ഡയലോഗുകൾ ക്ലിനൻ ന്യൂറോസി. 2013 Dec;15(4):431-43.
വേര്പെട്ടുനില്ക്കുന്ന
ധാരാളം മാനസിക വ്യവസ്ഥിതിയുടെ പ്രാധാന്യം അഭികാമ്യമാണെങ്കിലും, മയക്കുമരുന്ന് അടിമയ്ക്ക് ഒരു ജൈവ പ്രക്രിയയാണ് ഉള്ളത്: മയക്കുമരുന്ന് ആവശ്യപ്പെടുന്നതും മയക്കുമരുന്ന് എടുക്കുന്നതും ദുർബലമായ തലച്ചോറിലെ മാറ്റങ്ങൾക്ക് വഴിതെളിക്കുന്നതിനുള്ള ദുരുപയോഗവും മയക്കുമരുന്ന് ഉപയോഗം, അത് ആസക്തിയുടെ അവസ്ഥയെ നിർവചിക്കുന്നു. ഇവിടെ, മയക്കുമരുന്ന്, സെല്ലുലാർ അഡാപ്റ്റേഷനുകൾ, പ്രത്യേകിച്ചു തലച്ചോറിലെ പ്രദേശങ്ങളിൽ ഉണ്ടാകുന്ന, ആദിശക്തിയുമായി ബന്ധപ്പെട്ട പെരുമാറ്റ അസാധാരണത്വങ്ങൾ പരിഹരിക്കുന്നതിന് ഞങ്ങൾ അവലോകനം ചെയ്യുന്നു. ജൈൻ എക്സ്പ്രസിലുള്ള ചില മാറ്റങ്ങൾ, എപ്പിജെനറ്റിക്കൽ സംവിധാനങ്ങൾ, ന്യൂറോൺസ് ആൻഡ് സിനാപ്സസ് എന്നിവയിലെ ന്യൂറോഫിസയോളജിക്കൽ പ്രവർത്തനത്തിലും, ന്യൂറോണൽ ആൻഡ് സിനാപ്റ്റിക് മോർഫോളജിയിലെ പ്ലാസ്റ്റിക്സിറ്റിയുടെ ഫലമായി മധ്യസ്ഥതയായ ന്യൂറോട്രോപിക് ഘടകം സിഗ്നലിങ്ങിലൂടെയാണ്. ഈ തരം മയക്കുമരുന്ന് വ്യതിയാനങ്ങൾ വരുത്തുന്നത്, "സെല്ലുലാർ അല്ലെങ്കിൽ മോളികുലാർ മെമ്മറി" എന്ന രൂപത്തിൽ കാണാവുന്നതാണ്. കൂടാതെ, കൂടുതൽ ആസക്തിയുമായി ബന്ധപ്പെട്ട പ്ലാസ്റ്റിക്സിദ്ധാന്തം പ്ലാസ്റ്റിക്ക്സിക്ക് വളരെ സാമ്യമുള്ളതാണ്, അത് "സ്വഭാവസവിശേഷത" ന്റെ കൂടുതൽ ക്ലാസിക് രൂപങ്ങളുമായി ബന്ധപ്പെട്ടിരിക്കുന്നു. ഒരുപക്ഷേ, പരിസ്ഥിതിക്ക് അഭിമുഖീകരിക്കുമ്പോൾ ന്യൂറോണുകൾക്ക് ലഭ്യമായ അഡാപ്റ്റീവ് സംവിധാനങ്ങളുടെ പരിമിതമായ പ്രതിഫലനത്തെ പ്രതിഫലിപ്പിക്കുന്നു. വെല്ലുവിളികൾ. അവസാനമായി, ആസക്തി ബന്ധമുള്ള മോളികുലാർ ആന്റ് സെല്ലുലാർ അഡാപ്റ്റേഷനിൽ ഉൾക്കൊണ്ടിരിക്കുന്ന അതേ മസ്തിഷ്ക മേഖലകളിൽ മിക്കതും മെമ്മറി കൂടുതൽ ക്ലാസിക് രൂപങ്ങൾ ഉൾക്കൊള്ളുന്നു. അസാധാരണ ഓർമ്മകൾ ആസക്തി സിൻഡ്രോമുകളുടെ പ്രധാന ഡ്രൈവർമാരാണെന്ന് കണക്കിലെടുക്കുക. മയക്കുമരുന്നിന്റെയും കോശങ്ങളുടെയും അടിസ്ഥാനത്തിൽ മയക്കുമരുന്നിന്റെ അടിവയറ്റത്തെ വിശദീകരിക്കാനുള്ള ലക്ഷ്യം ഈ പഠനത്തിന്റെ ലക്ഷ്യം ജൈവശാസ്ത്രപരമായി അടിസ്ഥാനമാക്കിയുള്ള ഡയഗ്നോസ്റ്റിക് ടെസ്റ്റുകൾ വികസിപ്പിച്ചെടുക്കുക മാത്രമല്ല, ആസക്റ്റിവ് ഡിസോർഡേസിനുളള കൂടുതൽ ഫലപ്രദമായ ചികിത്സാരീതികളാണ്.
അവതാരിക
മയക്കുമരുന്നിന്റെ അടിമത്തം, മയക്കുമരുന്ന് ഉപയോഗം, മയക്കുമരുന്ന് ഉപയോഗം, മയക്കുമരുന്ന് ഉപയോഗിച്ചുള്ള നിയന്ത്രണം നഷ്ടപ്പെടൽ തുടങ്ങിയവയെ നിർവചിക്കാവുന്നതാണ്, ചില മസ്തിഷ്ക പ്രദേശങ്ങളിൽ നിലനിൽക്കുന്ന ദീർഘകാല മരുന്ന് വ്യതിയാനങ്ങളുണ്ടാകുന്ന മാറ്റങ്ങളാണ് ഇതിന് കാരണം.1 എന്നാൽ ചില വ്യക്തികൾ മാത്രമേ ആവർത്തിച്ച് മയക്കുമരുന്ന് exposure ഉള്ളപ്പോൾ ആസക്തി കുറയുന്നുള്ളൂ, മറ്റു ചിലർക്ക് മയക്കുമരുന്ന് ഉപയോഗിച്ച് മയക്കുമരുന്ന് ഉപയോഗിച്ച് ഒരു ആസക്തി സിൻഡ്രോം രക്ഷപെടാൻ കഴിയും. ആസക്തി വൈകല്യത്തിൽ ഈ വ്യത്യാസത്തിന്റെ ഏതാണ്ട് എൺപതു ശതമാനം ജനിതക കാരണങ്ങളാണുള്ളത്. ഉൽപാദനക്ഷമത, മയക്കുമരുന്ന്, നിക്കോട്ടിൻ, കാൻബാബിനോയിഡുകൾ.2 ഈ ജനിതക റിസ്ക് ഉൾപ്പെടുന്ന ഭൂരിഭാഗം ജീനുകളെയും തിരിച്ചറിയാൻ കഴിഞ്ഞിട്ടില്ല. ഒരുപക്ഷേ, ആസക്തിയുടെ അപര്യാപ്തത (അല്ലെങ്കിൽ, മറ്റ് വ്യക്തികളിൽ, പ്രതിരോധം) നൽകാൻ ഒരു വ്യക്തിയിൽ ഒരു നൂറുകണക്കിന് ജനിതക വ്യതിയാനങ്ങൾ ഉണ്ടായിരിക്കാം.
ആസക്തിയുടെ മറ്റ് 50% അപകടസാധ്യതകൾ കാരണം ഒരു ജീവിതകാലം മുഴുവൻ സംഭവിക്കുന്ന ഒരു വ്യക്തിയുടെ ജനിതകഘടനയുമായി ഇടപഴകുന്ന ഒരു വ്യക്തിയുടെ ജനിതകഘടനയുമായി ഇടപഴകുന്നതിനാലാണ് ആസക്തിയെ കൂടുതലോ കുറവോ ആയി ആസക്തിക്ക് ഇരയാക്കുന്നത്. മന os ശാസ്ത്രപരമായ സമ്മർദ്ദങ്ങൾ ഉൾപ്പെടെ നിരവധി തരത്തിലുള്ള പാരിസ്ഥിതിക ഘടകങ്ങൾ ആസക്തിയിൽ ഉൾപ്പെട്ടിട്ടുണ്ട്, എന്നാൽ ഏറ്റവും ശക്തമായ ഘടകം ദുരുപയോഗത്തിന്റെ ഒരു മയക്കുമരുന്ന് എക്സ്പോഷർ ആണ്. ചില “ഗേറ്റ്വേ” മരുന്നുകൾ, പ്രത്യേകിച്ചും, നിക്കോട്ടിൻ, മറ്റൊരു മയക്കുമരുന്നിന് അടിമപ്പെടാനുള്ള സാധ്യത വർദ്ധിപ്പിക്കുന്നു.3 കൂടാതെ, ജനസംഖ്യയ്ക്ക് ആനുപാതികമായി ജനിതക അപായസാധനങ്ങൾ ഉണ്ടാകുന്നിടത്തോളം കാലം, ഒരു മരുന്നിന്റെ അളവിൽ ഉയർന്ന അളവിൽ ഡോക്ടർമാർ ഉണ്ടാകുന്നത് ഒരു അടിമയാകുന്നതിനോടുള്ള താരതമ്യേന താഴ്ന്ന ജനിതക ലോഡിംഗ്.4
ഒരു ആസക്ഷൻ സിൻഡ്രോം മധ്യത്തിൽ പ്രാധാന്യം നൽകുന്ന മസ്തിഷ്കത്തിന്റെ പ്രത്യേക ഭാഗങ്ങളും, ഈ മേഖലകളിൽ മയക്കുമരുന്ന്, സെല്ലുലാർ അളവുകളിൽ മാറ്റങ്ങൾ വരുത്തുന്ന മാറ്റങ്ങളും തിരിച്ചറിയുന്നതിനായി കഴിഞ്ഞ രണ്ട് ദശാബ്ദങ്ങളിലായി വലിയ പുരോഗതി ഉണ്ടായിട്ടുണ്ട്. ആസക്തി1,5 വളരെയധികം ശ്രദ്ധ നേടിയ ഈ സർക്യൂട്ട് മെസൊംബൈബിക് ഡോപ്പമിൻ സംവിധാനമായി കണക്കാക്കപ്പെടുന്നു. മിഡ് സ്ട്രെയിനിന്റെ മധ്യഭാഗത്തെ ന്യൂട്രസ് അഗ്രംബൻസ് (നാഷണൽ സെന്റർ ഇൻഗ്രൂബൻസ്) എന്ന ന്യൂട്രസ് അൾട്രാവൻസ് ന്യൂട്രോണുകളിൽ ദപോമിൻ ന്യൂറോണുകൾ ഉൾപ്പെടുന്നു. ഈ VTA ന്യൂറോണുകൾ ഹിപ്പോകാമ്പസ്, അമാഗഡാല, പ്രീഫ്രോണൽ കോർട്ടക്സ് (പിഎഫ്എസി) തുടങ്ങി അനേകം മുൻപന്തിയിലുള്ള മേഖലകളെ സഹായിക്കുന്നു.
ഈ വോള്യത്തിൽ ഈ മയക്കുമരുന്ന് ഉത്തേജ ഉത്തേജ പ്രവർത്തനങ്ങളെ ഓർമ്മപ്പെടുത്തുന്നതിന് മൂന്ന് ഓവർലാപ്പിംഗ് കാരണങ്ങൾക്കുള്ള ഓർമ്മപ്പെടുത്തലാണ്.6
- ഒന്നാമതായി, മയക്കുമരുന്ന് അല്ലെങ്കിൽ സെല്ലുലാർ മെമ്മറി എന്ന തരത്തിലുള്ള മയക്കുമരുന്ന് ഉദ്ഗ്രഥിതമാതൃകകൾ കാണാൻ കഴിയും. മയക്കുമരുന്നിന്റെ ഫലമായി ഉണ്ടാകുന്ന നെയ്ൽ സെൽ വ്യത്യാസപ്പെട്ടിരിക്കുന്നു. അതിനാൽ ഒരേ മരുന്ന് വ്യത്യസ്തമായി പ്രതികരിക്കുന്നു. മറ്റ് മരുന്നുകൾ, അല്ലെങ്കിൽ മറ്റ് ഉത്തേജകത ഫലമായി.
