Front. Behuelen. Neurosci., 23. Mars 2016 | http://dx.doi.org/10.3389/fnbeh.2016.00054

 Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2,
Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2, 

- 1Iwwersetzungsfuerschungsinstitut an Institut fir Gesondheet a Biomedizinesch Innovatioun, Queensland University of Technology, Brisbane, QLD, Australien
- 2School of Biomedical Sciences, Universitéit vu Queensland, Brisbane, QLD, Australien
Déi modern Ernährung ass héich séiss ginn, wat zu onendlechen Niveauen vum Zockerverbrauch resultéiert, besonnesch bei Jugendlecher. Wärend chronesch laangfristeg Zockeropnahm bekannt ass fir zu der Entwécklung vu metabolesche Stéierungen bäizedroen, dorënner Adipositas an Typ II Diabetis, ass wéineg bekannt iwwer déi direkt Konsequenze vu laangfristeg, bingeähnlechen Zockerkonsum op d'Gehir. BWéinst Zocker kann d'Verëffentlechung vun Dopamin am Nucleus accumbens (NAc) verursaachen ähnlech wéi Drogen vu Mëssbrauch, hu mir Ännerungen an der Morphologie vun Neuronen an dëser Gehirregioun ënnersicht no kuerz- (4 Wochen) a laangfristeg (12 Wochen) Binge- wéi Saccharose Konsum mat engem intermittierenden zwee-Flaschen Choix Paradigma. Mir hunn Golgi-Cox-Faarwen benotzt fir mëttel spiny Neuronen (MSNs) aus dem NAc Kär a Schuel vu Kuerz- a laangfristeg Saccharose-konsuméierende Ratten ze imprägnéieren an dës mat alterspasste Waasserkontrolle verglach. Mir weisen datt de verlängerten Binge-ähnlechen Saccharoseverbrauch d'total dendritesch Längt vun den NAc Shell MSNs wesentlech reduzéiert huet am Verglach mat Altersmatcher Kontrollraten. Mir hunn och fonnt datt d'Restrukturéierung vun dësen Neuronen haaptsächlech aus reduzéierter distaler dendritescher Komplexitéit resultéiert. Ëmgekéiert hu mir verstäerkte Wirbelsäure bei den distale Branchebestellunge vun NAc Shell MSNs vu laangfristeg Saccharose konsuméiere Ratten observéiert. Kombinéiert beliicht dës Resultater d'neuronal Effekter vu verlängerter binge-ähnlecher Intake vun Saccharose op NAc Shell MSN Morphologie.
Aféierung
An de leschte 40 Joer ass et eng dokumentéiert Erhéijung vum Konsum vun Zocker-séisseg Getränker a Liewensmëttel mat Zousatz Zucker (Nielsen et al., 2002; Popkin, 2010; Ng et al., 2012), mat Berichter déi schätzen datt bis zu 75% vun alle Liewensmëttel a Gedrénks héich Quantitéiten un Zucker enthalen (Ford & Dietz, 2013; Bray & Popkin, 2014). Wärend dëser Period gouf et och eng gläichzäiteg Erhéijung vun der Prevalenz vun Adipositas an Typ II Diabetis, besonnesch bei Jugendlechen (Arslanian, 2002; Reiner, 2013; Dabelea et al., 2014; Fryer et al., 2014). Rezent Studien hu gewisen datt Iwwergewiicht an fettleibeg Kanner dacks héich Quantitéiten un Zousatz Zocker konsuméieren, awer de Bäitrag vun héijen Zocker enthale Diäten zu der erhéiter Heefegkeet vun Iwwergewiicht an fettleibeg Kanner bleift kontrovers (Huet, 2013; Bray & Popkin, 2014; Bucher Della Torre et al., 2015).
Wärend e wuessende Kierper vu Beweiser weist datt de Konsum vun héijen Zocker Diäten zum Deel zum Gewiichtsgewënn bei Kanner a Jugendlecher bäidroe kann (Malik et al., 2010; Te Morenga et al., 2013; Bray & Popkin, 2014), manner Opmierksamkeet gouf op negativ net-metabolesch Konsequenzen gegeben, déi aus exzessiver Zockeropnahm entstinn. Interessanterweis entstinn e puer allgemeng Verhalens- a psychologesch Musteren dacks ënner enger Ënnergrupp vun deenen, déi iwwer iessen an héich Zocker enthalen Diäten erhalen. Am meeschte bemierkenswäert sinn d'Entwécklung vun Iessstéierungen inklusiv Binge-Iessen, kombinéiert mat dem concurrenten Ufank vu psychologesche Symptomer inklusiv Mangel u Motivatioun an Depressioun (iwwerpréift an Sheehan and Herman, 2015). Zousätzlech, well Binge-Iessen Individuen dacks e Verloscht vu Kontroll an eng Onméiglechkeet hunn hir Zockeropnahm selwer ze limitéieren, ass et méiglech datt dës Verhalen entstinn als Resultat vun neurologeschen Adaptatiounen an Gehirregiounen, déi den hedonesche Wäert vun héich schmaachtem Iessen evaluéieren. (Saper et al., 2002; Lutter & Nestler, 2009; Kenny, 2011). Dës Begrënnung gëtt och ënnerstëtzt vu Beweiser bei Mënschen, déi weisen datt Zocker a Séissegkeet Verlaangen verursaache kënnen, déi ähnlech sinn wéi déi, déi duerch Suchtfaktoren wéi Alkohol an Nikotin induzéiert sinn (Volkow et al., 2012).
Och wann d'süchteg Eegeschafte vum Zocker nach ëmmer spekulativ sinn, sinn dës Observatioune kombinéiert mat Studien, déi de Bäitrag vun der exzessiver Zockeropnahm zu Verännerungen am Belounungskreeslaf an d'Entwécklung vun Suchtfaarf-ähnlechen Verhalen an emotionalen Zoustand an Déieremodeller weisen. (Avena et al., 2008; Benton, 2010; Ventura et al., 2014), garantéiert de Besoin fir weider Enquête. Virdrun Studien bei Nager hunn gewisen datt intermittierend Zougang zu Saccharose d'Aktivitéit vu verschiddenen Neurotransmitter am mesolimbesche System verännert, dorënner Dopamin, Opioiden an Acetylcholin (Iwwerpréiwung an Avena et al., 2008). Binge-ähnlech Konsum vun Saccharose gouf gewisen fir Dopamin Verëffentlechung am Nucleus accumbens (NAc) ze erliichteren, ähnlech wéi Drogen vu Mëssbrauch (Avena et al., 2008). Ausserdeem hu mir gewisen datt laangfristeg Verbrauch vun Saccharose mat engem 24 h intermittierenden Zougang zwee-Flasche Choix Paradigma (Simms et al., 2008) moduléiert den nikotineschen Acetylcholin Rezeptor (nAChR) Ausdrock am NAc (Shariff et al., Press). Interessanterweis hu mir och beobachtet datt nAChR Verbindunge bekannt fir Dopamin- an Acetylcholinaktivitéit am NAc ze moduléieren, verschidden Effekter op Saccharoseverbrauch no kuerz- a laangfristeg Intake hunn (Shariff et al., Press).
Wärend dës Studien Ähnlechkeeten an de Verhalens- an neurochemesche Verännerungen bewisen hunn, déi duerch intermittierend Zougang zu Zocker an Drogen vu Mëssbrauch verursaacht ginn, ass et net bekannt ob dës Effekter d'Verännerungen an der neuronaler Morphologie am NAc erliichteren. Tseng ass am Géigesaz zu Substanze vu Mëssbrauch abegraff Kokain, Amphetamin an Nikotin déi gutt charakteriséiert Verännerungen an der Morphologie vu mëttleren spiny Neuronen (MSNs) am NAc produzéieren, inklusiv erhéicht Wirbelsäule Dicht a verännert dendritesch Komplexitéit (Robinson a Kolb, 1999, 2004; Li et al., 2003; Crombag et al., 2005). Well mir virdru gewisen hunn datt laangfristeg Belaaschtung (12 Woch) fir Alkohol a Saccharose mat der intermittéierter Zwee-Flaschenwahlparadigma eng differentiell Äntwert op pharmakotherapeutesch Interventiounen am Verglach mat kuerzfristeg Intake (4 Wochen) produzéiert; Steensland et al., 2007; Shariff et al., Press), hu mir d'Effekter vu kuerz- a laangfristeg Saccharoseverbrauch op MSN Morphologie an der NAc bewäert. Mir hunn adolescent Ratten erlaabt Saccharose op eng binge-ähnlech Manéier fir 4 (kuerzfristeg) oder 12 (laangfristeg) Wochen ze konsuméieren an duerno d'Morphologie vun NAc MSNs vu Kuerz- a laangfristeg Saccharose-Konsuméiere Ratten analyséiert an dëst verglach mat Alter ugepasste Kontrollen déi nëmmen Zougang zu Waasser kritt hunn. Eis Resultater weisen datt MSNs aus der NAc Shell geännert ginn no laang- awer net kuerzfristeg Saccharoseverbrauch, mat reduzéierter dendritesch Längt, awer erhéicht distal dendritesch Wirbelsäule Dicht. Ausserdeem hu mir fonnt datt d'Morphologie vun MSNs aus dem NAc Kär relativ intakt bliwwen ass no kuerz- a laangfristeg Saccharosekonsum. Dës Resultater markéieren eng direkt neurologesch Konsequenz vu laangfristeg Saccharosekonsum op eng binge-ähnlech Manéier. Ausserdeem beweisen dës Donnéeën d'Noutwendegkeet fir weider Studien, déi d'molekulare an neurochemesch Verännerungen opklären, déi d'morphologesch Restrukturéierung vun NAc Shell MSNs begleeden, induzéiert duerch verlängert, binge-ähnlech Saccharoseaufnahme.
Materialien an Methoden
Ethik - Erklärung
All experimentell Prozeduren goufen am Aklang mat der Australian Code fir d'Betreiung an Notzung vun Déieren fir wëssenschaftlech Zwecker duerchgefouert, 8. Editioun (National Health and Medical Research Council, 2013). D'Protokoller goufen vun der Queensland University of Technology Animal Ethics Committee an der University of Queensland Animal Ethics Committee guttgeheescht.
