Фронт Бехав Неуросци. 2015 19. мај;9:128. дои: 10.3389/фнбех.2015.00128. еЗбирка 2015.
Андреата М1, Паули П1.
Апстрактан
У класичном условљавању, првобитно неутрални стимулус (условљени стимулус, ЦС) постаје повезан са биолошки истакнутим догађајем (безусловни стимулус, УС), који може бити бол (аверзивно условљавање) или храна (условљавање апетита). Након неколико асоцијација, ЦС је у стању да иницира или дефанзивне или конзумативне одговоре, респективно. За разлику од аверзивног условљавања, условљавање апетита се ретко истражује код људи, иако је његов значај за нормално и патолошко понашање (нпр. гојазност, зависност) неоспоран. Ова студија намерава да преведе налазе животиња о кондиционирању апетита на људе који користе храну као САД. Тридесет и троје учесника испитано је између 8 и 10 сати без доручка како би се уверило да су гладни. Током две фазе аквизиције, један геометријски облик (авЦС+) је предвидео аверзиван УС (болни електрични удар), други облик (аппЦС+) је предвиђао апетитиван УС (чоколадни или слани перец према жељи учесника), а трећи облик (ЦС– ) није предвидео ни САД. У фази изумирања, ова три облика плус нови облик (НОВО) поново су представљени без испоруке у САД. Оцене валенције и узбуђења, као и реакције проводљивости запрепашћења и коже (СЦР) су прикупљене као индекси учења. Пронашли смо успешно аверзивно и апетитивно условљавање. С једне стране, авЦС+ је оцењен као негативнији и узбудљивији од ЦС– и изазвао је потенцирање узбуне и побољшани СЦР. С друге стране, аппЦС+ је оцењен позитивнијим од ЦС– и изазвао је слабљење шока и већи СЦР. Укратко, успешно смо потврдили налазе на животињама код (гладних) људи демонстрирајући апетитивно учење и нормално аверзивно учење.
увод
Предвиђање опасности и хране је од пресудног значаја за опстанак сваког организма. У класичном кондиционирању (Павлов, КСНУМКС), знак претходи аверзивном догађају, као што је благи болни електрични шок (аверзивни безусловни стимулус, УС) или догађају са апетитом, као што је куглица хране (апетитивна САД), неколико пута. Након тога, сам овај стимуланс (сада означен са ЦС, ЦС+) може да изазове или одбрамбене или конзумативне одговоре, респективно. Прва врста асоцијативног учења назива се аверзивно условљавање, док се друга назива апетитивно условљавање.
Иако је предвиђање догађаја са апетитом једнако важно за преживљавање као и предвиђање аверзивних догађаја, условљавање апетита је значајно мање истражено код животиња (Боутон и Пецк, 1989; Коцх ет ал., КСНУМКС; МцДанналд ет ал., 2011, за преглед погледајте Мартин-Соелцх ет ал., КСНУМКС) као и код људи (Клуцкен ет ал., КСНУМКС, 2013; Аустин и Дука, КСНУМКС; Делгадо ет ал., КСНУМКС; Леви и Глимчер, 2011). Овај недостатак истраживања може бити последица сложености парадигме апетита у поређењу са аверзивном. На пример, храна као примарно појачање мора да се испоручује када је организам гладан да би био награђен (за недавни преглед погледајте Дицкинсон и Баллеине, КСНУМКС; Цларк ет ал., КСНУМКС). У истраживању људи, ова потешкоћа је превазиђена коришћењем новца (Аустин и Дука, КСНУМКС; Делгадо ет ал., КСНУМКС; Леви и Глимчер, 2011) или еротске слике (Клуцкен ет ал., КСНУМКС, 2013). Међутим, студије неуро-имагинга су истакле да примарни (тј. грицкалице или пића) и секундарни (тј. новац) појачавачи активирају неке уобичајене регије мозга (нпр. стријатум), али такође укључују различите обрасце активације (Делгадо ет ал., КСНУМКС; Леви и Глимчер, 2011).
Колико знамо, само неколико студија о кондиционирању код људи је испитало ефекте примарних појачавача апетита као што је мирис (Готтфриед ет ал., 2002), вода (Кумар ет ал., КСНУМКС), или храну (Превост ет ал., КСНУМКС). У првој студији, Готтфриед ет ал. (2002) повезана неутрална лица (ЦС) са непријатним, пријатним или неутралним мирисом. Занимљиво је да су открили већу активацију у орбитофронталном кортексу (ОФЦ) и у вентралном стриатуму као одговор на апетитивни ЦС+ у односу на аверзивни ЦС+. Закључили су да ОФЦ обрађује вредност мириса и учествује у преношењу афективне вредности са олфакторног (УС) на визуелни (ЦС) систем. (вентро)стриатална активација је протумачена као одраз апетитног ЦР-а који изазива апетитивни ЦС+. У другој студији, Кумар ет ал. (КСНУМКС) позвали своје учеснике (здраве контролне групе и пацијенте са великом депресијом) у лабораторију рано ујутру и замолили их да се уздрже од пића током ноћи како би били сигурни да су жедни пре скенирања. Фракталне слике су биле ЦС и 0.1 мл воде у САД. Занимљиво је да су, између осталих активација, здрави учесници (али не и пацијенти са депресијом) показали већу активацију у вентралном стриатуму до апетитног ЦС+, што сугерише да је овај стимулус обрађен као награда. У трећој студији, Превост и др. (2012) такође су представили фракталне слике као ЦС и слатке или слане грицкалице као УС према жељи учесника. ЦС је представљен 6 с, ау последњој секунди је додатно представљена слика хране. Сваки пут када би се слика хране појавила, експериментатор је стављао комад хране у руке учесника, којима је било дозвољено да одмах поједу ужину. Нажалост, аутори нису пријавили активацију мозга апетитном ЦС+ током класичне фазе кондиционирања; међутим, приметили су мање успоравање срца до награђеног ЦС+ у поређењу са ненаграђеним ЦС–, што указује на диференцијалне ефекте кондиционирања.