- രണ്ടാമതായി, ആസക്തിയുടെ അവസ്ഥയുമായി ബന്ധപ്പെട്ട മാറ്റങ്ങളുടെ പലതും, ഒരുപക്ഷേ, മിക്കതും രസകരമാണ് (ഉദാ: altered gene transcription, epigenetics, സിനാപ്റ്റിക്, മുഴുവൻ സെൽ പ്ലാസ്റ്റിഷ്യൻ, ന്യൂറോണൽ മോർഫോളജി, ന്യൂറോട്രോപിക് സംവിധാനങ്ങൾ) സ്പേഷ്യൽ മെമ്മറി, ഭയം കണ്ടീഷനിംഗ്, ഓപ്പറന്റ് കണ്ടീഷനിങ് തുടങ്ങിയ പാരമ്പര്യ രൂപങ്ങളിലുള്ളവയാണ് അവ.
- മൂന്നാമതായി, മാനസിക പീഡനം മൂലം മസ്തിഷ്കത്തെ ബാധിക്കുന്ന മസ്തിഷ്ക മേഖലകളിൽ ഹിപ്പോകാമ്പസ്, അമീഗഡാല, പി.എഫ്.സി തുടങ്ങിയ പെരുമാറ്റ സ്മരണകൾക്കുള്ള പ്രധാന ന്യൂറൽ സബ്ജാറ്റുകൾ. ആദ്ധ്യാത്മീകൃത മെമ്മറി സർക്യൂട്ടുകളിൽ അസാധാരണത്വങ്ങളെ പ്രതിഫലിപ്പിക്കുന്നതായി ക്ലിനിക്കലിയിൽ (ഉദാഹരണത്തിന്, മയക്കുമരുന്ന് വിരസവും പുനരാവിഷ്കരണവും) കാണിക്കുന്ന ഏറ്റവും ആധികാരികമായ ചില ഘടകങ്ങൾ വർദ്ധിച്ചുവരുന്ന യാഥാർഥ്യത്തോട് ചേർന്നു നിൽക്കുന്നു. ആഡംബര ജീവശാസ്ത്രത്തിന്റെ ശക്തമായ ഡ്രൈവർമാരുടെ ദീർഘകാല മെമ്മറികൾ.4,7,8 നേരെമറിച്ച്, തലച്ചോറിന്റെ പ്രതിഫല പ്രദേശങ്ങൾ (ഉദാ. വിടിഎ, എൻഎസി) പെരുമാറ്റ മെമ്മറിയെ സ്വാധീനിക്കുന്നു.
ഈ ലേഖനം ലഭ്യമായിട്ടുള്ള പ്രധാന തരം മോളികുലാർ സെല്ലുലാർ മാറ്റങ്ങളും, മസ്തിഷ്ക മോഡലുകളുമൊക്കെയുള്ള മസ്തിഷ്ക മോഡലുകളിലാണ് സംഭവിക്കുന്നത്. കൂടുതൽ വിവരങ്ങൾ നിലവിൽ ലഭ്യമാകുന്ന ന്യൂക്ലിയസ് അംബുംബൻസുകളിൽ ശ്രദ്ധ കേന്ദ്രീകരിക്കുന്നു. പ്രധാനമായും, പോസ്റ്റ്മോർട്ടം തലച്ചോറിലെ പഠനങ്ങളെ അടിസ്ഥാനമാക്കി മനുഷ്യരുടെ അടിമകളായ ഈ മാറ്റങ്ങളിൽ ചിലത് സാധൂകരിക്കാൻ കഴിയുന്നുണ്ട്. മയക്കുമരുന്ന് ദുരുപയോഗം വ്യത്യസ്ത രാസഘടനകൾ ഉള്ളതും വ്യത്യസ്ത പ്രോട്ടീൻ ലക്ഷ്യങ്ങളിൽ പ്രവർത്തിക്കുന്നുണ്ടെങ്കിലും, പല പ്രമുഖ ആദിവാസികളുടെ അനുബന്ധ വിനിമയങ്ങളും പലപ്പോഴും സാധാരണമാണ്, ചില കേസുകളിൽ മയക്കുമരുന്ന് ഉപയോഗം ആസക്തി സിൻഡ്രോം.4,9 ഇതിനു വിപരീതമായി, മരുന്ന് ഉദ്വമനത്തിനായുള്ള അനേകം അനലോഡുകൾ ഒരു പ്രത്യേക മരുന്നിന്റെ പ്രത്യേകതയാണ്. മയക്കുമരുന്ന് ഉത്തേജകവും മയക്കുമരുന്നു ദുരുപയോഗം ചെയ്യുന്നതിലും ഞങ്ങൾ ശ്രദ്ധ കേന്ദ്രീകരിക്കുന്നു, മറ്റ് മരുന്നുകളുമായി താരതമ്യം ചെയ്യുമ്പോൾ മൃഗം മാതൃകകളിൽ കൂടുതൽ നാടകീയമായ ഫലങ്ങൾ ഉണ്ടാക്കുന്നു. ഞങ്ങൾ ഭാവി ഗവേഷണത്തിനായി പ്രധാന മേഖലകളെ ഉയർത്തിക്കാട്ടുന്നു. അത് ആഡംബര സിൻഡ്രോമിനെക്കുറിച്ചുള്ള നമ്മുടെ അറിവ് കൂടുതൽ വർദ്ധിപ്പിക്കുകയും മെച്ചപ്പെടുത്തിയ ഡയഗണോസ്റ്റിക് പരിശോധനകൾക്കും ചികിത്സകൾക്കും ഈ പുരോഗതിയെ വിവർത്തനം ചെയ്യുകയും ചെയ്യും.
ട്രാൻസ്ക്രിപ്ഷണലും എപിജിനറ്റിക് സംവിധാനങ്ങളും
വർഷങ്ങളായി ശാരീരിക ശേഷി ഇല്ലാതാകുന്ന അവസ്ഥയിൽ ആസക്തിക്ക് സാധ്യത കൂടുതലാണ്, മയക്കുമരുന്നിൽ ഉണ്ടാകുന്ന മയക്കുമരുന്ന് വ്യതിയാനങ്ങൾ വളരെ സ്ഥിരതയുള്ളതാണെന്നതാണ്. ജനിതക എക്സ്പെഷനിലെ മാറ്റങ്ങൾ ആദ്വേഷങ്ങളുടെ ഒരു പ്രധാന ഘടകമായി കണക്കാക്കാൻ ഇത് നിരവധി ഗ്രൂപ്പുകളെ പ്രേരിപ്പിച്ചു (ചിത്രം 1). ആയതിനാൽ, ഡി.എൻ.എ മൈക്രോഗ്രേകൾ ഉൾപ്പെടുന്ന ജനിതക വൈറസ് അന്വേഷണങ്ങളും അടുത്തിടെ ആർഎൻഎ-സെക് (ഉയർന്ന RNA- ന്റെ ഉയർന്ന ഉൽപാദന സിഗ്നിക്കൽ) മനുഷ്യരുടെ ആസക്തിയുടേയും എലിപ്പനെപ്പറ്റിയും പ്രാകൃതമായ മാതൃസംബന്ധിയായ മോഡലുകളിൽ നൽകിയിരിക്കുന്ന ഒരു തലച്ചോറിൻറെ രൂപത്തിൽ വ്യത്യാസമുണ്ടാക്കുന്ന നിരവധി ജീനുകളെ തിരിച്ചറിഞ്ഞിട്ടുണ്ട് (ഉദാഹരണം, 10-17 എന്നത്). ഈ പുനരവലോകനത്തിന്റെ തുടർന്നുള്ള ഭാഗങ്ങളിൽ അത്തരം ജീനുകളുടെ ഉദാഹരണങ്ങൾ ചർച്ചചെയ്യപ്പെടുന്നു.
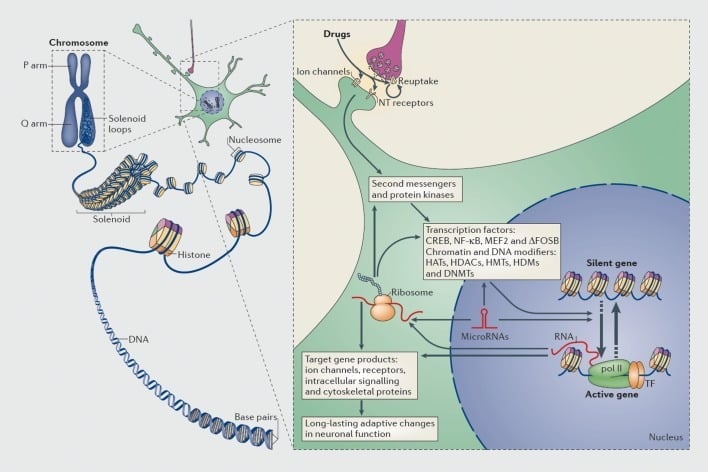
ദുരുപയോഗം ചെയ്യപ്പെട്ട മരുന്നുകൾ വഴി ട്രാൻസ്ക്രിപ്ഷണലും എപിജിനറ്റിറ്റി റെഗുലേഷന്റെ മാർഗ്ഗങ്ങളും. ഹിമാലയത്തിലെ ഒക്ടേമറുകൾ ചുറ്റിക്കൊണ്ട് ഡിസ്കോഎൻഎ സംഘടിപ്പിക്കുന്നത് ന്യൂക്ലിയോസോമുകൾ ഉണ്ടാക്കുന്നതിന് ഇടയാക്കി, അവ പിന്നീട് ഘടനാപരമായി ക്രോമസോം (ഇടത് ഭാഗം) രൂപം കൊള്ളുന്നു. ഒരു നിശ്ചിത ജീനിന്റെ ഡിഎൻഎ ട്രാൻസ്ക്രിപ്ഷണൽ മെഷിനറിയിലേക്ക് ആക്സസ് ചെയ്യുവാൻ കഴിയുമ്പോഴാണ് ക്രോമറ്റീന്റെ താൽക്കാലികമായി വിരളമായത്. മയക്കുമരുന്നിന്റെ സിഗ്നൽ കാസ്കേറ്റുകൾ (വലതുഭാഗം) മാറ്റാൻ പുനരാവിഷ് ഘടനകൾ, അയോൺ ചാനലുകൾ, ന്യൂറോ ട്രാൻസ്മിറ്റർ (എൻ.ടി) റിസപ്റ്ററുകൾ തുടങ്ങിയ സിനാപ്റ്റിക് ലക്ഷ്യങ്ങളിലൂടെ മയക്കുമരുന്നു ദുരുപയോഗം ചെയ്യുന്നു. ഇത് ട്രാൻസ്ക്രിപ്ഷൻ ഘടകങ്ങൾ (ടിഎഫ്എസ്), ക്രോമറ്റിൻ-റെഗുലേറ്ററി പ്രോട്ടീനുകൾ (കട്ടിയുള്ള അമ്പുകളുപയോഗിച്ച്) എന്നിവയുൾപ്പെടെയുള്ള മറ്റ് ആണവ ലക്ഷണങ്ങളുടെ പ്രവർത്തനക്ഷമതയോ അല്ലെങ്കിൽ തടസ്സപ്പെടുത്തുന്നതിലേക്കോ നയിക്കുന്നു; ക്രോമാറ്റിൻ-റെഗുലേറ്ററി പ്രോട്ടീനുകളുടെ സിനാപ്റ്റിക് നിയന്ത്രണത്തിൽ ഉൾക്കൊള്ളുന്ന വിശദമായ സംവിധാനങ്ങൾ വളരെ മോശമായി മനസ്സിലാക്കപ്പെടുന്നു. ഈ പ്രക്രിയകൾ അന്തിമമായി പ്രത്യേക ജീനുകളുടെ ഇൻഡക്ഷൻ അല്ലെങ്കിൽ അടിച്ചമർത്തലിന് കാരണമാകുന്നു, മൈക്രോ എൻ ആർ.എൻ. ഈ ജീനുകളിൽ ചില മാറ്റങ്ങളുടെ എക്സ്നെഷൻ ജീൻ ട്രാൻസ്ക്രിപ്ഷൻ തുടർന്നും നിയന്ത്രിക്കുവാൻ കഴിയും. ക്രോമറ്റിൻ തലത്തിലെ മയക്കുമരുന്നിന്റെ വ്യതിയാനം വരുത്തിയ മാറ്റങ്ങളിൽ ചിലത് വളരെ സ്ഥിരതയാർന്നതാണ്, അത് ആസക്തിയെ നിർവ്വചിക്കുന്ന ദീർഘകാല സ്വഭാവത്തിന് അടിവരയിടുന്നു എന്നാണ്. CREB, ചാക്രിക AMP- പ്രതികരിക്കുന്ന ഘടകമായ ബൈൻഡിംഗ് പ്രോട്ടീൻ; ഡിഎൻഎംടി, ഡിഎൻഎ മെത്തിyl ട്രാൻസ്ഫെറസ്; ഹാറ്റ്സ്, ഹിസ്റ്റോൺ അസറ്റൈൽ ട്രാൻസ്ഫറേഷനുകൾ; HDAC- കൾ, ഹിസ്റ്റോൺ ഡെയ്സിറ്റിലകൾ; HDM- കൾ, ഹിസ്റ്റോൺ ഡീമെട്രിക്സുകൾ; HMT- കൾ, ഹിസ്റ്റോൺ മിത്തീൽട്രാൻസ്ഫറൻസസ്; MEF2, മയോട്ടിപ്-സ്പെസിഫിക് എക്സൻസർ ഫാക്ടർ 2; NF-kB, ആണവ ഘടകം- KB; പോൾ രണ്ടാമൻ, ആർഎൻഎ പോളിളിമേസ് II. റിഫ്രഷ് 44 ൽ നിന്ന് പുനർനിർമ്മിച്ചത്: റോബൻസൺ എജെ, നെസ്റ്റ്ലർ ഇജെ. ആസക്തിയുടെ ട്രാൻസ്ക്രിപ്ഷണലും എപിജിനറ്റിക് സംവിധാനങ്ങളും. നാറ്റ് റെവ് ന്യൂറോസി. XXX- നം: 2011-12.
