼠系中交配。
除了在性行为和经历的情况下RHA和RLA大鼠的性活动的不同阶段期间细胞外多巴胺的上述差异外,该研究还显示两种罗马大鼠系之前的细胞外DOPAC浓度的意外重要和高度显着差异(即,基础值)和在接合装置中引入接收雌性之后。 因此,在RHA大鼠中发现mPFC透析液中细胞外DOPAC的基础值比在RLA大鼠中发现的2.6倍高约XNUMX倍。 尽管在这些实验条件下从两条罗马大鼠品系获得的mPFC透析液中细胞外多巴胺的基础值存在非常适度的差异,但RHA和RLA大鼠之间的这种差异在性初始和经历的条件下发生并持续存在。 总之,这些差异可能表明两种罗马鼠系之间mPFC水平的多巴胺转换率不同。 因此,DOPAC浓度通常反映释放的多巴胺的量,然后由多巴胺能神经末梢重新捕获并通过单氨基氧化酶转化为DOPAC(MAO; Carlsson,1975)。 因此,在RHA大鼠中发现较高的基础DOPAC浓度表明在RHA大鼠的mPFC中多巴胺能释放并通过多巴胺能神经末梢重新捕获,例如,与RLA相比,RHA大鼠的中皮质多巴胺能系统的基础活性更高。老鼠。 RHA大鼠中的中皮质多巴胺能张力高于RLA大鼠,也得到了性行为和经验丰富的RHA大鼠的mPFC透析液性活动期间发现的细胞外多巴胺浓度较高的RLA对应物。 在这方面,有必要回顾一下,更高的多巴胺能基调被认为至少部分地归因于RHA和RLA大鼠中存在的许多不同甚至相反的行为特征。 因此,RHA大鼠是活跃的大麻,高度冲动,新奇和感觉寻求者,并且倾向于摄入和滥用几类成瘾药物,而RLA大鼠是反应性的copers,hyperemotional并且易于发展抑郁样症状(Zeier等,1978; Giorgi等,1994, 2003b, 2007; Corda等,1997, 2014; Escorihuela等,1999; Steimer和Driscoll,2003; Lecca等,2004; Giménez-Llort等,2005; Carrasco等,2008; Fattore等,2009; Moreno等,2010; Coppens等,2012; Díaz-Morán等人,2012; Sabariego等,2013; Manzo等人,2014a,b; Oliveras等,2015),包括性生活和经验丰富的RHA大鼠与RHA大鼠相比更高的性动机和更好的交配表现(Sanna等,2015)。 有性幼稚和经验丰富的RHA大鼠的mPFC透析液中细胞外多巴胺浓度的较高增加类似于性暴力和经验丰富的RHA大鼠伏隔核中透析液中的细胞外多巴胺浓度较高的RLA对应物(Sanna等,2015)。 然而,与mPFC不同,来自性初始和经历过的RHA和RLA大鼠伏隔核的透析液中的细胞外多巴胺和DOPAC的基础水平没有显着差异(Sanna等,2015)。 因此,性接触和经验丰富的RHA大鼠的伏隔核中较高的中脑边缘多巴胺能基调可以仅通过RHA大鼠伏隔核中透析液中发现的较高细胞外多巴胺浓度来推断,与接受者存在时的RLA对应物相比较。女性,而不是由RHA大鼠和RLA大鼠中DOPAC浓度的较高和较低基础水平所揭示的不同的基础多巴胺能活性,发现在本研究中发生在mPFC中。 这也与先前报道的结果一致,该结果显示RHA大鼠的mPFC透析液中的细胞外多巴胺水平高于RLA大鼠,而伏隔核壳中透析液中的多巴胺水平相似。 RHA和RLA大鼠在不同实验条件下的作用(D'Angio等,1988; Scatton等,1988; Willig等,1991; Giorgi等人,2003a, 2007)。 尽管mPFC和伏隔核之间存在上述差异,但是当从性初始到经验状态时,性经验似乎在mPFC中产生更长时间的罗马鼠线中多巴胺释放的时间模式增加,尽管这种增加RHA大鼠的RAD大鼠通常高于RLA大鼠。 这与伏隔核中发现的有些不同,其中性经验倾向于将细胞外多巴胺增加至RHA大鼠的交配试验的第一部分,而在RLA大鼠的整个交配试验中增加它(Sanna等,2015).
