КОММЕНТАРИЙ: Абстинент приматтардагы допаминдик рецепторлордун деңгээлине кандай таасир этээрин камтыган бир нече изилдөөлөрдүн бири.
- D2 кабылдагычтары бир айга жетпеген убакытта кайра калыбына келет
- D1 рецепторлору бир айда өтө жогору, бирок 90 күндүн ичинде кайра калыбына келет.
- Жогорку же төмөн D1 кабылдагычтар курч алып салуу жана кумар ачкычы болушу мүмкүн
Thomas JR Beverridge1, Хилари Р Смит1, Майкл А Надер1 жана Линда Дж Поррино1
1Физиология жана фармакология бөлүмү, Баңгизатты кыянаттык менен пайдаланууну нейробиологиялык иликтөө борбору, Уэйк Форест университетинин Медицина мектеби, Уинстон-Сейлем, NC, АКШ
Корреспонденция: Dr LJ Porrino, Физиология жана фармакология бөлүмү, Баңгизатты кыянаттык менен пайдаланууну нейробиологиялык иликтөө борбору, Уэйк Форест университетинин Медицина мектеби, Медициналык борбор бульвары, Уинстон-Салем, NC 27157-1083, АКШ. Тел: +1 336 716 8575; Факс: +1 336 716 8501; E-mail: [электрондук почта корголгон]
29-жылдын 2008-апрелинде алынган; 25-жылдын 2008-июлунда кайра каралган; 30-жылдын 2008-июлунда кабыл алынган; 3-жылдын 2008-сентябрында онлайн жарыяланды.
жалпылаган
Допамин (DA) системасынын ичиндеги дисрегуляция кокаиндин өнөкөт таасиринин өзгөчөлүгү болсо да, бул өзгөртүүлөр абстинентке чейин сакталып калабы деген суроо жоопсуз бойдон калууда. Адамдык эмес приматтар өнөкөт кокаиндин таасиринен кийинки DA системасына абстиненттин таасирин баалоо үчүн идеалдуу моделди билдирет. Бул изилдөөдө эркек резус маймылдары кокаинди (0.3 mg/бир инъекцияга кг, ар бир сессияга 30 күчөткүч) белгиленген интервалда 3 мүнөт с.100 күн, андан кийин 30 же 90 күн бою баш тартуу. Кокаинди өзүн-өзү башкаруунун бул узактыгы мурда DA D2 сыяктуу рецепторлордун тыгыздыгын азайтып, D1 сыяктуу рецепторлордун жана DA транспортёрлорунун деңгээлин жогорулатат (DAT). Контролдук маймылдардын жооп берүүсү бирдей протокол жана ошол эле кармануу мөөнөттөрү боюнча тамак-аш көрсөтүү менен сакталган. [3H]SCH 23390 үчүн милдеттүү DA D1 рецепторлору 30 күн бою абстиненттен кийин стриатумдун бардык бөлүктөрүндө контролдук жаныбарларга салыштырмалуу кыйла жогору болгон, ал эми [3H]раклоприддин DA D2 рецепторлору менен байланышы топтордун ортосунда айырмаланган эмес. [3H]WIN 35 428дин DAT менен байланышы 30 күндүк абстиненттен кийин дорсалдык жана вентралдык стриатумдун дээрлик бардык бөлүктөрү боюнча кыйла жогору болгон. 90 күн бою абстиненттен кийин, DA D1 рецепторлорунун жана DAT деңгээли контролдук маанилерден айырмаланган эмес. Бул жыйынтыктар DA тутумунун айрым элементтеринин акыры калыбына келтирилгендигин көрсөтүп турат, бирок алар ошондой эле өнөкөт кокаинди өзүн-өзү башкаруудан баш тартуунун алгачкы фазаларында бул компоненттердин динамикалык мүнөзүн баса белгилешет.
Keywords:
кокаин, дофамин, авторрадиография, абстиненция, стриатум
КИРИШҮҮ
Адамдардын көз карандыларынын өнөкөт кокаинди колдонуу допамин (DA) системасындагы нейроадаптациялар менен байланыштырылды.Malison жана башкалар, 1998; Volkow жана башкалар, 1993, 1997). Аларга DA транспортёрлорунун тыгыздыгынын жогорулашы (DAT) жана DA D2 сыяктуу рецепторлордун концентрациясынын төмөндөшү кирет (кичине жана башкалар, 1999; Mash жана башкалар, 2002; Volkow жана башкалар, 1993). Мындан тышкары, DA чыгарууда өзгөрүүлөр да байкалган. Мисалы, позитрондук эмиссиялык томографияны (ПЭТ) колдонуп изилдөөчүлөр [11C]раклоприд жана метилфенидат кокаинди өнөкөт колдонуучулардын стриатумунда DA чыгаруунун төмөндөшүн көрсөттү (Volkow жана башкалар, 1997; Wong жана башкалар, 2006). Бирок бир көйгөй - башка мыйзамсыз жана мыйзамдуу баңгизаттарды колдонуу, баңги заттарды колдонуунун жана колдонуунун мурунку үлгүлөрүндөгү айырмачылыктар жана жашоо образындагы айырмачылыктар сыяктуу башка факторлордун таасирин жокко чыгаруу кыйын. Бул айырмачылыктар, ошондой эле баңгизаттарды колдонуудан мурун болгон шарттардын болушу, бейтаптардагы изилдөөлөрдүн чечмелөөнү чектеши мүмкүн.
Өзгөрмөлөрдү системалуу түрдө башкарууга мүмкүн болгон адамдык эмес примат моделдери кокаинди өнөкөт өзүн-өзү башкаруунун жана андан кийинки абстиненттин кесепеттерин изилдөөгө альтернативалуу мамилени билдирет. Мурунку изилдөөлөр көрсөткөндөй, кокаиндин өнөкөт таасири DA D2 рецепторлорунун концентрациясынын олуттуу төмөндөшү, ошондой эле D1 рецепторлорунун деңгээлинин жана DAT тыгыздыгынын жогорулашы менен коштолот.Letchworth жана башкалар, 2001; Мур жана башкалар, 1998a, 1998b; Надер жана башкалар, 2002, 2006). Бул эффекттер адамдарда байкалгандарды чагылдырат, ошентип, дары-дармектердин таасиринин бул моделдеринин пайдалуулугун далилдейт.
DA системаларынын дисрегуляциясы үчүн олуттуу далилдер бар болсо да, баңгизатты колдонууну токтоткондон кийин калыбына келтирүүгө кандайдыр бир далилдер бар же жок экендигин баалоо кыйыныраак болуп чыкты (Malison жана башкалар, 1998; Jacobsen жана башкалар, 2000; Volkow жана башкалар, 1993) же бул өзгөрүүлөр кокаинди колдонуунун убакыт алкагынан тышкары сакталып калабы. Дагы, адам эмес примат моделдери көз карандылыктын бул фазасы жөнүндө түшүнүк бере алат. Farfel жана башкалар (1992) маймылдардын стриатумунда DAT жана D1 сыяктуу рецепторлордун концентрациялары кокаиндин өнөкөт контингенттик таасиринен баш тарткандан кийин азайгандыгын билдирди. Бирок, абстиненттин өзгөчө ролун аныктоо кыйынга турду, анткени абстиненция мезгили жок топто өлчөөлөрдүн жоктугу. Ушундай эле жол менен, Мелега жана башкалар (2008) метамфетаминдин күчөгөн режиминен 3 жума бою кармангандан кийин вервет маймылдарынын стриатумунда DAT деңгээли бир кыйла төмөндөгөнүн билдирди. Бирок эки изилдөөдө стимуляторлорду башкаруу шарттуу эмес. Башкаруу маршруту (контингент vs контингенттик эмес) мээге DA релизине карата дифференциалдуу таасир этээри көрсөтүлдү (Хемби жана башкалар, 1997) жана глюкоза метаболизми (Грэм жана Поррино, 1995; Porrino жана башкалар, 2002). Ошентип, бул изилдөөдө өзүн-өзү башкарууну колдонуу бул маселени айланып өтөт. Мындан тышкары, узак мөөнөттүү кокаинди өзүн-өзү башкаруунун мээнин DA тутумдарына тийгизген таасири резус маймылдарында өзүн-өзү башкаруунун бул моделин колдонуу менен кеңири изилденген, ошону менен абстиненция учурунда пайда болгон нейроадаптацияларды баалоо үчүн базалык базаны камсыз кылган.
Бул изилдөөлөрдүн максаты, ошондуктан, DAT жана DA D1 жана D2 рецепторлордун концентрациясындагы өзгөрүүлөр кокаинди өз алдынча башкарууга дуушар болгон жаныбарларда мурда далилденгендигин аныктоо болгон.Letchworth жана башкалар, 2001; Мур жана башкалар, 1998a, 1998b; Надер жана башкалар, 2002) узакка созулган абстиненциядан кийин жокко чыгарылат. Баңги затын колдонуучулардын арасында жүргүзүлгөн изилдөөлөрдүн негизинде (кара Volkow жана башкалар, 1993), биз DA тутумундагы бул өзгөрүүлөр 3 айдан баш тартуудан кийин да сакталат деп божомолдодук. Бул максатта маймылдар 100 сеанс бою кокаинди өз алдынча ичишти, жалпы кабыл алуу 900 mg/кг, андан кийин же 30 же 90 күн дарыдан баш тартуу. DA D1 жана D2 рецепторлору, ошондой эле DAT, сандык менен ченелген экстракорпоралдык рецепторлордун авторрадиографиясы.