അതുപോലെ, പല തരത്തിലുള്ള ട്രാൻസ്ക്രിപ്ഷൻ ഘടകങ്ങൾ-ജീനുകളുടെ നിയന്ത്രിത മേഖലകളുമായി ബന്ധിപ്പിച്ച് പ്രോട്ടീൻ ജനിതകമാതൃകയിൽ ജനിതക പ്രയോഗത്തിൽ ദീർഘവീക്ഷണമുള്ള മരുന്നുകളുടെ ദീർഘകാല പ്രത്യാഘാതങ്ങളെ മധ്യസ്ഥതയിൽ ഉൾപ്പെടുത്തിയിട്ടുണ്ട്.. പ്രധാനപ്പെട്ട ഉദാഹരണങ്ങളിൽ CREB (CAMP പ്രതികരണ ഘടക ബാൻഡിംഗ് പ്രോട്ടീൻ), ΔFosB (ഒരു ഫാസ് കുടുംബത്തിന്റെ ട്രാൻസ്ക്രിപ്ഷൻ ഘടകം), എൻഎഫ്കെബി (ആണവോർജ്ജം കെ.ബി.), MEF2 (മയോസിറ്റ് എൻറോൾ ചെയ്ത ഫാക്ടർ- 2), ഗ്ലൂക്കോകോട്ടിക്കൈഡ് റിസപ്റ്ററുകൾ എന്നിവയും ഇതിൽ ഉൾപ്പെടുന്നു.5,10,18-22 ദുരുപയോഗം ചെയ്യുന്ന മരുന്നുകൾ തലച്ചോറിലെ ഒരു ട്രാൻസ്ക്രിപ്ഷൻ ഘടകത്തെ സജീവമാക്കുന്ന സെല്ലുലാർ സിഗ്നലിംഗ് പാതകളെ മനസിലാക്കുന്നതിനും അത്തരം ട്രാൻസ്ക്രിപ്ഷൻ ഘടകത്തിന്റെ ടാർഗെറ്റ് ജീനുകളുമായും ആസക്തിയുടെ പ്രത്യേക പെരുമാറ്റ വശങ്ങളുമായും അത്തരം ആക്റ്റിവേഷനെ ബന്ധിപ്പിക്കുന്നതിന് ഇത് കൂടുതൽ സാധ്യമാണ് (കാണുക ചിത്രം 1). ഈ പുരോഗതി ക്യുബി, ΔFosB എന്നിവയുടെ പരിഗണനയിലൂടെ ചിത്രീകരിക്കപ്പെടുന്നു. ആഡംബര മോഡലുകളിലെ മികച്ച പഠന ഗവേഷണ ഘടകങ്ങൾ ഇവയാണ്.
CAMP മറുപടി ഘടകം ബൈൻഡിംഗ് പ്രോട്ടീൻ
ദുരുപയോഗം ചെയ്യുന്ന ഉത്തേജക മരുന്ന് ഉത്തേജിത മരുന്നുകൾ സജീവമാക്കും ക്രെബി മസ്തിഷ്ക്കത്തിലെ പല ഭാഗങ്ങളിലും ആൺകുട്ടികളിൽ പ്രാധാന്യമുണ്ട്.23,24 ക്രാപ്, സി2+, വളർച്ച ഫാക്ടർ പാതകൾ,25 മയക്കുമരുന്ന് ദുരുപയോഗം ചെയ്തുകൊണ്ട് എൻഎസിയിൽ ഇത് പ്രവർത്തനക്ഷമമാക്കിയത് ഏത് മധ്യസ്ഥതയാണ്. NAC യിൽ CREB ൻറെ മയക്കുമരുന്ന് സജീവമാക്കൽ ഒരു ക്ലാസിക് നെഗറ്റീവ് ഫീഡ്ബാക്ക് സംവിധാനം സൂചിപ്പിക്കുന്നതാണ്, ഈ മരുന്നുകളുടെ പ്രതിഫലന ഫലങ്ങളോട് (സഹിഷ്ണുത) ഒരു മൃഗത്തിന്റെ സംവേദനക്ഷമത കുറയ്ക്കുന്നതിനും മയക്കുമരുന്ന് പിൻവലിക്കൽ (ആശ്രിതത്വം) സമയത്ത് ഒരു നെഗറ്റീവ് വൈകാരികാവസ്ഥയ്ക്ക് മധ്യസ്ഥത വഹിക്കുന്നതിനും CREB സഹായിക്കുന്നു..18,26,27 മയക്കുമരുന്ന് സ്വയം നിയന്ത്രിക്കലും പുനരാവിഷ്കരണവും വർദ്ധിപ്പിക്കാൻ ഈ ഫലങ്ങൾ സമീപകാലത്ത് കാണിച്ചു.28 ക്യുബിയുടെ ഈ പ്രവർത്തനങ്ങൾ എൻക മാദ്ധ്യമ സ്പൈനി ന്യൂറോണുകളുടെ പ്രധാന ഉപവിഭാഗങ്ങളും, പ്രധാനമായും ഡി1 ഡി2 ഡോപ്പാമൻ റിസപ്റ്ററുകൾ.24 Iഹിപ്പോകാമ്പസ്, അമാഗഡാല എന്നിവിടങ്ങളിൽ പ്രവർത്തിക്കുന്ന ക്രെബ്, പെരുമാറ്റ മെമ്മറിയിലെ ഒരു പ്രധാന തന്മാത്രയാണ്..29-31 ആസക്തിയുടെയും പെരുമാറ്റ സ്മരണയുടെയും ഈ വിശാലമായ പങ്കു് ഒരു ന്യൂറോണുകൾ നിരന്തരം മാറിക്കൊണ്ടിരിക്കുന്ന ഒരു പരിതസ്ഥിതിക്ക് അനുയോജ്യമായ ഒരു മോളിക്യൂളർ സംവിധാനത്തിലൂടെ ന്യൂറോണുകൾ കുടുങ്ങിക്കിടക്കുന്നു എന്ന വസ്തുത പ്രതിഫലിപ്പിക്കുന്നു.
ജനിതക അധിനിവേശം, തിരഞ്ഞെടുത്ത തിരഞ്ഞെടുപ്പുകൾ എന്നിവയിലൂടെ ഈ പെരുമാറ്റല്പരണിയിൽ ഇടപെടുന്ന ക്രെബിറ്റിനായുള്ള ടാർജെറ്റ് ജീനുകൾ തിരിച്ചറിഞ്ഞിട്ടുണ്ട്.10,18,32 ഒരു ഉദാഹരണം ഓപിഐയ്ഡ് പെപ്റ്റൈഡ് ഡൈനാഫീൻ: നാക് ന്യൂറോണുകളിൽ ഡൈൻഓർഫിൻ എക്സ്പ്രഷൻ ഉത്തേജിപ്പിക്കുന്ന ഉത്തേജനം, CREB മുഖേനയുള്ള മദ്ധ്യസ്ഥത, VTA ഡോപാമൈൻ ന്യൂറോണുകളിൽ കെ ഒപിഐഐഡിൻ റിസപ്റ്ററുകളുടെ ഡൈനാർഫിൻ സജീവമാക്കൽ ആക്കി, അതിനാൽ ഡോപാമമിർഗിക സംക്രമണം എൻ കിക്കു ഒപ്പം പ്രതിഫലം നൽകും.18 മയക്കുമരുന്ന് ഉത്തേജിത സിനാപ്റ്റിക് പ്ലാസ്റ്റിറ്റിക്ക് വേണ്ടി മറ്റ് നിരവധി ക്യുബിബി ടാർഗെറ്റുകൾ പ്രധാനപ്പെട്ടതാണെന്ന് കാണിച്ചു. മറ്റു തലച്ചോറ് പ്രദേശങ്ങളിലും ഉത്തേജക /23,24 ഈ ഫലത്തിന്റെ പെരുമാറ്റ പ്രത്യാഘാതങ്ങളെക്കുറിച്ചും അവ സംഭവിക്കുന്ന ടാർഗെറ്റ് ജീനുകളെക്കുറിച്ചും വളരെക്കുറച്ചേ അറിയൂ. അതുപോലെ, മറ്റ് മയക്കുമരുന്നുകളുടെ പ്രവർത്തനങ്ങൾക്ക് മധ്യസ്ഥത വഹിക്കുന്നതിൽ CREB വഹിക്കുന്ന പങ്കിനെക്കുറിച്ച് വളരെക്കുറച്ചേ അറിയൂ.19
ΔFosB
ഏതെങ്കിലും മരുന്ന് ദുരുപയോഗം ചെയ്യുന്നതിനുള്ള സൂക്ഷ്മമായ എക്സ്പോഷർ എൻ.സിയുടെയും മറ്റു തലച്ചോറിലെ എല്ലാ ഫാസ് കുടുംബത്തിൻറെ ട്രാൻസ്ക്രിപ്ഷൻ ഘടകങ്ങളെയും പ്രേരിപ്പിക്കുന്നു. ഈ ഉത്പാദനം ദ്രുതഗതിയിലുള്ളതും എന്നാൽ വളരെ താൽകാലികമായതുമാണ്, ഫോസ് പ്രോട്ടീൻ അളവ് സാധാരണ നിലയിലേക്ക് മാറുന്നു, ഇത് സാധാരണ മുതൽ രാവിലെ 9-10 വരെ മണിക്കൂറുകൾക്കുള്ളിൽ തുടരുന്നു. ഈ ഫോസ് ഫാമിലി പ്രോട്ടീനുകളിൽ വിവക്ഷിതമായത് ΔFosB ആണ്, FOSB ജീനിന്റെ ഒരു വെട്ടിയുണ്ടാക്കിയ ഉല്പന്നമാണ്, അത് അതിന്റെ അസാധാരണ സ്ഥിരതയുടെ ഫലമായി തുടർച്ചയായി മയക്കുമരുന്നിന്റെ അളവിലൂടെ സഞ്ചരിക്കുന്നു ഈ അവസ്ഥയിൽ പ്രകടമാക്കുന്ന പ്രബലമായ ഫോസ് പ്രോട്ടീൻ ആയിത്തീരുന്നു.22,33 മാത്രമല്ല, ഈ സ്ഥിരത കാരണം മരുന്നുകൾ പിൻവലിക്കാൻ ആഴ്ചകൾക്കുശേഷം ΔFosB നില കുറയ്ക്കുന്നു. എല്ലാ മയക്കുമരുന്ന് ദുരുപയോഗം ചെയ്യുന്നതിന് ΔFosB- യുടെ ഇത്തരം ദീർഘകാല പ്രവേശനം തെളിയിക്കപ്പെട്ടു34 മിക്ക മരുന്നുകളേയും Dl-type NAc ന്യൂറോണുകൾക്ക് തിരഞ്ഞെടുക്കാം.34,35 അതുതന്നെയായിരുന്നു മനുഷ്യ ആസക്തികളിൽ പ്രകടമാക്കി.35 ഡി എന്നതിൽ ΔFosB ഉളവാക്കുന്നതിൽ വലിയൊരു സാഹിത്യശാഖ കാണിച്ചിരിക്കുന്നു1-ടൈപ്പ് എൻഎസി ന്യൂറോണുകൾ മയക്കുമരുന്നിനോടുള്ള മൃഗത്തിന്റെ സംവേദനക്ഷമതയും സ്വാഭാവിക പ്രതിഫലവും വർദ്ധിപ്പിക്കുകയും മയക്കുമരുന്ന് സ്വയംഭരണത്തെ പ്രോത്സാഹിപ്പിക്കുകയും ചെയ്യുന്നു, ഇത് പോസിറ്റീവ് ബലപ്പെടുത്തൽ പ്രക്രിയയിലൂടെയാണ് (റെസ്പോൺസ്, 34 മുതൽ 38 വരെ കാണുക). രസകരമെന്നു പറയട്ടെ, എൻഎസിയിലെ ΔFOSB ന്റെ മരുന്നുകൾക്ക് കൗമാരപ്രായത്തിൽ കൂടുതൽ നാടകീയമായതിനാൽ,,39 നിക്കോട്ടിന്റെ പ്രേരണ നിക്കോട്ടിന്റെ ഗേറ്റ്വേ പോലുള്ള കൊക്കെയ്ൻ റിവാർഡ് വർദ്ധിപ്പിക്കുന്നതിന് മധ്യസ്ഥത വഹിക്കുന്നു.40
ക്രെബിബിനെ സംബന്ധിച്ചിടത്തോളം, ΔFosB- യ്ക്കായുള്ള ലക്ഷ്യം ജീൻ, ജിയോമോൺ-വൈഡ് സമീപനങ്ങളിലൂടെ NAC ൽ കണ്ടെത്തിയിട്ടുണ്ട്.10,32 CREB ഡൈനോർഫിൻ പ്രേരിപ്പിക്കുമ്പോൾ, osFosB അതിനെ അടിച്ചമർത്തുന്നു, ഇത് osFosB- യുടെ അനുകൂല പ്രതിഫല ഇഫക്റ്റുകൾക്ക് കാരണമാകുന്നു.38 മറ്റൊരു ΔFosB ടാർഗറ്റ് cFos ആണ്: ΔFosB തുടർച്ചയായി മയക്കുമരുന്നിന്റെ ഉൽപാദനക്ഷമതയോടെ സഞ്ചരിക്കുന്നു, ഇത് സി-ഫോസ് അടിച്ചമർത്തുന്നു, തന്മൂലം മയക്കുമരുന്നായി മാറുന്നു..41 എൻസി മാദ്ധ്യമ സ്പൈനി ന്യൂറോണുകളുടെ ഡൻഡറിറ്റിക്കൽ ആർബറൈസേഷനിൽ ഡി.എൻ.ആർ.ബി.ബി. ലക്ഷ്യങ്ങൾ എൻഎസിയിൽ സിനാപ്റ്റിക് പ്ലാസ്റ്റിറ്റിയിൽ ഉണ്ടാകാൻ സാധ്യതയുള്ള ചില മയക്കുമരുന്നുകളുടെ കഴിവ് മധ്യസ്ഥതയിൽ കാണിക്കുന്നു. താഴെ പറയുന്ന രീതിയിൽ ചർച്ച ചെയ്യപ്പെടും.