Déieren a Wunneng
Fënnef Woch al (adolescent) männlech Wistar Ratten (Kontroll: 176.4 ± 4.8 g; Saccharose: 178.3 ± 5.0 g) (ARC, WA, Australien), goufen individuell a gelëfter Dual Level Plexiglas gehal.® Käfeg. D'Ratten goufen 5 Deeg virum Start vun den Experimenter un déi individuell Wunnengsbedéngungen, Handhabung a Reverse-Light Zyklus acclimatiséiert. All Ratten goufen an engem klimat kontrolléierten 12-Stonnen ëmgedréint Liicht / Däischter Zyklus (Luuchten um 9 Auer aus) Raum mat Standard Ratchow a Waasser verfügbar ad libitum.
Intermittéierend-Zougang Zwee-Flaschen Choix Drénken Paradigma
Den intermittierenden Zougang 5% Saccharose Zwee-Flasche Choix Drénkparadigma (Simms et al., 2008) ugepasst gouf aus Wise (1973). All Flëssegkeete goufen an 300 ml graduéierter Plastiksfläschen mat Edelstahl-Drénkspäicher presentéiert, déi duerch zwee Grommets virun der Käfeg agefouert goufen no dem Ufank vum donkelen Liichtzyklus. Gewiichter vun all Fläsch goufen virun der Fläsch Presentatioun opgeholl. Zwou Fläschen goufen gläichzäiteg presentéiert: eng Fläsch mat Waasser; déi zweet Fläsch mat 5% (w/v) Saccharose. D'Placement vun der 5% (w/v) Saccharosefläsch gouf ofwiesselnd mat all Belaaschtung fir d'Kontroll fir Säit Virléiften. D'Flasche goufen 24 Stonnen no der Presentatioun vun de Flëssegkeete gewien, a Miessunge goufen op déi nootste 0.1 g geholl. D'Gewiicht vun all Rat gouf och gemooss fir d'Gramm vun der Saccharosezufuhr pro Kilogramm Kierpergewiicht ze berechnen. Am Dag 1 vun der Drénkzäit, Ratten (n = 6-9) kruten Zougang zu enger Fläsch vu 5% (w/v) Saccharose an enger Fläsch Waasser. No 24 Stonnen gouf d'Saccharosefläsch duerch eng zweet Waasserfläsch ersat, déi fir déi nächst 24 Stonnen verfügbar war. Dëst Muster gouf Mëttwochs a Freides widderholl. D'Ratten haten onlimitéiert Zougang zu Waasser op all aner Deeg. Binge-ähnlechen Konsum vun Saccharose huet zu enger Eskalatioun vun der Gesamt Saccharosenahme (ml) mat der Zäit gefouert (Ergänzungsbild 1) a gouf begleet vu stabile Baseline Drénkniveauen baséiert op Kierpergewiicht [20 ± 5 g / kg vun de 5% (w/v)] während der kuerzfristeg [~ 4 Wochen (13 Drénksessiounen)] a laangfristeg [ ~12 Wochen (37 Drénksessiounen)] Drénkperioden. Eng separat Grupp vu Kontroll Ratten (n = 6-9) goufen Zougang zu Waasser a béide Fläschen (dh keng Saccharose) ënner de selwechte Konditioune beschriwwen. Déi duerchschnëttlech Kierpergewiicht vu Kontroll- a Saccharose-konsuméierende Ratten um Enn vu kuerzer Beliichtung war 405.7 ± 40.8 g respektiv 426.4 ± 31.2 g. Um Enn vun der laangfristeg Belaaschtung war déi duerchschnëttlech Kierpergewiicht fir Kontroll- an Saccharosegruppen 578.8 ± 53.4 g a 600.2 ± 45.2 g.
Golgi-Cox Faarf
No der leschter Drénksessioun goufen d'Ratten vun der Déiereanlag transferéiert fir d'Veraarbechtung vun de Gehireproben an der Histologieanlag an der School of Biomedical Sciences, University of Queensland (St Lucia, Australien) z'erméiglechen. All genehmegt Moossname goufen geholl fir Stress beim Transport ze reduzéieren, duerno d'Ratten erlaabt iwwer Nuecht ze erholen. Den nächsten Dag goufen Ratten duerch Natriumpentobarbital Iwwerdosis geaffert (60-80 mg / kg, ip Vetcare, Brisbane, Australien) an intracardially perfuséiert mat ~ 300 ml kënschtlech cerebro-spinal Flëssegkeet déi enthält, (an mM): 130 NaCl, 3 KCl, 26 NaHCXNUMX31.25 NaH2PO4, 5 MgCl2, 1 CaCl2an 10 D-Glukose. All Déier gouf dunn ofgeschnidden an d'Gehir ewechgeholl an an der Däischtert an der Golgi-Cox Léisung inkubéiert déi 5% Kaliumdichromat, 5% Kaliumchromat, a 5% Quecksilberchlorid (all Chemikalien aus Sigma-Aldrich) enthält, déi 3 Deeg frësch gemaach gouf. virum Opfer wéi virdru beschriwwen (Rutledge et al., 1969). Golgi-Cox stain incubation an Post-Veraarbechtung Methoden goufen geännert aus Ranjan & Mallick (2010). Gehirer vu kuerzfristeg sucrose konsuméierend Déieren goufen fir 6 Deeg bei 37 ° C inkubéiert, wärend Gehirer vu laangfristeg sucrose konsuméierend Déieren fir 10 Deeg inkubéiert goufen, mat enger Ännerung op frësch Golgi-Cox Léisung no 4 Deeg Inkubatioun.
No der Inkubatioun goufen 300 μm Coronal Sektioune mat engem vibréierende Zeiss Hyrax V50 Mikrotom (Carl Zeiss, Däitschland) geschnidden. Scheiwen goufen dunn sequenziell an 24-Well Placke plazéiert, gefüllt mat 30% (w/v) Saccharose an 0.1 M phosphat gebufferter Salzlinn a veraarbecht wéi an (Ranjan & Mallick, 2010). Kuerz, goufen Sektiounen an 50% Ethanol fir 5 min dehydréiert, duerno an 0.1 M NH plazéiert4OH Léisung fir 30 min, zweemol mat destilléiert Waasser fir 5 min gespullt an an Fujihunt Film Fixer (Fujifilm, Singapur) fir 30 min am Däischteren gesat. D'Scheiwen goufen dann zweemol an destilléiert Waasser fir 2 min all gespullt an an 70, 90, 95, an 100% Ethanol zweemol fir 5 min all dehydréiert. D'Sektiounen goufen dunn an der CXA Léisung (1:1:1 Chloroform:Xylen:Alkohol) fir 10 min geläscht an an DPX (Sigma-Aldrich) op Superfrost Plus Rutschen (Menzel-Glaser, Lomb Scientific, Australien) montéiert an iwwerdeckt. (Menzel-Glaser, Däitschland). D'Rutschen goufen an der Däischtert gelooss fir iwwer Nuecht bei Raumtemperatur ze dréchen.
Neuronal Selektioun a Tracing am Nucleus Accumbens
Coronal Scheiwen tëscht bregma +2.8 an +1.7 goufen iwwerpréift fir MSNs am Kär an der Schuel vum NAc, mat der lateraler Ventrikel an der anteriorer Kommissioun als Landmarken mat der Hëllef vun engem Rat Gehir Atlas (Paxinos a Watson, 2007) (Bild 1). D'Konturfunktioun an Neurolucida 7 (MBF Bioscience, VT, USA) gouf benotzt fir den NAc Kär an NAc Shell an all Slice ze demarkéieren (Figure) 2). Tëscht 2 an 9 Neuronen pro Regioun pro Déier goufen fir dendritesch Längtparameter verfollegt mat engem 63x Objektiv oder fir Wirbeldensitéiten (gemellt als Wirbelen pro 100 μm) mat engem 100x Objektiv op engem Zeiss Axioskop II (Carl Zeiss, Däitschland) mat engem automatiséierte xyz Etapp gefuer vun Neurolucida® 7 Software (MBF Biosciences, VT, USA). All Tracing gouf op eng blann Manéier mat Respekt fir d'Behandlung gemaach. Morphologesch Parameter vu Golgi-Cox imprägnéierten Neuronen goufen analyséiert op eng Manéier ähnlech wéi fréier Berichter (Klenowski et al., 2015).
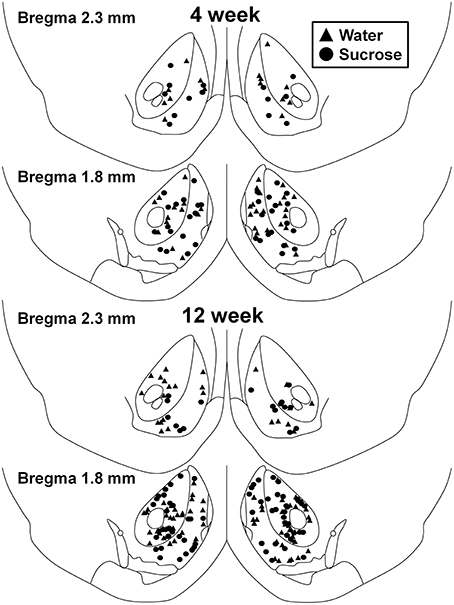
Figure 1. Kaart weist d'Plaze vu mëttleren spiny Neuronen, déi aus dem Nucleus accumbens Kär a Schuel vun 4 an 12 Wochen Saccharose konsuméierende Ratten an Alter ugepasste Kontrollen getest goufen. Top zwee Paneele weisen Plazen vun Neuronen, déi aus dem Nucleus accumbens Kär a Schuel vu 4 Woche Kontroll (Dräieck) an Saccharose (Krees) Déieren getest goufen. Ënnen zwee Paneele weisen Positiounen vun Neuronen, déi aus 12 Woche Kontroll (Dräieck) an Saccharose (Krees) Déieren gepréift goufen.
