Athugasemdir: Þar sem síðari rannsóknir munu leiða í ljós að DeltaFosB er algengi sameindarofi fyrir bæði fíkniefna- og atferlisfíkn. Það er umritunarstuðull sem þýðir að það hefur áhrif á hvaða gen eru kveikt eða óvirk. Eins og fram kemur annars staðar ræna ávanabindandi lyf aðeins venjulegum aðferðum. Þess vegna er kjánalegt að gefa í skyn að hegðunarfíkn geti ekki verið til.
Proc Natl Acad Sci US A. 2001 September 25; 98 (20): 11042-11046.
doi: 10.1073 / pnas.191352698.
Eric J. Nestler *, Michel Barrot og David W. Self
Department of Psychiatry og Center for Basic Neuroscience, University of Texas Southwestern Medical Center, 5323 Harry Hines Boulevard, Dallas, TX 75390-9070
Abstract
Langlífi sumra hegðunarafbrigðanna sem einkenna eiturlyfjafíkn hefur bent til þess að reglugerð um taugaþenslu geti verið þátt í því ferli þar sem eiturlyf misnotkun veldur fíkniefni. Égncreasing vísbendingar benda til þess að uppskrift þáttur ΔFosB táknar eitt kerfi sem misnotkun lyfja framleiðir tiltölulega stöðugar breytingar í heila sem stuðla að fíkniefnistöku. ΔFosB, sem er meðlimur Fos fjölskyldunnar á uppskriftarþáttum, safnast saman innan undirhóps taugafruma í kjarna accumbens og dorsal striatum (heila svæði mikilvæg fyrir fíkn) eftir endurtekna gjöf margs konar misnotkunarlyfja. Svipuð uppsöfnun ΔFosB á sér stað eftir þvingunarhlaup, sem bendir til þess að ΔFosB megi safnast til að bregðast við mörgum tegundum af þvingunarhegðun. Mikilvægt er að ΔFosB viðvarandi í taugafrumum í tiltölulega langan tíma vegna óvenjulegs stöðugleika þess. Þess vegna táknar ΔFosB sameindakerfi sem gæti byrjað og síðan viðhaldið breytingum á genþrýstingi sem er viðvarandi löngu eftir að lyfjaáhrif hætta. Rannsóknir á örvandi erfðabreyttum músum sem yfirfæra annaðhvort ΔFosB eða ríkjandi neikvæða hindrun próteinsins veita beinan sönnun þess að ΔFosB veldur aukinni næmi fyrir hegðunaráhrifum misnotkunarlyfja og hugsanlega aukinni eiturverkandi hegðun. Þessi vinna styður þá skoðun að ΔFosB virkar sem gerð viðvarandi "sameinda rofi" sem smám saman breytir bráðum eiturverkunum í tiltölulega stöðuga aðlögun sem stuðlar að langvarandi tauga- og hegðunarplástrum sem byggist á fíkn.
Fíknannsóknir eru lögð áhersla á að skilja flóknar leiðir þar sem eiturlyf misnotkunar breytir heilanum til að valda hegðunartruflunum sem einkennast af fíkn. Eitt af mikilvægum áskorunum á þessu sviði er að greina tiltölulega stöðug lyfjatengda breytingar í heila til að taka tillit til þeirra hegðunarvandamála sem eru sérstaklega langvarandi. Til dæmis getur mannlegur fíkill verið í aukinni hættu á afturfalli jafnvel eftir margra ára fráhvarf.
Stöðugleiki þessara hegðunarafbrigða hefur leitt til þess að þeir megi miðla, að minnsta kosti að hluta, með breytingum á genþrýstingi (1-3). Samkvæmt þessu sjónarmiði truflar endurtekin útsetning fyrir eiturlyfjasýki ítrekað sending á tilteknum synapses í heila sem eru viðkvæm fyrir lyfinu. Slíkar truflanir merki að lokum með innanfrumukrabba í kjarna, þar sem þeir byrja fyrst og síðan viðhalda breytingum á tjáningu tiltekinna gena. Helstu fyrirkomulag þar sem merki umleiðslunarleiðir hafa áhrif á genþrýsting er að mæla umritunarþætti, prótein sem bindast reglulegum svæðum gena og breyta umskrift þeirra.
Eitt markmið um fíknannsóknir hefur því verið að greina áletrunarþætti sem eru breytt í heila svæðum sem hafa áhrif á fíkn eftir langvarandi notkun lyfja af misnotkun. Nokkrar slíkar umritunarþættir hafa verið skilgreindir á síðasta áratug (1-6). Áherslan á þessari umfjöllun er á einum afskriftarþætti sem kallast ΔFosB.
Innleiðing ΔFosB með fíkniefni
ΔFosB, kóðað af fosB geninu, er meðlimur Fos fjölskyldunnar á uppskriftarþáttum, sem einnig inniheldur c-Fos, FosB, Fra1 og Fra2 (7). Þessar Fos fjölskyldu prótein heterodimerize með Jun fjölskyldu prótein (c-júní, JunB eða JunD) til að mynda virk AP-1 (virkjunarprótín-1) umritunarmörk sem bindast AP-1 stöðum (samruna röð: TGAC / GTCA) verkefnisstjórar tiltekinna gena til að stjórna uppskrift þeirra.
Þessar Fos fjölskylduprótein eru framkölluð hratt og tímabundið í sérstökum heila svæðum eftir bráða notkun margra fíkniefnanefna (mynd 1) (8-11). Áberandi svæði eru kjarninn og dorsal striatum sem eru mikilvægir miðlarar á hegðunarvandamálum við lyfin, einkum þeirra gefandi og staðbundnar virkjunaráhrifa (12, 13). Þessar prótein koma aftur til grunnþéttni innan klukkustunda eftir lyfjagjöf.