据我们所知,本研究还首次表明,与多巴胺相比,RHA和RLA大鼠的mPFC透析液在性活动过程中,细胞外多巴胺和细胞外NA均增加,但在这种情况下也是如此。与罗马鼠系和性经历有关的差异。 特别是,与来自RHA和RLA大鼠的mPFC的透析液中发现的基础多巴胺水平非常相似,来自RHA大鼠的mPFC的透析液中的细胞外NA的基础水平显着更高(约2,5倍) )比RLA大鼠的那些。 这些差异的原因尚不清楚,但如上文对多巴胺和DOPAC所讨论的,这些结果可能表明,与RLA大鼠相比,至少在RHA大鼠的mPFC皮质中存在更高的基础去甲肾上腺素能张力。 如上面针对多巴胺所讨论的,这种更高的去甲肾上腺素能基调也可能涉及RHA和RLA大鼠中存在的不同且通常相反的行为特征。 进一步支持RHA大鼠中的去甲肾上腺素能力高于mPFC中的RLA大鼠,结果表明,在性生活和经历的情况下,基础细胞外NA水平的差异在性活动期间也持续存在,例如,细胞外NA水平增加。当暴露于难以接近的雌性时,mPFC在RHA大鼠中进行透析,并且在所有交配期间比RLA大鼠更多,其中NA水平仅在交配期的第一个15分钟内增加,之后NA水平恢复到基础值。 。 在性生活和经历的情况下都观察到细胞外NA水平的这些变化,但是在性经历的RHA和RLA大鼠中,与其性天真的对应物相比,NA水平的增加更高。 因此,性经验显然诱导了RHA大鼠的mPFC中的去甲肾上腺素能活性的进一步增强,并且在较小程度上诱导了RLA大鼠,如已经讨论过的多巴胺。 与RLA大鼠相比,RHA大鼠的mPFC中伴随的更高的去甲肾上腺素能和多巴胺能音调值得一些评论。 事实上,据报道,mPFC中的去甲肾上腺素能活性可以通过不同的机制影响多巴胺释放(例如见 Carboni等,1990, 2006; Gresch等,1995; Westernik等,1998 和其中的参考文献)。 其中一个与NET的活性有关,其显示结合多巴胺的亲和力甚至高于从PFC匀浆获得的突触体中NA的亲和力(Horn,1973)。 如果这是在mPFC中发生的 体内在mPFC中释放的多巴胺不仅可以通过多巴胺能而且还可以通过去甲肾上腺素神经末梢以甚至高于NA的速度重新捕获。 这可能反过来导致多巴胺能终端的多巴胺释放增加(例如,增加的多巴胺能基调),以补偿从突触间隙中除去的多巴胺的量; 因此,与RLA大鼠相比,在RHA大鼠中发现的较高的去甲肾上腺素能基调可以至少部分地促成在RHA大鼠的mPFC中发现的较高的多巴胺能基调。 需要进一步的研究来验证这种可能性。 值得注意的是,本研究中在RHA和RLA大鼠之间的mPFC中鉴定的去甲肾上腺素能活性的差异也可能在它们对抗抑郁治疗的不同反应中起作用。 事实上,RHA大鼠被认为是弹性模型,而RLA大鼠则是易患抑郁症的模型。 因此,使用地昔帕明,氟西汀和氯米帕明等抗抑郁药进行急性或慢性治疗均可显着改善RLA大鼠强迫游泳试验中的行为反应,而不影响RHA大鼠的反应(Piras等,2010, 2014).
最后,这项研究证实并扩展了先前的研究结果,表明mPFC参与控制性行为,尽管这一大脑区域在性行为中的确切作用远未明确(Fernández-Guasti等人,1994; Agmo和Villalpando,1995; Agmo等人,1995; Hernández-Gonzáles等,1998, 2007; Kakeyama等,2003; Balfour等人,2006; Afonso等人,2007; 戴维斯等人,2010; Febo,2011)。 最近,使用单细胞发射记录范例显示了在接近雄性大鼠接近不可接近的性接受性雌性行为期间mPFC中的选择性细胞发射(Febo,2011)。 有趣的是,在第一次暴露男性到难以接近的接受女性期间没有反应的神经元在第二次暴露期间变得活跃,好像之前的经历能够诱导塑性变化导致行为反应的变化。 尽管如此,发现mPFC的具有ibotenic acid的病变不能改变具有性接受性的雌性的雄性大鼠的性行为,也不能改变条件性位置对性奖励的偏好的表达。 然而,在这些受损动物中,发现在与厌恶刺激配对时形成对性活动的条件厌恶的能力被完全消除(戴维斯等人,2010)。 这表明,性行为期间的mPFC激活在外部和内部信息的整合中起作用,用于执行和控制目标导向的行为,而不是表达对天然强化物的先天反应(见 Goto和Grace,2005)。 对于这项工作更为重要的是,一些研究表明,涉及性动机和性行为的大脑区域,例如伏隔核,腹侧被盖区,内侧视前区,纹状体末端的床核,基底外侧杏仁核和小脑下丘脑丘脑核接收输入来自性活动期间的mPFC(见 Balfour等人,2006)。 许多这些大脑区域参与了一个复杂的神经回路,涉及控制性行为,从性动机和奖励到性能力。 该电路包括起源于下丘脑(PVN)PVN的催产素神经元,并突出到腹侧被盖区,伏隔核,海马,杏仁核,PFC,纹状体末端,延髓和脊髓的床核,中脑边缘/中皮质多巴胺能。神经元从腹侧被盖区突出到伏隔核和PFC,下丘脑多巴胺能神经元和谷氨酸能神经元,参与电路的几个区域的局部和系统水平(见 Melis和Argiolas,1995, 2011; Melis等,2003, 2007, 2009, 2010; Succu等,2007, 2008, 2011)。 因此,在性活动期间观察到的mPFC透析液中细胞外多巴胺(以及NA)的增加可能表明mPFC是该电路的活跃部分,即中皮质多巴胺能神经元一起参与控制性活动。与去甲肾上腺素,中脑边缘多巴胺能,催产素和谷氨酸能神经元有关,虽然它似乎并不是表达性行为的必要条件(戴维斯等人,2010)。 根据这一假设,本研究表明,性活动过程中mPFC中多巴胺和NA释放的差异伴随着非接触性阴茎勃起次数和交配参数变化的差异,多巴胺和NA的释放更高。食欲和完成阶段与更高的性动机和更好的交配性能相关,在罗马鼠和有性生活和有经验的条件下。