Statistesch Analyse
Moyenne an Norm Feeler vun der Moyenne (SEM) sech fir all Daten Formatioun mat der Déier als berechent n, mat der mëttlerer Morphometrie Daten aus all de Kär oder Shell NAc MSNs (n = 7 fir NAc Shell an n = 6 fir NAc Kär 4-Woch, n = 9 fir 12-Wochengruppen). Wou uginn, ongepaart zwee tailed Student's t-Tester oder Zwee-Wee ANOVAs mat Bonferroni Post-Tester goufen fir all Analysë gemaach, déi de Verglach vu Gruppmëttelen involvéiert hunn, mat GraphPad Prism Versioun 6.02 (GraphPad Software, San Diego, CA). Statistesch Bedeitung gouf ugeholl um P < 0.05. All Daten an der Resultater Rubrik sinn als Mëttel ± SEM presentéiert. Prozentsaz Ännerungen ginn als relativ zu der Kontroll Wäert berechent.
Resultater
Medium Spiny Neuronen aus der Nucleus Accumbens Shell hunn eng Ofsenkung vun dendritesch Längt, verréngert Dendritesch Komplexitéit awer erhéicht duerchschnëttlech Dicht vun der Wirbelsäule bei Distale Branch Uerder no laanger awer net kuerzfristeg Sucrose Konsum
No kuerzfristeg (4 Wochen) Saccharoseverbrauch goufen et keng bedeitend Differenzen an NAc Shell MSN morphometresche Parameteren (Tabelle) 1). Et waren och keng bedeitend Differenzen tëscht kuerzfristeg Saccharoseverbrauch a Waasserkontroll NAc Shell MSNs an Analysen am Zesummenhang mat Zentrifugalbrancheuerdnung. Nämlech dendritesch Segmenter pro Brancheuerdnung (P = 0.4111), mëttel dendritesch Längt pro Brancheuerdnung (P = 0.5581) an duerchschnëttlech Wirbelsäule pro Brancheuerdnung (P = 0.2977, zwee-Wee ANOVAs) waren net wesentlech ënnerschiddlech tëscht Gruppen. Eng Standortkaart, déi d'ongeféier Positioune vun de gepréiften Neuronen weist, gëtt an der Figur gewisen 1.
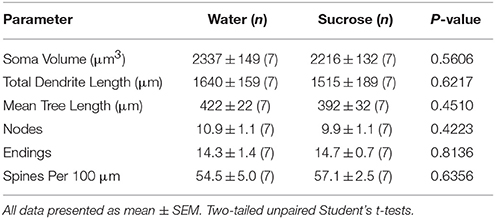
Table 1. Allgemeng morphologesch Parameter vu mëttleren spiny Neuronen aus der Nukleus accumbens Schuel vu kuerzfristeg Saccharose konsuméierende Ratten an Altersmatch Waasser Kontrollen.
No laangfristeg (12 Wochen) Saccharoseverbrauch gouf déi total dendritesch Arborlängt vun NAc Shell MSNs ëm 21% reduzéiert am Verglach mat Waasserverbrauchskontrollen (Waasser: 1827 ± 148 μm, n = 9; Saccharose 1449 ± 78 μm, n = 9,*P = 0.0384, zwee-tailed unpaired Student d' t-test, Figur 2, Table 2). Verglach vun der mëttlerer Zuel vun dendritesche Bifurkatiounen (Noden) an dendritesch Endungen tëscht de Waasser- an Saccharosegruppen huet e reduzéierten (och wann net bedeitend) Niveau vun der dendritescher Komplexitéit an NAc Shell MSNs (Noden: Waasser 12.9 ± 1.4) opgedeckt n = 9, Saccharose 10.1 ± 0.8 n = 9, P = 0.0879; Endungen: Waasser 17.9 ± 1.4 n = 9, Saccharose 14.8 ± 0.7 n = 9, P = 0.0657, zwee-tailed unpaired Student d' t-Test, Dësch 2). Et gouf keng Ännerung am Soma Volumen (P = 0.9400), mëttel dendritesch Bamlängt (P = 0.1646) oder total Wirbelsäule Dicht (P = 0.3662) an NAc Shell MSNs vu laangfristeg Saccharose konsuméiere Ratten am Verglach mat Waasserkontrollen. Dës morphometresch Parameter sinn an der Tabell detailléiert 2.
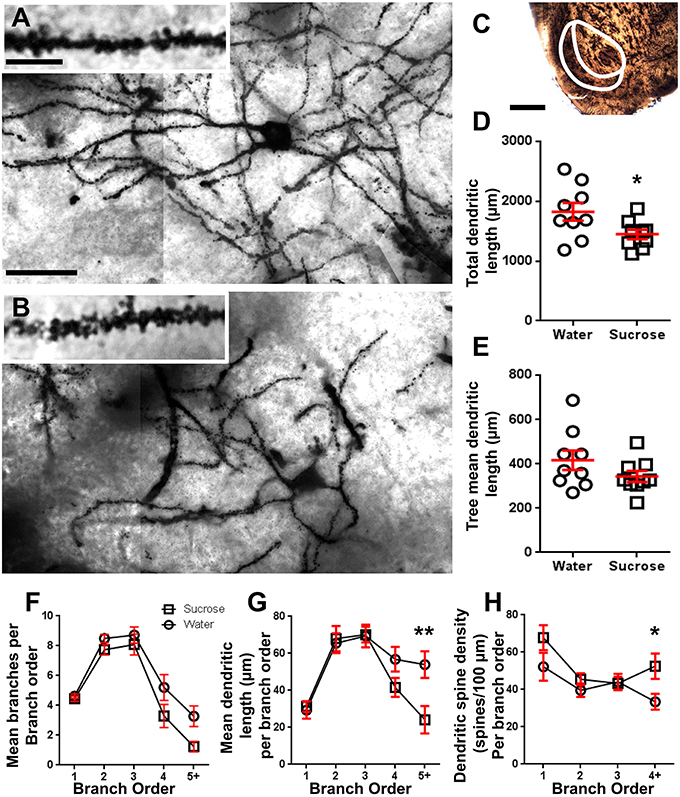 Figure 2. Verréngert dendritesch Arborlängt a verstäerkter distaler dendritesch Wirbelsäule Dicht vu mëttleren spiny Neuronen (MSNs) aus der Nucleus accumbens (NAc) Schuel vu laangfristeg sucrose behandelt Ratten am Verglach mat Kontrollraten. (A,B) weisen Representatioune vu Kontroll (uewen) a laangfristeg (12 Wochen) Saccharose (ënnen) behandelt Brightfield z-Stack Mosaik vu Golgi-imprägnéierten MSNs aus der NAc Shell (63x Vergréisserung). Inset vun (A, B) weist Kontroll a laangfristeg sucrose behandelt hellfeld Biller vu Golgi-impregnéierten MSN Dendriten an dendritesch Wirbelen aus der NAc Shell (100x Vergréisserung). (C) weist d'anatomesch Regiounen aus deenen d'MSNs an dëser Etude gepréift goufen. (D) weist e Scatter-Plot vu verréngert Gesamt MSN dendritesch Arbor (Moyenne ± SEM) vun der NAc Shell a laangfristeg Saccharose Déieren (Plaze) am Verglach mat Kontrollen (Krees), onpaarte Studenten t-test, *P <0.05, n = 9; kontrolléieren an n = 9; 12 Wochen Saccharose. (E) weist e Scatter-Plot vun onverännert mëttlerer MSN dendritescher Bamlängt (Moyenne ± SEM) vun der NAc Shell a laangfristeg Saccharose Déieren (Plaze) am Verglach mat Kontrollen (Krees), onpaarte Studenten t-test, P > 0.05, n = 9; kontrolléieren an n = 9; 12 Wochen Saccharose. Branch Uerdnung Analyse (Moyenne ± SEM) vun dendritic Segment Zuel pro Branche Uerdnung (F), heescht dendritesch Längt pro Brancheuerdnung (G) an dendritesch Wirbelsäule Dicht pro Brancheuerdnung (H). Laangfristeg Saccharoseverbrauch reduzéiert dendritesch Längt bei distale Branchebestellungen (5+) a erhéicht dendritesch Wirbelsäule Dicht bei distale Branchebestellungen (4+) am Verglach mat Kontrollen (G,H), Zwee-Wee ANOVAs mat Bonferroni Post-Tester, *P <0.05, **P <0.01, n = 9; kontrolléieren an n = 9; laangfristeg Saccharose. Skala Baren: (A, B) = 20 μm; inset vun (A, B) = 10 μm; (C) = 1 mm.
Figure 2. Verréngert dendritesch Arborlängt a verstäerkter distaler dendritesch Wirbelsäule Dicht vu mëttleren spiny Neuronen (MSNs) aus der Nucleus accumbens (NAc) Schuel vu laangfristeg sucrose behandelt Ratten am Verglach mat Kontrollraten. (A,B) weisen Representatioune vu Kontroll (uewen) a laangfristeg (12 Wochen) Saccharose (ënnen) behandelt Brightfield z-Stack Mosaik vu Golgi-imprägnéierten MSNs aus der NAc Shell (63x Vergréisserung). Inset vun (A, B) weist Kontroll a laangfristeg sucrose behandelt hellfeld Biller vu Golgi-impregnéierten MSN Dendriten an dendritesch Wirbelen aus der NAc Shell (100x Vergréisserung). (C) weist d'anatomesch Regiounen aus deenen d'MSNs an dëser Etude gepréift goufen. (D) weist e Scatter-Plot vu verréngert Gesamt MSN dendritesch Arbor (Moyenne ± SEM) vun der NAc Shell a laangfristeg Saccharose Déieren (Plaze) am Verglach mat Kontrollen (Krees), onpaarte Studenten t-test, *P <0.05, n = 9; kontrolléieren an n = 9; 12 Wochen Saccharose. (E) weist e Scatter-Plot vun onverännert mëttlerer MSN dendritescher Bamlängt (Moyenne ± SEM) vun der NAc Shell a laangfristeg Saccharose Déieren (Plaze) am Verglach mat Kontrollen (Krees), onpaarte Studenten t-test, P > 0.05, n = 9; kontrolléieren an n = 9; 12 Wochen Saccharose. Branch Uerdnung Analyse (Moyenne ± SEM) vun dendritic Segment Zuel pro Branche Uerdnung (F), heescht dendritesch Längt pro Brancheuerdnung (G) an dendritesch Wirbelsäule Dicht pro Brancheuerdnung (H). Laangfristeg Saccharoseverbrauch reduzéiert dendritesch Längt bei distale Branchebestellungen (5+) a erhéicht dendritesch Wirbelsäule Dicht bei distale Branchebestellungen (4+) am Verglach mat Kontrollen (G,H), Zwee-Wee ANOVAs mat Bonferroni Post-Tester, *P <0.05, **P <0.01, n = 9; kontrolléieren an n = 9; laangfristeg Saccharose. Skala Baren: (A, B) = 20 μm; inset vun (A, B) = 10 μm; (C) = 1 mm.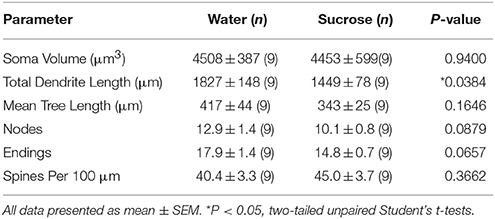
Table 2. Allgemeng morphologesch Parameter vu mëttleren spiny Neuronen aus der Nukleus accumbens Schuel vu laangfristeg Saccharose konsuméiere Ratten an Altersmatch Waasser Kontrollen.
