COMMENTS: While the study uses the term “Compulsive sexual behavior (CSB),” the subjects were porn addicts (see this press release). Compared to healthy controls CSB subjects had increased left amygdala volume and reduced functional connectivity during rest between the left amygdala and bilateral dorsolateral prefrontal cortex DLPFC. The authors conclude:
Our current findings highlight elevated volumes in a region implicated in motivational salience and lower resting state connectivity of prefrontal top-down regulatory control networks. Disruption of such networks may explain the aberrant behavioral patterns toward environmentally salient reward or enhanced reactivity to salient incentive cues. Although our volumetric findings contrast with those in substance use disorders (SUD), these findings may reflect differences as a function of the neurotoxic effects of chronic drug exposure.
Translation finding #1): “Less functional connectivity between the amygdala & dorsolateral prefrontal cortex.” The amygdala plays a key role in processing emotions, including our response to stress. The amygdala is strongly implicated in many aspects of addiction such as cravings, cue-reactivity and withdrawal symptoms. Reduced functional connectivity between the amygdala and the prefrontal cortex aligns with substance addictions. It is thought that poorer connectivity diminishes the prefrontal cortex’s control over a user’s impulse to engage in the addictive behavior.
Translation finding #2): “Increased amygdala volume” (which means more gray matter). Most drug addiction studies report a smaller amygdalae in addicts (less gray matter). This study suggests that drug toxicity may lead to less gray matter and thus reduced amygdala volume in drug addicts. No doubt this plays a role. It must be noted that the amygdala is consistently active during porn viewing, especially during initial exposure to a sexual cue. For example, clicking from tab to tab or searching for a video or image would light up the amygdala. Perhaps the constant sexual novelty and searching and seeking leads to a unique effect on the amygdala in compulsive porn users.
Alternative explanation for greater amygdala volume in porn addicts: Years of compulsive porn use can certainly be a stressor. Moreover, these CSB subjects were not only porn addicts; they also experienced severe negative consequences as a result of porn use (loss of job, relationship problems, development of porn-induced ED). Here’s a key point: Chronic social stress is related to increased amygdala volume:
Although the precise mechanisms of plasticity are still not fully understood, moderate to severe stress appears to increase the growth of several sectors of the amygdala, whereas the effects in the hippocampus and prefrontal cortex tend to be opposite.
We consider the above finding in light of this 2015 study that found that “sex addicts” have a hyperactive HPA axis (an overactive stress system). Could the chronic stress related to porn/sex addiction, along with factors that make sex unique, lead to greater amygdala volume? Finally, lower amygdala volume might be a pre-existing condition in alcoholics, as offspring in families with a high risk of alcoholism have smaller amygdalae.
LINK TO FULL STUDY
Casper Schmidt,1,2,3 Laurel S. Morris,1,4 Timo L. Kvamme,1,2,3 Paula Hall,5 Thaddeus Birchard,5 and Valerie Voon1,4,6*
- First published: 27 October 2016 Full publication history
- DOI: 10.1002/hbm.23447 View/save citation
The authors declare that they have no conflict of interests to declare.
Abstract
Background
Compulsive sexual behaviors (CSB) are relatively common and associated with significant personal and social dysfunction. The underlying neurobiology is still poorly understood. The present study examines brain volumes and resting state functional connectivity in CSB compared with matched healthy volunteers (HV).
Methods
Structural MRI (MPRAGE) data were collected in 92 subjects (23 CSB males and 69 age-matched male HV) and analyzed using voxel-based morphometry. Resting state functional MRI data using multi-echo planar sequence and independent components analysis (ME-ICA) were collected in 68 subjects (23 CSB subjects and 45 age-matched HV).
Results
CSB subjects showed greater left amygdala gray matter volumes (small volume corrected, Bonferroni adjusted P < 0.01) and reduced resting state functional connectivity between the left amygdala seed and bilateral dorsolateral prefrontal cortex (whole brain, cluster corrected FWE P < 0.05) compared with HV.
Conclusions
CSB is associated with elevated volumes in limbic regions relevant to motivational salience and emotion processing, and impaired functional connectivity between prefrontal control regulatory and limbic regions. Future studies should aim to assess longitudinal measures to investigate whether these findings are risk factors that predate the onset of the behaviors or are consequences of the behaviors.
Abbreviations
- ACC anterior cingulate cortex
- CSB compulsive sexual behavior
- CSF cerebrospinal fluid
- DLPFC dorsolateral prefrontal cortex
- GM gray matter
- GLM general linear model
- HV healthy volunteers
- MPRAGE magnetization prepared gradient-echo
- OFC orbitofrontal cortex
- ROI region of interest
- SPM Statistical Parametric Mapping
- TR repetition time
- TE echo time
- VBM voxel-based morphometry
- WM white matter.
INTRODUCTION
Compulsive sexual behavior (CSB), also known as hypersexual disorder or sexual addiction, is relatively common (estimated at 3%–6%) [Kraus et al., 2016] and associated with significant distress and psychosocial impairments, including being characterized by craving, impulsivity, and social and occupational impairment [Kraus et al., 2016]. Recent studies have focused on understanding underlying neurobiological correlates [Kraus et al., 2016] although the paucity of studies limits our understanding of underlying mechanisms and how we might conceptualize these disorders. CSB has been conceptualized as either an impulse control disorder or a behavioral addiction [Kraus et al., 2016]. However, although criteria for hypersexual disorder were proposed for the DSM-5 and validated in the field trial [Reid et al., 2012], this disorder along with the pathological use of the internet or video games, were not included in the main section of the DSM-5, in part due to limited data on the conditions. Thus, further studies on CSB are necessary to develop a greater understanding of these disorders. Although CSB can have a range of behaviors, here we focus on a group reporting predominant difficulties with compulsive pornography use. We have used the term CSB on the assumption that “compulsive” describes the repetitive phenomenology and is not intended to imply any mechanistic or etiological assumptions.