Недостатак студија које користе реакцију запрепашћења као индекс за условљавање апетита је изненађујући, посебно имајући у виду његову широку употребу у аверзивном условљавању. Реакција запрепашћења је прастари и аутоматски одбрамбени одговор на изненадне, неочекиване и јаке аверзивне догађаје (Коцх, КСНУМКС). Овај одбрамбени одговор је посредован релативно једноставним неуронским путем који укључује неуроне кохлеарног корена, каудално понтинско језгро ретикуларне формације (ПнЦ) и спиналне мотонеуроне (Фендт и Фанселов, 1999; Коцх, КСНУМКС). Студије на животињама су откриле да потенцирање страха зависи од пројекција од амигдале до ПнЦ (Фендт и Фанселов, 1999; Коцх, КСНУМКС), док слабљење страха зависи од интактног нуцлеус аццумбенс (НАцц, Коцх, КСНУМКС). Таква модулација одговора на стрепњу је корисна имплицитна мера за валентност представљених стимулуса у првом плану. Дакле, потенцирање указује на негативну валенцију, док слабљење указује на позитивну валенцу, оба без великог утицаја когнитивних процеса (Хам и Вејке, 2005; Андреатта ет ал., 2010). Према нашим сазнањима, само једна студија на животињама је истраживала условљавање апетита мерењем одговора на шок као зависну меру. Занимљиво је да су испитани пацови показали запањујуће слабљење ЦС+ (тј. светлости) повезано са испоруком раствора сахарозе. Нарочито, такво слабљење је нарушено код оних животиња са НАцц лезијама, али не и код оних са лезијама амигдале, што сугерише да НАцц игра специфичну улогу у изазивању апетитивних ЦР-а и у ублажавању одговора на препад (Коцх ет ал., КСНУМКС).
Садашња студија је, према нашем сазнању, прва која је ову парадигму кондиционирања апетита превела на људе користећи примарне појачиваче као што су УС, тј. слатко (чоколадни паметни®) или слана (мала слана переца) храна и модулација шока као мера ЦР. Очекивали смо да ће аппЦС+ покренути снажне ЦР-ове апетита што се огледа у пригушењу запрепаштења, побољшаном СЦР-у и позитивној оцени валентности у поређењу са другим стимулусима, тј. авЦС+ и ЦС–.
Материјал и метод
učesnici
Четрдесет два волонтера су прихватила учешће у студији и добила бодове за курс. Девет учесника је искључено из анализе, два због техничких проблема, три зато што су били кодирани као нереагујући (средња амплитуда запрепашћења < 5 μВ), а четири зато што нису показали довољно одговора запрепашћења по стању (минимум = 2; за детаље, погледајте Материјали и метод). На крају, у анализу су узета 33 учесника (16 мушкараца; средња старост: 22.09 година, SD: 2.84; распон: 18–29 година). Четири учесника нису били староседеоци Немци, а шест су били леворуки. Један учесник није био свестан асоцијација ЦС-УСс током експеримента (погледајте Процедуру), али смо одлучили да не искључимо овог учесника јер су његови одговори били нормални и нису утицали на резултате.
Materijali
Безусловни стимуланси (САД)
Коришћене су две врсте УС. Као аверзивни САД, применили смо благо болан електрични шок на недоминантну подлактицу учесника. Електрични удар је испоручен помоћу две електроде пречника 9 мм и размака од 30 цм. Електрични стимулус се састојао од импулсног стимулуса са фреквенцијом од 50 Хз и трајањем од 200 мс, који је генерисао струјни стимулатор (Дигитимер ДС7А, Дигитимер Лтд, Велвин Гарден Цити, УК, 400 В, максимално 9.99 мА). Интензитет електричног удара је одређен појединачно кроз поступак прага који је претходно описан (Андреатта ет ал., 2010). Укратко, учесници су прошли две серије узлазних и опадајућих интензитета у интервалима од 0.5 мА. Морали су да оцене сваки стимулус на визуелној скали у распону од 0 (не осећају се уопште) до 10 (заиста интензиван бол) са 4 као сидром за праг (само приметан бол). Средњи интензитет електричног стимулуса био је 2.12 мА (SD = 0.56) и оцењено је као болно (M = КСНУМКС, SD = 1.73). Апетитивна САД састојала се или од чоколаде (Смартиес®) или мала слана переца. Избор привлачне САД зависио је од индивидуалних преференција учесника као што је наведено током прелиминарног интервјуа. Наиме, учесници су морали да пријаве да ли иначе једу слану или слатку храну током доручка. Такође су могли слободно да бирају да ли им је дража чоколада или слани перец током самог експеримента. На крају су 22 учесника изабрала чоколаду, а 11 мали слани перец.
Условљени стимуланси (ЦС)
Геометријски облици (плави квадрат, жути круг, зелени троугао, црвени шестоугао) дијагонале 8 цм представљени су као ЦС. Облици су представљени у средини црног компјутерског екрана током 8 с. Један облик (авЦС+) је увек био повезан са аверзивним УС (болни електрични удар), један облик (аппЦС+) је увек био повезан са привлачним УС (чоколада или слана переца), један облик (ЦС–) никада није био повезан ни са једним од аверзивних или апетитни УСс, а четврти облик (НОВ) је представљен у фази изумирања, али не током фазе аквизиције да би се осигурала његова неутралност.
Старле Пробе
Бели шум јачине 103 дБ у трајању од 50 мс коришћен је као сонда за узбуну. Акустични стимуланси су представљени бинаурално преко слушалица и јављали су се насумично 4–6 с након појаве облика.
upitnici
Пре и после експеримента, учесници су морали да попуне немачке верзије Државног инвентара анксиозности (СТАИ, Лаук ет ал., 1981) и распоред позитивних негативних ефеката (ПАНАС, Крохне ет ал., 1996). СТАИ је инвентар за процену особине учесника и/или анксиозности стања и састоји се од 20 ставки и за верзију и за верзију стања. Ниво анксиозности учесника пре (M = КСНУМКС, SD = 7.80) и после (M = КСНУМКС, SD = 9.16) експеримент се није значајно променио [t(32) = КСНУМКС, p = 0.117]. Резултати анксиозности особина у тренутном узорку су се кретали између 20 и 58 (M = КСНУМКС, SD = 8.98), што је упоредиво са објављеним нормалним опсегом одраслих (Лаук ет ал., 1981). ПАНАС (Крохне ет ал., 1996) је индекс за позитивно и негативно расположење. Појединци са високим резултатима на скали позитивних утицаја (ПАС) склони су емоцијама као што је ентузијазам, док су појединци са високим резултатима на скали негативних утицаја (НАС) склони емоцијама као што је узнемиреност. Свака ставка се састоји од придева, а учесници указују на скали од 1 (веома мало) до 5 (изузетно) у којој мери придев одражава њихова осећања у том тренутку. Нису нађене значајне разлике у негативном утицају учесника као резултат експеримента [почетак: M = КСНУМКС, SD = 2.29; крај: M = КСНУМКС, SD = КСНУМКС; t(32) = КСНУМКС, p = 0.130]. Некако су учесници значајно променили своје позитивно расположење од почетка (M = КСНУМКС, SD = 4.70) до краја (M = КСНУМКС, SD = 7.02) експеримента [t(31) = КСНУМКС, p = 0.004]. Ово смањење позитивног расположења учесника могло би бити повезано са непријатношћу парадигме (представљени су болни електрични шокови као и аверзивни бели шум).
Поступак
По доласку у лабораторију, учесници су прочитали и потписали образац информисаног пристанка који је одобрила етичка комисија Одељења за психологију Универзитета у Вирцбургу. Нису били обавештени о непредвиђеним ситуацијама између ЦС и САД. Након попуњавања упитника, електроде су причвршћене и процедура прага бола је спроведена као што је горе описано.