No der Charakteriséierung vun der allgemenger dendritescher Morphologie vu laangfristeg Saccharose konsuméierende NAc Shell MSNs, hu mir dendritesch Arborisatiounen a Wirbelsdichten analyséiert wat hir Brancheuerdnungseigenschaften ugeet. Eis ëmfaassend Bewäertung vun den dendritesche Beem quantifizéiert d'Zuel vun dendritesche Segmenter pro Brancheuerdnung, déi mëttlere Längt vun dendritesch Segmenter pro Brancheuerdnung a mëttlerer Wirbelsäule pro Brancheuerdnung vun NAc Shell MSNs vu Waasserkontroll a laangfristeg Saccharose konsuméiere Ratten. E Resumé vun de Brancheuerdnungsdaten an Analyse gëtt an der Tabell presentéiert 3.
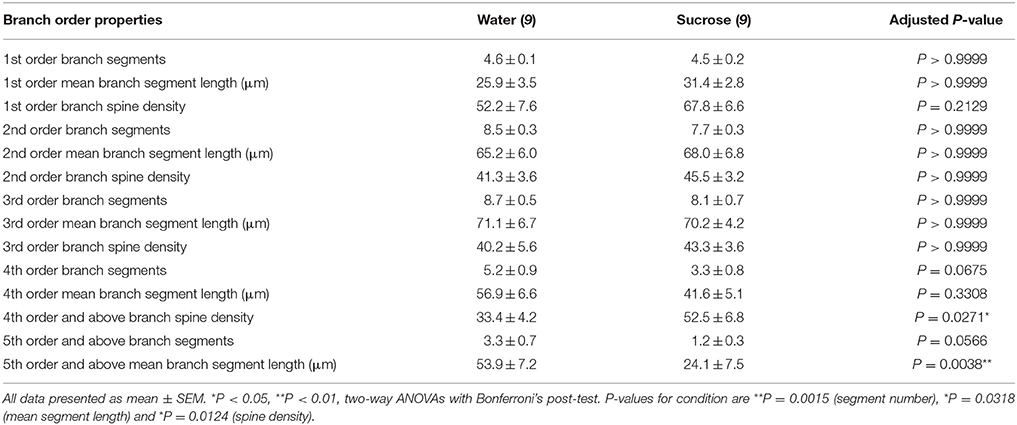 Table 3. Branch Uerdnung Charakteristiken vun mëttel- spiny Neuronen aus laangfristeg sucrose a Waasser drénken Ratten.
Table 3. Branch Uerdnung Charakteristiken vun mëttel- spiny Neuronen aus laangfristeg sucrose a Waasser drénken Ratten.Déi duerchschnëttlech dendritesch Branche Segment Zuel pro Brancheuerdnung vun NAc Shell MSNs gouf wesentlech reduzéiert a laangfristeg Saccharose konsuméiere Ratten am Verglach mat Waasserkontrollen (**P = 0.0015, zwee-Wee ANOVA). Bonferroni Post-Tester hunn en Trend zu enger reduzéierter Zuel vu Branchesegmenter um 4. (Waasser: 5.2 ± 0.9, n = 9; Saccharose 3.3 ± 0.8, n = 9, P = 0.0675, Figur 2F, Table 3), a 5. Uerdnung an iwwer Branche Uerderen (Waasser: 3.3 ± 0.7, n = 9; Saccharose 1.2 ± 0.3, n = 9, P = 0.0566, Figur 2F, Table 3). Déi duerchschnëttlech dendritesch Segmentlängt pro Brancheuerdnung vun NAc Shell MSNs gouf och wesentlech reduzéiert a laangfristeg sucrose konsuméiere Ratten am Verglach mat Waasserkontrollen (*P = 0.0444, zwee-Wee ANOVA). Bonferroni Post-Tester hunn eng Reduktioun vu 55% bei 5. n = 9; Saccharose 24.1 ± 7.5 μm, n = 9, **P = 0.0038, Figur 2G, Table 3).
Branch Order Analyse huet eng bedeitend Erhéijung vun der dendritescher Wirbelsäule Dicht vun NAc Shell MSNs vu laangfristeg Saccharose konsuméiere Ratten am Verglach mat Kontrollen gewisen (*P = 0.0124, zwee-Wee ANOVA). Bonferroni Post-Tester hunn eng Dicht vun der Wirbelsäule vun 57% bei distale 4. n = 9; Saccharose 52.5 ± 6.8, n = 9, P = 0.0271*, Inset vun Figuren 2A,B,H, Table 3). Representativ Biller vun der Gesamt MSN Architektur an distaler Wirbelsäule Dicht (Inset) sinn a Figuren duergestallt 2A, B.
Zesummegefaasst weisen dës Resultater datt de kuerzfristeg Saccharoseverbrauch wéineg Effekt op morphologesche Parameter vun MSNs an der NAc Shell huet. Wéi och ëmmer, nom längerem Konsum, gëtt et e wesentleche Réckgang vun der neuronaler Arborlängt a Komplexitéit, besonnesch an distalen dendritesche Branchen. Begleedend distaler Wirbelsäule Dicht Erhéijunge sinn och an der NAc Shell MSNs vu laangfristeg Saccharose konsuméiere Ratten anscheinend.
Medium Spiny Neuronen aus dem Nucleus Accumbens Core hunn d'Verzweigungskomplexitéit reduzéiert no laanger awer net kuerzfristeg Sucrose Konsum
No Kuerzfristeg vum Saccharoseverbrauch goufen et keng bedeitend Differenzen an NAc Kär MSN morphometresche Parameteren (Tabelle) 4). Et waren och keng bedeitend Differenzen tëscht 4-Wochen Saccharoseverbrauch a Waasserkontrollkär MSNs an Analysen am Zesummenhang mat Zentrifugalzweigeuerdnung. Nämlech dendritesch Segmenter pro Brancheuerdnung (P = 0.7717), mëttel dendritesch Längt pro Brancheuerdnung (P = 0.2096), a mëttlerer Wirbelsäule Dicht pro Brancheuerdnung (P = 0.3521, zwee-Wee ANOVAs) waren net ënnerschiddlech tëscht Gruppen.
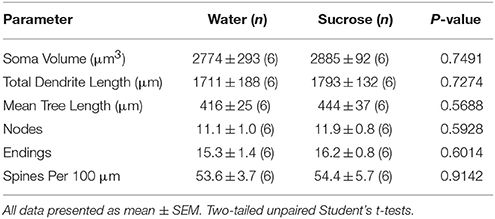 Table 4. Allgemeng morphologesch Parameter vu mëttlere spiny Neuronen aus dem Nukleus accumbens Kär vu kuerzfristeg Saccharose konsuméierende Ratten an Alter ugepasst Waasserkontrollen.
Table 4. Allgemeng morphologesch Parameter vu mëttlere spiny Neuronen aus dem Nukleus accumbens Kär vu kuerzfristeg Saccharose konsuméierende Ratten an Alter ugepasst Waasserkontrollen.Verlängert Saccharoseverbrauch hat och kee bedeitend op NAc Kär MSN morphometresche Parameteren (Tabelle 5). Déi duerchschnëttlech dendritesch Branche Segment Zuel pro Brancheuerdnung vun NAc Kär MSNs gouf wesentlech reduzéiert a laangfristeg Saccharose konsuméiere Ratten am Verglach mat Waasserkontrollen (*P = 0.0416, zwee-Wee ANOVA), awer et waren keng bedeitend Differenzen an der mëttlerer dendritescher Längt pro Brancheuerdnung (P = 0.0995) an duerchschnëttlech Wirbelsäule pro Brancheuerdnung (P = 0.4888, zwee-Wee ANOVAs) tëscht MSNs am NAc Kär vu laangfristeg Saccharose konsuméiere Ratten am Verglach mat Waasserkontrollen. Zesummegefaasst weisen eis Daten datt den NAc Kär net sou reaktiounsfäeger ass op laangfristeg Saccharosekonsum am Verglach mat MSNs aus der NAc Shell Regioun.
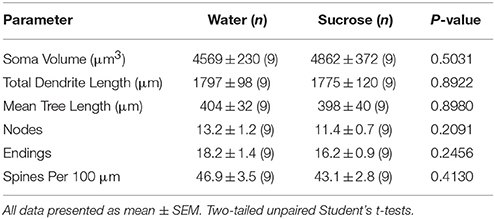 Table 5. Allgemeng morphologesch Parameter vu mëttleren spiny Neuronen aus dem Nukleus accumbens Kär vu laangfristeg Saccharose konsuméiere Ratten an Alter ugepasst Waasser Kontrollen.
