- 1સેન્ટ્રો ઇન્ટરડિસિપ્લિનિયો ડી ન્યુરોસિએન્સિયા ડી વાલ્પેરેસો, ફેકલ્ટી ઓફ સાયન્સ, યુનિવર્સિડેડ દ વાલ્પરíસો, વાલપેરેસો, ચિલી
- 2ડિપાર્ટમેંટો ડિ ન્યુરોસિએન્સિયા, ફેક્લટadડ દ મેડિસિના, યુનિવર્સિડેડ દ ચિલી, સેન્ટિયાગો, ચિલી
- 3ન્યુક્લેઓ મિલેનીઓ NUMIND બાયોલોજી Neફ ન્યુરોપ્સાયchiટ્રિક ડિસઓર્ડર્સ, યુનિવર્સિડેડ દ વાલ્પરíસો, વાલપરેસો, ચિલી
- 4સેલ્યુલર અને મોલેક્યુલર બાયોલોજી ડિપાર્ટમેન્ટ, બાયોલોજિકલ સાયન્સ ફેકલ્ટી, પોન્ટિફિયા યુનિવર્સિડેડ કેટટાલીકા ડે ચિલી, સેન્ટિયાગો, ચિલી
- 5ન્યુરોસાયન્સના ફાર્મસી અને ઇન્ટરડિસિપ્પ્લિનરી સેન્ટર વિભાગ, રસાયણશાસ્ત્રની ફેકલ્ટી, પોન્ટીફિયા યુનિવર્સિડેડ કેટટાલીકા દ ચિલી, સેન્ટિયાગો, ચિલી
ગોલલક્ષી વર્તણૂકની તાકાત મિડબ્રેઇન ડોપામાઇન ન્યુરોન્સ દ્વારા નિયંત્રિત થાય છે. ડોપામિનર્જિક સર્કિટ્સની તકલીફ ડ્રગ વ્યસન અને બાધ્યતા મનોગ્રસ્તિ વિકારમાં જોવા મળે છે. અનિવાર્ય વર્તન એ એક સુવિધા છે જે બંને ડિસઓર્ડર વહેંચે છે, જે ડોપામાઇનના ન્યુરોટ્રાન્સિસને વધારે છે. મિડબ્રેઇન ડોપામાઇન ન્યુરોન્સની પ્રવૃત્તિ મુખ્યત્વે ડી 2 રીસેપ્ટર્સ (ડી 2 આર) દ્વારા ડોપામાઇનની હોમિયોસ્ટેટિક ક્રિયા દ્વારા નિયમન કરવામાં આવે છે જે ચેતાકોષોના ફાયરિંગ તેમજ ડોપામાઇન સંશ્લેષણ અને પ્રકાશનમાં ઘટાડો કરે છે. ડોપામાઇન ટ્રાન્સમિશન, અન્ય લોકોમાં, કપ્પા ioપિઓઇડ સિસ્ટમ જેવી વિજાતીય ન્યુરોટ્રાન્સમીટર સિસ્ટમ્સ દ્વારા પણ નિયંત્રિત થાય છે. કપ્પા ioપિઓઇડ સિસ્ટમ વિશેના આપણા હાલના મોટાભાગના જ્ andાન અને ડોપામાઇન ટ્રાન્સમિશન પર તેના પ્રભાવ મગજની રોગોના પરાકાષ્ઠાવાળા પ્રાણીના મ modelsડેલોથી આવે છે. 1988 માં, મગજનો માઇક્રોડાયલિસીસનો ઉપયોગ કરીને, તે બતાવવામાં આવ્યું હતું કે કપ્પા Opપિઓઇડ રીસેપ્ટર્સ (કેઓઆર) ની તીવ્ર સક્રિયકરણ સ્ટ્રાઇટમમાં ડોપામાઇનના સિનેપ્ટિક સ્તરમાં ઘટાડો કરે છે. કેઓઆરનો આ અવરોધક પ્રભાવ ડોપામાઇન પ્રકાશન પરના દુરૂપયોગની દવાઓના સરળ પ્રભાવનો વિરોધ કરે છે, જેના કારણે કોર એગોનિસ્ટ્સને અનિવાર્ય ડ્રગના સેવન માટે ફાર્માકોલોજીકલ ઉપચાર તરીકે ઉપયોગ કરવાની દરખાસ્ત તરફ દોરી જાય છે. આશ્ચર્યજનક રીતે, 30 વર્ષ પછી, કેઓઆર વિરોધીને તેના બદલે ડ્રગ વ્યસનની સારવાર માટે દરખાસ્ત કરવામાં આવી છે. આ વર્ષોમાં શું બન્યું હશે જેણે આ દાખલામાં ધરખમ પરિવર્તન પેદા કર્યું? એકત્રિત પુરાવા સૂચવે છે કે સિનોપ્ટીક ડોપામાઇનના સ્તરો પર કેઓઆરની અસર જટિલ છે, કેઓઆર સક્રિયકરણની આવર્તન અને ડોપામાઇન ન્યુરોન્સમાં આવતા અન્ય ઉત્તેજના સાથેના સમય તેમજ લિંગ અને જાતિના તફાવતો પર આધાર રાખીને. તેના તીવ્ર પ્રભાવથી વિપરીત, ક્રોનિક કેઓઆર સક્રિયકરણ ડોપામાઇન ન્યુરોટ્રાન્સમિશન અને ડોપામાઇન-મધ્યસ્થી વર્તણૂકોને સગવડ કરે છે. તીવ્ર વિરુદ્ધ ક્રોનિક કેઓઆર સક્રિયકરણ દ્વારા કરવામાં આવતી વિરોધી ક્રિયાઓ દુરુપયોગની દવાઓના સંપર્કમાં હોવા દરમિયાન, પ્રારંભિક અવ્યવસ્થિત અને વિલંબિત લાભદાયી અસર સાથે સંકળાયેલી છે. ડી 2 આરના વારંવાર સક્રિયકરણ દ્વારા પ્રેરિત અનિવાર્ય વર્તણૂકો પણ કેઓઆરના સતત સહ-સક્રિયકરણ દ્વારા સંભવિત છે, જે ડોપામાઇન અને સેન્સેટાઇઝ્ડ ડી 2 આરના સિનેપ્ટિક સ્તરમાં ઘટાડો સાથે સુસંગત છે. આમ, KOR ની સમય-આધારિત સક્રિયકરણ, ડોપામાઇનના સ્તર પર સીધી અસર પ્રેરિત વર્તણૂકોના ટ્યુનિંગને અસર કરે છે. આ સમીક્ષા અનિવાર્ય વર્તણૂકના ડોપામિનર્જિક સહસંબંધમાં કપ્પા ioપિઓઇડ સિસ્ટમના યોગદાનનું વિશ્લેષણ કરે છે.
પરિચય
અનિવાર્ય વર્તણૂકોમાં ડોપામિનેર્જિક સિસ્ટમ
અનિવાર્યતા, પ્રતિકૂળ પરિણામો હોવા છતાં, જાણીતા પરિણામ સાથે રીualક ક્રિયા ચલાવવા માટે સ્વયં-બંધ થવાની અશક્યતા છે (રોબિન્સ એટ અલ., 2012). અનિયમિત વર્તણૂક એ મનોચિકિત્સાના અન્ય રોગોમાં બાધ્યતા મનોગ્રસ્તિ વિકાર (OCD) અને માદક દ્રવ્યોના વ્યસનનું લક્ષણ છે. બાધ્યતા મનોગ્રસ્તિ સ્પેક્ટ્રમ ડિસઓર્ડર્સમાં ત્રાસકારક વર્તણૂક ખૂબ સામાન્ય છે, ચોક્કસ રૂટિનની સતત પુનરાવર્તન દ્વારા, રૂ steિચુસ્ત અથવા કર્મકાંડની રીતે (વિલિયમ્સ એટ અલ., 2013). સામાન્ય વર્તણૂકો (દા.ત., તપાસ, સફાઈ, હાથ ધોવા, વગેરે) ની વિશાળ શ્રેણી, ઓસીડી દર્દીઓમાં અનિવાર્ય બની શકે છે અને સામાન્ય રીતે, બાધ્યતા અને ત્રાસદાયક વિચારોની ચિંતા માટેના પ્રતિભાવમાં ઉદ્ભવે છે. તેવી જ રીતે, દુરૂપયોગની દવાઓ લેવી અને તેનું સેવન કરવાથી ડ્રગ વ્યસનીમાં અનિવાર્ય બની જાય છે OCD ની જેમ, અસ્વસ્થતા અનુભવી ડ્રગના નશો કરનારાઓમાં અનિવાર્ય ડ્રગના વપરાશને ઉત્તેજીત કરવામાં મુખ્ય ભૂમિકા નિભાવે છે. ઇનામ અને સજા પ્રક્રિયામાં સમાન ક્ષતિઓ બંને સ્થિતિમાં જોવા મળે છે (ફીગી એટ અલ., 2016), જેના કારણે કેટલાક લેખકો OCD ને વર્તણૂકીય વ્યસન તરીકે ચર્ચા કરવા લાગ્યા છે (હોલ્ડન, 2001).
અનિવાર્ય વર્તણૂક તરફ દોરી શકે તેવું એક સંભવિત પદ્ધતિ વ્યસનની પ્રોત્સાહન-સંવેદના સિદ્ધાંતમાં ઘડવામાં આવે છે જે એ છે કે ડ્રગ માટે એક એમ્પ્લીફાઇડ પ્રેરણા ("ઇચ્છતા") એમ્પ્લીફાઇડ આનંદદાયક (“પસંદ”) અસર વિકસિત કર્યા વિના વ્યસન દરમિયાન વિકસે છે.બેરીજ એટ અલ., 1989; બેરીજ અને રોબિન્સન, 2016). ઇનામ / પ્રેરણા સર્કિટની ટકાઉ સંવેદનશીલતા ડ્રગની શોધ સાથે સંકળાયેલ પ્રોત્સાહન-સંવેદનાના સમાવેશમાં સામેલ છે. ઈનામ / પ્રેરણા સર્કિટ એ મિડબ્રેઇન ડોપામાઇન ન્યુરોન્સનો બનેલો છે સસ્તાં નિગ્રા (એસ.એન.) અને વેન્ટ્રલ ટેગમેન્ટલ એરિયા (વીટીએ), જે અનુક્રમે સ્ટ્રાઇટમના ડોર્સલ અને વેન્ટ્રલ સ્તરને લક્ષ્ય આપે છે. વેન્ટ્રલ સ્ટ્રાઇટમ અથવા ન્યુક્લિયસ એકમ્બમ્બન્સ (એનએસી) ને પ્રોજેકટ કરતી ડોપામાઇન ન્યુરોન્સ પરંપરાગત રીતે ધ્યેયલક્ષી વર્તણૂકથી સંબંધિત છે, જ્યારે ડોસોમાઇન ચેતાકોષો જે ડોર્સલ સ્ટ્રાઇટમને પ્રોજેકટ કરે છે તે ટેવ પ્રાપ્તિ સાથે સંકળાયેલા છે.એવરિટ અને રોબિન્સ, 2005; વાઈસ, 2009; યાજર એટ અલ., 2015; વોલ્કો એટ એટ., 2017).
ઈનામ / પ્રેરણા સર્કિટનું સંવેદનશીલતા ઉંદરોમાં જોવા મળે છે કારણ કે સંભવિત વ્યસનકારક દવાના ફિક્સ ડોઝના વારંવાર વહીવટ દ્વારા લોમમોટર પ્રવૃત્તિમાં ધીમે ધીમે વધારો થાય છે (પીઅર્સ અને કાલિવાસ, 1997; રોબિન્સન અને બેરીજ, 2001). લોકમોટર સેન્સેટાઇઝેશન એ એક ટકાઉ ઘટના છે કારણ કે તે અઠવાડિયા, મહિનાઓ અને ડ્રગના ઉપાડ પછી એક વર્ષ પછી પણ અવલોકનક્ષમ છે.રોબિન્સન અને બેરીજ, 1993). વહેલી તકે સૂચવવામાં આવ્યું હતું કે ઈનામ / પ્રેરણા સર્કિટની સંવેદનશીલતા અનિવાર્ય દવા શોધવામાં ફાળો આપે છે (રોબિન્સન અને બેરીજ, 1993). તદનુસાર, લોકોમોટર સંવેદના પુનizationસ્થાપનની શોધમાં સ્વ-વહીવટ કોકેનને સુવિધા આપે છે (ડી વેરીઝ એટ અલ., 2002). તદુપરાંત, કોકેઇન સ્વ-વહીવટની વિસ્તૃત withક્સેસવાળા ઉંદરો મર્યાદિત withક્સેસવાળા ઉંદરો કરતા કોકેઇન માટે વધુ સ્થાનિક લોકોનો પ્રતિસાદ દર્શાવે છે (ફેરારીયો એટ અલ., 2005). આ ઉપરાંત, મનોરોગનિરોધકોમાં અંતર્ગત લોમોમોટર સંવેદનશીલતાના ન્યુરોકેમિકલ ફેરફારો પણ અનિવાર્ય દવાઓની શોધમાં જોવા મળે છે (સ્ટેક્ટી અને કાલિવસ, 2011; જિયુલિયાનો એટ અલ., એક્સએન્યુએમએક્સ). આ ડેટા લોકોમોટર સંવેદના અને મનુષ્યમાં જોવાઈ રહેલી અનિવાર્ય દવાની માંગ વચ્ચેના પ્રારંભિક સૂચિત પત્રવ્યવહારને સમર્થન આપે છે (રોબિન્સન અને બેરીજ, 1993; વંડર્સચ્યુરેન અને કાલિવસ, 2000). યાંત્રિકરૂપે, દુરુપયોગની દવાઓનો વારંવાર એડમિનિસ્ટ્રેશન મેસોલીમ્બિક ડોપામાઇન સર્કિટ્સમાં ડોપામિનેર્જિક ન્યુરોટ્રાન્સમિશનમાં વધારો થાય છે. કોકોઈન અથવા એમ્ફેટામાઇન્સ જેવા સાયકોસ્ટીમ્યુલેન્ટ્સ, જે પ્લાઝ્મા મેમ્બ્રેન ડોપામાઇન ટ્રાન્સપોર્ટર (ડીએટી) ને અવરોધે છે, સ્ટ્રાઇટમ અને એનએસીમાં સિનેપ્ટિક જગ્યામાં ડોપામાઇનના મોટા પ્રમાણમાં વધારો કરે છે, આમ લોકમotionશનને સક્રિય કરે છે (સ્ટેક્ટી અને કાલિવસ, 2011). માદક પદાર્થના વ્યસનની જેમ, ડોપામાઇન ઇનામ / પ્રેરણા સર્કિટની સંવેદના ઓસીડીમાં જોવાયેલી અનિવાર્ય વર્તણૂકોને ફાળો આપે છે. ખરેખર, ડોપામાઇન ડી 2 રીસેપ્ટર્સ (ડી 2 આર) નું પુનરાવર્તિત સક્રિયકરણ, ઉંદરો અને ઉંદર બંનેમાં લોકમોટર સંવેદના અને વર્તન તપાસવા માટે પ્રેરિત કરવા માટે પૂરતું છે (સેશેટમેન એટ અલ., 1998; સેશેટમેન એટ અલ., 1999; સૂર્ય એટ અલ., 2019). ક્વિનપાયરોલનું વારંવાર વહીવટ, ડી 2 આર / ડી 3 આર એગોનિસ્ટ, તે ઓસીડીનું સ્વીકૃત મોડેલ છે કારણ કે તે ફરજિયાત તપાસ અને સ્ટીરિયોટાઇપ વર્તન, આગાહી માન્યતાના વધારા દ્વારા ચહેરાની માન્યતાને સુધારે છે, જેમ કે સેરોટોનિન રીપ્પેક સાથે લાંબી સારવાર પછી અનિવાર્ય વર્તણૂકોમાં ઘટાડો જોવા મળે છે. અવરોધકો (એસઆરઆઈ) અને માન્યતા નિર્માણ કરે છે કારણ કે આ મોડેલમાં સામેલ મગજની રચનાઓ પેથોલોજીમાંના લોકો સાથે વહેંચાયેલી છે (સ્ટુચલિક એટ અલ., 2016; સેશેટમેન એટ અલ., 2017). સારાંશમાં, ડોપામાઇન ટ્રાન્સમિશનનું વારંવાર સક્રિયકરણ, ક્યાં તો પૂર્વ-સિનેપ્ટિક (ડોપામાઇન પ્રકાશન) અથવા પોસ્ટ-સિનેપ્ટિક (ડી 2 આરનું સક્રિયકરણ) પદ્ધતિઓ દ્વારા લોકોમોટર સંવેદના અને અનિવાર્ય વર્તન તરફ દોરી જાય છે.
કપ્પા ioપિઓઇડ સિસ્ટમ એ ઇનામ / પ્રેરણા સર્કિટમાં ડોપામાઇન ટ્રાન્સમિશનને નિયંત્રિત કરતી સૌથી પ્રગા. સિસ્ટમ્સમાંની એક છે. પુરાવા બતાવે છે કે કપ્પા-ioપિઓઇડ ટ્રાન્સમિશન ડોપામાઇનની અસરોનો વિરોધ કરે છે; કપ્પા ioપિઓઇડ રીસેપ્ટર્સ (કેઓઆર) ની તીવ્ર સક્રિયકરણ મનોવિરોધીકરણ દ્વારા પ્રેરિત લોમોમોટર પ્રવૃત્તિનો પ્રતિકાર કરે છે. (ગ્રે એટ અલ., 1999). તેનાથી વિપરીત, પુનરાવર્તિત KOR સક્રિયકરણ અનિવાર્ય અને રીualો ડ્રગ લેવી જાળવી રાખે છે અને વધારે છે (કોઓબ, 2013). દુરૂપયોગની દવાઓનો વપરાશ હોમિયોસ્ટેટિક ઉન્નત કપ્પા opપિઓઇડ ટ્રાન્સમિશનને પ્રેરિત કરે છે, કદાચ ડિસફોરિયાની નકારાત્મક ભાવનાત્મક સ્થિતિમાં ફાળો આપે છે (કોઓબ, 2013) અનિવાર્ય દવાઓના ઉપયોગને ટ્રિગર કરવું (ચાવકીન અને કુબ, 2016). હકીકતમાં, કેઓઆર નાકાબંધી તણાવને અટકાવી હતી - પરંતુ નિકોટિનને ડ્રગ દ્વારા પ્રેરિત પુનstસ્થાપન નહીં (જેકસન એટ અલ., 2013), કોકેન (બીર્ડેલી એટ અલ., 2005) અને ઇથેનોલ (સ્પર્લિંગ એટ અલ., 2010). આ શોધની સાથે સાથે, કેઓઆર નાકાબંધી, એમ્ફેટેમાઈન સેન્સેટીનાઇઝ્ડ ઉંદરોના ડોર્સોનેટ્રલ સ્ટ્રાઇટમમાં ડોપામિનર્જિક ફેરફારોને બદલી નાંખે છે, ડ્રગના તેમના ઉન્નત લોકોમોટર પ્રતિસાદમાં ફેરફાર કર્યા વિના (એઝોકાર એટ અલ., 2019). આમ, કેઓઆર સિસ્ટમ નકારાત્મક મજબૂતીકરણમાં વધારો કરતી દવા-મૂલ્યમાં વૃદ્ધિ કરે તેવું લાગે છે. OCD માં, નકારાત્મક મજબૂતીકરણને મનોગ્રસ્તિઓ દ્વારા ઉત્તેજીત કરવામાં આવે છે, જે તે મનોગ્રસ્તિને ટાળવા માટે આપેલ મજબૂરીને મજબૂત બનાવે છે. તેમ છતાં તેની સીધી પરીક્ષણ કરવામાં આવી નથી, નકારાત્મક મજબૂતીકરણ ક્વિનપાયરોલ સંવેદના પર ભૂમિકા ભજવી શકે છે. ખરેખર, ડી 2 આર નકારાત્મક મજબૂતીકરણના નિર્માણમાં સામેલ છે. ઉદાહરણ તરીકે, મોર્ફિનમાં સ્થાન ટાળો- ઉપાડ-જોડી વિસ્તાર D2R ના લાંબા આઇસોફોર્મના અભાવમાં ઉંદરમાં વિકસિત થયો ન હતો (સ્મિથ એટ અલ., 2002) અને ત્યાગના સમયગાળા દરમિયાન પુનરાવર્તિત ક્વિનપાયરોલ ટ્રીટમેન્ટ, ઓટો-એડમિનિસ્ટ્રેશન દાખલાની શોધમાં રહેલા કોકેન અને હેરોઇનને ફરીથી સ્થાપિત કરે છે, આ અસર ક્વિનપાયરોલથી સંવેદનશીલ લોમમોશનથી સંબંધિત છે (ડી વેરીઝ એટ અલ., 2002), સાયકોસ્ટીમ્યુલેન્ટ અને ક્વિનપાયરોલ-પ્રેરિત સંવેદના વચ્ચે વહેંચાયેલ મિકેનિઝમ્સ સૂચવે છે. તદુપરાંત, ખુલ્લા મેદાનના ક્ષેત્રમાં હોમ કેજની રજૂઆત, પરંતુ નવલકથાની પાંજરું નહીં, લોકોમોટર સંવેદના અને ફરજિયાત ચેકિંગ વર્તન ઘટાડે છે (સેશેટમેન એટ અલ., 2001) સૂચવે છે કે સલામતી / પરિચિત સંકેતો નકારાત્મક પર્યાવરણીય સંકેતો સાથે સ્પર્ધા કરી શકે છે જે સંવેદનશીલતા તરફેણ કરે છે. તેવી જ રીતે સાયકોસ્ટીમ્યુલેન્ટ-પ્રેરિત સંવેદના માટે, કે.ઓ.આર.નું પુનરાવર્તિત સક્રિયકરણ, લોમોમોટર સેન્સેટાઇઝેશનને સગવડ કરે છે (એસ્કોબાર એટ અલ., 2017) અને ફરજિયાત ચેકિંગ વર્તન (પેરેલaultલ્ટ એટ અલ., એક્સએન્યુએમએક્સ) ક્વિનપાયરોલના વારંવારના વહીવટ દ્વારા પ્રેરિત. આ સંભવિત અસર ઉન્નત નકારાત્મક મજબૂતીકરણનું પરિણામ છે કે નહીં તે સ્પષ્ટ કરવાનું બાકી છે.