METHODS
субъекттери
Жалпысынан 17 эксперименталдык жактан таза бойго жеткен эркек резус маймылдары (Macaca mulatta) салмагы 7.7ден 13кө чейин кг (орточо±SD, 10.2±1.32) изилдөөнүн башында субъект катары кызмат кылган. Бардык жол-жоболор беренеде сүрөттөлгөн белгиленген практикага ылайык аткарылды Улуттук Саламаттыкты сактоо Институттары лабораториялык жаныбарларды багуу жана пайдалануу боюнча колдонмо. Мындан тышкары, бардык процедуралар Уэйк Форест университетинин Жаныбарларга кам көрүү жана пайдалануу комитети тарабынан каралып, бекитилген. Маймылдар жекече суу менен дат баспас болоттон жасалган капастарга жайгаштырылды ад либитум; жаныбарлар бири-бири менен физикалык жана визуалдык байланышта болгон. Алардын дене салмагы болжол менен 90-95 болуп сакталган% эксперименталдык сессияларда жана Lab Diet Monkey Chow кошумча азыктандыруу аркылуу табылган банандын жыты бар гранулдар менен эркин тамактандыруу салмактары, 30дан эрте эмес мин. сессиядан кийин. Мындан тышкары, аларга жумасына үч жолудан кем эмес жаңы мөмө же жержаңгак берилди. Ар бир маймылды жумасына бир жолу таразага тартып, керек болсо, алардын диетасы туруктуу салмакты кармап туруу үчүн туураланган.
Жүрүш-туруш аппараты
Эксперименталдык сеанстар желдетилген жана үн начарлатылган операнттык камераларда өткөрүлдү (1.5 × 0.74 × 0.76) м; Med Associates Inc., East Fairfield, VT) примат креслосун жайгаштыруу үчүн иштелип чыккан (Model R001; Primate Products, Redwood City, CA). Палатада чалгындоо панели болгон (48 × 69 см), ал эки тартылуучу рычагдан турган (5 см туурасы) жана үч стимул жарыктары. Рычактар примат креслосунда отурган маймылга оңой жете турган жерге жайгаштырылган. Банан даамы бар тамак-аш гранулдары (1 g; Bio-Serv, Frenchtown, NJ) камеранын үстүндө жайгашкан фидерден жеткирилген. Перистальтикалык инфузиялык насос (7531-10; Cole-Parmer Co., Chicago, IL) болжол менен 1 ылдамдыкта дары саймаларын жеткирүү үчүн колдонулган. 10 мл лар кокаинди өзү башкарган жаныбарларга. Камералардын иштеши жана маалыматтарды алуу интерфейси бар Power Macintosh компьютер системасы менен ишке ашырылган (Med Associates Inc.).
Хирургиялык жол-жоболору
Бардык маймылдар, анын ичинде башкаруу органдары, хирургиялык жол менен, стерилдүү шарттарда, тамырга катетер жана тамырга кирүү порттору (GPV модели; Access Technologies, Skokie, IL) менен даярдалган. Маймылдар кетаминдин кошулмасы менен анестезияланган (15 mg/кг, им) жана буторфанол (0.03 mg/кг, им) жана сан венасынын жанынан кесилген. Венаны туталак кесүүдөн жана изоляциялоодон кийин катетердин проксималдык учу венанын ичине төмөнкү кавада бүтүшү үчүн эсептелген аралыкка киргизилди. Катетердин алыскы учу тери астына арканын орто сызыгынан бир аз четте турган кесикке сайылган. Кан тамырга кирүү порту бул кесилген жердин жанындагы туюк диссекциядан пайда болгон чөнтөктүн ичинде жайгаштырылган. Маймылдарга 24–48 берилди h тамак-аш менен бекемделген жооп кайтаруу алдында калыбына келтирүү убактысы. Терминалдык процедурадан болжол менен 5 күн мурун ар бир маймылга өнөкөт катетер жанаша сан артериясына имплантацияланган, артериялык кандын үлгүлөрүн алуу үчүн. Хирургиялык процедуралар веналык катетерлер үчүн сүрөттөлгөндөй эле. Жыйынтыктоочу сессия күнү мээдеги глюкозанын метаболизминин терминалдык изилдөөсү жүргүзүлүп, анда маймылдарга 2-[14C]дезоксиглюкоза (2-DG) болжол менен 2 сессия аяктагандан кийин мин жана кан үлгүлөрү 45 ашуун артериялык катетер аркылуу алынган минимум мезгил (кара Beveridge жана башкалар, 2006 чоо-жайы үчүн). Бул изилдөөлөрдөн алынган метаболизм маалыматтары бул жерде берилген эмес.
Өзүн-өзү башкаруу процедуралары
Маймылдар алгач эки рычагдын бирине жооп берүүгө үйрөтүлгөн. Болжол менен 3 жумалык мезгилдин ичинде тамак-аш гранулдарынын жеткиликтүүлүгүнүн ортосундагы аралык 3 мүнөттүк интервалга жеткенге чейин акырындык менен көбөйтүлдү (б.а. бекитилген интервал 3 мүнөттүк күчөтүү графиги; FI 3-мин). Акыркы графиктин шарттарына ылайык, рычагдагы биринчи жооп 3төн кийин мин азык-тулук гранулунун жеткирилишине алып келди; сеанстар 30 тамак-аш көрсөтүүдөн кийин аяктады. Ар бир сессиянын аягында жооп рычагдары тартылып, үй жарыгы жана стимулдоочу жарыктар өчүп, жаныбарлар болжол менен 30 караңгы камерада калышты. мин алар өз үйлөрүнө кайтарылганга чейин. Бардык маймылдар FI 3 мүнөттүк тамак-аш көрсөтүү графигине ылайык, жок дегенде 20 сессияга жана туруктуу көрсөткүчтөргө жеткенге чейин жооп беришти (±20% үч сессиянын орточо көрсөткүчү, жооп ылдамдыгынын тенденциясы жок). Азык-түлүк менен камсыздалган жооп туруктуу болгондо, азыктандыруучу электрден ажыратылып, жок болуунун жооп кайтарууга тийгизген таасири беш ирет катары менен каралып, андан кийин жооп кайтаруу калыбына келтирилип, тамак-аш презентациясы менен колдоого алынган.
Негизги көрсөткүчтөр белгиленгенден кийин, бардык маймылдар хирургиялык жол менен жогоруда айтылгандай веноздук катетерлер менен даярдалган жана туш келди үч топтун бирине дайындалган. Маймылдардын бир тобу башкаруучу катары кызмат кылып, FI 3 мүнөттүк тамак-аш көрсөтүү графиги боюнча жалпысынан 100 сессияга жооп берүүнү улантышты (N=6). Калган 11 маймыл кокаинди өз алдынча башкаруу топторуна дайындалган (0.3 mg/кг сайынуу). Анткени 0.3 mg/0.1 кг кокаин бир сайынуу үчүн мурда кокаинди колдонбогон маймылдар үчүн жогорку доза деп эсептелген, көпчүлүк жаныбарлар үчүн бул дозага маймылга XNUMX өзүн өзү киргизүүгө уруксат берүү аркылуу эки сеанстын ичинде жетишилген. mg/кг кокаин. Кокаинди өзүн-өзү башкаруу сессиялары башталганга чейин, операциядан кийин (болжол менен 4-6 күн) тамак-аш менен камсыздалган көрсөткүчтөрдү турукташтырууга уруксат берилген. Ар бир эксперименттик сессиянын алдында жаныбардын арткы бөлүгү 95 менен тазаланган% этанол жана бетадин скрабы жана 22 калибрлүү Huber Point Needle (Модель PG20-125) веноздук катетерге алып баруучу портко киргизилип, кокаин эритмеси бар инфузиялык насосту катетерге туташтырды. Сеанс башталганга чейин насос болжол менен 3 саат иштетилди с, портту эксперименталдык сессия учурунда болгон кокаиндин дозасы менен толтуруу. Сеанстар 30 инъекциядан кийин аяктады; башкаруу шарттарында маймылдар болжол менен 30 караңгы бөлмөдө калышты мин. Ар бир сессиянын аягында порт гепаринизацияланган туз менен толтурулган (100 U/мл) кандын уюшун алдын алууга жардам берет.
