Front Behav Neurosci。 2015可能是19; 9:128。 doi:10.3389 / fnbeh.2015.00128。 eCollection 2015。
Andreatta M1, 保利P.1.
抽象
在经典调理中,最初的中性刺激(调理刺激,CS)与生物学上的显着事件(非调理刺激,US)相关,可能是疼痛(平均调理)或食物(开胃调理)。 经过几次联系后,CS可以分别发起防御或完善的响应。 与厌恶性调理相反,尽管对正常和病理行为(例如,肥胖,成瘾)的重要性,食欲性调理在人类中很少被研究。 本研究旨在以美国为食,通过动物对人类的食性条件进行动物实验。 上午8点至10点之间对XNUMX名参与者进行了调查,他们没有早餐,以确保他们感到饥饿。 在两个获取阶段中,一种几何形状(avCS +)可以预测令人反感的US(痛苦的电击),另一种形状(appCS +)可以预测令人食欲的US(根据参与者的喜好,巧克力或咸椒盐脆饼),而第三种形状(CS )均未预测美国。 在灭绝阶段,这三个形状加上一个新颖的形状(NEW)再次出现,而没有交付美国。 价和唤醒等级以及惊吓和皮肤电导(SCR)反应被收集作为学习指标。 我们发现成功的厌恶和开胃条件。 一方面,与CS-相比,avCS +被认为具有更大的消极作用和更强的诱发力,并引起惊吓增强和SCR增强。 另一方面,appCS +的评级比CS-更强,并且引起惊吓衰减和更大的SCR。 总之,我们通过展示食欲学习和正常厌恶学习成功地证实了(饥饿的)人类的动物发现。
介绍
预测威胁和食物对任何生物的生存至关重要。 在经典条件下(巴甫洛夫,1927),一个提示先于厌恶事件,例如温和的痛苦电击(厌恶的无条件刺激,美国),或食欲颗粒(美国食欲)等食欲事件,多次。 之后,这种刺激(现在标记为CS,CS +)分别能够引发防御或完成反应。 前一种联想学习被称为厌恶条件反射,后一种被称为食欲条件反射。
虽然对食欲事件的预测与厌食事件的预测一样重要,但食欲调节在动物中的研究显着减少(Bouton和Peck,1989; Koch等,1996; McDannald等,2011,评论见 Martin-Soelch等,2007)以及人类(Klucken等,2009, 2013; Austin和Duka,2010; Delgado等,2011; Levy和Glimcher,2011)。 缺乏研究可能是由于与厌恶模式相比,食欲范式的复杂性。 例如,当有机体饥饿时必须提供作为主要强化物的食物才能获得回报(最近的评论见 Dickinson和Balleine,1994; Clark等人,2012)。 在人类研究中,使用金钱克服了这一困难(Austin和Duka,2010; Delgado等,2011; Levy和Glimcher,2011)或色情图片(Klucken等,2009, 2013)。 然而,神经影像学研究指出,初级(即零食或饮料)和次级(即金钱)强化物激活一些常见的大脑区域(如纹状体),但也需要不同的激活模式(Delgado等,2011; Levy和Glimcher,2011).
据我们所知,只有少数人体调理研究检查了气味等原发性食欲增强物的作用(Gottfried等,2002), 水 (Kumar等人,2008),或食物(Prévost等,2012)。 在第一项研究中, 戈特弗里德等人。 (2002) 相关的中性面(CS)具有令人不快,愉快或中性的气味。 有趣的是,他们发现在眶额皮质(OFC)和腹侧纹状体中响应于食欲CS +与厌恶CS +的激活更大。 他们得出结论,OFC处理气味的价值,并参与将嗅觉(US)的情感价值转移到视觉(CS)系统。 (ventro)纹状体激活被解释为反映由食欲CS +引起的食欲CR。 在第二项研究中, 库马尔等人。 (2008) 邀请他们的参与者(健康对照和严重抑郁症患者)在清晨进入实验室并要求他们在夜间禁止饮酒,以确保他们在扫描前口渴。 分形图片是CS和美国的0.1毫升水。 有趣的是,在其他激活中,健康参与者(但不是抑郁症患者)表现出腹侧纹状体对食欲性CS +的更大激活,表明这种刺激被视为有益的。 在第三项研究中, Prévost等。 (2012) 根据参与者的喜好,分形图片也以CS表示,甜食或咸味小吃以US表示。 CS展示了6秒钟,在最后一秒钟内还展示了食物图片。 每当出现食物图片时,实验者就会将一块食物放在参与者的手中,参与者可以立即食用小吃。 不幸的是,作者没有在经典的调理阶段报告大脑对活化的CS +的激活。 然而,他们发现,与未获得奖励的CS–相比,获得奖励的CS +的心脏减速更少,这表明了不同的调节作用。
缺乏使用惊吓反应作为食欲调理指标的研究令人惊讶,特别是考虑到它在厌恶条件反射中的广泛应用。 惊吓反应是针对突发,意外和强烈厌恶事件的祖先和自动防御反应(Koch,1999)。 这种防御反应是由一个相对简单的神经通路介导的,涉及耳蜗根神经元,网状结构的尾椎桥核(PnC)和脊柱运动神经元(Fendt和Fanselow,1999; Koch,1999)。 动物研究表明,惊恐增强取决于杏仁核到PnC的预测(Fendt和Fanselow,1999; Koch,1999惊厥衰减依赖于完整的伏隔核(NAcc, Koch,1999)。 这种惊吓反应的调制是对所呈现的前景刺激的效价的有用的隐含测量。 因此,增强表示负价,而减弱表示正价,两者都不受认知过程的极大影响(哈姆和维克,2005; Andreatta等人,2010)。 据我们所知,只有一项动物研究通过测量惊吓反应作为依赖性测量来研究食欲调节。 有趣的是,检查的大鼠显示出与蔗糖溶液递送相关的CS +(即光)的惊吓衰减。 值得注意的是,这些衰减在NAcc病变的动物中受损,但在杏仁核损伤的动物中则没有,这表明NAcc在引发食欲CR和减弱惊恐反应方面起着特殊作用(Koch等,1996).