Mynd 1
Áætlun sem sýnir smám saman uppsöfnun ΔFosB móti hraðri og tímabundinni örvun annarra Fos fjölskyldu próteina til að bregðast við fíkniefnum. (A) Örvunarskírteinið sýnir mismunadreifingu þessara ýmissa próteina með bráðri örvun (1-2 klst. Eftir stakan útsetningu lyfja) í samanburði við langvarandi örvun (1 dagur eftir endurtekna lyfjaáhrif). (B) Nokkrir öldur Fos-eins próteina (samanstendur af c-Fos (52- til 58-kDa ísóformum), FosB (46- til 50-kDa ísóforms), ΔFosB (33-kDa ísóform) og Fra1 eða Fra2 40 kDa)] er framkallað í kjarnaabrúsa og dorsal striatal taugafrumum með bráðum lyfjagjöf um misnotkun. Einnig er framkallað lífefnafræðilega breytt ísóform af ΔFosB (35-37 kDa); Þeir eru líka framkallaðir (þó á litlum vöxtum) eftir bráða lyfjagjöf, en halda áfram í heilanum í langan tíma vegna stöðugleika þeirra. (C) Með endurtekinni (td tvisvar á dag) lyfjagjöf, veldur hver bráð hvati lágt stig af stöðugu ΔFosB ísóformunum, sem er gefið til kynna með lægri mengi af skarastum línum sem gefa til kynna ΔFosB af völdum hvers bráðrar hvata. Niðurstaðan er smám saman aukning á heildarmagni ΔFosB með endurteknum áreiti meðan á langvarandi meðferð stendur, sem er sýnt af vaxandi stigi á grafinu.
Mjög mismunandi svör sjást eftir langvarandi gjöf fíkniefnaneyslu (mynd 1). Lífefnafræðilega breyttar ísóformar ΔFosB (sameindamassi 35-37 kDa) safnast innan sömu heilaþátta eftir endurtekna útsetningu lyfsins, en allir aðrir Fos fjölskyldumeðlimir sýna umburðarlyndi (það er minni virkni samanborið við upphaf lyfjaskemmda). Slík uppsöfnun ΔFosB hefur komið fram fyrir kókaín, morfín, amfetamín, alkóhól, nikótín og fencyclidine (11, 14-18). Sumar vísbendingar eru um að þessi örvun sé sértæk fyrir dúkur / efnin P-innihaldsefnið af miðlungs hreinum taugafrumum sem staðsettir eru í þessum heila svæðum (15, 17), þótt meira sé þörf til að koma þessu með vissu. 35- til 37-kDa ísóforma ΔFosB dregur aðallega úr JunD til að mynda virkan og langvarandi AP-1 flókin innan þessara heila svæðum (19, 20). Þessar ΔFosB ísóformar safnast saman við langtímameðferð vegna vegna óvenju langvarandi helmingunartíma (21) og því halda áfram í taugafrumum í að minnsta kosti nokkrar vikur eftir að lyfjagjöf er hætt. Það er áhugavert að hafa í huga að þessar ΔFosB ísóformar eru mjög stöðugar vörur strax snemma gen (fosB). Stöðugleiki ΔFosB ísóformanna veitir nýtt sameindakerfi, þar sem lyfjafræðilegar breytingar á genþrýsting geta haldið áfram þrátt fyrir tiltölulega langan tíma frá meðhöndlun lyfja.
Þrátt fyrir að kjarninn sé áberandi gegnir mikilvægu hlutverki í gefandi áhrifum misnotkunarlyfja, er talið að það virki venjulega með því að stjórna svörun náttúrulegra efna, svo sem mat, drykk, kynlíf og félagsleg samskipti (12, 13). Þar af leiðandi er mikil áhugi á hugsanlegu hlutverki heila svæðisins í öðrum þvingunarhegðunum (td meinafræðilegu ofsöfnun, fjárhættuspil, æfing osfrv.). Af þessum sökum skoðuðum við hvort ΔFosB sé stjórnað í dýralíkani af þvingunarhlaupi. Reyndar eru stöðugar 35- til 37-kDa ísóformar ΔFosB framkallað sértækt innan kjarna accumbens hjá rottum sem sýna þvingunarhreyfingu. †
Lífefnafræðileg einkenni stöðugra ΔFosB ísóforma
Eins og getið er um hér að framan, sýna ΔFosB ísóform sem safnast eftir langvarandi gjöf lyfja sem eru misnotuð eða þvinguð í gangi sýna mólmassa 35-37 kDa. Þau geta verið frábrugðin 33-kDa ísóformi ΔFosB sem er framkallað hratt en tímabundið eftir einn útsetningu lyfja (mynd 1) (14, 19, 22). Núverandi sönnunargögn benda til þess að 33-kDa ísóformið sé innfæddur formur próteinsins, sem er breytt til að mynda stöðugri 35- til 37-kDa vörur (19, 21). Hins vegar hefur eðli lífefnafræðilegra breytinga sem umbreytir óstöðugan 33-kDa ísóform í stöðugar 35- til 37-kDa ísóformana verið óskýr. Það hefur verið í huga að fosfórýlering getur átt sér stað (11). Til dæmis er örvun á ΔFosB dregið úr í músum sem sakna DARPP-32, storkueinkað prótein (23, 24). Vegna þess að DARPP-32 stjórnar hvatvirkni próteinfosfatasa-1 og próteinkínasa A (25, 26) bendir krafan fyrir þetta prótein fyrir eðlilega uppsöfnun á stöðugu ΔFosB ísóformum hugsanleg hlutverk fosfórunar við kynslóð þessara stöðugra vara.