તાજેતરમાં કરવામાં આવેલા સંપૂર્ણ વિશ્લેષણ બતાવે છે કે ડોપામિનેર્જિક ટ્રાન્સમિશન પર કપ્પા-opપિઓઇડ સિસ્ટમની અસર જટિલ છે: તે ડોપામાઇનના માર્ગ પર શામેલ છે (માર્ગોલિસ એટ અલ., 2006; માર્ગોલિસ એટ અલ., 2008), અને કેઓઆર રીસેપ્ટરના સક્રિયકરણ અને ડોપામાઇન રીસેપ્ટરના સક્રિયકરણ વચ્ચેના સમય પર (ચાર્ટઓફ એટ અલ., 2016). આ જટિલતા સાથે સુસંગત, કેઓઆર લિગાન્ડ્સના સંભવિત રોગનિવારક ઉપયોગની વ્યાપક ચર્ચા કરવામાં આવી છે. એવું સૂચન કરવામાં આવ્યું છે કે ડ્રગના ઉપયોગના તબક્કા દરમિયાન કે.ઓ.આર. એગોનિસ્ટ ક્લિનિક રૂપે ઉપયોગી થઈ શકે છે, ડ્રગને પ્રોત્સાહનિત હાયપરડોપામિનર્જીયા (શિપ્પેનબર્ગ એટ અલ., 2007). બીજી તરફ, કેઆરએન વિરોધી વારંવાર ડ્રગના વપરાશ પછી ડાયનોર્ફિન અભિવ્યક્તિમાં વધારો દ્વારા પ્રેરિત ઉપાડ સિન્ડ્રોમની સારવારમાં ઉપયોગી થઈ શકે છે.વી અને કોઓબ, 2010). તદનુસાર, એવી દરખાસ્ત કરવામાં આવી છે કે કે.આર. આંશિક વ્યથિત (બ્યુગિન એટ અલ., 2012) વ્યસનીમાં અનિવાર્ય ડ્રગના સેવન અને ખસીના લક્ષણો બંનેની સારવાર માટે રોગનિવારક વિકલ્પ હોઈ શકે છે.ચાર્ટઓફ એટ અલ., 2016; કેલાઘાન એટ અલ., 2018). આ સમીક્ષામાં, અમે વર્તન સંવેદના અને ફરજિયાતતાના ડોપામિનર્જિક સહસંબંધના સમય / સંદર્ભ આધારિત મોડ્યુલેશનનું વિશ્લેષણ કરીએ છીએ.
સ્ટ્રાએટલ અને મિડબ્રેઇન પ્રદેશોમાં કપ્પા ઓપિયોઇડ અને ડોપામિનર્જિક સિસ્ટમ્સ વચ્ચેની એનાટોમિકલ અને ફંક્શનલ ક્રિસ્ટલstalક
સ્ટ્રાઇટલ પ્રદેશો
કેઓઆર જીઆઈ / ઓ પ્રોટીન-જોડી રીસેપ્ટર્સ છે જે મધ્યબ highlyન ડોપામાઇન સિસ્ટમમાં ખૂબ વ્યક્ત થાય છે (મનસુર એટ અલ., એક્સ્યુએનએક્સ). આ રીસેપ્ટર્સ મ્યુ (એમઓઆર), ડેલ્ટા (ડીઓઆર) અને કપ્પા (કેઓઆર) દ્વારા રચિત ઓપીયોઇડ રીસેપ્ટર્સના પરિવારના છે. આ રીસેપ્ટર્સ માટે એન્ડોજેનસ એગોનિસ્ટ્સ અનુક્રમે એન્ડોર્ફિન્સ, એન્કેફાલિન અને ડાયનોર્ફિન છે. સ્ટ્રાઇટમમાં, ડાયનોર્ફિનને ડોપામાઇન ડી 1 રીસેપ્ટર (ડી 1 આર) દ્વારા સંશ્લેષણ કરવામાં આવે છે -મધ્યમ કદના ન્યુરોન્સ (એમએસએન) સમાવે છે જે સમાન ન્યુક્લિય (કેઓઆર) ને પુનરાવર્તિત ચેતાક્ષ ધરાવે છે (મનસુર એટ અલ., એક્સ્યુએનએક્સ). ઉંદર એનએસીની ઇલેક્ટ્રોન માઇક્રોસ્કોપી છબીઓ બતાવે છે કે કેઓઆર મુખ્યત્વે ડીએટી ધરાવતા પ્રિસ્નાપ્ટિક માળખામાં જોવા મળે છે જ્યારે કેઓઆરનો નજીવો ભાગ ડેટ્રાઇટ્સ પર ડીએટીને નિમણૂકમાં સ્થાન આપે છે (સેવિઓસ એટ અલ., 2001; કિવેલ એટ અલ., 2014). એનએસી તરફથી પ્રેસિનપ્ટીક-સિનેપ્ટોસોમલ તૈયારીઓનું લક્ષણ દર્શાવતા ઇમ્યુનોફ્લોરોસેન્ટ અભ્યાસ દર્શાવે છે કે કેઓઆર અને ડી 2 આર પ્રાધાન્યમાં ડોપામાઇન સિન્થેટીઝિંગ એન્ઝાઇમ, ટાઇરોસિન હાઇડ્રોક્સિલેઝ (ટીએચ) ધરાવતા સિનેપ્ટોઝોમ્સમાં સહઅસ્તિત્વ ધરાવે છે.એસ્કોબાર એટ અલ., 2017). તદુપરાંત, એનએસી અને સ્ટ્રાઇટમના સેલ બ bodiesડીઓમાં કેઓઆર વિપુલ પ્રમાણમાં હોય છે, અને સેલ પેટા વસ્તીમાં ડી 2 આર સાથે એકરૂપ થાય છે (એસ્કોબાર એટ અલ., 2017). આનુવંશિક અને પરમાણુ આંતરદૃષ્ટિ સાથે, એવું સૂચવવામાં આવ્યું છે કે સ્ટ્રાઇટમમાં 20% કુલ KOR બંધનકર્તા DA ટર્મિનલ્સમાં અવલોકન કરવામાં આવે છે (વેનટ વીર એટ અલ., 2013). તદુપરાંત, તેજેડા એટ અલ. (2017) બતાવ્યું કે ડી 1 આર અને ડી 2 આર એમએસએન બંને એમએસએન ધરાવતા ડી 1 આર માટે preંચી પસંદગી સાથે કેઓઆર વ્યક્ત કરે છે (તેજેડા એટ અલ., 2017). આ એનાટોમિકલ ડેટા સૂચવે છે કે KORs ઇનામ / પ્રેરણા સર્કિટમાં ડોપામાઇન ન્યુરોટ્રાન્સમિશનને નિયમન કરીને પૂર્વ અને અનુસૂચિત રૂપે હાજર છે.
કેટલાંક પ્રાયોગિક અભિગમો બતાવે છે કે કેઓઆરનું સક્રિયકરણ ડોપામાઇન રિલેઝને અટકાવે છેઇ. પ્રણાલીગત ઇંજેક્શન દ્વારા કેઓઆરની તીવ્ર સક્રિયકરણ અથવા એગોનિસ્ટ્સના સ્થાનિક પ્રેરણા, એનએસીમાં ડોપામાઇનના બાહ્ય સ્તરને ઘટાડે છે (દી ચીરા અને ઇમ્પેરોટો, 1988; સ્પેનેગેલ એટ અલ., 1992; ફુએન્ટેઆલ્બા એટ અલ., 2006) અને ડોર્સલ સ્ટ્રાઇટમ (ગેહર્કે એટ અલ., 2008). ડોપામાઇન ન્યુરોટ્રાન્સમિશન પર KORs ની ટોનિક અવરોધક ક્રિયાને ટેકો આપવો, લાંબા સમયથી ચાલતા અને પસંદગીયુક્ત KOR વિરોધી નોર-બાયનોલ્ટરફાઇમિન (ન-બીએનઆઈ) નો સીધો પ્રભાવબ્રોડબેઅર એટ અલ., 1994) એનએસીમાં ડોપામાઇનનું મૂળભૂત સ્તર વધે છે (સ્પેનેગેલ એટ અલ., 1992) અને ડોર્સિન સ્ટ્રાઇટમમાં ડોપામાઇન પ્રકાશન (એઝોકાર એટ અલ., 2019). કેઓઆર નોકઆઉટ ઉંદરમાં કેઓઆર ટોનિકના નિષેધના અંતિમ પુરાવા બતાવવામાં આવ્યા હતા, જેમાં સ્ટ્રાઇટમ અને એનએસીમાં ડોપામાઇનના બાહ્ય સેલના સ્તરને દર્શાવવામાં આવ્યા હતા (ચેફર એટ અલ., 2005). ડોપામાઇન પ્રકાશનના કેઓઆર નિષેધ માટે જવાબદાર મિકેનિઝમ્સ સંપૂર્ણપણે સ્પષ્ટ નથી. જો કે, તે જાણીતું છે કે કેઓઆરના સક્રિયકરણથી કે + નો વધારો થાય છે અને સીએ 2 + વાહકતામાં ઘટાડો થાય છે, આમ કોષના અતિસંવેદનશીલતા અને વેસિકલર ન્યુરોટ્રાન્સમીટર પ્રકાશનને અવરોધિત કરે છે (બ્રુહાસ અને ચેકીન, 2010; માર્ગોલીસ અને કારખાનીસ, 2019).
વધુમાં, ઈન વિટ્રોમાં અને વિવો માં વિધેયાત્મક ડેટા સૂચવે છે કે કેઓઆર DAT ની પ્રવૃત્તિમાં ફેરફાર કરીને ડોપામાઇન એક્સ્ટ્રાસેલ્યુલર સ્તરોમાં ફેરફાર કરે છે. હમણાં પૂરતું, ઇએમ 4 સેલ્સમાં કેઓઆરનું સક્રિયકરણ જે કેઓઆર અને ડીએટીને સહ-વ્યક્ત કરે છે, વોલ્ટેમેટ્રી દ્વારા માપવામાં આવેલા ડોપામાઇનનું પ્રમાણ વધે છે (કિવેલ એટ અલ., 2014). એન ભૂતપૂર્વ વિવો વિશ્લેષણમાં પણ અસંતુલિત પેશીઓમાં વોલ્ટેમેટ્રીનો ઉપયોગ કરીને બતાવ્યું કે કેઓઆર એગોનિસ્ટ યુ-69593 ના પ્રણાલીગત ઇન્જેક્શનમાં એનએસીમાં ડોપામાઇનનો વપરાશ વધ્યો (થોમ્પસન એટ અલ., 2000). આ જ પ્રકારનો તાજેતરનો લેખ બતાવે છે કે નોર-બીએનઆઈ એમપી 1104 ના તીવ્ર પ્રણાલીગત ઇન્જેક્શન દ્વારા પ્રેરિત, વેન્ટ્રલ અને ડોર્સલ સ્ટ્રાઇટમમાં ડોપામાઇનના વપરાશના વધારાને અવરોધે છે, મિશ્ર કપ્પા / ડેલ્ટા ioપિઓઇડ રીસેપ્ટર એગોનિસ્ટ (એટિગારી એટ અલ., 2019). તેમ છતાં, ડોપામાઇનના ઉપભોગ પર કેઓઆર સક્રિયકરણની અસર હજી સંપૂર્ણ રીતે સ્પષ્ટ કરી શકાતી નથી. કેઓઆરના આંશિક એગોનિસ્ટ નલ્ફેફેનના પ્રણાલીગત વહીવટ, સ્ટ્રિએટલ ડોપામાઇનનો ઉપભોગ આશ્રિત રીતે ઘટાડ્યો, ઝડપી સ્કેન સાયક્લિક વોલ્ટેમેટ્રી (એફએસસીવી) દ્વારા પ્રમાણિત (ગુલાબ એટ અલ., 2016). પુખ્ત પુરૂષ ઉંદરોમાં નો-નેટ ફ્લક્સ માઇક્રોડાયલિસીસનો ઉપયોગ કરીને, કેઓઆરને અવરોધિત કરવા સાથે નિષ્કર્ષણ અપૂર્ણાંક (એડ) માં વધારો થયો હતો, જે ડોપામાઇન અપટેકનું પરોક્ષ પગલું છે (ચેફર એટ અલ., 2006; એઝોકાર એટ અલ., 2019) સૂચવે છે કે કે.ઓ.આર. નું ટોનિક સક્રિયકરણ DAT પ્રવૃત્તિ (ડોપામાઇન અપટેક) પર અવરોધક નિયંત્રણ લાવે છે. આ પરિણામો ડોપામાઇન એક્સ્ટ્રા સેલ્યુલર સ્તરને નિયંત્રિત કરવા માટે ડોપામાઇનના ઉપભોગ પર અંતર્જાત કેઓઆર પ્રવૃત્તિની જટિલ ભૂમિકાને પ્રકાશિત કરે છે. એફએસસીવી જેવા ઉચ્ચ ટેમ્પોરલ રિઝોલ્યુશન અભિગમો ડોપામાઇનના ઉપભોગ પર કેઓઆરની અસર દર્શાવવામાં નિષ્ફળ થયા છે (ઇબનર એટ અલ., 2010; એહરિચ એટ અલ., 2015; હોફમેન એટ અલ., 2016), સૂચવે છે કે સ્ટ્રિએટલ પ્રદેશોમાં DAT પ્રવૃત્તિ વધારવા માટે KOR એ સેવન અવધિની જરૂર છે., DAT પ્રવૃત્તિના KOR- મધ્યસ્થી વૃદ્ધિને KOR સક્રિયકરણ દ્વારા પ્રેરિત સેલ મેમ્બ્રેન પર DAT ની સંખ્યામાં વધારો દ્વારા સમજાવી શકાય છે, સ્ટ્રિએટલ સિનેપ્ટોઝોમ્સ અને સેલમાં અહેવાલ આપ્યો છે. રેખાઓ (કિવેલ એટ અલ, 2014).
મિડબ્રેઇન પ્રદેશો
ઉંદરના મિડબ્રેઇનમાં કરવામાં આવેલ oraટોરાડીયોગ્રાફિક એસિઝ એસએન અને વીટીએ (રોટરોકudડલ અક્ષ) પર કેઓઆર માટે નોંધપાત્ર બંધનકર્તા દર્શાવે છે.સ્પેસિએલ એટ અલ., 1993). બીજી બાજુ, ઇલેક્ટ્રોન માઇક્રોસ્કોપી ડેટા દર્શાવે છે કે ડાયનોર્ફિન ધરાવતા ટર્મિનલ્સ સીએનએચ એસ પોઝિટિવ ડેંડ્રિટ્સ પર સીએનએસ અને વીટીએ (સેસેક અને પીકેલ, 1992) સૂચવે છે કે કેઓઆર ડોપામાઇન ન્યુરોન્સના સોમેટોડેન્ડ્રિટિક ભાગોમાં સ્થાનિક કરે છે. સ્ટ્રિએટલ ડી 1 આર ધરાવતા એમએસએનએસ એ મિડબ્રેઇન ડોપામાઇન ન્યુરોન્સ માટેનો ડાયનોર્ફિન ઇનપુટ્સ છે. રસપ્રદ વાત એ છે કે, કે.ઓ.એસ. ના અવરોધથી ડી.એ.આર.-એમ.એસ.એન. ની પ્રતિબંધિત અસરને વી.ટી.એ. ડોપામાઇન ન્યુરોન્સમાં સુધારવામાં આવતી નથી, જે દર્શાવે છે કે આ અવરોધ GABA દ્વારા મધ્યસ્થી છે (એડવર્ડ્સ એટ અલ, 2017). કેઓઆર ડોપામાઇન મિડબ્રેઇન ન્યુરોન્સના સોમેટોડેન્ડ્રિટિક જવાબોને મોડ્યુલેટ કરે છે. ઇલેક્ટ્રોફિઝિયોલોજીકલ અધ્યયન દર્શાવે છે કે વીટીએ હાયપરપોલારિઝમાં કેઓઆરનું સક્રિયકરણ અને ડોપામાઇન ન્યુરોન્સના સ્વયંસ્ફુરિત ફાયરિંગ રેટમાં ઘટાડો થાય છે (માર્ગોલિસ એટ અલ., 2003). પરિણામે, કેઓઆર એગોનિસ્ટ્સનું પ્રેરણા સોમેટોડેન્ડ્રિટિક ડોપામાઇન ફ્લxક્સમાં ઘટાડો કરે છે (સ્મિથ એટ અલ., 1992; ડાલમેન અને ઓ'માલે, 1999). જો કે, ડોપામાઇન ન્યુરોન્સ પર કેઓઆરની આ અવરોધક અસર સર્કિટ આધારિત હોવાનું જણાય છે. વીટીએમાં કપ્પા-ioપિઓઇડ એગોનિસ્ટ્સનું પ્રેરણા મેડિયલ પ્રિફ્રન્ટલ કોર્ટેક્સ (એમપીએફસી) માં ડોપામાઇન પ્રકાશનમાં ઘટાડો કરે છે.માર્ગોલિસ એટ અલ., 2006) પરંતુ એનએસીમાં નથી (ડેવીન એટ અલ., 1993; માર્ગોલિસ એટ અલ, 2006). તદુપરાંત, માર્ગોલીસ એટ અલ. (2006) મળ્યું કે કેઓઆરઓ વીટીએ ડોપામાઇન ન્યુરોન્સને અવરોધે છે જે એમપીએફસી અને બેસોલ્ટ્રલ એમીગડાલાને પ્રોજેકટ કરે છે, પરંતુ જે તે એનએસીને પ્રોજેક્ટ કરે છે. તે જ વર્ષે, ફોર્ડ એટ અલ. (2006) બતાવ્યું છે કે માઉસ વીટીએ ટુકડાઓમાં કેઓઆર એગોનિસ્ટ્સની બાથ એપ્લિકેશન એ એનએસીને પ્રોજેકટ કરેલા ડોપામાઇન ન્યુરોન્સમાં outંચી બાહ્ય પ્રવાહ પ્રેરિત કરે છે, જે દર્શાવે છે કે કેઓઆરઝ ડોપામાઇન ન્યુરોન્સના એનએસી કરતા વધુ પ્રોજેક્ટ કરે છે. એમીગડાલાને. તદુપરાંત, કેઓઆરનું સક્રિયકરણ ઉત્તેજનાના કંપનવિસ્તારમાં ઘટાડો કરે છે (માર્ગોલિસ એટ અલ., 2005) અને અવરોધક (ફોર્ડ એટ અલ., 2007) મિડબ્રેઇન ડોપામાઇન ન્યુરોન્સમાં પોસ્ટસિએપ્ટિક પ્રવાહો. પ્રજાતિઓ અને એમ.પી.એફ.સી. અને એન.એ.સી. (વી.ટી.એ.) ના જટિલ પ્રોફિરેન્ટ પ્રોક્કેશન્સ વચ્ચેના તફાવતો (વાન બોકસ્ટેલે અને પિકેલ, 1995; કાર અને સેસૅક, 2000) તે સ્થાપિત કરવું પડકારજનક બનાવે છે કે કે.ઓ.આર.એસ. વી.ટી.એ.માં ન્યુરોનલ ડોપામાઇન વસ્તીમાંથી કેટલાકને પસંદગીયુક્ત રીતે અટકાવે છે. તેમ છતાં, અહીં સારાંશ આપવામાં આવેલા ડેટા સૂચવે છે કે કેઓઆર ડોપામાઇન ન્યુરોન્સના સોમા અને ટર્મિનલ્સમાં છે, તેમજ ઇનપુટ્સ કે જે તેમને નિયમન કરે છે, આમ મધ્યબbraન ડોપામાઇન ન્યુરોન્સની સિનેપ્ટિક પ્રવૃત્તિને નિયંત્રિત કરવા માટે ઉત્કૃષ્ટ સ્થિતિમાં છે.
સાયકોસ્ટીમ્યુલન્ટ્સ-પ્રેરિત સેન્સિટાઇઝેશન અને અનિવાર્ય વર્તણૂકોમાં ડોપામાઇન ન્યુરોટ્રાન્સમિશનને નિયંત્રિત કરતી KORs ની ભૂમિકા.
