Framan. Behav. Neurosci., 23 March 2016 | http://dx.doi.org/10.3389/fnbeh.2016.00054

 Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2,
Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2, 

- 1Þýðingarannsóknarstofnun og Institute for Health and Biomedical Innovation, Queensland University of Technology, Brisbane, QLD, Ástralíu
- 2Líffræðifræðideild, Háskólinn í Queensland, Brisbane, QLD, Ástralíu
Nútíma mataræði hefur orðið mjög sætt, sem leiðir til óvenjulegs sykursnotkunar, einkum hjá unglingum. Þótt langvarandi sykurskammtur á langvinnan hátt sé þekktur fyrir að stuðla að þróun efnaskiptatruflana, þ.mt offitu og sykursýki af tegund II, er lítið vitað um bein afleiðing langvarandi, binge-eins sykurs neyslu á heilanum. BVegna þess að sykur getur valdið losun dópamíns í kjarna accumbens (NAc) á sama hátt og misnotkunartímar, rannsóknum við breytingar á formgerð taugafrumna í heila svæðinu eftir skammtíma (4 vikur) og langvarandi (12 vikur) eins og súkrósa neysla með því að nota hlé á tveggja flöskum valháttum. Við notuðum Golgi-Cox litun til að meðhöndla miðlungs taugaþrengsli (MSN) úr NAc kjarnanum og skelum skammta og langs tíma súkrósa neyslu rottum og borið saman við aldurstengda vatnsstýringu. Við sýnum að langvarandi binge-líkur súkrósa neysla minnkaði verulega heildar dendritic lengd NAc skel MSNs samanborið við aldurstengda stjórn rottur. Við komumst að því að endurskipulagning þessara taugafrumna stafaði fyrst og fremst af minni dreifingu dendritic flókið. Hins vegar sáum við aukinn hryggþéttleiki í fjarlægum útibúaviðskiptum NAc skel MSNs úr langtíma súkrósa neyslu rottum. Samanlagt bendir þessi árangur á taugafrumum áhrifum langvarandi binge-eins og inntaka súkrósa á NAc skel MSN formgerð.
Hvernig gengur lífið dag frá degi? Er það í jafnvægi og allt eins og það á að vera? Er jafnvægi hvort sem litið er á veraldlega stöðu eða andlega? Lífið er eins og það er. Það er ekki alltaf sólskyn. Það koma reglulega lægðir með rok og rigningu. Við vitum að í heildar samhenginu er lægð hluti af vistkerfi að leita að jafnvægi. Stundum erum við stödd í miðju lægðarinnar. Þar er logn og gott veður, sama hvað gengur á þar sem stormurinn er mestur. Sama lögmál gildir varðandi þitt eigið líf. Ef þú ert í þinn miðju, þínum sannleik þá heldur þú alltaf jafnvægi átakalaust. Sama hvað gustar mikið frá þér þegar þú lætur til þín taka. Huldufólk hefur gefið okkur hugleiðslu sem hjálpar okkur að finna þessa miðju, finna kjarna okkar og sannleikann sem í honum býr. Þegar þú veist hver þú ert og hvers vegna þú ert hér, mun líf þitt vera í flæðandi jafnvægi. Hugleiðslan virkjar þekkinguna sem er í vitund jarðar og færir hana með lífsorkunni inn í líkama okkar. Þar skoðar hún hugsana og hegðunar munstrið og athugar hvort það myndar átakalausu flæðandi jafnvægi. Hinn möguleikinn er falskt jafnvægi sem hafa þarf fyrir að viðhalda með tilheyrandi striti, áhyggjum og ótta. Síðan leiðbeinir þessi þekking okkur að því jafnvægi sem er okkur eðlilegt. Við blómstrum átakalaust, líkt og planta sem vex átakalaut frá fræi í fullþroska plöntu sem ber ávöxt.
Á síðustu 40 árum hefur verið skráð skjöl í neyslu sykursósuðu drykkja og matvæla sem innihalda viðbættan sykur (Nielsen o.fl., 2002; Popkin, 2010; Ng et al., 2012), með skýrslum sem meta að allt að 75% allra matvæla og drykkja innihalda mikið magn af viðbættum sykrum (Ford og Dietz, 2013; Bray og Popkin, 2014). Á þessu tímabili hefur einnig verið samhliða aukning á tíðni offitu og sykursýki af tegund II, sérstaklega hjá unglingum (Arslanian, 2002; Reinehr, 2013; Dabelea et al., 2014; Fryar et al., 2014). Nýlegar rannsóknir hafa sýnt að of þungar og offitusjúklingar nota oft mikið magn af viðbættum sykri, en framlag hás sykurs sem inniheldur fæði á aukinni tíðni ofþyngdar og offitu barna er enn umdeild (Hu, 2013; Bray og Popkin, 2014; Bucher Della Torre o.fl., 2015).
Þó að vaxandi líkamsyfirlýsing bendir til þess að neysla sykursýkis megi stuðla að þyngdaraukningu meðal barna og unglinga (Malik et al., 2010; Te Morenga o.fl., 2013; Bray og Popkin, 2014) hefur minna athygli verið lögð á aukaverkanir sem ekki eru umbrotnar vegna óhóflegs sykurs inntöku. Athyglisvert koma nokkrar algengar hegðunar- og sálfræðilegar mynstur fram undir þeim hópi þeirra sem elda og viðhalda háum sykur sem innihalda fæði. Mest áberandi eru þróun átraskana, þar á meðal binge-borða, ásamt samtímis upphaf sálfræðilegra einkenna þ.mt skortur á hvatningu og þunglyndi (endurskoðuð í Sheehan og Herman, 2015). Þar að auki, vegna þess að binge-eating einstaklinga sýna oft tjón á eftirliti og vanhæfni til að takmarka sykurinntöku sín er líklegt að þessi hegðun myndist vegna taugafræðilegra aðlögunar í heilaþáttum sem meta hedonic gildi mjög mætanlegan mat (Saper et al., 2002; Lutter og Nestler, 2009; Kenny, 2011). Þessi rök eru einnig studd af sönnunargögnum hjá mönnum sem sýna að sykur og sætindi geta valdið þráum sem eru svipaðar þeim sem valda fíkniefnum eins og áfengi og nikótíni (Volkow et al., 2012).
Þrátt fyrir að ávanabindandi eiginleikar sykurs eru enn íhugandi, eru þessar athuganir ásamt rannsóknum sem sýna fram á að of mikið af sykri sé tekið til breytinga á verðlaunakröfum og þróun ávanabindandi hegðunar og tilfinningalegra ríkja í dýraformum (Avena o.fl., 2008; Benton, 2010; Ventura o.fl., 2014), ábyrgist þörfina fyrir frekari rannsókn. Fyrrverandi rannsóknir á nagdýrum hafa sýnt að hléum aðgengi að súkrósa breytir virkni nokkurra taugaboðefna innan mesólimbísku kerfisins, þ.mt dópamín, ópíóíðar og asetýlkólín (endurskoðað í Avena o.fl., 2008). Binge-eins og súkrósa neysla hefur verið sýnt fram á að auðvelda losun dópamíns í kjarna accumbens (NAc), á sama hátt og fíkniefniAvena o.fl., 2008). Ennfremur höfum við sýnt að langtíma neysla súkrósa með því að nota 24 h hléum aðgangsleiðbeiningarkerfi tveggja flúsa (Simms o.fl., 2008) stuðlar að nikótín acetýlkólínviðtaka (nAChR) tjáningu í NAc (Shariff o.fl., í stuttu). Athyglisvert höfum við einnig komist að því að nAChR efnasambönd sem vitað er að móta dópamín og asetýlkólínvirkni í NAc, hafa mismunandi áhrif á súkrósa neyslu eftir skammt og langtíma inntaka (Shariff o.fl., í stuttu).
Þrátt fyrir að þessar rannsóknir hafi sýnt líkt í hegðunar- og taugafræðilegum breytingum sem stafa af truflun á sykursýkingu og fíkniefnaneyslu, er ekki vitað hvort þessi áhrif auðvelda breytingar á taugafrumumyndun í NAc. THann er í mótsögn við misnotkunarefni, þar á meðal kókaín, amfetamín og nikótín sem framleiða vel einkenndu breytingar á formgerð miðlungs taugaþrengsli (MSN) í NAc, þar með talin aukin þéttleiki í hrygg og breytt dendritic flókið (Robinson og Kolb, 1999, 2004; Li et al., 2003; Crombag et al., 2005). Vegna þess að við höfum áður sýnt fram á að langtímaáhrif (12 viku) á áfengi og súkrósa með því að nota tímabundna tveggja flaska valparadísið, býr við mismunadrif við lyfjameðferð í samanburði við skammtímainntöku (4 vikur; Steensland o.fl., 2007; Shariff o.fl., í stuttu), metum við áhrif skamms og langtíma súkrósa neyslu á MSN formgerð í NAc. Við leyfðum unglingum rottum að neyta súkrósa á svipaðan hátt fyrir 4 (skammtíma) eða 12 (langan tíma) vikur og síðan greindar formgerð NAc MSNs úr skammtíma og langtíma súkrósa neyslu rottum og borið saman við þetta Aldurstengdir stýringar sem einungis voru veittir til vatns. Niðurstöður okkar sýna að MSNs úr NAc skelnum eru breytt eftir langvarandi en ekki skamms tíma súkrósa neyslu, með minnkað dendritic lengd, en aukin fjarlægur dendritic hryggþéttni. Ennfremur komumst við að formgerð MSNs úr NAc kjarnanum haldist tiltölulega ósnortinn eftir skammvinn og langvarandi súkrósa neyslu. Þessar niðurstöður vekja athygli á beinum taugafræðilegum afleiðingum langvarandi súkrósa neyslu á binge-svipaðan hátt. Enn fremur sýndu þessar upplýsingar þörfina fyrir frekari rannsóknir sem miða að því að lýsa mól- og taugafræðilegum breytingum sem fylgja formfræðilegri endurskipulagningu NAc skelja MSNs sem framkallað er með langvarandi, binge-eins súkrósa inntöku.