Hlutverk ΔFosB í hegðunar plasticity við misnotkunartilfelli
Innsýn í hlutverk ΔFosB í fíkniefni hefur að mestu komið frá rannsókn á erfðabreyttum músum þar sem hægt er að örva ΔFosB sértækt innan kjarnans og annarra fóstursvæða fullorðinsdýra (27, 28). Mikilvægt er að þessi mýs ofmeta ΔFosB sértækt í díhormón / efninu P-innihaldsefninu, sem eru örlítið taugafrumur, þar sem lyfin eru talin örva próteinið. Hegðunarfíknin af ΔFosB-overexpressing músunum, sem á margan hátt líkist dýrum eftir langvarandi lyfjameðferð, er tekin upp í töflu 1. Músasýningin aukið staðbundna svörun við kókaíni eftir bráða og langvinnan gjöf (28). Þeir sýna einnig aukið næmi fyrir launandi áhrifum kókaíns og morfíns í staðbundnar prófanir (11, 28) og mun sjálfstætt stjórna lægri skammta af kókaíni en littermates sem ekki yfirfæra ΔFosB. ‡ Hins vegar sýna þessi dýr venjulega kælivökva næmi fyrir kókaíni og eðlilegt staðbundið nám í Morris vatninu völundarhúsinu (28). Tþessar upplýsingar benda til þess að ΔFosB auki næmi dýra fyrir kókaíni og kannski öðrum misnotkunarlyfjum og geti táknað fyrirkomulag fyrir tiltölulega langvarandi næmi fyrir lyfjunum.
 striatum
striatum
| Aukin virkjun á staðsetningum til að bregðast við bráðri og endurtekinni kókaín gjöf. |
| Aukin gefandi svör við kókaíni og morfíni í staðbundnar prófanir. |
| Aukin sjálfs gjöf lágskammta kókaíns. |
| Aukin áhugi á kókaíni í stigvaxandi prófi. |
| Aukin kvíðastillandi viðbrögð við áfengi. |
| Aukin þvingunarhreyfing. |
Byggt á gögnum í refs. 28 og 29.† ‡ §¶
Hegðunar plasticity miðlað af ΔFosB í kjarna accumbens-dorsal striatum
In viðbót, eru forkeppni sannanir fyrir því að áhrif ΔFosB geta verið vel umfram reglur um næmni lyfja í sjálfu sér til flóknari hegðun sem tengist fíkninni. Mýs sem tjá ΔFosB vinna erfiðara með að gefa kókaíni sjálfstætt, í sjálfvirkri prófunarferliggesting sem ΔFosB getur næmt dýrum í hvatningarvaldandi eiginleika kókaíns og þannig leitt til tilhneigingu til að koma aftur eftir að lyf hefur dregið úr1. ‡ ΔFosB tjáandi mýs sýna einnig aukið kvíðastillandi áhrif áfengis, § svipgerð sem hefur verið tengd aukinni áfengisneyslu hjá mönnum. Saman benda þessar fyrstu niðurstöður að ΔFosB, auk þess að auka næmi fyrir fíkniefnum, framleiðir eigindlegar breytingar á hegðun sem stuðla að eiturverkandi hegðun. Þannig getur ΔFosB virkað sem viðvarandi "sameindarrof" sem hjálpar til við að hefja og viðhalda mikilvægum þáttum hávaða. Mikilvæg spurning samkvæmt þessari rannsókn er hvort ΔFosB uppsöfnun við lyfjameðferð stuðlar að eiturverkandi hegðun eftir langan afturköllunartíma, jafnvel þótt ΔFosB gildi hafi verið eðlileg (sjá hér að neðan).
Adult mýs sem yfirfæra ΔFosB valkvætt innan kjarna accumbens og dorsal striatum sýndu einnig meiri þvingunarhreyfingu samanborið við fullorðinsfræðslu. Þessar athuganir vekja áhugaverðan möguleika á að ΔFosB uppsöfnun innan þessara taugafrumna þjónar almennt hlutverki í myndun og viðhaldi venja minningar og þvingunar hegðun, kannski með því að styrkja virkni tauga hringrásar þar sem þessir taugafrumur virka.
ΔFosB safnast í ákveðnum heila svæðum utan kjarna accumbens og dorsal striatum eftir langvarandi útsetningu fyrir kókaíni. Áberandi meðal þeirra svæði eru amygdala og medial prefrontal heilaberki (15). Helstu markmið núverandi rannsókna er að skilja framlag ΔFosB framkalla á þessum svæðum til fíkniefna.
Fyrri vinna við fosB útsláttar mýs leiddi í ljós að þessi dýr ná ekki að mynda næmi fyrir hreyfisáhrifum kókaíns, sem er í samræmi við niðurstöður ΔFosB-oftjáandi músa sem nefndar eru hér að ofan (22). Hins vegar sýndu fosB stökkbrigðin aukið næmi fyrir bráðum áhrifum kókaíns, sem er í ósamræmi við þessar aðrar niðurstöður. Túlkun niðurstaðna með fosB stökkbrigðunum er þó flókin af því að þessi dýr skortir ekki aðeins ΔFosB, heldur einnig FosB í fullri lengd. Ennfremur skortir stökkbrigðin bæði prótein um heilann og frá fyrstu þroskastigum. Reyndar styður nýleg vinna ályktanir frá ΔFosB ofþjöppandi músum: framkallandi oftjáningu styttrar stökkbreytingar af c-Jun, sem virkar sem ráðandi neikvæður andstæðingur ΔFosB, sértækt í nucleus accumbens og dorsal striatum sýnir skert næmi fyrir gefandi áhrifum kókaíns. .¶ Þessar niðurstöður leggja áherslu á varúð sem verður að nota við túlkun á niðurstöðum músa með stökkbreytingar og sýna fram á mikilvægi músa með hvata og frumutegundarsértækar stökkbreytingar í rannsóknum á plastleika í heila fullorðinna.