ડ્રગ વ્યસન એ એક પ્રક્રિયા છે જેમાં શરૂઆતમાં આવેગજનક દવા લેવી પડે છે જેની સકારાત્મક અસરકારક અસરો સાથે સંકળાયેલ હોય છે. બીજી બાજુ, અનિવાર્યતા ડ્રગ વ્યસનીમાં નિરીક્ષણયોગ્ય એક વ્યક્તિત્વ લક્ષણ છે. ડોપામિનેર્જિક માર્ગોના કેટલાક ન્યુરોઆડેપ્ટેશન્સ, દુરુપયોગની દવાઓના વારંવાર સંપર્કમાં આવતા, અનિવાર્ય ડ્રગની શોધ અને ઇન્ટેક માટેના હિસાબ માટે સૂચન કર્યું છે (એવરિટ અને રોબિન્સ, 2005; કોઓબ અને વોલ્કો, 2016). અનિવાર્ય ડ્રગનું સેવન ડ્રાઇવિંગ સૂચિત પૂર્વધારણાઓમાંથી એક એ તેના નકારાત્મક-દબાણયુક્ત અસરોની સંવેદના છે (કોઓબ, 2013). ડોપામાઇન પ્રકાશન પર કપ્પા ioપિઓઇડ સિસ્ટમનું અવરોધક નિયંત્રણ દુરુપયોગની દવાઓના નકારાત્મક-પ્રબલિત ગુણધર્મોને ફાળો આપી શકે છે. જો કે, ડોપામાઇન ન્યુરોટ્રાન્સમિશન અને ફરજિયાત દવાઓ માંગતી દવાઓ પર કેઓઆર સક્રિયકરણના પરિણામો જટિલ અને દેખીતી રીતે વિરોધાભાસી લાગે છે. ખરેખર, એમ્ફેટામાઇન અને કોકેઇન દ્વારા પ્રેરિત ડોપામાઇન પ્રકાશન, કે.ઓ.આર. એકોનિસ્ટ્સના સહવર્તી વહીવટ દ્વારા ઘટાડવામાં આવે છે (હીડબ્રેડર અને શિપ્નબર્ગ, 1994; મેઇઝન્યુવ એટ અલ., 1994; થોમ્પસન એટ અલ., 2000) અને તે પણ ઘટાડે છે કોકેઇન સ્વ-વહીવટ (નેગસ એટ અલ., 1997). તદુપરાંત, સાયકોસ્ટીમ્યુલેન્ટ્સના વારંવાર સંપર્કમાં આવતાં પોસ્ટ-સિનેપ્ટિક ડી 1 આરના સતત સક્રિયકરણના જવાબમાં મેસોલીમ્બીક માર્ગના ડોપામાઇન પ્રકાશન અંગે કેઓઆરઓ અવરોધક પ્રતિસાદ આપે છે.કોલ એટ અલ., 1995; નેસ્લેર, 2001). વિચિત્ર રીતે, કેઓઆરનું સક્રિયકરણ, ઇનામ / પ્રેરણા માર્ગમાં ડોપામાઇન પ્રકાશનની સુવિધા પણ કરી શકે છે (ફુએન્ટેઆલ્બા એટ અલ., 2006; ફુએન્ટેઆલ્બા એટ અલ., 2007) અને સાયકોસ્ટીમ્યુલેન્ટ્સ વપરાશ (વી એટ અલ., 2009). ફુએન્ટેઆલ્બા એટ અલ. (2007) બતાવ્યું કે ચાર દિવસ U69593 વહીવટ કર્યા પછી, એક KOR એગોનિસ્ટ, એનએસીમાં એમ્ફેટેમાઇન પ્રેરિત ડોપામાઇન પ્રકાશનમાં વધારો કર્યો. તાજેતરમાં, એવું દર્શાવવામાં આવ્યું હતું કે કેઓઆરને અવરોધિત કરવાથી ડોફેમાઇન પ્રકાશનમાં ફેરફાર અને ડોર્સ્ટલ સ્ટ્રાઇટમમાં વધારો થાય છે જે એમ્ફેટામાઇન દ્વારા પ્રેરિત લોકોમોટર સંવેદના દરમિયાન થાય છે (એઝોકાર એટ અલ., 2019). એકસાથે, આ ડેટા સૂચવે છે કે કેઓઆરનું સક્રિયકરણ દુરુપયોગની દવાના સકારાત્મક-પ્રબલિત ગુણધર્મોને પણ ફાળો આપી શકે છે (ચાર્ટઓફ એટ અલ., 2016).
આ ઉપરાંત, કેઓઆરએસ સક્રિયકરણ પણ અનિવાર્ય ડ્રગની શોધમાં ફાળો આપે તેવું લાગે છે; કેઓઆર નાકાબંધી કોકેન ઘટાડે છે (વી એટ અલ., 2009), હેરોઈન (સ્ક્લોસબર્ગ એટ અલ., 2013) અને મેથામ્ફેટામાઇન (વ્હિટફિલ્ડ એટ અલ., 2015) ડ્રગની અમર્યાદિત withક્સેસવાળા ઉંદરોમાં સેવન (વી એટ એટલ, 2009). આ અસર તાણ-પ્રેરિત ડ્રગની શોધમાં પણ પુરાવા છે. હમણાં પૂરતું, કેઓઆર નોકઆઉટ ઉંદરને દબાણયુક્ત તરણ પછી કોકેઇન સ્થાનની પસંદગી દર્શાવતી નથી (મેક્લોફ્લિન એટ અલ., 2006 એ). કેઓઆરને અવરોધિત કરવાથી મજબૂતાઇથી તનાવના તાણના સંપર્કમાં આવતાં નિકોટિન સ્થળની પસંદગીને ઘટાડવામાં આવે છે (સ્મિથ એટ અલ., 2012). રસપ્રદ વાત એ છે કે કે.ઓ.આર. અવરોધિત થવાથી તનાવથી પ્રેરિત કોકેન અને નિકોટિન સચેત થાય છે, પરંતુ ડ્રગ ચેલેન્જ દ્વારા પ્રેરિત શોધવામાં અસર થતી નથી.બીર્ડેલી એટ અલ., 2005; જેકસન એટ અલ., 2013). તણાવ દ્વારા પ્રેરિત આ સુવિધાયુક્ત કેઓઆર અસર પુરસ્કાર / પ્રેરણા સર્કિટ દ્વારા મધ્યસ્થી હોવાનું લાગે છે (શિપ્પેનબર્ગ એટ અલ., 2007; વી અને કોઓબ, 2010). ડો. કૌર અને તેના જૂથ દ્વારા કરવામાં આવેલા એક ભવ્ય અધ્યયનમાં, તે બતાવવામાં આવ્યું હતું કે વીટીએમાં કેઓઆરને અવરોધિત કરવું, અગાઉ અથવા તીવ્ર તણાવ પછી, કોકેન-શોધવાની પુનstસ્થાપન અટકાવે છે, જે અસર લાંબા ગાળાના બચાવ સાથે સંકળાયેલ છે. ડોપામાઇન ચેતાકોષોમાં અવરોધક સિનેપ્સનું સંમિશ્રણ (ગ્રાઝિયન એટ અલ., 2013; પોટર એટ અલ., 2014).
કેઓઆર દ્વારા હાથ ધરવામાં આવેલા સાયકોસ્ટીમ્યુલેન્ટ્સના સેવનની સુવિધા ડ્રગના સંપર્કમાં સંબંધિત ટાઇમ વિંડો પર આધારિત હોવાનું લાગે છે. કોકિનના સંપર્કમાં આવતાં પહેલાં KOR એગોનિસ્ટ U50488 1 કલાકનો વહીવટ, બંને કોકેન સ્થાનની પસંદગીને સંભવિત કરે છે અને એનએસીમાં કોપેઇન દ્વારા સંબંધિત ડોપામાઇનને મુક્ત કરે છે, જ્યારે 15 મિનિટ પહેલાં આપવામાં આવે ત્યારે વિપરીત અસરો જોવા મળે છે (મેક્લોફ્લિન એટ અલ., 2006 એ; એહરિચ એટ અલ., 2014). ઇન્ટ્રાકાર્નિયલ સ્વ-ઉત્તેજનાનો ઉપયોગ કરવો ચાર્ટોફ એટ અલ. (2016) જોયું કે કેઓઆર એગોનિસ્ટ સાલ્વિનોરીન એ, પ્રારંભિક અવ્યવસ્થિત અને વિલંબિત લાભદાયી અસર ધરાવે છે, તેની સાથે અનુક્રમે એનએસીમાં ઉત્તેજિત ડોપામાઇન પ્રકાશનમાં ઘટાડો અને વધારો થયો છે. આ બધા ડેટા મળીને કોરના લાભદાયી ગુણધર્મો પર કેઓઆર સક્રિયકરણની સમય-આધારિત અસર સૂચવે છે, અને અનિવાર્ય ડ્રગની શોધના વિકાસ માટેના મુખ્ય ખેલાડી તરીકે તાણ-મધ્યસ્થી કેઓઆર સક્રિયકરણ તરફ ધ્યાન દોરે છે.
ક્વિનપાયરોલ-પ્રેરિત લોકમોટર સંવેદના અને અનિવાર્ય વર્તણૂક
D2R એગોનિસ્ટ, ક્વિનપાયરોલ સાથેના ઉંદરોમાં નિરીક્ષણ કરવામાં આવતા વર્તનથી ડોપામાઇન સિસ્ટમ સંવેદનશીલતા અને અનિવાર્યતાના પે inીમાં સામેલ છે તે તથ્યો મજબૂત બને છે. ટૂંકમાં, ડી 2 આર એ જી કપ્લ્ડ રીસેપ્ટર્સ છે જેનો પુરસ્કાર / પ્રેરણા સર્કિટમાં વ્યાપકપણે વ્યક્ત કરવામાં આવે છે; તેઓ સોમેટોડેન્ડ્રિટિકલી અને ડોપામાઇન ન્યુરોન્સના એક્સન ટર્મિનલ્સ પર વ્યક્ત થાય છે (સેસાક એટ અલ., 1994), અને તેના સક્રિયકરણથી ડોપામાઇન એક્સ્ટ્રા સેલ્યુલર સ્તરમાં ઘટાડો થાય છે (ઇમ્પ્રેટો અને દી ચીરા, 1988). સ્ટ્રાઇટમમાં, ડી 2 આર એ મધ્યમ સ્પાઇન ન્યુરોન્સ પર પણ પોસ્ટપ્લેસથી સ્થિત છે.સેસાક એટ અલ., 1994) અને તેનું સક્રિયકરણ એ આડકતરી માર્ગને અવરોધિત કરે છે જેમાં લોકમોટર પ્રવૃત્તિને મંજૂરી મળે છે.
ડ Dr.. હેનરી શેચેમેને 1980 ના દાયકાના અંતમાં ઉંદરોની વર્તણૂક પર ક્વિનપાયરોલના પ્રભાવોનો અભ્યાસ કરવાનું શરૂ કર્યું. તેમના પ્રારંભિક તારણોથી જાણવા મળ્યું કે ક્વિનપાયરોલનું તીવ્ર વહીવટ લોમમોટર પ્રવૃત્તિ પર ડોઝ-આધારિત અસર ધરાવે છે. ઓછા ડોઝ (0.03 મિલિગ્રામ / કિગ્રા) પર તે લોકોમોટર પ્રવૃત્તિમાં ઘટાડો કરે છે, જ્યારે વધારે ડોઝ (> 0.5 મિલિગ્રામ / કિગ્રા) માં, તે વધે છે. (ઇલામ અને શેચેમેન, 1989). આ અસરો અનુક્રમે હાઇ-એફિનીટી પ્રિસ્નેપ્ટિક ડી 2 આર અને લો-એફિનીટી પોસ્ટસિનેપ્ટિક ડી 2 આરના સક્રિયકરણ સાથે સંકળાયેલા છે (યુસીએલો એટ અલ., 2000). અણધારી રીતે, ક્વિનપાયરોલનું પુનરાવર્તિત (દર બીજા દિવસે) વહીવટ, લોકોમોશનમાં ધીમે ધીમે અને સતત વધારો પ્રેરિત કરે છે, જે મનોરોગ ઉત્તેજના દ્વારા પ્રેરિત લોમોમોટર સંવેદનાને મળતા આવે છે (સેશેટમેન એટ અલ., 1993; સેશેટમેન એટ અલ., 1994). લોકમોટર સંવેદનાત્મક અસર D2R પર આધારિત હોવાનું દર્શાવવામાં આવ્યું હતું, કેમ કે આ રીસેપ્ટરમાં ઉંદરની ઉણપ ક્વાનપાયરોલમાં લોકોમોટર સંવેદનાનો વિકાસ કરતી નથી (એસ્કોબાર એટ અલ., 2015).
90 ના દાયકાની શરૂઆતમાં, શેચેમેન અને આઈલમે અહેવાલ આપ્યો કે લોમોમોટર સંવેદનાની સાથે, ઉંદરોએ એક સ્ટીરિયોટાઇપ વર્તન વિકસાવી, જે ક્વિનપાયરોલના દરેક વહીવટથી મજબૂત બને છે (ઇલામ અને શેચેમેન, 1989; સેશેટમેન એટ અલ., 1993). આજે, ક્વિનપાયરોલનું પુનરાવર્તિત વહીવટ એ OCD માટે માન્ય મોડેલ છે (સેશેટમેન એટ અલ., 1999; સેશેટમેન એટ અલ., 2001; ઇલામ અને શેચેમેન, 2005; સ્ટુચલિક એટ અલ., 2016; સેશેટમેન એટ અલ., 2017), ઉંદરોની વર્તણૂક વધુને વધુ માળખાગત અને જટિલ બને છે તે અવલોકનના આધારે, અનિવાર્ય ચેકિંગ વર્તનની વિધિની વર્તણૂકની યાદ અપાવે છે (સેશેટમેન એટ અલ., 1998; સેશેટમેન એટ અલ., 2017). તાજેતરના અધ્યયનો દર્શાવે છે કે પુનરાવર્તિત ક્વિનપાયરોલ પણ ઉંદરમાં ફરજિયાત વર્તણૂકોને પ્રેરે છે, જેમ કે અનિવાર્ય તપાસ (સૂર્ય એટ અલ., 2019), વર્તણૂકની અસ્થિરતા અને અનિવાર્ય ચ્યુઇંગ (Asaoka એટ અલ., 2019), સ્ટ્રિઆટમમાં D2R નાકાબંધી દ્વારા બાદમાં ફેરવાયેલ, આગળની તરફેણ કરે છે કે અનિવાર્ય વર્તણૂકોને પ્રેરિત કરવા માટે વારંવાર D2Rs સક્રિયકરણ જરૂરી છે. એકસાથે ડેટા મિડબ્રેઇન ડોપામાઇન માર્ગોની અંદર ડી 2 આરની નિર્ણાયક ભૂમિકા તરફ દોરી જાય છે, જેમાં લોકોમોટર સંવેદના અને અનિવાર્યતાને પ્રેરિત કરે છે. પુનરાવર્તિત ક્વિનપાયરોલ એડમિનિસ્ટ્રેશન પ્રાઇમ્સ કોકેન-પ્રેરિત સ્ટીરિયોટાઇપ વર્તન (થોમ્પસન એટ અલ., 2010) અને એમ્ફેટેમાઇનની લોકોમોટર અસરો (કોપ એટ અલ., 2010), D2Rs એક્ટિવેશનને સાયકોસ્ટીમ્યુલેન્ટ-પ્રેરિત સંવેદનાને ધ્યાનમાં રાખીને અને ક્વિનપાયરોલ અને સાયકોસ્ટીમ્યુલેન્ટ્સ-પ્રેરિત સંવેદના વચ્ચેના વહેંચાયેલ મિકેનિઝમ સૂચવે છે તે વિચારને મજબૂત બનાવશે. રસપ્રદ વાત એ છે કે પુનરાવર્તિત ડી 2 આર સક્રિયકરણની સંવેદનાત્મક અસર મનોવૈજ્timાનિક તત્વો દ્વારા પ્રેરિત કરતા વધુ મજબૂત લાગે છે, કેમ કે ક્વિનપાયરોલ સાથે ઉપચાર કરવામાં આવતા દરેક ઉંદરો લોકોમોટર સંવેદના વિકસાવે છે (એસ્કોબાર એટ અલ., 2015), જ્યારે લગભગ સાઠ ટકા ઉંદરો એમ્ફેટેમાઇન પ્રત્યે સંવેદનશીલ હોય છે (એસ્કોબાર એટ અલ., 2012; કેસોનોવા એટ અલ., 2013).
ડી 2 આર ના વારંવાર સક્રિયકરણ દ્વારા પ્રેરિત વર્તણૂક સંવેદના, ઇનામ / પ્રેરણા સર્કિટમાં અનુકૂલન સાથે છે. ક્વિનપાયરોલ સાથે સંવેદનશીલ ઉંદરો, એનએસીમાં નીચલા ડોપામિનર્જિક સ્વર ધરાવે છે, જે મૂળભૂત ઘટાડો તરીકે જોવાય છે (કોલ્ટઝો એટ અલ., 2003) અને ઉત્તેજિત ટોનિક અને ફેસિક ડોપામાઇન પ્રકાશન (એસ્કોબાર એટ અલ., 2015), ડોપામાઇન મિડબ્રેઇન સર્કિટની ડોપામાઇન પ્રકાશન ક્ષમતામાં ઘટાડો સૂચવે છે. એનએસીમાં સિનેપ્ટિક ડોપામાઇન લેવલ, ડીએટી અને ડોપામાઇન ન્યુરોન્સ પ્રવૃત્તિ બંનેની પ્રવૃત્તિ દ્વારા નિયંત્રિત થાય છે (ગોટો અને ગ્રેસ, 2008), જે વિવો માં ટોનિક અને બર્સ્ટ ફાયરિંગનો સમાવેશ કરે છે (વિલ્સન એટ અલ., 1977; ગ્રેસ અને બૂની, 1980). પહેલાનાં અહેવાલો બતાવે છે કે ક્વિનપાયરોલ-સંવેદનાત્મક ઉંદરો, વી.ટી.એ. માં ટોનિક અને બર્સ્ટ ફાયરિંગમાં નાની સંખ્યામાં ડોપામાઇન ન્યુરોન્સ દર્શાવે છે (સેસિયા એટ અલ., 2013). આ ડેટા સાથે મળીને સૂચવે છે કે ક્વાનપાયરોલ સંવેદના પછી જોવા મળતા ડોપામાઇનના પ્રકાશનમાં ઘટાડો એ ડોપામાઇન ન્યુરોન્સની એકંદર પ્રવૃત્તિમાં ઘટાડોનું પરિણામ છે. ક્વિનપાયરોલ સાથે વારંવાર સારવાર દ્વારા પ્રેરિત અનિવાર્ય વર્તન અને સંવેદનાત્મક લોમોમોટર પ્રવૃત્તિ, એનએસીમાં ડોપામિનેર્જિક સ્વરમાં ઘટાડો થવાને કારણે, ડી 2 આરની સંવેદનાનું પરિણામ હોઈ શકે છે. ખરેખર, ક્વિનપાયરોલ-સંવેદનાત્મક ઉંદરો ડોપામાઇન ડી 2 આરના બંધનકર્તામાં વધારો દર્શાવે છે (કલ્વર એટ અલ., 2008) અને આ રીસેપ્ટર્સની જોડાણની સ્થિતિમાં વધારો (પેરેલaultલ્ટ એટ અલ., એક્સએન્યુએમએક્સ), આ પૂર્વધારણાને ટેકો આપે છે.