Efni og aðferðir
Siðareglur Yfirlýsing
Allar tilraunir voru gerðar í samræmi við ástralska reglurnar um umönnun og notkun dýra í vísindalegum tilgangi, 8th Edition (National Health and Medical Research Council, 2013). Samskiptareglurnar voru samþykktar af Dýragarðanefnd Háskóla Íslands í Dýragarði og Háskólanum í dýraheilbrigðiskerfinu í Queensland.
Dýr og húsnæði
Fimm vikna gamlar (unglingar) karlkyns Wistar rottur (Control: 176.4 ± 4.8 g; Súkrósa: 178.3 ± 5.0 g) (ARC, WA, Ástralía), voru hýst í loftræstum, tvíþættu plexiglasi® búr. Rotturnar voru lofaðar við einstaklingsbundnar húsnæðisaðstæður, meðhöndlun og afturljós hringrás 5 daga fyrir upphaf tilrauna. Öllum rottum var hýst í 12-klukku með loftslagsstýrðri ljós / dökkri hringrás (ljósin á 9 am) herbergi með venjulegu rottum og vatn í boði ad libitum.
Intermittent-Access Two-Bottle Choice Drinking Paradigm
Tímabundin aðgangur 5% súkrósa tveggja flaska valdreifingarháttur (Simms o.fl., 2008) var aðlöguð frá Vitur (1973). Öll vökvi var kynnt í 300 ml fluttu plastflöskum með ryðfríu stáli drykkjarstútum sett í gegnum tvær grommets framan við búrið eftir upphaf dimmu ljóssins. Þyngd hverrar flösku var skráð fyrir flaska kynningu. Tvær flöskur voru gefin samtímis: Einn flösku sem inniheldur vatn; seinni flöskan sem inniheldur 5% (w / v) súkrósa. Staðsetningin á 5% (w / v) súkrósa flöskunni var til skiptis með hverri útsetningu fyrir stjórn á hliðarvalkostum. Flöskur voru vigtaðar 24 klst eftir að vökvinn var kynntur og mælingar voru teknar til næsta 0.1 g. Þyngd hverrar rottu var einnig mældur til að reikna út gramm af súkrósainntöku á hvert kílógramm líkamsþyngdar. Á degi 1 í drykkjartímabilinu, rottum (n = 6-9) fengu aðgang að einum flösku af 5% (w / v) súkrósa og einum flösku af vatni. Eftir 24 klst var súkrósaflaska skipt út fyrir annan vatnsflaska sem var í boði fyrir næsta 24 klst. Þetta mynstur var endurtekið á miðvikudögum og föstudögum. Rotturnar höfðu ótakmarkaðan aðgang að vatni á öllum öðrum dögum. Binge-eins og súkrósa neysla leiddi til aukinnar heildar súkrósa neyslu (ml) með tímanum (aukatekjur 1) og fylgdist með stöðugum drekkaþéttni miðað við grunnþyngd [20 ± 5 g / kg af 5% (w / v)] á skammtíma [4 vikur (13 drykkir)] og langtíma [ ~ 12 vikur (37 drykkir)] drekka tímabil. Sérstakur hópur af rottum með stjórn (n = 6-9) fengu aðgang að vatni í báðum flöskum (þ.e. ekki súkrósa) við sömu skilyrði sem lýst er hér að ofan. Meðalþyngd eftirlits með rottum og súkrósa sem neytt var í lok tímabilsins var 405.7 ± 40.8 g og 426.4 ± 31.2 g. Í lok langtíma útsetningar var meðalþyngd eftirlits og súkrósahópa 578.8 ± 53.4 g og 600.2 ± 45.2 g.
Golgi-Cox litun
Eftir síðasta drykkjutímann voru rottur fluttar frá dýragarðinum til að unnt sé að vinna úr heilasýnum í vefjafræðilegum aðferðum við læknadeild Háskólans í Queensland (St Lucia, Ástralíu). Allar samþykktar ráðstafanir voru gerðar til að draga úr streitu meðan á flutningi stóð og eftir það voru rottur heimilt að batna yfir nótt. Daginn eftir var rottum fórnað með ofskömmtun af natríumpentóbarbítali (60-80 mg / kg, ip Vetcare, Brisbane, Ástralía) og hjartað með hjartsláttartruflunum ~ 300 ml sem innihélt (í mM): 130 NaCl, 3 KCl, 26 NaHCO3, 1.25 NaH2PO4, 5 MgCl2, 1 CaCl2, og 10 D-glúkósa. Hvert dýr var síðan deytt og heilinn fjarlægður og ræktaður í myrkri í Golgi-Cox lausn sem innihélt 5% kalíumdíkrómetat, 5% kalíumkrómat og 5% kvikasilfurklóríð (öll efni frá Sigma-Aldrich) sem voru gerðar ferskar 3 dagar fyrir fórn eins og áður var lýst (Rutledge o.fl., 1969). Golgi-Cox blettur ræktun og eftirvinnslu aðferðir voru breytt frá Ranjan og Mallick (2010). Hjörtu frá skammtíma súkrósa neyslu dýra voru ræktuð í 6 daga við 37 ° C, en heilar frá langtíma súkrósa neyslu dýr voru ræktuð í 10 daga, með einum breytingu á ferskum Golgi-Cox lausn eftir 4 daga ræktunar.
Eftir ræktun voru 300 μm kóróna köflunum skorin með því að nota titringur Zeiss Hyrax V50 míkrótróma (Carl Zeiss, Þýskaland). Skeri voru síðan sett í röð í 24-brunni fyllt með 30% (w / v) súkrósa í 0.1 M fosfat bólusettri saltlausn og unnin eins og lýst er í (Ranjan og Mallick, 2010). Í stuttu máli voru þurrkaðir hlutar í 50% etanóli fyrir 5 mín, síðan sett í 0.1 M NH4OH lausn fyrir 30 mín, skolað tvisvar með eimuðu vatni fyrir 5 mín og sett í Fujihunt kvikmynd fixer (Fujifilm, Singapore) fyrir 30 mín í myrkrinu. Skerðin voru síðan skoluð tvisvar í eimuðu vatni fyrir 2 mín. Hver og þurrkaðir í 70, 90, 95 og 100% etanól tvisvar fyrir 5 mín. Köflunum var síðan hreinsað í CXA lausn (1: 1: 1 klóróform: xýlen: alkóhól) fyrir 10 mín og sett í DPX (Sigma-Aldrich) á Superfrost Plus glærum (Menzel-Glaser, Lomb Scientific, Ástralíu) og kápa (Menzel-Glaser, Þýskaland). Skyggnurnar voru eftir í myrkrinu og þorna við stofuhita yfir nótt.
Neuronal Val og rekja innan Nucleus Accumbens
Coronal sneiðar milli bregma + 2.8 og + 1.7 voru könnuð fyrir MSN innan kjarna og skel af NAc, með því að nota hliðarmyndun og fremri kommissúr sem kennileiti með hjálp hjartastaðsPaxinos og Watson, 2007) (Mynd 1). Aðferðin í Neurolucida 7 (MBF Bioscience, VT, USA) var notuð til að afmarka NAc kjarna og NAc skel í hverri sneið (Mynd 2). Milli 2 og 9 taugafrumna á hverja svæði á dýrum voru reknar til breytinga á dendritic lengd með því að nota 63x markmið eða hryggþéttni (greint sem spines per 100 μm) með því að nota 100x markmið á Zeiss Axioskop II (Carl Zeiss, Þýskalandi) með sjálfvirkum xyz stigi ekið af Neurolucida® 7 hugbúnaður (MBF Biosciences, VT, USA). Öll rekja var gerð á blönduðum hátt með tilliti til meðferðar. Morfological breytur Golgi-Cox gegndreypt taugafrumum voru greind á svipaðan hátt og fyrri skýrslur (Klenowski et al., 2015).
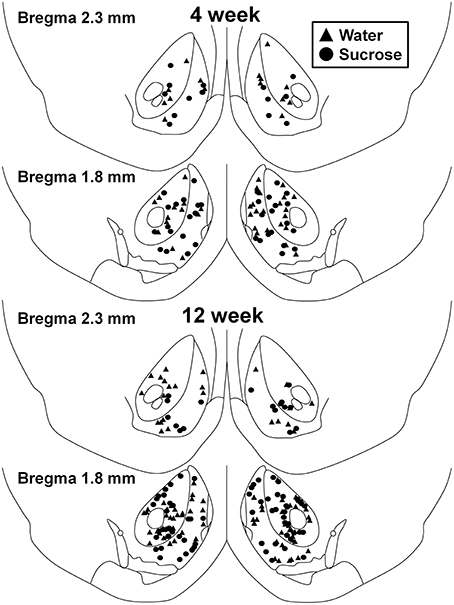
Mynd 1. Kort sem sýnir staði miðlungs hvítum taugafrumum, sem eru sýndar úr kjarnanum, sem er áberandi kjarna og skel af 4 og 12 viku, súkrósa neyslu rottum og aldurstengdum samanburði. Top tveir spjöld sýna staðsetningar taugafrumna sem eru sýndar úr kjarnanum sem er áberandi kjarna og skel af 4 viku eftirlit (þríhyrningur) og súkrósa (hringi) dýr. Neðri tveir spjöld sýna stöðu taugafrumna sem eru sýndar úr 12 viku eftirlit (þríhyrningur) og súkrósa (hringi) dýr.