上面给出的讨论主要基于众所周知的在性活动期间雄性大鼠伏隔核中发生的细胞外多巴胺的增加(Pfaus等,1990; Pleim等,1990; Pfaus和Phillips,1991; Damsma等,1992; Wenkstern等,1993)。 然而,多巴胺和其他神经递质可能在其他实验条件下释放在mPFC(和伏隔核)中,例如在压力期间或应对它时(Thierry等,1976; Fadda等人,1978),与性行为并不严格相关。 因此,有人可能会争辩说,在mPFC(本研究)或伏核中细胞外多巴胺中发现的多巴胺增加(Sanna等,2015),更多地与次要的压力有关,或与应对性接受的女性的存在和/或相互作用而不是性活动有关。 这可能与RHA和RLA大鼠有一定的相关性,它们表现出明显不同的应对方式和不同的下丘脑 - 垂体 - 肾上腺轴激活以应对厌恶条件(RLA大鼠显示该轴的激活远高于RHA大鼠;见 Carrasco等,2008; Díaz-Morán等人,2012)。 这也提高了应激激素(即皮质类固醇)可能影响mPFC和其他大脑区域的多巴胺能活性的可能性。 事实上,RLA大鼠是反应性的copers并且表现出以低运动和冷冻为特征的高度激动行为,而RHA大鼠表现出主动应对行为,旨在获得对压力源的控制(Driscoll和Bättig,1982; Willig等,1991; Escorihuela等,1999; Steimer和Driscoll,2003; Giorgi等,2007)。 尽管不可能完全排除这种情况可能发生在未接受过性行为的大鼠(之前从未与雌性受体发生过相互作用)中,尤其是在未接受过性行为的RLA大鼠中(见上文),但这是不可能的。 在我们的实验条件下,已尽一切可能避免各种压力(请参阅“材料和方法”部分),并且组织实验的方式是雄性不会立即与雌性互动,而只是在暴露于雌性的30分钟之后,在此期间,测得的多巴胺增加量远低于性交时发生的增加量。 如果在罗马大鼠的mPFC(以及伏隔核)中发现多巴胺增加(天真的RHA和RLA大鼠在mPFC和伏隔核的透析液中具有基础水平的细胞外多巴胺,非常相似)实际上是继发于(i)多巴胺的增加会在第一个时间段内(当雄性动物与雌性动物并存时而不是后来)被发现要高得多,如在交配时发现的; (ii)在获得稳定的性活动后,即完成学习过程(包括应对新奇现象)并且在性行为的执行中仅扮演较小的角色后,多巴胺的增加预计将消失或被强烈减少。活动,而相反,当重复性活动时,它继续发生。 换句话说,即使在已经了解性行为之后,mPFC(以及伏隔核)中的细胞外多巴胺始终会增加。 需要进一步的研究来阐明多巴胺活性增加在mPFC(和伏隔核;请参见其中的“简介和参考”部分)中的功能作用。
总之,这项研究首次表明,与性接受性女性的相互作用的存在,甚至更多,导致性暴力的mPFC透析液中的细胞外多巴胺和NA的增加(从未暴露于性刺激) )和性经历(经历了五次初步交配测试并表现出稳定的性表现)RHA和RLA大鼠,其表现出明显不同的应对方式,以应对厌恶条件和与性接纳女性不同的交配行为模式。 因此,在性活动的预期和完成阶段期间,与性RBA大鼠相比,在性暴力和经历过的RHA大鼠的透析液中发现多巴胺和NA浓度更高。 这些结果可能部分是由于RHA大鼠的mPFC中的多巴胺能和去甲肾上腺素能高于RLA大鼠,如伏隔核中的多巴胺所报道的(Sanna等,2015)。 此外,在性经历的RHA中发现细胞外mPFC多巴胺和NA,并且在较小程度上,RLA大鼠与基础条件下的性天真对应物以及性行为的食欲和完成阶段期间相比较高。 这表明性经验诱导了塑料过程,进一步加强了mPFC中的多巴胺和NA神经传递,正如罗马鼠线伏核中的多巴胺所发现的那样(Sanna等,2015)。 两种罗马品系的mPFC中多巴胺能和去甲肾上腺素能神经传递的上述差异不仅可以在性行为的不同模式中起作用,而且可以在表征这两种大鼠品系的不同且通常相反的行为特征中起作用。
作者贡献
FS,AA,MRM,OG和MGC设计了该项目。 FS和JB设计,执行和分析了性行为和微透析实验的数据。 MAP,OG和MGC选择和繁殖罗马大鼠。 FS,AA,MRM,OG和MGC对该研究进行了监督。 FS,AA,MRM,OG和MGC撰写了手稿。 所有作者都讨论了结果,并对手稿进行了评论。
资金
这项工作部分得到意大利大学和研究部(MIUR)(年轻研究人员的科学独立,SIR 2014,代码编号RBSI14IUX7)给予撒丁岛自治区(ARS)FS(LR 7 / 2007)的资助。 ,“在撒丁岛推广科学研究和技术创新”,欧洲社会基金,2007-2013,项目代号CRP-59842)到OG,从卡利亚里大学到AA,MRM,FS,OG和MGC。
利益冲突声明
作者声明,研究是在没有任何可被解释为潜在利益冲突的商业或金融关系的情况下进行的。
致谢
卡利亚里大学Barbara Tuveri博士,负责动物住房和护理。
参考资料
Afonso,VM,Sison,M.,Lovic,V。和Fleming,AS(2007)。 雌性大鼠的内侧前额叶皮层损伤影响性和母性行为及其顺序组织。 Behav。 神经科学。 121,515-526。 doi:10.1037 / 0735-7044.121.3.515
Agmo,A。和Villalpando,A。(1995)。 中枢神经兴奋剂促进具有内侧前额叶皮质损伤的雄性大鼠的性行为。 Brain Res。 696, 187–193. doi: 10.1016/0006-8993(95)00853-i
Agmo,A.,Villalpando,A.,Picker,Z。和Fernández,H。(1995)。 内侧前额叶皮质的损伤和雄性大鼠的性行为。 Brain Res。 696, 177–186. doi: 10.1016/0006-8993(95)00852-h