മറ്റ് തലച്ചോറിലെ പ്രദേശങ്ങളിൽ ΔFosB ഉദ്വമനത്തിന്റെ ഫലമായ പരിണാമം കുറച്ചുകൂടി നന്നായി മനസ്സിലാക്കാം, എന്നാൽ ഓർബിഫ്രോൺറ്റൽ കോർട്ടക്സിൽ (OFC) അതിന്റെ ഉദ്ധാരണം വിശദമായി പഠിച്ചിട്ടുണ്ടെങ്കിലും. ഇവിടെ, ΔFOSB ദീർഘകാല എക്സ്പോഷർ കാലഘട്ടത്തിൽ കൊക്കൈൻ മനഃശാസ്ത്രവിശകലനം-ഹാനികരമായ പ്രത്യാഘാതങ്ങൾക്ക് ഇടയാക്കുന്ന, സഹിഷ്ണുത നിലനിർത്താനും ഈ കൊളോണിയൽ വർദ്ധിപ്പിക്കുന്നത് കൊക്കെയ്ൻ സ്വയം നിയന്ത്രിക്കാനും.42,43
ജനിതക വൈവിധ്യമാർന്ന പരിണതഫലങ്ങൾ ഈ ഫലങ്ങളെ മധ്യസ്ഥമാക്കുന്ന നിരവധി സാധ്യതയുള്ള ജീനുകൾ നിർദ്ദേശിച്ചിട്ടുണ്ട്.42 Os ഫോസ്ബിയുടെ തനതായ താൽക്കാലിക സവിശേഷതകളും പരമ്പരാഗത മെമ്മറി സർക്യൂട്ടുകളിൽ (ഉദാ. ഹിപ്പോകാമ്പസ്) ഇത് പ്രചോദിപ്പിക്കപ്പെട്ടുവെന്ന അറിവും ഉണ്ടായിരുന്നിട്ടും, ഭാവിയിലെ ഗവേഷണത്തിനുള്ള രസകരമായ വിഷയമായ ബിഹേവിയറൽ മെമ്മറിയിൽ os ഫോസ്ബിയുടെ പങ്കിനെക്കുറിച്ച് ഇതുവരെ ഒരു പര്യവേക്ഷണവും നടന്നിട്ടില്ല.
എപിജിനറ്റിക് സംവിധാനങ്ങൾ
അടുത്തകാലത്തായി, എപ്പിജെനെറ്റിക്സിന് ഒരു പടി കൂടി മുന്നോട്ടകീഴെപ്പറ്റിയുള്ള പഠനം മുന്നോട്ടുവച്ചിട്ടുണ്ട്44 (കാണുക ചിത്രം 1), ഡിഎൻഎ ശ്രേണിയിലെ ഒരു മാറ്റത്തിന്റെ അഭാവത്തിൽ സംഭവിക്കുന്ന ജനിതക എക്സ്പ്രഷനിലെ മാറ്റമായി ഇത് വിശകലനം ചെയ്യാൻ സാധിക്കും. എലിജിനറ്റിക് സംവിധാനങ്ങൾ ഹൈഡ്രോണുകളുമായും മറ്റു പല തരം ആണവ പ്രോട്ടീനുകളുമായുള്ള പ്രതിപ്രവർത്തനം വഴി ഒരു സെൽ ന്യൂക്ലിസിലുള്ള ഡി.എൻ.എയുടെ പാക്കേജിംഗിനെ നിയന്ത്രിക്കുന്നു. ഇവ ക്രോമറ്റിൻ ഉൾക്കൊള്ളുന്നു. ഹിസ്റ്റോണുകൾ, മറ്റ് പ്രോട്ടീനുകൾ, ഡിഎൻഎ എന്നിവയുടെ സ്വയംഭരണമാറ്റത്തിലൂടെ ജീൻ പദപ്രയോഗമാണ് ഈ പാക്കേജിംഗ് സംവിധാനത്തിലൂടെ നിയന്ത്രിക്കുന്നത്. ചില ഉദാഹരണങ്ങൾ പോലെ, ഹിസ്റ്റോണുകളുടെ അസറ്റലീലേഷനും ജീൻ ആക്ടിവേറ്റേഷനെ പ്രോത്സാഹിപ്പിക്കുന്നതിനുമാണ്, ഹിസ്റ്റോകളുടെ മിഥിലേഷൻ ഈ മാറ്റം വരുത്തുന്നതിന് ലിസി അവശിഷ്ടം അനുസരിച്ച് ജീൻ ആക്ടിവേഷൻ അല്ലെങ്കിൽ അടിച്ചമർത്തൽ പ്രോത്സാഹിപ്പിക്കുകയും, ഡിഎൻഎയുടെ മിഥിലൈസേഷൻ സാധാരണയായി ജനിതക സമ്മർദ്ദവുമായി ബന്ധപ്പെട്ടതാണെങ്കിലും ചില വ്യത്യസ്ത രൂപമാതൃക മിഥിലേഷനുകൾ ഉദാഹരണത്തിന്, 5- ഹൈഡ്രോക്സൈമൈലീലൈഷൻ) ജീൻ ആക്റ്റിവേഷനുമായി ബന്ധപ്പെട്ടിരിക്കുന്നു.
എപ്പിജെനിറ്റിക്സ് ഒരു ആകർഷക സംവിധാനമാണ്, കാരണം മറ്റ് സിസ്റ്റങ്ങളിൽ, വികസനവും കാൻസറി ബയോളജിയും, ചില എപ്പീജനിറ്റി മാറ്റങ്ങൾക്ക് സ്ഥിരമായതായിരിക്കാം. ഈ കാരണത്താൽ, എപ്പിജെനിറ്റിക്സ് പഠനത്തിലും മെമ്മറി മോഡലുകളിലും (ഉദാഹരണം, 45-48) അതുപോലെ തന്നെ ആസക്തിയിലും പിന്തുടരുകയും ചെയ്തു;44,49 രണ്ടും സിസ്റ്റങ്ങളിൽ വളരെയധികം മാറ്റങ്ങൾ ഹിസ്റ്റോൺ അസീസിലേയും മിഥിലേഷനിലെയും ഡിഎൻഎയുടെ മിഥിലേഷനിൽ റിപ്പോർട്ട് ചെയ്തിട്ടുണ്ട്. ഒരു ഉദാഹരണം പോലെ, ഹിസ്റ്റോൺ methyltransferase, G9a, രണ്ട് മെമ്മറിയിൽ ഇൻക്രിപ്റ്റേറ്റഡ്50 ആസക്തി.51,52 ആഡംബര മോഡലുകളിൽ, G9a എക്സ്പ്രഷൻ ഒഴിവാക്കപ്പെട്ടിരിക്കുന്നു ഉത്തേജകവും ഉത്തേജകവും ഉത്തേജിതവും മയക്കുമരുന്ന് ഉപയോഗിച്ചുള്ള മരുന്നും പ്രതികരണമായി എൻഎസിയിൽഈ മരുന്നുകളുടെ പ്രതിഫലന ഫലങ്ങൾ മെച്ചപ്പെടുത്തുന്നതിന് അദ്ദേഹത്തിനു കഴിഞ്ഞിട്ടുണ്ട്.51,52 രസകരമായാൽ, G9A ന് കോകൈൻ അടിച്ചമർത്തലാണ് ΔFosB ഉപയോഗിച്ച് മധ്യസ്ഥത നൽകുന്നത്. ജീൻ അടിച്ചമർത്തലിന്റെ ഒരു പ്രധാന മധ്യസ്ഥനായ എച്ച്എസ്ടോൺ H9 (H9K3MXXX) ന്റെ ലൈസൻസിന്റെ അളവെടുപ്പ് G3a ആവുന്നു. Chip-chip അല്ലെങ്കിൽ ChIP-seq (പ്രോമോട്ടർ ചിപ്സ് അല്ലെങ്കിൽ ഹൈ-ഔട്ട്പുട്ട് സീക്വൻസിങ് വഴി യഥാക്രമം ക്രോമസറ്റ് ഇമിനോപ്പ്രസിപ്പിഷൻ പിന്തുടരുക) NAAC ലെ ജീനുകളുടെ വിന്യാസമാതൃകകൾ ലഭ്യമാക്കുന്നതിനായി ഉപയോഗിക്കുന്നു, ഇത് ഉത്തേജക / ഉത്തേജക പ്രഭാവത്തിനു ശേഷം H9K2XXX ൽ മാറ്റം വരുത്തുന്നു.32,52,53 ജനിതക എക്സ്പ്രഷൻ മാറ്റങ്ങളുടെ ജനിതക വൈവിധ്യമാർഗ ലിസ്റ്റുകളുമൊത്ത് ഈ ജീൻ ലിസ്റ്റുകൾ ഓവർലാപ്പുചെയ്യുന്നതിലൂടെ, എപ്പീഎൻഇജി രൂപത്തിലുള്ള മറ്റ് നിരവധി രൂപങ്ങളുടെ ജിയോമോൺ മാപ്പിംഗ് മാപ്പുകൾ (ഉദാ: ΔFosB ബൈൻഡിംഗ്, ക്യുബിബി ബൈൻഡ്, മറ്റ് ഹിസ്റ്റോൺ മാറ്റങ്ങൾ മുതലായവ)32,53 ദുരുപയോഗം നടത്തുന്ന മയക്കുമരുന്നുകൾ നിയന്ത്രിക്കുന്നതും പൂർണ്ണമായ അസ്ഥീജനിസം സംവിധാനങ്ങളെക്കുറിച്ചും മനസ്സിലാക്കുന്നതിനായി കൂടുതൽ വിപുലമായ ജീനുകളെ തിരിച്ചറിയാനും സാദ്ധ്യതയുണ്ട്.