Tölfræðileg greining
Meðaltal og staðalfrávik meðalgildis (SEM) voru reiknuð fyrir hvert gagnasett með dýrum sem n, með því að nota meðaltal morphometry gögn frá öllum kjarna eða skel NAc MSNs (n = 7 fyrir NAc skel og n = 6 fyrir NAc kjarna 4 viku, n = 9 fyrir 12 vikna hópa). Þar sem það er gefið til kynna, eru ekki paraðir tveir hali námsmanna tprófanir eða tvíhliða ANOVAs með Bonferroni eftirprófum voru gerðar fyrir allar greiningar sem felur í sér samanburð á hópnum með því að nota GraphPad Prism útgáfa 6.02 (GraphPad Software, San Diego, CA). Tölfræðilega þýðingu var samþykkt á P <0.05. Öll gögn í niðurstöðukaflanum eru sett fram sem meðaltal ± SEM. Prósentubreytingar eru reiknaðar út miðað við stjórngildið.
Niðurstöður
Medium Spiny Neurons frá Nucleus Accumbens Shell hafa minnkað Dendritic lengd, minnkað Dendritic Complexity en aukin Meinhryggþéttni við Distal Branch Pantanir Eftir Long-en ekki skammtíma Súkrósi Neysla
Eftir skammtíma (4 vikur) súkrósa neyslu, voru engin marktækur munur á NAc skel MSN morphometric breytur (tafla 1). Einnig voru engar marktækar munur á skammtíma súkrósa neyslu og vatnalegu NAc skel MSNs í greiningar sem tengjast miðflótta útibú röð. Nefnilega, dendritic hluti eftir útibú röð (P = 0.4111), meðaltal dendritic lengd á útibú röð (P = 0.5581) og meðalhryggþéttni fyrir hverja greinP = 0.2977, tvíhliða ANOVA) voru ekki marktækt mismunandi milli hópa. Staðsetningarkort sem sýnir áætlaða stöðu sýndu taugafrumna er sýnt á myndinni 1.
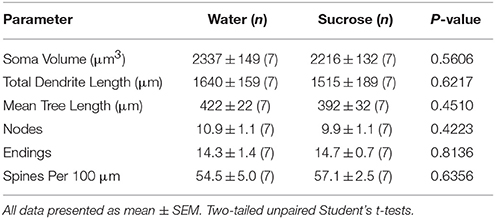
Tafla 1. Almennar morphologic breytur miðlungs spiny neurons úr kjarnanum accumbens skel af skammtíma súkrósa neyta rottur og aldursstilla vatn stjórna.
Eftir langtíma (12 vikur) súkrósa neyslu, minnkaði heildar dendritic arbor lengd NAc skel MSNs með 21% samanborið við vatnsnotkun stjórna (Vatn: 1827 ± 148 μm, n = 9; Súkrósi 1449 ± 78 μm, n = 9, *P = 0.0384, tveggja hala ópöraður námsmaður tpróf, mynd 2, Töflu 2). Samanburður á meðalfjölda dendritic bifurcations (nodes) og dendritic endings milli vatns og súkrósa hópa leiddi í ljós minni (þó ekki marktæk) stig dendritic flókið í MSc skel (NAc shell) (hnútar: Water 12.9 ± 1.4 n = 9, súkrósa 10.1 ± 0.8 n = 9, P = 0.0879; endir: Vatn 17.9 ± 1.4 n = 9, súkrósa 14.8 ± 0.7 n = 9, P = 0.0657, tveggja hala ópöraður námsmaður t-test, Tafla 2). Það var engin breyting á sumu rúmmáli (P = 0.9400), meðal dendritic tré lengd (P = 0.1646) eða heildarþéttleiki í hryggP = 0.3662) í NAc skel MSN frá langtíma súkrósa neyslu rottum samanborið við vatn stjórna. Þessar morphometric breytur eru ítarlegar í töflu 2.
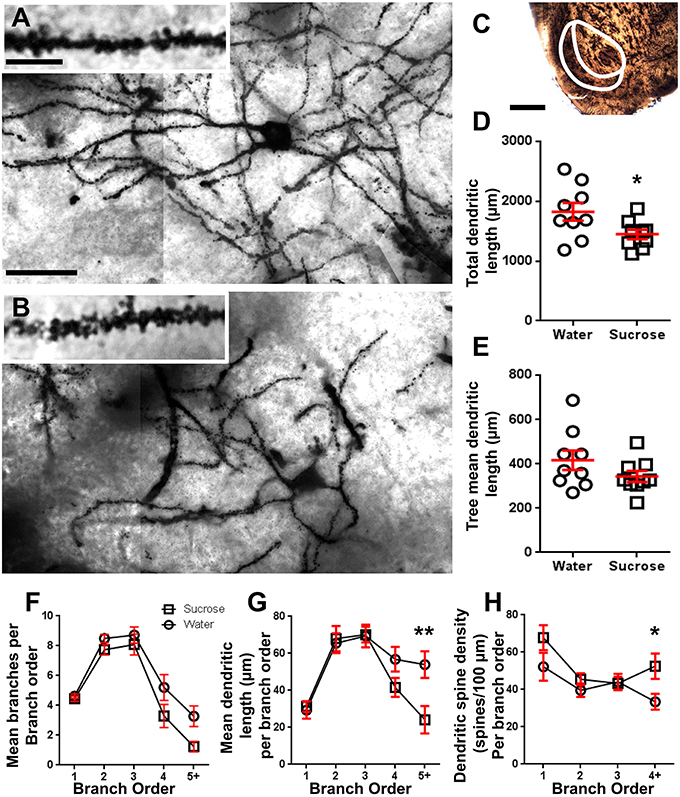 Mynd 2. Minnkað dendritic arbor lengd og aukin fjarlægur dendritic hryggþéttleiki miðlungs hryggjarnarefnum (MSN) úr kjarnanum accumbens (NAc) hylkinu á langvarandi súkrósaháðum rottum samanborið við samanburðarrottna. (A, B) Sýnið framsetning stjórnunar (efst) og langtíma (12 viku) súkrósa (botn) meðhöndluð björt svæði z-stack mósaík af Golgi-gegndreypt MSNs úr NAc skelinni (63x stækkun). Inset of (A, B) Sýnir stjórn og langtíma súkrósa meðhöndluðum björgunarmyndum af Golgi-gegndreyptum MSN dendrites og dendritic spines úr NAc skelinni (100x stækkun). (C) sýnir líffærafræði svæðin sem MSNs voru sýndar úr í þessari rannsókn. (D) sýnir dreifingarrit af minnkaðri heildar MSN dendritic arbor (meðaltal ± SEM) úr NAc skelinu í langtíma súkrósdýrum (ferninga) samanborið við stýringar (hringi), ópaired nemendur t-test, *P <0.05, n = 9; stjórn og n = 9; 12 viku súkrósa. (E) sýnir dreifbreytileika óbreyttrar meðaltals MSN dendritic tré lengd (meðal ± SEM) frá NAc skelinu í langtíma súkrósdýrum (ferninga) samanborið við stýringar (hringi), ópaired nemendur tpróf, P > 0.05, n = 9; stjórn og n = 9; 12 viku súkrósa. Greining á greiningu á útibúum (meðaltal ± SEM) af dendritic hluti númeri fyrir hverja grein (F), meðaltal dendritic lengd á útibú röð (G) og dendritic hryggþéttni fyrir hverja grein (H). Langtíma súkrósa neysla minnkaði dendritic lengd við fjarskiptapantanir (5 +) og aukin dendritic hryggþéttni við fjarskiptapantanir (4 +) samanborið við eftirlit (G, H), tvíhliða ANOVAs með Bonferroni eftir prófun, *P <0.05, **P <0.01, n = 9; stjórn og n = 9; langtíma súkrósa. Vogir: (A, B) = 20 μm; inntak af (A, B) = 10 μm; (C) = 1 mm.