ક્વિનપાયરોલ-પ્રેરિત અનિવાર્ય વર્તણૂકોમાં કેઓઆર-ડોપામાઇન ક્રિયાપ્રતિક્રિયા
ડી 2 આર-પ્રેરિત અનિવાર્ય વર્તણૂકોમાં કેઓઆરની ભૂમિકા સંબંધિત પ્રારંભિક અભ્યાસ પણ શેચમેનની પ્રયોગશાળામાંથી આવ્યા હતા. આ જૂથે કેઓઆર એગોનિસ્ટ યુ 69593 ના સહવર્તી વહીવટની તપાસ લોનમોટર પ્રવૃત્તિ પર ક્વિનપાયરોલથી કરી. ખાસ કરીને, લેખકોએ 69593 થી 8 ઇન્જેક્શન પૂર્ણ ન થાય ત્યાં સુધી, U10 અને ક્વિનપાયરોલ મિશ્રણ સાથે ઉંદરોને સબક્યુટેનીયસ ઇન્જેક્શન્સ આપ્યા. એકલા યુ 69593 ની હાઈપોલોકોમોટર અસરથી વિપરીત, જ્યારે ક્વિનપાયરોલના નીચા (પ્રેઝિનેટicપ્ટિક) અને ઉચ્ચ (પોસ્ટસaptનaptપ્ટિક) ડોઝ સાથે એકસાથે સંચાલિત કરવામાં આવે ત્યારે હાઇપરલોકોમોશન જોવા મળ્યું. યુ 69593 એ ક્વિનપાયરોલના પ્રિસ્નેપ્ટિક ડોઝની હાયપોલોકોમોટર અસરને હાયપરલોકોમોશનમાં બદલી અને ક્વિનપાયરોલના પોસ્ટસિનાપ્ટિક ડોઝની હાયપરલોકોમોટર અસરમાં વધારો કર્યો (પેરેલaultલ્ટ એટ અલ., એક્સએન્યુએમએક્સ). કે.ઓ.આર.એસ.ના સહ-સક્રિયકરણથી પણ લોકોમોટર સેન્સેટાઇઝેશનના સમાવેશને વેગ મળ્યો અને ડી 2 આર સક્રિયકરણની અસરને સંભવિત કરવામાં આવી, કારણ કે ડબલ ટ્રીટમેન્ટ દ્વારા પ્રાપ્ત મહત્તમ લોમમોશન એકલા ક્વિનપાયરોલ દ્વારા પ્રેરિત લોકમોટર અસરની નકલ કરે છે (પેરેલaultલ્ટ એટ અલ., એક્સએન્યુએમએક્સ; એસ્કોબાર એટ અલ., 2017). કેઓઆરના સહ-સક્રિયકરણ, ફરજિયાત ચેકિંગ વર્તનના સંપાદનને પણ વેગ આપે છે (પેરેલaultલ્ટ એટ અલ., એક્સએન્યુએમએક્સ). ક્વોનપાયરોલ-પ્રેરિત વર્તણૂકો પર કેઓઆરની આ સંભવિત અસરો માટે KOR ની પુનરાવર્તિત સક્રિયકરણની જરૂર છે. હકીકતમાં, કેઓઆર એગોનિસ્ટ યુ 69593 ના તીવ્ર ઇન્જેક્શનમાં ક્વિનપાયરોલ સાથે સંવેદનાવાળી ઉંદરોમાં લોકોમોટર પ્રવૃત્તિમાં વધુ સુધારો થયો નથી (એસ્કોબાર એટ અલ., 2017). ડી.આર.આર.-પ્રેરિત સંવેદનાને સંભવિત કરવા માટે કે.ઓ.આર.ની પદ્ધતિ અજાણ છે. એક શક્યતા એ છે કે એન્ડોજેનસ કપ્પા ioપિઓઇડ સિસ્ટમ પોતે D2R- આધારિત સંવેદનામાં મધ્યસ્થી કરી રહી છે. જો કે, નોર્બીએનઆઈના પૂર્વ-વહીવટ દ્વારા, કpનપાયરોલમાં લોકોમોટર સંવેદનાને સુધારવામાં ન આવે તે બતાવીને આ શક્યતાને નકારી કા wasી હતી, સૂચવે છે કે ડાયનોર્ફિન ડાઉનસ્ટ્રીમ ડી 2 આર એક્ટિવેશન પ્રકાશિત નથી થતું (એસ્કોબાર એટ અલ., 2017). આ ડેટાને નકારી કા doesતો નથી કે ડાઈનોર્ફિન અનિવાર્ય વર્તણૂકોને સંવેદનામાં લાવવાની ભૂમિકા હોઈ શકે છે, ઉદાહરણ તરીકે, તાણ ડાયનોર્ફિનને મુક્ત કરવા અને કેઓઆરના સક્રિયકરણને પ્રેરે છે જે અનિવાર્ય વર્તણૂકોને સરળ બનાવે છે (મેકલોગલીન એટ અલ., 2003; મેક્લોફ્લિન એટ અલ., 2006 એ; મેક્લોફ્લિન એટ અલ., 2006 બી).
ડી 2 આર અને કેઓઆર વચ્ચેનો ક્રોસલેક જટિલ છે અને તે બંને રીસેપ્ટર્સનું સક્રિયકરણ યોગાનુયોગી છે કે અસ્થાયી રૂપે અલગ છે કે કેમ તેના પર નિર્ભર છે. એનાટોમિકલ ડેટા સૂચવે છે કે ડી 2 આર અને કેઓઆર વચ્ચેનો ક્રોસલેક પૂર્વનિર્ધારણ રીતે ચેતાક્ષમાં અને ડોપામાઇન ન્યુરોનના સોમામાં, તેમજ સ્ટ્રાઇટમના એમએસએસમાં પોસ્ટસિએપ્ટલી રીતે થઈ શકે છે. તેમ છતાં તે અન્ય ન્યુરોકેમિકલ સિસ્ટમ્સના ચેતાક્ષ પર સ્થિત કેઓઆરની ભૂમિકાને નકારી શકતો નથી, તેમ છતાં, એનાટોમિકલ ડેટા, ડી 2 આરને નિયંત્રિત કરતી KORs ની સીધી ભૂમિકા તરફ ધ્યાન આપે છે. ક્યાં તો તીવ્ર અથવા પુનરાવર્તિત, કેઓઆરનું સક્રિયકરણ ડોપામાઇન ન્યુરોન્સ પર અવરોધક ડી 2 આર કાર્ય ઘટાડે છે. ઇલેક્ટ્રોફિઝિયોલોજી અધ્યયન દર્શાવે છે કે વીટીએ અને એસ.એન.ના ડોપામાઇન ન્યુરોન્સમાં કેઓઆરની તીવ્ર સક્રિયતા, ડી 2 આર-મધ્યસ્થી અવરોધક પોસ્ટસિનાપ્ટિક વર્તમાનને અટકાવે છે, કેઆરએલ ડોપામિન પ્રકાશનમાં ઘટાડો કરે છે અને ડાયનોર્ફિન અવરોધિત ડોપામાઇનના અવરોધક અસરને અવરોધે છે.ફોર્ડ એટ અલ., 2007). ન્યુરોકેમિકલ અધ્યયનોએ દર્શાવ્યું હતું કે કેઓઆર્સના પુનરાવર્તિત સક્રિયકરણ, એનએસીમાં ડોપામાઇન પ્રકાશનને અવરોધિત ડી 2 આર-પ્રેરિત અવરોધ છે (ફુએન્ટેઆલ્બા એટ અલ., 2006). તદુપરાંત, સંયોગી ડી 2 આર અને કેઓઆર તીવ્ર સક્રિયકરણ એકલા દરેક રીસેપ્ટરની અસરની તુલનાએ એનએસીમાં ડોપામાઇન પ્રકાશનનું અવરોધ ઘટાડે છે (એસ્કોબાર એટ અલ., 2017). આમ, પ્રિઝેનેપ્ટિક કેઓઆરઓ પ્રેઝિનેપ્ટિક ડી 2 આર સાથે, verseલટું, કેઓઆર ક્યાં તો અવરોધે છે અથવા ડી 2 આર અવરોધક અસરને બાકાત રાખે છે. આ મિકેનિઝમ, ક્વોનપાયરોલની ઓછી માત્રા સાથે સુસંગત KOR એગોનિસ્ટ્સની તીવ્ર માત્રાના લોકોમોટર સક્રિયકરણ અસરને સમજાવી શકે છે.પેરેલaultલ્ટ એટ અલ., એક્સએન્યુએમએક્સ).
તાજેતરના એક અભ્યાસ બતાવે છે કે વીટીએમાં કેઓઆર સક્રિયકરણ વર્તણૂક અવરોધ અને આરસની દફન તરીકે માપવામાં આવતી અનિવાર્ય વર્તનને મધ્યસ્થ કરે છે (અબ્રાહમ એટ અલ., 2017), KORs સક્રિયકરણ એ અનિવાર્યતા માટે ખરેખર ટ્રિગર છે તે વિચારને મજબૂત બનાવવી. દ્વારા પ્રકાશિત ડેટા માર્ગોલીસ એટ અલ. (2006); 2008) સૂચવે છે કે એમ.પી.એફ.સી. ને લક્ષ્ય બનાવતા ડોપામાઇન ન્યુરોન્સ પર કેઓઆર અને ડી 2 આર ક્રિયાપ્રતિક્રિયા થવી જોઈએ (માર્ગોલિસ એટ અલ., 2006; માર્ગોલિસ એટ અલ., 2008). તેમ છતાં, ફોર્ડ એટ અલ. (2006); 2007) શોધી કા that્યું કે ડી 2 આર મધ્યસ્થી આઇપીએસસીનું કેઓઆર નિષેધ એ એનએસીને લક્ષ્યાંકિત કરતા ડોપામાઇન ન્યુરોન્સ પર થાય છે (ફોર્ડ એટ અલ., 2006; ફોર્ડ એટ અલ., 2007). આ ડેટા સાથે મળીને બતાવે છે કે ડોપામાઇન ન્યુરોન્સના સોમાટોોડેન્ડ્રિટિક ડબ્બામાં ડી 2 આર સાથે કેઓઆર ક્રિયાપ્રતિક્રિયા એ જ ડોપામાઇન ન્યુરોનમાં ક્રોસસ્ટkકના પરિણામે couldભી થઈ શકે છે. આ મેસોલીમ્બિક અથવા મેસોકોર્ટિકલ અંદાજોમાં થાય છે તે હજી વિવાદાસ્પદ છે.
નોંધપાત્ર રીતે, કે.એ.આર. એન.એ.સી. ના એમ.એસ.એન. માં મળી આવ્યા હતા (એસ્કોબાર એટ અલ., 2017; તેજેડા એટ અલ., 2017), આમ સૂચવે છે કે ડોપામાઇન ન્યુરોન્સના લક્ષ્ય કોષો પર સીધી ક્રિયાઓ દ્વારા D2R- પ્રેરિત અનિવાર્ય વર્તનની શક્તિ પણ canભી થઈ શકે છે. આ સંદર્ભમાં, તે ઉલ્લેખનીય છે કે યુ 69593 ના વારંવારના વહીવટથી ઉચ્ચ જોડાણની સ્થિતિમાં D2R ની માત્રામાં વધારો થાય છે (પેરેલaultલ્ટ એટ અલ., એક્સએન્યુએમએક્સ). ન્યુરોકેમિકલ ડેટા સૂચવે છે કે ડોપામાઇન એક્સ્ટ્રા સેલ્યુલર સ્તરમાં ઘટાડો એ ડી 2 આર સંવેદના સાથે સંકળાયેલ છે. કેઓઆરએસના સહ-સક્રિયકરણમાં એનએસીમાં ડોપામાઇનના બાહ્યકોષીય સ્તરમાં વધુ ઘટાડો થતો નથી, જે પહેલાથી જ D2R ની વારંવાર સક્રિયતા દ્વારા ઘટાડો થયો છે (એસ્કોબાર એટ અલ., 2017), આ મિકેનિઝમ દ્વારા એનએસીમાં ડી 2 આરની સંવેદનાને વેગ આપવા અથવા સંભવિત બનાવવાની પ્રેસિનેપ્ટિક કેઓઆરની ભૂમિકાને નકારી કા .ો. તેથી, કેઓઆરએસ ધીમી પરમાણુ મિકેનિઝમ્સને ટ્રિગર કરે છે જે ડી 2 આર ની ન્યુરોસાયકલ અને વર્તણૂકીય અસરોને વધુ સંવેદનશીલ બનાવે છે, સૂચવે છે કે લોમમોશન સેન્સિટાઇઝેશન વૃદ્ધિ કોઈ પ્રિસ્નેપ્ટીક અસરને બદલે અનુકૂલનશીલ પોસ્ટસિનેપ્ટિકને કારણે હોઈ શકે છે. આ સંદર્ભમાં કેઓઆરની વારંવાર સક્રિયતા D2R પરોક્ષ સ્ટ્રિએટલ પાથવે D1R / D2R સંતુલનને D1R પર અનિર્ણિત કરનારી અનિવાર્યતાને અટકાવી શકે છે (આકૃતિ 1).
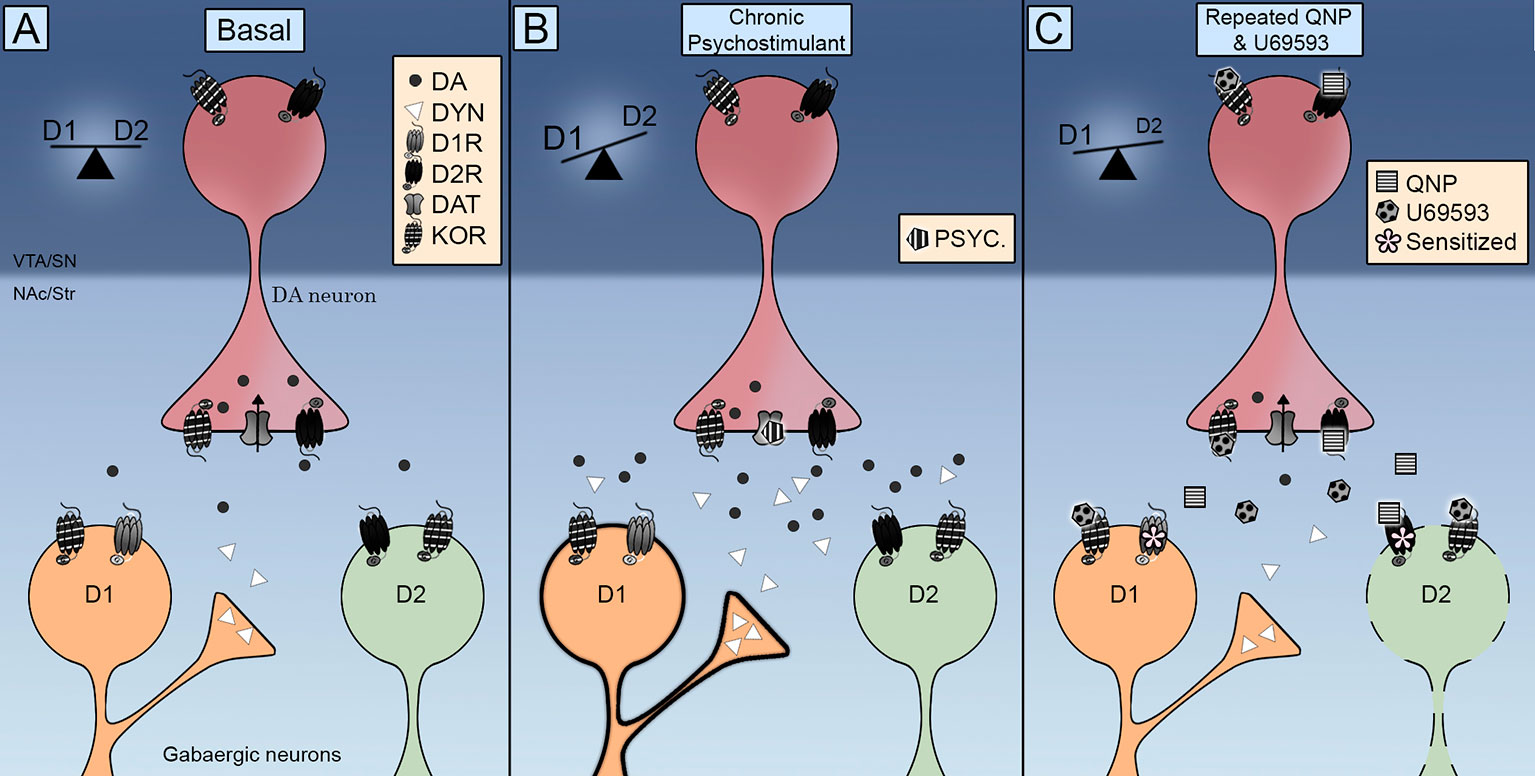 આકૃતિ 1 ડાયરેક્ટર (ડી 1 આર) અને આડકતરી (ડી 2 આર) સ્ટ્રિએટલ ફાટવે પર કપ્પા ઓપિઓઇડ રીસેપ્ટર્સ (કેઓઆર) નિયંત્રણની એકીકૃત યોજના. (એ) કેઓઆર ડોપામાઇન ટર્મિનલ્સ પર પૂર્વ-સિનાપ્ટિટિકલી સ્થિત છે અને મધ્યમ કદના ન્યુરોન્સ (એમએસએન) માં સિનાપ્ટિકલી પોસ્ટ કરે છે. તેનું સક્રિયકરણ ડોપામાઇન એક્સ્ટ્રા સેલ્યુલર સ્તરને નિયંત્રિત કરે છે અને તેનું સ્થાનિકીકરણ ડોપામાઇન ટ્રાન્સપોર્ટર (DAT) અને ડોપામાઇન ડી 2 રીસેપ્ટર્સ સાથેના સંપર્કને પ્રોત્સાહન આપે છે. (બી) સાયકોસ્ટીમ્યુલેન્ટમાં વારંવાર સંપર્કમાં આવતા ડોપામાઇન એક્સ્ટ્રા સેલ્યુલર સ્તર અને ડાયનોર્ફિન બંનેમાં વધારો થાય છે. ડી 1 અને ડી 2 રીસેપ્ટર્સનું સક્રિયકરણ એ સંતુલનને ડી 1 આર ડાયરેક્ટ પાથવે પર ફેરવે છે જે લોકમોટર સંવેદનાને પ્રોત્સાહન આપે છે. (સી) ક્વિનપાયરોલ અને યુ 69593 નો સહ-વહીવટ ડોપામાઇન એક્સ્ટ્રા સેલ્યુલર સ્તરમાં ઘટાડો સાથે છે. કે.ઓ.આર. અને ડી 2 રીસેપ્ટર્સની સાથે મળીને સક્રિયકરણ ડી 2 અપ્રત્યક્ષ માર્ગને અનિવાર્ય વર્તણૂકનું નિર્માણ કરે છે.
આકૃતિ 1 ડાયરેક્ટર (ડી 1 આર) અને આડકતરી (ડી 2 આર) સ્ટ્રિએટલ ફાટવે પર કપ્પા ઓપિઓઇડ રીસેપ્ટર્સ (કેઓઆર) નિયંત્રણની એકીકૃત યોજના. (એ) કેઓઆર ડોપામાઇન ટર્મિનલ્સ પર પૂર્વ-સિનાપ્ટિટિકલી સ્થિત છે અને મધ્યમ કદના ન્યુરોન્સ (એમએસએન) માં સિનાપ્ટિકલી પોસ્ટ કરે છે. તેનું સક્રિયકરણ ડોપામાઇન એક્સ્ટ્રા સેલ્યુલર સ્તરને નિયંત્રિત કરે છે અને તેનું સ્થાનિકીકરણ ડોપામાઇન ટ્રાન્સપોર્ટર (DAT) અને ડોપામાઇન ડી 2 રીસેપ્ટર્સ સાથેના સંપર્કને પ્રોત્સાહન આપે છે. (બી) સાયકોસ્ટીમ્યુલેન્ટમાં વારંવાર સંપર્કમાં આવતા ડોપામાઇન એક્સ્ટ્રા સેલ્યુલર સ્તર અને ડાયનોર્ફિન બંનેમાં વધારો થાય છે. ડી 1 અને ડી 2 રીસેપ્ટર્સનું સક્રિયકરણ એ સંતુલનને ડી 1 આર ડાયરેક્ટ પાથવે પર ફેરવે છે જે લોકમોટર સંવેદનાને પ્રોત્સાહન આપે છે. (સી) ક્વિનપાયરોલ અને યુ 69593 નો સહ-વહીવટ ડોપામાઇન એક્સ્ટ્રા સેલ્યુલર સ્તરમાં ઘટાડો સાથે છે. કે.ઓ.આર. અને ડી 2 રીસેપ્ટર્સની સાથે મળીને સક્રિયકરણ ડી 2 અપ્રત્યક્ષ માર્ગને અનિવાર્ય વર્તણૂકનું નિર્માણ કરે છે.
અનિવાર્ય વર્તણૂકોમાં કેઓઆર-ડોપામાઇન ક્રિયાપ્રતિક્રિયાના લૈંગિક તફાવતો
ક્લિનિકલ અધ્યયનોએ અનિવાર્ય દવાઓની શોધ સહિત અનિવાર્ય વર્તનમાં જાતીય તફાવતો દર્શાવ્યા છે. સ્ત્રીઓની તુલનામાં પુરુષોમાં OCD લક્ષણોની શરૂઆતની શરૂઆત જોવા મળે છે (મેથિસ એટ અલ., 2011), સ્ત્રીઓ દૂષિતતા અને સફાઇના લક્ષણોનું વધુ પ્રમાણ દર્શાવે છે.લાબાડ એટ અલ., 2008). માદક પદાર્થના વ્યસનમાં લૈંગિક તફાવતો અંગે, ક્લિનિકલ પુરાવા સૂચવે છે કે પુરુષોમાં ડ્રગનો ઉપયોગ વધુ પ્રચલિત છે, જ્યારે સ્ત્રીઓ અનિવાર્ય ડ્રગની શોધમાં પુરુષો કરતા ઝડપી પ્રગતિ દર્શાવે છે (હર્નાન્ડેઝ-એવિલા એટ અલ., 2004; ફટ્ટોર અને મેલિસ, 2016).