根据我们的知识,目前的研究首先将这种食欲调节范式转化为人类,使用初级增强剂作为美国,即甜食(巧克力巧克力)®)或咸(小咸椒盐脆饼)食物,以及作为CRs量度的惊吓调制。 我们期望appCS +触发强烈的食欲CR,与其他刺激(即avCS +和CS-)相比,反映在惊吓衰减,增强的SCR和正价等级中。
材料和方法
参与者成员
四十二名志愿者接受了参加研究并获得了学分。 有5名参与者被排除在分析之外,其中两名是由于技术问题,三名是因为他们被编码为无反应者(平均惊吓幅度<2μV),还有四名是因为他们在每种情况下都没有表现出足够的惊吓反应(最低= 33;对于详细信息,请参见材料和方法。 最后,我们分析了16位参与者(22.09位男性;平均年龄:XNUMX岁, SD:2.84; 范围:18-29年)。 四名参与者不是土生土长的德国人,六名是左撇子。 在整个实验过程中,一位参与者仍未意识到CSs-USs协会(参见程序),但我们决定不排除该参与者,因为他的回答是正常的,并且不影响结果。
材料
无条件刺激(美国)
使用了两种美国。 作为厌恶美国,我们对参与者的非优势前臂施加了轻度痛苦的电击。 通过9 mm直径和间距30 cm的两个电极传递电击。 电刺激由脉冲刺激组成,其频率为50Hz,持续时间为200ms,由电流刺激器(Digitimer DS7A,Digitimer Ltd,Welwyn Garden City,UK,400 V,max of 9.99 mA)产生。 通过前面描述的阈值程序单独确定电击的强度(Andreatta等人,2010)。 简而言之,参与者在0.5 mA的间隔内经历了两个系列的上升和下降强度。 他们不得不在视觉范围内对每个刺激进行评分,范围从0(一无所有)到10(非常剧烈的疼痛),4作为阈值的锚点(只是明显的疼痛)。 电刺激的平均强度为2.12 mA(SD = 0.56)它被评为痛苦(M = 6.45, SD = 1.73)。 食欲美国由巧克力(Smarties)组成®)或小椒盐脆饼。 如初步面试中所述,选择具有吸引力的美国取决于参与者的个人偏好。 即,参与者必须报告他们在早餐期间通常吃咸的还是甜的食物。 他们还可以自由选择在实验过程中自己喜欢巧克力还是咸椒盐脆饼。 最后,有22位参与者选择了巧克力,有11位参与者选择了小椒盐脆饼。
条件刺激(CS)
具有8 cm对角线的几何形状(蓝色方形,黄色圆形,绿色三角形,红色六边形)呈现为CS。 这些形状呈现在8的黑色计算机屏幕中间。 一种形状(avCS +)总是与厌恶的美国(痛苦的电击)相关联,一种形状(appCS +)总是与食欲的美国(巧克力或咸椒盐脆饼)相关联,一种形状(CS-)从未与厌恶相关联。或者是食欲的美国,并且第四种形状(NEW)在灭绝阶段呈现,但在收购阶段没有,以确保其中立性。
惊吓探针
持续时间为103 ms的50 dB白噪声用作惊吓探头。 声音刺激通过耳机通过双耳发出,并且在形状发作后4-6 s内随机发生。
问卷调查
在实验之前和之后,参与者必须填写德国版的状态 - 特质焦虑量表(STAI, Laux等人,1981)和积极的负面影响时间表(PANAS, Krohne等,1996)。 STAI是一种评估参与者特征和/或状态焦虑的清单,由20个特征和状态版本的项目组成。 参与者之前的焦虑水平(M = 37.06, SD = 7.80)之后(M = 39.33, SD = 9.16)实验没有显着变化[t(32) = 1.61, p = 0.117]。 当前样本中的特质焦虑评分介于20和58之间(M = 36.6, SD = 8.98),与公布的正常成人范围相当(Laux等人,1981)。 PANAS(Krohne等,1996)是积极和消极情绪的指标。 在积极情绪量表(PAS)上得分高的人容易产生诸如热情之类的情绪,而在负面情绪量表(NAS)上得分高的人容易产生诸如痛苦之类的情绪。 每个项目都包含一个形容词,参与者在从1(非常轻微)到5(极度)的范围内指示该形容词在那个特定时刻反映他们的感受的程度。 实验结果显示,参与者的负面影响没有显着差异[开始: M = 11.67, SD = 2.29; 结束: M = 12.88, SD = 4.69; t(32) = 1.55, p = 0.130]。 不知何故,参与者从一开始就大大改变了他们的积极情绪(M = 26.72, SD = 4.70)到最后(M = 23.66, SD = 7.02)实验[t(31) = 3.11, p = 0.004]。 参与者积极情绪的下降可能与范式的不愉快有关(出现了痛苦的电击以及厌恶的白噪声)。
程序
抵达实验室后,参与者阅读并签署了维尔茨堡大学心理学系伦理委员会批准的知情同意书。 他们没有被告知CS和美国之间的意外情况。 在填写问卷后,连接电极并如上所述进行疼痛阈值程序。
在 适应阶段,四个几何形状呈现两次,试验间间隔(ITI)在18和25之间变化(平均值:21.5 s)。 在此阶段没有交付US或惊吓探针。
在采集阶段之前,每个7-15都会发送七次白噪声,以降低初始的惊吓反应性。