Током фаза навикавања, четири геометријска облика су представљена два пута са интервалом између покушаја (ИТИ) који варира између 18 и 25 с (средња вредност: 21.5 с). Током ове фазе нису испоручене никакве сонде за САД или шок.
Пре фазе аквизиције, седам напада беле буке је испоручено сваких 7–15 с како би се смањила почетна реактивност запрепашћења.
Следеће две фазе стицања били су идентични (Сл 1). Свака фаза стицања састојала се од 24 испитивања: 8 ЦС– испитивања, 8 авЦС+ испитивања и 8 аппЦС+ испитивања. ЦС секвенца је била псеудослучајна са јединим ограничењем да исти стимулус није могао бити представљен више од два пута заредом. Значајно је да је авЦС+ представљен заједно са муњом као симболом струјног удара, а болни САД је испоручен у офсету. Апликација ЦС+ је представљена у комплексу са сликом Смартиес или сланог переца и учесник је могао да изабере Смартиес или перец из тегле. ЦС– је представљен заједно са симболом забране и није испоручен ниједан УС. Током три од 8 ЦС презентација сваког типа, сонда за шок је испоручена између 4 и 6 с након почетка стимулуса. Три додатне сонде за шок су представљене током ИТИ да би се осигурала њихова непредвидљивост и да би се смањило навикавање на узнемиреност. ИТИ, који се састоји од црног екрана, варирао је између 18 и 25 с са средњом 21.5 с.
СЛИКА КСНУМКС
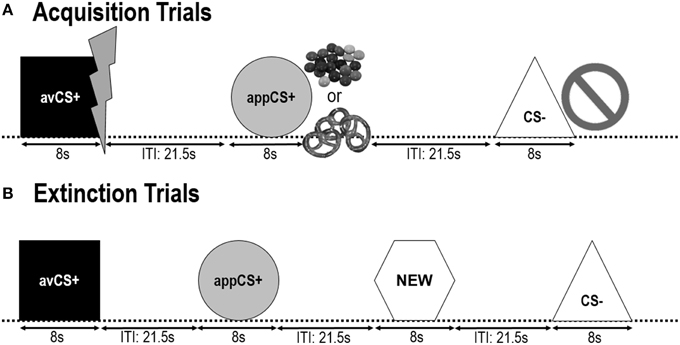
Слика 1. Пробе током две фазе аквизиције (А) и фазе изумирања (Б). Учесници су сазнали да један облик (авЦС+) предвиђа благи болни електрични удар, један облик (аппЦС+) предвиђа или комад чоколаде или малу слану перецу (према њиховој жељи), а трећи облик (ЦС–) није предвиђао никакав биолошки значајан догађај. Сваки облик је представљен заједно са сликом која приказује струјни удар, паметне/слане переце или ништа у зависности од повезаности са САД. Током фазе изумирања, три геометријска облика су поново представљена, али ниједан УС није испоручен. Поред тога, четврти геометријски облик (НОВ) је представљен као неутрална контрола.
Током фаза изумирања, учесници су поново видели три геометријска облика (тј. авЦС+, аппЦС+ и ЦС–) плус нови неутрални облик (НОВО). Ниједан УС није испоручен и облици нису представљени у комбинацији са сликама блица, чоколадних/сланих переца или забрана. Сваки стимулус је представљен осам пута псеудослучајним редоследом (тј. исти стимулус није представљен више од два пута заредом), стварајући 32 испитивања. Стимулуси сонде за шок су представљени током 4 од 8 презентација стимулуса сваког типа ЦС. Као иу фази аквизиције и навикавања, ИТИ-ови су варирали између 18 и 25 с, а 4 додатне сонде за шок су испоручене непредвидиво током ИТИ-а.
Након сваке фазе, учесници су оцењивали валенцију (пријатност) и узбуђење (узбудљиво) ЦС-а визуелно аналогним скалама (ВАС) у распону од 1 до 9. Скала равнотеже се кретала од „1” што указује на „веома непријатно” до „9” означавајући „веома пријатно“; скала узбуђења се кретала од „1“ што означава „смирено“ до „9“ што указује на „узбудљиво“. Поред тога, оцене непредвиђених околности су процењене након две фазе аквизиције и фазе изумирања. Учесници су видели геометријски облик 1 с, а затим су морали да назначе да ли је овај облик повезан са електричним ударом, са чоколадним/сланим перецем, ни са чим, или нису могли да направе било какву асоцијацију. Приметно је да су сви учесници (осим једног) били свесни непредвиђених околности након аквизиције 2.
Смањење података
Физиолошки одговори су забележени помоћу В-Амп 16 појачала и софтвера Висион Рецордер В-Амп Едитион (верзија 1.03.0004, БраинПродуцтс Инц., Минхен, Немачка). Примењена је брзина узорковања од 1000 Хз и 50 Хз зарезни филтер. Офлајн анализе су спроведене помоћу Браин Висион Анализер-а (верзија 2.0; БраинПродуцтс Инц., Минхен, Немачка).
Страшни одговор
Реакција на узнемиреност је мерена помоћу електромиографије (ЕМГ) са леве стране орбицуларис оцули мишића са две 5 мм Аг/АгЦл електроде. У складу са смерницама (Блументхал ет ал., 2005), једна електрода је постављена испод зенице, а друга 1 цм бочно. Уземљена и референтна електрода постављене су на десни и леви мастоид, респективно. Пре причвршћивања електрода, кожа је лагано истргнута и очишћена алкохолом како би импеданса била испод 10 кΩ. Електромиографски сигнал је ван мреже филтриран филтером ниске границе од 28 Хз и филтером високе границе од 500 Хз. Затим је ЕМГ сигнал исправљен и примењен је покретни просек од 50 мс. Користили смо 50 мс пре почетка сонде за узбуну као основну линију (Гриллон ет ал., КСНУМКС). Одговори на сонде за шок су оцењивани ручно, а испитивања са прекомерним помацима основне линије (±5 μВ) или артефакти покрета су искључени из даље анализе. Реакције запрепашћења ниже од 5 μВ су кодиране као нула и узете у обзир за израчунавање магнитуде запрепашћења (Блументхал ет ал., 2005). Свеукупно, 10.4% испитивања је одбијено, а најмање 2 од 3 одговора запрепашћења у фазама аквизиције и 4 од 8 одговора запрепаштења у фази изумирања за свако стање су била потребна да би се учесник задржао у групи анализа. Из тог разлога, четири учесника су искључена. Амплитуда пика је дефинисана као максимални пик у односу на основну линију током временског прозора од 20–120 мс након почетка сонде за узбуну. Сирови подаци су затим нормализовани унутар субјеката користећи z-сцоре па Т-сцоре у циљу смањења утицаја индивидуалне варијабилности и бољег откривања психолошких процеса. Т-резултати су упросечени за свако стање (авЦС+, аппЦС+, ЦС–, НЕВ и ИТИ). Да би се истражила потенцирање пренеражености или пригушивање запрепаштења, резултати за ИТИ реакције на препад су одузети од одговора на препад за свако стање.