Mynd 2. Minnkað dendritic arbor lengd og aukin fjarlægur dendritic hryggþéttleiki miðlungs hryggjarnarefnum (MSN) úr kjarnanum accumbens (NAc) hylkinu á langvarandi súkrósaháðum rottum samanborið við samanburðarrottna. (A, B) Sýnið framsetning stjórnunar (efst) og langtíma (12 viku) súkrósa (botn) meðhöndluð björt svæði z-stack mósaík af Golgi-gegndreypt MSNs úr NAc skelinni (63x stækkun). Inset of (A, B) Sýnir stjórn og langtíma súkrósa meðhöndluðum björgunarmyndum af Golgi-gegndreyptum MSN dendrites og dendritic spines úr NAc skelinni (100x stækkun). (C) sýnir líffærafræði svæðin sem MSNs voru sýndar úr í þessari rannsókn. (D) sýnir dreifingarrit af minnkaðri heildar MSN dendritic arbor (meðaltal ± SEM) úr NAc skelinu í langtíma súkrósdýrum (ferninga) samanborið við stýringar (hringi), ópaired nemendur t-test, *P <0.05, n = 9; stjórn og n = 9; 12 viku súkrósa. (E) sýnir dreifbreytileika óbreyttrar meðaltals MSN dendritic tré lengd (meðal ± SEM) frá NAc skelinu í langtíma súkrósdýrum (ferninga) samanborið við stýringar (hringi), ópaired nemendur tpróf, P > 0.05, n = 9; stjórn og n = 9; 12 viku súkrósa. Greining á greiningu á útibúum (meðaltal ± SEM) af dendritic hluti númeri fyrir hverja grein (F), meðaltal dendritic lengd á útibú röð (G) og dendritic hryggþéttni fyrir hverja grein (H). Langtíma súkrósa neysla minnkaði dendritic lengd við fjarskiptapantanir (5 +) og aukin dendritic hryggþéttni við fjarskiptapantanir (4 +) samanborið við eftirlit (G, H), tvíhliða ANOVAs með Bonferroni eftir prófun, *P <0.05, **P <0.01, n = 9; stjórn og n = 9; langtíma súkrósa. Vogir: (A, B) = 20 μm; inntak af (A, B) = 10 μm; (C) = 1 mm.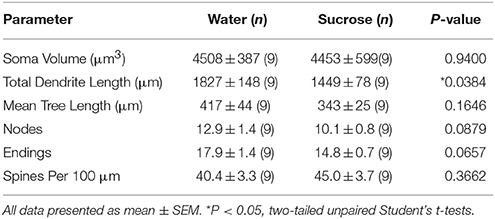
Tafla 2. Almennar morphologic breytur miðlungs spiny neurons úr kjarnanum accumbens skel af langtíma súkrósa neyslu rottum og aldursstilla vatn stjórna.
Í kjölfar einkenna almennrar dendritic formgerð langvarandi súkrósa neyta NAc skel MSNs, greindum við dendritic arborizations og hryggþéttleiki með tilliti til útibú röð eiginleika þeirra. Alhliða mat okkar á dendritískum trjám töldu fjölda dendritískra tegunda fyrir hverja greinafjölda, meðal lengd dendrítískra tegunda fyrir hverja greinafjölda og meðalhryggþéttni fyrir hverja útibú röð NAc skelja MSNs vatnsstýringar og langtíma súkrósa neyslu rottum. Samantekt á útibúargögnum og greiningu er að finna í töflu 3.
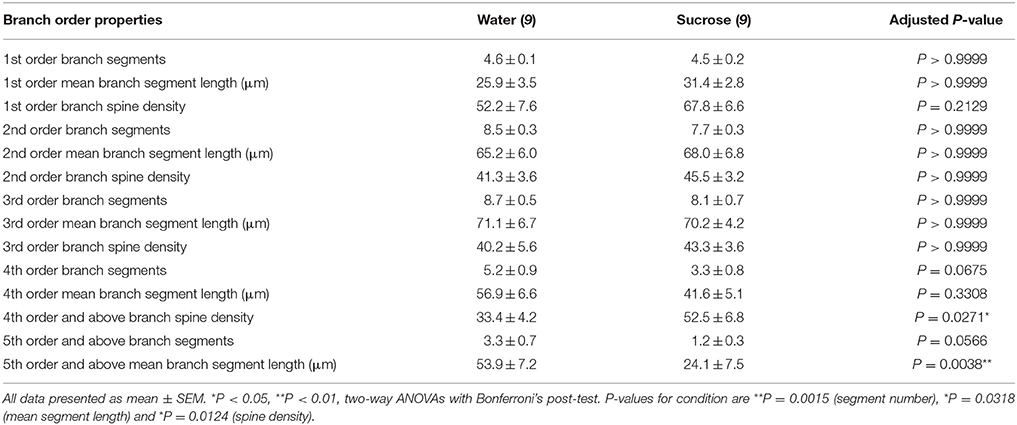 Tafla 3. Branch röð einkenni miðlungs spiny neurons frá langtíma súkrósa og vatni drekka rottum.
Tafla 3. Branch röð einkenni miðlungs spiny neurons frá langtíma súkrósa og vatni drekka rottum.Meðaltal dendritic útibúsþáttaröðunar á hverri grein röð NAc skelja MSNs var marktækt minni hjá langtíma súkrósa neyslu rottum samanborið við eftirlit með vatni (**P = 0.0015, tvíhliða ANOVA). Bonferroni eftir prófanir leiddu í ljós að stefnt var að fækkun útibúa í 4th (Vatn: 5.2 ± 0.9, n = 9; Súkrósi 3.3 ± 0.8, n = 9, P = 0.0675, Mynd 2F, Töflu 3) og 5th röð og yfir útibú pantanir (Vatn: 3.3 ± 0.7, n = 9; Súkrósi 1.2 ± 0.3, n = 9, P = 0.0566, Mynd 2F, Töflu 3). Meðal dendritic hluti lengd á hverri grein röð NAc skel MSNs var einnig marktækt minni hjá langtíma súkrósa neyslu rottum samanborið við vatn stjórna (*P = 0.0444, tvíhliða ANOVA). Bonferroni eftir prófun sýndi lækkun á 55% við 5th röð útibúa og víðar (Vatn: 53.9 ± 7.2 μm, n = 9; Súkrósi 24.1 ± 7.5 μm, n = 9, **P = 0.0038, Mynd 2G, Töflu 3).
Greining á greinargildi sýndi veruleg aukning á dendritic hryggþéttni NAc skelja MSNs af langtíma súkrósa neyslu rottum samanborið við eftirlit (*P = 0.0124, tvíhliða ANOVA). Bonferroni eftir prófanir sýndu aukningu á hryggþéttni 57% í fjarlægum 4th röð útibúum og víðar (Vatn: 33.4 ± 4.2, n = 9; Súkrósi 52.5 ± 6.8, n = 9, P = 0.0271 *, inntak mynda 2A, B, H, Töflu 3). Fulltrúar myndir af heildar MSN arkitektúr og þéttleiki í þéttum hrygg (innskot) eru sýndar í myndum 2A, B.
Samanlagt benda þessar niðurstöður til þess að skammtíma súkrósa neysla hafi lítil áhrif á fjölgunarmörk MSNs innan NAc skelarinnar. Eftir langvarandi neyslu er hins vegar marktæk lækkun á lengd og flóknu taugafrumum, sérstaklega í fjarlægum dendritískum greinum. Samhliða aukning á þéttni hryggja í þvagi er einnig sýnileg í NAc skel MSNs af langtíma súkrósa neyslu rottum.
Miðlungs Spiny Neurons frá Nucleus Accumbens Core hafa minnkað útibúaþéttni eftir langan tíma en ekki skammtíma súkrósa neyslu
Eftir skammtíma súkrósa neyslu, voru engar marktækar munur á mælingum á MSc morðfræðilegum NAc kjarna MSN (tafla 4). Það var einnig engin marktækur munur á notkun súkrósa neyslu á 4 viku og vatnstjórnun algerlega MSN í greiningar sem tengjast miðflóttaútibúum. Nefnilega, dendritic hluti eftir útibú röð (P = 0.7717), meðaltal dendritic lengd á útibú röð (P = 0.2096) og meðaltal hryggþéttni fyrir hverja greinP = 0.3521, tvíhliða ANOVA) voru ekki mismunandi milli hópa.
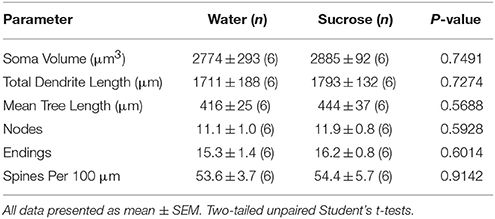 Tafla 4. Almennar morphologic breytur miðlungs spiny neurons úr kjarnanum accumbens kjarna skamms tíma súkrósa neyta rottur og aldursstilla vatn stjórna.
Tafla 4. Almennar morphologic breytur miðlungs spiny neurons úr kjarnanum accumbens kjarna skamms tíma súkrósa neyta rottur og aldursstilla vatn stjórna.Langvarandi súkrósa neysla hafði einnig ekki marktæk áhrif á MSF morphometric breytur (tafla 5). Meðaltal dendritic útibúsþáttaröðunar á hverri grein röð NAc algerlega MSNs var marktækt minni hjá langtíma súkrósa neyslu rottum samanborið við vatnsstýringu (*P = 0.0416, tvíhliða ANOVA), hins vegar var enginn marktækur munur á meðaldendritic lengd fyrir hverja greinastærð (P = 0.0995) og meðalhryggþéttni fyrir hverja greinP = 0.4888, tvíhliða ANOVA) milli MSNs í NAc kjarna langtíma súkrósa neyslu rottum samanborið við vatn stjórna. Samanlagt sýnir gögnin okkar að NAc kjarnainn er ekki eins viðbrögð við langtíma súkrósa neyslu miðað við MSN frá NAc skel svæðinu.