以下两个 收购阶段 是相同的(图 1)。 每个采集阶段包括24试验:8 CS-试验,8 avCS +试验和8 appCS +试验。 CS序列是伪随机的,唯一的限制是相同的刺激不能连续两次呈现。 值得注意的是,avCS +与闪电一起作为电击的象征而出现,而痛苦的美国则是在抵消时提供的。 appCS +以复合物的形式展示了Smarties或咸椒盐脆饼干,参与者可以从罐子里挑选Smarties或椒盐卷饼。 CS-与禁令符号一起呈现,没有美国交付。 在每种类型的三个8 CS呈现期间,在刺激开始后在4和6之间递送惊吓探针。 在ITI期间提供了三个额外的惊吓探针,以确保它们的不可预测性并减少惊吓习惯。 ITI由黑屏组成,在18和25之间变化,平均值为21.5。
图1
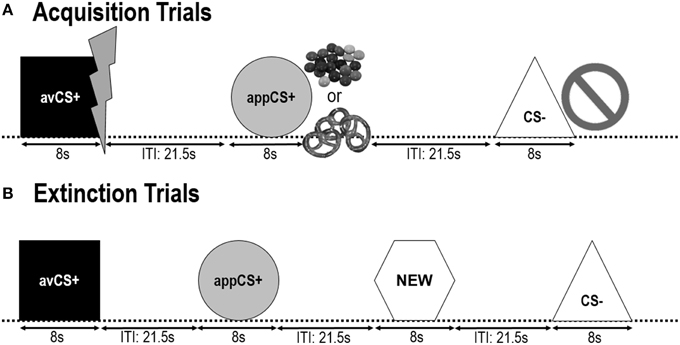
图1。 两个采集阶段(A)和灭绝阶段(B)的试验。 参与者了解到一种形状(avCS +)预测会出现轻微的疼痛性电击,一种形状(appCS +)预测一块巧克力或一小块咸椒盐脆饼(根据他们的喜好),而第三种形状(CS-)则无法预测任何形状生物学上的显着事件。 每个形状都与描绘电击,智能/咸味椒盐脆饼的图片一起呈现,或者根据与美国的关联,没有任何形状。 在灭绝阶段,再次呈现三种几何形状,但没有交付美国。 此外,第四个几何形状(NEW)被呈现为中性对照。
在 灭绝阶段参与者再次看到了三种几何形状(即avCS +,appCS +和CS-)以及一种新颖的中性形状(NEW)。 没有美国交付,形状没有与闪光,巧克力/咸椒盐脆饼或禁令的图片一起呈现。 每个刺激以伪随机顺序呈现八次(即,相同的刺激不连续呈现两次以上),从而创建32试验。 在每种CS类型的4刺激呈现中,在8期间呈现惊恐探针刺激。 在采集和习惯阶段,ITI在18和25之间变化,并且在ITI期间4额外的惊吓探测器是不可预测的。
在每个阶段之后,参与者用从1到9的视觉模拟量表(VAS)评价CS的化合价(愉悦度)和觉醒(兴奋性)。 价值范围从“1”表示“非常不愉快”到“9”表示“非常愉快”;唤醒量表从“1”表示“平静”到“9”表示“令人兴奋”。此外,应急评级为在两个收购阶段和灭绝阶段后进行评估。 参与者看到了1的几何形状,然后他们必须指出这种形状是否与电击,巧克力/咸椒盐脆饼,没有任何东西,或者他们是否无法进行任何关联。 值得注意的是,所有参与者(除一人)都意识到收购2后的意外情况。
数据减少
用V-Amp 16放大器和Vision Recorder V-Amp Edition软件(版本1.03.0004,BrainProducts Inc.,Munich,Germany)记录生理反应。 采用1000 Hz和50 Hz陷波滤波器的采样率。 使用Brain Vision Analyzer(版本2.0; BrainProducts Inc.,Munich,Germany)进行离线分析。
惊吓反应
通过左侧的肌电图(EMG)测量惊恐反应 眼轮匝肌 肌肉有两个5 mm Ag / AgCl电极。 按照指南(Blumenthal等,2005),一个电极位于瞳孔下方,第二个电极位于横向1 cm。 将接地电极和参比电极分别放置在右侧和左侧乳突上。 在连接电极之前,皮肤轻微磨损并用酒精清洁,以保持阻抗低于10kΩ。 使用28 Hz低截止滤波器和500 Hz高截止滤波器对肌电信号进行离线滤波。 然后,对EMG信号进行整流,并应用50 ms的移动平均值。 我们在惊吓探针发作之前使用50 ms作为基线(Grillon等人,2006)。 手动对惊恐探针的反应进行评分,并将进一步分析排除具有过度基线偏移(±5μV)或运动伪影的试验。 低于5μV的惊吓反应被编码为零并考虑用于计算惊吓幅度(Blumenthal等,2005)。 总而言之,10.4%的试验被拒绝,并且在采集阶段中2惊吓反应中的最小3和在每个条件的消退阶段的4惊吓反应中的8都需要使参与者保持在分析池中。 因此,四名参与者被排除在外。 峰值振幅定义为在惊吓探针开始后的20-120 ms时间窗口期间相对于基线的最大峰值。 然后使用原始数据在受试者内标准化 z - 分数,然后是T分数,以减少个体差异的影响,并更好地检测心理过程。 对每种条件(avCS +,appCS +,CS-,NEW和ITI)平均T分数。 为了研究惊恐增强或惊恐衰减,从每种情况的惊吓反应中减去ITI惊恐反应的分数。
皮肤电导响应(SCR)
使用放置在非惯用手掌上的两个5 mm Ag / AgCl电极记录皮肤电导响应(SCR)。 使用1 Hz高截止滤波器对电流响应进行离线滤波。 SCR被定义为响应开始(刺激发作后1-3 s)与响应峰值之间的差异(以μS为单位)(Tranel和Damasio,1994; Delgado等,2011)。 在SCR的分析中未考虑含有惊恐探针的试验。 低于0.02μS的响应被编码为零。 另外五名参与者被排除在SCR分析之外,因为他们的平均SCR低于0.02μS。 对原始皮肤电导数据进行平方根变换以使分布标准化,并且对于两个采集阶段(avCS +,appCS +,CS-)和灭绝阶段(avCS +,appCS +,CS-和NEW)分别对每个条件的分数取平均值。 )。