Argiolas,A。和Melis,MR(1995)。 阴茎勃起的神经调节:概述神经递质和神经肽的作用。 PROG。 神经生物学。 47, 235–255. doi: 10.1016/0301-0082(95)00020-8
Argiolas,A。和Melis,MR(2005)。 阴茎勃起的中枢控制:下丘脑室旁核的作用。 PROG。 神经生物学。 76,1-21。 doi:10.1016 / j.pneurobio.2005.06.002
Argiolas,A。和Melis,MR(2013)。 神经肽和从过去到现在的性行为的中央控制:审查。 PROG。 神经生物学。 108,80-107。 doi:10.1016 / j.pneurobio.2013.06.006
Balfour,ME,Brown,JL,Yu,L。和Coolen,LM(2006)。 来自内侧前额叶皮层的传出物对雄性大鼠性行为后神经活化的潜在贡献。 神经 137,1259-1276。 doi:10.1016 / j.neuroscience.2005.11.013
Balfour,ME,Yu,L。和Coolen,LM(2004)。 性行为和性相关的环境因素激活了雄性大鼠的中脑边缘系统。 神经精神药理学 29,718-730。 doi:10.1038 / sj.npp.1300350
Beloate,LN,Omrani,A.,Adan,RA,Webb,IC和Coolen,LM(2016)。 在雄性大鼠性行为期间腹侧被盖区多巴胺细胞活化调节性别禁欲后的神经可塑性和d-苯异丙胺交叉致敏。 J.Neurosci。 36,9949-9961。 doi:10.1523 / JNEUROSCI.0937-16.2016
Berridge,K。和Robinson,T。(1998)。 多巴胺在奖励中的作用是什么:享乐效应,奖励学习或激励突显? Brain Res。 启示录 28, 309–369. doi: 10.1016/s0165-0173(98)00019-8
Broadhurst,PL和Bignami,G。(1965)。 心理遗传选择的相关效应:大鼠罗马高低回避菌株的研究。 Behav。 RES。 疗法。 3, 273–280. doi: 10.1016/0005-7967(64)90033-6
Carboni,E.,Silvagni,A.,Vacca,C。和Di Chiara,G。(2006)。 去甲肾上腺素和多巴胺载体阻断对伏隔核,纹状体末端和前额叶皮层细胞外多巴胺增加的累积作用。 J. Neurochem。 96,473-481。 doi:10.1111 / j.1471-4159.2005.03556.x
Carboni,E.,Tanda,GL,Frau,R。和Di Chiara,G。(1990)。 阻断去甲肾上腺素载体增加前额皮质中的细胞外多巴胺浓度:多巴胺被摄取的证据 体内 由noradrenergic终端。 J. Neurochem。 55, 1067–1070. doi: 10.1111/j.1471-4159.1990.tb04599.x
Carrasco,J.,Márquez,C.,Nadal,R.,Tobeña,A.,Fernández-Teruel,A。和Armario,A。(2008)。 在近交罗马大鼠品系中表征下丘脑 - 垂体 - 肾上腺轴的中心和外周组分。 心理神经 33,437-445。 doi:10.1016 / j.psyneuen.2008.01.001
Coppens,CM,de Boer,SF,Steimer,T。和Koolhaas,JM(2012)。 罗马高低回避大鼠的冲动和攻击行为:基线差异和青少年社会压力引起的变化。 生理学。 Behav。 105,1156-1160。 doi:10.1016 / j.physbeh.2011.12.013
Corda,MG,Lecca,D.,Piras,G.,Di Chiara,G。和Giorgi,O。(1997)。 罗马高回避和罗马低回避大鼠中枢神经系统多巴胺能和GABA能神经传递的生化参数。 Behav。 遗传学。 27,527-536。 doi:10.1023 / A:1021452814574
Corda,MG,Piras,G.,Piludu,MA和Giorgi,O。(2014)。 自愿乙醇消耗对罗马高回避和低回避大鼠伏核中多巴胺输出的不同影响:行为和脑微透析研究。 世界J.Neurosci。 4,279-292。 doi:10.4236 / wjns.2014.43031
Dahlstrom,A。和Fuxe,K。(1964)。 中枢神经系统中含有单胺的神经元存在的证据。 I.脑干神经元细胞体中单胺的证明。 Acta Physiol。 SCAND。 增刊。 62,1-54。
Damsma,G.,Pfaus,J.,Wenkstern,D.,Phillips,A。和Fibiger,H。(1992)。 性行为增加雄性大鼠伏隔核和纹状体中的多巴胺传递:与新奇和运动的比较。 Behav。 神经科学。 106,181-191。 doi:10.1037 / 0735-7044.106.1.181
D'Angio,M.,Serrano,A.,Driscoll,P。和Scatton,B。(1988)。 压力环境刺激增加了前额叶皮层的低血压(罗马高回避)但不是高度消退(罗马低回避)大鼠的细胞外DOPAC水平。 一个 体内 伏安研究。 Brain Res。 451, 237–247. doi: 10.1016/0006-8993(88)90768-8
Davis,JF,Loos,M.,Di Sebastiano,AR,Brown,JL,Lehman,MN和Coolen,LM(2010)。 内侧前额叶皮层的损伤导致雄性大鼠的适应不良性行为。 生物学。 精神病学 67,1199-1204。 doi:10.1016 / j.biopsych.2009.12.029