We conducted a review of the literature on behavioral addictions using either voxel-based morphometry (VBM) or cortical thickness. We used the following search words on PubMed (http://www.ncbi.nlm.nih.gov/pubmed): ‘[(“voxel-based morphometry” or “cortical thickness”) and],’ followed by either “[pathological gambling],” “[internet addiction],” “[internet disorder],” “,” or “[gaming addiction].” In total, 13 studies were found within behavioral addictions related to gambling, internet use, or video gaming that assessed either VBM or cortical thickness. The review of the literature is presented in Table 1 and discussed below.
Table 1. Literature review of volumetric and cortical thickness studies on behavioral addictions
|
Title |
Behavioral addiction |
Subjects (P/HV) |
Measure |
Regions implicated |
|---|---|---|---|---|
|
||||
| [Grant et al., 2015] | Gambling disorder | 16/17 | Cortical thickness | Decreased cortical thickness in r-SFC, RMFC, MOFC, PCG and bl-IPC |
| [Joutsa et al., 2011] | Pathological gambling | 12/12 | Voxel-based morphometry | No volumetric differences in GM or WM between HV and patients |
| [Koehler et al., 2013] | Pathological gambling | 20/21 | Voxel-based morphometry | Increased GM volume in bl-VS and r-PFC |
| [van Holst et al., 2012] | Problem gambling | 40/54 | Voxel-based morphometry | No volumetric differences in GM or WM between problem gamblers and HV |
| [Hong et al., 2013] | Internet addiction | 15/15 | Cortical thickness | Decreased cortical thickness in r-LOFC |
| [Yuan et al., 2011] | Internet addiction | 18/18 | Voxel-based morphometry | Decreased GM volume in DLPFC, SMA, OFC, CB, RACC |
| [Zhou et al., 2011] | Internet addiction | 15/18 | Voxel-based morphometry | Decreased GM density in l-ACC, PCC, IC, LING |
| [Lin et al., 2014] | Internet gaming addiction | 35/36 | Voxel-based morphometry | Decreased GM density in the IFG, l-CG, IC, and r-HIPP
Decreased WM density in IFG, IC, IPC, ACC |
| [Sun et al., 2014] | Internet gaming addiction | 18/21 | Voxel-based morphometry | Increased GM volume in the r-ITG, MTG, PHG
Decreased GM volume in l-PrCG |
| [Wang et al., 2015] | Internet gaming disorder | 28/28 | Voxel-based morphometry | Decreased GM volume in ACC, PCUN, SMA, SPC, and l-DLPFC, IC, CB |
| [Cai et al., 2015] | Internet gaming disorder | 27/30 | Subcortical volume, FreeSurfer | Increased volumes of CN and VS |
| [Weng et al., 2013] | Online game addiction | 17/17 | Voxel-based morphometry | Decreased GM volume in r-OFC, SMA and bl-IC |
| [Yuan et al., 2013] | Online game addiction | 18/18 | Cortical thickness | Increased cortical thickness in l-PrCG, PCUN, MFC, ITG, MTG
Decreased cortical thickness in l-LOFC, IC, r-PCG, IPC |
Insight into neural disturbances in addictions comes from studies of substance use disorders (SUD). Individuals with SUD show decreases in cortical brain volume and thickness particularly in prefrontal cortical regions that subserve flexible behavioral control. A recent meta-analysis of 9 studies and 296 alcohol dependent individuals found significantly reduced prefrontal gray matter (GM) volumes, including anterior cingulate cortex (ACC) [Xiao et al., 2015], with frontal cortical GM volume being negatively associated with lifetime alcohol use [Taki et al., 2006]. Prefrontal GM volumes were similarly reduced in cocaine dependent individuals, including in orbitofrontal cortex (OFC) [Rando et al., 2013; Tanabe et al., 2009], anterior prefrontal cortex [Rando et al., 2013] and ACC [Connolly et al., 2013], the latter associated with years of drug use [Connolly et al., 2013].
Group differences in cortical volumes and thickness have been less clear in behavioral addictions (reviewed in Table 1). Three small studies of gambling disorder showed inconsistent findings with either decreased cortical thickness in multiple prefrontal and parietal regions [Grant et al., 2015], increased volumes in the right prefrontal cortex [Koehler et al., 2013] or no group differences [Joutsa et al., 2011]. In a large study of less severe problem gamblers, no group differences were observed in brain volumes [van Holst et al., 2012]. One small study in internet addiction showed lower cortical thickness in the OFC [Hong et al., 2013], with another reporting lower volume in dorsolateral prefrontal cortex (DLPFC) [Yuan et al., 2011] and two studies suggesting lower ACC volumes [Yuan et al., 2011; Zhou et al., 2011]. Two small studies in internet gaming disorders reported decreased volumes in OFC [Weng et al., 2013; Yuan et al., 2013], and two larger studies reported smaller volumes in cingulate cortex [Lin et al., 2014; Wang et al., 2015] with single studies reporting decreases in DLPFC [Wang et al., 2015], inferior frontal [Lin et al., 2014], superior parietal [Wang et al., 2015] and inferior parietal [Yuan et al., 2013] cortices. With respect to subcortical structures, one small study reported higher ventral striatal (VS) volumes in gambling disorder [Koehler et al., 2013] with no subcortical differences reported in the other studies. In internet gaming disorder, findings were similarly inconsistent with either greater parahippocampal [Sun et al., 2014], lower hippocampal [Lin et al., 2014] or no differences [Wang et al., 2015; Weng et al., 2013]. One study with a reasonable sample size focusing on subcortical volumes reported greater caudate and VS volumes associated with cognitive control deficits [Cai et al., 2015]. Taken together, the findings of cortical or subcortical abnormalities in gambling disorder are highly inconsistent. In contrast, reports of cortical abnormalities in internet use or internet gaming more consistently report decreased volumes with decreased ACC and OFC volume replicated across at least two studies.