Одзив проводљивости коже (СЦР)
Одзив проводљивости коже (СЦР) је забележен коришћењем две 5 мм Аг/АгЦл електроде постављене на длан недоминантне руке. Галвански одговор је био ван мреже филтриран помоћу филтера високе границе од 1 Хз. СЦР је дефинисан као разлика (у μС) између почетка одговора (1-3 с након почетка стимулуса) и врхунца одговора (Транел и Дамасио, 1994; Делгадо ет ал., КСНУМКС). У анализи СЦР-а нису узета у обзир испитивања која су садржавала сонде за шок. Одговори испод 0.02 μС су кодирани као нула. Још пет учесника је искључено из СЦР анализе јер су имали средњи СЦР мањи од 0.02 μС. Необрађени подаци о проводљивости коже трансформисани су квадратним кореном да би се нормализовала дистрибуција, а резултати су упросечени за свако стање посебно за две фазе аквизиције (авЦС+, аппЦС+, ЦС–) и фазу изумирања (авЦС+, аппЦС+, ЦС– и НЕВ ).
Статистичка анализа
Сви подаци су анализирани помоћу СПСС за Виндовс (верзија 20.0, СПСС Инц.). За физиолошке одговоре, израчунате су одвојене мултиваријантне анализе варијансе (АНОВА) за две фазе аквизиције и фазу изумирања. АНОВА за фазе аквизиције је имала стимуланс (авЦС+, аппЦС+, ЦС–) и фазу (Ацкуиситион 1, Ацкуиситион 2) као факторе унутар испитаника. АНОВА за фазу изумирања имала је само стимуланс (авЦС+, аппЦС+, ЦС–, НЕВ) као фактор унутар испитаника. Оцене валенције, узбуђења и контингентности су анализиране са одвојеним АНОВА-ама које садрже факторе стимуланса унутар субјекта (авЦС+, аппЦС+, ЦС– и НЕВ) и фазу. Овај фактор је имао четири нивоа за оцене валенце и узбуђења (Т1: након фазе навикавања, Т2: након прве фазе стицања, Т3: након друге фазе аквизиције, Т4: након фазе изумирања), али три нивоа за оцене непредвиђених околности (Т1: након прва фаза аквизиције, Т2: након друге фазе аквизиције, Т3: након фазе изумирања).
Ниво алфа (α) је постављен на 0.05 за све анализе. Величина ефекта је приказана као делимични η2.
Резултати
Оцене валенције и узбуђења за сваку фазу су приказане на слици 2; реакције на запрепашћење и СЦР-ови су приказани на слици 3.
СЛИКА КСНУМКС
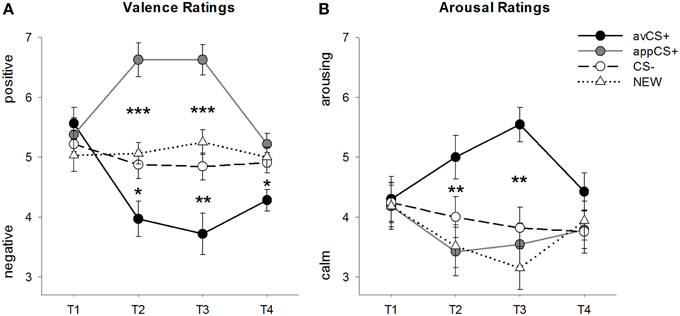
Слика 2. Оцене за валентност (А) и узбуђење (Б). Линије (са стандардним грешкама) приказују оцене након фазе навикавања (Т1), аквизиције 1 (Т2), аквизиције 2 (Т3) и фазе изумирања (Т4). Аверзивни ЦС+ (црна пуна линија) је стекао негативну валенцију и високо узбуђење након две фазе аквизиције у поређењу са ЦС– (црна испрекидана линија) и НЕВ (црна тачкаста линија). Важно је да је привлачни ЦС+ (сива пуна линија) добио позитивну валенцу у поређењу са ЦС– и НОВИМ. *p <0.05, **p > 0.01, ***p <0.001.
СЛИКА КСНУМКС
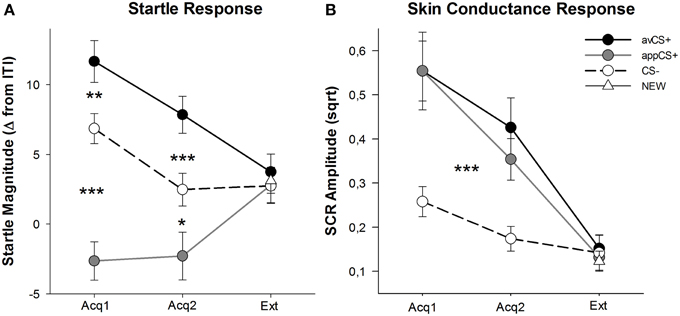
Слика 3. Реакције на узнемиреност (А) и проводљивост коже (Б) (са стандардним грешкама) током прве фазе аквизиције (Ацк1), друге фазе аквизиције (Ацк2) и фазе изумирања (Ект). Реакције запрепашћења су значајно потенциране на аверзивни ЦС+ (црна пуна линија) и значајно ослабљене на привлачни ЦС+ (сива пуна линија) у поређењу са ЦС– (црна испрекидана линија) током фаза аквизиције. СЦР је био значајно већи код авЦС+ и аппЦС+ у поређењу са ЦС–. Током фазе изумирања нису откривене разлике. *p <0.05, **p > 0.01, ***p <0.001.
Оцене
АНОВА за валентне оцене током аквизиције откривени су значајни главни ефекти стимулуса [F(КСНУМКС, КСНУМКС) = 17.26, ГГ-ε = 0.801, p < 0.001, парцијални η2 = 0.358] и фаза [F(КСНУМКС, КСНУМКС) = 3.30, ГГ-ε = 0.731, p = КСНУМКС, парцијално η2 = 0.096] као и значајна интеракција између стимулуса и фазе [F(КСНУМКС, КСНУМКС) = 9.54, ГГ-ε = 0.463, p < 0.001, парцијални η2 = 0.235]. Пратити t-тестови показују да су валенце четири геометријска облика на почетку експеримента биле идентичне (pс > 0.19), док је и након аквизиције 1 и аквизиције 2 авЦС+ оцењен као посебно негативан, а аппЦС+ као посебно позитиван. Конкретно, авЦС+ је оцењен као негативно валентнији у поређењу са ЦС– [Ацк1: t(31) = КСНУМКС, p = 0.026; Ацк2: t(31) = КСНУМКС, p = 0.004], НОВИ [Ацк1: t(31) = КСНУМКС, p = 0.011; Ацк2: t(31) = КСНУМКС, p < 0.001], а аппЦС+ [Ацк1: t(31) = КСНУМКС, p < 0.001; Ацк2: t(31) = КСНУМКС, p < 0.001]. АппЦС+ је оцењен као значајно позитивнији од ЦС– [Ацк1: t(31) = КСНУМКС, p < 0.001; Ацк2: t(31) = КСНУМКС, p < 0.001] и НОВИ [Ацк1: t(31) = КСНУМКС, p < 0.001; Ацк2: t(31) = КСНУМКС, p < 0.001]. Разлике између ЦС– и НЕВ никада нису биле значајне (pс > 0.18).