હમણાં હમણાં, પૂર્વ-ક્લિનિકલ પુરાવાએ ક્લિનિકલ અધ્યયનમાં જોવા મળેલા ડ્રગના દુરૂપયોગમાં લૈંગિક તફાવતને ધ્યાનમાં રાખતા ન્યુરોબાયોલોજીકલ પાયાને ભારપૂર્વક પ્રકાશિત કર્યા છે (બેકર અને ચાર્ટોફ, 2019). નો-નેટ ફ્લક્સ માઇક્રોડિઆલિસિસનો ઉપયોગ કરીને પ્રારંભિક અવલોકનોએ દર્શાવ્યું હતું કે ડોર્સિન સ્ટ્રાઇટમમાં ડોપામાઇન એક્સ્ટ્રાસેલ્યુલર એકાગ્રતા ડાયેટ્રસની તુલનામાં પ્રોસ્ટ્રસ અને એસ્ટ્રસમાં ઉચ્ચ સ્તર સાથેના એસ્ટ્રસ ચક્ર દરમિયાન બદલાય છે. તદુપરાંત, જ્યારે ovariectomy સ્ત્રી ઉંદરોમાં સ્ટ્રાઇટલ ડોપામાઇન એક્સ્ટ્રાસેલ્યુલર સાંદ્રતામાં ઘટાડો કરે છે, ત્યારે પુરુષ ઉંદરોના અવકાશીકરણ ડોપામાઇન સ્ટ્રાઇટલ એક્સ્ટ્રા સેલ્યુલર એકાગ્રતામાં ફેરફાર કરતા નથી (ઝિઓ અને બેકર, 1994), ડોપામાઇન પ્રવૃત્તિ પર અંડાશયના હોર્મોન્સની મહત્વપૂર્ણ ભૂમિકા સૂચવતા. આ ઉપરાંત, સ્ત્રી હોર્મોન્સ સાયકોસ્ટીમ્યુલેન્ટ્સના પ્રતિભાવને નિયંત્રિત કરે છે. વહેલી ઈન વિટ્રો સ્થિતિએ પ્રયોગો દર્શાવે છે કે એસ્ટ્રાડિયોલ પ્લસ પ્રોજેસ્ટેરોન અંડાશયનાશક સ્ત્રી ઉંદરો દ્વારા પ્રાપ્ત સ્ટ્રિએટલ પેશીમાંથી એમ્ફેટેમાઇન-પ્રેરિત ડોપામાઇન પ્રકાશનને પુનર્સ્થાપિત કરે છે (બેકર અને રામિરેઝ, 1981). તાજેતરમાં જ, ઝડપી સ્કેન ચક્રીય વોલ્ટેમેટ્રી અભ્યાસ દર્શાવે છે કે સ્ત્રીઓ પુરુષોની તુલનામાં વધુ વિદ્યુત-ઉત્તેજિત ડોપામાઇન પ્રકાશન અને ઉપભોગ દર્શાવે છે (વોકર એટ અલ., 2000). ડોપામાઇન ન્યુરોટ્રાન્સમિશનમાં આ લૈંગિક તફાવત સ્ત્રીઓમાં જોવાયેલી cંચી કોકેન અને એમ્ફેટેમાઇન માટેનો હિસ્સો હોઈ શકે છે. (રોબર્ટ્સ એટ અલ., 1989; કોક્સ એટ અલ., 2013).
ડોપામાઇન એક્સ્ટ્રા સેલ્યુલર સ્તરો પર કેઓઆરનું નિયમન પણ જાતીય તફાવત બતાવે છે (ચાર્ટોફ અને માવરિકાકી, 2015). ઇન્ટ્રાકાર્નિયલ સ્વ-ઉત્તેજના અને ચક્રીય વોલ્ટેમેટ્રીનો ઉપયોગ કરીને, કોનવે એટ અલ. (2019) પુરુષ ઉંદરોની તુલનામાં સ્ત્રી ઉંદરોમાં જોવા મળતા KOR એગોનિસ્ટની તીવ્ર એનેહેડોનિક અસરની ઓછી સંવેદનશીલતા, એનએસીમાં ઉત્તેજીત ડોપામાઇન પ્રકાશનની તીવ્ર અવરોધ સાથે છે (કોનવે એટ અલ., 2019). એવું સૂચવવામાં આવ્યું છે કે એ.એસ.ટી.ઓ.આર. (KOR) સક્રિયકરણ પછી સ્ત્રી ઉંદરોમાં જોવા મળતા ડોપામાઇનના પ્રકાશનના નિષ્ફળ અવરોધમાં ફાળો આપે છે.અબ્રાહમ એટ અલ., 2018). જ્યારે કેઓઆર અને ડોપામાઇન સિગ્નલિંગ વચ્ચેનો ક્રોસલેકનો અભ્યાસ પુરુષોમાં કરવામાં આવ્યો છે (તેજદા અને બોંસી, 2019), આ ક્રિયાપ્રતિક્રિયા પર સંશોધન અને સ્ત્રીઓમાં વ્યસન પ્રક્રિયામાં તેના પ્રભાવનો અભાવ છે (ચાર્ટોફ અને માવરિકાકી, 2015). સ્ત્રી ઉંદરોમાં, કેઓઆર એગોનિસ્ટ યુ 69593 ના તીવ્ર વહીવટ, નિયંત્રણ અને અંડાશયના ઉંદરો બંનેમાં કોકેન-પ્રેરિત હાયપરલોકોમોશનને આકર્ષિત કરે છે. રસપ્રદ વાત એ છે કે, U69593 પુનરાવર્તિત વહીવટ એસ્ટ્રાડિયોલ-આશ્રિત રીતે કોકેન-પ્રેરિત હાઇપરલોકોમેશનને નબળી પાડે છે (પુગ-રામોસ એટ અલ., 2008). આ ડેટા સૂચવે છે કે સ્ત્રી ઉંદરોમાં એસ્ટ્રાડીયોલ પ્રિ.ઓ.ઓ.આર. ક્રિયાઓ કરે છે, એક અસર જે તણાવના પ્રતિભાવમાં લૈંગિક તફાવતોથી સંબંધિત હોઈ શકે છે (પુગ-રામોસ એટ અલ., 2008). સ્ત્રી ઉંદરોમાં કેઓઆરનું વારંવાર સક્રિયકરણ પુરુષમાં જોવા મળેલ સ્ટ્રાઇટલ ડોપામાઇન પ્રકાશનની સુવિધા આપે છે તે અનુત્તરિત પ્રશ્ન છે.
પુરૂષ ઉંદરોની તુલનામાં સ્ત્રીમાં મનોવિરોધક પ્રેરિત ડોપામાઇન પ્રકાશનની સુવિધા જોવામાં આવે છે, તેમ છતાં, એમ્ફેટામાઇન લોમોમોટર સેન્સેટાઇઝેશન હેઠળના ડોપામાઇન મિકેનિઝમ્સમાં લૈંગિક તફાવત સંપૂર્ણ રીતે સ્પષ્ટ કરાયા નથી (બેકર, 1999). એમ્ફેટામાઇનમાં વારંવાર સંપર્કમાં, કિશોર વયે બંનેમાં વધુને વધુ મોટું પ્રવૃત્તિ પ્રેરિત કરે છે (મેથ્યુસ અને મેકકોર્મિક, 2007) અને પુખ્ત સ્ત્રી ઉંદરો (માઇલેસી-હોલી એટ અલ., 2007), એમ્ફેટામાઇનના વારંવાર સંપર્કમાં આવ્યા પછી સ્ત્રી કિશોરવયના ઉંદરો વધુ મજબૂત લોકોમોટર સંવેદના દર્શાવે છે. ડી 2 રીસેપ્ટરના નવજાત સક્રિયકરણ માત્ર સ્ત્રી ઉંદરોમાં એમ્ફેટામાઇન પ્રેરણાત્મક વર્તણૂકીય સંવેદનાને બળવાન બનાવે છે (બ્રાઉન એટ અલ., 2011). અગાઉ જણાવ્યા મુજબ, પુરૂષ ઉંદરોમાં તે જોવા મળ્યું છે કે ડી 2 એગોનિસ્ટના વારંવાર સંપર્કમાં લોકોમોટર સંવેદના અને અનિવાર્ય જેવી વર્તણૂક પ્રેરિત કરે છે (ડ્વોર્કિન એટ અલ., 2006). તદુપરાંત, કે.ઓ.આર. ની સહ-સક્રિયકરણ, ક્વિનપાયરોલના વારંવાર સંપર્કમાં આવવાથી લોમમોટર સંવેદનાને સંભવિત કરે છે, એનએસીમાં ડી.એ. રિલીઝ પર ડી 2 રીસેપ્ટર્સના અવરોધક નિયંત્રણની સુવિધા આપે છે (એસ્કોબાર એટ અલ., 2017). સ્ત્રીઓમાં ડોપામાઇન પ્રકાશન પર કેઓઆરની અવરોધક અસરની અવલોકન ઓછી સંવેદનશીલતા જેવા લિંગ તફાવત (કોનવે એટ અલ., 2019) અનિવાર્ય દવાઓની શોધમાં કેઓઆરના વિભિન્ન યોગદાન માટેનો હિસાબ કરી શકે છે.
નિષ્કર્ષ
પ્રોત્સાહિત વર્તણૂકોને વિસ્તૃત કરવા માટે કે.ઓ.આર. ડોપામાઇન સિગ્નલને કેવી રીતે મોડ્યુલેટ કરે છે અને તે સંવેદનશીલ અનિવાર્ય વર્તનનું પરિણામ ક્યારે આવે છે? એનાટોમિકલ ડેટા બતાવે છે કે મિડબ્રેઇન ડોપામાઇન ન્યુરોન્સની સિનેપ્ટિક પ્રવૃત્તિને નિયંત્રિત કરવા માટે કેઓઆર ઉત્કૃષ્ટપણે સ્થિત છે. કાર્યાત્મક ડેટા સૂચવે છે કે કેઓઆરએસ DAT અને D2R ની કામગીરી તેમજ ડોપામાઇન ન્યુરોન્સ ફાયરિંગ રેટને નિયંત્રિત કરે છે. પ્રારંભિક પુરાવા દર્શાવે છે કે કેઓઆરની તીવ્ર સક્રિયતા દુરૂપયોગની દવાઓ દ્વારા પ્રેરિત ડોપામાઇન પ્રકાશનમાં ઘટાડો થાય છે તે ડેટા સાથે પૂરક કરવામાં આવ્યું છે જે દર્શાવે છે કે કે.ઓ.આર. ની વારંવાર સક્રિયતા ડોપામાઇન પ્રકાશન અને ફરજિયાત દવા શોધવાની સુવિધા આપે છે. સ્ટ્રાઇટલ વિસ્તારોમાંથી ડોપામાઇન સિગ્નલિંગ બેલેન્સ ડાયરેક્ટ અને પરોક્ષ આઉટપુટ માર્ગો (આકૃતિ 1A). ક્યાં તો સાયકોસ્ટીમ્યુલેન્ટ્સ સાથે ક્રોનિક ઉત્તેજના કે જે ડોપામાઇન પ્રકાશનમાં વધારો કરે છે D1R અને D2R બંનેને સક્રિય કરે છે (આકૃતિ 1B) અથવા ક્વિનપાયરોલ જે ફક્ત D2R ને સક્રિય કરે છે (આકૃતિ 1C) નબળા D2R પરોક્ષ માર્ગ દ્વારા લોકોમોટર સંવેદના અને અનિવાર્ય વર્તણૂંકમાં પરિણમે છે, આમ સંતુલનને ડી 1 આર સીધા માર્ગ પર ફેરવે છે. સ્ટ્રિએટલ ડી 1 ન્યુરોન્સમાં ડાયનોર્ફિનના વધારા દ્વારા ક્રોનિક સાયકોસ્ટીમ્યુલેન્ટ ઇન્ટેક દરમિયાન કેઓઆર ટ્રાન્સમિશન વધારવામાં આવે છે (આકૃતિ 1B). ઉન્નત કેઓઆર ટ્રાન્સમિશન U69593 વહીવટ દ્વારા OCD ના ફાર્માકોલોજીકલ મોડેલમાં અનુકરણ કરવામાં આવે છે. આ સાથોસાથ કેઓઆર સક્રિયકરણ D2 પરોક્ષ માર્ગને વધુ નબળા બનાવે છે (આકૃતિ 1C). ડીએટી પ્રવૃત્તિ પરના કે.ઓ.આર.ના સક્રિયકરણના પરિણામોને સંપૂર્ણ રીતે સ્પષ્ટ કરવા, ક્વિનપાયરોલ પ્રેરિત અનિવાર્યતામાં અંતર્ગત કousર સિસ્ટમની ભૂમિકાને સમજવા અને અનિવાર્ય વર્તણૂકમાં જોવા મળેલી જાતિ વિભિન્નતામાં કે.ઓ.આર. સિસ્ટમનો ફાળો નક્કી કરવા માટે ભાવિ સંશોધન હાથ ધરવું જોઈએ.
લેખક ફાળો
હસ્તપ્રતની વિભાવનામાં એ.ઇ., એમ.એ અને જે.એફ.એ ફાળો આપ્યો. એ.એ અને જે.એફ.એ એમ.એ. ના ઇનપુટ સાથે હસ્તપ્રતનો પહેલો ડ્રાફ્ટ લખ્યો. એમ.એ. અને જે.સી.એ હસ્તપ્રતની ટીકાત્મક સમીક્ષા અને સંપાદન કરવામાં ફાળો આપ્યો. બધા લેખકોએ તેને પ્રકાશન માટે મંજૂરી આપી.
ભંડોળ
આ સમીક્ષામાં ટાંકવામાં આવેલા લેખકોના કામને ફOંડસીવાયટી ગ્રાન્ટ નંબરો: 1110352 અને 1150200 થી એમએ દ્વારા સમર્થન આપવામાં આવ્યું છે; 1141088 થી જેએફ; ડીપોજીએ જેએફને 391340281 ગ્રાન્ટ; જેસીથી ફ3170497ન્ડેસાઇટ પોસ્ટડોકટરલ સાથી 3190843 અને એઇને XNUMX.
રસ સંઘર્ષ
લેખકો જાહેર કરે છે કે આ સંશોધન કોઈ વ્યાપારી અથવા નાણાકીય સંબંધોની ગેરહાજરીમાં હાથ ધરવામાં આવ્યો હતો જેને સંભવિત રૂચિના સંઘર્ષ તરીકે ગણવામાં આવી શકે છે.
હેન્ડલિંગ સંપાદક હાલમાં લેખકો જે.એફ. સાથે સંશોધન વિષયનું આયોજન કરી રહ્યું છે, અને અન્ય કોઈ સહયોગની ગેરહાજરીની પુષ્ટિ આપે છે.
સંદર્ભ
અબ્રાહમ, એડી, ફontન્ટાઇન, એચએમ, સોંગ, એજે, એન્ડ્ર્યૂઝ, એમએમ, બેર્ડ, એમએ, કાઇફર, બીએલ, એટ અલ. (2017). ડોપામાઇન ન્યુરોન્સમાં કપ્પા ioપિઓઇડ રીસેપ્ટર સક્રિયકરણ વર્તણૂક અવરોધમાં ખલેલ પહોંચાડે છે. ન્યુરોસાયકોફોર્માકોલોજી 43 (2), 362–372. doi: 10.1038 / npp.2017.133
અબ્રાહમ, એડી, શેટ્ટાઉઅર, એસએસ, રેચાર્ડ, કેએલ, કોહેન, જેએચ, ફોન્ટાઇન, એચએમ, સોંગ, એજે, એટ અલ. (2018). જીઆરકે 2 નું એસ્ટ્રોજન રેગ્યુલેશન કપ્પા ioપિઓઇડ રીસેપ્ટરને ઇન્ટિએટ કરે છે મેડિએટિંગ એનલજેસીયા, પરંતુ અવગણના નહીં. જે ન્યુરોસી. 38 (37), 8031-8043. doi: 10.1523 / JNEUROSCI.0653-18.2018
અસૌકા, એન., નિશીતાની, એન., કિનોશિતા, એચ., નાગાઈ, વાય., હટકામા, એચ., નાગાયસુ, કે., એટ અલ. (2019) એડેનોસિન એ 2 એ રીસેપ્ટર વિરોધી પુનરાવર્તિત ક્વિનપાયરોલ-પ્રેરિત માનસિકતાના બહુવિધ લક્ષણોમાં સુધારો કરે છે. ઈન્યુરો 6 (1), 1–16. ENEURO.0366-18.2019. doi: 10.1523 / ENEURO.0366-18.2019
અટીગારી, ડીવી, ઉપ્રેટી, આર., પેસ્ટર્નક, જીડબ્લ્યુ, મજુમદાર, એસ., કિવેલ, બીએમ (2019) એમપી 1104, મિશ્રિત કપ્પા-ડેલ્ટા ioપિઓઇડ રીસેપ્ટર એગોનિસ્ટમાં ઉંદરોમાં ઓછી આડઅસરોવાળી એન્ટી-કોકેન ગુણધર્મો છે. ન્યુરોફર્મકોલોજી 150, 217-228. ડોઇ: 10.1016 / જે. ન્યુરોફાર્મ.2019.02.010
એઝોકર, વી.એચ., સેપ્લેવેદ, જી., રુઇઝ, સી., Ileગુઇલેરા, સી., Éન્ડ્રેસ, એમઇ, ફુએન્ટેઆલ્બા, જેએ (2019). કપ્પા-sensપિઓઇડ રીસેપ્ટરને અવરોધિત કરવું એમ્ફેટેમાઇન સેન્સેટીનાઇઝેશન દરમિયાન ડોર્સોટલલ સ્ટ્રાઇટમ ડોપામાઇન ગતિશીલતામાં ફેરફારને વિરુદ્ધ બનાવે છે. જે. ન્યુરોકેમ 148, 348–358. doi: 10.1111 / jnc.14612
બ્યુગિન, સી., પોટુઝાક, જે., ઝુ, ડબલ્યુ., લિયુ-ચેન, એલવાય, સ્ટ્રેઇચર, જેએમ, ગ્રોઅર, સીઈ, એટ અલ. (2012). 12-એપિ-સ salલ્વિનોરીન એ અને તેના એનાલોગ્સના કપ્પા ioપિઓઇડ રીસેપ્ટર પર વિશિષ્ટ સંકેત ગુણધર્મો. બાયોર્ગ. મેડ. રસાયણ. લેટ. 15; 22 (2), 1023–1026. doi: 10.1016 / j.bmcl.2011.11.128
બેર્ડસ્લે, પીએમ, હોવર્ડ, જેએલ, શેલ્ટન, કેએલ, કેરોલ, એફઆઇ (2005) નવલકથા કાપ્પા ioપિઓઇડ રીસેપ્ટર વિરોધી, જેડીટીક, વિશિષ્ટ અસરો, ફુલોના શોક તનાવ વિ, કોકેન પ્રાઈમ્સ અને તેના ઉંદરોમાં એન્ટીડિપ્રેસન્ટ જેવી અસરો દ્વારા પ્રેરિત કોકેન-શોધવાની પુન seekingસ્થાપન પર. સાયકોફાર્માકોલ. (બર્લ) 183, 118–126. doi: 10.1007/s00213-005-0167-4
બેકર, જેબી, ચાર્ટોફ, ઇ. (2019) ઇનામ અને વ્યસનની મધ્યસ્થતા કરતી ન્યુરલ મિકેનિઝમ્સમાં લૈંગિક તફાવત. ન્યુરોસાયકોફોર્માકોલોજી 44, 166–183. doi: 10.1038/s41386-018-0125-6
બેકર, જેબી, રેમિરેઝ, વીડી (1981) એમ્ફિટામાઇનમાં લિંગ તફાવત, વિટ્રોમાં ઉંદરો સ્ટ્રાઇટલ પેશીમાંથી કેટેકોલેમિન્સના પ્રકાશનને ઉત્તેજિત કરે છે. મગજનો અનાદર 204, 361–372. doi: 10.1016/0006-8993(81)90595-3