Markmið fyrir ΔFosB
Vegna þess að ΔFosB er umritunarþáttur, veldur líklega prótein hegðunargleði með breytingum á tjáningu annarra gena. ΔFosB er myndað með annarri splicing á fosB geninu og skortir hluta af C-enda transactivation léninu sem er til staðar í fullri lengd FosB. Þess vegna var upphaflega lagt til að ΔFosB virki sem transkriptrar (29). Hins vegar hefur vinna í frumurækt sýnt fram á að ΔFosB getur annaðhvort valdið eða stutt AP-1-miðlað uppskrift eftir því hvaða tilteknu AP-1 síða er notað (21, 29-31). FosB í fullri lengd hefur sömu áhrif og ΔFosB á ákveðnum stökkbrigðum, en mismunandi áhrif á aðra. Nauðsynlegt er að vinna að því að skilja kerfi sem liggja að baki þessum fjölbreyttu aðgerðum ΔFosB og FosB.
Hópurinn okkar hefur notað tvær aðferðir til að greina markgen fyrir ΔFosB. Ein er framboð gena nálgun. Við töldum upphaflega α-amínó-3-hýdroxý-5-metýl-4-ísoxasólprópíónsýru (AMPA) glútamatviðtaka sem afleidd markmið, miðað við mikilvægu hlutverki glútamatergískrar smitunar í kjarnanum. Vinna til þessa hefur gefið til kynna að ein sérstök AMPA glútamatviðtaka undireining, GluR2, geti verið bona fide markmið fyrir ΔFosB (mynd 2). GluR2 tjáning, en ekki tjáning annarra AMPA viðtaka undireininga, er aukin í nucleus accumbens (en ekki dorsal striatum) við oftjáningu ΔFosB (28) og tjáning ráðandi neikvæðrar stökkbreytingar dregur úr getu kókaíns til að framkalla próteinið .¶ Að auki inniheldur hvatamaður GluR2 genins samstöðu AP-1 stað sem bindur ΔFosB (28). Yfirtjáning á GluR2 í kjarna accumbens, með því að nota veiru-miðlað genaflutning, eykur næmi dýrs fyrir gefandi áhrif kókaíns og hermir þar með eftir hluta af svipgerðinni sem sést í ΔFosB-tjáandi músum (28). Framleiðsla á GluR2 gæti skýrt frá skertri raflífeðlisfræðilegri næmni taugafrumna í kjarna fyrir AMPA viðtakaörva eftir langvarandi gjöf kókaíns (32), vegna þess að AMPA viðtakar sem innihalda GluR2 sýna minni heildarleiðni og skerta Ca2 + gegndræpi. Minni svörun þessara taugafrumna við spennandi aðföngum getur þá aukið viðbrögð við misnotkun lyfs. Hvernig sem dópamínvirk og glútamatergísk merki í kjarna accumbens stjórna ávanabindandi hegðun eru enn óþekkt; til þess þarf taugakerfisskilningsstig, sem ekki er enn í boði.
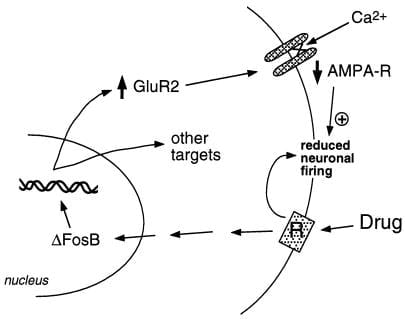
Mynd 2
AMPA glútamatviðtakaeiningin, GluR2, er afleit markmið fyrir ΔFosB. Sýnt er hvernig ΔFosB miðill örvun GluR2 getur breytt lífeðlisfræðilegri svörun taugafrumna í kjarna og leitt til næmra viðbragða við misnotkun lyfja. Samkvæmt þessu kerfi framleiða misnotkun lyf bráða styrkjandi áhrif þeirra með því að hindra taugafrumur í kjarna. Við endurtekna útsetningu framkalla lyfin ΔFosB, sem stýrir fjölmörgum markgenum, þar með talið GluR2. Þetta eykur hlutfall AMPA viðtaka (AMPA-R) á taugafrumum í kjarna sem innihalda GluR2 undireininguna sem veldur minni AMPA straumi og minni Ca2 + straumi. Þessi skerta spennu gæti gert taugafrumurnar næmari fyrir bráðum hamlandi áhrifum lyfjanna og þar með fyrir styrkjandi áhrif lyfjanna.
Annað formlegt markmið fyrir ΔFosB er genið sem kóðar dynorphin. Eins og fram hefur komið er dynorphin gefið upp í undirhóp kjarnans accumbens miðlungs spiny neurons sem sýna framköllun ΔFosB. Dynorphin virðist virka í intercellular feedback loop: losunin hindrar dopamínvirka taugafrumurnar sem innihalda miðlungs hryggjarlið, með κ ópíóíðviðtökum sem eru til staðar á dopamínvirka taugaskiptum í kjarnanum og einnig á frumum og dendritum í ventral tegmental svæðinu (Mynd 3) (33-35). Þessi hugmynd er í samræmi við getu κ viðtakaörva, við gjöf í annað hvort þessara tveggja heilahluta, til að minnka eiturlyfd (35).
Recent vinnu hefur gefið til kynna að ΔFosB dregur úr tjáningu dynorphins, sem gæti stuðlað að aukinni verðlaunakerfi sem sést með ΔFosB örvun. Athyglisvert er að önnur lyfjafræðilegur umritunarþáttur, CREB (cAMP svarhluta bindandi prótein) (2, 3), hefur hið gagnstæða áhrif: það veldur þunglyndi tjáningu í kjarnanum accumbens og dregur úr gefandi eiginleika kókaíns og morfíns (4). **
Bvegna þess að lyfjaeinkenni örvunarvirkni CREB dreifist hratt eftir lyfjagjöf, svo gagnkvæm gjöf dynorphins með CREB og ΔFosB gæti útskýrt gagnkvæma hegðunarbreytingar sem eiga sér stað á fyrstu og síðari stigum afturkalls, með neikvæðum tilfinningalegum einkennum og minnkaðri næmi í lyfjum í upphafi fasa af afturköllun og næmni fyrir gefandi og hvatandi hvatningaráhrif lyfja sem ráða yfir síðari tímapunkti.