Важно је да нису нађене никакве значајне разлике када се упореде оцене валенције за аппЦС+ повезану са чоколадом у односу на слану перецу [Ацк1: t(30) = КСНУМКС, p = 0.477; Ацк2: t(30) = КСНУМКС, p = КСНУМКС].
Након фазе изумирања, авЦС+ је и даље оцењен као негативнији у поређењу са ЦС– [t(31) = КСНУМКС, p = 0.023], НОВИ [t(31) = КСНУМКС, p = 0.002], а аппЦС+ [t(31) = КСНУМКС, p = 0.002]. Напротив, валенција аппЦС+ се више није разликовала од ЦС– [t(31) = КСНУМКС, p = 0.086] или НОВИ [t(31) = КСНУМКС, p = КСНУМКС].
АНОВА за оцене узбуђења током аквизиције откривен је значајан главни ефекат стимулуса [F(КСНУМКС, КСНУМКС) = 7.07, ГГ-ε = 0.737, p = КСНУМКС, парцијално η2 = 0.181], али не фазе [F(КСНУМКС, КСНУМКС) = 1.27, ГГ-ε = 0.805, p = КСНУМКС, парцијално η2 = 0.038], и значајна интеракција између стимулуса и фазе [F(КСНУМКС, КСНУМКС) = 4.53, ГГ-ε = 0.582, p = КСНУМКС, парцијално η2 = 0.124]. Пратити t-тестови нису показали значајне разлике међу стимулусима у погледу њиховог почетног узбуђења (pс > 0.74). Међутим, након две фазе аквизиције, авЦС+ је оцењен као узбудљивији од НОВЕ [Ацк1: t(32) = КСНУМКС, p = 0.005; Ацк2: t(32) = КСНУМКС, p < 0.001] и аппЦС+ [Ацк1: t(32) = КСНУМКС, p = 0.013; Ацк2: t(32) = КСНУМКС, p < 0.001], и незнатно узбудљивије од ЦС – након прве фазе аквизиције [t(32) = КСНУМКС, p = 0.058], али значајно више узбуђује након друге фазе аквизиције [t(32) = КСНУМКС, p = 0.001]. За разлику од валентних оцена, узбуђење аппЦС+ се није разликовало од узбуђења ЦС– и НЕВ (ps > 0.13) након фаза аквизиције.
Слично као и са валентним оценама, ЦС– и НЕВ се нису разликовали по свом узбуђењу (pс > 0.07).
Као што је био случај са валентношћу, нису откривене разлике за аппЦС+ повезан са чоколадом и аппЦС+ повезан са сланим перецем [Ацк1: t(31) = КСНУМКС, p = 0.797; Ацк2: t(31) = КСНУМКС, p = КСНУМКС].
Након фазе изумирања, нису откривене значајне разлике у оценама узбуђења (pс > 0.08).
За додатне пост-хоц т-тестови који упоређују оцене након фазе навикавања, прве и друге фазе стицања и фазе изумирања, видети Додатни материјали.
Страшни одговор
АНОВА за фазе аквизиције вратила је главни ефекат стимулуса [F(КСНУМКС, КСНУМКС) = 49.92, ГГ-ε = 0.964, p < 0.001, парцијални η2 = 0.609], али не и фаза [F(КСНУМКС, КСНУМКС) = КСНУМКС, p = КСНУМКС, парцијално η2 = 0.090], и значајна интеракција Стимулус × Фаза [F(КСНУМКС, КСНУМКС) = 3.37, ГГ-ε = 0.875, p = КСНУМКС, парцијално η2 = 0.095]. Пратити t-тестови су открили значајну потенцирање запрепаштења за авЦС+ у поређењу са ЦС– током оба прва [t(32) = КСНУМКС, p = 0.003] и други [t(32) = КСНУМКС, p < 0.001] фазе аквизиције. Запањујући одговори на авЦС+ такође су били значајно потенцирани у поређењу са онима на аппЦС+, поново након оба набавка 1 [t(32) = КСНУМКС, p < 0.001] и аквизиција 2 [t(32) = КСНУМКС, p < 0.001]. Важно је да је величина запрепашћења аппЦС+ била значајно ослабљена у поређењу са ЦС– током обе набавке 1 [t(32) = КСНУМКС, p < 0.001] и аквизиција 2 [t(32) = КСНУМКС, p = 0.007]. Поново иу складу са оценама, нису откривени различити одговори запрепашћења за аппЦС+ када су повезани са чоколадом или сланим перецем [Ацк1: t(31) = КСНУМКС, p = 0.309; Ацк2: t(31) = КСНУМКС, p = 0.947]. Током фазе изумирања није пронађен значајан ефекат [F(КСНУМКС, КСНУМКС) = 0.26, ГГ-ε = 0.906, p = КСНУМКС, парцијално η2 = КСНУМКС].
Одзив проводљивости коже (СЦР)
Из АНОВА за СЦР током две фазе аквизиције, главни ефекти стимуланс [F(КСНУМКС, КСНУМКС) = 18.04, ГГ-ε = 0.908, p < 0.001, парцијални η2 = 0.401] и фаза [F(КСНУМКС, КСНУМКС) = КСНУМКС, p < 0.001, парцијални η2 = 0.436] показало се значајно, али не и њихова интеракција [F(КСНУМКС, КСНУМКС) = 0.68, ГГ-ε = 0.637, p = КСНУМКС, парцијално η2 = КСНУМКС]. Пост-хоц т-тестови су показали значајно већи СЦР у односу на авЦС+ [t(27) = КСНУМКС, p < 0.001] и на аппЦС+ [t(27) = КСНУМКС, p < 0.001] у поређењу са ЦС–, док су учесници показали упоредив СЦР са авЦС+ и аппЦС+ [t(27) = КСНУМКС, p = 0.527]. Нарочито није пронађена никаква разлика у СЦР-овима за чоколадну аппЦС+ и за слану перецу аппЦС+ [Ацк1: t(26) = КСНУМКС, p = 0.120; Ацк2: t(26) = КСНУМКС, p = 0.210]. Као што је било тачно за оцене и одговор запрепашћења, нису пронађени значајни ефекти за фазу изумирања [F(КСНУМКС, КСНУМКС) = 0.28, ГГ-ε = 0.634, p = КСНУМКС, парцијално η2 = КСНУМКС].