统计分析
用SPSS for Windows(版本20.0,SPSS Inc.)分析所有数据。 对于生理反应,计算两个采集阶段和消退阶段的分离的多变量方差分析(ANOVA)。 采集阶段的ANOVA具有刺激(avCS +,appCS +,CS-)和阶段(采集1,采集2)作为受试者内因子。 灭绝阶段的ANOVA仅具有刺激(avCS +,appCS +,CS-,NEW)作为受试者内因子。 使用包含受试者内因子刺激(avCS +,appCS +,CS-和NEW)和相位的单独ANOVA分析化合价,唤醒和偶然性评级。 该因子对于化合价和唤醒评级有四个级别(T1:适应期后,T2:在第一次采集阶段后,T3:在第二次采集阶段之后,T4:在消退阶段之后),但是三个级别的应急评级(T1:之后)第一个采集阶段,T2:第二个采集阶段后,T3:灭绝后阶段)。
对于所有分析,α(α)水平设定为0.05。 效果大小报告为部分η2.
成果
每相的化合价和唤醒评级如图所示 2; 惊吓反应和SCR如图所示 3.
图2
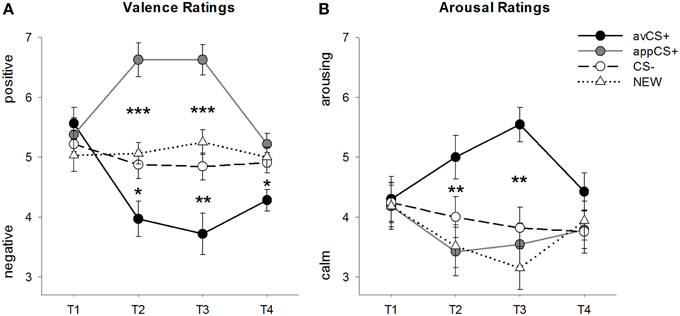
图2。 化合价(A)和唤醒(B)的评级。 线(具有标准误差)描绘了适应阶段(T1),采集1(T2),采集2(T3)和消光阶段(T4)之后的评级。 与CS-(黑色虚线)和NEW(黑色虚线)相比,厌恶CS +(黑色实线)在两个采集阶段之后获得负价和高唤醒。 重要的是,与CS-和NEW相比,食欲CS +(灰色实线)获得了正价。 *p <0.05,**p > 0.01,***p <0.001。
图3
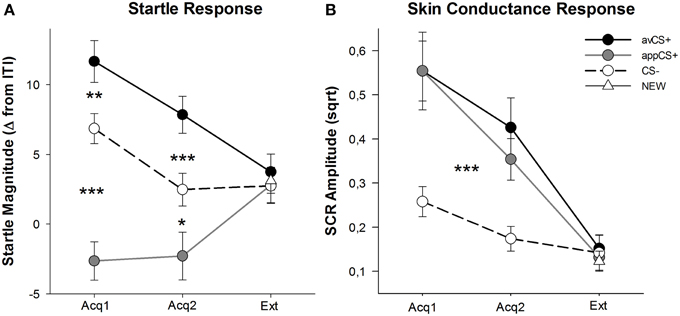
图3。 在第一个采集阶段(Acq1),第二个采集阶段(Acq2)和灭绝阶段(Ext)期间的惊吓反应(A)和皮肤电导(B)(标准误差)。 与采集阶段期间的CS-(黑色虚线)相比,惊恐响应显着增强了厌恶CS +(黑色实线)并且显着衰减到食欲CS +(灰色实线)。 与CS-相比,SCR对于avCS +和appCS +显着更大。 在灭绝阶段没有发现任何差异。 *p <0.05,**p > 0.01,***p <0.001。
评价
方差分析 价格评级 在收购期间发现刺激的重大主要影响[F(3,93) = 17.26,GG-ε= 0.801, p <0.001,部分η2 = 0.358]和阶段[F(3,93) = 3.30,GG-ε= 0.731, p = 0.039,部分η2 = 0.096]以及刺激和相位之间的显着相互作用[F(9,279) = 9.54,GG-ε= 0.463, p <0.001,部分η2 = 0.235]。 跟进 t- 测试表明实验开始时四种几何形状的化合价是相同的(ps> 0.19),而在采集1和采集2之后,avCS +被评为特别负,而appCS +被评为特别正。 具体来说,与CS– [Acq1: t(31) = 2.34, p = 0.026; Acq2: t(31) = 3.07, p = 0.004],新[Acq1: t(31) = 2.70, p = 0.011; Acq2: t(31) = 3.89, p <0.001]和appCS + [Acq1: t(31) = 5.41, p <0.001; Acq2: t(31) = 6.11, p <0.001]。 appCS +的评分比CS– [Acq1: t(31) = 4.99, p <0.001; Acq2: t(31) = 5.31, p <0.001]和新的[Acq1: t(31) = 4.92, p <0.001; Acq2: t(31) = 4.14, p <0.001]。 CS–和NEW之间的差异从未显着(ps> 0.18)。
重要的是,在比较appCS +与巧克力和咸椒盐脆饼相关的效价时,没有发现显着差异[Acq1: t(30) = 0.03, p = 0.477; Acq2: t(30) = 0.29, p = 0.775]。
在灭绝阶段之后,与CS-相比,avCS +仍然被评为更负面[t(31) = 2.40, p = 0.023],新[t(31) = 3.40, p = 0.002]和appCS + [t(31) = 3.35, p = 0.002]。 相反,appCS +的效价与CS-没有任何不同 - [t(31) = 1.77, p = 0.086]或新[t(31) = 1.07, p = 0.293]。