બેકર, જેબી (1999) સ્ટ્રાઇટમ અને ન્યુક્લિયસ એકમ્બેન્સમાં ડોપામિનર્જિક ફંક્શનમાં લિંગ તફાવત. ફાર્માકોલ બાયોકેમ પાછળ 64, 803–812. doi: 10.1016/S0091-3057(99)00168-9
બેરીજ, કેસી, રોબિન્સન, ટીઇ (2016). ગમવું, ઇચ્છવું અને વ્યસનની પ્રોત્સાહન-સંવેદનાનો સિદ્ધાંત. એમ. મનોવિજ્ઞાન. 71, 670-679. doi: 10.1037 / amp0000059
બેરીજ, કેસી, વેનિયર, આઈએલ, રોબિન્સન, ટીઇ (1989) 6-હાઇડ્રોક્સિડોપેમાઇન-પ્રેરિત અફેગિયાના સ્વાદની પ્રતિક્રિયાશીલતા વિશ્લેષણ: ડોપામાઇનના કાર્યના ઉત્તેજના અને એનેહોડિયા પૂર્વધારણા માટે સૂચિતાર્થ. બિહાવ ન્યુરોસી. 103, 36-45. ડોઇ: 10.1037 / 0735-7044.103.1.36
બ્રોડબિયર, જેએચ, નેગસ, એસએસ, બુટલેમેન, ઇઆર, ડી કોસ્ટા, બીઆર, વુડ્સ, જેએચ (1994). પદ્ધતિસર સંચાલિત ન norન-બાયનોલ્ટોર્ફિમાઇન (ન-બીએનઆઈ) ની વિશિષ્ટ અસરો મ mouseસ રિથિંગ એસિમાં કપ્પા-ioપિઓઇડ એડોનિસ્ટ્સ પર. સાયકોફોર્માકોલોજી 115, 311-319. ડોઇ: 10.1007 / BF02245071
બ્રાઉન, આરડબ્લ્યુ, પર્ના, એમ.કે., નોએલ, ડી.એમ., વ્હાઇટમોર, જેડી, લેહમેન, જે., સ્મિથ, એમ.એલ. (2011). એમ્ફ્ટેમાઈન લોકમોટર સંવેદના અને કુંડપાયરોલથી નવજાત રીતે સારવાર આપતી કિશોર પુરૂષ અને સ્ત્રી ઉંદરોમાં કન્ડિશન્ડ પ્લેસ પ્રેફરન્સ. બિહાવ ફાર્માકોલ. 22, 374–378. doi: 10.1097/FBP.0b013e328348737b
બ્રુહાસ, એમઆર, ચાવકિન, સી. (2010) કપ્ના ઓપીયોઇડ રીસેપ્ટર પર કિનાસે કાસ્કેડ્સ અને લિગાન્ડ-ડાયરેક્ટ કરેલા સંકેત. સાયકોફાર્માકોલ. (બર્લ) 210, 137–147. doi: 10.1007/s00213-010-1806-y
કlaલ્ઘન, સી.કે., રineઇન, જે., ઓ'મારા, એસ.એમ. (2018). પ્રેરણા અને મુખ્ય ડિપ્રેસિવ ડિસઓર્ડરમાં ioપિઓઇડ રીસેપ્ટર્સ માટેની સંભવિત ભૂમિકાઓ. પ્રોગ. મગજ રિઝ. 239, 89–119. doi: 10.1016 / bs.pbr.2018.07.009
કાર, ડીબી, સેસesક, એસઆર (2000) પ્રીફ્રન્ટલ કોર્ટેક્સથી ઉંદર વેન્ટ્રલ ટેગમેન્ટલ એરિયા પ્રોજેક્ટમાં GABA- ધરાવતા ન્યુરોન્સ. સિનેપ્સ 38 (2), 114–123. doi: 10.1002/1098-2396(200011)38:2<114::AID-SYN2>3.0.CO;2-R
કેસોનોવા, જેપી, વેલિસ, જી.પી., ફુએન્ટેઆલ્બા, જેએ (2013) એમ્ફેટામાઇન લોકોમોટર સેન્સેટાઇઝેશન ઉંદર મેડિયલ પ્રિફ્રન્ટલ કોર્ટેક્સમાં ઉન્નત ઉચ્ચ કે + -સ્ટિમેટેડ ડોપામાઇન પ્રકાશન સાથે છે. પાછળ મગજનો અનાદર 237, 313-317. ડોઇ: 10.1016 / j.bbr.2012.09.052
ચાર્ટોફ, ઇએચ, માવરિકાકી, એમ. (2015) કપ્પા ioપિઓઇડ રીસેપ્ટર ફંક્શનમાં સેક્સ તફાવત અને વ્યસન પર તેમની સંભવિત અસર. આગળ. ન્યુરોસી. 9, 466. doi: 10.3389 / fnins.2015.00466
ચાર્ટોફ, ઇએચ, ઇબનર, એસઆર, સ્પેરો, એ., પોટર, ડી., બેકર, પીએમ, રાગોઝિનો, એમઇ, એટ અલ. (2016). કપ્પા ioપિઓઇડ રીસેપ્ટર એક્ટિવેશન અને કોકેઇન વચ્ચેનો સંબંધિત સમય ઇનામ અને ડોપામાઇન પ્રકાશન પરની અસર નક્કી કરે છે. ન્યુરોસાયકોફોર્માકોલોજી 41, 989-1002. ડોઇ: 10.1038 / npp.2015.226
ચાવકિન, સી., કુબ, જીએફ (2016). ડાયનોર્ફિન, ડિસફોરિયા અને પરાધીનતા: વ્યસનનું તાણ. ન્યુરોસાયકોફોર્માકોલોજી 41, 373-374. ડોઇ: 10.1038 / npp.2015.258
ચેફર, VI, Czyzyk, T., Bolan, EA, Moron, J., Pintar, JE, Shippenberg, T. S. (2005). એન્ડોજેનસ કપ્પા-ioપિઓઇડ રીસેપ્ટર સિસ્ટમ્સ મેસોએકમ્બમ્બલ ડોપામાઇન ગતિશીલતા અને કોકેનની સંવેદનશીલતાને નિયંત્રિત કરે છે. જે ન્યુરોસી. 25, 5029-5037. ડોઇ: 10.1523 / JNEUROSCI.0854-05.2005
ચેફર, છઠ્ઠો, ઝપાટા, એ. શિપ્નબર્ગ, ટી.એસ., બુંગાય, પી.એમ. (2006) ક્વોન્ટિટેટિવ નો-નેટ-ફ્લક્સ માઇક્રોડાયલિસિસ માઉસ ન્યુક્લિયસ એક્મ્બમ્બન્સમાં ડોપામાઇન અપટેકમાં વધારો અને ઘટાડવાની તપાસની મંજૂરી આપે છે. જે ન્યુરોસી. પદ્ધતિઓ 155, 187-193. ડોઇ: 10.1016 / j.jneumeth.2005.12.018
કોલ, આરએલ, કોનરાડી, સી., ડગ્લાસ, જે., હાઇમેન, એસઇ (1995). એમ્ફેટામાઇન અને ડોપામાઇનમાં ચેતાકોષીય અનુકૂલન: ઉંદર સ્ટ્રાઇટમમાં પ્રોડિનોર્ફિન જનીન નિયમનની પરમાણુ પદ્ધતિઓ. ચેતાકોષ 14, 813–823. doi: 10.1016/0896-6273(95)90225-2
કોનવે, એસ.એમ., પુટિક, ડી., રસેલ, એસ., પોટર, ડી., રોઇટમેન, એમ.એફ., ચાર્ટોફ, ઇ. એચ. (2019). કપ્પા ioપિઓઇડ રીસેપ્ટર એક્ટિવેશનની પ્રેરણાત્મક અને ડોપામાઇન-સપ્રેસિંગ ઇફેક્ટિવ સ્ત્રીઓ માટે પુરુષો ઓછી સંવેદનશીલ હોય છે. ન્યુરોફર્મકોલોજી 146, 231-241. ડોઇ: 10.1016 / જે. ન્યુરોફાર્મ.2018.12.002
કોપ, ઝેડએ, હ્યુગિન્સ, કેએન, શેપ્પાર્ડ, એબી, નોએલ, ડીએમ, રોને, ડીએસ, બ્રાઉન, આરડબ્લ્યુ (2010). નવજાત ક્વિનપાયરોલ ટ્રીટમેન્ટ, પુખ્તાવસ્થામાં એમ્ફેટેમાઇન સારવારના જવાબમાં ન્યુક્લિયસ accમ્બેન્સ કોરમાં લોકોમોટર એક્ટિવેશન અને ડોપામાઇન પ્રકાશનમાં વધારો કરે છે. સિનેપ્સ 64, 289-300. ડોઇ: 10.1002 / syn.20729
કોક્સ, બીએમ, યંગ, એબી, જુઓ, આરઇ, રીશેલ, સીએમ (2013) ઉંદરોમાં શોધતા મેથેમ્ફેટેમાઇનમાં લિંગ તફાવત: xyક્સીટોસિનની અસર. સાયકોરોયુરોએન્ડ્રોક્રિનોલોજી 38, 2343-2353. ડોઇ: 10.1016 / j.psyneuen.2013.05.005
કલ્વર, કેઇ, શેચેટમેન, એચ., લેવંટ, બી. (2008) બદલાયેલ ડોપામાઇન ડી 2 જેવા રીસેપ્ટર, ક્વિનપાયરોલને વર્તણૂકીય સંવેદનશીલતાવાળા ઉંદરોમાં બંધનકર્તા: રો 41-1049 સાથે પૂર્વ-સારવારની અસરો. EUR. જે. ફાર્માકોલ 592, 67-72. ડોઇ: 10.1016 / j.ejphar.2008.06.101
ડાલમેન, એફસી, ઓ'માલે, કેએલ (1999) કપ્પા-ઓપીયોઇડ સહિષ્ણુતા અને ડોપામિનેર્જિક મિડબ્રેઇન ન્યુરોન્સની સંસ્કૃતિમાં અવલંબન. જે ન્યુરોસી. 19, 5750–5757. doi: 10.1523/JNEUROSCI.19-14-05750.1999
ડી વિરીઝ, ટીજે, શોફેલમીર, એએન, બિન્નેકેડે, આર., રાસે, એચ., વેન્ડરશ્યુરન, એલજે (2002). ડોકેમાઇન ડી 2 રીસેપ્ટર્સ દ્વારા મધ્યસ્થી અને કોરોઇનની હેરોઈન-શોધતી વર્તણૂક સાથે ફરીથી જોડાણ સમય આધારિત અને વર્તણૂકીય સંવેદના સાથે સંકળાયેલું છે. ન્યુરોસાયકોફોર્માકોલોજી 26, 18–26. doi: 10.1016/S0893-133X(01)00293-7
ડિવાઇન, ડી.પી., લિયોન, પી., પોકોક, ડી., વાઈઝ, આરએ (1993). બેસલ મેસોલીમ્બિક ડોપામાઇન પ્રકાશનના મોડ્યુલેશનમાં વેન્ટ્રલ ટેગમેન્ટલ મ્યુ, ડેલ્ટા અને કપ્પા ioપિઓઇડ રીસેપ્ટર્સની વિશિષ્ટ સંડોવણી: વિવો માઇક્રોડાયલિસીસ અભ્યાસમાં. જે. ફાર્માકોલ સમાપ્તિ થર. 266, 1236-1246
દી ચિઆરા, જી., ઇમ્પેરાટો, એ. (1988). ન્યુક્લિયસના અભિયાનમાં અને મુક્તપણે ફરતા ઉંદરોના ડોર્સલ પુજન્યમાં ડોપામાઇનના પ્રકાશન પર મ્યુ અને કપ્પાના અસ્પષ્ટ એગોનિસ્ટ્સની વિરુદ્ધ અસરો. જે. ફાર્માકોલ સમાપ્તિ થર. 244, 1067-1080. ડોઇ: 10.1073 / pnas.85.14.5274
ડ્વોર્કિન, એ., પેરીઓલ્ટ, એમ.એલ., શેચેટમેન, એચ. (2006) બાધ્યતા મનોગ્રસ્તિ વિકારના પ્રાણીના મોડેલમાં ડોપામાઇન એગોનિસ્ટ ક્વિનપાયરોલના વારંવાર ઇન્જેક્શન દ્વારા પ્રેરિત અનિવાર્ય ચેકિંગના વિકાસ અને ટેમ્પોરલ સંસ્થા. પાછળ મગજનો અનાદર 169, 303-311. ડોઇ: 10.1016 / j.bbr.2006.01.024
ઇબેનર, એસઆર, રોઇટમેન, એમએફ, પોટર, ડી.એન., રચલિન, એબી, ચાર્ટોફ, ઇએચ (2010) કપ્પા ioપિઓઇડ રીસેપ્ટર એગોનિસ્ટ સાલ્વિનોરીન એ ની ડિપ્રેસિવ જેવી અસરો ન્યુક્લિયસ accમ્બબેન્સમાં ફાસિક ડોપામાઇન પ્રકાશનમાં ઘટાડો સાથે સંકળાયેલ છે. સાયકોફાર્માકોલ. (બર્લ) 210, 241–252. doi: 10.1007/s00213-010-1836-5
એડવર્ડ્સ, એનજે, તેજેડા, એચ.એ., પિગ્નેટેલી, એમ., ઝાંગ, એસ., મ Mcકડેવિટ, આરએ, વુ, જે., એટ અલ. (2017). વીટીએના અવરોધક આર્કિટેક્ચરમાં સર્કિટની વિશિષ્ટતા કોકેન-પ્રેરિત વર્તનને નિયંત્રિત કરે છે. નાટ. ન્યુરોસી. 20 (3), 438–448. doi: 10.1038 / nn.4482
એહરિચ, જેએમ, ફિલિપ્સ, પીઈએમ, ચાવકિન, સી. (2014) કપ્પા ioપિઓઇડ રીસેપ્ટર એક્ટિવેશન, માઉસ ન્યુક્લિયસ એક્મ્બમ્બન્સમાં વિવોમાં નોંધાયેલ ઇવોક્ટેડ ડોપામાઇન પ્રકાશનમાં કોકેન-પ્રેરિત વધારાને સંભવિત કરે છે. ન્યુરોસાયકોફોર્માકોલોજી 39, 3036-3048. ડોઇ: 10.1038 / npp.2014.157
એહરિચ, જેએમ, મેસિન્જર, ડીઆઈ, નાકલ, સીઆર, કુહર, જેઆર, સ્કટ્ટાઉર, એસએસ, બ્રુહાસ, એમઆર, એટ અલ. (2015). કપ્પા ioપિઓઇડ રીસેપ્ટર-પ્રેરિત એવર્ઝનને વીટીએ ડોપામાઇન ન્યુરોન્સમાં પી 38 એમએપીકે સક્રિયકરણની જરૂર છે. જે ન્યુરોસી. 35, 12917-12931. ડોઇ: 10.1523 / JNEUROSCI.2444-15.2015
આઈલમ, ડી., શેચેટમેન, એચ. (1989) લોકો-ગતિ અને હલનચલન પર ડી -2 એગોનિસ્ટ ક્વિનપાયરોલની બિફેસિક અસર. EUR. જે. ફાર્માકોલ 161, 151–157. doi: 10.1016/0014-2999(89)90837-6
આઈલમ, ડી., શેચેટમેન, એચ. (2005) બાધ્યતા મનોગ્રસ્તિ વિકારના પ્રાણીના મોડેલ તરીકે સાયકોસ્ટીમ્યુલેન્ટ-પ્રેરિત વર્તન: અનિવાર્ય ધાર્મિક વિધિઓના સ્વરૂપમાં નૈતિક અભિગમ. સી.એન.એસ. સ્પેક્ટર. 10, 191–202. doi: 10.1017 / S109285290001004X
એસ્કોબાર, એપી, કોર્નેજો, એફએ, એન્ડ્રેસ, એમઇ, ફુએન્ટેઆલ્બા, જેએ (2012) કપ્પા ioપિઓઇડ રીસેપ્ટર એગોનિસ્ટ યુ 69593 સાથે વારંવાર સારવાર કરવાથી ન્યુક્લિયસ accમ્બેન્સમાં ઉન્નત કે + પ્રેરિત ડોપામાઇન પ્રકાશન વિરુદ્ધ થાય છે, પરંતુ એમ્ફેટામાઇન-સંવેદી ઉંદરોમાં લોકોમોટર સંવેદનાની અભિવ્યક્તિ નહીં. ન્યુરોકેમ. Int. 60 (4), 344–349. doi: 10.1016 / j.neuint.2012.01.014
એસ્કોબાર, એપી, કોર્નેજો, એફએ, ivલિવારેસ-કોસ્ટા, એમ., ગોંઝેલેઝ, એમ., ફુએન્ટેઆલ્બા, જેએ, ગિસલિંગ, કે., એટ અલ. (2015). અવરોધક ડી 2 oreટોરિસેપ્ટર ફંક્શનમાં ક્વિનપાયરોલ-સંવેદનાત્મક ઉંદરોના મધ્યવર્તી કેન્દ્રમાં ડોપામાઇન અને ગ્લુટામેટ ન્યુરોટ્રાન્સમિશનમાં ઘટાડો. જે. ન્યુરોકેમ 134, 1081–1090. doi: 10.1111 / jnc.13209
એસ્કોબાર, એપી, ગોંઝલેઝ, સાંસદ, મેઝા, આરસી, નોચેસ, વી., હેન્ની, પી., ગિસલિંગ, કે., એટ અલ. (2017). ઉંદરોમાં ક્વિનપાયરોલ-પ્રેરિત લોકોમોટર સંવેદનામાં ડોપામાઇન ડી 2 રીસેપ્ટર ફંક્શનની કપ્પા opપિઓઇડ રીસેપ્ટર પentiન્ટિએશનની પદ્ધતિઓ. Int. જે ન્યુરોપ્સિકોફાર્માકોલ. 20, 660–669. doi: 10.1093 / ijnp / pyx042
એવરિટ, બીજે, રોબિન્સ, ટીડબ્લ્યુ (2005) ડ્રગના વ્યસન માટે મજબૂતીકરણની ન્યુરલ સિસ્ટમ્સ: ક્રિયાઓથી માંડીને મજબૂરી સુધી. નાટ. ન્યુરોસી. 8, 1481-1489. ડોઇ: 10.1038 / nn1579
ફટ્ટોર, એલ., મેલિસ, એમ. (2016) આવેગજન્ય અને અનિવાર્ય વર્તણૂંકમાં લિંગ તફાવત: ડ્રગ વ્યસન પર ધ્યાન કેન્દ્રિત. વ્યસની બાયોલ. 21 (5), 1043–1051. doi: 10.1111 / adb.12381
ફેરારીયો, સીઆર, ગોર્ની, જી., ક્રોમ્બેગ, એચએસ, લિ, વાય., કોલબ, બી., રોબિન્સન, ટીઇ (2005). નિયંત્રિતથી વધેલા કોકેઇનના ઉપયોગમાં સંક્રમણ સાથે સંકળાયેલ ન્યુરલ અને વર્તણૂકીય પ્લાસ્ટિસિટી. બાયોલ. માનસિક. 58 (9), 751-9.