Эксперименттик сессиялар күн сайын болжол менен бир убакта жүргүзүлүп, жалпысынан 100 сессияга чейин уланды. 100 сессия аяктагандан кийин, 30 же 90 күндүк абстиненция мезгили киргизилди, бул убакыттын ичинде катетерлер күн сайын гепаринизацияланган туз менен жуулчу, бирок кокаин же тамак-ашты өз алдынча башкаруу сессиялары өткөрүлгөн эмес. Контролдук топ үчүн төрт жаныбарга 30 күндүк жана калган экөөнө 90 күндүк абстиненттик мөөнөт киргизилген. Кокаин тобу үчүн сегиз жаныбарга 30 күндүк жана үч жаныбарга 90 күндүк абстиненция мөөнөтү белгиленген. Абстиненция мезгилинин аягында бир жыйынтыктоочу өзүн-өзү башкаруу сессиясы (тамак-ашты көзөмөлдөө же кокаин) өткөрүлдү жана сессиядан кийин дароо эле 2-DG процедурасы башталды. 30 күндүк абстиненттик топтун эки көзөмөлүндө жана төрт кокаинди өз алдынча башкаруучу жаныбарларда акыркы сессияда кокаин алынган эмес. Жаныбарлар пентобарбиталдын ашыкча дозасы менен өлтүрүлгөн (100 mg/кг, iv) 45-жылдын аягында мин тракерди кабыл алуу мезгили.
Tissue Processing
Өлтүргөндөн кийин, мээлер дароо алынып, бөгөттөлгөн жана изопентанда -35тен -55°Cге чейин тоңдурулган жана андан кийин -80°Cде сакталган. Андан кийин стриатумду камтыган кыртыш блоктору криостатта -20°Cде короналдык тегиздикте 20га кесилген. μm секциялар электростатикалык заряддалган слайддарга чогултулуп, түнү бою вакуумда 4°C температурада кургатылган, андан кийин авторадиография үчүн иштетилгенге чейин -80°Cде сакталган. Мээ бөлүктөрү каудат өзөкчөлөрүнүн, путамендердин жана алдыңкы комиссиянын rostral жагында жаткан өзөкчөлөрүнүн бөлүктөрүнөн чогултулган. Бул аймак precommissural striatum деп аталат. Андан ары, precommissural стриатумдун ростралдык жана каудалдык деңгээлдери аккумбенс ядросуна шилтеме менен белгиленген. rostral precommissural striatum ядросу айкын кабыкча жана өзөк суббөлүмдөрүнө айырмаланбаган аймак болуп саналат. Каудалдык precommissural striatum жыт сезүүчү туберкулездун чыгышынан артта жайгашкан аккумбенс ядросунун кабыгынын жана өзөгүнүн көрүнүшүнө дал келген аймак. Байланыштуу изилдөөлөрдүн ар бири үчүн ар бир жаныбарга жалпысынан 10 бөлүктөн турган прекоммиссуралдык стриатум аркылуу беш деңгээлден (эки ростралдык жана үч каудалдык) эки чектеш бөлүм алынды.
D1 рецептор менен байланышуу
DA D1 кабылдагычты байланыштыруучу сайттын тыгыздыгы менен аныкталган [3H]SCH 23390 (өзгөчө иш-85 Ci/ммоль; PerkinElmer, Бостон, MA) сандык боюнча экстракорпоралдык ылайыкталган жол-жоболор боюнча кабылдагыч autoradiography Lidow жана башкалар (1991) жана Надер жана башкалар (2002). Бөлүмдөр 20га чейин алдын ала инкубацияланган буферде мин (50 mM Tris, 120 mM NaCl, 5 mM KCl, 2 mM CaCl2, 1 mM MgCl2, рН 7.4, 25°C) эндогендик DA, кокаинди жана [14C] 2-DG процедурасынан. Андан кийин бөлүмдөр 30 инкубацияланган мин ошол эле буферде, рН 7.4, 25°C, 1 камтыган мм аскорбин кислотасы, 40 nM кетансерин жана 1 nM [3H]SCH 23390. Инкубациялоодон кийин бөлүмдөр 20 жолу чайкалат s камтыган буферде 1 mM аскорбин кислотасы рН 7.4, 4°С, андан кийин 4°С дистилденген сууга малып, муздак аба агымынын астында кургатылган. Спецификалык эмес байланыш инкубация эритмесинде чектеш бөлүмдөрдүн 5 катышуусунда инкубациялоо менен аныкталган. μM (+)-бутакламол. Бөлүмдөр, калибрленген менен бирге [3H] авторадиографиялык стандарттар (Amersham, Piscataway, NJ) Kodak Biomax MR тасмасына (Fisher Scientific, Pittsburgh, PA) 6 жума бою бекитилген.
D2 рецептор менен байланышуу
DA D2 кабылдагычтарды байланыштыруучу жерлердин тыгыздыгы жана бөлүштүрүлүшү менен аныкталган [3H]раклоприд (өзгөчө активдүүлүгү, 87 Ci/ммоль; PerkinElmer) ылайыкташтырылган жол-жоболор боюнча Lidow жана башкалар (1991) жана Надер жана башкалар (2002). Бөлүмдөр 20га чейин алдын ала инкубацияланган буферде мин (50 mM Tris, 120 mM NaCl, 5 mM KCl, pH 7.4, 25°C) эндогендик DA, кокаин жана [14C] 2-DG процедурасынан. Андан кийин слайддар 30 инкубацияланган мин бир эле буферде, камтыган 5 мм аскорбин кислотасы жана 2 nM [3H]раклоприд. Бөлүктөр 3 × 2 чайкалат мин буферде рН 7.4, 4°С, андан кийин 4°C дистилденген сууга малып, муздак аба агымы астында кургатылган. Спецификалык эмес байланыш инкубация эритмесинде чектеш бөлүмдөрдүн инкубациялоо жолу менен аныкталды. μM (+)–бутакламол. Бөлүмдөр, калибрленген менен бирге [3H] авторадиографиялык стандарттар, Kodak Biomax MR тасмасына 8 жумага бекитилген.
Допамин ташуучу байланыш
DAT байланыш сайттарынын тыгыздыгы колдонуу менен аныкталган [3H]WIN 35,428 (конкреттүү иш, 87 Ci/ммоль; PerkinElmer) ылайыкташтырылган жол-жоболор боюнча авторадиография Canfield жана башкалар (1990) жана Letchworth жана башкалар (2001). Ткань бөлүмдөрү буферде алдын ала инкубацияланган (50 mM Tris, 100 mM NaCl, pH 7.4, 4°C) 20 үчүн мин ар кандай калдык DA алып салуу, кокаин, жана [14C] 2-DG процедурасынан. Андан кийин бөлүмдөр 2 инкубацияланган h 4°C 5 камтыган ошол эле буферде nM [3H]WIN 35 428. Бөлүктөр жалпысынан 2 жолу чайкалды мин буферде 4°С, андан кийин 4°C дистилденген сууга малып, муздак аба агымы астында кургатылган. Спецификалык эмес байланыш инкубация эритмесинде чектеш бөлүмдөрдү 30 μМ кокаин. Бөлүмдөр, калибрленген менен бирге [3H] авторадиографиялык стандарттар, Kodak Biomax MR тасмасына 6 жумага бекитилген.
Денситометрия жана маалыматтарды талдоо
Тасмалар Kodak GBX иштеп чыгуучусу, токтоочу ванна жана Rapid Fixer (VWR, West Chester, PA) менен иштелип чыккан, андан кийин чайкалып алынган. Авторадиограммалардын анализи сүрөттү компьютерлештирилген иштетүү системасы менен сандык денситометрия менен жүргүзүлгөн (MCID, Imaging Research; InterFocus Imaging Ltd, Кембридж, Улуу Британия). Оптикалык тыгыздыктын маанилери fmolга айландырылды/мг (нымдуу салмактагы кыртыштын) калибрленген шилтеме менен [3H] стандарттар. Спецификалык байланыш жалпы байланыштын үстү-үстүнө коюлган чектеш сүрөттөрдөн спецификалык эмес байланыштын сүрөттөрүн санарип алуу менен аныкталган. Структуралар рецептордук байланыш үчүн талдангандарга чектеш бөлүмдөрдүн Nissl боёосу менен аныкталган. Ар бир анализден алынган маалыматтар дисперсияны бир тараптуу талдоо, андан кийин эң аз квадраттык айырмачылыктар аркылуу өз алдынча талданган Постту туз бир нече салыштыруу үчүн тесттер. Ар бир аймак өзүнчө анализден турат. Анткени 30 жана 90 күн бою абстинент болгон контролдук жаныбарлардан алынган милдеттүү маалыматтар мурунку изилдөөлөргө окшош, бири-биринен олуттуу айырмаланган эмес (Надер жана башкалар, 2002), контролдук топтордун маалыматтары бириктирилди. Мындан тышкары, кокаин алган жаныбарлардын маалыматтары менен акыркы сессиясында албагандардын ортосунда олуттуу айырмачылыктар болгон эмес, ошондуктан бул топтордун маалыматтары да бириктирилген.
ЖЫЙЫНТЫКТАРЫ
30 күн бою өнөкөт кокаинди өзүн-өзү башкаруудан баш тартуунун кесепеттери
концентрациялары [3H]SCH 23390 прекоммиссуралдык стриатумдагы DA D1 рецепторлору менен байланышта көрсөтүлгөн. стол 1. Өзгөчө байланыш [3H]SCH 23390 90дон ашыкты түзгөн% жалпы байланыш. Мурунку отчетторго ылайык (Мур жана башкалар, 1998a; Надер жана башкалар, 2002), байлоо [3H]SCH 23390 D1 кабылдагычтарга дары-дармексиз таасир этпеген контролдук жаныбарларда стриатумдун субрегиондорунун ортосундагы байланыш даражасындагы олуттуу айырмачылыктар менен гетерогендүү болгон. Маркировка striatum боюнча көбүрөөк ростралдык жана ортоңку бөлүктөрүндө тыгызыраак болгон.