Díaz-Morán,S.,Palència,M.,Mont-Cardona,C.,Cañete,T.,Blázquez,G.,Martínez-Membrives,E.,et al。 (2012)。 遗传异质大鼠的应对方式和应激激素反应:与罗马大鼠品系比较。 Behav。 Brain Res。 228,203-210。 doi:10.1016 / j.bbr.2011.12.002
Driscoll,P。和Bättig,K。(1982)。 “大鼠的行为,情绪和神经化学特征,选择主动,双向回避表现的极端差异,” 大脑的遗传学,编辑。 I. Lieblich(阿姆斯特丹:Elsevier),95-123。
Escorihuela,RM,Fernández-Teruel,A.,Gil,L.,Aguilar,R.,Tobeña,A。和Driscoll,P。(1999)。 近交罗马高低回避大鼠:焦虑,新奇寻求和穿梭行为的差异。 生理学。 Behav。 67, 19–26. doi: 10.1016/S0031-9384(99)00064-5
Everitt,BJ(1990)。 性动机:对雄性大鼠食欲和交配反应机制的神经和行为分析。 神经科学。 Biobehav。 启示录 14, 217–232. doi: 10.1016/s0149-7634(05)80222-2
Fadda,F.,Argiolas,A.,Melis,MR,Tissari,AH,Onali,PL和Gessa,GL(1978)。 应激诱导的大脑皮层和n.accumbens中3,4-二羟基苯乙酸(DOPAC)水平的增加:地西泮逆转。 Life Sci. 23, 2219–2224. doi: 10.1016/b978-0-08-023768-8.51934-4
Fattore,L.,Piras,G.,Corda,MG和Giorgi,O。(2009)。 罗马高回避和低回避大鼠系在静脉注射可卡因自我给药的获得,维持,消退和恢复方面存在差异。 神经精神药理学 34,1091-1101。 doi:10.1038 / npp.2008.43
Fernández-Guasti,A.,Omaña-Zapata,I.,Luján,M。和Condés-Lara,M。(1994)。 坐骨神经结扎对性经验和缺乏经验的雄性大鼠性行为的影响:额极剥脱的影响。 生理学。 Behav。 55, 577–581. doi: 10.1016/0031-9384(94)90119-8
Fernández-Teruel,A.,Driscoll,P.,Gil,L.,Aguilar,R.,Tobeña,A。和Escorihuela,RM(2002)。 环境富集对两种大鼠系(RHA / Verh和RLA / Verh)的新奇寻求,糖精和乙醇摄入的持久影响在激励寻求行为方面存在差异。 药理学。 生物化学。 Behav。 73, 225–231. doi: 10.1016/s0091-3057(02)00784-0
Giménez-Llort,L.,Cañete,T.,Guitart-Masip,M.,Fernández-Teruel,A。和Tobeña,A。(2005)。 两种独特的阿扑吗啡诱导表型在罗马高回避和低回避大鼠.. 生理学。 Behav。 86,458-466。 doi:10.1016 / j.physbeh.2005.07.021
Giorgi,O.,Lecca,D.,Piras,G。和Corda,MG(2005)。 急性或重复苯丙胺注射后伏隔核和壳中多巴胺释放的差异激活:罗马高回避和低回避大鼠系的比较研究。 神经 135,987-998。 doi:10.1016 / j.neuroscience.2005.06.075
Giorgi,O.,Lecca,D.,Piras,G.,Driscoll,P。和Corda,MG(2003a)。 在两种心理遗传选择的大鼠系中,中脑皮质多巴胺释放与恐惧相关行为之间的分离,这些大鼠的应对策略与厌恶条件不同。 欧元。 J.Neurosci。 17,2716-2726。 doi:10.1046 / j.1460-9568.2003.02689.x
Giorgi,O.,Piras,G.,Lecca,D.,Hansson,S.,Driscoll,P。和Corda,MG(2003b)。 罗马高回避和低回避大鼠中枢5-羟色胺能传递的差异神经化学特性。 J. Neurochem。 86,422-431。 doi:10.1046 / j.1471-4159.2003.01845.x
Giorgi,O.,Orlandi,M.,Escorihuela,RM,Driscoll,P.,Lecca,D。和Corda,MG(1994)。 罗马高回避和罗马低回避大鼠脑内GABA能和多巴胺能传递。 Brain Res。 638, 133–138. doi: 10.1016/0006-8993(94)90642-4
Giorgi,O.,Piras,G。和Corda,MG(2007)。 精神遗传学选择的罗马高回避和低回避鼠系:研究个体对药物成瘾的脆弱性的模型。 神经科学。 Biobehav。 启示录 31,148-163。 doi:10.1016 / j.neubiorev.2006.07.008
Gresch,PJ,Sved,AF,Zigmond,MJ和Finlay,JM(1995)。 内源性去甲肾上腺素对大鼠内侧前额叶皮层细胞外多巴胺的局部影响。 J. Neurochem。 65,111-116。 doi:10.1046 / j.1471-4159.1995.65010111.x