方差分析 觉醒等级 收购期间发现了刺激的重要主要影响[F(3,96) = 7.07,GG-ε= 0.737, p = 0.001,部分η2 = 0.181],但不是相位[F(3,96) = 1.27,GG-ε= 0.805, p = 0.289,部分η2 = 0.038],以及刺激和相位之间的显着相互作用[F(9,288) = 4.53,GG-ε= 0.582, p = 0.001,部分η2 = 0.124]。 跟进 t- 试验表明刺激对其初始唤醒没有显着差异(ps> 0.74)。 但是,在两个采集阶段之后,对avCS +的评价比对新的[Acq1: t(32) = 2.99, p = 0.005; Acq2: t(32) = 5.97, p <0.001]和appCS + [Acq1: t(32) = 2.62, p = 0.013; Acq2: t(32) = 4.42, p <0.001],并且在第一个采集阶段之后比CS–的唤醒程度要高[t(32) = 1.96, p = 0.058],但在第二次采集阶段后更加引人注目[t(32) = 3.65, p = 0.001]。 与效价评级相反,appCS +的唤醒与CS-和NEW的唤醒没有区别(ps > 0.13)。
与化合价评级相似,CS-和NEW的唤醒没有差异(ps> 0.07)。
与化合价一样,没有显示与巧克力相关的appCS +和与椒盐脆饼相关的appCS + [Acq1: t(31) = 0.26, p = 0.797; Acq2: t(31) = 0.33, p = 0.724]。
灭绝阶段后,唤醒评级没有显着差异(ps> 0.08)。
另外 事后t- 比较适应阶段,第一和第二个采集阶段和灭绝阶段后的评级,参见补充材料。
惊吓反应
收购阶段的ANOVA回归了刺激的主要影响[F(2,64) = 49.92,GG-ε= 0.964, p <0.001,部分η2 = 0.609],但不是阶段[F(1,32) = 3.16, p = 0.085,部分η2 = 0.090]和显着的相互作用刺激×相[F(2,64) = 3.37,GG-ε= 0.875, p = 0.048,部分η2 = 0.095]。 跟进 t- 测试结果表明,与CS-相比,avCS +在第一次[t(32) = 3.27, p = 0.003]和第二[t(32) = 4.00, p <0.001]的获取阶段。 同样,在两次采集1之后,对avCS +的惊吓反应也比对appCS +的惊吓反应显着增强。t(32) = 8.20, p <0.001]和采集2 [t(32) = 5.74, p <0.001]。 重要的是,与CS–相比,appCS +的惊吓幅度在两次采集1 [t(32) = 6.34, p <0.001]和采集2 [t(32) = 2.91, p = 0.007]。 再次与评级一致,当与巧克力或咸椒盐脆饼相关时,appCS +没有显示差异惊吓反应[Acq1: t(31) = 1.04, p = 0.309; Acq2: t(31) = 0.07, p = 0.947]。 在灭绝阶段,没有发现明显的效果[F(3,96) = 0.26,GG-ε= 0.906, p = 0.833,部分η2 = 0.008]。
皮肤电导响应(SCR)
从两个采集阶段的SCR的ANOVA,主效应刺激[F(2,54) = 18.04,GG-ε= 0.908, p <0.001,部分η2 = 0.401]和阶段[F(1,27) = 20.91, p <0.001,部分η2 = 0.436]结果显着,但不是他们的互动[F(2,54) = 0.68,GG-ε= 0.637, p = 0.451,部分η2 = 0.024]。 事后t- 测试表明SCR对AVCS +的显着增加[t(27) = 6.46, p <0.001]和appCS + [t(27) = 4.84, p 与CS–相比,<0.001],而参与者的SCR与avCS +和appCS + [t(27) = 0.64, p = 0.527]。 值得注意的是,没有发现SCR与巧克力appCS +和咸椒盐饼干appCS +的差异[Acq1: t(26) = 2.55, p = 0.120; Acq2: t(26) = 1.29, p = 0.210]。 对于评级和惊吓反应都是如此,对于灭绝阶段没有发现显着影响[F(3,81) = 0.28,GG-ε= 0.634, p = 0.743,部分η2 = 0.010]。
讨论
这项研究的目的是通过使用经典的食欲调节范例和主要增强剂作为无条件刺激来将动物发现转化为人类(美国)。 为了做到这一点,参与者一大早就来到实验室,没有早餐,以确保他们饿了,根据他们的喜好,巧克力或咸椒盐脆饼被用作食欲的美国人。 在采集阶段,一个几何形状(avCS +)与轻度痛苦的电击(厌恶的美国),另一个形状(appCS +)与食欲的美国和第三种形状(CS-)相关联,既没有食欲美国也没有厌恶美国。 结果表明在明确的言语水平(即评级),隐性行为水平(即惊吓反应)和生理水平(即SCR)上成功地进行厌恶和食欲调节。 具体而言,avCS +与CS相比,引起更多的负价评级,更高的唤醒评级,惊吓增强和更高的SCR。 最重要的是,appCS +与CS触发的更高正价值,惊吓衰减和更高的SCR相比。 我们关于厌恶条件反射的研究结果符合预期,因为之前的研究发现预测威胁的刺激(avCS +)被评为厌恶,引发更大的恐惧反应,并增加生理唤醒(Fendt和Fanselow,1999; 哈姆和维克,2005; Andreatta等人,2010, 2013)。 