To date, there is sparse evidence of structural neural changes in individuals with CSB. Studies of healthy individuals with excessive pornography use without a diagnosis of CSB show lower GM volumes in the right caudate [Kühn and Gallinat, 2014]. A small diffusion MRI study of individuals with CSB (N = 8 per group) showed reduced mean diffusivity in superior frontal white matter (WM) tracts compared with HV [Miner et al., 2009]. With respect to functional activity, male HV show enhanced habituation processes with lower left putaminal BOLD activity to static erotic images [Kühn and Gallinat, 2014] and lower late positive potential to explicit images [Prause et al., 2015]. In contrast, in a task-based fMRI study comparing CSB with HV, explicit sexual videos elicited higher VS, amygdala and dorsal ACC BOLD responses in CSB [Voon et al., 2014]. Functional connectivity between these regions correlated with an index of sexual desire or “wanting” but not “liking” in CSB subjects suggesting the role of incentive motivation, paralleling substance addiction. Similarly, in another study in internet pornography addiction, the preferred sexual image was associated with greater ventral striatal activity and correlated only with self-reported symptoms of internet pornography addiction and not with other measures of sexual behavior or depression [Brand et al., 2016]. Another recent study also shows that individuals with problematic hypersexual behavior experienced more frequent and enhanced sexual desire during exposure to sexual stimuli, and that greater activation was observed in the caudate, inferior parietal lobe, dorsal anterior cingulate gyrus, thalamus, and DLPFC in this group [Seok and Sohn, 2015]. CSB individuals further show greater early attentional bias to explicit sexual stimuli [Mechelmans et al., 2014] that correlated with choice preferences for cues conditioned to sexual images [Banca et al., 2016]. In response to repeated exposure of static erotic images, CSB subjects showed greater habituation in the dorsal ACC to sexual outcomes, which correlated with choice preferences for novel sexual images [Banca et al., 2016], an effect that may be explained by either habituation but might also be consistent with the concept of tolerance in addictions.
The current study examines volumetric GM in CSB and review the current literature on volumetric and cortical thickness studies in gambling disorder and in internet and gaming use disorders. We also examine resting state functional connectivity of individuals with CSB and matched HV with a novel multi-echo planar sequence and independent components analysis (ME-ICA) wherein BOLD signals are identified as independent components with linear echo time (TE)-dependent signal change whereas non-BOLD signals are identified as TE-independent components [Kundu et al., 2012]. We expect a disrupted network of salience and reward-related systems subserved by amygdala, VS and dorsal ACC.
METHODS
Participants
CSB subjects were recruited via internet-based advertisements and from referrals from therapists. Age-matched male HV were recruited from community-based advertisements in the East Anglia area. All CSB subjects were interviewed by a psychiatrist to confirm they fulfilled diagnostic criteria for CSB (met proposed diagnostic criteria for both hypersexual disorder [Kafka, 2010; Reid et al., 2012] and sexual addiction [Carnes et al., 2007], focusing on compulsive use of online sexually explicit material. This was assessed using a modified version of the Arizona Sexual Experiences Scale (ASES) [Mcgahuey et al., 2011], in which questions were answered on a scale of 1–8, with higher scores representing greater subjective impairment. Given the nature of the cues, all CSB subjects and HV were male and heterosexual. All HV were age-matched (±5 years of age) with CSB subjects. Subjects were also screened for compatibility with the MRI environment as we have done previously [Banca et al., 2016; Mechelmans et al., 2014; Voon et al., 2014]. Exclusionary criteria included being under 18 years of age, having a history of SUD, being a current regular user of illicit substances (including cannabis), and having a serious psychiatric disorder, including current moderate-severe major depression or obsessive-compulsive disorder, or history of bipolar disorder or schizophrenia (screened using the Mini International Neuropsychiatric Inventory) [Sheehan et al., 1998]. Other compulsive or behavioral addictions were also exclusions. Subjects were assessed by a psychiatrist regarding problematic use of online gaming or social media, pathological gambling or compulsive shopping, childhood or adult attention deficit hyperactivity disorder, and binge-eating disorder diagnosis. Subjects completed the UPPS-P Impulsive Behavior Scale [Whiteside and Lynam, 2001] to assess impulsivity, and the Beck Depression Inventory [Beck et al., 1961] to assess depression. Two of 23 CSB subjects were taking antidepressants or had comorbid generalized anxiety disorder and social phobia (N = 2) or social phobia (N = 1) or a childhood history of ADHD (N = 1). Written informed consent was obtained, and the study was approved by the University of Cambridge Research Ethics Committee. Subjects were paid for their participation.
Neuroimaging
Data acquisition and processing
Structural.
Structural images were collected including full magnetization prepared gradient-echo (MPRAGE) using a Siemens Tim Trio 3T-scanner with a 32-channel head coil using a T1 weighted MPRAGE sequence (176 sagittal slices, 9 minute scans; repetition time (TR) = 2,500 ms; echo time (TE) = 4.77 ms; inversion time = 1,100 ms; acquisition matrix = 256 × 256 × 176; flip angle = 7°; voxel size 1 × 1 × 1 mm). Scanning took place at The Wolfson Brain Imaging Centre at the University of Cambridge.