Table 5. Allgemeng morphologesch Parameter vu mëttleren spiny Neuronen aus dem Nukleus accumbens Kär vu laangfristeg Saccharose konsuméiere Ratten an Alter ugepasst Waasser Kontrollen.Diskussioun
Déi verstäerkte Verfügbarkeet vun héich séiss Liewensmëttel an der westlecher Ernährung huet net nëmmen zu der verstäerkter Prevalenz an der wirtschaftlecher Belaaschtung vun Adipositas an Typ II Diabetis bäigedroen, et huet och zu engem Ufank vun Iessstéierunge gefouert wéi Binge Iessen (Schwanson et al., 2011; Kessler et al., 2013; Davis, 2015). Och wann déi süchteg Eegeschafte vun Zucker inklusiv Fruktose an Saccharose spekulativ bleiwen, gëtt et eng markant Ähnlechkeet an de Verhalens- an Neurale Korrelaten, déi sech manifestéieren als Resultat vun iwwer Iessen a längerer Drogekonsum. (Avena et al., 2008, 2011). Zousätzlech aktivéiert Zocker d'Belounungskreesser vum Gehir op eng Manéier déi ähnlech wéi Drogen vu Mëssbrauch ass (Volkow et al., 2012), a Resultater vu mënschleche Studien suggeréieren datt Zocker a Séiss Verlaangen induzéieren, déi an der Gréisst vergläichbar sinn mat deenen, déi duerch Suchtfaktoren wéi Alkohol an Nikotin induzéiert sinn. (Volkow et al., 2012). Mir hunn dofir e Modell vum Binge-Saccharose Konsum bei Ratten benotzt fir d'Effekter vu kuerzen (4 Wochen) a laangfristeg (12 Wochen) Saccharoseverbrauch op neuronal Morphologie vun MSNs an der NAc ze bestëmmen, e Schlësselkomponent vun der iwwerlappende Belounungsschaltung dat moduléiert gëtt duerch Zocker an süchteg Drogen. Mir weisen datt MSNs aus der NAc Shell vu chronesche laangfristeg Saccharose konsuméiere Ratten wesentlech dendritesch Längt a Komplexitéit erofgeholl hunn, awer d'distale dendritesch Wirbelsäule Dicht erhéicht hunn. Laangfristeg Saccharoseverbrauch hat keen Effekt op d'Morphologie vun MSNs aus dem NAc Kär, während de Kuerzfristeg Saccharose Konsum och kee signifikanten Effekt op d'MSN Morphologie vum NAc Kär oder Shell hat. Dës Resultater weisen net nëmmen en direkten Effekt vu verlängerter binge-ähnlecher Saccharose-Intake op neuronal Morphologie vun NAc Shell MSNs, awer si beliichten och déi potenziell schiedlech Konsequenze vum verlängerten Konsum vun héijen Zocker enthalen Diäten.
Den NAc, deen en Deel vum ventrale Striatum ass, besteet haaptsächlech aus MSNs, déi morphologesch charakteriséiert sinn als mëttelgrouss Neuronen mat extensiv dendriteschen Arborisatiounen an héijer Wirbelsäule (Kemp and Powell, 1971; Graveland and DiFiglia, 1985; Rafols et al., 1989; Kawaguchi et al., 1990). Glutamatergesch an dopaminergesch Neuronen sinn déi zwee primär afferent Inputen zum NAc, haaptsächlech kontaktéieren d'dendritesch Wellen a Wirbelsäule vun MSNs (Groves, 1980; Kaiya an Namba, 1981; Groves et al., 1994). Speziell kréien d'NAc Schuel a Kär glutamatergesch Input vu funktionell ënnerschiddleche kortikale Gebidder (Brog et al., 1993). D'NAc Schuel ass och innervéiert vun excitatoreschen Afferenten aus subkortikale Regiounen wéi Hippocampus, Thalamus a basolateral Amygdala (Brog et al., 1993; Wright and Groenewegen, 1995). Virdrun Studien hunn bewisen datt dës glutamatergesch Inputen eng pivotal Roll spillen an der Motivatioun an Zilgeriicht Verhalen wéi Iessen a Belounungssich (Maldonado-Irizarry et al., 1995; Kelley and Swanson, 1997; Reynolds a Berridge, 2003; Richard a Berridge, 2011). Déi aner predominant Input op NAc MSNs ass vun dopaminergesche Afferenten déi aus dem ventrale tegmentale Beräich projizéieren (Lindvall et Björklund, 1978; Veening et al., 1980; Kalivas & Miller, 1984). Interessanterweis hu fréier Studien déi ähnlech Modeller vum intermittierenden Zockerzougang benotzt hunn gewisen datt de resultéierende binge-ähnlechen Konsum zu enger Erhéijung vun extrazellulärer Dopamin am NAc ähnlech (och a mannerem Ausmooss) zu Drogen vu Mëssbrauch resultéiert (Rada et al., 2005; Avena et al., 2006), a kann Dopamin Rezeptor Ausdrock moduléieren (Colantuoni et al., 2001, 2002) am NAc Kär a Shell. Interessanterweis verursaacht binge-ähnlechen Konsum vun Saccharose eng Eskalatioun vun der Intake mat der Zäit ähnlech wéi Selbstverwaltung vun Drogen vu Mëssbrauch wéi Kokain an Heroin (Ahmed a Koob, 1998; Ahmed et al., 2000, 2003) déi mat der Entwécklung vun engem "süchteg wéi" Staat assoziéiert ass.
Eis Analyse vun der Brancheuerdnungsmorphometrie weist datt d'Gesamtreduktioun vun der dendritescher Längt vun den NAc Shell MSNs verursaacht duerch laangfristeg Saccharoseaufnahme primär aus Reduktiounen an der Komplexitéit vun distale Branchebestellungen resultéiert. Mir hunn reduzéiert distal Verzweigung observéiert (4. a 5. Uerdnung an iwwer Branchebestellungen) a wesentlech reduzéierter mëttlerer Längt op 5. E gemeinsame Faktor, dee méiglecherweis dës Zort vun dendritescher Restrukturéierung beaflosst, enthält Ännerungen an der synaptescher Konnektivitéit an / oder der Funktioun (Russo et al., 2010). Virdrun Studien hu gewisen datt glutamatergesch Synapsen op MSNs haaptsächlech op Wirbelsäule geformt ginn, besonnesch bei distalen Dendriten (Groenewegen et al., 1999). Zousätzlech, Co-Lokaliséierung vun Dopamin a glutamatergesch Inputen aus der prefrontaler Cortex (Sesack a Pickel, 1992), hippocampus (Totterdell a Smith, 1989; Sesack a Pickel, 1990), an amygdala (Johnson et al., 1994) goufen op dendritesche Wirbelen vun MSNs observéiert. Dës Observatioune kombinéiert mat der verstäerkter Dicht vun der Wirbelsäule no laangfristeg Saccharoseverbrauch an eiser Etude gesi, ënnerstëtzen d'Bildung vu verstäerkten excitatoreschen Inputen. Dofir entsteet d'Méiglechkeet, wou persistent Effekter verursaacht duerch verlängert binge-ähnlech Saccharoseaufnahme erhéicht excitatoresch synaptesch Aktivitéit bei den distale Dendriten vun MSNs an der NAc Shell erliichteren. Dofir kann d'Reduktioun an / oder d'Retraktioun vun distalen Dendriten iwwer e synaptesche homeostatesche Mechanismus entstoen. (Reissner a Kalivas, 2010), awer dëst bleift ze bestëmmen.
Et ass interessant ze bemierken datt Crombag a Kollegen gewisen hunn datt et keng Wirbelsäure Dicht Erhéijung an der NAc Shell no 4-Wochen Saccharose Konsum iwwer d'Nues-Poke Selbstverwaltungsparadigma trotz enger méi robuster Acquisitioun an enger méi héijer Äntwert op Saccharose am Verglach mat Amphetamin (Crombag et al., 2005). Hir Observatioun vun engem Fehlen vun der Verännerung vun der Wirbelsäule op 4 Wochen spigelt eis Erkenntnisser. Am Géigesaz, awer, weist eis Studie datt no laangfristeg (12 Wochen) Belaaschtung fir chronesch Saccharose Konsum eng bedeitend Erhéijung vun der distaler Wirbelsäule Dicht op den MSNs vun den Saccharose-Erliefnes Ratten ass. Ausserdeem huet eise Laboratoire virdru gewisen datt laangfristeg (12 Wochen) Saccharosekonsum eng differenziell pharmakologesch Äntwert op Pharmakotherapeutik erliichtert, déi gewisen hunn d'Dopamin- an Acetylcholinreaktiounen um Niveau vun der NAc ze moduléieren.Shariff et al., Press). Zesummegefaasst suggeréiert dëst datt laangfristeg (12 Wochen an doriwwer eraus) Saccharose Belaaschtung, déi méi präzis reflektéiert vun real-Welt Szenarien, zu Morphologie Adaptatiounen um Niveau vum NAc resultéiert.
Am Sënn vun Drogen vu Mëssbrauch produzéiert widderholl Belaaschtung vu verschiddenen Drogen laang dauerhafte Verännerungen an der Struktur vun Dendriten an dendritesche Wirbelen. Zum Beispill, Amphetaminen a Kokain erhéijen béid Wirbelsäule Dicht am NAc a béid Shell a Kär (Robinson a Kolb, 2004). Nikotin Belaaschtung ass och gewise ginn fir d'Dicht vun der Wirbelsäule an der NAc Shell ze erhéijen. Ëmgekéiert féiert d'Morphinbelaaschtung zu enger Ofsenkung vun der Wirbelsäule an der dendritescher Branche Komplexitéit (Robinson a Kolb, 2004). Wat de laangfristeg Saccharoseverbrauch ugeet, hu mir eng Erhéijung vun der Wirbelsäule beobachtet ähnlech wéi Amphetamin, Kokain an Nikotin an am Géigesaz zum Effekt vum Morphin. Wéi och ëmmer, am Géigesaz zu Amphetamin a Kokain, awer ähnlech wéi Nikotin, ass d'Erhéijung vun der Wirbelsäule Dicht op laangfristeg Belaaschtung fir Saccharose limitéiert op d'NAc Shell. Et ass och interessant datt Ännerungen a béid dendritesch Verzweigung (Robinson a Kolb, 1999) an Wirbelsäule Dicht (Li et al., 2003) produzéiert vun Amphetamin oder Kokain sinn op distal Dendriten vun MSNs an der NAc agespaart, wat d'Resultater an eiser Etude reflektéiert. Ausserdeem, a korroborativ zu den uewe beschriwwenen Ännerungen, ass d'Saccharosekonsum och virdru gewise ginn fir excitatoresch synaptesch Kraaft op accumbal Dopaminneuronen ze verbesseren (Stuber et al., 2008b) wéi och aner Komponente vum mesolimbesche Belounungswee (Stuber et al., 2008a; Chen et al., 2010). Zesummegefaasst poséiert dëst Saccharose als e mächtege Modulator vun der Neuronmorphologie no längerer schwéierer Notzung, wat ähnlech ass mat den Effekter, déi vun Drogen vu Mëssbrauch observéiert ginn.