![1-таблица - Өнөкөт кокаинди өзүн-өзү башкаруудан кийинки 30 же 90 күндүк абстиненттин [lsqb]3H[rsqb]SCH 23390 тыгыздыгына таасири. бул. Бул сүрөткө кирүү үчүн жардам керек болсо, help@nature.com же автор менен байланышыңыз](https://yourbrainonporn.com/wp-content/uploads/2020/01/table_thumb.gif)
Кокаиндин таасиринен 30 күн бою сактангандан кийин, D1 рецепторлору менен байланышы прекоммиссуралдык стриатумдун бүт ростралдык-каудалдык кеңдиги боюнча кеңири таралган бийиктиктер менен мүнөздөлгөн, бул дары-дармек менен байланышпаган контролдук жаныбарлардагы байланыштын тыгыздыгы менен салыштырганда (стол 1; Figure 1). Көбүрөөк ростралдык стриатумда, концентраттар каудаттык ядродо, анын ичинде дорсолатералдууда (+27%), Борбордук (+27%), dorsomedial (+27%) жана вентромедиалдык (+23%) бөлүктөрү, ошондой эле дорсалдык (+17%), Борбордук (+22%) жана вентралдык (+23%) путамендердин бөлүктөрү, дары-дармексиз контролдоочу жаныбарлардын тыгыздыгы менен салыштырганда. Маанилүү бийиктиктер ядронун аккументтеринде да байкалды (+23%) ушул деңгээлде. Ядронун өзөгү жана кабыгы эң көп дифференцияланган стриатумдун деңгээлинде, D1 сыяктуу рецепторлордун тыгыздыгы да арткы каптал боюнча бир кыйла жогору болгон (+31%), Борбордук (+29%), dorsomedial (+30%) жана вентромедиалдык (+18%) куйруктуу ядро, ошондой эле арка (+23%), Борбордук (+29%) жана вентралдык (+28%) путамендер, дары-дармексиз контролдоолордун тыгыздыгы менен салыштырганда. Бул деңгээлдеги вентралдык стриатумдун ичинде Д1 кабылдагычтарды байланыштырган жерлер ядронун өзөгүндө жогору болгон (+45%) Жана кабыгы (+20%), ошондой эле жыт сезүүчү туберкулезде (+26%), башкаруу элементтериндеги тыгыздыктарга салыштырмалуу.
Figure 1.
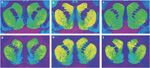 Өкүл авторадиограммалар [3H] D23390 кабылдагычтарга SCH 1 байланышы (жогорку панели) жана [3H]WIN 35428 резус маймылынын стриатумунун короналдык бөлүмдөрүндөгү дофамин ташуучуларга (төмөнкү панель) байланыштырат. (а, г) Жаныбарлардын тамак-ашты күчөтүү үчүн жооп кайтарышын көзөмөлдөө. (б, д) Кокаинди өз алдынча башкаруу жаныбары, 30 күн бою карманбоо. (c, f) Кокаинди өз алдынча башкаруу жаныбары, 90 күн бою карманбоо.
Өкүл авторадиограммалар [3H] D23390 кабылдагычтарга SCH 1 байланышы (жогорку панели) жана [3H]WIN 35428 резус маймылынын стриатумунун короналдык бөлүмдөрүндөгү дофамин ташуучуларга (төмөнкү панель) байланыштырат. (а, г) Жаныбарлардын тамак-ашты күчөтүү үчүн жооп кайтарышын көзөмөлдөө. (б, д) Кокаинди өз алдынча башкаруу жаныбары, 30 күн бою карманбоо. (c, f) Кокаинди өз алдынча башкаруу жаныбары, 90 күн бою карманбоо.
концентрациялары [3H]раклоприддин прекоммиссуралдык стриатумдагы DA D2 рецепторлору менен байланышы көрсөтүлгөн. стол 2. менен конкреттүү байланыштыруу [3H]раклоприд 90дон ашыкты түзгөн% жалпы байланыш. бөлүштүрүү [3H]раклоприддин D2 рецепторлору менен байланышы да мурунку отчеттордогудай арка жана вентралдык стриатумдун субрегиондору боюнча гетерогендүү болгон.Мур жана башкалар, 1998b; Надер жана башкалар, 2002). Дары-дармектин таасири тийбеген контролдордо вентралдык стриатумга салыштырмалуу дорсалда D2 байланышуучу жерлердин жогорку концентрациясы болгон. Мындан тышкары, striatum дагы каптал бөлүктөрүндө бар милдеттүү сайттардын жогорку концентрациясы менен каптал градиенттин ортодон далилдер бар.
30 күндүк абстиненттен кийин, кокаинге дуушар болгон жана тамак-аш менен бекемделген жаныбарларда D2 рецепторунун байланышынын деңгээли стриатумдун көпчүлүк аймактарында бири-биринен олуттуу айырмаланган эмес. Байланыштуу жерлердин жогорку концентрациясы вентралдык путамендерде байкалган (+10%) жана алдыңкы өзөкчөлөр (+12%) контроль менен салыштырганда кокаинге дуушар болгон маймылдардын кыртышында. Башка олуттуу айырмачылыктар байкалган жок.
концентрациялары [3H]WIN 35428 прекоммиссуралдык стриатумда DAT менен байланышта көрсөтүлгөн стол 3. Мурунку отчетторго ылайык (Letchworth жана башкалар, 2001), дары-дармексиз жаныбарлардын DAT сайттары менен байланышы ventral striatum менен салыштырганда дорсалда жогору болгон. Ядронун аккумбенстеринин ичинде кабык бөлүктөрүнө салыштырмалуу өзөктө жогорку тыгыздык байкалган. Акыр-аягы, спецификалык эмес байланыш 10дон азды түзгөн% жалпысынан.
Кокаиндин таасиринен 30 күн бою сактангандан кийин, DAT менен байланыш стриатум ростралынын көпчүлүк аймактарында алдыңкы комиссияга чейин дары-дармек менен байланышпаган контролдук жаныбарларга салыштырмалуу бир кыйла жогору болгон (Figure 1). Тактап айтканда, ростралдык деңгээлдеги DAT байланыш жерлеринин концентрациясы борбордук (+22%), dorsomedial (+25%) жана вентромедиалдык (+28%) куйруктуу ядро, жана аркасында (+16%) жана борбордук (+23%) путамен, тамак-аш менен бекемделген контролдоо менен салыштырганда. Мындан тышкары, алдынкы ядродо DAT менен байланышы кыйла жогору болгон (+37%) кокаинде- тамак-аш менен бекемделген маймылдарга салыштырмалуу. Precommissural striatum көбүрөөк каудалдык бөлүктөрдүн ичинде, DAT байланыш сайттардын тыгыздыгы борбордук каудат ядросунда бир кыйла жогору болгон (+21%) жана путамен, борбордук (+20%) жана вентралдык (+19%; Figure 1). Бул деңгээлдеги вентралдык стриатумдун ичинде, DAT менен байланышуу ядронун өзөгүндө кыйла жогору болгон (+20%) Жана жыт tubercle (+24%) кокаиндин кыртышында- vs тамак-аш менен бекемделген маймылдар.
90 күн бою өнөкөт кокаинди өзүн-өзү башкаруудан баш тартуунун кесепеттери
1 күндөн кийин кокаинге дуушар болгон жаныбарларда байкалган D30 рецепторлорун байланыштыруучу участоктордун тыгыздыгынын кеңири таралган айырмачылыктарынан айырмаланып, 90 күн бою абстиненциядан кийин, комиссиянын алдындагы комиссиянын кайсы бир бөлүгүндө тамак-аш менен күчөтүлгөн көзөмөлгө салыштырмалуу олуттуу айырмачылыктар болгон жок. стриатум (стол 1; Figure 1). Ошо сыяктуу эле, концентрациялары [3H]раклоприддин DA D2 рецепторлору менен 90 күн бою абстиненттен кийин прекоммиссуралдык стриатумдагы байланышы да дары-дармек менен байланышпаган жаныбарлардан олуттуу айырмаланган эмес (стол 2).
концентрациялары [3H]WIN 35428 DAT менен байланышкан D1 жана D2 рецепторлор менен байкалган окшош үлгү көрсөткөн. Кокаинге дуушар болгон маймылдардын ортосунда 90 күн бою абстиненттен кийин DAT тыгыздыгында баңги заты тийбеген контролдордогу деңгээлдерге салыштырмалуу олуттуу айырмачылыктар болгон эмес (стол 3; Figure 1), бул алдынкы өзөкчөлөр менен байланыштыруу жогорку денгээлде карай тенденция бар экенин белгилей кетүү керек.