Structural data was processed with Statistical Parametric Mapping (SPM8; http://www.fil.ion.ucl.ac.uk/spm) (Wellcome Trust Centre for Neuroimaging, London, UK). Anatomical images were manually re-oriented, placing the origin at the anterior commissure. Images were segmented (using New Segment for SPM) into GM, WM and cerebrospinal fluid (CSF) based on standard tissue probability maps for each tissue type. The three tissue class volumes were summed to produce estimated total intracranial volume. A custom template was created using DARTEL [Ashburner, 2007], which defines the parameters necessary to fit each individuals’ native GM image to a common space, in an iterative manner. This DARTEL template was then registered to the tissue probability maps with affine transformations, bringing images into MNI space. Images were smoothed spatially with a full width at half maximum kernel of 8 mm3.
Resting state.
Resting state fMRI data were acquired for 10 minutes with eyes open with a Siemens 3T Tim Trio scanner with a 32-channel head coil at the Wolfson Brain Imaging Centre, University of Cambridge. A multi-echo echo planar imaging sequence was used with online reconstruction (repetition time, 2.47 s; flip angle, 78°; matrix size 64 × 64; in-plane resolution, 3.75 mm; FOV, 240 mm; 32 oblique slices, alternating slice acquisition slice thickness 3.75 mm with 10% gap; iPAT factor, 3; bandwidth = 1,698 Hz/pixel; echo time (TE) = 12, 28, 44, and 60 ms).
Multi-echo independent component analysis (ME-ICAv2.5 beta6; http://afni.nimh.nih.gov) was used for analysis and de-noising of the multi-echo resting state fMRI data. ME-ICA decomposes multi-echo fMRI data into independent components with FastICA. BOLD signal percent signal change is linearly dependent on TE, a characteristic of the T2* decay. This TE-dependence is measured using the pseudo-F-statistic, kappa, with components that scale strongly with TE having high kappa scores [Kundu et al., 2012]. Non-BOLD components are identified by TE independence measured by the pseudo-F-statistic, rho. Components are thus categorized as BOLD or non-BOLD based on their kappa and rho value weightings, respectively [Kundu et al., 2012]. Non-BOLD components are removed by projection, de-noising data for motion, physiological and scanner artifacts in a robust manner based on physical principles. Each individual’s de-noised echo planar images were coregistered to their MPRAGE and normalized to the Montreal Neurological Institute (MNI) template. Spatial smoothing was conducted with a Gaussian kernel (full width half maximum = 6 mm). The time course for each voxel was temporally band-pass filtered (0.008 < f < 0.09 Hz). Each individual’s anatomical scan was segmented into GM, WM, and CSF. Significant principal components of the signals from WM and CSF were removed.
Functional connectivity analysis was performed using a region of interest (ROI)-driven approach with CONN-fMRI Functional Connectivity toolbox [Whitfield-Gabrieli and Nieto-Castanon, 2012] for SPM (http://www.fil.ion.ucl.ac.uk/spm/software/spm8/).
Statistical Analysis
Subjects’ characteristics and questionnaire scores were compared between groups with two-tailed t-tests without assuming equal variance. All statistical analyses were performed using R version (3.2.0) [RC Team, 2014].
Structural
For group comparisons, GM volumes for CSB subjects and HV were entered into a general linear model (GLM). Data was corrected for participants’ total intracranial volume using proportional scaling and an explicit mask in SPM. Group comparisons were adjusted for both age and depression scores as covariates. We focused on a priori hypothesized regions of interest identified in our previous study [Voon et al., 2014] and in meta-analyses of drug cue reactivity studies [Kühn and Gallinat, 2011], namely left and right VS, left and right amygdala, and dorsal ACC using small volume corrected (SVC) family-wise error (FWE) corrected P < 0.01 (Bonferroni corrected for multiple comparisons). For these SVC analyses, we used a VS anatomical ROI, previously described [Murray et al., 2008] which was hand drawn using MRIcro based on the definition of VS by Martinez et al. [2003]. The amygdala ROI was obtained from the Automated Anatomical Labelling (AAL) atlas. The dorsal ACC was manually altered using MarsBaR ROI toolbox [Brett et al., 2002] and based on the cingulate cortex ROI from the AAL atlas. It was modified such that the anterior border was the tip of the genu of the corpus callosum [Cox et al., 2014; Desikan et al., 2006] and the posterior was the posterior end of the genu of the corpus callosum [Desikan et al., 2006]. Additional analyses adjusting for BDI scores were performed.
Resting state
To compare connectivity between CSB subjects and HV, ROI-to-voxel whole brain connectivity maps were computed for the left amygdala seed region of interest based on volumetric group difference findings. Resultant connectivity maps were entered into full factorial GLM’s to compare whole-brain connectivity between groups adjusting for age with a subsequent analysis adjusting for both age and depression. Whole brain cluster corrected FWE P < 0.05 was considered significant for group differences.
RESULTS
Characteristics
Twenty-three heterosexual men with CSB (age 26.9; SD 6.22 years) and 69 age-matched (age 25.6; SD 6.55 years) heterosexual male HV participated in the study (Table 2), of which 19 CSB subjects and 55 HV completed behavioral questionnaires. CSB subjects had higher BDI (P = 0.006) and UPPS-P (P < 0.001) scores compared with HV. Other behavioral scores including pattern and severity of pornography and internet use have been reported elsewhere [Mechelmans et al., 2014; Voon et al., 2014].
Table 2. Demographic and behavioral data for compulsive sexual behavior subjects and healthy volunteers
|
Group |
Age |
BDI |
UPPS-P |
|---|---|---|---|
Missing 4 participants out of 23.
Missing 14 participants out of 69.
|
|||
| CSB (N = 23) | 26.9 (6.22) | 14.82 (11.85)a | 152.21 (16.50)a |
| HV (N = 69) | 25.6 (6.55) | 6.03 (7.20)b | 124.87 (20.73)b |
| T-value (P-value) | 0.88 (P = 0.380) | 3.04 (P = 0.006) | 5.81 (P < 0.001) |
Structural
The ROI analyses of left and right amygdala, left and right VS and dorsal ACC revealed that left amygdala grey matter volume was increased in CSB compared with matched HV (SVC FWE-corrected, P = 0.0096, Z = 3.37, xyz = −28, −4, −15) (Bonferroni corrected for SVC FWE-corrected P < 0.01) (Fig. 1). All other ROI analyses were not significant. Adjusting for depression did not change the group difference findings.