我们关于食欲调节的结果也与先前的人类和动物研究一致,表明刺激预测奖励(appCS +)被评为阳性,抑制恐惧反应,并增加生理唤醒(Koch等,1996; Gottfried等,2002; Kumar等人,2008; Klucken等,2009, 2013; Austin和Duka,2010; Prévost等,2012)。 据我们所知,这是第一项研究,证明人类的条件性惊吓衰减对预测主要奖励的刺激。 重要的是,我们能够转移并确认动物研究的结果(Koch等,1996)。 这项动物研究表明,大鼠的惊恐衰减取决于NAcc(腹侧纹状体的一部分)到PnC的预测。 因此,我们研究中的惊吓衰减可能意味着NAcc活动,这也与fMRI结果一致(Gottfried等,2002; Kumar等人,2008; Klucken等,2009, 2013; Delgado等,2011; Levy和Glimcher,2011)。 因此,我们得出结论,我们的食欲条件范式是成功的,因为显性(评级)和隐含(惊吓衰减)正价指示。
除了这个新的但相当可预测的发现之外,另外两个有趣的结果值得一提。 首先,对appCS +的言语和生理唤醒反应分离。 其次,与厌恶CR相比,我们发现食欲CR的消退速度更快。
SCR指示交感神经激活,其增加至奖励相关刺激(appCS +)和威胁相关刺激(avCS +)。 该结果与先前的调理研究一致,其中色情图片被用作食欲美国(Klucken等,2013),以及一项研究揭示可比较的SCR与预测货币的提示和预测厌恶噪音的提示(Austin和Duka,2010)。 值得注意的是,SCR是与交感神经系统的激活相关的定向响应。 这种反应被认为反映了对动机突出事件的行为反应的准备(布拉德利,2009)。 基于此,我们认为威胁相关刺激和奖励相关刺激都引发了对连续行为反应的预备反应。 换句话说,食物和疼痛信号分别开始准备接近和避免行为。 与高生理唤醒相反,言语反应表明对奖励相关刺激的唤醒较低。 言语唤醒可能更多地受到美国唤起性质的影响,而不是生理激活 本身。 事实上,我们认为appCS +与avCS +的评价相对较低而非高度唤起,因为它们分别与低迷和高度兴奋的美国联系在一起。 不幸的是,我们没有收集美国的唤醒评级,因此未来的研究将明确地检验这一假设。
在灭绝阶段,没有美国交付。 这可能会引发一种新的抑制性学习,称为灭绝(对于评论,请参阅 Milad和Quirk,2012因此,厌恶性和食欲性CR均下降。 显然,在行为(即惊吓反应)和生理(即SCR)水平上,不再能检测到对avCS +,appCS +和CS-的差异反应。 类似地,在消退阶段评估的条件刺激的明确觉醒在低水平均衡,表明成功的消退学习。 然而,avCS +的评级仍显着高于CS-,而appCS +效价与CS-价不再相同。 厌恶的明确反应的消失可能是由于进化保守主义,这意味着威胁信号特别难以忘记,因为对威胁信号的不响应可能会危及生命。
还有一个问题是:为什么在灭绝阶段,惊恐反应(即隐性价)完全消失? 首先,这些响应是在整个阶段计算的。 因此,可以想象在灭绝阶段的第一次试验期间仍然可以检测到辨别性CR。 以探索的方式,我们遵循这个假设,并在整个灭绝阶段查看惊吓反应和SCR(参见补充材料)。 虽然我们没有发现显着差异,但我们观察到与CS-和新型对照刺激相比,avCS +的惊吓幅度略高。 我们还观察到与灭活阶段的CS-和新型对照刺激相比,appCS +的惊吓衰减略微增加,然而,在几次试验中消失了。 在2nd灭绝试验之后,SCR到appCS +已经下降,而与SCR相比,SCR到avCS +几乎全部消失了。 虽然我们应该非常谨慎地解释这些结果,但是惊人的反应似乎与效价等级相似。 此外,惊吓反应和SCR都进一步支持了对威胁刺激的进化保守主义的观点。
最后,我们必须承认这项研究的一些局限性。 首先,由于技术问题,我们无法报告巧克力和咸椒盐脆饼的愉悦(和唤醒)评级。 然而,食欲性CRs表明参与者确实经历了两个美国的食欲。 其次,厌恶和美国食欲的持续时间大不相同。 因此,痛苦的电击是在avCS +的偏移处传递的,并且恰好持续了200毫秒,而巧克力和咸椒盐饼干在appCS +发病后大约在2时被呈现给参与者,并且持续时间是不可确定的,因为它取决于多快个人吃了他们。 以这种方式提供食欲美国的选择是基于先前对人类的研究(Prévost等,2012)。 然而,在方法上更优雅的是,提供一个比厌恶的美国更具可比性的美食,例如在appCS +偏移时喝一口果汁或姜汁。 为了弥补这种巨大的感知差异,我们提出了几何形状以及象征美国的视觉刺激。 第三,我们不能完全排除在灭绝阶段迅速消失的生理反应是由于方法学方面的可能性。 实际上,这一阶段的视觉刺激并未与美国符号一起呈现,就像采集阶段的情况一样。 单独介绍CS可能会影响CR,因此在第一次灭绝试验中没有明显的差异。 然而,我们的快速反应与先前研究中的熄灭反应一致,其中CS +(面部)在获得期间呈现在与美国的化合物(尖叫)中,但在灭绝期间不存在(Lissek等,2008).