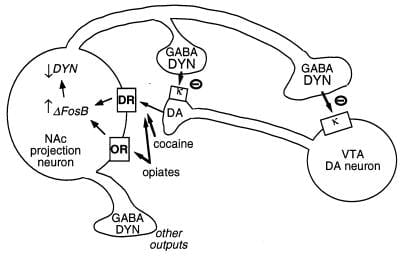
Mynd 3
Dynorphin er formlegt markmið fyrir ΔFosB. Sýnt er taugafruma (VTA) dopamín (DA) taugafruma sem innheimtir kjarna accumbens (NAc) GABAergic vörpun taugafrumum sem lýsir dynorphin (DYN). Dynorphin þjónar viðbrögðarkerfi í þessari hringrás: Dynorphin, losað úr skautum NAc taugafrumum, virkar á κ opioid viðtaka sem staðsettir eru á taugaskermum og frumum líkama DA taugafrumna til að hamla starfsemi þeirra. ΔFosB, með því að hamla dynorphin tjáningu, getur dregið úr þessum endurgjöfslóða og aukið verðandi lyf misnotkunar. Ekki er sýnt fram á gagnkvæm áhrif CREB á þessu kerfi: CREB eykur tjáningu dynorphins og dregur þannig úr ávinningi eiginleika misnotkunarlyfja (4). GABA, Y-amínósmjörsýra; DR, dópamínviðtaka; OR, ópíóíð viðtaka.
Önnur nálgunin sem notuð er til að bera kennsl á gen gena fyrir ΔFosB felur í sér DNA örflagsgreiningu. Óleiðanleg tjáning á ΔFosB eykur eða dregur úr tjáningu fjölmargra gena í kjarna accumbens (36). Þrátt fyrir að töluverð vinna sé nú nauðsynleg til að staðfesta hvert þessara gena sem lífeðlisfræðileg markmið ΔFosB og til að skilja framlag þeirra til fíkniefnisgerðarinnar, virðist eitt mikilvægt markmið vera Cdk5 (sýklín háð kínasi-5). Þannig var Cdk5 upphaflega auðkenndur sem ΔFosB-stjórnaður með því að nota örflokka og síðar sýnt fram á það í kjarna accumbens og dorsal striatum eftir langvarandi gjöf kókaíns (37). ΔFosB virkjar cdk5 genið um AP-1 stað sem er til staðar innan hvatamanns erfða (36). Saman styðja þessi gögn kerfi þar sem kókaín framkallar Cdk5 tjáningu á þessum heilasvæðum um ΔFosB. Framleiðsla á Cdk5 virðist breyta dópamínvirkum boðefnum að minnsta kosti að hluta til með aukinni fosfórun á DARPP-32 (37), sem er breytt úr hemli próteinsfosfatasa-1 í hemil á próteinkínasa A við fosfórun þess með Cdk5 (26).
Hlutverk ΔFosB í að miðla "varanlegri" plasticity við misnotkunartæki
Þótt ΔFosB merki sé tiltölulega langvarandi, er það ekki varanlegt. ΔFosB lækkar smám saman og er ekki lengur hægt að greina í heilanum eftir að 1-2 er tekið af lyfjameðferð, jafnvel þó að ákveðin hegðunartruflun sé í langan tíma. Þess vegna virðist ΔFosB í sjálfu sér ekki vera fær um að miðla þessum semipermanent hegðunartruflunum. Erfiðleikarnir við að finna sameindaaðlögunina sem liggja að baki hinna mjög stöðuga hegðunarbreytingar í tengslum við fíkn er hliðstæð þeim áskorunum sem blasa við í nám og minni. Þrátt fyrir að það séu glæsileg frumu- og sameindalíkön af nám og minni, hefur það ekki verið mögulegt að greina sameinda- og frumnauðlög sem eru nægilega langvarandi til að taka tillit til mjög stöðugar hegðunar minningar. Reyndar er ΔFosB langlífast aðlögun sem vitað er um að eiga sér stað í heila fullorðinna, ekki aðeins til að bregðast við misnotkun lyfja, heldur einnig við allar aðrar truflanir (sem ekki fela í sér meiðsli). Tvær tillögur hafa þróast, bæði á sviði fíknar og náms og minni, til að gera grein fyrir þessu misræmi.
Einn möguleiki er að tímabundin breyting á genþrýstingi, svo sem þeim sem miðlað er með ΔFosB eða öðrum uppskriftarþáttum (td CREB), getur miðlað fleiri langvarandi breytingum á taugafrumumyndun og synaptic uppbyggingu. Til dæmis, aukning á þéttleika dendritic spines (einkum aukning á tveir-headed spines) fylgir aukin verkun glutamatergic synapses í pýramíddar taugafrumum með hippocampal við langvarandi stækkun (38-40) og samhliða aukinni hegðunarvandamál við kókaín sem miðlað er á vettvangi miðlungs taugaþrenginga í kjarnanum (41). Ekki er vitað hvort slíkar skipulagsbreytingar séu nægilega langvarandi til að taka tillit til mjög stöðugra breytinga á hegðun, þótt hið síðarnefnda haldist í að minnsta kosti 1 mánaðar eftir að lyfið hefur verið hætt. Nýlegar vísbendingar vekur möguleika á því að ΔFosB og örvun þess Cdk5 er eini miðillinn af lyfjafræðilegum breytingum á synaptic uppbyggingu í kjarna accumbens (mynd 4). ‡ Þannig kemur innrennsli Cdk5 hemils í kjarnann í veg fyrir að hæfni endurtekinna kókaínviðtaka til að auka dendritic hryggþéttni í þessu svæði. Þetta er í samræmi við þá skoðun að Cdk5, sem er auðgað í heilanum, stjórnar taugauppbyggingu og vexti (sjá refs. 36 og 37). Það er mögulegt, þó að það sé ekki sannað, að slíkar breytingar á taugafræðilegu formgerðinni gætu umfram ΔFosB merki sjálft.