ТАЛКУУЛОО
Биздин топтун мурунку изилдөөлөрү көрсөткөндөй, кокаиндин өзүн-өзү башкаруусунун өнөкөт таасири адам эмес приматтардын DA системасынын олуттуу дисрегуляциясы менен коштолот.Letchworth жана башкалар, 2001; Мур жана башкалар, 1998a, 1998b; Надер жана башкалар, 2002, 2006). Бул изилдөөнүн натыйжалары бул дисрегуляция кокаинди колдонууну токтоткондон кийин ачык бойдон калаарын көрсөтүп турат. 30 күн бою абстиненттен кийин, DA D1 рецепторлорунун жана DAT концентрациясы маймылдардын стриатумунда тамак-аш менен бекемделген контролго салыштырмалуу өнөкөт кокаинди өз алдынча башкаруу тарыхы менен бир кыйла жогорулаган. Бирок, бул изилдөө ошондой эле узак убакыт бою карманбоо (90 күн) кийин DA системасынын ичинде калыбына келтирүү үчүн ачык далилдерди берет, бул учурда кокаин дуушар болгон жана контролдоочу жаныбарлардын ортосунда олуттуу айырмачылыктардын жоктугу менен далилденген. Бул маалыматтар кокаиндин таасири DA системасында туруктуу өзгөрүүлөрдү жаратпашы мүмкүн экенин көрсөтүп турат, бирок бул калыбына келтирүү баңги затын колдонуудан узак убакыт бою баш тартуу менен болушу мүмкүн.
Абстиненттен кийин бул жерде көрсөтүлгөн DAT концентрациясынын дисрегуляциясы адам эмес приматтардагы мурунку отчетторго шайкеш келет (Letchworth жана башкалар, 2001), бул вентралдык жана дорсалдык стриатумдагы DAT байланышуу жерлеринин тыгыздыгынын олуттуу жогорулашын көрсөттү. Ачык текшерилбесе да, дары-дармектин таасири токтотулгандан кийин DAT байланышуучу жердин тыгыздыгынын жогорулоосу, жок эле дегенде, стриатумдун аймактарында эч кандай алып салуу мезгили жок билдирилгендерге караганда чоңураак жана кеңири таралган.Letchworth жана башкалар, 2001). Ошо сыяктуу эле, бул жерде 1 күн бою абстиненттен кийин байкалган D30 рецепторлорун байланыштыруучу жерлердин жогору концентрациясы кокаинди өз алдынча башкаруунун бирдей режимине дуушар болгон адам эмес приматтардын стриатумунда D1 сыяктуу рецепторлорду байланыштыруу тыгыздыгын көрсөткөн мурунку изилдөөлөр менен шайкеш келет (Надер жана башкалар, 2002). Ал эми кокаинге дуушар болгон жана контролдоочу жаныбарлардын стриатумундагы D2 сыяктуу кабылдагычтарды бириктирүү тыгыздыктарынын деңгээлдеринин ортосунда олуттуу айырмачылыктар байкалган эмес. Бул дисрегуляциянын жоктугу D2 рецепторлорунун концентрациясынын чоң төмөндөшүнө карабастан, эки адамда тең көз каранды болгондордо да болгон (Volkow жана башкалар, 1993) жана кокаинди өз алдынча башкаруунун жаныбарлар моделдери (Мур жана башкалар, 1998a, 1998b; Надер жана башкалар, 2002, 2006). Учурдагы маалыматтар, анда, D1 сыяктуу кабылдагычтарга жана DAT салыштырганда, бул системанын денгээлин башкаруу үчүн тезирээк нормалдаштыруу сунуш кылат. Чогуу алганда, DA рецепторлорундагы жана DATдагы өзгөрүүлөр кокаинди өз алдынча башкарууну токтоткондон кийинки мезгил өтө туруктуу экенин, DA системасын жөнгө салууда олуттуу өзгөрүүлөр болуп жатканын, бирок андан кийин кайра жөнгө салуу менен коштолорун ачык көрсөтүп турат. узакка созулган абстиненттен кийин DA рецепторлорунун жана DATтын нормалдуу бөлүштүрүлүшүнө жакындаган система.
D1 рецепторунун өзгөрүүлөрү
Кокаинди колдонууну токтоткондон кийин бул жерде байкалган D1 рецепторлорунун тыгыздыгынын кеңири таралган көтөрүлүшү D1 рецепторлорунун сезгичтигин көрсөткөн отчетторго шайкеш келет. D1 сезгичтигин өлчөө изилдөөдө жүргүзүлгөн Генри жана Уайт (1991), анда ядронун аккумбенсиндеги нейрондордун бирдик жазуулары туз менен иштетилген контролдорго салыштырмалуу кокаинди өнөкөт күнүмдүк инъекциялардан кийин D1 рецептордук агонист SKF 38393 үчүн көбүрөөк сезгич экени аныкталган. Бул таасир туруктуу болгон, анткени жогорулаган сезгичтик бир айга чейин байкалган. Авторлор D1 рецепторунун сезгичтиги somatodendritic A2 аймагындагы D10 авторецепторунун сезгичтигинен улам келип чыккан, ошону менен mesoaccumbens DA тутумундагы ингибитордук импульстун агымын азайтат деп болжолдогон.Генри менен Ак, 1991). Бул изилдөөнүн маалыматтары D1 рецепторлорун байланыштыруучу жерлердин абстиненциянын көбөйүшүн сунуштайт, бул допаминергиялык нейрондордун түздөн-түз таасир этүүчү D1 рецепторунун агонистине болгон сезгичтигин түшүндүрүшү мүмкүн. Мындан тышкары, SKF 38393 ядросунун нейрондорго күчөтүлгөн таасири, D1 кабылдагыч сезгичтигин калыбына келтирүү бар экенин көрсөтүп, эки айдан кийин айкын болгон эмес.Генри менен Ак, 1991); бул изилдөөдө белгиленген D1 кабылдагыч тыгыздыгын калыбына келтирүү менен шайкеш натыйжа 90 күн бою абстиненттүү кийин. Башка отчеттор да рецидивде D1 рецепторлорунун маанилүү ролун колдойт. Ядронун кабыгындагы D1 рецепторлорун түз стимулдаштыруу абстинент кемирүүчүлөрдүн кокаинди издөөнү калыбына келтириши мүмкүн (Schmidt жана башкалар, 2006). Бирок, адабияттар D1 агонисттери да, антагонисттери да кокаиндин примерлери же кокаин менен байланышкан стимулдардан келип чыккан баңги заттарын издөөнү басаңдата ала тургандыгы менен бир аз карама-каршы келет.Alleweireldt жана башкалар, 2002; Де жана башкалар, 1999; Хроян жана башкалар, 2000; өз алдынча башкаруу жана башкалар, 1996; Weiss жана башкалар, 2001). Жакында, Хроян жана башкалар (2003) D1 агонисттери жана антагонисттери кокаинди издөөнүн адамдык эмес примат моделиндеги рецидивди азайтат деп билдирди. Бул авторлор кокаинди издөө үчүн зарыл болгон D1 рецептордук активдүүлүгүнүн критикалык диапазону болушу мүмкүн жана антагонисттер да, агонисттер да бул терезеден активдүүлүктү жылдырышы мүмкүн деп божомолдошот. Абстиненция менен коштолгон D1 рецепторлорунун концентрациясынын жогорулашы бул диапазонду өзгөртүшү мүмкүн, натыйжада бул системанын сезгичтигинин өзгөрүшүнө алып келет. Дагы бир жагдай, D1 активдүүлүгү D2 рецепторлорунун активдүүлүгүн модуляциялоо үчүн аракет кылышы мүмкүн (Nolan жана башкалар, 2007; Раскин жана башкалар, 1999; Уолтерс жана башкалар, 1987). Бул маалыматтар D1 жана D2 рецепторлорунун катышы абстиненттин жүрүшүндө өзгөрүп, ошону менен бул модуляциянын эффективдүүлүгүн өзгөртүшү мүмкүн экенин көрсөтүп турат.
Азыркы маалыматтардан айырмаланып, өнөкөт кокаинди өзүн-өзү башкаруудан кийин D1 рецепторлорунун деңгээли төмөндөгөндүгү жөнүндө кабарлар бар (Мур жана башкалар, 1998a), бул изилдөөлөрдүн ортосунда олуттуу айырмачылыктар бар, мисалы, кокаиндин дозасы жана узактыгы, жалпы кабыл алуу жана салыштырмалуу контролдук топтор. Ошентип, бириктирилген далилдер D1 системасы өнөкөт кокаинди башкаруудан чыккандан кийин олуттуу агымда экенин көрсөтүп турат.
Допамин ташуучу өзгөрүүлөр
Баңги затын колдонууну токтоткондон кийин кокаинге дуушар болгон жаныбарлардын стриатумундагы DAT концентрациясынын жогорулашынын табылгалары адам эмес приматтарда кокаинди өз алдынча башкаруу менен коштолгон DAT байланыш сайттарынын деңгээли жогорулаганын көрсөткөн мурунку изилдөөлөрүбүздү кеңейтет. Учурдагы маалыматтар бул дисрегуляция абстиненциянын алгачкы фазаларында сакталып турганын көрсөтүп турат. Андан тышкары, алар контролдук деңгээлге чейин калыбына келтирүү салыштырмалуу узакка созулат (учурдагы изилдөөдө 90 күнгө чейин). Биздин мурунку изилдөөлөрүбүздө биз алгач негизинен вентралдык стриаталдык аймактар менен чектелсе да, DAT байланыш жерлеринин тыгыздыгынын өзгөрүшү кокаиндин узак мөөнөттүү таасири менен стриатумдун дорсалдык жана ростралдык бөлүктөрүн камтышы үчүн кеңейгендигин көрсөттүк.Letchworth жана башкалар, 2001; Porrino жана башкалар, 2004). Бул изилдөөдө абстиненция учурунда DAT концентрациясынын контролдук деңгээлдерине кайтуу вентралдык стриатумга караганда дорсалдык стриатумда көбүрөөк жана тезирээк болуп көрүндү жана ошентип, кокаиндин өнөкөт таасири менен шартталган эффекттердин үлгүсүнө тескери анатомиялык траекторияны ээрчип жаткандай көрүндү. .