总之,我们发现成功的厌恶和食欲条件反应分别与威胁和与奖励相关的刺激相关的刺激。 有趣的是,显性(评级)和隐性(惊恐反射)水平的反应以协同方式起作用,因为avCS +被报告为阴性和诱导的惊恐增强,并且appCS +被报告为阳性和诱导的惊恐衰减。 此外,appCS +的显性(评级)和生理(SCR)唤醒分离,反映了两个不同的过程。
利益冲突声明
作者声明,研究是在没有任何可被解释为潜在利益冲突的商业或金融关系的情况下进行的。
致谢
这项工作得到了协作研究中心“恐惧,焦虑和焦虑症”的支持,SFB-TRR 58项目B1。
补充材料
本文的补充材料可在以下网址找到: http://journal.frontiersin.org/article/10.3389/fnbeh.2015.00128/abstract
参考资料
Andreatta,M.,Mühlberger,A.,Glotzbach-Schoon,E。和Pauli,P。(2013)。 疼痛可预测性逆转了与缓解相关的刺激的价态评级。 面前。 SYST。 神经科学。 7:53。 doi:10.3389 / fnsys.2013.00053
PubMed摘要 | CrossRef全文 | Google Scholar
Andreatta,M.,Mühlberger,A.,Yarali,A.,Gerber,B。和Pauli,P。(2010)。 人类疼痛缓解学习后隐性和显性条件价之间的裂痕。 PROC。 生物学。 科学。 277,2411-2416。 doi:10.1098 / rspb.2010.0103
PubMed摘要 | CrossRef全文 | Google Scholar
Austin,AJ和Duka,T。(2010)。 巴甫洛夫条件下对食欲和厌恶结果的关注机制。 Behav。 Brain Res。 213,19-26。 doi:10.1016 / j.bbr.2010.04.019
PubMed摘要 | CrossRef全文 | Google Scholar
Blumenthal,TD,Cuthbert,BN,Filion,DL,Hackley,S.,Lipp,OV和van Boxtel,A。(2005)。 委员会报告:人类惊恐眨眼肌电图研究指南。 心理生理学 42,1-15。 doi:10.1111 / j.1469-8986.2005.00271.x
PubMed摘要 | CrossRef全文 | Google Scholar
Bouton,ME和Peck,CA(1989)。 环境对食欲调理准备中的调理,消退和恢复的影响。 动画。 学习。 Behav。 17,188-198。 doi:10.3758 / BF03207634
布拉德利,MM(2009)。 自然选择性注意:定向和情感。 心理生理学 46,1-11。 doi:10.1111 / j.1469-8986.2008.00702.x
PubMed摘要 | CrossRef全文 | Google Scholar
Clark,JJ,Hollon,NG和Phillips,PEM(2012)。 巴甫洛夫的学习和决策评估系统。 CURR。 奥平。 神经生物学。 22,1054-1061。 doi:10.1016 / j.conb.2012.06.004
PubMed摘要 | CrossRef全文 | Google Scholar
Delgado,MR,Jou,RL和Phelps,EA(2011)。 具有初级和次级增强物的人类厌恶调节的神经系统。 面前。 神经科学。 5:71。 doi:10.3389 / fnins.2011.00071
PubMed摘要 | CrossRef全文 | Google Scholar
Dickinson,A。和Balleine,B。(1994)。 目标导向行动的动机控制。 动画。 学习。 Behav。 22,1-18。 doi:10.3758 / BF03199951
Fendt,M。和Fanselow,MS(1999)。 条件性恐惧的神经解剖学和神经化学基础。 神经科学。 Biobehav。 启。 23,743-760。 doi:10.1016 / j.ijpsycho.2012.09.006
PubMed摘要 | CrossRef全文 | Google Scholar
Gottfried,JA,O'Doherty,J.和Dolan,RJ(2002)。 使用与事件相关的功能磁共振成像研究了人类的食欲和厌恶嗅觉学习。 J.Neurosci。 22,10829-10837。 在线提供: http://www.jneurosci.org/content/22/24/10829.full
Grillon,C.,Baas,JM,Cornwell,B。和Johnson,L。(2006)。 虚拟现实环境中的上下文调节和行为规避:可预测性的影响。 生物学。 精神病学 60,752-759。 doi:10.1016 / j.biopsych.2006.03.072
PubMed摘要 | CrossRef全文 | Google Scholar
Hamm,AO和Weike,AI(2005)。 恐惧学习和恐惧调节的神经心理学。 诠释。 J. Psychophysiol。 57,5-14。 doi:10.1016 / j.ijpsycho.2005.01.006
PubMed摘要 | CrossRef全文 | Google Scholar
Klucken,T.,Schweckendiek,J.,Merz,CJ,Tabbert,K.,Walter,B.,Kagerer,S.,et al。 (2009)。 获得条件性性唤起的神经激活:应变意识和性别的影响。 J.性别。 地中海。 6,3071-3085。 doi:10.1111 / j.1743-6109.2009.01405.x
PubMed摘要 | CrossRef全文 | Google Scholar
Klucken,T.,Wehrum,S.,Schweckendiek,J.,Merz,CJ,Hennig,J.,Vaitl,D.,et al。 (2013)。 5-HTTLPR多态性与食欲调节期间血流动力学反应的改变有关。 哼。 Brain Mapp。 34,2549-2560。 doi:10.1002 / hbm.22085