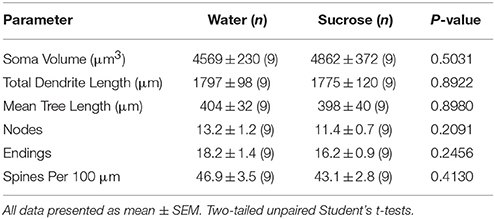 Tafla 5. Almennar morphologic breytur miðlungs spiny taugafrumum úr kjarnanum accumbens kjarna langvarandi súkrósa neyslu rottum og aldursstilla vatn stjórna.
Tafla 5. Almennar morphologic breytur miðlungs spiny taugafrumum úr kjarnanum accumbens kjarna langvarandi súkrósa neyslu rottum og aldursstilla vatn stjórna.Discussion
Aukin framboð á mjög sætuðum matvælum í vestrænu mataræði hefur ekki aðeins stuðlað að aukinni útbreiðslu og efnahagslegan byrði offitu og sykursýki af tegund II, það hefur einnig leitt til þess að sjúkdómar á borð við binge-ávexti (Swanson o.fl., 2011; Kessler et al., 2013; Davis, 2015). Þrátt fyrir að ávanabindandi eiginleikar sykurs, þar með talin frúktósa og súkrósa, eru enn íhugandi, þá er sláandi líkt í hegðunar- og tauga tengslunum sem koma fram vegna þess að borða og langvarandi notkun lyfja (Avena o.fl., 2008, 2011). Að auki virkjar sykur verðlaunahringrás heilans á svipaðan hátt og misnotkunarlyf (Volkow et al., 2012) og niðurstöður úr mönnum rannsóknum benda til þess að sykur og sætindi geti valdið þráhyggju sem er sambærileg í samræmi við þá sem valda fíkniefnum eins og áfengi og nikótíni (Volkow et al., 2012). Við notuðum því líkan af binge-súkrósa neyslu í rottum til að ákvarða áhrif skammta (4 vikur) og langtíma (12 vikur) súkrósa neyslu á taugafrumumyndun MSNs í NAc, lykilþáttur í skarastála sem er breytt af sykri og ávanabindandi lyfjum. Við sýnum að MSNs úr NAc skelinni á langvarandi, langvarandi súkrósa neyslu rottum hafa verulega dendritic lengd og flókið, en aukin fjarlægur dendritic hryggþéttni. Langtíma súkrósa neysla hafði engin áhrif á formgerð MSNs úr NAc kjarnanum, en skammtíma súkrósa neysla hafði einnig engin marktæk áhrif á MSN formgerð frá NAc kjarna eða skel. Þessar niðurstöður sýna ekki aðeins bein áhrif á langvarandi binge-eins súkrósa neyslu á taugafrumumyndun NAc skel MSNs, en einnig er lögð áhersla á hugsanlega skaðleg afleiðingar langvarandi neyslu sykursýru sem innihalda fæði.
The NAc, sem er hluti af ventral striatum, samanstendur fyrst og fremst af MSNs, sem eru einkennandi fyrir miðlungsmörk taugafrumum með víðtæka dendritic arborisations og hár hryggþéttleikiKemp og Powell, 1971; Graveland og DiFiglia, 1985; Rafols o.fl., 1989; Kawaguchi o.fl., 1990). Glutamatergic og dopaminergic taugafrumur eru tvö aðal afferent inntak til NAc, fyrst og fremst að hafa samband við dendritic stokka og spines af MSNs (Groves, 1980; Kaiya og Namba, 1981; Groves et al., 1994). Nánar tiltekið, NAc skel og kjarna fá glutamatergic inntak frá virkni greinilega cortical svæði (Brog et al., 1993). The NAc skel er einnig innervated af excitatory afferents frá subcardical svæðum eins og hippocampus, thalamus og basolateral amygdala (Brog et al., 1993; Wright og Groenewegen, 1995). Fyrri rannsóknir hafa sýnt að þessi glutamatergic inntak gegna lykilhlutverki í hvatningu og markvissri hegðun eins og mat og verðlaun (Maldonado-Irizarry et al., 1995; Kelley og Swanson, 1997; Reynolds og Berridge, 2003; Richard og Berridge, 2011). Önnur ríkjandi inntak á NAc MSNs er frá dópamínvirkum afferðum sem eru verkefni frá ventral tegmental svæðinu (Lindvall og Björklund, 1978; Veening o.fl., 1980; Kalivas og Miller, 1984). Athyglisvert hefur verið að fyrri rannsóknir með svipuðum gerðum af hléum sykursaðgangi hafi sýnt að binge-eins neysla sem leiðir af sér leiðir til aukningar á utanfrumu dópamíni í NAc á sama hátt (þó að minnsta kosti) að misnotkunartímum (Rada o.fl., 2005; Avena o.fl., 2006) og geta mótað dópamínviðtaka tjáningu (Colantuoni et al., 2001, 2002) í NAc kjarna og skel. Athyglisvert er að binge-eins og súkrósa neysla veldur aukningu í inntöku með tímanum á sama hátt og sjálfstjórn eiturlyfja af misnotkun eins og kókaín og heroin (Ahmed og Koob, 1998; Ahmed et al., 2000, 2003) sem tengist þróun ávanabindandi eins og ástands.
Greining á greiningartöflu útibúa sýnir að heildarlækkun á dendritic lengd NAc skelja MSNs vegna langvarandi súkrósa neyslu, leiðir fyrst og fremst af lækkun á flókið fjarskiptaskipti. Við sáum minnkað fjarlægur útibú (4th og 5th röð og yfir útibú pantanir) og marktækt minni meðal lengd við 5th röð og yfir dendrites, ásamt aukinni þéttleiki í hryggjum við þessar greinar. Algeng þáttur sem líklegt er að hafa áhrif á þessa tegund endurskipulagningar í dendritum felur í sér breytingar á synaptic tengingu og / eða virkni (Russo o.fl., 2010). Fyrri rannsóknir hafa sýnt að glutamatergic synapses á MSNs myndast fyrst og fremst á spines, einkum í fjarlægum dendrites (Groenewegen o.fl., 1999). Að auki er samhliða staðsetning dópamíns og glutamatergic innganga frá prefrontal heilaberki (Sesack og Pickel, 1992), hippocampus (Totterdell og Smith, 1989; Sesack og Pickel, 1990), og amygdala (Johnson et al., 1994) hafa komið fram á dendritic spines of MSNs. Þessar athuganir ásamt aukinni hryggþéttni eftir langvarandi súkrósa neyslu sem sjást í rannsókninni okkar, styðja myndun aukinnar spennandi inntaks. Þess vegna myndast möguleiki þar sem viðvarandi áhrif af völdum langvarandi binge-eins og súkrósainntaka gætu auðveldað aukna segulmagnaðir virkni í fjarlægum dendritum MSNs í NAc skelinni. Þar af leiðandi getur minnkun og / eða afturköllun fjarlægra dendrites komið fram með synaptic hemostatic kerfi (Reissner og Kalivas, 2010), þó að þetta sé enn fremur ákvörðuð.
Það er athyglisvert að hafa í huga að Crombag og samstarfsmenn sýndu að engin aukning á hryggþéttni náðist í NAc skelinu eftir að súkrósa neysla á 4 viku var tekin í gegnum neyðarpokann sjálfs gjöf, þrátt fyrir öflugri kaup og hærri svörun við súkrósa þegar borið var saman með amfetamíni (Crombag et al., 2005). Athugun þeirra á að engin breyting sé á þéttleika hryggja á 4 vikum endurspeglar niðurstöður okkar. Hins vegar sýnir rannsóknin okkar að eftirfarandi langvarandi (12 viku) útsetning fyrir langvarandi súkrósa neyslu, það er veruleg aukning á þéttleiki í þéttum hryggjum á MSN súkrósa-reynslu rottum. Enn fremur hefur rannsóknarstofan okkar sýnt fram á að langtíma (12 viku) súkrósa neysla auðveldar mismunandi lyfjafræðilega svörun við lyfjameðferð sem hefur verið sýnt fram á að móta dópamín og asetýlkólínviðbrögð við stigi NAcShariff o.fl., í stuttu). Samanlagt bendir þetta til þess að langtíma (12 vikur og víðar) súkrósaáhrif, sem endurspeglar nákvæmlega raunverulegar aðstæður, leiðir til breytingar á formgerð á stigi NAc.
Hvað varðar eiturlyf af misnotkun, veldur endurtekin útsetning fyrir ýmsum lyfjum langvarandi breytingum á uppbyggingu dendrites og dendritic spines. Til dæmis auka amfetamín og kókaín bæði hryggþéttni í NAc bæði í skel og kjarna (Robinson og Kolb, 2004). Sýnt hefur verið fram á að útsetning nikótíns hefur aukið hryggþéttni í NAc skelinni. Hins vegar veldur morfín útsetning lækkun á þéttleika hryggs og dendritískrar útibúa (Robinson og Kolb, 2004). Að því er varðar langtíma súkrósa neyslu, sáum við aukningu á þéttleika hryggs, svipað og amfetamíns, kókaíns og nikótíns og gagnstæða áhrifum morfíns. Hins vegar, ólíkt amfetamíni og kókaíni, en svipað nikótíni, er aukning á þéttleika hryggja við langtímaáhrif á súkrósa takmörkuð við NAc skel. Það er líka athyglisvert að breytingar á bæði dendritic branching (Robinson og Kolb, 1999) og hryggþéttleikiLi et al., 2003) framleiddar með amfetamíni eða kókaíni eru bundin við fjarlægum dendrites MSNs í NAc sem endurspeglar niðurstöðurnar í rannsókninni. Enn fremur, og staðfesting á breytingum sem lýst er hér að framan, hefur súkrósa neysla einnig áður verið sýnt fram á að auka sveigjanleg synaptic styrk á accumbal dópamín taugafrumum (Stuber o.fl., 2008b) auk annarra þætti í mesolimbic verðlaunahópnum (Stuber o.fl., 2008a; Chen et al., 2010). Samanlagt setur þetta súkrósa sem öflugt mótamælir taugafræðinnar eftir langvarandi notkun, sem er í sambandi við þau áhrif sem koma fram við lyf við misnotkun.