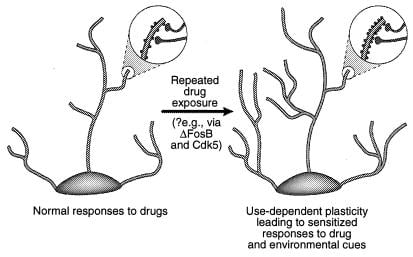
Mynd 4
Stjórnun dendrítískrar uppbyggingar með misnotkun lyfja. Sýnt er stækkun dendrítitrés taugafrumu eftir langvarandi útsetningu fyrir misnotkun lyfs, eins og sést hefur með kókaíni í kjarna kjölfestu og heilaberki fyrir framan (41). Stækkunarsvæðin sýna aukningu á dendritískum hryggjum, sem sagt er að eigi sér stað í tengslum við virkjar taugaenda. Þessari aukningu á þéttni hryggjarliðar má miðla um ΔFosB og afleidda framköllun Cdk5 (sjá texta). Slíkar breytingar á dendrítískri uppbyggingu, sem eru svipaðar þeim sem komu fram í sumum lærdómslíkönum (td langtímavirkni), gætu haft milligöngu um langvarandi næm viðbrögð við misnotkun lyfja eða umhverfisvísum. [Afritað með leyfi viðskrh. 3 (Copyright 2001, Macmillian Magazines Ltd.)].
Annar möguleiki er að tímabundin örvun uppritunarþáttar (td ΔFosB, CREB) leiðir til fleiri varanlegra breytinga á genatjáningu með breytingu á chromatin. Þessar og margar aðrar uppskriftarþættir teljast virkja eða bæla uppskrift á miða geninu með því að stuðla að asetýleringunni eða deacetyleringen í históni í nágrenni gensins (42). Þrátt fyrir að slík asetýlering og deacetylering históns geti komið fram mjög hratt er hugsanlegt að ΔFosB eða CREB gæti valdið langvarandi aðlögun í ensímatækinu sem stýrir histónsetýlerun. ΔFosB eða CREB getur einnig stuðlað að langvarandi breytingum á genþrýstingi með því að stjórna öðrum breytingum á chromatin (td DNA eða histónmetýleringu) sem hafa verið fólgin í varanlegri breytingu á genskriftir sem eiga sér stað við þróun (sjá refs. 42 og 43) . Þrátt fyrir að þessar möguleikar séu íhugandi gætu þau búið til kerfi þar sem tímabundin aðlögun að misnotkunartilvikum (eða einhverjum öðrum truflunum) leiða til í raun lífshættulegar afleiðingar.
Heimildir
- ↵
- Nestler EJ,
- Vona að BT,
- Widnell KL
(1993) Neuron 11: 995-1006.
CrossRefMedlineweb af vísindum
- ↵
- Berke JD,
- Hyman SE
(2000) Neuron 25: 515-532.
CrossRefMedlineweb af vísindum
- ↵
- Nestler EJ
(2001) Nat Rev Neurosci 2: 119-128.
CrossRefMedlineweb af vísindum
- ↵
- Carlezon WA Jr,
- Thome J,
- Olson VG,
- Lane-Ladd SB,
- Brodkin ES,
- Hiroi N,
- Duman RS,
- Neve RL,
- Nestler EJ
(1998) Vísindi 282: 2272-2275.
-
- O'Donovan KJ,
- Tourtellotte WG,
- Millbrandt J,
- Baraban JM
(1999) Stefna Neurosci 22: 167-173.
- ↵
- Mackler SA,
- Korutla L,
- Cha XY,
- Koebbe MJ,
- Fournier KM,
- Bowers MS,
- Kalivas PW
(2000) J Neurosci 20: 6210-6217.
- ↵
- Morgan JI,
- Curran T
(1995) Stefna Neurosci 18: 66-67.
- ↵
- Young ST,
- Porrino LJ,
- Iadarola MJ
(1991) Proc Natl Acad Sci USA 88: 1291-1295.
-
- Graybiel AM,
- Moratalla R,
- Robertson HA
(1990) Proc Natl Acad Sci USA 87: 6912-6916.
-
- Vona að B,
- Kosofsky B,
- Hyman SE,
- Nestler EJ
(1992) Proc Natl Acad Sci USA 89: 5764-5768.
- ↵
- Kelz MB,
- Nestler EJ
(2000) Curr Opin Neuról 13: 715-720.
CrossRefMedlineweb af vísindum
- ↵
- Koob GF,
- Sanna PP,
- Bloom FE
(1998) Neuron 21: 467-476.
CrossRefMedlineweb af vísindum
- ↵
- Vitur RA
(1998) Lyf Alcohol Dependence 51: 13-22.
CrossRefMedlineweb af vísindum
- ↵
- Vona að BT,
- Nye HE,
- Kelz MB,
- Sjálf DW,
- Iadarola MJ,
- Nakabeppu Y,
- Duman RS,
- Nestler EJ
(1994) Neuron 13: 1235-1244.
- ↵
- Nye H,
- Vona að BT,
- Kelz M,
- Iadarola M,
- Nestler EJ
(1995) J Pharmacol Exp Ther 275: 1671-1680.
-
- Nye HE,
- Nestler EJ
(1996) Mol Pharmacol 49: 636-645.
- ↵
- Moratalla R,
- Elibol B,
- Vallejo M,
- Graybiel AM
(1996) Neuron 17: 147-156.
CrossRefMedlineweb af vísindum
- ↵
- Pich EM,
- Pagliusi SR,
- Tessari M,
- Talabot-Ayer D,
- Hooft van Huijsduijnen R,
- Chiamulera C
(1997) Vísindi 275: 83-86.
- ↵
- Chen JS,
- Nye HE,
- Kelz MB,
- Hiroi N,
- Nakabeppu Y,
- Vona að BT,
- Nestler EJ
(1995) Mol Pharmacol 48: 880-889.