Дискусија
Циљ ове студије је био да се резултати животиња преведу на људе коришћењем класичне парадигме кондиционирања апетита са примарним појачалом као безусловним стимулусом (УС). Да би то урадили, учесници су долазили у лабораторију рано ујутру без доручка како би се уверили да су гладни, а према жељи, комадићи чоколаде или слане переце су коришћени као укусни УС-ови. Током фазе аквизиције, један геометријски облик (авЦС+) је постао повезан са благо болним електричним ударом (аверзивни УС), други облик (аппЦС+) са апетитним УС, а трећи облик (ЦС–) ни са привлачним УС нити са аверсиве УС. Резултати указују на успешно аверзивно и апетитивно условљавање на експлицитном вербалном нивоу (тј. оцене), на имплицитном нивоу понашања (тј. реаговање на препад) и на физиолошком нивоу (тј. СЦР). Конкретно, авЦС+ у поређењу са ЦС– изазвао је више негативних оцена валентности, веће оцене узбуђења, потенцирање запрепашћења и већи СЦР. Оно што је најважније, аппЦС+ у поређењу са ЦС– покренуо је више позитивних оцена валентности, слабљење запрепашћења и већи СЦР. Наши налази о аверзивном условљавању били су у складу са очекивањима, пошто су претходне студије откриле да је претња која предвиђа стимуланс (авЦС+) оцењена као аверзивна, изазива веће реакције страха и повећава физиолошко узбуђење (Фендт и Фанселов, 1999; Хам и Вејке, 2005; Андреатта ет ал., 2010, 2013). Наши резултати о кондиционирању апетита су такође у складу са претходним студијама на људима и животињама које показују да је награда за предвиђање стимулуса (аппЦС+) оцењена као позитивна, инхибира реакције на страх и повећава физиолошко узбуђење (Коцх ет ал., КСНУМКС; Готтфриед ет ал., 2002; Кумар ет ал., КСНУМКС; Клуцкен ет ал., КСНУМКС, 2013; Аустин и Дука, КСНУМКС; Превост ет ал., КСНУМКС). Према нашим сазнањима, ово је прва студија која демонстрира условљено слабљење шока код људи на стимулус који предвиђа примарну награду. Важно је да смо успели да пренесемо и потврдимо резултате студије на животињама (Коцх ет ал., КСНУМКС). Ова студија на животињама је показала да слабљење запрепашћења код пацова зависи од пројекција од НАцц (део вентралног стриатума) до ПнЦ. Стога би запрепаштење у нашој студији могло да имплицира активност НАцц, што је такође у складу са фМРИ налазима (Готтфриед ет ал., 2002; Кумар ет ал., КСНУМКС; Клуцкен ет ал., КСНУМКС, 2013; Делгадо ет ал., КСНУМКС; Леви и Глимчер, 2011). Дакле, закључујемо да је наша парадигма условљавања апетита била успешна, као што су и експлицитна (оцене) и имплицитна (слабљење запрепашћења) позитивна валентност указивале.
Поред овог новог, али прилично предвидљивог налаза, заслужују да се спомену још два занимљива резултата. Прво, вербални и физиолошки одговори узбуђења на аппЦС+ су се раздвојили. Друго, открили смо брже изумирање апетитних ЦР у поређењу са аверзивним ЦР.
СЦР указује на симпатичку активацију, која се повећава и на стимулус повезан са наградом (аппЦС+) и стимулус повезан са претњом (авЦС+). Овај резултат је у складу са претходном студијом кондиционирања, у којој су еротске слике коришћене као привлачне САД (Клуцкен ет ал., КСНУМКС), и студија која открива упоредиву СЦР са знаком који предвиђа новац и знаком који предвиђа аверзивну буку (Аустин и Дука, КСНУМКС). Посебно, СЦР је оријентациони одговор који се односи на активацију симпатичког система. Предложено је да овај одговор одражава припрему бихејвиоралне реакције на мотивационо истакнуте догађаје (Бредли, 2009). На основу овога, мислимо да су и стимуланси повезани са претњом и наградом изазвали припремни одговор за узастопне бихејвиоралне одговоре. Другим речима, сигнали хране и бола иницирали су припрему за приступ, односно избегавање понашања. За разлику од високог физиолошког узбуђења, вербални одговори су указивали на ниско узбуђење на стимулус повезан са наградом. Могуће је да на вербално узбуђење може више утицати побудна природа САД, а не физиолошка активација по себи. У ствари, мислимо да су аппЦС+ у односу на авЦС+ оцењени као ниско наспрам веома узбуђујућих јер су били повезани са ниским и веома узбуђујућим САД, респективно. Нажалост, нисмо прикупили оцене узбуђења за САД, и стога ће на будућим студијама бити да експлицитно тестирају ову хипотезу.
Током фазе изумирања, ниједна УС није испоручена. Ово је можда изазвало ново инхибиторно учење звано изумирање (за преглед погледајте Милад и Квирк, 2012), при чему се и аверзивни и апетитивни ЦР смањују као последица. Очигледно, на бихејвиоралним (тј. реакција на препад) и на физиолошким (тј. СЦР) нивоима, више се нису могле детектовати диференцијалне реакције на авЦС+, аппЦС+ и ЦС–. Слично томе, експлицитно узбуђење условљених стимулуса процењених након фазе изумирања изједначило се на ниском нивоу, што сугерише успешно учење о изумирању. Међутим, авЦС+ је и даље оцењен знатно негативнијим од ЦС–, док се валенција аппЦС+ више није разликовала од ЦС– валенце. Спорије изумирање експлицитног аверзивног одговора могло би бити последица еволуционог конзервативизма, што значи да је сигнале претње посебно тешко заборавити јер нереаговање на сигнал претње може бити опасно по живот.
Једно питање и даље остаје: зашто се онда реакција запрепашћења (тј. имплицитна валенција) потпуно угасила током фазе изумирања? Прво, ови одговори су израчунати током читаве фазе. Стога је могуће да би се дискриминативни ЦР и даље могли открити током првих испитивања фазе изумирања. На истраживачки начин, пратили смо ову хипотезу и погледали и реакције на запрепашћење и СЦР током фазе изумирања (погледајте Додатни материјал). Иако нисмо пронашли значајне разлике, приметили смо нешто већу магнитуду запрепашћења за авЦС+ у поређењу са ЦС– и новим контролним стимулусом. Такође смо приметили нешто више запањујуће слабљење аппЦС+ у поређењу са ЦС– и новим контролним стимулусом на самом почетку фазе изумирања, који је, међутим, нестао у року од неколико покушаја. СЦР за аппЦС+ је опао већ након 2. покушаја изумирања, док је СЦР за авЦС+ остао већи за скоро све изумирање у поређењу са СЦР за НЕВ. Иако ове резултате треба тумачити са великим опрезом, чинило се да су запрепашћени одговори паралелни са валентним оценама. Штавише, и запрепашћени одговори и СЦР-ови даље подржавају идеју еволуционог конзервативизма према претњи стимулансима.