സൂക്ഷ്മപരിശോധനയുടെ മറ്റൊരു രൂപമാണ് മെമ്മറിയിലും ആഡംബരത്തിലും ഉൾപ്പെട്ടിരിക്കുന്നത്. ഈ ചെറിയ, നോൺകോഡ് ചെയ്യുന്ന ആർഎൻഎകൾ mRNAs ന്റെ പരിമിതമായ പ്രദേശങ്ങളുമായി ബന്ധപ്പെടുത്തി, അവയുടെ വിവർത്തനം അടിച്ചമർത്തുന്നു, അല്ലെങ്കിൽ അവരുടെ തരം താഴ്ത്തലിനെ പ്രോത്സാഹിപ്പിക്കുന്നു. മൈഗ്എൻഎൻഎസുകളുടെ പ്രോസസ്സിംഗിനുള്ള ഒരു പ്രോട്ടീൻ ആർഗാനോട്ട് നീക്കം ചെയ്യൽ, കൊക്കെയ്നുമായുള്ള പെരുമാറ്റ പ്രതികരണങ്ങളെ മാറ്റിമറിക്കുന്നു. D1- നും D2- ടൈപ്പ് ഇടത്തരം സ്പൈനി ന്യൂറോണുകൾക്കും ബാധകമായ വ്യതിയാനങ്ങളാണവ.54 മയക്കുമരുന്നിന്റെ ഉപയോഗം നിയന്ത്രിക്കുന്നതിന് അനേകം പ്രത്യേക മിറാനുകൾ നൽകിയിട്ടുണ്ട്, കൂടാതെ മരുന്നുകൾക്ക് പെരുമാറ്റ പ്രതികരണങ്ങളെ സ്വാധീനിക്കാൻ (ഉദാഹരണം, 55,56). ഈ മിറാൻറുകളുടെ MRNA ടാർഗെറ്റുകളെ തിരിച്ചറിയാനും ഭാവിയിലെ ആസക്തി പ്രക്രിയയെ എങ്ങനെ ബാധിക്കുമെന്ന് തിരിച്ചറിയാനും ഇത് ഭാവിയിൽ പഠിക്കാം.
സിനാപ്റ്റിക് പ്ലാസിറ്റി
ഹിപ്പോകാറ്റഗ്രാസിനും അമാഗഡാലയിലെയും സ്വഭാവസവിശേഷതകളിലെ (ഈ വിഷയത്തിലെ മറ്റു ലേഖനങ്ങൾ കാണുക) ഉൾപ്പെട്ടിരിക്കുന്ന ഗ്ലൂറ്റാമറ്റജിക് സിനാപ്സുകളിൽ സമാന സിനാപ്ടിക് മാറ്റങ്ങൾ, സമാനമായ രീതിയിൽ ആസക്തി മോഡുകളിൽ തലച്ചോറുള്ള റിവാർഡ് പ്രദേശങ്ങളിൽ ഉണ്ടാകുന്നതിനും മധ്യസ്ഥതയിൽ പ്രാധാന്യം നൽകുന്നതിനും ആസക്തി പ്രക്രിയ.57,58 ഇത്തരം മയക്കുമരുന്ന് സിനാപ്റ്റിക് പ്ലാസ്റ്റിറ്റി പല തലച്ചോറ് പ്രദേശങ്ങളിലും വിവരിച്ചിട്ടുണ്ട്. എങ്കിലും, നാഷണൽ സെന്ററിൽ (ചിത്രം 2).
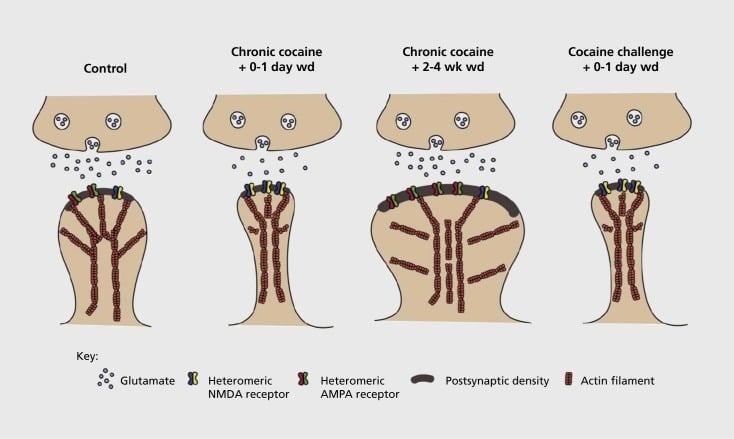
ആക്സിഡൻസിനോട് ബന്ധപ്പെട്ട സിനാപ്റ്റിക്, ഘടനാപരമായ പ്ലാസ്റ്റിക്സിറ്റി ന്യൂക്ലിയസ് അംബുംബൻസ് (എൻഎസി) മോഡൽ. കൊക്കെയ്നുമായുള്ള ദീർഘകാലമായുള്ള പ്രഭാവം α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA), എൻ-മെത്വി-ഡി-ആർപ്പാർഡിക് ആസിഡ് (എൻഎംഡി) ഗ്ലൂറ്റമാറ്റ് റിസപ്റ്ററുകൾ സ്പൈനി ന്യൂറോൺ (എംഎസ്എൻ) സിനാപ്സ്, അതുപോലെ നാവിഎംഎൻ എംഎസ്എസ്സിന്റെ നട്ടെല്ല് തലത്തിലുള്ള ഘടനാപരമായ മാറ്റങ്ങളാണ്. ഉദാഹരണത്തിന്, ദീർഘകാല കോക്കെയ്ൻ എൻഎംഡിഎ റിസപ്റ്ററുകൾ, നിശബ്ദമായ സിനാപ്സ് രൂപീകരണം, ദീർഘകാല വിഷാദരോഗം (ലിമിറ്റഡ്) എന്നിവയുടെ ആദ്യകാല പിൻവലിക്കൽ സമയ സൂചകങ്ങളിൽ ഉപരിതല expression തുടങ്ങുന്നു. കൂടുതൽ ദീർഘവീക്ഷണത്തോടെയുള്ള പിൻവലിക്കൽ (wd) സമയത്ത്, ഈ സിനാപ്റ്റിക് മാറ്റങ്ങൾ ഫലമായി റിവേഴ്സ് ഓഫ് എക്സ്പ്രഷൻ ഓഫ് എക്സ്പ്രെഷൻ ഓഫ് പ്ലാറ്റ്ഫോം AMPA റിസപ്റ്ററുകൾ, സിനാപ്സുകളുടെ ഏകീകരണം കൂൺ ആകൃതിയിലുള്ള നട്ടെല്ല്, ദീർഘകാല ഊർജ്ജം (LTP). ഈ പ്രതിഫലങ്ങൾ കൊക്കെയ്ൻ വെല്ലുവിളി ഉയർത്താൻ വീണ്ടും വീണ്ടും തിരിച്ച്, നട്ടെല്ല് മുള്ളുകളിലേക്ക് പുനർനിർമ്മിക്കുന്നതിനും സിനാപ്റ്റിക് ശക്തിയെ വിഷാദമാക്കുന്നതിനും കാരണമാകുന്നു.
ദുരുപയോഗം ഉത്തേജിപ്പിക്കുന്ന മയക്കുമരുന്നുകളെക്കുറിച്ചുള്ള ആവർത്തിച്ചുള്ള വെളിപ്പെടുത്തൽ ഒരു ലിമിറ്റഡ് (ദീർഘകാല വിഷാദരോഗം) -നാഷണൽ ലെ ഗ്ലൂറ്റാമറ്റജിക് സിനാപ്സുകളിൽ സംസ്ഥാനത്തെ ഉളവാക്കുന്നു എന്ന് ആദ്യകാല പരീക്ഷണങ്ങൾ തെളിയിച്ചു.59 എന്നിരുന്നാലും, അടുത്ത കാലത്ത് ഇത്തരം പ്ലാസ്റ്റിക്ക് കൂടുതൽ സമയാധിഷ്ഠിതമായിരിക്കണമെന്ന് തെളിഞ്ഞു. കഴിഞ്ഞ കോകൈൻ എക്സ്പോഷർ കൂടുതൽ LTC (ദീർഘകാല ഊർജ്ജം) ആയി മാറുന്ന ഉടൻതന്നെ, LDL ദീർഘകാല പിൻവലിക്കൽ ശേഷിക്ക് ശേഷം സംഭവിച്ചുകൊണ്ടിരിക്കുകയായിരുന്നു.60,61 സ്വയം നിയന്ത്രിക്കപ്പെട്ട മരുന്നുകൾക്ക് എതിരായി നൽകിയ ഗവേഷണത്തെയാണ് പ്രധാനമായും ഉപയോഗിക്കുന്നത്. ഈ സിനാപ്റ്റിക് പ്ലാസ്റ്റിക്ക് രൂപകൽപ്പന ചെയ്യുന്ന സ്വയംഭരണ സംവിധാനങ്ങളിൽ കൂടുതൽ സിസ്റ്റമാറ്റിക് അന്വേഷണങ്ങൾ ആവശ്യമാണെന്ന് നിർവചിക്കപ്പെട്ടിട്ടുണ്ട്. സ്വയംഭരണം നിലനിർത്തുന്നതിൽ നിന്നും, പിൻവലിക്കൽ, വംശനാശം എന്നിവയ്ക്കായി വിവിധ സമയങ്ങളിലൂടെ, പുനരാവിഷ്കരണത്തിനും ഉത്തേജനം നൽകുന്നതിനുമുള്ള വിശദമായ സമയ പരിധിയിൽ എൻ.ഇ.സി. ഈ മയക്കുമരുന്ന് ഉത്തേജിത സിനാപ്റ്റിക് പ്ലാസ്റ്റിറ്റിയിലേയ്ക്ക് സംഭാവന ചെയ്യുന്ന ചില മോളികുലാർ മെക്കാനിസങ്ങൾ കാലാകാലങ്ങളിൽ നിർവചിക്കപ്പെട്ടിട്ടുണ്ട്. ഇതിൽ AMPA റിസപ്റ്ററുകളെ കടത്തലിലേക്ക് കടത്തുക, ചിലപ്പോൾ മധ്യഭാഗം CaMKII (Ca2+/ calmodulin-dependent protein kinase II) ചില AMPA റിസപ്റ്ററുകളുടെ ഉപോയ്സ്റ്റുകളുടെ ഫോസ്ഫോരിലേഷൻ, AMPA റിസപ്റ്ററിന്റെ സബ്ട്ടൂട്ടുകളുടെ മാറ്റം വരുത്തിയ എക്സ്പ്രഷൻ (ഉദാഹരണത്തിന്, 60,62-65, ചിത്രങ്ങളും 2, 3). ക്രെബിബിയ്ക്കും ΔFosB യ്ക്കും ഈ പ്രതിഭാസത്തിൽ പങ്കുണ്ട്. അതുപോലെ ഗ്ലൂറ്റമാറ്റർസിക് സിനാപ്സുകളുടെ മോർഫോളജിയിൽ (താഴെ കാണുക) ബന്ധപ്പെട്ട മാറ്റങ്ങളുമായി ബന്ധപ്പെട്ടിരിക്കുന്നു. ഉദാഹരണത്തിന്, എൻഎസിയിലെ ക്യുബിഎൽ ലക്ഷ്യം ഗ്ലുവൽ ഗ്ലോഗ്എൻഎൻഎക്സ്എക്സ്എക്സ്എക്സ്എക്സ്.എക്സ്.എഫ്.എഫ്.ഒ.യുടെ രണ്ട് ലക്ഷ്യങ്ങളാണുള്ളത്. ഈ തലച്ചോറിൽ .35,36,66,67 മുന്നോട്ട് നീങ്ങുന്നതിനു്, സിനാപ്റ്റിക് ഫംഗ്ഷനിൽ സമയബന്ധിതമായ മാറ്റങ്ങൾ, ആസക്തിയുടെ പെരുമാറ്റ വിശേഷതകൾ എന്നിവയിൽ പ്രത്യേക ആധികാരികതകൾ ലിങ്ക് ചെയ്യുന്നതു് പ്രധാനമായിരിയ്ക്കും.