Figure 1.
{kind=link}
Voxel-based morphometry in compulsive sexual behaviors. Greater left amygdala volume is shown in compulsive sexual behaviors relative to healthy volunteers. The image is thresholded at P < 0.005 uncorrected for illustration. [Color figure can be viewed at wileyonlinelibrary.com]
Resting State
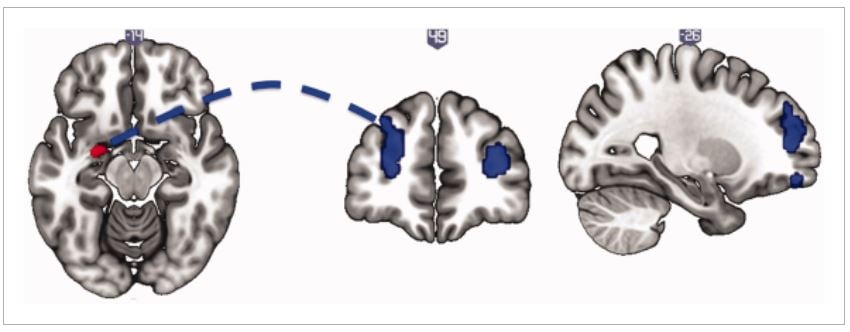
Based on the structural results, we examined resting state functional connectivity with a seed in the left amygdala. We found reduced connectivity with bilateral DLPFC (Right DLPFC: P = 0.012, Z = 4.11, xyz = 31 42 16; Left DLPFC: P = 0.003, Z = 3.96, xyz = −27 52 23) (Fig. 2). Adjusting for BDI did not alter the significance of the findings (Right DLPFC: P = 0.001, Z = 4.54, xyz = 31 61 23; Left DLPFC: P = 0.003, Z = 4.26, xyz = −29 49 35).

Figure 2.
{kind=link}
Resting state functional connectivity of the left amygdala. Compulsive sexual behavior is associated with decreased resting state functional connectivity of the left amygdala (seed, left) with bilateral dorsolateral prefrontal cortex (middle and right), relative to healthy volunteers. The image is thresholded at P < 0.005 uncorrected for illustration. [Color figure can be viewed at wileyonlinelibrary.com]
DISCUSSION
We investigated structural and functional neural differences in individuals with CSB compared with matched HV. CSB subjects had increased left amygdala volume and reduced functional connectivity during rest between the left amygdala and bilateral DLPFC.
The amygdala is implicated in environmental salience processing that directs behavior. The nuclei of the amygdala link previously neutral environmental or internal stimuli with associative representations of affective value, propagating cue-induced motivational salience [Everitt et al., 2003], as well as processing of emotional control [Cardinal et al., 2002; Gottfried et al., 2003]. The finding of increased amygdala volume is in opposition with several studies on alcohol use disorders [Makris et al., 2008; Wrase et al., 2008], as studies across this type of addiction report decreased amygdala volumes, where volumetric measures have been assessed. A potential explanation for this discrepancy is that long-term substance use result in long-lasting neuroplastic changes and toxicity [Kovacic, 2005; Reissner and Kalivas, 2010] that may contribute to the perseveration of drug-seeking behavior [Gass and Olive, 2008]. Such neurotoxicity may certainly contribute to the widespread atrophy observed in substance addictions [Bartzokis et al., 2000; Carlen et al., 1978; Mechtcheriakov et al., 2007]. Such drug-related neurotoxicity is likely a highly relevant issue in SUD but less of an issue in behavioral addictions. In a recent CSB study using fMRI, exposure to sexually explicit cues in CSB compared with non-CSB subjects was associated with activation of the amygdala [Voon et al., 2014]. Whether the difference in amygdala volume is a pre-existing trait predisposing individuals to CSB or related to excessive exposure remains to be established.
The functioning of the DLPFC is well known to be associated with broad aspects of cognitive control [MacDonald et al., 2000] and working memory [Petrides, 2000]. Our finding of decreased functional connectivity between amygdala and DLPFC converges with the existing literature on the connectivity in these regions. This functional connectivity is important for emotion regulation, which has previously been reported in that reduced connectivity between amygdala and DLPFC in individuals with internet gaming disorder is associated with higher levels of impulsivity [Ko et al., 2015]. Another study measuring the capacity to modulate negative emotional responses through the use of cognitive strategies showed that activity in specific areas of the frontal cortex, including DLPFC, covaried with amygdala activity, and that functional connectivity between these regions was dependent on applying cognitive strategies in the regulation of negative emotion [Banks et al., 2007]. Amygdala and DLPFC connectivity has similarly been associated with unipolar depression [Siegle et al., 2007]. CSB has been associated with depressive and anxiety symptoms and stress may trigger such activities; however, our findings were unrelated to depression scores. The DLPFC was also implicated in a study of male HV in which greater pornography use was associated with lower functional connectivity between the DLPFC and striatum when viewing explicit imagery [Kühn and Gallinat, 2014].
We caution that these findings are preliminary given the small sample size of CSB subjects although notably we compare this group with a large sample size of matched HV. One limitation of the study is the homogeneity of the population. As we did not include subjects with other comorbid psychiatric disorders which might play a mechanistic role, these results should be cautiously extrapolated to CSB subjects with other comorbidities. Furthermore, the observed structural and functional abnormalities among the CSB subjects may be related to pre-existing traits or may be a result of the effects of CSB, and as such this study cannot make causal inferences about the effects of CSB. Future studies should aim to assess longitudinal measures to determine differences between state and trait tendencies and potential pre-morbid neural abnormalities in larger sample sizes and with mixed genders.