Hernández-Gonzáles,M.,Guevara,MA,Cervantes,M.,Morali,G。和Corsi-Cabrera,M。(1998)。 作为因子分析的结果,在雄性大鼠的性交互过程中皮质 - 前额脑电图的特征频带。 生理学 92, 43–50. doi: 10.1016/s0928-4257(98)80022-3
Hernández-Gonzáles,M.,Prieto-Beracoechea,CA,Arteaga-Silva,M。和Guevara,MA(2007)。 在大鼠的性动机任务期间内侧和眼眶前额叶皮层的不同功能。 生理学。 Behav。 90,450-458。 doi:10.1016 / j.physbeh.2006.10.006
Horn,AS(1973)。 在大鼠脑匀浆中抑制儿茶酚胺摄入去甲肾上腺素能和多巴胺能神经元的突触体的结构 - 活性关系。 BR。 J. Pharmacol。 47, 332–338. doi: 10.1111/j.1476-5381.1973.tb08331.x
Hull,EM,Du,J.,Lorrain,DS和Matuszewich,L。(1995)。 内侧视前区的细胞外多巴胺:对性交动和交配激素控制的影响。 J.Neurosci。 15,7465-7471。
Hull,EM,Lorrain,DS,Du,J.,Matuszewich,L.,Lumley,LA,Putnam,SK,et al。 (1999)。 激素 - 神经递质相互作用控制性行为。 Behav。 Brain Res。 105, 105–116. doi: 10.1016/s0166-4328(99)00086-8
Hull,EM,Weber,MS,Eaton,RC,Dua,R.,Markowski,VP,Lumley,L.,et al。 (1991)。 腹侧被盖区域中的多巴胺受体影响雄性大鼠的运动,但不是动机或反射性交配的组成部分。 Brain Res。 554, 72–76. doi: 10.1016/0006-8993(91)90173-S
Ikemoto,S。和Panksepp,J。(1999)。 伏隔核多巴胺在动机行为中的作用:一种统一的解释,特别参考奖励寻求。 Brain Res。 启示录 31, 6–41. doi: 10.1016/S0165-0173(99)00023-5
Kakeyama,M.,Sone,H.,Miyabara,Y。和Tohyama,C。(2003)。 围产期暴露于2,3,7,8-四氯二苯并对二恶英改变了新皮质中BDNF mRNA的活性依赖性表达和成年期雄性大鼠的性行为。 神经毒理学 24, 207–217. doi: 10.1016/S0161-813x(02)00214-0
Lecca,D.,Piras,G.,Driscoll,P.,Giorgi,O。和Corda,MG(2004)。 伏隔核壳和核心中多巴胺输出的差异激活与运动对成瘾药物的反应有关:罗马高回避和低回避大鼠的脑透析研究。 神经药理学 46,688-699。 doi:10.1016 / j.neuropharm.2003.11.011
Manzo,L.,Gómez,MJ,Callejas-Aguilera,JE,Donaire,R.,Sabariego,M.,Fernández-Teruel,A.,et al。 (2014a)。 乙醇偏好与感觉/新奇寻求的关系。 生理学。 Behav。 133,53-60。 doi:10.1016 / j.physbeh.2014.05.003
Manzo,L.,Gómez,MJ,Callejas-Aguilera,JE,Fernández-Teruel,A.,Papini,MR和Torres,C。(2014b)。 大鼠激励损失引起的抗焦虑自我药疗。 生理学。 Behav。 123,86-92。 doi:10.1016 / j.physbeh.2013.10.002
Masana,M.,Bortolozzi,A。和Artigas,F。(2011)。 去甲肾上腺素能药物对中皮质多巴胺能传递的选择性增强:精神分裂症的治疗机会。 诠释。 J. Neuropsychopharmacol。 14,53-68。 doi:10.1017 / s1461145710000908
Meisel,RL和Sachs,BD(1994)。 “男性性行为的生理学” 生殖生理学,卷。 2,2nd Edn。编辑E. Knobil和J. Neil(纽约,纽约:Raven Press),3-96。
Melis,MR和Argiolas,A。(1995)。 多巴胺和性行为。 神经科学。 Biobehav。 启示录 19, 19–38. doi: 10.1016/0149-7634(94)00020-2
Melis,MR和Argiolas,A。(2011)。 阴茎勃起的中枢控制:重新探讨催产素在雄性大鼠中的作用及其与多巴胺和谷氨酸的相互作用。 神经科学。 Biobehav。 启示录 35,939-955。 doi:10.1016 / j.neubiorev.2010.10.014
Melis,MR,Melis,T.,Cocco,C.,Succu,S.,Sanna,F.,Pillolla,G.,et al。 (2007)。 注射到腹侧被盖区域的催产素诱导阴茎勃起并增加雄性大鼠下丘脑伏核和室旁核的细胞外多巴胺。 欧元。 J.Neurosci。 26,1026-1035。 doi:10.1111 / j.1460-9568.2007.05721.x
Melis,MR,Succu,S.,Cocco,C.,Caboni,E.,Sanna,F.,Boi,A.,et al。 (2010)。 当注射到腹侧下托室时,催产素诱导阴茎勃起:一氧化氮和谷氨酸的作用。 神经药理学 58,1153-1160。 doi:10.1016 / j.neuropharm.2010.02.008