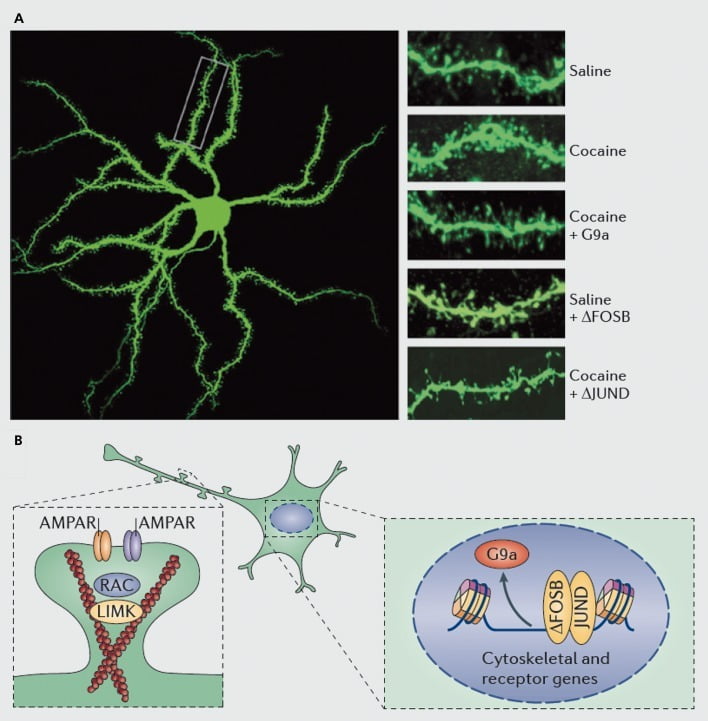
ന്യൂക്ലിയസ് അക്കുമ്പെൻസിൽ (എൻഎസി) ഇടത്തരം സ്പൈനി ന്യൂറോണുകളിൽ ഡെൻഡ്രൈറ്റിക് മുള്ളുകളുടെ കൊക്കെയ്ൻ ഇൻഡക്ഷന് അടിസ്ഥാനമായ തന്മാത്രാ സംവിധാനങ്ങൾ. എ) ഡെൻഡ്രിറ്റിക് നട്ടെല്ല് സംഖ്യയിൽ കൊക്കെയ്ൻ-ഇൻഡ്യൂസ്ഡ് വർദ്ധനവ് കാണിക്കുന്നു, അത് ജി 9 എ അല്ലെങ്കിൽ ജുൻഡിയുടെ വൈറൽ അമിതപ്രയോഗം വഴി തടയാൻ കഴിയും (എപി 1-മീഡിയേറ്റഡ് ട്രാൻസ്ക്രിപ്ഷന്റെ എതിരാളി), അല്ലെങ്കിൽ ഫോസ്ബിയുടെ വൈറൽ അമിതപ്രയോഗം അനുകരിക്കുന്നു. ബി) എഎംപിഎ റിസപ്റ്റർ (എഎംപിആർ) കടത്തലിന്റെയും ആക്റ്റിൻ സൈറ്റോസ്ക്ലെട്ടന്റെയും (ഇടത്) നിയന്ത്രണം, അതുപോലെ ഗ്ലൂട്ടാമേറ്റ് റിസപ്റ്ററുകളുടെയും ആക്റ്റിൻ റെഗുലേറ്ററി പ്രോട്ടീനുകളുടെയും ട്രാൻസ്ക്രിപ്ഷൻ നിയന്ത്രണം (ഉദാ. Os ഫോസ്ബി വഴി മധ്യസ്ഥത വഹിക്കുന്നത്, വലത്) പ്രധാന പങ്ക് വഹിക്കുന്നു. എൻഎസി ഡെൻഡ്രിറ്റിക് നട്ടെല്ല് സാന്ദ്രതയെ കൊക്കെയ്ൻ നിയന്ത്രിക്കുന്നതിന് മധ്യസ്ഥത വഹിക്കുന്നതിൽ. യുഎംകെ, എൽഐഎം ഡൊമെയ്ൻ കൈനാസ്; ആർഎസി, റാസുമായി ബന്ധപ്പെട്ട സി 3 ബോട്ടുലിനം ടോക്സിൻ സബ്സ്ട്രേറ്റ്.
പുതിയ പരീക്ഷണാത്മക ഉപകരണങ്ങൾ ആദ്യ ഘട്ടത്തിൽ കൃത്യമായ സർക്യൂട്ടുകൾ ഈ സിനാപ്റ്റിക് പ്ലാസ്റ്റിക്ക് പ്രകടമാക്കുകയും അവർ എങ്ങനെ ഇടപെടുന്ന പെരുമാറ്റ വൈകല്യങ്ങൾ പ്രദർശിപ്പിക്കുകയും ചെയ്യുന്നു എന്നത് കൃത്യമായി വ്യക്തമാക്കുന്നു. ഉദാഹരണം, tമയക്കുമരുന്ന് ഉത്തേജിത സിനാപ്റ്റിക് പ്ലാസ്റ്റിറ്റിയിൽ നാക് ഡിസ്പ്ലെയിലെ വ്യത്യാസങ്ങളെ അദ്ദേഹം ഷെല്ലും കോർ സബ്മിരിയനുകളും ചെയ്യുന്നു. ഓരോ subregion ലെ D1- നും D2- ടൈപ്പ് ഇടത്തരം സ്പൈനി ന്യൂറോണുകൾ പോലെ.60,63,64,67 അതുപോലെ, optogenetic പരീക്ഷണങ്ങൾ NAAC ലെ ഗ്ലൂറ്റാമറ്റജിക് സിനാപ്സുകളുടെ പ്രത്യേക ജനസംഖ്യയിൽ ഒരു പ്രത്യേക സിനാപ്റ്റിക് പ്ലാസിറ്റി (ഉദാഹരണത്തിന്, എ.ടി.,) യുടെ സംഭാവനയെക്കുറിച്ചുള്ള നോവൽ ഉൾക്കാഴ്ച നൽകുകയും, ഉദാഹരണത്തിന്, ഇടത്തരം PFC ഉം ബഷോലറ്റൽ അമീഗഡാല ആൻഡ് വ്രെൽറൽ സബ്ക്യുലവും (പ്രധാന ഔട്പുട്ട് ഹിപ്പോകാമ്പസ്).68-70 ആത്യന്തികമായി, ഈ അനുബന്ധ ന്യൂറോണുകളിൽ മയക്കുമരുന്ന്-പ്രേരിപ്പിച്ച തന്മാത്രാ അഡാപ്റ്റേഷനുകളെ അവയുടെ പോസ്റ്റ്നാപ്റ്റിക് ഡെൻഡ്രൈറ്റുകളിൽ സംഭവിക്കുന്ന സിനാപ്സ്-നിർദ്ദിഷ്ട അഡാപ്റ്റേഷനുകളുപയോഗിച്ച് ഓവർലേ ചെയ്യേണ്ടത് ആവശ്യമാണ്. ആസക്തി നിറഞ്ഞ അവസ്ഥ. ഇന്നുവരെ വളരെ കുറച്ച് ശ്രദ്ധ നേടിയ ഈ പ്രദേശമായ ഇതേ മസ്തിഷ്ക മേഖലകളിലെ തടസ്സം സൃഷ്ടിക്കുന്ന സിനാപ്സുകളിൽ മയക്കുമരുന്ന് പ്രേരണയുള്ള പ്ലാസ്റ്റിറ്റിയെക്കുറിച്ച് ഈ ശ്രമത്തിന് കൂടുതൽ വിലമതിപ്പ് ആവശ്യമാണ്.65
മുഴുവൻ സെൽ പ്ലാറ്റിവിറ്റി
മയക്കുമരുന്ന് ദുരുപയോഗം ചെയ്യുന്ന പ്രതിഭാസങ്ങളിലെ ന്യൂറോഫിസിയോളജിക്കൽ മാറ്റങ്ങളെ സംബന്ധിച്ച ഗവേഷണങ്ങളിൽ ഭൂരിഭാഗവും പഠനത്തിലും മെമ്മറി പ്രതിഭാസത്തിലുമുള്ള പോലെ സിനാപ്റ്റിക് പ്ലാസ്റ്റിറ്റിയിൽ ശ്രദ്ധ കേന്ദ്രീകരിച്ചിട്ടുണ്ട്. മുഴുവൻ സെൽ പ്ലാസിറ്റി പ്രാധാന്യം വർദ്ധിക്കുന്നതിനുള്ള തെളിവുകൾ ലഭ്യമാണ്. ഹോമിയോസ്റ്റാറ്റി പ്ലാസ്റ്റിക്ക് എന്ന് വിളിക്കപ്പെടുന്ന മുഴുവൻ സെൽ പ്ലാസ്റ്റിറ്റി,71 ഒരു സിനാപ്സ് സ്പെക്ട്രം അല്ലാത്ത രീതിയിലുള്ള ഒരു മുഴുവൻ സെല്ലിന്റെ ആന്തരിക ആവേശത്തിൽ മാറ്റം വരുത്തുന്നത്. മയക്കുമരുന്നിന്റെ ചില പ്രത്യേകതകൾ മയക്കുമരുന്നിന് മെച്ചപ്പെട്ടതോ കുറച്ചതോ ആയ സെൻസിറ്റിവിറ്റി ഉൾപ്പെട്ടതാകുമ്പോൾ, ചില നാഡികളിലെ ഇലക്ട്രോണിക് അമറലൈറ്റിന്റെ മെച്ചപ്പെട്ടതോ അല്ലെങ്കിൽ കുറച്ചതോ ആയ പെരുമാറ്റ രീതികൾ.5
ദി ഒരു സെൽ പ്ലാസിറ്റിയെ മുഴുവനായും വിശകലനം ചെയ്യുന്ന മയക്കുമരുന്നിന്റെ ഏറ്റവും മികച്ച ഉദാഹരണമാണ് ലോക്കസ് കോറോലൈസിന്റെ നഡാഡ്രൻസർക് ന്യൂറോണുകളുടെ ആന്തരിക ആവേശം വർദ്ധിപ്പിക്കുന്നതിന് വിട്ടുമാറാത്ത ഓപിയേറ്റുകൾ).72 ഈ വർദ്ധിച്ച ഉണർവ്വ് CREB മുഖേനയും മധ്യത്തിലുള്ള Adenylyl സൈക്ലെയ്സിന്റെ ചില ഐസോഫോറുകളുടെ പ്രചോദനവും, N + ചാനലുകളുടെ ഉദ്ധരണികൾ വഴി LC ന്യൂറോണുകളുടെ വെടിവയ്പ്പ് വർദ്ധിപ്പിക്കും.72-75 LC ന്യൂറോണുകളുടെ ഈ ഹൈപ്രെക്സ്സൈറ്റിറ്റൽ സഹിഷ്ണുതയുടെയും ആശ്രിതത്വത്തിന്റെയും ക്ലാസിക് സംവിധാനത്തെ പ്രതിനിധാനം ചെയ്യുന്നു, ഒപ്പം ഒപ്പിട്ട് പിൻവലിക്കലിന്റെ ലക്ഷണങ്ങളും ലക്ഷണങ്ങളും വലിക്കുന്നു. സൂക്ഷ്മതരംഗ നാഷണൽ സ്പിനീഷ് ന്യൂറോണുകളിൽ സമാനമായ ഒരു സെൽ പ്ലാസ്റ്റിറ്റിയിൽ ക്രെബിറ്റ് മധ്യസ്ഥത വഹിക്കുന്നുണ്ട്, ക്രെബിറ്റ് ഉപയോഗിച്ചുള്ള മയക്കുമരുന്ന് രോഗബാധിതമായ ദീർഘദൂരങ്ങളിലൂടെ ഇവ വ്യാപകമാക്കപ്പെടുന്നു.76 സൂക്ഷ്മതലത്തിലുള്ള സ്പൈനി ന്യൂറോണുകളിൽ ഗ്ലൂറ്റ്മാറ്റേജിക് സിനാപ്സുകളുടെ ക്രെബ്-മദ്ധ്യേഷ്യൻ സിനാപ്റ്റിക് പ്ലാസ്റ്റിറ്റി എങ്ങിനെ മനസ്സിലാക്കുന്നു എന്ന് മനസ്സിലാക്കാൻ ഭാവിയിലെ അന്വേഷണങ്ങൾ ഇത് വളരെ വിമർശനാത്മകമായിരിക്കും65,66 ഈ ന്യൂറോണുകളുടെ ക്രെംബിക-മധ്യസ്ഥത ആന്തരിക ഹൈപ്പർക്രീസിബലിറ്റിയുമായി ഉദ്ധരിക്കുന്നു76 ആസക്തിയുടെ സ്വഭാവ സവിശേഷതകളെ നിയന്ത്രിക്കുന്നതിന്.
ആഡംബര മോഡലുകളുടെ മുഴുവൻ സെൽ പ്ലാസിറ്റിയുടെ മറ്റൊരു ഉദാഹരണം VTA ഡോപ്പാമൈൻ ന്യൂറോണുകളുടെ ഹൈപ്പറൈസിസിറ്റബിളിറ്റാണ്, ഇത് ദീർഘകാലത്തെ മയക്കുമരുന്നുകൾe (ചിത്രം 4).77,78 ഈ നാഡീകോശങ്ങളിലുള്ള മോർഫോളജിക്കൽ മാറ്റങ്ങളുമായി ബന്ധപ്പെട്ടിരിക്കുന്ന ഈ ഉപവിഭാഗം (അടുത്ത വിഭാഗം കാണുക), CREB ക്ക് മധ്യസ്ഥതയല്ല, പകരം ന്യൂറോട്രോപിക് സിഗ്നലിംഗ് കാസ്കറ്റുകൾ നിയന്ത്രണം വഴി നേടിയെടുക്കുകയാണ്, താഴെ വിവരിച്ചിരിക്കുന്നത് പോലെ.