Þó að frekari rannsóknir séu nauðsynlegar til að afhjúpa frumu og synaptic kerfi sem stuðla að formfræðilegum breytingum sem sjást í þessari rannsókn, sýna niðurstöður okkar veruleg taugafræðileg áhrif sem stafa af langvarandi súkrósa neyslu. Einkum er umfjöllun sem ekki er rannsökuð í rannsókninni okkar hvort hægt sé að vekja fram morphological áhrif súkrósa með sótthreinsiefnum sem ekki eru kaloría eins og sakkarín. Í þessu sambandi er mikilvægt að hafa í huga að Lenoir og samstarfsmenn hafa sýnt að mikil sælgæti nær yfir kókaínverðlaun, hvort sem það myndast af sakkaríni eða súkrósa (Lenoir o.fl., 2007). Enn fremur, nýleg rannsókn sem birt var af Lab okkar (Shariff o.fl., í stuttu) sýnir að vareniclín, nikótíns acetýlkólínviðtaka hluta örva minnkað bæði súkrósa og sakkarín inntöku hjá nagdýrum, eftir sömu langtímameðferð, sem var notað í þessari rannsókn. Athyglisvert er að fyrri rannsóknir hafa sýnt líkt við bráðaáhrif sótthreinsiefna sem ekki eru kaloría eins og sakkarín og súkrósi á stigi NAc (Scheggi o.fl., 2013; Tukey o.fl., 2013; Carelli og Vestur, 2014). Hins vegar þarf frekari rannsóknir að ákvarða hvort sótthreinsiefni sem ekki eru kaloría geta valdið langtímaáhrifum svipað og breytingar á formgerð NAc skelinni MSNs vegna langvarandi súkrósa neyslu sem hér er greint frá.
Skortur á áhrifum á MSc-formgerð NAc eftir skammtíma súkrósa neyslu, er lögð áhersla á mikilvægi þess að framkvæma langtímarannsóknir til að meta áhrif langvarandi misnotkunar á fíkniefnum eða náttúrulegum ávinningi eins og súkrósa. Að því er varðar ósjálfstæði eru ekki aðeins endurteknar hringrásir binge inntöku og fráhvarfseinkennum í fíkniefninu, en vaxandi líkamsyfirlýsing hefur leitt í ljós að breytingin í ósjálfstæði er framsækið ferli sem oft á sér stað um langan tíma. Þrátt fyrir að ávanabindandi eiginleikar sykurs séu óviss eru líkurnar á því að fíkn á öðrum ónæmisbæðum, svo sem kynlíf, fjárhættuspil og mat, sífellt rannsökuð. Niðurstöðurnar úr þessari rannsókn bætast við þeirri forsendu að sykur eins og súkrósa hafi hugsanlega ávanabindandi eiginleika eftir langvarandi, binge-eins og neysla. Niðurstöður okkar hafa einnig áhrif á vaxandi fjölda barna og unglinga sem viðhalda óheilbrigðum matarvenjum (mikilli sykursnotkun og binge eating) til fullorðinsárs. Í takt við aukna hættu á að fá efnaskiptaáhrif er einnig mögulegt að taugakerfi og geðræn afleiðingar sem hafa áhrif á skap og hvatningu geta einnig stafað af þessum hegðun.
Höfundur Framlög
Taka þátt í rannsóknarhönnun: PK, SB. Framkvæmdar tilraunir: PK, MS, AB, MF, EM. Gögnargögn: PK, MF, MS. Túlkað gögnin og stuðlað að því að skrifa handritið: PK, MS, MF, EM, MB, SB. Allir höfundar lesa og samþykktu endanlegt handrit fyrir uppgjöf.
Hagsmunaárekstur
Höfundarnir lýsa því yfir að rannsóknirnar hafi farið fram án þess að viðskiptabundin eða fjárhagsleg tengsl gætu talist hugsanleg hagsmunaárekstur.
Gagnrýnendur SC, SA og meðhöndlun ritstjóri lýsti hlutdeild sinni og meðhöndlun ritstjóri segir að aðferðin uppfylli engu að síður viðmiðanir um sanngjarna og hlutlæga endurskoðun.
Acknowledgments
Þessi vinna var studd með fjármögnun frá styrkjum frá Australian Research Council (FT1110884) til SB og National Health and Medical Research Council (1061979) til SB og MB.
Viðbótarefni
The viðbótarefni fyrir þessa grein er að finna á netinu á: http://journal.frontiersin.org/article/10.3389/fnbeh.2016.00054
Aukategund 1. Inntaka súkrósa og ákvarða frá 4 og 12 viku súkrósa neyslu rottum. (A, B) sýna aukningu í heildarinnihald súkrósa (ml) yfir 4 og 12 vikur útsetningar. (C, D) sýna mikla áherslu á súkrósa yfir vatni meðan á súkrósa kynningu stendur.
Meðmæli
Ahmed, SH og Koob, GF (1998). Breyting frá miðlungs til mikilli inntöku af völdum lyfsins: breyting á blæðingarhættu. Vísindi 282, 298-300. doi: 10.1126 / science.282.5387.298
Ahmed, SH, Lin, D., Koob, GF og Parsons, LH (2003). Aðlögun kókaíns sjálfs gjafar fer ekki eftir breytingum á kókaínvöldum kjarna sem dregur úr dópamíni. J. Neurochem. 86, 102-113. doi: 10.1046 / j.1471-4159.2003.01833.x
Ahmed, SH, Walker, JR og Koob, GF (2000). Viðvarandi aukning á hvatningu til að taka heróíni í rottum með sögu um lyfjameðferð. Neuropsychopharmacology 22, 413–421. doi: 10.1016/S0893-133X(99)00133-5
Arslanian, S. (2002). Gerðu 2 sykursýki hjá börnum: klínískar hliðar og áhættuþættir. Horm Res 57 (Suppl. 1), 19-28. gera: 10.1159 / 000053308
Avena, NM, Bocarsly, ME, Hoebel, BG og Gull, MS (2011). Skarast í nosology um misnotkun á efnaskipti og overeating: þýðingu afleiðingar af "fíkniefni". Curr. Misnotkun eiturlyfja. 4, 133-139. gera: 10.2174 / 1874473711104030133
Avena, NM, Rada, P. og Hoebel, BG (2008). Vísbendingar um sykurfíkn: Hegðunarvandamál og taugafræðileg áhrif af hléum, óhóflegri sykursnotkun. Neurosci. Biobehav. Rev. 32, 20-39. doi: 10.1016 / j.neubiorev.2007.04.019
Avena, NM, Rada, P., Moise, N. og Hoebel, BG (2006). Súkrósasamdráttur í binge áætlun gefur frá sér dopamín endurtekið endurtekið og útilokar asetýlkólín mætingarviðbrögð. Neuroscience 139, 813-820. doi: 10.1016 / j.neuroscience.2005.12.037
Benton, D. (2010). Ásættanlegt sykursýkni og hlutverk þess í offitu og átröskunum. Clin. Nutr. 29, 288-303. doi: 10.1016 / j.clnu.2009.12.001
Bray, GA og Popkin, BM (2014). Matarsykur og líkamsþyngd: höfum við náð kreppu í faraldri offitu og sykursýki ?: Heilsa vera fordæmdur! Hellið á sykri. Sykursýki 37, 950-956. doi: 10.2337 / dc13-2085
Brog, JS, Salyapongse, A., Deutch, AY og Zahm, DS (1993). Mynstur afferent innervation kjarna og skel í "accumbens" hluta rottum ventral striatum: ónæmissvörfræðileg greining retrogradely flutt flúor-gull. J. Comp. Neuról. 338, 255-278. doi: 10.1002 / cne.903380209
Bucher Della Torre, S., Keller, A., Laure Depeyre, J., og Kruseman, M. (2015). Sykursýktar drykkir og offitaáhætta hjá börnum og unglingum: kerfisbundin greining á því hvernig aðferðafræðileg gæði getur haft áhrif á niðurstöður. J. Acad. Nutr. Mataræði. [Epub á undan prenta]. doi: 10.1016 / j.jand.2015.05.020
Carelli, RM og West, EA (2014). Þegar góður smekkur verður slæmur: taugakerfi sem undirliggjandi eru tilkomu neikvæðra áhrifa og tilheyrandi náttúruverðlauna af kókaíni. Neuropharmacology 76 (Pt B), 360-369. doi: 10.1016 / j.neuropharm.2013.04.025
Chen, BT, Hopf, FW og Bonci, A. (2010). Synaptic plasticity í mesolimbic kerfi: meðferðaráhrif fyrir misnotkun efna. Ann. NY Acad. Sci. 1187, 129-139. doi: 10.1111 / j.1749-6632.2009.05154.x
Colantuoni, C., Rada, P., McCarthy, J., Patten, C., Avena, NM, Chadeayne, A., et al. (2002). Vísbendingar um að hléum, óhóflega sykurskammtur veldur innrænum ópíóíðfíkn. Obes. Res. 10, 478-488. doi: 10.1038 / oby.2002.66
Colantuoni, C., Schwenker, J., McCarthy, J., Rada, P., Ladenheim, B., Cadet, JL, et al. (2001). Óþarfa sykurinntaka breytir bindingu við dópamín og mu-ópíóíðviðtaka í heilanum. Neuroreport 12, 3549–3552. doi: 10.1097/00001756-200111160-00035
Crombag, HS, Gorny, G., Li, Y., Kolb, B. og Robinson, TE (2005). Andstæða áhrif amfetamíns sjálfs gjafar reynslu á dendritic spines í miðgildi og hringlaga barkakrabbameini. Cereb. Heilaberki 15, 341-348. doi: 10.1093 / cercor / bhh136
Dabelea, D., Mayer-Davis, EJ, Saydah, S., Imperatore, G., Linder, B., Divers, J., et al. (2014). Algengi tegund 1 og tegund sykursýki af tegund 2 hjá börnum og unglingum frá 2001 til 2009. Jama 311, 1778-1786. doi: 10.1001 / jama.2014.3201
Davis, C. (2015). Faraldsfræði og erfðafræði binge eating disorder (BED). CNS Spectr. 20, 522-529. doi: 10.1017 / s1092852915000462
Ford, ES og Dietz, WH (2013). Stefna í orkunotkun meðal fullorðinna í Bandaríkjunum: niðurstöður frá NHANES. Am. J. Clin. Nutr. 97, 848-853. doi: 10.3945 / ajcn.112.052662
Fryar, CD, Carroll, MD og Ogden, CL (2014). Algengi ofþyngdar, offitu og mikillar offitu meðal fullorðinna: Bandaríkin, 1960-1962 gegnum 2011-2012. Atlanta, GA: CDC.