ફીગી, એમ., પટ્ટીજ, ટી., વિલુહ્ન, આઇ., લુઇગજેસ, જે., વેન ડેન બ્રિંક, ડબલ્યુ., ગૌદ્રીઆન, એ., એટ અલ. (2016). બાધ્યતા મનોગ્રસ્તિ-અનિવાર્ય વિકાર અને વ્યસનોમાં અનિવાર્યતા. EUR. ન્યુરોસાયક્ફોર્માકોલ 26, 856-868. ડોઇ: 10.1016 / j.euroneuro.2015.12.003
ફોર્ડ, સીપી, માર્ક, જી.પી., વિલિયમ્સ, જેટી (2006) લક્ષ્ય સ્થાન અનુસાર મેસોલીમ્બીક ડોપામાઇન ન્યુરોન્સના ગુણધર્મો અને opપિઓઇડ નિષેધ. જે ન્યુરોસી. 26, 2788-2797. ડોઇ: 10.1523 / JNEUROSCI.4331-05.2006
ફોર્ડ, સીપી, બેકસ્ટીડ, એમજે, વિલિયમ્સ, જેટી (2007) સોપ્ટોોડેન્ડ્રિટિક ડોપામાઇન ઇનહિબિટોરી પોસ્ટસિનાપ્ટિક પ્રવાહોનું કપ્પા ઓપીયોઇડ અવરોધ. જે ન્યુરોફિઝિઓલ. 97, 883-891. ડોઇ: 10.1152 / jn.00963.2006
ફુએન્ટેઆલ્બા, જે.એ., ગેસલિંગ, કે., મેજેન્દોઝો, કે., એન્ડ્રેસ, એમઈ (2006). પસંદગીયુક્ત કપ્પા-ioપિઓઇડ રીસેપ્ટર એગોનિસ્ટ યુ-69593 ના વારંવારના વહીવટથી ઉંદરીના મધ્યવર્તી કેન્દ્રમાં ઉત્તેજિત ડોપામાઇન એક્સ્ટ્રા સેલ્યુલર સ્તર વધે છે. જે ન્યુરોસી. Res. 84, 450-459. ડોઇ: 10.1002 / jnr.20890
ફુએન્ટેઆલ્બા, જેએ, ગિસલિંગ, કે., એન્ડ્રેસ, એમઇ (2007) પસંદગીયુક્ત કપ્પા-ioપિઓઇડ રીસેપ્ટર એગોનિસ્ટ યુ-69593 ના વારંવારના વહીવટ દ્વારા પ્રેરિત એમ્ફેટામાઇનનો વધતો લોકોમોટર પ્રતિસાદ. સિનેપ્સ 61, 771-777. ડોઇ: 10.1002 / syn.20424
ગેહર્કે, બી.જે., ચેફર, છઠ્ઠો, શિપ્નબર્ગ, ટી.એસ. (2008) ઉંદર ડોર્સલ સ્ટ્રાઇટમમાં ડોપામાઇન ફંક્શન પર સલ્વિનોરીન એના તીવ્ર અને વારંવારના વહીવટની અસરો. સાયકોફાર્માકોલ. (બર્લ) 197, 509–517. doi: 10.1007/s00213-007-1067-6
ગિયુલિયાનો, સી., બેલિન, ડી., એવરિટ, બીજે (2019). વર્તન પર ડોર્સોટલલ સ્ટ્રાઇટલ કંટ્રોલને છૂટા કરવામાં નિષ્ફળતાના પરિણામ રૂપે અનિયમિત આલ્કોહોલ એઇકિંગ. જે ન્યુરોસિ. 39 (9), 1744-1754. doi: 10.1523 / JNEUROSCI.2615-18.2018
ગોટો, વાય., ગ્રેસ, એએ (2008) મધ્યવર્તી કેન્દ્રમાં લિંબિક અને કોર્ટીકલ માહિતી પ્રક્રિયા. પ્રવાહો ન્યૂરોસી 31, 552-558. ડોઇ: 10.1016 / j.tins.2008.08.002
ગ્રેસ, એએ, બન્ની, બીએસ (1980) નિગ્રલ ડોપામાઇન ન્યુરોન્સ: ઇન્ટ્રાસેલ્યુલર રેકોર્ડિંગ અને એલ-ડોપા ઇંજેક્શન અને હિસ્ટોફ્લોરોસેન્સ સાથે ઓળખ. વિજ્ઞાન 210, 654-656. ડોઇ: 10.1126 / વિજ્ઞાન.7433992
ગ્રે, એએમ, રlsલ્સ, એસ.એમ., શિપ્નબર્ગ, ટી.એસ., મGકિંટી, જે.એફ. (1999) Κ-opioid agonist, U-69593, વેન્ટ્રલ સ્ટ્રાઇટમમાં ડોફેમાઇન અને ગ્લુટામેટનું તીવ્ર એમ્ફેટામાઇન-ઇવોક્ટેડ વર્તણૂક અને કેલ્શિયમ આધારિત ડાયાલીસેટ સ્તરમાં ઘટાડો કરે છે. જે. ન્યુરોકેમ 73, 1066-1074. ડોઇ: 10.1046 / j.1471-4159.1999.0731066.x
ગ્રેઝિએન, એનએમ, પોટર, એએમ, બ્રાયન્ડ, એલએ, પિયર્સ, આરસી, કૌર, જેએ (2013) કપ્પા ioપિઓઇડ રીસેપ્ટર્સ તાણ-પ્રેરિત કોકેઇનની શોધ અને સિનેપ્ટિક પ્લાસ્ટિસિટીને નિયંત્રિત કરે છે. ચેતાકોષ 77, 942-954. ડોઇ: 10.1016 / j.neuron.2012.12.034
હીડબ્રેડર, સીએ, શિપ્નબર્ગ, ટીએસ (1994). યુ -69593 બેઝલ umbમ્બમ્બન્સ ડોપામાઇનને સામાન્ય કરીને કોકેન સંવેદનાને અટકાવે છે. ન્યૂરોરપોર્ટ 5, 1797–1800. doi: 10.1097/00001756-199409080-00028
હર્નાન્ડેઝ-અવિલા, સીએ 1., રnsન્સવિલે, બી.જે., ક્રેનઝ્લર, એચઆર (2004). ઓપિઓઇડ-, કેનાબીસ- અને આલ્કોહોલ આધારિત મહિલાઓ પદાર્થના દુરૂપયોગની સારવારમાં વધુ ઝડપથી પ્રગતિ દર્શાવે છે. ડ્રગ આલ્કોહોલ ડીપેન્ડ. 74 (3), 265–272. doi: 10.1016 / j.drugalcdep.2004.02.001
હોફમેન, એએફ, સ્પિવાક, સીઈ, લ્યુપિકા, સીઆર (2016) પ્રતિબંધિત પ્રસાર મોડેલ અને ફાસ્ટ-સ્કેન ચક્રીય વોલ્ટેમેટ્રી દ્વારા વર્ણવેલ ડોપામાઇન ટ્રાન્સપોર્ટ અવરોધકો દ્વારા ઉન્નત ડોપામાઇન પ્રકાશન. એસીએસ કેમ. ન્યુરોસી. 7, 700-709. doi: 10.1021 / acchemneuro.5b00277
હોલ્ડન, સી. (2001) "વર્તન" વ્યસનો: શું તે અસ્તિત્વમાં છે? વિજ્ઞાન 294, 980-982. ડોઇ: 10.1126 / વિજ્ઞાન.294.5544.980
ઇમ્પેરાટો, એ., ડી ચિઆરા, જી. (1988) મગજ ડાયાલિસિસ સાથે અભ્યાસ કરેલ સ્થાનિક રીતે લાગુ ડી -1 અને ડી -2 રીસેપ્ટર એગોનિસ્ટ્સ અને વિરોધી લોકોની અસરો. EUR. જે. ફાર્માકોલ 156, 385–393. doi: 10.1016/0014-2999(88)90284-1
જેક્સન, કેજે, મLકલોફ્લિન, જેપી, કેરોલ, એફઆઈ, દમાજ, એમઆઈ (2013) કપ્પા ioપિઓઇડ રીસેપ્ટર વિરોધી, નોર્બિનેટોલ્ફિમાઇન, ઉનાળામાં નિકોટિન-કન્ડિશન્ડ સ્થળ પસંદગીને તાણ અને ડ્રગ-પ્રેરિત પુનstસ્થાપન પરની અસરો. સાયકોફાર્માકોલ. (બર્લ) 226, 763–768. doi: 10.1007/s00213-012-2716-y
કિવેલ, બી., ઉઝેલેક, ઝેડ., સુંદરમૂર્તિ, એસ., રાજામણિકમ, જે., ઇવાલ્ડ, એ., ચેફર, વી., એટ અલ. (2014). સાલ્વિનોરીન એ, કપ્પા ioપિઓઇડ રીસેપ્ટર અને ERK1 / 2-આધારિત આ પદ્ધતિ દ્વારા ડોપામાઇન ટ્રાન્સપોર્ટર કાર્યને નિયંત્રિત કરે છે. ન્યુરોફર્મકોલોજી 86, 228-240. ડોઇ: 10.1016 / જે. ન્યુરોફાર્મ.2014.07.016
કોલ્ટઝો, ટીઇ, inસ્ટિન, જેડી, વેઝિના, પી. (2003) ક્વિનપાયરોલ પ્રત્યે વર્તણૂક સંવેદના, ન્યુક્લિયસ umbક્યુબેન્સ ડોપામાઇન ઓવરફ્લો સાથે સંકળાયેલ નથી. ન્યુરોફર્મકોલોજી 44, 102–110. doi: 10.1016/S0028-3908(02)00328-3
કુબ, જીએફ, વોલ્કો, એનડી (2016). વ્યસનની ન્યુરોબાયોલોજી: એક ન્યુરોસિક્ટીટ્રી વિશ્લેષણ. લેન્સેટ સાઇકિયાટ્રી 3, 760–773. doi: 10.1016/S2215-0366(16)00104-8
કુબ, જીએફ (2013). વ્યસન એ ઈનામની ખામી અને તાણની સર્ફિટ ડિસઓર્ડર છે. આગળ. મનોચિકિત્સા 4, 72. ડોઇ: 10.3389 / fpsyt.2013.00072
લાબાબાદ, જે. 1., મેંચોન, જેએમ, એલોન્સો, પી., સેગાલસ, સી., જીમેનેઝ, એસ., જૌરીતા, એન., એટ અલ. (2008). બાધ્યતા મનોગ્રસ્તિ લક્ષણ પરિમાણોમાં લિંગ તફાવત. ડિપ્રેશન ચિંતા 25 (10), 832–838. doi: 10.1002 / da.20332
મેઇઝન્યુવ, આઇએમ, આર્ચર, એસ., ગ્લિક, એસડી (1994) U50,488, એક કપ્પા ioપિઓઇડ રીસેપ્ટર એગોનિસ્ટ, ઉંદરોના મધ્યવર્તી કેન્દ્રમાં એક્સ્ટ્રા સેલ્યુલર ડોપામાઇનમાં કોકેન-પ્રેરિત વધારાને ઘટાડે છે. ન્યુરોસી. લેટ. 181, 57–60. doi: 10.1016/0304-3940(94)90559-2
મન્સુર, એ., ફોક્સ, સીએ, અકિલ, એચ., વોટસન, એસજે (1995) ઉંદર સી.એન.એસ. માં Opપિઓઇડ-રીસેપ્ટર એમઆરએનએ અભિવ્યક્તિ: શરીરરચનાત્મક અને કાર્યાત્મક અસરો. પ્રવાહો ન્યૂરોસી 18 (1), 22–29. doi: 10.1016/0166-2236(95)93946-U
મન્સૌર, એ., બર્ક, એસ., પેવલિક, આરજે, અકિલ, એચ., વોટસન, એસજે (1996). ઉંદરો સી.એન.એસ. અને કફોત્પાદક માં ક્લોન થયેલ કપ્પા 1 રીસેપ્ટરનું ઇમ્યુનોહિસ્ટોકેમિકલ સ્થાનિકીકરણ. ન્યુરોસાયન્સ 71, 671–690. doi: 10.1016/0306-4522(95)00464-5
માર્ગોલીસ, ઇબી, કારખાનિસ, એએન (2019) ડોપામિનેર્જિક સેલ્યુલર અને કપ્પા ioપિઓઇડ રીસેપ્ટર મધ્યસ્થી વિરુદ્ધમાં સર્કિટ યોગદાન. ન્યુરોકેમ. Int. 129, 104504. doi: 10.1016 / j.neuint.2019.104504
માર્ગોલીસ, ઇબી, હેલ્લ્મસ્ટાડ, જીઓ, બોંસી, એ., ફીલ્ડ્સ, એચએલ (2003) કપ્પા-ioપિઓઇડ એગોનિસ્ટ્સ મિડબ્રેઇન ડોપામિનેર્જિક ન્યુરોન્સને સીધા અવરોધે છે. જે ન્યુરોસી. 23, 9981–9986. doi: 10.1523/JNEUROSCI.23-31-09981.2003
માર્ગોલીસ, ઇબી, હેલ્લ્મસ્ટાડ, જીઓ, બોંસી, એ., ફીલ્ડ્સ, એચએલ (2005) કપ્પા અને મ્યુ ઓપીઓઇડ એગોનિસ્ટ્સ બંને ગ્લુટામેટર્જિક ઇનપુટથી વેન્ટ્રલ ટેગમેન્ટલ એરિયા ન્યુરોન્સને અટકાવે છે. જે ન્યુરોફિઝિઓલ. 93, 3086-3093. ડોઇ: 10.1152 / jn.00855.2004
માર્ગોલીસ, ઇબી, લોક, એચ., ચેફર, છઠ્ઠી, શિપનબર્ગ, ટીએસ, હેલ્મસ્ટેડ, જીઓ, ફીલ્ડ્સ, એચએલ (2006). કપ્પા ioપિઓઇડ્સ ડોફામિનેર્જિક ન્યુરોન્સને પ્રીફ્રન્ટલ કોર્ટેક્સમાં રજૂ કરતા પસંદગીયુક્ત રીતે નિયંત્રિત કરે છે. પ્રોક. નાટ. એકાદ વિજ્ઞાન. યૂુએસએ 103, 2938-2942. ડોઇ: 10.1073 / pnas.0511159103
માર્ગોલીસ, ઇબી, મિશેલ, જેએમ, ઇશિકાવા, જે., હેલમસ્ટાડ, જીઓ, ફીલ્ડ્સ, એચએલ (2008). મિડબ્રેઇન ડોપામાઇન ન્યુરોન્સ: પ્રક્ષેપણ લક્ષ્ય ક્રિયા સંભવિત અવધિ અને ડોપામાઇન ડી (2) રીસેપ્ટર અવરોધ નક્કી કરે છે. જે ન્યુરોસી. 28, 8908-8913. ડોઇ: 10.1523 / JNEUROSCI.1526-08.2008
મેથ્યુઝ, આઇઝેડ, મCકકોર્મિક, સીએમ (2007) કિશોરાવસ્થાના અંતમાં સ્ત્રી અને પુરુષ ઉંદરો એમ્ફેટામાઇન-પ્રેરણિત લોકોમોટર પ્રવૃત્તિમાં પુખ્ત વયના લોકો કરતા અલગ હોય છે, પરંતુ એમ્ફેટામાઇન માટે શરતી સ્થાનની પસંદગીમાં નહીં. બિહાવ ફાર્માકોલ. 18, 641–650. doi: 10.1097/FBP.0b013e3282effbf5
મેથિસ, એમ.એ. 1., પીડી, એ., ફનારો, જી., આરસી, ટી., મોરેસ, આઇ., એઆર, ટી., એટ અલ. (2011). બાધ્યતા મનોગ્રસ્તિ વિકારમાં લિંગ તફાવત: સાહિત્યિક સમીક્ષા. બ્રાઝ. જે મનોચિકિત્સા 33 (4), 390–399. doi: 10.1590 / S1516-44462011000400014
મેકલોફ્લિન, જેપી, માર્ટન-પોપોવિસિ, એમ., ચાવકિન, સી. (2003) કપ્પા ioપિઓઇડ રીસેપ્ટર વિરોધી અને પ્રોડિનોર્ફિન જનીન વિક્ષેપ તાણ-પ્રેરિત વર્તણૂકીય પ્રતિસાદને અવરોધિત કરે છે. જે ન્યુરોસી. 23 (13), 5674–5683. doi: 10.1523 / JNEUROSCI.23-13-05674.2003
મેકલફ્લિન, જેપી, લેન્ડ, બીબી, લી, એસ., પિન્ટાર, જેઈ, ચાવકિન, સી. (2006 એ). U50,488 મીમિક્સ દ્વારા કપ્પા ioપિઓઇડ રીસેપ્ટર્સની પહેલાંની સક્રિયકરણ, કોકેઇન પ્લેસ પ્રેફરન્સ કન્ડીશનિંગને સંભવિત કરવા માટે વારંવાર દબાણયુક્ત તરણ તણાવને દબાણ આપે છે. ન્યુરોસાયકોફોર્માકોલોજી 31, 787-794. ડોઇ: 10.1038 / sj.npp.1300860
મLકલોફલિન, જેપી, લી, એસ., વાલ્ડેઝ, જે., ચvકિન, ટી.એ., ચvકિન, સી. (2006 બી). સામાજિક પરાજિત તાણ-પ્રેરિત વર્તણૂકીય પ્રતિભાવો અંતર્ગત કપ્પા ioપિઓઇડ સિસ્ટમ દ્વારા મધ્યસ્થી છે. ન્યુરોસાયકોફોર્માકોલોજી 31 (6), 1241–1248. doi: 10.1038 / sj.npp.1300872
માઇલેસી-હéલé, એ., મMકમિલેન, ડીઇ, લureરેન્ઝના, ઇએમ, બાયર્ન્સ-બ્લેક, કેએ, ઓવેન્સ, એસએમ (2007) (+) - એમ્ફેટેમાઇન- અને (+) - જાતિના તફાવતોમાં પુરુષ અને સ્ત્રી સ્પ્રેગ-ડawલી ઉંદરોમાં મેથેમ્ફેટેમાઇન-પ્રેરિત વર્તણૂકીય પ્રતિસાદ. ફાર્માકોલ બાયોકેમ પાછળ 86, 140-149. ડોઇ: 10.1016 / j.pbb.2006.12.018
નેગસ, એસએસ, મેલ્લો, એનકે, પોર્ટોગીઝ, પીએસ, લિન, સીઈ (1997). રીસસ વાંદરાઓ દ્વારા કોકેઇન સ્વ-વહીવટ પર કપ્પા ઓપીયોઇડ્સની અસરો. જે. ફાર્માકોલ સમાપ્તિ થર. 282, 44-55
નેસ્ટલર, ઇજે (એક્સએનએમએક્સ). લાંબા ગાળાના પ્લાસ્ટિકિટી અંતર્ગત વ્યસનનો પરમાણુ આધાર. નેટ રેવ. ન્યૂરોસી 2, 119-128. ડોઇ: 10.1038 / 35053570
પેરેલaultલ્ટ, એમ.એલ., ગ્રેહામ, ડી., બિસ્નારે, એલ., સિમ્સ, જે., હેટન, એસ., શેચેટમેન, એચ. (2006). કપ્પા-ioપિઓઇડ એગોનિસ્ટ યુ 69593 એ ડી 2 / ડી 3 એગોનિસ્ટ ક્વિનપાયરોલ માટે લોમોમોટર સંવેદનાને સંભવિત કરે છે: પૂર્વ- અને પોસ્ટસynનaptપ્ટિક મિકેનિઝમ્સ. ન્યુરોસાયકોફોર્માકોલોજી 31, 1967-1981. ડોઇ: 10.1038 / sj.npp.1300938
પેરેલaultલ્ટ, એમ.એલ., સીમેન, પી., શેચમેન, એચ. (2007) કપ્પા-ઓપીયોઇડ રીસેપ્ટર સ્ટીમ્યુલેશન બાધ્યતા-અનિવાર્ય ડિસઓર્ડર (ઓસીડી) ના ક્વિનપાયરોલ સેન્સેટાઇઝેશન મોડેલમાં અનિવાર્ય ચેકિંગના પેથોજેનેસિસને ઝડપી બનાવે છે. બિહાવ ન્યુરોસી. 121, 976-991. ડોઇ: 10.1037 / 0735-7044.121.5.976
પિયર્સ, આરસી, કાલિવાસ, પીડબ્લ્યુ (1997). એમ્ફેટેમાઇન જેવા સાયકોસ્ટીમ્યુલન્ટ્સ પ્રત્યે વર્તણૂકીય સંવેદનાની અભિવ્યક્તિનું એક સર્કિટરી મોડેલ. મગજ રિઝ. મગજ રિઝ. રેવ. 25, 192–216. doi: 10.1016/S0165-0173(97)00021-0
પોટર, એએમ, બિશપ, આરએ, બ્રાયંડ, એલએ, ગ્રાઝિયાને, એનએમ, પિયર્સ, આરસી, કૌર, જેએ (2014). કપ્પા ioપિઓઇડ રીસેપ્ટર્સનો પોસ્ટટ્રેસ બ્લોક ઇનહિબિરી સિનેપ્સનો લાંબા ગાળાના સંભવણાને બચાવે છે અને કોકેનની શોધના પુન: સ્થાપનાને અટકાવે છે. બાયોલ. મનોચિકિત્સા 76, 785-793. ડોઇ: 10.1016 / j.biopsych.2014.04.019
પુઇગ-રામોસ, એ., સેન્ટિયાગો, જીએસ, સેગરા, એસી (2008) યુ-69593, કેપ્પા ioપિઓઇડ રીસેપ્ટર એગોનિસ્ટ, સ્ત્રી ઉંદરોમાં કોકેન-પ્રેરિત વર્તણૂકીય સંવેદના ઘટાડે છે. બિહાવ ન્યુરોસી. 122, 151-160. ડોઇ: 10.1037 / 0735-7044.122.1.151
રોબિન્સ, ટીડબ્લ્યુ, ગિલાન, સીએમ, સ્મિથ, ડીજી, ડી વિટ, એસ., અર્શે, કેડી (2012) આવેગ અને અનિવાર્યતાના ન્યુરોકognગ્નિટીવ એન્ડોફેનોટાઇપ્સ: પરિમાણીય મનોચિકિત્સા તરફ. વલણ સંજ્ઞા. વિજ્ઞાન. 16, 81-91. ડોઇ: 10.1016 / j.tics.2011.11.009
રોબર્ટ્સ, ડીસીએસ, બેનેટ, એસએએલ, વિકર્સ, જીજે (1989) એસ્ટ્રોસ ચક્ર ઉંદરોના પ્રગતિશીલ ગુણોત્તરના સમયપત્રક પર કોકેઇન સ્વ-વહીવટને અસર કરે છે. સાયકોફાર્માકોલ. (બર્લ) 98, 408-411. ડોઇ: 10.1007 / BF00451696
રોબિન્સન, ટીઇ, બેરીજ, કેસી (1993). ડ્રગની તૃષ્ણાના ન્યુરલ આધાર: વ્યસનની પ્રોત્સાહન-સંવેદનાનો સિદ્ધાંત. મગજ રિઝ. મગજ રિઝ. રેવ. 18, 247–291. doi: 10.1016/0165-0173(93)90013-P
રોબિન્સન, ટીઇ, બેરીજ, કેસી (2001) પ્રોત્સાહન-સંવેદના અને વ્યસન. વ્યસન 96, 103-114. ડોઇ: 10.1046 / j.1360-0443.2001.9611038.x
ગુલાબ, જેએચ, કારખાનીસ, એએન, સ્ટીનીગર-બ્રchચ, બી., જોન્સ, એસઆર (2016). ક્રોનિક અંતરાલ ઇથેનોલના સંપર્કમાં આવતા ન્યુક્લિયસમાં ડોપામાઇન અપટેક રેટ અને કપ્પા opપિઓઇડ રીસેપ્ટર પ્રવૃત્તિ પર નાલ્મેફેનની વિશિષ્ટ અસરો. Int. જે. મોલ. વિજ્ઞાન. 17, 1216. doi: 10.3390 / ijms17081216
સ્ક્લોસબર્ગ, જેઈ, વ્હિટફિલ્ડ, ટીડબ્લ્યુ, જુનિયર, પાર્ક, પીઇ, ક્રોફોર્ડ, ઇએફ, જ્યોર્જ, ઓ., વેન્દ્રુસ્કોલો, એલએફ, એટ અલ. (2013). Κ ioપિઓઇડ રીસેપ્ટર્સની લાંબા ગાળાની દુશ્મનાવટ, હિરોઇનના સેવનની વૃદ્ધિ અને પ્રેરણામાં વધારો અટકાવે છે. જે ન્યુરોસી. 33 (49), 19384-19392. doi: 10.1523 / JNEUROSCI.1979-13.2013
સેસackક, એસઆર, પિકલ, વીએમ (1992) ઉંદર વેન્ટ્રલ ટેગમેન્ટલ ક્ષેત્રમાં એન્કેફાલિન અને ટાઇરોસિન હાઇડ્રોક્સિલેઝ ઇમ્યુનોએરેક્ટિવિટીનું ડ્યુઅલ અલ્ટ્રાસ્ટ્રક્ચરલ સ્થાનિકીકરણ: iateફીટ-ડોપામાઇન ક્રિયાપ્રતિક્રિયા માટેના બહુવિધ સબસ્ટ્રેટ્સ. જે ન્યુરોસિ. 12, 1335–a1350. doi: 10.1523/JNEUROSCI.12-04-01335.1992
સેસackક, એસઆર, okઓકી, સી., પિકેલ, વીએમ (1994). મિડબ્રેઇન ડોપામાઇન ન્યુરોન્સ અને તેમના સ્ટ્રાઇટલ લક્ષ્યોમાં ડી 2 રીસેપ્ટર જેવી ઇમ્યુનોરેક્ટિવિટીનું અલ્ટ્રાસ્ટ્રક્ચરલ સ્થાનિકીકરણ. જે ન્યુરોસી. 14, 88–106. doi: 10.1523/JNEUROSCI.14-01-00088.1994
સેસિયા, ટી., બીઝઅપ, બી. ગ્રેસ, એએ (2013). બાધ્યતા મનોગ્રસ્તિ વિકારના પ્રાણીઓના મોડેલોનું મૂલ્યાંકન: ફાસિક ડોપામાઇન ન્યુરોન પ્રવૃત્તિ સાથે સહસંબંધ. Int. જે ન્યુરોપ્સિકોફાર્માકોલ. 16, 1295–1307. doi: 10.1017 / S146114571200154X
શિપ્નબર્ગ, ટી.એસ., ઝપાટા, એ., ચેફર, છઠ્ઠી (2007) ડાયનોર્ફિન અને ડ્રગના વ્યસનની પેથોફિઝિયોલોજી. ફાર્માકોલ. Ther. 116, 306–321. doi: 10.1016 / j.pharmthera.2007.06.011
સ્મિથ, જેએ, લોફલિન, એસઇ, લેસ્લી, એફએમ (1992). [એચ] ડોપામાઇનનું ઉંદરોના ક્ષેપક મેસેન્સાફેલિક ડિસઓસેટેડ કોષ સંસ્કૃતિઓમાંથી મુક્ત થવા માટેના કપ્પા-ઓપિઓઇડ અવરોધ. મોલ. ફાર્માકોલ 42.