Melis,MR,Succu,S.,Mascia,MS,Cortis,L。和Argiolas,A。(2003)。 在性活动期间,雄性大鼠的室旁核中的细胞外多巴胺增加。 欧元。 J.Neurosci。 17,1266-1272。 doi:10.1046 / j.1460-9568.2003.02558.x
Melis,MR,Succu,S.,Sanna,F.,Boi,A。和Argiolas,A。(2009)。 注射到腹侧下托或杏仁核的后内侧皮质核中的催产素诱导阴茎勃起并增加雄性大鼠伏隔核中的细胞外多巴胺水平。 欧元。 J.Neurosci。 30,1349-1357。 doi:10.1111 / j.1460-9568.2009.06912.x
Moreno,M.,Cardona,D.,Gómez,MJ,Sánchez-Santed,F.,Tobeña,A.,Fernández-Teruel,A.,et al。 (2010)。 罗马高回避和低回避大鼠品系的冲动特性:行为和神经化学差异。 神经精神药理学 35,1198-1208。 doi:10.1038 / npp.2009.224
Oliveras,I.,Río-Álamos,C.,Cañete,T.,Blázquez,G.,Martínez-Membrives,E.,Giorgi,O。,et al。 (2015)。 Preulse抑制预测近交罗马高回避和低回避大鼠和遗传异质NIH-HS大鼠的空间工作记忆表现:与研究精神分裂症的预先注意力和认知异常相关。 面前。 Behav。 神经科学。 9:213。 doi:10.3389 / fnbeh.2015.00213
Pfaus,JG(2010)。 多巴胺:帮助男性至少交换200百万年:对Kleitz-Nelson等人的理论评论。 (2010)。 Behav。 神经科学。 124,877-880。 doi:10.1037 / a0021823
Pfaus,JG,Damsma,G.,Nomikos,GG,Wenkstern,DG,Blaha,CD,Phillips,AG,et al。 (1990)。 性行为增强雄性大鼠的中枢多巴胺传递。 Brain Res。 530, 345–348. doi: 10.1016/0006-8993(90)91309-5
Pfaus,JG和Phillips,AG(1991)。 多巴胺在雄性大鼠性行为的预期和完善方面的作用。 Behav。 神经科学。 105,727-747。 doi:10.1037 / 0735-7044.105.5.727
Piras,G.,Giorgi,O。和Corda,MG(2010)。 抗抑郁药对两种经心理选择的大鼠的强迫游泳试验的表现的影响,这些大鼠的应对策略与厌恶条件不同。 精神药理学 211, 403–414. doi: 10.1007/s00213-010-1904-x
Piras,G.,Piludu,MA,Giorgi,O。和Corda,MG(2014)。 慢性抗抑郁治疗在假定的脆弱性遗传模型(罗马低回避大鼠)和抵抗(罗马高回避大鼠)对压力诱导的抑郁症的影响。 精神药理学 231, 43–53. doi: 10.1007/s00213-013-3205-7
Pitchers,KK,Frohmader,KS,Vialou,V.,Mouzon,K.,Nestler,EJ,Lehman,MN,et al。 (2010)。 伏隔核中的ΔFosB对于增强性奖励的效果至关重要。 基因脑行为。 9, 831–840. doi: 10.1111/j.1601-183X.2010.00621.x
Pitchers,KK,Coppens,CM,Beloate,LN,Fuller,J.,Van,S.,Frohmader,KS,et al。 (2014)。 内源性阿片类药物诱导的腹侧被盖区多巴胺能神经元的神经可塑性影响自然和鸦片剂的奖赏。 J.Neurosci。 34,8825-8836。 doi:10.1523 / JNEUROSCI.0133-14.2014
Pitchers,KK,Vialou,V.,Nestler,EJ,Laviolette,SR,Lehman,MN和Coolen,LM(2013)。 自然和药物奖励作用于共同神经可塑性机制,ΔFosB作为关键介质。 J.Neurosci。 33,3434-3442。 doi:10.1523 / JNEUROSCI.4881-12.2013
Pleim,E.,Matochik,J.,Barfield,R。和Auerbach,S。(1990)。 伏隔核多巴胺释放与大鼠男性性行为的相关性。 Brain Res。 524, 160–163. doi: 10.1016/0006-8993(90)90507-8
Robbins,TW和Arnsten,AFT(2009)。 前额执行功能的神经精神药理学:单胺能调节。 安。 Rev. Neurosci。 32,267-287。 doi:10.1146 / annurev.neuro.051508.135535
Sabariego,M.,Morón,I.,Gómez,MJ,Donaire,R.,Tobeña,A.,Fernández-Teruel,A.,et al。 (2013)。 近交罗马高(RHA-I)和罗马低(RLA-I)回避大鼠的激励损失和海马基因表达。 Behav。 Brain Res。 257,62-70。 doi:10.1016 / j.bbr.2013.09.025
Sachs,BD(2000)。 勃起功能,勃起功能障碍和性唤起的生理学和分类的背景方法。 神经科学。 Biobehav。 启示录 24, 541–560. doi: 10.1016/S0149-7634(00)00022-1