Our current findings highlight elevated volumes in a region implicated in motivational salience and lower resting state connectivity of prefrontal top-down regulatory control networks. Disruption of such networks may explain the aberrant behavioral patterns toward environmentally salient reward or enhanced reactivity to salient incentive cues. Although our volumetric findings contrast with those in SUD, these findings may reflect differences as a function of the neurooxic effects of chronic drug exposure. Emerging evidence suggests potential overlaps with an addiction process particularly supporting incentive motivation theories. We have shown that activity in this salience network is then enhanced following exposure to highly salient or preferred sexually explicit cues [Brand et al., 2016; Seok and Sohn, 2015; Voon et al., 2014] along with enhanced attentional bias [Mechelmans et al., 2014] and desire specific to the sexual cue but not generalized sexual desire [Brand et al., 2016; Voon et al., 2014]. Enhanced attention to sexually explicit cues is further associated with preference for sexually conditioned cues thus confirming the relationship between sexual cue conditioning and attentional bias [Banca et al., 2016]. These findings of enhanced activity related to sexually conditioned cues differ from that of the outcome (or the unconditioned stimulus) in which enhanced habituation, possibly consistent with the concept of tolerance, increases the preference for novel sexual stimuli [Banca et al., 2016]. Together these findings help elucidate the underlying neurobiology of CSB leading toward a greater understanding of the disorder and identification of possible therapeutic markers.
ACKNOWLEDGMENTS
We would like to thank the WBIC staff for their expertise and assistance with collecting the imaging data, and our participants for their time and commitment. Also, we would like to thank Thaddeus Birchard and Paula Hall for referral of patients for the study. The Behavioural and Clinical Neuroscience Institute (BCNI) is supported by the Wellcome Trust and Medical Research Council.
REFERENCES
- Ashburner J (2007): A fast diffeomorphic image registration algorithm. Neuroimage 38:95–113.
- Banca P, Morris LS, Mitchell S, Harrison NA, Potenza MN, Voon V (2016): Novelty, conditioning and attentional bias to sexual rewards. J Psychiatr Res 72:91–101.
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 2
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 307
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 75
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times Cited: 18323
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 1
- Banks SJ, Eddy KT, Angstadt M, Nathan PJ, Phan KL (2007): Amygdala–frontal connectivity during emotion regulation. Soc Cogn Affect Neurosci 2:303–312.
- CrossRef |
- Web of Science® Times Cited: 1
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 1087
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times Cited: 342 |
- ADS
- Bartzokis G, Beckson M, Lu PH, Edwards N, Rapoport R, Wiseman E, Bridge P (2000): Age-related brain volume reductions in amphetamine and cocaine addicts and normal controls: Implications for addiction research. Psychiatry Res Neuroimaging 98:93–102.
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 16 |
- ADS
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 7
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 1782
- Wiley Online Library |
- PubMed |
- Web of Science® Times Cited: 245
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times Cited: 217
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times Cited: 597 |
- ADS
- CrossRef |
- PubMed |
- Web of Science®
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 19
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 27
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 21
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 172
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 8
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 5
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times Cited: 30
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times Cited: 1
- Wiley Online Library |
- PubMed |
- Web of Science® Times Cited: 76
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 23
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 63
- Beck AT, Ward C, Mendelson M (1961): Beck depression inventory (BDI). Arch Gen Psychiatry 4:561–571.
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times Cited: 1895 |
- ADS
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times Cited: 134
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times Cited: 313
- Brand M, Snagowski J, Laier C, Maderwald S (2016): entral striatum activity when watching preferred pornographic pictures is correlated with symptoms of internet pornography addiction. Neuroimage 129:224–232.
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 7 |
- ADS
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 70
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 28
- CrossRef |
- CAS |
- Web of Science® Times Cited: 196
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times Cited: 255
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 3
- Wiley Online Library |
- PubMed |
- Web of Science® Times Cited: 20
- Brett M, Anton J-L, Valabregue R, Poline J-B (2002): Region of interest analysis using the MarsBar toolbox for SPM 99. Neuroimage 16:S497.
- Wiley Online Library |
- PubMed |
- Web of Science® Times Cited: 43
- CrossRef |
- PubMed |
- CAS |
- Web of Science® Times Cited: 63
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 1
- PubMed |
- Web of Science® Times Cited: 7675
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 383
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 4
- Wiley Online Library |
- PubMed |
- Web of Science® Times Cited: 38
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 110
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 25
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 3
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 25
- CrossRef |
- Web of Science® Times Cited: 1108
- CrossRef |
- PubMed
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 92
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 3
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 72 |
- ADS
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 31 |
- ADS
- CrossRef |
- PubMed |
- Web of Science® Times Cited: 66
- Cai C, Yuan K, Yin J, Feng D, Bi Y, Li Y, Yu D, Jin C, Qin W, Tian J (2015): Striatum morphometry is associated with cognitive control deficits and symptom severity in internet gaming disorder. Brain Imaging Behav 10:12.
- Cardinal RN, Parkinson JA, Hall J, Everitt BJ (2002): Emotion and motivation: The role of the amygdala, ventral striatum, and prefrontal cortex. Neurosci Biobehav Rev 26:321–352.
- Carlen PL, Wortzman G, Holgate RC, Wilkinson DA, Rankin JC (1978): Reversible cerebral atrophy in recently abstinent chronic alcoholics measured by computed tomography scans. Science 200:1076–1078.
- Carnes P, Delmonico DL, Griffin E (2007): In the Shadows of the Net: Breaking Free of Compulsive Online Sexual Behavior, 2nd ed. Center City, MN: Hazelden Publishing.