Бул маалыматтар адам кокаин колдонуучулардын отчетторуна да дал келет (кичине жана башкалар, 1999; Malison жана башкалар, 1998; Mash жана башкалар, 2002; Стейли жана башкалар, 1994) башкаруу органдарына салыштырмалуу стриатумдагы DAT сайттары менен байланыштын жогорку деңгээлин көрсөттү, эң байкаларлык жогорулаш вентралдык стриатумда жайгашкан. Жакында эле, бул бийиктиктер везикулярдык моноамин ташуучу 2 (VMAT2) менен байланыштын олуттуу төмөндөшү менен коштолгон.кичине жана башкалар, 2003), DA нейрондорунун иш жүзүндө жоголушу жөнүндө. Авторлор DATтин жогорулашы кокаин менен фармакологиялык блокадага компенсациялык жоопко түздөн-түз байланыштуу деген тыянакка келишкен, ал эми VMAT2дин төмөндөшү DA метаболизминдеги жалпы өзгөрүүлөрдү чагылдырып, гиподопаминергиялык функцияга алып келет.
Кокаинге көз каранды адамдардын дени сак адамдар менен салыштырганда, метилфенидат чакырыгына жооп катары, ПЭТ менен өлчөнгөн вентралдык стриатумда DA концентрациялары төмөндөгөн.Volkow жана башкалар, 1997). Жакында, Martinez жана башкалар (2007) кокаин колдонуучулар вентралдык стриатум жана путамендеги амфетамин чакырыгына катуу жооп беришкенин билдирди. Андан тышкары, амфетамин менен шартталган DA чыгаруунун төмөндөшү өзүнчө өзүн-өзү башкаруу сессияларында кокаинди тандоо менен байланыштырылды, андыктан амфетаминге жооп катары DA-нын эң төмөн деңгээли бар колдонуучулар кокаинди альтернативалуу күчөткүчкө караганда тандашат.Martinez жана башкалар, 2007). Кокаинди колдонуунун кемирүүчүлөрдүн моделдериндеги акыркы изилдөөлөр да бул идеяны колдойт. Mateo жана башкалар (2005), мисалы, өнөкөт кокаиндин өз алдынча башкаруусунун таасири DAT функциясынын өзгөрүшү менен байланышкан. Бул изилдөөчүлөр базалык DA алуу көбөйгөнүн көрсөттү, натыйжада синаптикалык DA тезирээк тазаланат, демек, клеткадан тышкаркы DAнын базалдык деңгээли же гиподопаминергиялык абал төмөндөдү. Ошентип, бул изилдөөдө байкалган өнөкөт кокаинди өзүн-өзү башкаруудан чыккандан кийин DAT концентрациясынын жогорулашы компенсациялык жоопту көрсөтүп, клеткадан тышкаркы DAнын төмөнкү баштапкы деңгээлине алып келиши мүмкүн.
D2 рецепторунун өзгөрүүлөрү
Ушул изилдөөнүн бир натыйжасы, D2 рецепторлорунун концентрациялары 30 күн бою абстиненттен кийин контролдук маанилерге кайтып келген, ал эми алып салуу мезгили жок жаныбарларда байкалган олуттуу төмөндөө менен салыштырганда (Надер жана башкалар, 2002). Азыркы иликтөөдөн айырмаланып, адамдын сүрөтүн изилдөөлөр жалпысынан D2 рецепторлордун деңгээли кокаиндин өнөкөт таасиринен узак убакыт бою кармангандан кийин контролдукка караганда төмөн экенин аныкташкан (Martinez жана башкалар, 2004; Volkow жана башкалар, 1993). Бул адамдык изилдөөлөр менен азыркы адамдык эмес приматтык иликтөөнүн ортосундагы айырмачылыктардын потенциалдуу түшүндүрмөсү кокаинди кабыл алуунун үлгүсүндөгү жана узактыгындагы айырмачылыктарды, ошондой эле адам көз карандыларында D2 рецепторлорунун төмөнкү деңгээлдеринин алдын ала болушу мүмкүндүгүн камтыйт.
Акыркы идеяга ылайык, дени сак адамдарда D2 рецепторлорунун төмөнкү базалдык деңгээли стимуляторлордун күчөтүлгөн эффективдүүлүгүн болжолдойт деген далилдер бар. метилфенидате (Volkow жана башкалар, 1999Маймылдардагы D2 рецепторлорунун баштапкы деңгээли кокаинди өз алдынча башкарууга ыктуулукту болжолдогон.Морган жана башкалар, 2002; Надер жана башкалар, 2006). Түрлөр боюнча бул табылгалардын параллелдүү мүнөзүнө ылайык, эки адам тең (Volkow жана башкалар, 1993) жана адам эмес примат (Надер жана башкалар, 2006) Сүрөттөө изилдөөлөрү кокаиндин таасиринен баш тарткандан кийин D2 рецепторлорунун жеткиликтүүлүгүнүн төмөн деңгээлин көрсөттү. Белгилей кетсек, акыркы экспериментте кокаинди өз алдынча башкаруунун графиги (Надер жана башкалар, 2006) бул изилдөөдө колдонулган күчөтүү графигине окшош болгон. Ошентип, бул эки изилдөөнүн окшош эмес натыйжалары, мисалы, кокаинди өз алдынча башкаруу учурунда күчөтүү графиги же кумулятивдүү кабыл алуу сыяктуу методологиялык айырмачылыктарга байланыштуу болушу күмөн.
Көбүрөөк түшүндүрмө DA системасынын функционалдык динамикасын камтыйт. PET менен D2 рецепторлорунун жеткиликтүүлүгүнүн чаралары "функционалдык" деп мүнөздөлгөн, анткени сигнал белоктун көлөмүнө (бул учурда D2 рецепторлорунун тыгыздыгы) жана циркуляциядагы нейротрансмиттердин деңгээлине байланыштуу. (кара Laruelle, 2000; Надер жана Czoty, 2008 андан ары талкуулоо үчүн). Иn карама-каршы, кабылдагыч autoradiography DA жүгүртүүдөгү денгээлде булганган эмес. Ошентип, бул изилдөө, биздин мурунку иш менен бирге, D2 кабылдагыч тыгыздыгы кокаин өзүн-өзү башкаруу менен азайган, бирок кабылдагыч денгээлдери абстинент учурунда калыбына келет деп болжолдойт. Ушундай эле жүргүзүлгөн PET изилдөөсүндө (Надер жана башкалар, 2006), калыбына келтирүү беш маймылдардын үчөө байкалган. Учурдагы табылгалар бул маймылдардын D2 рецепторлордун тыгыздыгы боюнча айырмаланбагандыгын көрсөтүп турат, бирок, балким, DA тутумунун жооп берүү жөндөмдүүлүгү (б.а. абстиненция учурунда айлануучу DA деңгээли) "калыбына келтирилген" жана "айыкпаган" субъекттердин ортосунда айырмаланган.
чектөөлөр
Учурдагы изилдөөлөрдүн бир маанилүү чектөөсү, биздин изилдөөлөр D1 жана D2 рецепторлорунун же DATтын иштешин чече албайт. Тескерисинче, биз кабылдагыч белоктордун тыгыздыгынын өзгөрүшүн гана карап чыктык. Натыйжалар бул системалардын потенциалдуу ролуна таасир этсе да, бул жерде көрсөтүлгөн өзгөрүүлөрдүн жүрүм-турумдук кесепеттерин чагылдыруу үчүн мындан аркы изилдөөлөр талап кылынат. Бул изилдөөнүн дагы бир чектөөсү, авторадиографиялык лиганддар көп учурда өз максаттарынын клетка ичиндеги жана цитоплазмалык жерлеринин ортосунда ажырай алышпайт, анткени радиобелгиленген антагонисттер көбүнчө мембрана өткөрүүчү. кичине жана башкалар (2002) кокаиндин өнөкөт таасири трансфекцияланган клеткаларда клетка ичиндеги DAT концентрациясынын төмөндөшү менен бир убакта мембрана бетинде DATтын олуттуу жогорулашына алып келерин көрсөттү. Жакында, Самувел жана башкалар (2008) келемиш стриаталдык синаптосомалык препараттарда ушундай эле табылганы билдирди. Бул жыйынтыктар ушул изилдөөдө байкалган DAT бөлүштүрүү өзгөртүүлөр клетка ичиндеги сайттарга эмес, мембрана бетиндеги өзгөрүүлөрдү чагылдырышы мүмкүн экенин көрсөтүп турат.