Коначно, морамо признати нека ограничења ове студије. Прво, због техничког проблема нисмо могли да пријавимо оцене пријатности (и узбуђења) за чоколаду и слани перец. Међутим, привлачни ЦР сугеришу да су учесници заиста доживели две САД као привлачне. Друго, трајање аверзивног и привлачног САД се увелико разликовало. Дакле, болни електрични шок је испоручен на офсету авЦС+ и трајао је тачно 200 мс, док су чоколада и слана переца представљени учесницима око 2 с након почетка аппЦС+, а трајање је било недефинисано јер је зависило од тога колико брзо појединац их је појео. Избор да се на овај начин испоруче привлачни САД заснован је на претходној студији на људима (Превост ет ал., КСНУМКС). Међутим, методолошки би било елегантније испоручити привлачни САД који су упоредивији са аверзивним САД, као што је гутљај сока или ђумбирово пиво уз аппЦС+ оффсет. Да бисмо надокнадили ову велику разлику у перцепцији, представили смо геометријске облике у комбинацији са визуелним стимулусом који симболизује САД. Треће, не можемо дефинитивно искључити могућност да су брзо угашени физиолошки одговори у фази изумирања последица методолошког аспекта. У ствари, визуелни стимуланси у овој фази нису били представљени заједно са симболом САД као што је то био случај у фазама аквизиције. Могуће је да је само презентација ЦС-а утицала на ЦР и из тог разлога нису биле видљиве значајне разлике током првих испитивања изумирања. Међутим, наши брзи угашени одговори су у складу са угашеним одговорима у претходној студији у којој је ЦС+ (лице) представљен као спој са САД (врисак) током аквизиције, али не и током изумирања (Лиссек ет ал., 2008).
У закључку, пронашли смо успешне аверзивне и апетитивне условљене одговоре на стимулус повезан са претњом и на стимулус повезан са наградом, респективно. Занимљиво је да су експлицитни (оцене) и имплицитни (рефлекс пренеражености) нивои одговора функционисали на синергијски начин, тако што је авЦС+ пријављен као негативна и индукована потенцираност запрепашћења, а аппЦС+ је пријављен као позитивно и индуковано пригушење запрепашћења. Штавише, експлицитно (оцене) и физиолошко (СЦР) узбуђење аппЦС+ се раздвојило, одражавајући два различита процеса.
Изјава о сукобу интереса
Аутори изјављују да је истраживање проведено у одсуству било каквих комерцијалних или финансијских односа који би се могли тумачити као потенцијални сукоб интереса.
priznanja
Овај рад је подржао Центар за колаборативно истраживање „Страх, анксиозност и анксиозни поремећаји“, СФБ-ТРР 58, пројекат Б1.
Додатни материјал
Додатни материјал за овај чланак можете пронаћи на адреси: http://journal.frontiersin.org/article/10.3389/fnbeh.2015.00128/abstract
Референце
Андреатта, М., Мухлбергер, А., Глотзбацх-Сцхоон, Е., анд Паули, П. (2013). Предвидљивост бола преокреће валентне оцене стимулуса повезаног са олакшањем. Фронт. Сист. Неуросци. 7:53. дои: 10.3389/фнсис.2013.00053
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Андреатта, М., Мухлбергер, А., Иарали, А., Гербер, Б., анд Паули, П. (2010). Расцеп између имплицитне и експлицитне условљене валенције након учења за ублажавање болова код људи. Проц. Биол. Сци. 277, 2411–2416. дои: 10.1098/рспб.2010.0103
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Аустин, АЈ, и Дука, Т. (2010). Механизми пажње за апетитивне и аверзивне исходе у павловском условљавању. Бехав. Браин Рес. КСНУМКС, КСНУМКС – КСНУМКС. дои: КСНУМКС / ј.ббр.КСНУМКС
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Блументхал, ТД, Цутхберт, БН, Филион, ДЛ, Хацклеи, С., Липп, ОВ, и ван Боктел, А. (2005). Извештај комитета: смернице за електромиографске студије трептаја очију код људи. Псицхопхисиологи КСНУМКС, КСНУМКС – КСНУМКС. дои: КСНУМКС / ј.КСНУМКС-КСНУМКС.к
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Боутон, МЕ, и Пецк, ЦА (1989). Контекст ефекти на условљавање, изумирање и поновно успостављање у припреми за кондиционирање апетита. Аним. Научите. Бехав. КСНУМКС, КСНУМКС – КСНУМКС. дои: КСНУМКС / БФКСНУМКС
ЦроссРеф Фулл Тект | Гоогле Сцхолар
Бредли, ММ (2009). Природна селективна пажња: оријентација и емоција. Псицхопхисиологи КСНУМКС, КСНУМКС – КСНУМКС. дои: КСНУМКС / ј.КСНУМКС-КСНУМКС.к
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Цларк, ЈЈ, Холлон, НГ и Пхиллипс, ПЕМ (2012). Павловски системи вредновања у учењу и одлучивању. Цурр. Опин. Неуробиол. 22, 1054–1061. дои: 10.1016/ј.цонб.2012.06.004
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Делгадо, МР, Јоу, РЛ, и Пхелпс, ЕА (2011). Неурални системи који леже у основи аверзивног условљавања код људи са примарним и секундарним појачањима. Фронт. Неуросци. КСНУМКС: КСНУМКС. дои: КСНУМКС / фнинс.КСНУМКС
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Дицкинсон, А., анд Баллеине, Б. (1994). Мотивациона контрола циљане акције. Аним. Научите. Бехав. КСНУМКС, КСНУМКС – КСНУМКС. дои: КСНУМКС / БФКСНУМКС
ЦроссРеф Фулл Тект | Гоогле Сцхолар
Фендт, М., и Фанселов, МС (1999). Неуроанатомска и неурохемијска основа условљеног страха. Неуросци. Биобехав. Рев. 23, 743–760. дои: 10.1016/ј.ијпсицхо.2012.09.006
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Готтфриед, ЈА, О'Дохерти, Ј., и Долан, РЈ (2002). Апетитивно и аверзивно учење мириса код људи проучавано је коришћењем функционалне магнетне резонанце повезане са догађајима. Ј. Неуросци. 22, 10829–10837. Доступно на мрежи на: http://www.jneurosci.org/content/22/24/10829.full