સ્મિથ, જેડબ્લ્યુ, ફેટ્સકો, એલએ, ઝૂ, આર., વાંગ, વાય. (2002) ડોપામાઇન ડી 2 એલ રીસેપ્ટર નોકઆઉટ ઉંદરને મોર્ફિનની સકારાત્મક અને નકારાત્મક પ્રબલિત ગુણધર્મો અને અવગણના શીખવાની તંગીમાં. ન્યુરોસાયન્સ 113 (4), 755–765. doi: 10.1016/S0306-4522(02)00257-9
સ્મિથ, જેએસ, શિન્ડલર, એજી, માર્ટિનેલી, ઇ., ગુસ્ટિન, આરએમ, બ્રુહાસ, એમઆર, ચાવકિન, સી. (2012). એમીગડાલામાં નિકોટિન કન્ડિશન્ડ પ્લેસ પ્રેફરન્સીસમાં ડાયનોર્ફિન / κ-ioપિઓઇડ રીસેપ્ટર સિસ્ટમની તાણ-પ્રેરિત સક્રિયકરણ. જે ન્યુરોસી. 32, 1488-1495. ડોઇ: 10.1523 / JNEUROSCI.2980-11.2012
સ્પેનાગેલ, આર., હર્ઝ, એ. શિપ્નબર્ગ, ટીએસ (1992) ટોનિકલી એક્ટિવ એન્ડોજેનસ ઓપિઓઇડ સિસ્ટમ્સનો વિરોધ કરવો મેસોલીમ્બિક ડોપામિનર્જિક માર્ગને મોડ્યુલેટ કરે છે. પ્રોક. નાટ. એકાદ વિજ્ઞાન. 89, 2046-2050. ડોઇ: 10.1073 / pnas.89.6.2046
સ્પેસિએલ, એસજી, મનાયે, કેએફ, સાદેક, એમ., જર્મન, ડીસી (1993). ઉંદરોના મધ્યબbraન ડોપામિનર્જિક પ્રદેશોમાં ioપિઓઇડ રીસેપ્ટર્સ. II. કપ્પા અને ડેલ્ટા રીસેપ્ટર oraટોરાડીયોગ્રાફી. જે ન્યુરલ ટ્રાન્સમ. જનરલ પંથ. 91, 53-66. ડોઇ: 10.1007 / BF01244918
સ્પર્લિંગ, આરઇ, ગોમ્સ, એસ.એમ., સીપેક, ઇઆઈ, કેરી, એએન, મLકલોફ્લિન, જેપી (2010) ઇથેનોલ-કન્ડિશન્ડ પ્લેસ પ્રેફરન્સ અને સ્વ-વહીવટની તાણ-પ્રેરિત પentiન્ટેશનની એન્ડોજેનસ કપ્પા-ioપિઓઇડ મધ્યસ્થતા. સાયકોફાર્માકોલ. (બર્લ) 210 (2), 199–209. doi: 10.1007/s00213-010-1844-5
સ્ટેકીટી, જેડી, કાલિવસ, પીડબ્લ્યુ (2011). ડ્રગ વોન્ડીંગ: વર્તણૂક સંવેદના અને ડ્રગ-શોધતી વર્તણૂકથી ફરીથી sથલો કરવો સિબીલી ડી. ફાર્માકોલ. રેવ. 63, 348–365. doi: 10.1124 / pr.109.001933
સ્ટુચલિક, એ., રેડોસ્તોવ, ડી., હટાલોવા, એચ., વેલ્સ, કે., નેકોવારોવા, ટી., કોપ્રીવોવા, જે., એટ અલ. (2016). ઓસીડીના ક્વિનપાયરોલ સેન્સિટાઇઝેશન ઉંદર મોડેલની માન્યતા: પ્રાણી અને ક્લિનિકલ અભ્યાસના પુરાવાને જોડતા. આગળ. બિહાવ ન્યુરોસી. 10, 209. doi: 10.3389 / fnbeh.2016.00209
સન, ટી., સોંગ, ઝેડ., ટિયન, વાય., ટિયાન, ડબલ્યુ., ઝુ, સી. જી, જી., એટ અલ. (2019) મેડિયલ પ્રિફ્રન્ટલ કોર્ટેક્સમાં બાસોલેટ્રલ એમીગડાલા ઇનપુટ બાધ્યતા મનોગ્રસ્તિ વિકાર જેવી તપાસની વર્તણૂકને નિયંત્રિત કરે છે. પ્રોક. નાટ. એકાદ વિજ્ઞાન. યૂુએસએ 116, 3799-3804. ડોઇ: 10.1073 / pnas.1814292116
સિવિન્ગોઝ, એએલ, ચાવકીન, સી., કોલાગો, ઇઇઓ, પિકેલ, વીએમ (2001). ?? નું મુખ્ય સહઅંગ્રેપીકરણ - ન્યુક્લિયસમાં opપિઓઇડ રીસેપ્ટર્સ અને ડોપામાઇન ટ્રાન્સપોર્ટર એક્કોનલ પ્રોફાઇલ. સિનેપ્સ 42, 185-192. ડોઇ: 10.1002 / syn.10005
શેચેટમેન, એચ., તાલંગબાયન, એચ., આઈલમ, ડી. (1993). ડોપામાઇન એગોનિસ્ટ ક્વિનપાયરોલ દ્વારા પ્રેરિત સંવેદનાના પર્યાવરણીય અને વર્તનશીલ ઘટકો. બિહાવ ફાર્માકોલ. 4, 405–410. doi: 10.1097/00008877-199308000-00014
શેચેટમેન, એચ., તાલંગબાયન, એચ., કેનારાન, જી., ડાઇ, એચ., આઈલમ, ડી. (1994). ડોપામાઇન એગોનિસ્ટ ક્વિનપાયરોલ અને સૂચિત કેન્દ્રીય energyર્જા નિયંત્રણ મિકેનિઝમ દ્વારા પ્રેરિત વર્તણૂકીય સંવેદનાની ગતિશીલતા. સાયકોફાર્માકોલ. (બર્લ) 115, 95-104. ડોઇ: 10.1007 / BF02244757
શેચેટમેન, એચ., સુલિસ, ડબલ્યુ., આઈલમ, ડી. (1998). ક્વિનપાયરોલ ઉંદરોમાં અનિયમિત ચેકિંગ વર્તણૂકને પ્રેરે છે: બાધ્યતા મનોગ્રસ્તિ વિકાર (ઓસીડી) નું સંભવિત પ્રાણી મોડેલ. બિહાવ ન્યુરોસી. 112, 1475-1485. ડોઇ: 10.1037 / 0735-7044.112.6.1475
શેચેટમેન, એચ., કલ્વર, કે., આઈલમ, ડી. (1999) બાધ્યતા મનોગ્રસ્તિ વિકાર (OCD) માં ડોપામાઇન સિસ્ટમોની ભૂમિકા: નવલકથા સાયકોસ્ટીમ્યુલેન્ટ-પ્રેરિત પ્રાણી મોડેલના સૂચિતાર્થ. પોલ. જે ફાર્માકોલ. 51, 55-61
શેચેટમેન, એચ., એકર્ટ્ટ, એમજે, ત્સે, ડબલ્યુએસ, બોઅર્સમા, જેટી, બોનુરા, સી એ, જેઝેડ, એમ., એટ અલ. (2001). ઓબ્સેસીવ-કમ્પલસિવ ડિસઓર્ડર (ઓસીડી) ના પ્રાણીના મ asડલ તરીકે ક્વિનપાયરોલ-સંવેદનાત્મક ઉંદરોની અનિયમિત ચેકિંગ વર્તન: ફોર્મ અને નિયંત્રણ. બીએમસી ન્યુરોસી. 2, 4. doi: 10.1186/1471-2202-2-4
શેચેટમેન, એચ., અહમારી, એસઇ, બેનિન્ગર, આરજે, આઈલમ, ડી. હાર્વે, બીએચ, એડિમન-કાલેસેન, એચ., એટ અલ. (2017). બાધ્યતા મનોગ્રસ્તિ વિકાર: પ્રાણીના મ modelsડેલોની આંતરદૃષ્ટિ. ન્યુરોસી બ્રીબહેવ રેવ. 76, 254-279. ડોઇ: 10.1016 / j.neubiorev.2016.04.019
તેજેડા, એચ.એ., બોંસી, એ. (2019) ડાયનોર્ફિન / કપ્પા-ioપિઓઇડ રીસેપ્ટર ડોપામાઇનની ગતિશીલતાનું નિયંત્રણ: નકારાત્મક લાગણીશીલ અવસ્થાઓ અને માનસિક વિકારો માટે અસર. મગજનો અનાદર 1713, 91-101. ડોઇ: 10.1016 / j.brainres.2018.09.023
તેજેડા, એચ.એ., વુ, જે., કોર્નસ્પૂન, એ.આર., પિગ્નેટેલી, એમ., કાશ્તીલિયન, વી., ક્રેશેસ, એમ. જે., એટ અલ. (2017). પાથવે- અને સેલ-વિશિષ્ટ કપ્પા-ioપિઓઇડ રીસેપ્ટર મોડ્યુલેશન ઓફ ઉત્તેજના-અવરોધ સંતુલન વિભિન્ન રીતે ગેટ્સ ડી 1 અને ડી 2 ન્યુરોન પ્રવૃત્તિને વધારે છે. ચેતાકોષ 93 (1), 147–163. doi: 10.1016 / j.neuron.2016.12.005
થomમ્પસન, એસી, ઝપાટા, એ. જસ્ટિસ, જેબી, વauન, આરએ, શાર્પ, એલજી, શીપનબર્ગ, ટીએસ (2000). કપ્પા-ioપિઓઇડ રીસેપ્ટર એક્ટિવેશન, ન્યુક્લિયસના લોકોમાં ડોપામાઇનના વપરાશમાં ફેરફાર કરે છે અને કોકેનની અસરોનો વિરોધ કરે છે. જે ન્યુરોસી. 20, 9333–9340. doi: 10.1523/JNEUROSCI.20-24-09333.2000
થomમ્પસન, ડી., માર્ટિની, એલ., વ્હિસ્લર, જેએલ (2010). માઉસ સ્ટ્રાઇટમમાં ડી 1 અને ડી 2 ડોપામાઇન રીસેપ્ટર્સનો બદલાયેલ ગુણોત્તર, કોકેઇન પ્રત્યે વર્તણૂકીય સંવેદના સાથે સંકળાયેલ છે. પ્લોસ વન 5, e11038. doi: 10.1371 / Journal.pone.0011038
યુસિલો, એ., બાઇક, જે. એચ., ર Rouગા-પોન્ટ, એફ., પિક્ટી, આર., ડીરીચ, એ., લેમ્યુર, એમ., એટ અલ. (2000). ડોપામાઇન ડી 2 રીસેપ્ટર્સના બે આઇસોફોર્મ્સના વિશિષ્ટ કાર્યો. કુદરત 408, 199-203. ડોઇ: 10.1038 / 35041572
વેન બોકસ્ટેઇલ, ઇજે, પિકેલ, વીએમ (1995). ઉંદરોના મગજમાં ન્યુક્લિયસને toમ્બેબલ્સમાં વેન્ટ્રલ ટેગમેન્ટલ એરિયા પ્રોજેક્ટમાં GABA- ધરાવતા ન્યુરોન્સ. મગજનો અનાદર 682 (1-2), 215–221. doi: 10.1016/0006-8993(95)00334-M
વેનટ વીર, એ., બેચથોલ્ટ, એજે, ઓન્વની, એસ., પોટર, ડી. વાંગ, વાય., લિયુ-ચેન, એલવાય, એટ અલ. (2013). મગજ ડોપામાઇન ન્યુરોન્સમાંથી કપ્પા-ioપિઓઇડ રીસેપ્ટર્સનું એબિલેશન એનિસિઓલિટીક જેવી અસરો ધરાવે છે અને કોકેન-પ્રેરિત પ્લાસ્ટિસિટીને વધારે છે. ન્યુરોસાયકોફોર્માકોલોજી 38 (8), 1585–1597. doi: 10.1038 / npp.2013.58
વેન્ડરશ્યુરેન, એલજે, કાલિવાસ, પીડબ્લ્યુ (2000) વર્તણૂકીય સંવેદનાના સમાવેશ અને અભિવ્યક્તિમાં ડોપામિનેર્જિક અને ગ્લુટામેટરજિક ટ્રાન્સમિશનમાં ફેરફાર: પૂર્વજ્ precાનિક અભ્યાસની નિર્ણાયક સમીક્ષા. સાયકોફાર્માકોલ. (બર્લ) 151 (2-3), 99-120. doi: 10.1007 / s002130000493
વોલ્કો, એનડી, વાઈઝ, આરએ, બેલેર, આર. (2017) ડોપામાઇન હેતુ સિસ્ટમ: ડ્રગ અને ખોરાકના વ્યસન માટેના સૂચિતાર્થ. નેટ રેવ. ન્યૂરોસી 18, 741-752. doi: 10.1038 / nrn.2017.130
વkerકર, ક્યૂડી, રૂની, એમબી, વિટમેન, આરએમ, કુહન, સીએમ (2000) ઝડપી ચક્રીય વોલ્ટેમેટ્રી દ્વારા માપવામાં આવેલા પુરુષ ઉંદર સ્ટ્રાઇટમ કરતાં ડોપામાઇનનું પ્રકાશન અને ઉપભોગ સ્ત્રીઓમાં વધારે છે. ન્યુરોસાયન્સ 95, 1061–1070. doi: 10.1016/S0306-4522(99)00500-X
વી, એસ., કુબ, જીએફ (2010) દુરૂપયોગની દવાઓ પર અસરકારક અસરોમાં ડાયનોર્ફિન κ io κઓપિઇડ સિસ્ટમની ભૂમિકા. સાયકોફાર્માકોલ. (બર્લ) 210, 121–135. doi: 10.1007/s00213-010-1825-8
વી, એસ., ઓરિઓ, એલ., ગિરમi, એસ., કેશમેન, જેઆર, કુબ, જીએફ (2009). કાપ્પા ioપિઓઇડ રીસેપ્ટર્સના નિષેધને કારણે ઉંદરોમાં કોકેઇનની વિસ્તૃત વપરાશ સાથે વધેલા કોકેનની માત્રામાં ઘટાડો થયો છે. સાયકોફાર્માકોલ. (બર્લ) 205, 565–575. doi: 10.1007/s00213-009-1563-y
વ્હિટફિલ્ડ, ટીડબ્લ્યુ, જુનિયર, શ્લોસબર્ગ, જેઈ, વી, એસ., ગોલ્ડ, એ., જ્યોર્જ, ઓ., ગ્રાન્ટ, વાય., એટ અલ. (2015). nuc ન્યુક્લિયસમાં κપિઓઇડ રીસેપ્ટર્સ મેથેમ્ફેટેમાઇન ઇનટેકના શેલ મધ્યસ્થ વૃદ્ધિ. જે ન્યુરોસી. 35 (10), 4296-4305. doi: 10.1523 / JNEUROSCI.1978-13.2015
વિલિયમ્સ, એમટી, મુગ્નો, બી., ફ્રેન્કલિન, એમ., ફેબર, એસ. (2013) બાધ્યતા મનોગ્રસ્તિ વિકારમાં લક્ષણ પરિમાણો: પ્રદર્શન અને ધાર્મિક નિવારણ સાથે ઘટના અને ઉપચારના પરિણામો. મનોવિકૃતિશાસ્ત્ર 46, 365-376. ડોઇ: 10.1159 / 000348582
વિલ્સન, સીજે, યંગ, એસજે, ગ્રોવ્સ, પીએમ (1977). સબસ્ટન્ટિયા નિગ્રામાં ન્યુરોનલ સ્પાઇક ટ્રેનોના આંકડાકીય ગુણધર્મો: કોષના પ્રકારો અને તેમની ક્રિયાપ્રતિક્રિયા. મગજનો અનાદર 136, 243–260. doi: 10.1016/0006-8993(77)90801-0
વાઈઝ, આરએ (2009). ઇનામ અને વ્યસનમાં ડોગામિન - માત્ર મેસોકોર્ટિકોલિમ્બિક માટે નિગ્રોસ્ટ્રિએટલની ભૂમિકાઓ. પ્રવાહો ન્યૂરોસી 32, 517-524. ડોઇ: 10.1016 / j.tins.2009.06.004
ઝિઓ, એલ., બેકર, જેબી (1994) પુરુષ અને સ્ત્રી ઉંદરોમાં એક્સ્ટ્રાસેલ્યુલર સ્ટ્રાઇટલ ડોપામાઇનની સાંદ્રતાનો જથ્થાત્મક માઇક્રોડાયલિસીસ નિર્ધારણ: એસ્ટ્રોસ ચક્ર અને ગોનાડેક્ટોમીની અસરો. ન્યુરોસી. લેટ. 180, 155–158. doi: 10.1016/0304-3940(94)90510-X
કીવર્ડ્સ: કપ્પા ioપિઓઇડ રીસેપ્ટર, ડોપામાઇન, અનિવાર્યતા, એમ્ફેટામાઇન, ક્વિનપાયરોલ, લોકોમોટર સંવેદના
પ્રશસ્તિપત્ર: એસ્કોબાર એડીપી, કેસોનોવા જેપી, éન્ડ્રેસ એમઇ અને ફુએન્ટિલ્બા જેએ (2020) કોપ્સ્પ્લિવ બિહેવીઅર્સમાં કપ્પા ioપિઓઇડ અને ડોપામાઇન સિસ્ટમ્સ વચ્ચે ક્રોસ્ટલેક. આગળ. ફાર્માકોલ. 11: 57. ડોઇ: 10.3389 / fphar.2020.00057
પ્રાપ્ત: 16 Octoberક્ટોબર 2019; સ્વીકૃત: 22 જાન્યુઆરી 2020;
પ્રકાશિત: 18 ફેબ્રુઆરી 2020.
દ્વારા સંપાદિત:
ગોન્ઝાલો ઇ. યેવીન્સ, ચિલી, યુનિવર્સિટી ઓફ કન્સેપ્શન
દ્વારા ચકાસાયેલ:
લુઇસ ગેરાડો એગ્યુઆયો, ચિલી, યુનિવર્સિટી ઓફ કન્સેપ્શન
હુગો તેજેડા, નેશનલ ઇન્સ્ટિટ્યૂટ Drugન ડ્રગ એબ્યુઝ (NIDA), યુનાઇટેડ સ્ટેટ્સ
સેસિલિયા સ્કોર્ઝા, ઇન્સ્ટિટ્યુટો ડી ઈન્વેસ્ટિક્વન્સ બાયોલóજિકિસ ક્લેમેન્ટે ઇસ્ટ (બલ (IIBCE), ઉરુગ્વે
ક Copyrightપિરાઇટ 2020 XNUMX એસ્કોબાર, કેસોનોવા, éન્ડ્રેસ અને ફુએન્ટાલ્બા. ની શરતો હેઠળ વિતરિત આ એક ખુલ્લો-accessક્સેસ લેખ છે ક્રિએટિવ કૉમન્સ એટ્રિબ્યુશન લાયસન્સ (સીસી દ્વારા). અન્ય ફોરમમાં ઉપયોગ, વિતરણ અથવા પ્રજનનની પરવાનગી છે, સિવાય કે મૂળ લેખક (ઓ) અને કૉપિરાઇટ માલિક (ઓ) ને શ્રેય આપવામાં આવે છે અને સ્વીકારેલ શૈક્ષણિક અભ્યાસ અનુસાર, આ જર્નલમાં મૂળ પ્રકાશનનો ઉલ્લેખ કરવામાં આવે છે. કોઈ ઉપયોગ, વિતરણ અથવા પ્રજનનની પરવાનગી નથી જે આ શરતોનું પાલન કરતી નથી.
* પત્રવ્યવહાર: જોસ એન્ટોનિયો ફુએન્ટાલબ, [ઇમેઇલ સુરક્ષિત]
†ઓરસીડ: જોસ એન્ટોનિયો ફુએન્ટાલ્બા, orcid.org/0000-0003-0775-0675