- Connolly CG, Bell RP, Foxe JJ, Garavan H (2013): Dissociated grey matter changes with prolonged addiction and extended abstinence in cocaine users. PLoS One 8:e59645.
- Cox SR, Ferguson KJ, Royle NA, Shenkin SD, MacPherson SE, MacLullich AMJ, Deary IJ, Wardlaw JM (2014): A systematic review of brain frontal lobe parcellation techniques in magnetic resonance imaging. Brain Struct Funct 219:1–22.
- Desikan RS, Ségonne F, Fischl B, Quinn BT, Dickerson BC, Blacker D, Buckner RL, Dale AM, Maguire RP, Hyman BT (2006): An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. Neuroimage 31:968–980.
- Everitt BJ, Cardinal RN, Parkinson JA, Robbins TW (2003): Appetitive behavior: Impact of amygdala-dependent mechanisms of emotional learning. Ann N Y Acad Sci 985:233–250.
- Gass JT, Olive MF (2008): Glutamatergic substrates of drug addiction and alcoholism. Biochem Pharmacol 75:218–265.
- Gottfried JA, O’Doherty J, Dolan RJ (2003): Encoding predictive reward value in human amygdala and orbitofrontal cortex. Science 301:1104–1107.
- Grant JE, Odlaug BL, Chamberlain SR (2015): Reduced cortical thickness in gambling disorder: A morphometric MRI study. Eur Arch Psychiatry Clin Neurosci 265:655–661.
- van Holst RJ, de Ruiter MB, van den Brink W, Veltman DJ, Goudriaan AE (2012): A voxel-based morphometry study comparing problem gamblers, alcohol abusers, and healthy controls. Drug Alcohol Depend 124:142–148.
- Hong S-B, Kim J-W, Choi E-J, Kim H-H, Suh J-E, Kim C-D, Klauser P, Whittle S, Yűcel M, Pantelis C, Yi S-H (2013): Reduced orbitofrontal cortical thickness in male adolescents with internet addiction. Behav Brain Funct 9:11.
- Joutsa J, Saunavaara J, Parkkola R, Niemelä S, Kaasinen V (2011): Extensive abnormality of brain white matter integrity in pathological gambling. Psychiatry Res – Neuroimaging 194:340–346.
- Kafka MP (2010): Hypersexual disorder: A proposed diagnosis for DSM-V. Arch Sex Behav 39:377–400.
- Ko C-H, Hsieh T-J, Wang P-W, Lin W-C, Yen C-F, Chen C-S, Yen J-Y (2015): Altered gray matter density and disrupted functional connectivity of the amygdala in adults with internet gaming disorder. Prog Neuropsychopharmacol Biol Psychiatry 57:185–192.
- Koehler S, Hasselmann E, Wüstenberg T, Heinz A, Romanczuk-Seiferth N (2013): Higher volume of ventral striatum and right prefrontal cortex in pathological gambling. Brain Struct Funct 220:469–477.
- Kovacic P (2005): Unifying mechanism for addiction and toxicity of abused drugs with application to dopamine and glutamate mediators: Electron transfer and reactive oxygen species. Med Hypotheses 65:90–96.
- Kraus SW, Voon V, Potenza MN (2016): Neurobiology of compulsive sexual behavior: Emerging science. Neuropsychopharmacology 41:385–386.
- Kühn S, Gallinat J (2011): Common biology of craving across legal and illegal drugs–a quantitative meta-analysis of cue-reactivity brain response. Eur J Neurosci 33:1318–1326.
- Kühn S, Gallinat J (2014): Brain structure and functional connectivity associated with pornography consumption: The brain on porn. JAMA Psychiatry 71:827–834.
- Kundu P, Inati SJ, Evans JW, Luh WM, Bandettini PA (2012): Differentiating BOLD and non-BOLD signals in fMRI time series using multi-echo EPI. Neuroimage 60:1759–1770.
- Lin X, Dong G, Wang Q, Du X (2014): Abnormal gray matter and white matter volume in “Internet gaming addicts.” Addict Behav 40C:137–143.
- MacDonald AW, Cohen JD, Stenger VA, Carter CS (2000): Dissociating the role of the dorsolateral prefrontal and anterior cingulate cortex in cognitive control. Science 288:1835–1838.
- Makris N, Oscar-Berman M, Jaffin SK, Hodge SM, Kennedy DN, Caviness VS, Marinkovic K, Breiter HC, Gasic GP, Harris GJ (2008): Decreased volume of the brain reward system in alcoholism. Biol Psychiatry 64:192–202.
- Martinez D, Slifstein M, Broft A, Mawlawi O, Chatterjee R, Hwang D-R, Huang Y, Cooper T, Kegeles L, Zarahn E, Abi-Dargham A, Haber SN, Laruelle M (2003): Imaging human mesolimbic dopamine transmission with positron emission tomography. Part II: Amphetamine-induced dopamine release in the functional subdivisions of the striatum. J Cereb Blood Flow Metab 23:285–300.
- Mcgahuey CA, Gelenberg AJ, Cindi A, Moreno FA, Delgado PL, Mcknight KM, Manber R (2011): Journal of sex & marital the arizona sexual experience scale (asex): reliability and validity. J Sex Marital Ther 26:37–41.
- Mechelmans DJ, Irvine M, Banca P, Porter L, Mitchell S, Mole TB, Lapa TR, Harrison NA, Potenza MN, Voon V (2014): Enhanced attentional bias towards sexually explicit cues in individuals with and without compulsive sexual behaviours. PLoS One 9:e105476.
- Mechtcheriakov S, Brenneis C, Egger K, Koppelstaetter F, Schocke M, Marksteiner J (2007): A widespread distinct pattern of cerebral atrophy in patients with alcohol addiction revealed by voxel-based morphometry. J Neurol Neurosurg Psychiatry 78:610–614.