Sachs,BD,Akasofu,K.,Citro,JK,Daniels,SB和Natoli,JH(1994)。 来自发情期雌性的非接触性刺激引起大鼠阴茎勃起。 生理学。 Behav。 55, 1073–1079. doi: 10.1016/0031-9384(94)90390-5
Sachs,BD和Barfield,RJ(1976)。 大鼠男性交配行为的功能分析。 进阶 梭哈。 Behav。 7, 91–154. doi: 10.1016/s0065-3454(08)60166-7
Salamone,JD,Pardo,M.,Yohn,SE,López-Cruz,L.,San Miguel,N。和Correa,M。(2016)。 中脑边缘多巴胺和动机行为的调节。 CURR。 最佳。 Behav。 神经科学。 27, 231–257. doi: 10.1007/7854_2015_383
Sanna,F.,Corda,MG,Melis,MR,Piludu,MA,Giorgi,O。和Argiolas,A。(2014a)。 雄性罗马高低避孕大鼠表现出不同的交配行为模式:与sprague dawley大鼠比较。 生理学。 Behav。 127,27-36。 doi:10.1016 / j.physbeh.2014.01.002
Sanna,F.,Piludu,MA,Corda,MG,Argiolas,A.,Giorgi,O。和Melis,MR(2014b)。 多巴胺参与罗马高低避孕大鼠的不同交配行为模式:阿扑吗啡和氟哌啶醇的研究。 药理学。 生物化学。 Behav。 124,211-219。 doi:10.1016 / j.pbb.2014.06.012
Sanna,F.,Piludu,MA,Corda,MG,Melis,MR,Giorgi,O。和Argiolas,A。(2015)。 多巴胺参与罗马高和低回避大鼠之间性行为的差异:脑内微透析研究。 Behav。 Brain Res。 281,177-186。 doi:10.1016 / j.bbr.2014.12.009
Scatton,B.,D'Angio,M.,Driscoll,P。和Serrano,A。(1988)。 一个 体内 伏安研究中脑皮质和中胚层多巴胺能神经元对具有不同情绪水平的大鼠品系的环境刺激的反应。 安。 纽约阿卡德。 科学。 537, 124–137. doi: 10.1111/j.1749-6632.1988.tb42101.x
Steimer,T。和Driscoll,P。(2003)。 精神遗传学选择的罗马高(RHA)和低(RLA)回避大鼠中的不同应激反应和应对方式:行为,神经内分泌和发育方面。 应力 6,87-100。 doi:10.1080 / 1025389031000111320
Succu,S.,Sanna,F.,Argiolas,A。和Melis,MR(2011)。 注射到海马腹侧下托中的催产素通过增加腹侧被盖区域的谷氨酸能神经传递诱导雄性大鼠阴茎勃起。 神经药理学 61,181-188。 doi:10.1016 / j.neuropharm.2011.03.026
Succu,S.,Sanna,F.,Cocco,C.,Melis,T.,Boi,A.,Ferri,GL,et al。 (2008)。 当注射到雄性大鼠的腹侧被盖区时,催产素诱导阴茎勃起:一氧化氮和环状GMP的作用。 欧元。 J.Neurosci。 28,813-821。 doi:10.1111 / j.1460-9568.2008.06385.x
Succu,S.,Sanna,F.,Melis,T.,Boi,A.,Argiolas,A。和Melis,MR(2007)。 雄性大鼠下丘脑室旁核中的多巴胺受体的刺激诱导阴茎勃起并增加伏隔核中的细胞外多巴胺:中枢催产素的参与。 神经药理学 52,1034-1043。 doi:10.1016 / j.neuropharm.2006.10.019
Thierry,AM,Tassin,JP,Blanc,G。和Glowinski,J。(1976)。 应激选择性激活中皮质DA系统。 自然 263,242-244。 doi:10.1038 / 263242a0
Wenkstern,D.,Pfaus,J。和Fibiger,H。(1993)。 雄性大鼠在首次暴露于性接受性雌性大鼠期间,多巴胺在伏隔核中的传播增加。 Brain Res。 618, 41–46. doi: 10.1016/0006-8993(93)90426-N
Westernik,BHC,De Boer,P.,De Vries,JB和Long,S。(1998)。 抗精神病药物对大鼠大脑内侧前额叶皮质中多巴胺和去甲肾上腺素的释放产生类似的作用。 欧元。 J. Pharmacol。 361, 27–33. doi: 10.1016/s0014-2999(98)00711-0
Willig,F.,M'Harzi,M.,Bardelay,C.,Viet,D。和Delacour,J。(1991)。 罗曼菌株作为研究工作记忆的心理遗传模型:行为和生化数据。 药理学。 生物化学。 Behav。 40, 7–16. doi: 10.1016/0091-3057(91)90313-q
关键词:性行为,多巴胺,去甲肾上腺素,mPFC,微透析,RHA和RLA大鼠
引用:Sanna F,Bratzu J,Piludu MA,Corda MG,Melis MR,Giorgi O和Argiolas A(2017)多巴胺,去甲肾上腺素和罗马高低避孕雄性大鼠性行为的差异:内侧前额叶皮层的微透析研究。 面前。 Behav。 神经科学。 11:108。 doi:10.3389 / fnbeh.2017.00108
收到:22十二月2016; 接受:19 May 2017;
发布时间:07 June 2017。