Акыр-аягы, узакка созулган (90 күн) абстинентти изилдөөнүн чечмелөөсүнө этият болуу керек, анткени бул жыйынтыктар салыштырмалуу азыраак жаныбарлар тобуна негизделген (N=3). Субъекттердин саны аз болгонуна карабастан, бул топтон алынган маалыматтар ырааттуу болгон. Figure 2. Striatum боюнча D1 кабылдагычтарды байланыштыруучу сайттардын концентрациясы топтордун ичинде аз өзгөрмөлүүлүгүн көрсөттү, бул табылгалардын ишенимдүүлүгүн көрсөтүп турат. Окшош ырааттуулук D2 кабылдагыч жана DAT байланыш анализдеринин маалыматтарында да айкын болгон. Этияттык керек болсо да, бул маалыматтар стриатумдагы DAT жана DA рецепторлорунун концентрациясын калыбына келтирүү узакка созулган абстиненция менен пайда болушу мүмкүн экенин көрсөтүп турат.
Figure 2.
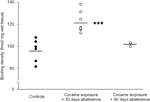 Жеке жаныбарлар үчүн D1 рецепторлорунун байланыш тыгыздыгы тамак-ашты күчөтүүдөн (көзөмөлдөө) же өнөкөт кокаинди өз алдынча башкаруудан 30 же 90 күн бою баш тартуудан кийин бүт стриатум боюнча орточо. топтор үчүн каражаттар кара тилкелер менен сүрөттөлгөн, ***p<0.001 тамак-аш менен күчөтүлгөн контролдоо менен салыштырганда.
Жеке жаныбарлар үчүн D1 рецепторлорунун байланыш тыгыздыгы тамак-ашты күчөтүүдөн (көзөмөлдөө) же өнөкөт кокаинди өз алдынча башкаруудан 30 же 90 күн бою баш тартуудан кийин бүт стриатум боюнча орточо. топтор үчүн каражаттар кара тилкелер менен сүрөттөлгөн, ***p<0.001 тамак-аш менен күчөтүлгөн контролдоо менен салыштырганда.
КОРУТУНДУ
Жыйынтыктап айтканда, кокаинди өз алдынча башкаруунун таасири DA системаларын жөнгө салууда олуттуу өзгөрүүлөрдү жаратты, алар абстиненттин алгачкы фазаларында (биринчи 30 күн) сакталды. Бул D1 рецепторлорунун жана DATтын концентрациясын жөнгө салууда, алардын өзгөрүшүнүн чоңдугу жана топографиялык масштабы боюнча эң айкын болгон. Тескерисинче, кокаиндин таасиринен баш тартуунун узактыгы менен нормалдашуу үчүн далилдер бар, муну менен 1 күндөн кийин DAT, D2 жана D90 рецепторлорунун концентрациялары дары-дармекке дуушарланбаган контролдордон айырмаланган эмес. Tбирок бул системалар сөзсүз эле калыбына келүүнүн бирдей убактылуу курсун ээрчибейт, бул DA деңгээлин жөнгө салууда айрым туруксуздук болушу мүмкүн деп болжолдойт, айрыкча абстиненциянын башында. Бул допаминергиялык дисрегуляция кокаинге көз каранды болгондорго берилүүчү ар кандай потенциалдуу фармакотерапиянын натыйжалуулугуна таасирин тийгизиши мүмкүн, айрыкча, эгерде дары анын иш-аракет механизми үчүн DA тутумуна таянса.
жазуулар
АЧЫККА ЧЫГАРУУ/ЧЫР пайыздык
Авторлор ачыкка чыгара турган кызыкчылыктардын кагылышуусу жок.
шилтемелер
- Alleweireldt AT, Weber SM, Kirschner KF, Bullock BL, Neisewander JL (2002). D1 дофамин рецепторлорун блокадалоо же стимулдаштыруу келемиштерде өчүп калган кокаинди издөө жүрүм-турумунун калыбына келишин басаңдатат. Psychopharmacology (Берл) 159: 284–293. | макала | PubMed | ChemPort |
- Beverridge TJ, Smith HR, Daunais JB, Nader MA, Porrino LJ (2006). Өнөкөт кокаинди өзүн-өзү башкаруу адам эмес приматтардын убактылуу бөлүкчөлөрүндөгү функционалдык активдүүлүктүн өзгөрүшү менен байланыштуу. Eur J Neurosci 23: 3109–3118. | макала | PubMed |
- Канфилд ДР, Спиалман РД, Кауфман МЖ, Мадрас БК (1990). Маймылдын мээсинде [3H] CFT ([3H] WIN 35428) тарабынан кокаинди байланыштырган жерлердин авторадиографиялык локализациясы. синапс 6: 189–195. | макала | PubMed | ChemPort |
- De Vries TJ, Schoffelmeer AN, Binnekade R, Vanderschuren LJ (1999). Допаминергиялык механизмдер IV баңги затын өз алдынча башкарууну узак мөөнөткө алып салгандан кийин кокаин менен героинди издөөгө түрткү берет. Psychopharmacology (Берл) 143: 254–260. | макала | PubMed | ChemPort |
- Farfel GM, Kleven MS, Woolverton WL, Seiden LS, Perry BD (1992). Кокаинди кайталап инъекциялоонун катехоламиндик рецепторлорду байланыштырган жерлерине, допаминди ташыгычты бириктирүүчү жерлерге жана резус маймылынын жүрүм-турумуна тийгизген таасири. Brain Рез 578: 235–243. | макала | PubMed | ChemPort |
- Graham J, Porrino LJ (1995). Кокаиндин өзүн-өзү башкаруусунун нейроанатомиялык субстраттары. CRC: Бока Ратон, Флорида.
- Hemby SE, Co C, Koves TR, Smith JE, Dworkin SI (1997). Келемиште жоопко көз каранды жана жоопко көз каранды эмес кокаинди башкаруу учурунда ядродогу допаминдин клеткадан тышкаркы концентрациясындагы айырмачылыктар. Psychopharmacology (Берл) 133: 7–16. | макала | PubMed | ChemPort |
- Henry DJ, White FJ (1991). Кокаинди кайталап башкаруу келемиш ядросунун ичиндеги D1 допаминдик рецепторлордун сезгичтигинин туруктуу жогорулашына алып келет. J Pharmacol Exp Ther 258: 882–890. | PubMed | ChemPort |
- Jacobsen LK, Staley JK, Malison RT, Zoghbi SS, Seibyl JP, Kosten TR жана башкалар (2000). Кокаинге көз каранды болгон бейтаптарда борбордук серотонинди байланыштыргычтын болушу жогорулаган. Am J психиатрия 157: 1134–1140. | макала | PubMed | ISI | ChemPort |
- Khroyan TV, Barrett-Larimore RL, Rowlett JK, Spealman RD (2000). Кокаин издеген жүрүм-турумдун кайталанышында допамин D1 жана D2 сыяктуу рецепторлордун механизмдери: тандалма антагонисттердин жана агонисттердин таасири. J Pharmacol Exp Ther 294: 680–687. | PubMed | ISI | ChemPort |
- Khroyan TV, Platt DM, Rowlett JK, Spealman RD (2003). Адам эмес приматтарда допамин D1 рецепторунун агонисттери жана антагонисттери тарабынан кокаинди издөөгө болгон рецидивдин басаңдашы. Psychopharmacology (Берл) 168: 124–131. | макала | PubMed | ChemPort |
- Laruelle M (2000). менен синаптикалык нейротрансмиссияны сүрөттөө бододо, милдеттүү атаандаштык ыкмалары: сын карап чыгуу. J Cereb Blood Flow Metab 20: 423–451. | макала | PubMed | ISI | ChemPort |
- Летчворт СР, Надер М.А., Смит HR, Фридман ДП, Поррино Л.Ж. (2001). Резус маймылдарында кокаинди өз алдынча башкаруунун натыйжасында допаминди ташыгычтын тыгыздыгынын өзгөрүшүнүн жүрүшү. J Neurosci 21: 2799–2807. | PubMed | ISI | ChemPort |
- Лидоу МС, Голдман-Ракич П.С., Галлагер Д.У., Ракич П (1991). Примат мээ кыртышында допаминергиялык рецепторлордун бөлүштүрүлүшү: [3H] raclopride, [3H] спиперон жана [3H] SCH 23390 колдонуу менен сандык autoradiographic талдоо. Neuroscience 40: 657–671. | макала | PubMed | ChemPort |
- Little KY, Elmer LW, Zhong H, Scheys JO, Zhang L (2002). Кокаиндин плазма мембранасына допамин ташуучу трафиктин индукциясы. Mol Pharmacol 61: 436–445. | макала | PubMed | ISI | ChemPort |
- Little KY, Krolewski DM, Zhang L, Cassin BJ (2003). Адам кокаин колдонуучулардын striatal везикулярдуу моноамин ташуучу протеинди (VMAT2) жоготуу. Am J психиатрия 160: 47–55. | макала | PubMed |
- Кичинекей KY, Чжан Л, Десмонд Т, Фрей К.А., Далак ГВ, Кассин БДж (1999). Кокаинди колдонуучулардын стриаталдык допаминергиялык аномалиялары. Am J психиатрия 156: 238–245. | PubMed | ChemPort |