- ↵
- Hiroi N,
- Brown J,
- Þér H,
- Saudou F,
- Vaidya VA,
- Duman RS,
- Greenberg ME,
- Nestler EJ
(1998) J Neurosci 18: 6952-6962.
- ↵
- Chen J,
- Kelz MB,
- Vona að BT,
- Nakabeppu Y,
- Nestler EJ
(1997) J Neurosci 17: 4933-4941.
- ↵
- Hiroi N,
- Brown J,
- Haile C,
- Þér H,
- Greenberg ME,
- Nestler EJ
(1997) Proc Natl Acad Sci USA 94: 10397-10402.
- ↵
- Fienberg AA,
- Hiroi N,
- Mermelstein P,
- Song WJ,
- Snyder GL,
- Nishi A,
- Cheramy A,
- O'Callaghan JP,
- Miller D,
- Cole DG,
- et al.
(1998) Vísindi 281: 838-842.
- ↵
- Hiroi N,
- Feinberg A,
- Haile C,
- Greengard P,
- Nestler EJ
(1999) Eur J Neurosci 11: 1114-1118.
CrossRefMedlineweb af vísindum
- ↵
- Greengard P,
- Allen PB,
- Nairn AC
(1999) Neuron 23: 435-447.
CrossRefMedlineweb af vísindum
- ↵
- Bibb JA,
- Snyder GL,
- Nishi A,
- Yan Z,
- Meijer L,
- Fienberg AA,
- Tsai LH,
- Kwon YT,
- Girault JA,
- Czernik AJ,
- et al.
(1999) Náttúra (London) 402: 669-671.
- ↵
- Chen JS,
- Kelz MB,
- Zeng GQ,
- Sakai N,
- Steffen C,
- Shockett PE,
- Picciotto M,
- Duman RS,
- Nestler EJ
(1998) Mol Pharmacol 54: 495-503.
- ↵
- Kelz MB,
- Chen JS,
- Carlezon WA,
- Whisler K,
- Gilden L,
- Beckmann AM,
- Steffen C,
- Zhang YJ,
- Marotti L,
- Self SW,
- et al.
(1999) Náttúra (London) 401: 272-276.
- ↵
- Dobrazanski P,
- Noguchi T,
- Kovary K,
- Rizzo CA,
- Lazo PS,
- Bravo R
(1991) Mol Cell Biol 11: 5470-5478.
-
- Nakabeppu Y,
- Nathans D
(1991) Cell 64: 751-759.
CrossRefMedlineweb af vísindum
- ↵
- Yen J,
- Visku RM,
- Tratner ég,
- Verma IM
(1991) Proc Natl Acad Sci USA 88: 5077-5081.
- ↵
- White FJ,
- Hu XT,
- Zhang XF,
- Wolf ME
(1995) J Pharmacol Exp Ther 273: 445-454.
- ↵
- Hyman SE
(1996) Neuron 16: 901-904.
-
- Kreek MJ
(1997) Pharmacol Biochem Behav 57: 551-569.
CrossRefMedlineweb af vísindum
- ↵
- Shippenberg TS,
- Rea W
(1997) Pharmacol Biochem Behav 57: 449-455.
CrossRefMedlineweb af vísindum
- ↵
- Chen JS,
- Zhang YJ,
- Kelz MB,
- Steffen C,
- Ang ES,
- Zeng L,
- Nestler EJ
(2000) J Neurosci 20: 8965-8971.
- ↵
- Bibb JA,
- Chen JS,
- Taylor JR,
- Svenningsson P,
- Nishi A,
- Snyder GL,
- Yan Z,
- Sagawa ZK,
- Nairn AC,
- Nestler EJ,
- et al.
(2001) Náttúra (London) 410: 376-380.
- ↵
- Luscher C,
- Nicoll RA,
- Malenka RC,
- Muller D
(2000) Nat Neurosci 3: 545-550.
CrossRefMedlineweb af vísindum
-
- Malinow R,
- Helstu ZF,
- Hayashi Y
(2000) Curr Opin Neurobiol 10: 352-357.
CrossRefMedlineweb af vísindum
- ↵
- Scannevin RH,
- Huganir RL
(2000) Nat Rev Neurosci 1: 133-141.
CrossRefMedlineweb af vísindum
Robinson, TE & Kolb, B. (1999) (1997) Eur. J. Neurosci.11, 1598-1604.
- ↵
- Carey M,
- Smale ST
(2000) Transkriptunarreglugerð í eukaryotes (Cold Spring Harbor Lab. Press, Plainview, NY).
- ↵
- Spencer VA,
- Davie JR
(1999) Gene 240: 1-12.