Och wa weider Ermëttlungen erfuerderlech sinn fir déi cellulär a synaptesch Mechanismen z'entdecken, déi zu de morphologesche Verännerungen an dëser Etude bäidroen, weisen eis Resultater bedeitend neuronal Effekter entstinn duerch laangfristeg Saccharosekonsum. Besonnesch eng Iwwerleeung, déi net an eiser Etude iwwerpréift gouf, ass ob déi observéiert morphologesch Effekter vun der Saccharose och mat net-kaloresche Séissstoffer wéi Saccharin ausgeléist kënne ginn. An dëser Hisiicht ass et wichteg ze bemierken datt Lenoir a Kollegen gewisen hunn datt intensiv Séiss Kokainbelounung iwwerschreift, sief et duerch Saccharin oder Saccharose generéiert (Lenoir et al., 2007). Ausserdeem, eng rezent Etude publizéiert vun eisem Labo (Shariff et al., Press) weist datt Varenicline, en nikotineschen Acetylcholin-Rezeptor partiellen Agonist souwuel d'Saccharose- wéi och d'Saccharin-Aufnahme bei Nager reduzéiert huet no deemselwechte laangfristeg intermittierenden Zougangsregime, deen an der aktueller Studie benotzt gouf. Interessanterweis hu fréier Studien Ähnlechkeeten an den akuten Effekter vun net-kaloresche Séissstoffer wéi Saccharin an Saccharose um Niveau vun der NAc gewisen.Scheggi et al., 2013; Tesch et al., 2013; Carelli & West, 2014). Wéi och ëmmer, weider Studien musse bestëmmen ob net-kaloresch Séissstoffer laangfristeg Effekter induzéieren kënnen ähnlech wéi Ännerungen an der Morphologie vun den NAc Shell MSNs verursaacht duerch laangfristeg Saccharosekonsum hei gemellt.
De Mangel un Effekt op NAc MSN Morphologie nom kuerzfristeg Saccharosekonsum, beliicht d'Wichtegkeet vun der Ëmsetzung vun laangfristeg Studien fir den Impakt vu längerer Mëssbrauch vun Drogen oder natierleche Belounungen wéi Saccharose ze bewäerten. Wat d'Ofhängegkeet ugeet, sinn net nëmmen widderholl Zyklen vu Binge-Intake an Abstinenz Schlësselkomponente vum Suchtzyklus, e wuessende Kierper vu Beweiser huet opgedeckt datt den Iwwergank zu Ofhängegkeet e progressive Prozess ass deen dacks iwwer eng verlängert Zäit geschitt. Och wann d'Suchteigenschaften vun Zucker onsécher bleiwen, gëtt d'Plausibilitéit vun der Sucht un aner net-Drogenbelounungen wéi Sex, Spillen a Liewensmëttel ëmmer méi ënnersicht. D'Resultater vun dëser Etude addéieren Verdéngschter zu der Hypothese datt Zucker wéi Saccharose potenziell Suchtfaktoren hunn no laangfristeg, bingeähnlechen Konsum. Eis Resultater hunn och Implikatioune fir d'wuessend Zuel vu Kanner a Jugendlecher, déi ongesonde Iessgewunnechten (héigen Zockerverbrauch a Binge-Iessen) an Erwuessenen erhalen. Am Aklang mat dem erhéicht Risiko fir metabolesch Effekter z'entwéckelen ass et och méiglech datt neurologësch a psychiatresch Konsequenzen, déi Stëmmung a Motivatioun beaflossen, och aus dëse Verhalen entstinn.
Autor Contributeuren
Un der Fuerschungsdesign deelgeholl: PK, SB. Experimenter duerchgefouert: PK, MS, AB, MF, EM. Datenanalyse: PK, MF, MS. Interpretéiert d'Donnéeën an huet zum Schreiwen vum Manuskript bäigedroen: PK, MS, MF, EM, MB, SB. All Autoren liesen an guttgeheescht der Finale Manuskript fir Soumissioun.
Konflikt vun der Zënssazéierung
D'Auteuren deklaréieren datt d'Fuerschung an der Verôffentlechung vu kommerziellen oder finanzielle Bezéiungen, déi als potenzielle Konflikt vun Interesse entwéckelt ginn kënne gemaach ginn.
D'Rezensoren SC, SA an den Handling Editor hunn hir gemeinsame Vereenegung deklaréiert, an den Handling Editor seet datt de Prozess trotzdem de Standarde vun enger fairer an objektiver Iwwerpréiwung entsprécht.
Arbeschterlidder
Dës Aarbecht gouf ënnerstëtzt duerch Finanzéierung vu Stipendien vum Australian Research Council (FT1110884) op SB an dem National Health and Medical Research Council (1061979) op SB a MB.
Ergänzungsmaterial
Den Ergänzungsmaterial fir dësen Artikel fannt Dir online op: http://journal.frontiersin.org/article/10.3389/fnbeh.2016.00054
Zousaz Figur 1. Sucrose ofgeroden an Preferenze vun 4 an 12 Woch sucrose Konsuméiere Ratten. (A,B) weisen Eskalatioun vun der totaler Saccharoseaufnahme (ml) iwwer 4 an 12 Woche vun der Belaaschtung. (C, D) weisen héich Präferenz fir Saccharose iwwer Waasser während Perioden vun der Saccharose Presentatioun.
Referenze
Ahmed SH, Koob GF (1998). Iwwergank vu moderéierter bis exzessiv Drogenintak: Ännerung vum hedonesche Setpunkt. Science 282, 298-300. Doi: 10.1126 / science.282.5387.298
Ahmed SH, Lin D, Koob GF, Parsons LH (2003). Eskalatioun vu Kokain Selbstverwaltung hänkt net vun der verännerter Kokain-induzéierter Nukleus accumbens Dopaminniveauen of. J. Neurochem. 86, 102-113. Doi: 10.1046 / j.1471-4159.2003.01833.x
Ahmed SH, Walker JR, Koob GF (2000). Persistent Erhéijung vun der Motivatioun fir Heroin bei Ratten ze huelen mat enger Geschicht vun der Drogeneskalatioun. Neuropsychopharmacologie 22, 413–421. doi: 10.1016/S0893-133X(99)00133-5
Arslanian, S. (2002). Typ 2 Diabetis bei Kanner: Klinesch Aspekter a Risikofaktoren. Horm Res 57 (Suppl. 1), 19–28. doi: 10.1159/000053308
Avena, NM, Bocarsly, ME, Hoebel, BG, and Gold, MS (2011). Iwwerlappungen an der Nosologie vu Substanzmëssbrauch an Iwwereess: d'Iwwersetzungsimplikatioune vun der "Liewensmëttel Sucht". Curr. Drogenmëssbrauch Rev. 4, 133-139. Doi: 10.2174 / 1874473711104030133
Avena, NM, Rada, P., and Hoebel, BG (2008). Beweiser fir Zocker Sucht: Verhalens- an neurochemesch Effekter vun intermittierend, exzessive Zockeropnahm. Neurosci. Biobehav. Rev. 32, 20-39. Doi: 10.1016 / j.neubiorev.2007.04.019
Avena NM, Rada P, Moise N, Hoebel BG (2006). Sucrose Sham Ernierung op engem Binge Zäitplang befreit accumbens Dopamin ëmmer erëm an eliminéiert d'Acetylcholin Sattheetsreaktioun. Neurologie 139, 813-820. Doi: 10.1016 / j.neuroscience.2005.12.037
Benton, D. (2010). D'Plausibilitéit vun der Zockersucht a seng Roll bei Adipositas an Iessstéierungen. Clin. Nutr. 29, 288–303. doi: 10.1016/j.clnu.2009.12.001
Bray GA, Popkin BM (2014). Diätzocker a Kierpergewiicht: hu mir eng Kris an der Epidemie vun Adipositas an Diabetis erreecht?: Gesondheet verdammt! Den Zocker opgoen. Diabetis Care 37, 950-956. doi: 10.2337/dc13-2085
Brog JS, Salyapongse A, Deutch AY, Zahm DS (1993). D'Muster vun der afferenter Innervatioun vum Kär an der Schuel am "Accumbens" Deel vum Ratten ventrale Striatum: Immunhistochemesch Detektioun vu retrograd transportéiert Fluor-Gold. J. Comp. Neurol. 338, 255 – 278. doi: 10.1002 / cne.903380209
Bucher Della Torre, S., Keller, A., Laure Depeyre, J., and Kruseman, M. (2015). Zocker-séisseg Getränker an Adipositasrisiko bei Kanner a Jugendlecher: eng systematesch Analyse iwwer wéi methodologesch Qualitéit d'Conclusiounen beaflosse kann. J. Acad. Nutr. Diät. [Epub virum Drock]. doi: 10.1016/j.jand.2015.05.020
Carelli RM, West EA (2014). Wann e gudde Goût schlecht gëtt: neurale Mechanismen, déi d'Entstoe vun negativen Afloss ënnersträichen an assoziéiert natierlech Belounungsdevaluatioun vu Kokain. Neuropharmakologie 76 (Pt B), 360–369. doi: 10.1016/j.neuropharm.2013.04.025
Chen BT, Hopf FW, Bonci A. (2010). Synaptesch Plastizitéit am mesolimbesche System: therapeutesch Implikatioune fir Substanzmëssbrauch. Ann. NY Acad. Sci. 1187, 129-139. Doi: 10.1111 / j.1749-6632.2009.05154.x
Colantuoni, C., Rada, P., McCarthy, J., Patten, C., Avena, NM, Chadeayne, A., et al. (2002). Beweiser datt intermittéiert, exzessiv Zockeropnahm endogen opioid Ofhängegkeet verursaacht. Obes. Res. 10, 478 – 488. doi: 10.1038 / oby.2002.66
Colantuoni, C., Schwenker, J., McCarthy, J., Rada, P., Ladenheim, B., Cadet, JL, et al. (2001). Exzessiv Zockeropnahm verännert d'Bindung un Dopamin a Mu-Opioid Rezeptoren am Gehir. Neuroreport 12, 3549–3552. doi: 10.1097/00001756-200111160-00035
Crombag, HS, Gorny, G., Li, Y., Kolb, B., and Robinson, TE (2005). Opposite Effekter vun Amphetamin Selbstverwaltungserfahrung op dendritesche Wirbelen an der medialer an orbitaler prefrontaler Cortex. Cereb. Cortex 15, 341–348. doi: 10.1093/cercor/bhh136
Dabelea, D., Mayer-Davis, EJ, Saydah, S., Imperatore, G., Linder, B., Divers, J., et al. (2014). Prävalenz vum Typ 1 an Typ 2 Diabetis bei Kanner a Jugendlecher vun 2001 bis 2009. JAMA 311, 1778 – 1786. doi: 10.1001 / jama.2014.3201
Davis, C. (2015). D'Epidemiologie an d'Genetik vun der Binge Iessstéierung (BED). CNS Spectr. 20, 522-529. Doi: 10.1017 / s1092852915000462
Ford ES, Dietz WH (2013). Trends an der Energieaufnahme bei Erwuessener an den USA: Resultater vun NHANES. Am. J. Clin. Nutr. 97, 848-853. doi: 10.3945/ajcn.112.052662
Fryar, CD, Carroll, MD, Ogden, CL (2014). Prävalenz vun Iwwergewiicht, Adipositas an Extrem Adipositas ënner Erwuessener: USA, 1960-1962 duerch 2011-2012. Atlanta, GA: CDC.