Гриллон, Ц., Баас, ЈМ, Цорнвелл, Б., анд Јохнсон, Л. (2006). Условљавање контекста и избегавање понашања у окружењу виртуелне реалности: ефекат предвидљивости. Биол. Псицхиатри КСНУМКС, КСНУМКС – КСНУМКС. дои: КСНУМКС / ј.биопсицх.КСНУМКС
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Хамм, АО, и Веике, АИ (2005). Неуропсихологија учења страха и регулације страха. Инт. Ј. Псицхопхисиол. 57, 5–14. дои: 10.1016/ј.ијпсицхо.2005.01.006
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Клуцкен, Т., Сцхвецкендиек, Ј., Мерз, ЦЈ, Табберт, К., Валтер, Б., Кагерер, С., ет ал. (2009). Неуралне активације стицања условљеног сексуалног узбуђења: ефекти свести о непредвиђеним околностима и секс. Ј. Сек. Мед. КСНУМКС, КСНУМКС – КСНУМКС. дои: КСНУМКС / ј.КСНУМКС-КСНУМКС.к
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Клуцкен, Т., Вехрум, С., Сцхвецкендиек, Ј., Мерз, ЦЈ, Хенниг, Ј., Ваитл, Д., ет ал. (КСНУМКС). Полиморфизам КСНУМКС-ХТТЛПР је повезан са промењеним хемодинамским одговорима током кондиционирања. Зујати. Браин Мапп. 34, 2549–2560. дои: 10.1002/хбм.22085
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Коцх, М. (1999). Неуробиологија запрепашћења. Прог. Неуробиол. 59, 107–128. doi: 10.1016/S0301-0082(98)00098-7
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Коцх, М., Сцхмид, А., анд Сцхнитзлер, Х.-У. (1996). Задовољство-слабљење стрепње је нарушено лезијама нуцлеус аццумбенса. Неурорепорт 7, 1442–1446. doi: 10.1097/00001756-199605310-00024
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Крохне, ХВ, Еглофф, Б., Кохманн, Ц.-В., и Таусцх, А. (1996). Унтерсуцхунген мит еинер деутсцхен версион дер „Распоред позитивних и негативних ефеката“ (ПАНАС). Диагностица КСНУМКС, КСНУМКС-КСНУМКС.
Кумар, П., Ваитер, Г., Ахеарн, Т., Милдерс, М., Реид, И., анд Стееле, ЈД (2008). Абнормална временска разлика сигнали награђивања за учење у великој депресији. Мозак 131, 2084–2093. дои: 10.1093/браин/авн136
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Лаук, Л., Гланзманн, П., Сцхаффнер, П., анд Спиелбергер, ЦД (1981). Дас Стате-Траит Ангстинвентар. Вајнхајм: Белцов тест.
Леви, ДЈ, и Глимцхер, ПВ (2011). Поређење јабуке и поморанџе: коришћење субјективног представљања вредности у мозгу специфичне за награду и генералну награду. Ј. Неуросци. КСНУМКС, КСНУМКС – КСНУМКС. дои: КСНУМКС / ЈНЕУРОСЦИ.КСНУМКС-КСНУМКС
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Лиссек, С., Левенсон, Ј., Биггс, АЛ, Јохнсон, ЛЛ, Амели, Р., Пине, ДС, ет ал. (2008). Повишено условљавање страха друштвено релевантним безусловним стимулансима код социјалног анксиозног поремећаја. Сам. Ј. Псицхиатри КСНУМКС, КСНУМКС – КСНУМКС. дои: КСНУМКС / аппи.ајп.КСНУМКС
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Мартин-Соелцх, Ц., Линтхицум, Ј., и Ернст, М. (2007). Условљавање апетита: неуронске основе и импликације за психопатологију. Неуросци. Биобехав. Рев. КСНУМКС, КСНУМКС – КСНУМКС. дои: КСНУМКС / ј.неубиорев.КСНУМКС
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
МцДанналд, МА, Луцантонио, Ф., Бурке, КА, Нив, И., анд Сцхоенбаум, Г. (2011). Вентрални стријатум и орбитофронтални кортекс су потребни за учење засновано на моделу, али не и без модела. Ј. Неуросци. КСНУМКС, КСНУМКС – КСНУМКС. дои: КСНУМКС / ЈНЕУРОСЦИ.КСНУМКС-КСНУМКС
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Милад, МР, и Куирк, Г. (2012). Страх од изумирања као модел за транслациону неуронауку: десет година напретка. Анну. Рев. Псицхол. КСНУМКС, КСНУМКС – КСНУМКС. дои: КСНУМКС / аннурев.псицх.КСНУМКС
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Павлов, ИП (1927). Условљени рефлекси: истраживање физиолошке активности мождане коре. Лондон: Окфорд Университи Пресс.
Превост, Ц., Лиљехолм, М., Тисзка, ЈМ, и О'Дохерти, ЈП (2012). Неурални корелати специфичног и општег павловског преноса на инструментални у оквиру људских амигдаларних подрегиона: фМРИ студија високе резолуције. Ј. Неуросци. 32, 8383–8390. дои: 10.1523/јнеуросци.6237-11.2012
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Транел, Д., и Дамасио, Х. (1994). Неуроанатомски корелати одговора електродермалне проводљивости коже. Псицхопхисиологи 31, 427–438. doi: 10.1111/j.1469-8986.1994.tb01046.x
ПубМед Абстрацт | ЦроссРеф Фулл Тект | Гоогле Сцхолар
Кључне речи: класично условљавање, награда, казна, рефлекс запрепашћења, реакција проводљивости коже
Цитирање: Андреатта М и Паули П (2015) Аппетитиве вс. Аверсиве цондитионинг ин хуман. Фронт. Бехав. Неуросци. 9: КСНУМКС. дои: КСНУМКС / фнбех.КСНУМКС
Примљено: 11. фебруара 2015. године; Прихваћено: 05. маја 2015.;
Објављено: 19. маја 2015.
Едитед би:
Ниелс Бирбаумер, Университи оф Туебинген, Германи
Рецензија:
Тим Клуцкен, Јустус Лиебиг Универзитет Гисен, Немачка
Алессандро Ангрилли, Универзитет у Падови, Италија
Цопиригхт © 2015 Андреатта и Паули. Ово је чланак отвореног приступа који се дистрибуира под условима Цреативе Цоммонс лиценца за признање (ЦЦ БИ). Употреба, дистрибуција или репродукција на другим форумима је дозвољена, под условом да су изворни аутор (и) или давалац лиценце признати и да је изворна публикација у овом часопису наведена, у складу са прихваћеном академском праксом. Није дозвољена употреба, дистрибуција или репродукција која није у складу са овим условима.
*Кореспонденција: Марта Андреатта, Одсек за психологију (биолошка психологија, клиничка психологија и психотерапија), Универзитет у Вирцбургу, Марцусстраßе 9-11, Д-97070 Вурзбург, Немачка, [емаил заштићен]