ദ്ട്രാമിൻ ന്യൂറോണുകൾ ventral tegmental പ്രദേശത്ത് (VTA) ലെ വിട്ടുമാറാത്ത മോർഫിൻ-ഇൻഡുഡന്റ് അഡാപ്റ്റേഷന്റെ വർക്കിങ് മോഡൽ. വിട്ടുമാറാത്ത മോർഫിൻ കുറയുന്നു VTA dopamine (DA) soma size ഇനിയും ന്യൂറോണൽ ആവേശം വർദ്ധിക്കുന്നു, അതേസമയം ന്യൂക്ലിയസ് accumbens ലേക്കുള്ള ഡോപ്പാമൻ സംക്രമണം കുറയുന്നു. മോർഫിൻ എന്നതിന്റെ ഫലത്തിൽ, പ്രതികൂല പ്രതിഫലനപാത, അതായത് പ്രതിഫലന ടോളറൻസ് ആണ്. VTA ൽ IRS2-AKT സിഗ്നലിങ്ങിൽ ഡ്രോഗ്ഗ്രൂപ്പ് ചെയ്യുന്നത് സോമാ സൈറ്റിലും ഇലക്ട്രിക്കൽ ആവൃത്തിയിലും വിട്ടുമാറാത്ത മോർഫിൻ ഫലങ്ങളെ ഇടപെടുക്കുന്നു; കുറഞ്ഞ γ- അമിനോബ്യൂട്ടിക് ആസിഡ് (GABA) വഴിയാണ് എക്സിബിറ്റബിളിറ്റിയുടെ പ്രഭാവം മധ്യസ്ഥമാക്കുന്നത്. കെ ചാനൽ എക്സ്പ്രഷന്റെ പ്രവാഹങ്ങളും അടിച്ചമർത്തലും. ഈ മോർഫിൻ ഉളവാക്കുന്ന മോർഫോളജിക്കൽ, ഫിസിയോളജിക്കൽ ലാപ്ടോപ്പുകൾക്കും റിവാർ ടോളറൻസ് നൽകുന്നവർക്കും VTA ലെ mTORC2 പ്രവർത്തനത്തിന്റെ മോർഫിൻ ഉൽപാദനം നിർണ്ണായകമാണ്. MT0RC2 വിരുദ്ധമായി, കോർണിൻ മോർഫിൻ mTORCI പ്രവർത്തനം വർദ്ധിപ്പിക്കുന്നു, ഇത് ഈ മോർഫിൻ പ്രേരിപ്പിച്ച അഡാപ്റ്റേഷനുകളെ സ്വാധീനിക്കുന്നില്ല. ബി.ഡി.എൻ.എഫ്, ബ്രെയിൻ ഡിറൈവ്ഡ് ന്യൂറോട്രോപിക് ഘടകം; ഇൻസുലിൻ റിസപ്റ്റർ വസ്തുവായ IRS; mTORC, mTOR സങ്കീർണ്ണത; എ.കെ.ടി, പ്രോട്ടീൻ കിനിസ് ബി ഉപ്പ് 77 ൽ നിന്ന് പുനർനിർമ്മിച്ചത്
ശാരീരിക പ്ലാസ്റ്റിറ്റി, ന്യൂറോട്രോപിക് സംവിധാനങ്ങൾ
ഹിപ്പോകാമ്പാലിന്റെയും സെറിബ്രൽ കോർട്ടിക്കൽ ന്യൂറോണുകളുടെയും പഠനങ്ങളിൽ നിന്നും അതിലേറെ തെളിവുകൾ വർദ്ധിക്കുന്നതായി സിനാപ്റ്റിക് പ്ലാസ്റ്റിക്യിൽ സംഭവിച്ച മാറ്റങ്ങൾ സിനാപ്സസിലെ മോർഫോളജിക്കൽ മാറ്റങ്ങളുമായി ബന്ധപ്പെട്ടതാണെന്ന് തെളിയിച്ചിരിക്കുന്നു. ഉദാഹരണത്തിന്, ലിമിറ്റഡ്, നിശബ്ദമായ സിനാപ്സുകളുടെ ഉത്പന്നം എന്നിവ നേർത്തതോ മണ്ടത്തരമോ ആയ ഡൈൻറിക്റ്റിക് മുള്ളുകളുടെ രൂപവത്കരണവുമായി ബന്ധപ്പെട്ടിരിക്കുന്നു. എൽ.ടി.പി വലിയ, കൂൺ ആകൃതിയിലുള്ള മുള്ളുകളുമായി ബന്ധപ്പെട്ടിരിക്കുന്നു.79,80 > 15 വർഷമായി ഡെൻട്രിറ്റിക് മുള്ളുകളിൽ മയക്കുമരുന്ന് പ്രേരണയുള്ള മാറ്റങ്ങളിൽ മയക്കുമരുന്ന് ഉപയോഗ മേഖല ശ്രദ്ധ കേന്ദ്രീകരിച്ചിരിക്കുന്നു എന്നത് രസകരമാണ്. ദീർഘകാല എക്സ്പോഷർ ദുരുപയോഗം ഉത്തേജിപ്പിക്കുന്ന മരുന്നുകൾ ഡിഎൻ-ടൈപ്പ് ന്യൂറോണുകൾക്ക് മുൻതൂക്കമുള്ള ഒരു മാറ്റത്തിന്റെ നാഷണൽ സ്പിന്നിക്ക് ന്യൂറോണുകളുടെ ഡൻഡറിക് സ്പിൻ ഡെൻസിറ്റി വർദ്ധിപ്പിക്കുന്നു..67,81,82 മരുന്നുകളുടെ സ്പന്ദന സ്പെസിഫിക്കേഷൻ പ്രതികരണങ്ങളുള്ളതിനാൽ മുള്ളുകളുടെ ഇൻഡിക്കേറ്റർ അനേകം ഭാഗങ്ങളായി ബന്ധപ്പെട്ടിട്ടുണ്ട്. ചില തെളിവുകൾ ഈ കാഴ്ചപ്പാടുമായി വൈരുദ്ധ്യമുണ്ടാക്കുന്നു.
സിനാപ്റ്റിക് പ്ലാസ്റ്റിറ്റിയിൽ പഠിക്കുന്നതുപോലെ, മയക്കുമരുന്ന് സ്വയം നിയന്ത്രിക്കൽ, പിൻവലിക്കൽ, പുനരധിവാസം എന്നീ ഘട്ടങ്ങളിൽ ഉണ്ടാകുന്ന dendritic spines ലെ മാറ്റം വ്യവസ്ഥാപിതമായി നിർവചിക്കേണ്ടതുണ്ട്. Sഅന്വേഷണക്കാരനും സ്വയം നിയന്ത്രിക്കപ്പെട്ട മയക്കുമരുന്നും ഉൾപ്പെടുന്ന ട്യൂഡുകളിലൂടെ, വ്യത്യസ്ത പിൻവലിക്കൽ സമയ പോയിന്റുകളിൽ ഉണ്ടാകുന്ന വളരെ വ്യത്യസ്തമായ നട്ടെല്ല് മാറ്റങ്ങളും നാഷണൽ ഷെല്ലുകൾക്കും കോർ സബ് റീജിയുകളിലും.83-86 കൊക്കൈൻ അല്ലെങ്കിൽ മറ്റൊരു ഉത്തേജനം ഈ സമയ-ആശ്രിതവും സെൽ-ടൈപ്പ് പ്രത്യേക സവിശേഷതകളും ഉൽപാദിപ്പിക്കുന്ന കൃത്യമായ മോളികുലാർ സംവിധാനങ്ങളെ നിർവചിക്കുന്നതും പ്രധാനമാണ്. ΔFOSB ഡിഎൽ-ടൈപ്പ് NAc ന്യൂറോണുകളിൽ പക്വതയില്ലാത്ത മുള്ളുകൾ ഉണ്ടാക്കുന്നതിനാവശ്യമായ പര്യാപ്തവും പര്യാപ്തവുമാണ്..35,51,67 ആക്ടിൻ സൈക്കോസ്കെലിടൺ പുനർക്രമീകരിക്കുന്നത് നിയന്ത്രിക്കുന്നതിന് ധാരാളം പ്രോട്ടീനുകളുടെ കൊക്കൈൻ, ΔFOSB എന്നിവയുടെ നിയന്ത്രണത്തിലാണ് ഇത്തരം നിയന്ത്രണം ഉണ്ടാകുന്നത്. ഒരു ഉദാഹരണം എന്ന നിലയിൽ, നിരവധി ഗ്വാളിൻ ന്യൂക്ലിയോടൈഡ് എക്സ്ചേഞ്ച് ഘടകങ്ങളുടെ ട്രാൻസ്ക്രിപ്ഷണൽ നിയന്ത്രണവും, GTPase പ്രോട്ടീനുകൾ പ്രോത്സാഹിപ്പിക്കുന്നതും ഓരോ കൊക്കൈൻ എക്സ്പോഷണലിനുമുള്ള പ്രവർത്തനത്തിൽ തകരാറിലായി കുറയ്ക്കാൻ റേകൺഎൻഎൻഎക്സ്, ഒരു ചെറിയ GTPase, ഒപ്പം, റേക്എക്സ്എക്സ്എക്സ് പ്രവർത്തനത്തിലെ ഇത്തരം pulsatile കുറവ്, optogenetic control റാങ്കിംഗിൽ നിന്നുള്ള, മിതത്വ മുള്ളുകളുടെ ഇൻഡക്ഷൻ.87 റാഫ്റ്റിന്റെ ഈ പ്രഭാവം കോഫിയിനുകളുടെയും മറ്റ് ആക്ടിൻ റെഗുലേറ്ററി പ്രോട്ടീനുകളുടെയും നിയന്ത്രണം മൂലം ഉണ്ടാകുമായിരുന്നു, അത് നട്ടെല്ലിൻറെ വളർച്ചയുടെ കൊക്കെയ്ൻ നിയന്ത്രണം ഇടപെടുന്നതായി കാണിക്കപ്പെട്ടിരിക്കുന്നു.87,88 എന്നിരുന്നാലും, കൊക്കെയ്ൻ പക്വതയില്ലാത്ത മുള്ളുകളെ നിയന്ത്രിക്കുന്നതിൽ ഉൾപ്പെട്ടിട്ടുള്ള ഒരു പാത മാത്രമാണിതെന്ന് to ന്നിപ്പറയേണ്ടത് പ്രധാനമാണ്, കാരണം മറ്റ് നിരവധി പ്രോട്ടീനുകളും സിഡികെ 5 (സൈക്ലിൻ-ആശ്രിത കൈനാസ് -5), കാംകെഐഐ, എൻഎഫ്കെബി എന്നിവയുൾപ്പെടെ ഒരു പ്രധാന പങ്ക് വഹിക്കുന്നു. , MEF2, CREB, G9a, DNMT3 (DNA methyltransf മായ്ക്കൽ 3a), കുറച്ച് പേരിടാൻ.20,21,35,51,67,89,90 സി.എക്സ്.കെ.എക്സ്.എം.എക്സ്, ക്യു.എൻ.യു.ഐ., എൻ.എഫ്.കികൾ, എൻഎക്സ്എസിബി ഉൾപ്പെടുത്തൽ, ജി.എക്സ്.എൻ.എ.എൻ.എ. അടിച്ചമർത്തൽ എന്നിവയും ഉൾപ്പെടെയുള്ള ജീനുകളുടെ കൊക്കെയ്ൻ നിയന്ത്രണം ΔFosB വഴിയാണ് സാധ്യമാകുന്നത്.20,35,51,91
അതിശയകരമെന്നു പറയട്ടെ, അമിതമായ മയക്കുമരുന്നുകൾ വിപരീത ഫലത്തെ സ്വാധീനിക്കുകയും എൻഎസി മാദ്ധ്യമ സ്പൈനി ന്യൂറോണുകളുടെ ഡൻഡറിക് സ്പൈൻ സാന്ദ്രത കുറയ്ക്കുകയും ചെയ്യുന്നു.81 ഈ അനുകൂലത്തിന്റെ സ്വഭാവം, ആന്തരികമായ മോളികുലാർ സംവിധാനം എന്നിവയെക്കുറിച്ച് വളരെക്കുറച്ചു അറിവ് മാത്രമേയുള്ളൂ. ഈ പ്രതിഭാസമാണ്, എന്നിരുന്നാലും, ക്രെബിറ്റും ΔFosB ഉം ഉത്തേജകവും ഉത്തേജകവും സൃഷ്ടിക്കുന്നതിൽ അത്ഭുതവും, NAAC ഡെൻഡറിക് സ്പിൻ ഡെൻസിറ്റിയുടെ ഉത്തേജക-മധ്യേയുള്ള ഉദ്ദീപനത്തിലാണ്. ഈ ഘടകങ്ങളെ അവഗണിച്ച്, നാക് സങ്കേതത്തിന്റെ സാന്ദ്രത അടിച്ചമർത്തുന്നതിനെക്കുറിച്ചാണ് ഇത് ഉയർത്തിക്കാട്ടുന്നത്.