- Miner MH, Raymond N, Mueller B. a, Lloyd M, Lim KO (2009): Preliminary investigation of the impulsive and neuroanatomical characteristics of compulsive sexual behavior. Psychiatry Res – Neuroimaging 174:146–151.
- Murray GK, Corlett PR, Clark L, Pessiglione M, Blackwell AD, Honey G, Jones PB, Bullmore ET, Robbins TW, Fletcher PC (2008): Substantia nigra/ventral tegmental reward prediction error disruption in psychosis. Mol Psychiatry 13:267–276.
- Petrides M (2000): The role of the mid-dorsolateral prefrontal cortex in working memory. Exp Brain Res 133:44–54.
- Prause N, Steele VR, Staley C, Sabatinelli D, Proudfit GH (2015): Modulation of late positive potentials by sexual images in problem users and controls inconsistent with “porn addiction”. Biol Psychol 109:192–199.
- Rando K, Tuit K, Hannestad J, Guarnaccia J, Sinha R (2013): Sex differences in decreased limbic and cortical grey matter volume in cocaine dependence: A voxel-based morphometric study. Addict Biol 18:147–160.
- RC Team (2014): R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing. ISBN 3-900051-07-0.
- Reid RC, Carpenter BN, Hook JN, Garos S, Manning JC, Gilliland R, Cooper EB, McKittrick H, Davtian M, Fong T (2012): Report of findings in a DSM-5 field trial for hypersexual disorder. J Sex Med 9:2868–2877.
- Reissner KJ, Kalivas PW (2010): Using glutamate homeostasis as a target for treating addictive disorders. Behav Pharmacol 21:514.
- Seok JW, Sohn JH (2015): Neural substrates of sexual desire in individuals with problematic hypersexual behavior. Front Behav Neurosci 9:1–11.
- Sheehan DV, Lecrubier Y, Sheehan KH, Amorim P, Janavs J, Weiller E, Hergueta T, Baker R, Dunbar GC (1998): The mini-international neuropsychiatric interview (mini): the development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J Clin Psychiatry 59:22–33.
- Siegle GJ, Thompson W, Carter CS, Steinhauer SR, Thase ME (2007): Increased amygdala and decreased dorsolateral prefrontal BOLD responses in unipolar depression: Related and independent features. Biol Psychiatry 61:198–209.
- Sun Y, Sun J, Zhou Y, Ding W, Chen X, Zhuang Z, Xu J, Du Y (2014): Assessment of in vivo microstructure alterations in gray matter using DKI in internet gaming addiction. Behav Brain Funct 10:37.
- Taki Y, Kinomura S, Sato K, Goto R, Inoue K, Okada K, Ono S, Kawashima R, Fukuda H (2006): Both global gray matter volume and regional gray matter volume negatively correlate with lifetime alcohol intake in non-alcohol-dependent Japanese men: A volumetric analysis and a voxel-based morphometry. Alcohol Clin Exp Res 30:1045–1050.
- Tanabe J, Tregellas JR, Dalwani M, Thompson L, Owens E, Crowley T, Banich M (2009): Medial orbitofrontal cortex gray matter is reduced in abstinent substance-dependent individuals. Biol Psychiatry 65:160–164.
- Voon V, Mole TB, Banca P, Porter L, Morris L, Mitchell S, Lapa TR, Karr J, Harrison NA, Potenza MN, Irvine M (2014): Neural correlates of sexual cue reactivity in individuals with and without compulsive sexual behaviours. PLoS One 9:e102419.
- Wang H, Jin C, Yuan K, Shakir TM, Mao C, Niu X, Niu C, Guo L, Zhang M (2015): The alteration of gray matter volume and cognitive control in adolescents with internet gaming disorder. Front Behav Neurosci 9:1–7.
- Weng C-B, Qian R-B, Fu X-M, Lin B, Han X-P, Niu C-S, Wang Y-H (2013): Gray matter and white matter abnormalities in online game addiction. Eur J Radiol 82:1308–1312.
- Whiteside SP, Lynam DR (2001): The five factor model and impulsivity: Using a structural model of personality to understand impulsivity. Pers Individ Diff 30:669–689.
- Whitfield-Gabrieli S, Nieto-Castanon A (2012): A functional connectivity toolbox for correlated and anticorrelated brain networks. Brain Connect 2:125–141.
- Wrase J, Makris N, Braus DF, Mann K, Smolka MN, Kennedy DN, Caviness VS, Hodge SM, Tang L, Albaugh M, Ziegler D. a, Davis OC, Kissling C, Schumann G, Breiter HC, Heinz A (2008): Amygdala volume associated with alcohol abuse relapse and craving. Am J Psychiatry 165:1179–1184.
- Xiao P, Dai Z, Zhong J, Zhu Y, Shi H, Pan P (2015): Regional gray matter deficits in alcohol dependence: A meta-analysis of voxel-based morphometry studies. Drug Alcohol Depend 153:22–28.
- Yuan K, Qin W, Wang G, Zeng F, Zhao L, Yang X, Liu P, Liu J, Sun J, von Deneen KM, Gong Q, Liu Y, Tian J (2011): Microstructure abnormalities in adolescents with internet addiction disorder. PLoS One 6:e20708.
- Yuan K, Cheng P, Dong T, Bi Y, Xing L, Yu D, Zhao L, Dong M, von Deneen KM, Liu Y, Qin W, Tian J (2013): Cortical thickness abnormalities in late adolescence with online gaming addiction. PLoS One 8:e53055.
- Zhou Y, Lin FC, Du YS, Qin LD, Zhao ZM, Xu JR, Lei H (2011): Gray matter abnormalities in Internet addiction: A voxel-based morphometry study. Eur J Radiol 79:92–95.