CrossRefMedlineweb af vísindum
 Facebook
Facebook twitter
twitter- Google+
 CiteULike
CiteULike Delicious
Delicious Digg
Digg Mendeley
Mendeley
HighWire Press-hýst greinar vitna þessa grein
- Natural og Drug Rewards lögum um sameiginlegar tauga plástur kerfi með {Delta} FosB sem lykill sáttasemjari J. Neurosci. 2013 33 (8) 3434-3442
- Eiturlyf, glæpastarfsemi og epigenetics af heillandi alostasis Journal of Contemporary Criminal Justice 2012 28 (3) 314-328
- Abstract
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Abstract
- Fullur texti (HTML)
- Fullur texti (PDF)
- Morfín virkjar E Tuttugu og sex-eins Transcription Factor-1 / Serum Response Factor Pathway gegnum utanfrumulaga segulmagnaðir Kinases 1 / 2 í F11 frumum afleidd úr Dorsal Root Ganglia Neurons J. Pharmacol. Exp. Ther. 2012 342 (1) 41-52
- Molecular Mechanism for Gateway Drug: Epigenetic breytingar sem hefjast með nikótín Prime Gene Tjáning með Cocaine Sci Þýðing með 2011 3 (107) 107ra109
- Aukin súkrósa og kókain sjálfsstjórnun og kúgunartilboðandi lyf Leitað eftir tapi VGLUT2 í Midbrain dópamín taugafrumum í músum J. Neurosci. 2011 31 (35) 12593-12603
- Langvarandi truflandi blóðþrýstingur eykur blóðþrýsting og tjáningu FosB / {Delta} FosB í miðlægum sjálfstjórnarsvæðum Am. J. Physiol. Regla. Integr. Comp. Physiol. 2011 301 (1) R131-R139
- Frávik GPR37 / PAEL viðtakanna dregur úr striatal Akt og ERK2 fosfórýlering, {Delta} FosB tjáningu og skilyrt staðvalmöguleika á amfetamíni og kókaíni FASEB J. 2011 25 (6) 2071-2081
- Sambandið milli tímalengdar upphafs áfengislosunar og þrávirkrar mólþols er markvisst ólínulegt J. Neurosci. 2011 31 (7) 2436-2446
- In vivo Bioluminescence Imaging afhjúpar Redox-Regulated Activator Protein-1 Virkjun í Paraventricular Nucleus of Mice Með Renovascular Háþrýstingur Háþrýstingur 2011 57 (2) 289-297
- Striatal Yfirþynning á {Delta} FosB endurtekur langvarandi lifrarfrumukrabbamein J. Neurosci. 2010 30 (21) 7335-7343
- Epigenetísk miðlun umhverfisáhrifa í meiriháttar geðrofi Schizophr Bull 2009 35 (6) 1045-1056
- DNA-undirstaða MRI rannsóknir til sérstakrar greiningar á langvinnri útsetningu fyrir amfetamíni í lifandi heila J. Neurosci. 2009 29 (34) 10663-10670
- Breytt Dendritic Spine Plastity í kókain-Retracted Rottur J. Neurosci. 2009 29 (9) 2876-2884
- Yfirþrýstingsskjár í Drosophila skilgreinir taugahóp Hlutverk GSK-3 {beta} / shaggy sem eftirlitsstofnanna um AP-1-afleiddan þroskaþroska Erfðafræði 2008 180 (4) 2057-2071
- Uppskrift MRI: Nýtt útsýni yfir lifandi heila Neuroscientist 2008 14 (5) 503-520
- (Delta) FosB innleiðing í sporbrautarskotalyfjum Tolerance to Cocaine-induced Cognitive Dysfunction J. Neurosci. 2007 27 (39) 10497-10507
- Varanlegur varnarleysi við að hefja metamfetamín-leitarniðurstaðan í glæru frumuafleiddum taugafrumum stökkbreyttum músum FASEB J. 2007 21 (9) 1994-2004
- {Delta} FosB í Nucleus Accumbens stjórnar matvælaörvandi hljóðfæraleik og hvatningu J. Neurosci. 2006 26 (36) 9196-9204
- Reglugerð um {Delta} FosB Stöðugleiki með fosfórýleringu. J. Neurosci. 2006 26 (19) 5131-5142
- Tjáning á stökkbreytt NMDA-viðtaka í dópamín D1-viðtaka-innihaldsefnum kemur í veg fyrir könínskynjun og dregur úr kjúklingastillingu J. Neurosci. 2005 25 (28) 6651-6657
- D1 dópamínviðtaka Modulate (Delta) FosB innleiðing í Striatum rati eftir tímabundna notkun Morphine J. Pharmacol. Exp. Ther. 2005 314 (1) 148-154
- Neurobiology of Mice Valin fyrir High Voluntary Wheel-hlaupandi Virkni Integr. Comp. Biol. 2005 45 (3) 438-455
- Áhrif vatnsskorts og vökvaþurrðunar á c-Fos og FosB litun í rottum upptöku kjarnans og Lamina Terminalis svæðinu Am. J. Physiol. Regla. Integr. Comp. Physiol. 2005 288 (1) R311-R321
- Þrýstingur innleiðsla FosB / {Delta} FosB gen með vélrænni streitu í osteoblasts J Biol Chem 2004 279 (48) 49795-49803
- Innleiðing á {Delta} FosB í verðlaunasvæðinu eftir langvarandi streitu J. Neurosci. 2004 24 (47) 10594-10602
- Sim1 genaskammtur módelar heimaóstaðandi svörun við aukinni fæðufitu í músum Am. J. Physiol. Endókrinól. Metab. 2004 287 (1) E105-E113
- DNA microarray greining á genþrýstingi í geislalyfjum í sjóntaugum taugafrumum sem svar við vatnsþrýstingi Physiol. Genomics 2004 17 (2) 157-169
- Superoxíð er fólgið í miðtaugakerfi virkjun og meðvitundarleysi vegna hjartadreps í hjarta- og æðakerfi Circ. Res. 2004 94 (3) 402-409
- Adenósín A2A viðtaka í taugaaðlögun að endurtekinni dópamínvirkri örvun: Áhrif til meðferðar við hreyfitruflunum í Parkinsonsveiki Neurology 2003 61 (90116) S74-81
- Cytoplasmic Versus Nuclear Localization af Fos-tengdum próteinum í frosknum, Rana esculenta, Testis: In vivo og Bein In Vitro Áhrif gonadótrópín-losunarhormóns Agonist Biol. Reprod. 2003 68 (3) 954-960
- Periadolescent Mýs sýna aukið Delta FosB Upregulation í svörun við kókain og amfetamíni J. Neurosci. 2002 22 (21) 9155-9159
- Delta FosB stjórnar hjólum J. Neurosci. 2002 22 (18) 8133-8138
- CREB virkni í kjarnanum fylgir skel með því að stjórna hegðunarvandamálum við tilfinningalega áreiti Proc. Natl. Acad. Sci. USA 2002 99 (17) 11435-11440
- Psychogenomics: Tækifæri til að skilja fíkn J. Neurosci. 2001 21 (21) 8324-8327