Graveland, GA og DiFiglia, M. (1985). Tíðni og dreifing miðlungs stórra taugafrumna með inndælingu kjarnum í frumu og nagdýr neostriatum. Brain Res. 327, 307–311. doi: 10.1016/0006-8993(85)91524-0
Groenewegen, HJ, Wright, CI, Beijer, AV og Voorn, P. (1999). Samleitni og sundurliðun á inntökum og útgangi á innrennsli. Ann. NY Acad. Sci. 877, 49–63. doi: 10.1111/j.1749-6632.1999.tb09260.x
Groves, PM (1980). Synaptic endingar og postsynaptic markmið þeirra í neostriatum: Synaptic sérhæfingar komu fram frá greiningu á raðgreinum. Proc. Natl. Acad. Sci. Bandaríkin 77, 6926-6929. doi: 10.1073 / pnas.77.11.6926
Groves, PM, Linder, JC og Young, SJ (1994). 5-hýdroxýdóprómer-merkt dópamínvirka axón: þrívíð enduruppbygging axons, synapses og postsynaptic markmið í rottum neostriatum. Neuroscience 58, 593–604. doi: 10.1016/0306-4522(94)90084-1
Hu, FB (2013). Leyst: Það er nægjanlegt vísindaleg merki um að minnkandi sykur-sætt drykkur neysla muni draga úr útbreiðslu offita og offita sem tengjast sjúkdómum. Obes. Rev. 14, 606-619. doi: 10.1111 / obr.12040
Johnson, LR, Aylward, RL, Hussain, Z, og Totterdell, S. (1994). Inntak frá amygdala í rottum kjarnanum accumbens: tengsl þess við týrósín hýdroxýlasa ónæmissvörun og greind taugafrumum. Neuroscience 61, 851–865. doi: 10.1016/0306-4522(94)90408-1
Kaiya, H. og Namba, M. (1981). Tvær gerðir af dópamínvirkum taugaskiptum í rottum neostriatum. Yfirbyggingarfræði. Neurosci. Lett. 25, 251–256. doi: 10.1016/0304-3940(81)90400-6
Kalivas, PW og Miller, JS (1984). Neurotensín taugafrumur í ventral tegmental svæðinu eru í miðju kjarna accumbens. Brain Res. 300, 157–160. doi: 10.1016/0006-8993(84)91351-9
Kawaguchi, Y., Wilson, CJ og Emson, PC (1990). Útsetningar undirhvarfa af fósturfrumum úr fósturfrumum sem komu fram með inndælingu í blóðfrumum í blóði. J. Neurosci. 10, 3421-3438.
Kelley, AE og Swanson, CJ (1997). Feeding framkölluð af blokkun AMPA og kainate viðtaka innan ventral striatum: rannsóknaráætlun um örverufræðslu. Behav. Brain Res. 89, 107–113. doi: 10.1016/S0166-4328(97)00054-5
Kemp, JM og Powell, TP (1971). The synaptic skipulag á caudate kjarnanum. Philos. Trans. R. Soc. Lond. B Biol. Sci. 262, 403-412. doi: 10.1098 / rstb.1971.0103
Kenny, PJ (2011). Verðlaunakerfi í offitu: nýjar upplýsingar og framtíðarstefnur. Taugafruma 69, 664-679. doi: 10.1016 / j.neuron.2011.02.016
Kessler, RC, Berglund, PA, Chiu, WT, Deitz, AC, Hudson, JI, Shahly, V., et al. (2013). Algengi og fylgni binge eating disorder í World Health Organization World Mental Health Kannanir. Biol. Geðlækningar 73, 904-914. doi: 10.1016 / j.biopsych.2012.11.020
Klenowski, PM, Fogarty, MJ, Belmer, A., Noakes, PG, Bellingham, MC og Bartlett, SE (2015). Uppbygging og virkni einkenna dendritic arbors og GABAergic synaptic inntak á interneurons og helstu frumur í rottum basolateral amygdala. J. Neurophysiol. 114, 942-957. doi: 10.1152 / jn.00824.2014
Lenoir, M., Serre, F., Cantin, L., og Ahmed, SH (2007). Mikil sælgæti nær yfir kókaínverðlaun. PLoS ONE 2: e698. doi: 10.1371 / journal.pone.0000698
Li, Y., Kolb, B. og Robinson, TE (2003). Staðsetning viðvarandi breytinga á amfetamíni í þéttleika dendritic spines á miðlungs hreinum taugafrumum í kjarnanum accumbens og caudate-putamen. Neuropsychopharmacology 28, 1082-1085. doi: 10.1038 / sj.npp.1300115
Lindvall, O. og Björklund, A. (1978). Líffærafræði dopamínvirkra taugakerfisins í rottum heilanum. Adv. Biochem. Psychopharmacol. 19, 1-23.
Lutter, M., og Nestler, EJ (2009). Hjartasjúkdómseinkenni og vöðvaspennur hafa áhrif á mat á inntöku. J. Nutr. 139, 629-632. doi: 10.3945 / jn.108.097618
Maldonado-Irizarry, CS, Swanson, CJ og Kelley, AE (1995). Glutamat viðtaka í kjarnanum bætir skelinni með því að stjórna fóðrun hegðun með hliðarhimnubólgu. J. Neurosci. 15, 6779-6788.
Malik, VS, Popkin, BM, Bray, GA, Després, JP og Hu, FB (2010). Sykursósuðu drykkir, offita, tegund 2 sykursýki og áhættu á hjarta og æðasjúkdómi. Hringrás 121, 1356-1364. gera: 10.1161 / CIRCULATIONAHA.109.876185
Ng, SW, Slining, MM, og Popkin, BM (2012). Notkun á kalorískum og noncaloric sætuefnum í bandarískum neysluvörum, 2005-2009. J. Acad. Nutr. Mataræði. 112, 1828-1834 e1821-e1826. doi: 10.1016 / j.jand.2012.07.009
Nielsen, SJ, Siega-Riz, AM og Popkin, BM (2002). Stefna í orkunotkun í Bandaríkjunum milli 1977 og 1996: svipuð breyting séð yfir aldurshópa. Obes. Res. 10, 370-378. doi: 10.1038 / oby.2002.51
Paxinos, G. og Watson, C. (2007). Hjörtur Hjarta í hornhimnu. Amsterdam; Boston, MA: Academic Press / Elsevier.