മയക്കുമരുന്ന് ദുരുപയോഗം ചെയ്യുന്ന മോഡലുകളിൽ കാണപ്പെടുന്ന മോർഫോളജിക്കൽ പ്ലാസ്റ്റിറ്റിയിലെ മറ്റ് പ്രമുഖ രൂപങ്ങൾ സെൽഫ് സൈറ്റുകളുടെ സെൽ വ്യാപ്തിയാണ്. ഇത് വൈറ്റ് ഡോപ്പാമൻ ന്യൂറോണുകളുടെ ദീർഘകാല ഓപിഐറ്റാണ്..77,92,93 കാൻബാബിനോയ്ഡുകൾക്ക് സമാനമായ രീതിയിൽ സമാനമായ ഒരു രൂപാന്തരം സംഭവിക്കുന്നു.94 VTA ഡോപാമൈൻ ന്യൂറോണുകളുടെ സങ്കലനം, അത് സ്വയം നിയന്ത്രിക്കാനുള്ള നിയന്ത്രണം ആണ്93 പോസ്റ്റ്മോർട്ടത്തെ പരിശോധിച്ച മനുഷ്യ ഹെറിൻ ആഡിക്കറ്റിൽ രേഖപ്പെടുത്തപ്പെട്ടിട്ടുണ്ട്,77 പ്രതിഫലം സഹിഷ്ണുത പുലർത്തുന്നതായി തോന്നുന്നു, എൻകിൽ കുറച്ചു ഡോപ്പാമിൻ ഉൽപാദനവുമായി ബന്ധപ്പെട്ടിരിക്കുന്നു. ഈ ന്യൂറോണുകൾക്കുള്ളിൽ ബ്രോഡ്-ഡിറൈവ്ഡ് ന്യൂറോട്രോപിക് ഫാക്റ്റർ (ബിഡിഎൻഎഫ്) എക്സ്പ്രഷനുകളുടെ ഓപിഐറ്റേറ്റ് അടിച്ചമർത്തലിലൂടെ സെൽ സോമയുടെ വലിപ്പത്തിൽ കുറവുണ്ടാകുമെന്നാണ് ഇപ്പോൾ കാര്യമായ തെളിവുകൾ സൂചിപ്പിക്കുന്നത്. VD ഡോപ്പാമൻ ന്യൂറോണുകളിൽ താഴെയുള്ള BDNF സിഗ്നലിംഗ് കാസ്കേഡ്സിന്റെ പ്രവർത്തനം കുറയ്ക്കാൻ BDNF പിന്തുണയും VTA ന്യൂറോൺ ചുരുക്കലും ഈ ഓപിഐറ്റേറ്റ്-ഇൻഡുഡ് പിൻവലിക്കൽ നേരിട്ട് നേരിട്ട് ബന്ധപ്പെടുത്തിയിരിക്കുന്നു, IRS2 ഇൻസുലിൻ റിസപ്റ്റർ ഉപരിതല -83 ന്റെ പ്രവർത്തനം, AKT (ഒരു സെറിൽ- threonine kinase), കൂടാതെ TORC2 (റാറ്റാമിൻ- 2- യുടെ ലക്ഷ്യം, ഇത് റാമമിക്സിനിലേക്ക് ബോധരഹിതമാണ്).77,93 മുമ്പ് സൂചിപ്പിച്ചതുപോലെ മോർഫിൻ ഉൾക്കൊള്ളുന്ന വർദ്ധിത ഉത്പാദനക്ഷമതയെ നേരിട്ട് ബി.ഡി.എൻ.എഫ്.77,78 തീർച്ചയായും, കുറച്ചുകൂടി വലിപ്പമുള്ള സെൽ വലിപ്പം, വർദ്ധിച്ചുവരുന്ന ആവേശം എന്നിവ ദൃഡമായി കൂടിച്ചേർന്നു. സെൽ എക്സിറ്റബിളിറ്റിയെ കുറിച്ചുള്ള ഈ നിയന്ത്രണം K ന്റെ അടിച്ചമർത്തലാണ്+ ചാനലുകൾ, GABA എന്നിവA ഈ ന്യൂറോണുകൾ ഇപ്പോൾ നിലവിലുണ്ട്.
മോർഫിൻ പ്രതികരണങ്ങൾ നിയന്ത്രിക്കുന്നതിൽ ഡി.ടി.ടിയുടെ തലത്തിൽ ബിഡിഎൻഎഫ് പങ്കുവെക്കുന്നതിൽ ഈ പങ്ക് കോകൈൻ, മറ്റ് ഉത്തേജക നടപടികളുടെ കാര്യത്തിൽ വ്യത്യസ്തമായ പങ്കു വഹിക്കുന്നു. ബി.ടി.എൻ.എഫ്.യുടെ പ്രാദേശിക സിൻസിസിനും അതുപോലെതന്നെ വിവിധ മേഖലകളിൽ നിന്നും കൂടുതൽ പ്രകാശനം ചെയ്യപ്പെട്ടതിനാലുമാണ് ഉത്തേജക പരിപാടികൾ എൻ ഡി യ്ക്ക് ബി.ഡി.എൻ.എഫ്. സിഗ്നൽ നൽകുന്നത്.95 കൂടാതെ, എൻഎസിയിൽ ബി.ഡി.എൻ.എഫ്. സിഗ്നലിങ് ഉയർത്തുകയും ചെയ്തു. എന്നാൽ വി.ടി.എ.യിൽ ഈ സ്വയം മരുന്നുകൾ ഉൾപ്പെടെയുള്ള മരുന്നുകളുടെ പെരുമാറ്റത്തിലെ പ്രഭാവം പ്രോത്സാഹിപ്പിക്കുന്നതിന് കഴിഞ്ഞിട്ടുണ്ട്.95,96 വിടിഎ-എൻഎസി പാതയിലെ ബിഡിഎൻഎഫ് സിഗ്നലിംഗിന്റെ വിപരീത നിയന്ത്രണം ഓപിയേറ്റുകളും വേഴ്സസ് ഉത്തേജകങ്ങളും അത്തരം വ്യത്യാസങ്ങൾ മരുന്നുകളുടെ എൻഎസി ഡെൻഡ്രിറ്റിക് മുള്ളുകളുടെ വിപരീത നിയന്ത്രണത്തിന് മധ്യസ്ഥത വഹിക്കാനുള്ള സാധ്യത ഉയർത്തുന്നു, ഇത് ഇപ്പോൾ അന്വേഷണത്തിലാണ്.
ഭാവിയിലേക്കുള്ള വഴികൾ
മയക്കുമരുന്ന് പ്രതിഫലം ദുരുപയോഗം ചെയ്യപ്പെട്ട പ്രദേശങ്ങളിൽ സംഭവിക്കുന്ന തന്മാത്രകളുടെയും സെല്ലുലാർ അഡാപ്റ്ററുകളെയും മനസ്സിലാക്കുന്നതിൽ വിപുലമായ പുരോഗതികൾ മുകളിൽ വിവരിക്കുന്നുണ്ട്. മയക്കുമരുന്നിന്റെ അപര്യാപ്തതയുടെ ആവർത്തിച്ചുള്ള പ്രതിപ്രവർത്തനം, മൃഗചികിത്സയിലെ ആഡംബര സിൻഡ്രോമിന്റെ ചില സ്വഭാവ സവിശേഷതകളിൽ . ഈ പുരോഗതി ഉണ്ടായിട്ടും പ്രധാന ചോദ്യങ്ങൾ ഇപ്പോഴും നിലനിൽക്കുന്നു. നമ്മുടെ നിലവിലുള്ള അറിവുകൾക്ക് VTA, NAC എന്നിവയിൽ ശ്രദ്ധ കേന്ദ്രീകരിക്കുന്നു. മയക്കുമരുന്ന് അടിമകളിലെ മറ്റ് പ്രധാന ലെബിക് തലച്ചോർ പ്രദേശങ്ങളെ കുറിച്ചുള്ള വിവരങ്ങൾ വളരെ കുറവാണ്. കൂടാതെ, ഒരു മയക്കുമരുന്നു സംബന്ധമായ പെരുമാറ്റത്തിലെ തന്മാത്ര-സെല്ലുലാർ അഡാപ്റ്റേഷന്റെ കാരണാവയവത്തിന്റെ എല്ലാ പരീക്ഷണാത്മക പ്രകടനങ്ങളും ഒറ്റയടിക്ക് ഒരു വ്യക്തിഗത അഡാപ്റ്റേഷനെ ഒന്നാക്കി മാറ്റിയിട്ടുണ്ട്. ഒരേ സമയം അനേകം അനുകരണങ്ങളെ കൈകാര്യം ചെയ്യുന്നത് വളരെ ബുദ്ധിമുട്ടുള്ള കാര്യമാണ്, പക്ഷേ അത് അത്യന്താപേക്ഷിതമാണ്. വ്യക്തിഗത ന്യൂറോണുകളിൽപ്പോലും മരുന്നുകൾ വളരെ വ്യത്യസ്തമായ വ്യത്യാസങ്ങൾ സൃഷ്ടിക്കുന്നുണ്ടെന്ന് നമുക്കറിയാം, കാരണം പെരുമാറ്റത്തെ സ്വാധീനിക്കാൻ സങ്കീർണ്ണമായ രീതിയിൽ ഇത് സമാഹരിക്കാറുണ്ട്. അത്തരം ഒരു വ്യവസ്ഥ ജീവശാസ്ത്ര സമീപനം ആത്യന്തികമായി ആസക്തിയുടെ ജീവശാസ്ത്രപരമായ അടിത്തറ തകർക്കുക വളരെ നിർണായകമാകും. ഒടുവിൽ, ആസക്തിയുമായി ബന്ധപ്പെട്ട ഓർമ്മകളിലെ തന്മാത്രകളുടെ സെല്ലുലാർ മെക്കാനിസം മനസ്സിലാക്കാൻ പരിശ്രമിക്കുക, പെരുമാറ്റ സ്മരണത്തിന്റെ ജൈവപരമായ അടിത്തറയെക്കുറിച്ച് മനസിലാക്കാനുള്ള മറ്റെല്ലാ ശ്രമങ്ങളും ഈ ഘട്ടത്തിലുണ്ട്: ജീവശാസ്ത്രപരമായ പ്രതിഭാസങ്ങളെ സങ്കീർണമായ സ്വഭാവമയമായ ഓർമ്മപ്പെടുത്തലുമായി ബന്ധപ്പെടുത്താനുള്ള ഞങ്ങളുടെ കഴിവ് വളരെ പ്രയാസകരമാണ്. ഈ വിഭജനം മറികടക്കുക എന്നത് ഒരുപക്ഷെ ന്യൂറോസയൻസസിലെ ഏറ്റവും വലിയ വെല്ലുവിളി ആയിട്ടാണ്.
അക്നോളജ്മെന്റ്
നാഷണൽ ഇൻസ്റ്റിറ്റ്യൂട്ട് ഓൺ ഡ്രഗ്സ് അബ്യൂസ് നൽകിയ ഗ്രാന്റുകൾ ഈ ജോലിക്ക് സഹായകമായി.
തെരഞ്ഞെടുത്ത ചുരുക്കങ്ങളും ചുരുക്കെഴുത്തുകളും
- നാക്
- ന്യൂക്ലിയസ് accumbens
- ക്രെബി
- cAMP പ്രതികരണ ഘടകമായ ബൈൻഡിംഗ് പ്രോട്ടീൻ
- ΔFosB
- ഒരു Fos കുടുംബ ട്രാൻസ്ക്രിപ്ഷൻ ഘടകം
- VTA
- വാൽട്രൽ ടെഗ്ഗിക്കൽ ഏരിയ
- Ampa
- α-amino-xNUMx-hydroxy-xNUMx-methyl-3-isoxazolepropionic ആസിഡ്
- ലിമിറ്റഡ്
- ദീർഘകാല വിഷാദരോഗം
- LTP
- ദീർഘകാല ആധിപത്യം
- ബി.ഡി.എൻ.എഫ്
- ബ്രെയിൻ ഡിറൈവ്ഡ് ന്യൂറോട്രോപിക് ഘടകം
- NKkB
- ആണവ ഘടകം kB
അവലംബം