Graveland, GA, DiFiglia, M. (1985). D'Frequenz an d'Verdeelung vu mëttelgrousse Neuronen mat indented Kären am Primat an Nager Neostriatum. Brain Res. 327, 307–311. doi: 10.1016/0006-8993(85)91524-0
Groenewegen, HJ, Wright, CI, Beijer, AV, and Voorn, P. (1999). Konvergenz a Segregatioun vu ventralen striatalen Inputen an Ausgänge. Ann. NY Acad. Sci. 877, 49–63. doi: 10.1111/j.1749-6632.1999.tb09260.x
Groves, PM (1980). Synaptesch Endungen an hir postsynaptesch Ziler am Neostriatum: synaptesch Spezialisatiounen aus der Analyse vu Serien Sektiounen opgedeckt. Proc. Natl. Acad. Sci. USA 77, 6926-6929. Doi: 10.1073 / pnas.77.11.6926
Groves PM, Linder JC, Young SJ (1994). 5-Hydroxydopamin-markéiert dopaminergesch Axonen: dreidimensional Rekonstruktioune vun Axonen, Synapsen a postsynapteschen Ziler am Rattenneostriatum. Neurologie 58, 593–604. doi: 10.1016/0306-4522(94)90084-1
Hu, FB (2013). Geléist: et gëtt genuch wëssenschaftlech Beweiser datt d'Ofsenkung vun Zocker-séisseg Getränksverbrauch d'Prévalenz vun Adipositas an Adipositas-relatéierte Krankheeten reduzéiere wäert. Obes. Rev. 14, 606-619. doi: 10.1111/obr.12040
Johnson, LR, Aylward, RL, Hussain, Z., and Totterdell, S. (1994). Input vun der Amygdala zum Rattenkär accumbens: seng Relatioun mat Tyrosinhydroxylase Immunreaktivitéit an identifizéierten Neuronen. Neurologie 61, 851–865. doi: 10.1016/0306-4522(94)90408-1
Kaiya, H., and Namba, M. (1981). Zwou Aarte vun dopaminergesche Nerveterminalen am Rattenneostriatum. Eng ultrastrukturell Studie. Neurosci. Lett. 25, 251–256. doi: 10.1016/0304-3940(81)90400-6
Kalivas PW, Miller JS (1984). Neurotensin Neuronen am ventralen Tegmentalgebitt projizéieren op de medialen Nukleus accumbens. Brain Res. 300, 157–160. doi: 10.1016/0006-8993(84)91351-9
Kawaguchi, Y., Wilson, CJ, and Emson, PC (1990). Projektiounssubtypen vu Ratten neostriatal Matrixzellen opgedeckt duerch intrazellulär Injektioun vu Biozytin. J. Neurosci. 10, 3421-3438.
Kelley AE, Swanson CJ (1997). Fütterung induzéiert duerch Blockade vun AMPA a Kainat Rezeptoren am ventralen Striatum: eng Mikroinfusiounsmappingstudie. Behav. Brain Res. 89, 107–113. doi: 10.1016/S0166-4328(97)00054-5
Kemp JM, Powell TP (1971). D'synaptesch Organisatioun vum Caudatkär. Philos. Trans. R. Soc. London. B Biol. Sci. 262, 403-412. Doi: 10.1098 / rstb.1971.0103
Kenny, PJ (2011). Belounungsmechanismen an der Adipositas: nei Abléck an zukünfteg Richtungen. Neuron 69, 664-679. Doi: 10.1016 / j.neuron.2011.02.016
Kessler RC, Berglund PA, Chiu WT, Deitz AC, Hudson JI, Shahly V, et al. (2013). D'Prévalenz an d'Korrelate vun der Binge Iessstéierung an der Weltgesondheetsorganisatioun World Mental Health Surveys. Biol. Psychiatrie 73, 904-914. Doi: 10.1016 / j.biopsych.2012.11.020
Klenowski, PM, Fogarty, MJ, Belmer, A., Noakes, PG, Bellingham, MC, and Bartlett, SE (2015). Strukturell a funktionell Charakteriséierung vun dendriteschen Arbors a GABAergesche synaptesche Inputen op Interneuronen a Haaptzellen an der Ratten basolateral Amygdala. J. Neurophysiol. 114, 942-957. Doi: 10.1152 / jn.00824.2014
Lenoir, M., Serre, F., Cantin, L., and Ahmed, SH (2007). Intens Séiss iwwerschreift d'Kokainbelounung. PLoS ONE 2: e698. Doi: 10.1371 / journal.pone.0000698
Li, Y., Kolb, B., and Robinson, TE (2003). De Standort vu persistent Amphetamin-induzéierte Verännerungen an der Dicht vun dendritesche Wirbelen op mëttlere spiny Neuronen am Nucleus accumbens a caudate-putamen. Neuropsychopharmacologie 28, 1082-1085. Doi: 10.1038 / sj.npp.1300115
Lindvall, O., Björklund, A. (1978). Anatomie vun den dopaminergeschen Neuronsystemer am Ratgehir. Adv. Biochem. Psychopharmacol. 19, 1-23.
Lutter, M., and Nestler, E. J. (2009). Homeostatesch an hedonesch Signaler interagéieren an der Reguléierung vun der Nahrungsaufnahme. J. Nutr. 139, 629-632. Doi: 10.3945 / jn.108.097618
Maldonado-Irizarry CS, Swanson CJ, Kelley AE (1995). Glutamat Rezeptoren am Nukleus accumbens Shell kontrolléieren d'Fütterungsverhalen iwwer de lateralen Hypothalamus. J. Neurosci. 15, 6779-6788.
Malik, VS, Popkin, BM, Bray, GA, Després, JP, and Hu, FB (2010). Zocker-séisseg Getränker, Adipositas, Typ 2 Diabetis mellitus, a Risiko vu kardiovaskuläre Krankheeten. Circulatioun 121, 1356–1364. doi: 10.1161/CIRCULATIONAHA.109.876185
Ng SW, Slining MM, Popkin BM (2012). Benotzung vu kaloreschen an net-kaloresche Séissstoffer an US Konsument verpackte Liewensmëttel, 2005-2009. J. Acad. Nutr. Diät. 112, 1828–1834 e1821–e1826. doi: 10.1016/j.jand.2012.07.009
Nielsen SJ, Siega-Riz AM, Popkin BM (2002). Trends an der Energiezufuhr an den USA tëscht 1977 an 1996: ähnlech Verréckelung iwwer Altersgruppen gesi ginn. Obes. Res. 10, 370 – 378. doi: 10.1038 / oby.2002.51
Paxinos, G., a Watson, C. (2007). De Rat Brain an de Stereotaxesche Koordinaten. Amsterdam; Boston, MA: Academic Press/Elsevier.