Popkin, BM (2010). Hvað er athugavert við nálgun Bandaríkjanna á offitu? Virtual Mentor 12, 316–320. doi: 10.1001/virtualmentor.2010.12.4.pfor2-1004
Rada, P., Avena, NM og Hoebel, BG (2005). Daglegt bingeing á sykur losar endurtekið dópamín í sólgleraugu. Neuroscience 134, 737-744. doi: 10.1016 / j.neuroscience.2005.04.043
Rafols, JA, Cheng, HW og McNeill, TH (1989). Golgi rannsókn á striatum músinni: aldurstengd dendritic breytingar á mismunandi taugafrumum. J. Comp. Neuról. 279, 212-227. doi: 10.1002 / cne.902790205
Ranjan, A. og Mallick, BN (2010). Breytt aðferð fyrir samkvæm og áreiðanleg Golgi-cox litun á verulega minni tíma. Framan. Neuról. 1: 157. doi: 10.3389 / fneur.2010.00157
Reinehr, T. (2013). Sláðu inn 2 sykursýki hjá börnum og unglingum. Heimur J. Sykursýki 4, 270-281. doi: 10.4239 / wjd.v4.i6.270
Reissner, KJ og Kalivas, PW (2010). Notkun glutamat hemostasis sem markmið fyrir meðhöndlun ávanabindandi sjúkdóma. Behav. Pharmacol. 21, 514–522. doi: 10.1097/FBP.0b013e32833d41b2
Reynolds, SM og Berridge, KC (2003). Glutamat áhugasamstæður í kjarnanum accumbens: rostrocaudal skel stig af ótta og fóðrun. Eur. J. Neurosci. 17, 2187-2200. doi: 10.1046 / j.1460-9568.2003.02642.x
Richard, JM og Berridge, KC (2011). Nucleus accumbens dopamín / glutamat samskipti skiptir stillingar til að búa til löngun á móti ótta: D (1) einn fyrir matarlyst en D (1) og D (2) saman fyrir ótta. J. Neurosci. 31, 12866-12879. doi: 10.1523 / JNEUROSCI.1339-11.2011
Robinson, TE og Kolb, B. (1999). Breytingar á formgerð dendrites og dendritic spines í kjarnanum accumbens og prefrontal heilaberki eftir endurtekna meðferð með amfetamíni eða kókaíni. Eur. J. Neurosci. 11, 1598-1604. doi: 10.1046 / j.1460-9568.1999.00576.x
Robinson, TE og Kolb, B. (2004). Structural plasticity í tengslum við útsetningu fyrir fíkniefnum. Neuropharmacology 47 (Suppl. 1), 33-46. doi: 10.1016 / j.neuropharm.2004.06.025
Russo, SJ, Dietz, DM, Dumitriu, D., Morrison, JH, Malenka, RC og Nestler, EJ (2010). The hávaði synapse: kerfi synaptic og uppbyggingu plasticity í kjarnanum accumbens. Stefna Neurosci. 33, 267-276. doi: 10.1016 / j.tins.2010.02.002
Rutledge, LT, Duncan, J., og Beatty, N. (1969). Rannsókn á pýramíddarfrumuæxlfrumum í ósnortnum og að hluta til einangruðum heila heilaberki. Brain Res. 16, 15–22. doi: 10.1016/0006-8993(69)90082-1
Saper, CB, Chou, TC og Elmquist, JK (2002). Þörfin á að fæða: heimavinnandi og heillandi stjórn á að borða. Taugafruma 36, 199–211. doi: 10.1016/S0896-6273(02)00969-8
Scheggi, S., Secci, ME, Marchese, G., De Montis, MG og Gambarana, C. (2013). Áhrif á sælgæti á hvatningu til að starfa fyrir kalorískan og ekki kalorískan mat í ófæddum matvælum og matarleysum rottum. Neuroscience 236, 320-331. doi: 10.1016 / j.neuroscience.2013.01.027
Sesack, SR og Pickel, VM (1990). Í miðtaugakerfi rottum samanstendur hippocampal og catecholaminergic skautanna á spiny neurons og eru í sambandi við hvert annað. Brain Res. 527, 266–279. doi: 10.1016/0006-8993(90)91146-8
Sesack, SR og Pickel, VM (1992). Prefrontal cortical efferents í rottum synapse á ómerktum taugafrumum catecholamine skautanna í kjarnanum accumbens septi og á dópamín taugafrumum í ventral tegmental svæði. J. Comp. Neuról. 320, 145-160. doi: 10.1002 / cne.903200202
Shariff, M., Quik, M., Holgate, JY, Morgan, M., Patkar, OL, Tam, V., et al. (í stuttu). Neuronal nikótín acetýlkólínviðtaksmælir draga úr sykursýkingu. PLoS ONE.
Sheehan, DV og Herman, BK (2015). Sálfræðilegir og sjúklegir þættir sem tengjast ómeðhöndluðu binge eating disorder. Prim. Umhirðu félagsþjónustufulltrúa. 17. doi: 10.4088 / PCC.14r01732
Simms, JA, Steensland, P., Medina, B., Abernathy, KE, Chandler, LJ, Wise, R., et al. (2008). Bráðan aðgang að 20% etanóli veldur mikilli etanól neyslu í Long-Evans og Wistar rottum. Áfengi. Clin. Exp. Res. 32, 1816-1823. doi: 10.1111 / j.1530-0277.2008.00753.x
Steensland, P., Simms, JA, Holgate, J., Richards, JK, og Bartlett, SE (2007). Varenicline, alfa4beta2 nikótín acetýlkólínviðtaka hluta örvi, veldur sértækum neyslu etanóls og leitar. Proc. Natl. Acad. Sci. Bandaríkin 104, 12518-12523. doi: 10.1073 / pnas.0705368104
Stuber, GD, Hopf, FW, Hahn, J., Cho, SL, Guillory, A. og Bonci, A. (2008a). Sjálfgefin etanólinntaka eykur sveigjanlegan synaptískan styrk á kviðarholi. Áfengi. Clin. Exp. Res. 32, 1714-1720. doi: 10.1111 / j.1530-0277.2008.00749.x
Stuber, GD, Klanker, M., de Ridder, B., Bowers, MS, Joosten, RN, Feenstra, MG, et al. (2008b). Reward-predictive cues auka spennandi synaptic styrk á midbrain dópamín taugafrumum. Vísindi 321, 1690-1692. doi: 10.1126 / science.1160873
Swanson, SA, Crow, SJ, Le Grange, D., Swendsen, J. og Merikangas, KR (2011). Algengi og fylgni með átröskunum hjá unglingum. Niðurstöður úr þjóðhagsskýrslu eftirlíkingar unglinga viðbót. Arch. Geðlækningar 68, 714-723. doi: 10.1001 / archgenpsychiatry.2011.22
Te Morenga, L., Mallard, S. og Mann, J. (2013). Mataræði og líkamsþyngd: kerfisbundin endurskoðun og meta-greining á slembiröðuðum samanburðarrannsóknum og hóprannsóknum. BMJ 346: e7492. doi: 10.1136 / bmj.e7492
Totterdell, S. og Smith, AD (1989). Samleitni hippocampal og dopaminergic inntak á greind taugafrumum í kjarnanum accumbens af rottum. J. Chem. Neuroanat. 2, 285-298.
Tukey, DS, Ferreira, JM, Antoine, SO, D'Amour, JA, Ninan, I., Cabeza de Vaca, S., et al. (2013). Inntöku súkrósa framkallar hratt AMPA viðtaka. J. Neurosci. 33, 6123-6132. doi: 10.1523 / JNEUROSCI.4806-12.2013
Veening, JG, Cornelissen, FM og Lieven, PA (1980). Staðbundin stofnun afferents til caudatoputamen á rottum. Peroxidasa rannsókn á piparrótrót. Neuroscience 5, 1253–1268. doi: 10.1016/0306-4522(80)90198-0
Ventura, T., Santander, J., Torres, R. og Contreras, AM (2014). Neurobiologic grundvelli þrá fyrir kolvetni. Næring 30, 252-256. doi: 10.1016 / j.nut.2013.06.010
Volkow, ND, Wang, GJ, Fowler, JS, Tomasi, D. og Baler, R. (2012). Matur og eiturlyf laun: skarast hringrás í offitu og fíkn manna. Curr. Efst. Behav. Neurosci. 11, 1–24. doi: 10.1007/7854_2011_169
Vitur, RA (1973). Taka á fúsum etanóli í rottum eftir að hafa verið sýnt fram á etanól á ýmsum tímum. Psychopharmacologia 29, 203-210. gera: 10.1007 / BF00414034
Wright, CI og Groenewegen, HJ (1995). Mynstur samleitni og aðgreiningar í miðgildum kjarnanum sem fylgir rottum: Sambönd af prefrontal cortical, midline thalamic og basal amygdaloid afferents. J. Comp. Neuról. 361, 383-403. doi: 10.1002 / cne.903610304
Leitarorð: Binge-eins og neysla, langtíma, miðlungs spiny neuron, kjarna accumbens, súkrósa
Tilvitnun: Klenowski PM, Shariff MR, Belmer A, Fogarty MJ, Mu EWH, Bellingham MC og Bartlett SE (2016) Langvarandi neysla súkrósa í binge-svipaðri hátt, breytir formgerð miðlungs taugaþyrpinga í Nucleus Accumbens Shell. Framan. Behav. Neurosci. 10: 54. doi: 10.3389 / fnbeh.2016.00054
Móttekið: 03 Desember 2015; Samþykkt: 07 mars 2016;
Útgefið: 23 mars 2016.
Breytt af:
Djoher Nora Abrous, Institut des Neurosciences de Bordeaux, Frakklandi
Yfirfarið af:
Serge H. Ahmed, Centre National de la Recherche Scientifique, Frakklandi
Stéphanie Caille, Centre National de la Recherche Scientifique, Frakklandi
Höfundarréttur © 2016 Klenowski, Shariff, Belmer, Fogarty, Mu, Bellingham og Bartlett. Þetta er opið aðgangs grein sem er dreift samkvæmt skilmálum þess Creative Commons Attribution License (CC BY). Notkun, dreifing eða fjölgun á öðrum vettvangi er leyfileg, að því tilskildu að upphaflegir höfundar eða leyfisveitendur séu látnir í té og að frumritið í þessari dagbók sé vitnað í samræmi við viðurkenndan fræðilegan starfsvenja. Ekki er heimilt að nota, dreifa eða endurskapa sem uppfyllir ekki þessa skilmála.
* Bréfaskipti: Selena E. Bartlett, [netvarið]