- Malison RT, Best SE, van Dyck CH, McCance EF, Wallace EA, Laruelle M жана башкалар (1998). [123I] бета-CIT SPECT менен өлчөнгөн курч кокаин абстиненция учурунда көтөрүлгөн striatal дофамин ташыгычтары. Am J психиатрия 155: 832–834. | PubMed | ChemPort |
- Мартинес Д, Брофт А, Фолтин РВ, Слифштейн М, Хванг ДР, Хуанг Ы жана башкалар (2004). Кокаинге көз карандылык жана striatum функционалдык бөлүмдөрүндө d2 кабылдагыч болушу: кокаин издөө жүрүм-туруму менен байланышы. Neuropsychopharmacology 29: 1190–1202. | макала | PubMed | ISI | ChemPort |
- Мартинес Д, Нарендран Р, Фолтин РВ, Слифштейн М, Хванг ДР, Брофт А жана башкалар (2007). Амфетамин менен шартталган допаминдин чыгарылышы: кокаинге көз карандылыктын кескин начарлашы жана кокаинди өз алдынча башкарууну тандоону болжолдоо. Am J психиатрия 164: 622–629. | макала | PubMed | ISI |
- Mash DC, Pablo J, Ouyang Q, Hearn WL, Izenwasser S (2002). Допаминди ташуу функциясы кокаинди колдонуучуларда жогорулайт. J Neurochem 81: 292–300. | макала | PubMed | ChemPort |
- Mateo Y, Lack CM, Morgan D, Roberts DC, Jones SR (2005). Кокаинди өз алдынча башкаруудан жана ажыратуудан кийин допаминдин терминалдык функциясы жана кокаинге сезимсиздик кыскарган. Neuropsychopharmacology 30: 1455–1463. | макала | PubMed | ISI | ChemPort |
- Melega WP, Jorgensen MJ, Lacan G, Way BM, Pham J, Morton G жана башкалар (2008). Вервет маймылында узак мөөнөттүү метамфетаминди башкаруу адамдын таасиринин аспектилерин моделдейт: мээнин нейротоксиктиги жана жүрүм-турум профилдери. Neuropsychopharmacology 33: 1441–1452. | макала | PubMed | ChemPort |
- Мур РДЖ, Винсант С.Л., Надер МА, Поррино Л.Ж., Фридман ДП (1998a). Кокаинди өз алдынча башкаруунун резус маймылдарындагы стриаталдык допамин D1 рецепторлоруна тийгизген таасири. синапс 28: 1–9. | макала | PubMed | ChemPort |
- Мур РДЖ, Винсант С.Л., Надер М.А., Поррино Л.Ж., Фридман ДП (1998b). Кокаинди өз алдынча башкаруунун резус маймылдарындагы допамин D2 рецепторлоруна тийгизген таасири. синапс 30: 88–96. | макала | PubMed | ChemPort |
- Морган Д, Грант К.А., Гейдж HD, Mach RH, Каплан Дж.Р., Приоло О жана башкалар (2002). Маймылдардагы социалдык үстөмдүк: допамин D2 рецепторлору жана кокаинди өз алдынча башкаруу. Nat Neurosci 5: 169–174. | макала | PubMed | ISI | ChemPort |
- Nader MA, Czoty PW (2008). Адамдык эмес приматтарда мээнин сүрөтү: баңгизатка көз карандылыкты түшүнүү. ИЛАР Ж 49: 89–102. | PubMed | ChemPort |
- Nader MA, Daunais JB, Moore T, Nader SH, Moore RJ, Smith HR жана башкалар (2002). Кокаинди өзүн-өзү башкаруунун резус маймылдарындагы стриаталдык допаминдик системаларга тийгизген таасири: баштапкы жана өнөкөт экспозиция. Neuropsychopharmacology 27: 35–46. | макала | PubMed | ISI | ChemPort |
- Nader MA, Morgan D, Gage HD, Nader SH, Calhoun TL, Buchheimer N жана башкалар (2006). Маймылдарда өнөкөт кокаинди өз алдынча башкаруу учурунда допамин D2 рецепторлорунун PET сүрөтү. Nat Neurosci 9: 1050–1056. | макала | PubMed | ChemPort |
- Нолан Э.Б., Харрисон Л.М., Лахосте ГДж., Рускин Д.Н. (2007). Чычкандардагы D (1) жана D (2) дофамин рецепторлорунун ортосундагы жүрүм-турум синергизми боштук түйүндөрүнөн көз каранды эмес. синапс 61: 279–287. | макала | PubMed | ChemPort |
- Porrino LJ, Lyons D, Miller MD, Smith HR, Friedman DP, Daunais JB жана башкалар (2002). Адам эмес приматтагы өзүн-өзү башкаруунун баштапкы этаптарында кокаиндин таасиринин метаболикалык картасы. J Neurosci 22: 7687–7694. | PubMed | ChemPort |
- Porrino LJ, Lyons D, Smith HR, Daunais JB, Nader MA (2004). Кокаинди өз алдынча башкаруу лимбиялык, ассоциациялык жана сенсомотордук стриаталдык домендердин прогрессивдүү катышуусун жаратат. J Neurosci 24: 3554–3562. | макала | PubMed | ISI | ChemPort |
- Ruskin DN, Bergstrom DA, Walters JR (1999). Globus pallidusдагы атуу ылдамдыгынын көп секунддук термелүүсү: D1 жана D2 допаминдик рецепторлор тарабынан синергетикалык модуляция. J Pharmacol Exp Ther 290: 1493–1501. | PubMed | ISI | ChemPort |
- Samuvel DJ, Jayanthi LD, Manohar S, Kaliyaperumal K, карагыла RE, Ramamoorthy S (2008). Келемиштерде кокаинди өзүн-өзү башкаруудан баш тарткандан кийин допамин ташуучу трафигинин дисрегуляциясы жана иштеши: каудат путамендеринде жана ядронун аккументтеринде дифференциалдык жөнгө салуу үчүн далилдер. J Pharmacol Exp Ther 325: 293–301. | макала | PubMed | ChemPort |
- Шмидт HD, Андерсон СМ, Пирс RC (2006). Кабыктагы D1 сыяктуу же D2 допаминдик рецепторлорду стимулдаштыруу, бирок ядронун ядросунун өзөгү эмес, келемиште кокаинди издөө жүрүм-турумун калыбына келтирет. Eur J Neurosci 23: 219–228. | макала | PubMed |
- Self DW, Barnhart WJ, Lehman DA, Nestler EJ (1996). D1 жана D2 сыяктуу допаминдик рецепторлордун агонисттери тарабынан кокаин издөө жүрүм-турумунун карама-каршы модуляциясы. илим 271: 1586–1589. | макала | PubMed | ISI | ChemPort |
- Staley JK, Hearn WL, Ruttenber AJ, Wetli CV, Mash DC (1994). Кокаиндин ашыкча дозасын колдонуунун курмандыктарында дофаминди ташуучу жогорку жакындыктагы кокаинди таануу сайттары жогорулайт. J Pharmacol Exp Ther 271: 1678–1685. | PubMed | ChemPort |
- Volkow ND, Fowler JS, Wang Г.Ж., Hitzemann R, Логан J, Schlyer DJ жана башкалар (1993). Допамин D2 рецепторлорунун азайышы кокаинди кыянаттык менен пайдалануучулардын фронталдык метаболизминин төмөндөшү менен байланыштуу. синапс 14: 169–177. | макала | PubMed | ISI | ChemPort |
- Volkow ND, Wang GJ, Fowler JS, Logan J, Gatley SJ, Gifford A жана башкалар (1999). Мээ допамин D2 кабылдагыч деңгээли боюнча адамдарда психостимуляторлорго реакцияларды күчөтүү. Am J психиатрия 156: 1440–1443. | PubMed | ISI | ChemPort |
- Volkow ND, Wang Г.Ж., Fowler JS, Логан J, Gatley SJ, Hitzemann R жана башкалар (1997). Детоксикацияланган кокаинге көз каранды субъекттерде стриаталдык допаминергикалык реакциянын төмөндөшү. жаратылыш 386: 830–833. | макала | PubMed | ISI | ChemPort |
- Walters JR, Bergstrom DA, Carlson JH, Chase TN, Braun AR (1987). D1 допамин кабылдагыч активдештирүү D2 агонист эффекттеринин постсинаптикалык билдирүүсү үчүн талап кылынат. илим 236: 719–722. | макала | PubMed | ChemPort |
- Weiss F, Martin-Fardon R, Ciccocioppo R, Kerr TM, Smith DL, Ben-Shahar O (2001). Баңгизатка байланыштуу сигналдар менен шартталган кокаинди издөө жүрүм-турумунун жок болушуна туруктуу каршылык. Neuropsychopharmacology 25: 361–372. | макала | PubMed | ISI | ChemPort |
- Вонг ДФ, Кувабара Н, Шретлен диджей, Бонсон КР, Чжоу Ю, Нанди А жана башкалар (2006). Кокаинди кумарлануу учурунда адамдын стриатумунда дофамин рецепторлорунун көбөйүшү. Neuropsychopharmacology 31: 2716–2727. | макала | PubMed | ChemPort |