Popkin, BM (2010). Wat ass falsch mat der US Approche fir Adipositas? Virtuelle Mentor 12, 316–320. doi: 10.1001/virtualmentor.2010.12.4.pfor2-1004
Rada P, Avena NM, Hoebel BG (2005). Deeglech Bingeing op Zocker verëffentlecht ëmmer erëm Dopamin an der Accumbens Shell. Neurologie 134, 737-744. Doi: 10.1016 / j.neuroscience.2005.04.043
Rafols JA, Cheng HW, McNeill TH (1989). Golgi-Studie vum Mausstriatum: Altersbezunnen dendritesch Verännerungen a verschiddene neuronale Populatiounen. J. Comp. Neurol. 279, 212 – 227. doi: 10.1002 / cne.902790205
Ranjan, A., and Mallick, BN (2010). Eng modifizéiert Method fir konsequent an zouverlässeg Golgi-cox Färzen a wesentlech reduzéierter Zäit. Front. Neurol. 1:157. doi: 10.3389/fneur.2010.00157
Reinehr, T. (2013). Typ 2 Diabetis mellitus bei Kanner a Jugendlecher. Welt J. Diabetis 4, 270–281. doi: 10.4239/wjd.v4.i6.270
Reissner KJ, Kalivas PW (2010). Benotzt Glutamat Homeostasis als Zil fir Suchtkrankheeten ze behandelen. Behënnert. Pharmacol. 21, 514–522. doi: 10.1097/FBP.0b013e32833d41b2
Reynolds SM, Berridge KC (2003). Glutamate motivational Ensemblen am Nucleus accumbens: Rostrocaudal Shell Gradienten vun Angscht a Fudder. EUR. J. Neurosci. 17, 2187-2200. Doi: 10.1046 / j.1460-9568.2003.02642.x
Richard JM, Berridge KC (2011). Nucleus accumbens Dopamin / Glutamat Interaktioun wiesselt Modus fir Wonsch versus Angscht ze generéieren: D (1) eleng fir appetitiv Iessen awer D (1) an D (2) zesummen aus Angscht. J. Neurosci. 31, 12866-12879. Doi: 10.1523 / JNEUROSCI.1339-11.2011
Robinson, TE, and Kolb, B. (1999). Ännerungen an der Morphologie vun Dendriten an dendritesche Wirbelen am Nukleus accumbens a prefrontal Cortex no widderholl Behandlung mat Amphetamin oder Kokain. EUR. J. Neurosci. 11, 1598-1604. Doi: 10.1046 / j.1460-9568.1999.00576.x
Robinson, TE, and Kolb, B. (2004). Strukturell Plastizitéit verbonne mat der Belaaschtung vun Drogen vu Mëssbrauch. Neuropharmakologie 47 (Suppl. 1), 33–46. doi: 10.1016/j.neuropharm.2004.06.025
Russo, SJ, Dietz, DM, Dumitriu, D., Morrison, JH, Malenka, RC an Nestler, EJ (2010). Dee süchteg Synapse: Mechanismen vun synaptescher a struktureller Plastizitéit am Nukleus accumbens. Trends Neurosci. 33, 267–276. doi: 10.1016/j.tins.2010.02.002
Rutledge, LT, Duncan, J., & Beatty, N. (1969). Eng Studie vu pyramideschen Zell Axon Collateralen an intakten an deelweis isoléierten erwuessene zerebrale cortex. Brain Res. 16, 15–22. doi: 10.1016/0006-8993(69)90082-1
Saper CB, Chou TC, Elmquist JK (2002). De Besoin fir ze ernähren: homeostatesch an hedonesch Kontroll vum Iessen. Neuron 36, 199–211. doi: 10.1016/S0896-6273(02)00969-8
Scheggi, S., Secci, ME, Marchese, G., De Montis, MG, and Gambarana, C. (2013). Afloss vun der Geschmaach op d'Motivatioun fir fir kaloresch an net-kaloresch Nahrung an net-Liewensmëttel entzunnen a Liewensmëttel entzunnen Ratten ze bedreiwen. Neurologie 236, 320-331. Doi: 10.1016 / j.neuroscience.2013.01.027
Sesack SR, Pickel VM (1990). Am Rat medialen Nukleus accumbens, hippocampal a katecholaminergesch Terminaler konvergéieren op spiny Neuronen a sinn an der Astellung zueneen. Brain Res. 527, 266–279. doi: 10.1016/0006-8993(90)91146-8
Sesack SR, Pickel VM (1992). Prefrontal kortikale Efferenter an der Rattensynapse op onbekannte neuronalen Ziler vu Katecholamin-Terminaler am Nucleus accumbens septi an op Dopaminneuronen am ventralen Tegmentalgebitt. J. Comp. Neurol. 320, 145 – 160. doi: 10.1002 / cne.903200202
Shariff, M., Quik, M., Holgate, JY, Morgan, M., Patkar, OL, Tam, V., et al. (an der Press). Neuronal nikotinesch Acetylcholin Rezeptor Modulatoren reduzéieren Zockerzufuhr. PLoS ONE.
Sheehan, DV, and Herman, BK (2015). Déi psychologesch a medizinesch Faktore verbonne mat onbehandelt Binge-Iessstéierung. Prim. Pfleeg Begleeder CNS Disord. 17. doi: 10.4088/PCC.14r01732
Simms JA, Steensland P, Medina B, Abernathy KE, Chandler LJ, Wise R, et al. (2008). Intermittéierend Zougang zu 20% Ethanol induzéiert héije Ethanolverbrauch bei Long-Evans a Wistar Ratten. Alkohol. Clin. Exp. Res. 32, 1816-1823. Doi: 10.1111 / j.1530-0277.2008.00753.x
Steensland, P., Simms, JA, Holgate, J., Richards, JK, and Bartlett, SE (2007). Varenicline, en alpha4beta2 nikotineschen Acetylcholin Rezeptor partiell Agonist, reduzéiert selektiv den Ethanolkonsum a sichen. Proc. Natl. Acad. Sci. USA 104, 12518-12523. Doi: 10.1073 / pnas.0705368104
Stuber, GD, Hopf, FW, Hahn, J., Cho, SL, Guillory, A., and Bonci, A. (2008a). Fräiwëlleg Ethanolopnahm verbessert excitatoresch synaptesch Kraaft am ventrale tegmentale Beräich. Alkohol. Clin. Exp. Res. 32, 1714-1720. Doi: 10.1111 / j.1530-0277.2008.00749.x
Stuber GD, Klanker M, de Ridder B, Bowers MS, Joosten RN, Feenstra MG, et al. (2008b). Belounung-predictive Cues verbesseren excitatoresch synaptesch Kraaft op Midbrain Dopamin Neuronen. Science 321, 1690-1692. Doi: 10.1126 / science.1160873
Swanson SA, Crow SJ, Le Grange D, Swendsen J, Merikangas KR (2011). Prävalenz a Korrelate vun Iessstéierunge bei Jugendlechen. Resultater vun der nationaler Komorbiditéit Ëmfro Replikatioun Adolescent Supplement. Arch. Gen. Psychiatrie 68, 714 – 723. doi: 10.1001 / archgenpsychiatry.2011.22
Te Morenga, L., Mallard, S., and Mann, J. (2013). Diätzucker a Kierpergewiicht: Systematesch Iwwerpréiwung a Metaanalyse vu randomiséierter kontrolléierter Studien a Kohortstudien. BMJ 346:e7492. doi: 10.1136/bmj.e7492
Totterdell, S., and Smith, AD (1989). Konvergenz vun hippocampalen an dopaminergeschen Input op identifizéiert Neuronen am Nukleus accumbens vun der Rat. J. Chem. Neuroanat. 2, 285-298.
Tukey DS, Ferreira JM, Antoine SO, D'Amour JA, Ninan I, Cabeza de Vaca S, et al. (2013). Sucrose-Intake induzéiert séier AMPA-Rezeptorhandel. J. Neurosci. 33, 6123-6132. Doi: 10.1523 / JNEUROSCI.4806-12.2013
Veening JG, Cornelissen FM, Lieven PA (1980). Déi topesch Organisatioun vun den Afferenten zum Caudatoputamen vun der Rat. Eng Meerrettich Peroxidase Studie. Neurologie 5, 1253–1268. doi: 10.1016/0306-4522(80)90198-0
Ventura, T., Santander, J., Torres, R., and Contreras, AM (2014). Neurobiologesch Basis vu Verlaangen no Kuelenhydrater. Ernährung 30, 252 – 256. doi: 10.1016 / j.nut.2013.06.010
Volkow, ND, Wang, GJ, Fowler, JS, Tomasi, D., and Baler, R. (2012). Liewensmëttel an Drogenbelounung: iwwerlappend Kreesleef an der mënschlecher Adipositas a Sucht. Curr. Top. Behav. Neurosci. 11, 1–24. doi: 10.1007/7854_2011_169
Wise, RA (1973). Fräiwëlleg Ethanolopnahm bei Ratten no Belaaschtung fir Ethanol op verschiddene Flugplang. Psychopharmacologia 29, 203-210. Doi: 10.1007 / BF00414034
Wright CI, Groenewegen HJ (1995). Muster vun der Konvergenz an der Segregatioun am medialen Nukleus accumbens vun der Rat: Bezéiunge vu prefrontal cortical, midline thalamesch a basal Amygdaloid Afferenten. J. Comp. Neurol. 361, 383 – 403. doi: 10.1002 / cne.903610304
Schlësselwieder: binge-ähnlechen Konsum, laangfristeg, mëttel spiny Neuron, nucleus accumbens, sucrose
Zitat: Klenowski PM, Shariff MR, Belmer A, Fogarty MJ, Mu EWH, Bellingham MC a Bartlett SE (2016) Verlängert Verbrauch vu Saccharose op enger Binge-Like Manéier, Ännert d'Morphologie vu mëttlere Spiny Neuronen an der Nucleus Accumbens Shell. Front. Behav. Neurosci. 10: 54. Doi: 10.3389 / fnbeh.2016.00054
Empfang: 03 Dezember 2015; Akzeptéiert: 07 Mäerz 2016;
Verëffentlecht: 23 March 2016.
Edited by:
Djoher Nora Abrous, Institut des Neurosciences de Bordeaux, Frankreich
Review vun:
Serge H. Ahmed, Centre National de la Recherche Scientifique, Frankräich
Stéphanie Caille, Centre National de la Recherche Scientifique, Frankräich
Copyright © 2016 Klenowski, Shariff, Belmer, Fogarty, Mu, Bellingham a Bartlett. Dëst ass en Open-Access Artikel verdeelt ënner de Bedéngungen vun der Creative Commons Attribution License (CC BY). D'Benotzung, Verdeelung oder Vervillfäegung an anere Foren ass erlaabt, wann de ursprénglechen Auteur oder de Lizenzgeber kritiséiert gëtt an datt déi originell Publikatioun an dëser Zeitung zitéiert ass, anhand vun akzeptéierter akademescher Praxis. Kee Benotzung, Verdeelung oder Reproduktioun ass net erlaabt déi dës Conditioune net erhalen.
*Korrespondenz: Selena E. Bartlett, [Email geschützt]