PubMed摘要 | CrossRef全文 | Google Scholar
Koch,M。(1999)。 惊吓的神经生物学。 PROG。 神经生物学. 59, 107–128. doi: 10.1016/S0301-0082(98)00098-7
PubMed摘要 | CrossRef全文 | Google Scholar
Koch,M.,Schmid,A。和Schnitzler,H.-U。 (1996)。 伏隔核的损伤会破坏惊恐的快感减弱。 Neuroreport 7, 1442–1446. doi: 10.1097/00001756-199605310-00024
PubMed摘要 | CrossRef全文 | Google Scholar
Krohne,HW,Egloff,B.,Kohmann,C.-W。和Tausch,A。(1996)。 Untersuchungen mit einer deutschen version der“Positive and Negative Affect Schedule”(PANAS)。 Diagnostica公司 42,139-156。
Kumar,P.,Waiter,G.,Ahearn,T.,Milders,M.,Reid,I。和Steele,JD(2008)。 重度抑郁症中的时间差异奖励 - 学习信号异常。 大脑 131,2084-2093。 doi:10.1093 / brain / awn136
PubMed摘要 | CrossRef全文 | Google Scholar
Laux,L.,Glanzmann,P.,Schaffner,P。和Spielberger,CD(1981)。 Das State-Trait Angstinventar。 Weinheim:Beltz测试。
Levy,DJ和Glimcher,PW(2011)。 比较苹果和橘子:在大脑中使用奖励特定和奖励一般的主观价值表示。 J.Neurosci。 31,14693-14707。 doi:10.1523 / JNEUROSCI.2218-11.2011
PubMed摘要 | CrossRef全文 | Google Scholar
Lissek,S.,Levenson,J.,Biggs,AL,Johnson,LL,Ameli,R.,Pine,DS,et al。 (2008)。 在社交焦虑障碍中,恐惧条件升级为社会相关的无条件刺激。 上午。 J.精神病学 165,124-132。 doi:10.1176 / appi.ajp.2007.06091513
PubMed摘要 | CrossRef全文 | Google Scholar
Martin-Soelch,C.,Linthicum,J。和Ernst,M。(2007)。 食欲调节:神经基础和对精神病理学的影响。 神经科学。 Biobehav。 启。 31,426-440。 doi:10.1016 / j.neubiorev.2006.11.002
PubMed摘要 | CrossRef全文 | Google Scholar
McDannald,MA,Lucantonio,F.,Burke,KA,Niv,Y。和Schoenbaum,G。(2011)。 腹侧纹状体和眶额皮质都是基于模型的,但不是无模型的强化学习所必需的。 J.Neurosci。 31,2700-2705。 doi:10.1523 / JNEUROSCI.5499-10.2011
PubMed摘要 | CrossRef全文 | Google Scholar
Milad,MR和Quirk,G。(2012)。 害怕灭绝作为转化神经科学的典范:十年的进步。 Annu。 Rev. Psychol。 63,129-111。 doi:10.1146 / annurev.psych.121208.131631
PubMed摘要 | CrossRef全文 | Google Scholar
巴甫洛夫,IP(1927)。 条件反射:大脑皮层生理活动的调查。 伦敦:牛津大学出版社。
Prévost,C.,Liljeholm,M.,Tyszka,JM和O'Doherty,JP(2012)。 神经和相关的人类杏仁核亚区域内特定和一般的巴甫洛夫到仪器的转移:高分辨率fMRI研究。 J.Neurosci。 32,8383-8390。 doi:10.1523 / jneurosci.6237-11.2012
PubMed摘要 | CrossRef全文 | Google Scholar
Tranel,D。和Damasio,H。(1994)。 皮肤皮肤电导反应的神经解剖学相关性。 心理生理学 31, 427–438. doi: 10.1111/j.1469-8986.1994.tb01046.x
PubMed摘要 | CrossRef全文 | Google Scholar
关键词:经典调理,奖励,惩罚,惊恐反射,皮肤电导反应
引用:Andreatta M和Pauli P(2015)食欲与人类的厌恶调节。 面前。 Behav。 神经科学. 9:128。 doi:10.3389 / fnbeh.2015.00128
收到:11二月2015; 接受:05 May 2015;
发布时间:19 May 2015。
编辑:
Niels Birbaumer,德国图宾根大学
点评人:
蒂姆克鲁肯,Justus Liebig University Giessen,德国
亚历山德罗·安格里利,意大利帕多瓦大学
版权所有©2015 Andreatta和Pauli。 这是一份根据条款分发的开放获取文章 知识共享署名许可(CC BY)。 允许在其他论坛中使用,分发或复制,前提是原始作者或许可人被记入贷方,并且根据公认的学术惯例引用本期刊中的原始出版物。 不允许使用,分发或复制,不符合这些条款。
*通讯:Marta Andreatta,维尔茨堡大学心理学系(生物心理学,临床心理学和心理治疗),Marcusstraße9-11,D-97070Würzburg,德国, [电子邮件保护]