Fronto. Psikiatrio, 25 Februaro 2016 | http://dx.doi.org/10.3389/fpsyt.2016.00024
Jarid Goodman kaj Mark G. Packard*
- Fako de Psikologio, Teksasa A&M-Instituto por Neŭroscienco, Teksasa A&M-Universitato, College Station, TX, Usono
La vidpunkto, ke anatomie apartaj memorsistemoj malsame kontribuas al disvolviĝo de drogmanio kaj reaperado ricevis ampleksan subtenon. La nuna mallonga recenzo recenzas ĉi tiun hipotezon, ĉar ĝi estis origine proponita antaŭ 20 jaroj (1) kaj elstarigas plurajn lastatempajn evoluojn. Vasta esplorado uzanta diversajn paradigmojn de bestaj lernado indikas, ke disaj neŭralaj sistemoj mediacias malsamajn specojn de lernado kaj memoro. Ĉiu memorsistemo potenciale kontribuas unikajn komponentojn al la lernita konduto subtenanta drogmanion kaj recesion. Precipe, la ŝanĝo de distra uzado de drogoj al deviga droguzado eble reflektas neŭroatomikan movon de kognitiva kontrolo de konduto mediata de hipokampo / dorsomedia striato al kutima kontrolo de konduto mediata de la dorsolatera striatum (DLS). Krome, streĉo / angoro povas konsistigi kofaktoron, kiu faciligas DLS-dependan memoron, kaj tio povas servi kiel neŭrohavora mekanismo sub la pliigita droguzo kaj reaperado en homoj post streĉaj vivokazaĵoj. La evidenteco kiu subtenas la vidpunktojn de multnombraj sistemoj de drogmanio venas ĉefe de studoj pri lernado kaj memoro, kiuj utiligis kiel plifortikigajn substancojn ofte konsideratajn en la kunteksto de esplorado pri drogmanio, inkluzive de kokaino, alkoholo kaj amfetaminoj. Krome, lastatempaj evidentecoj sugestas, ke la memoro-sistemoj povas ankaŭ esti helpaj por kompreni aktualajn fontojn de toksomanio, kiuj reflektas emerĝantajn zorgojn pri sano, inkluzive de marijuuana uzo, alta grasa dieto, kaj videoludado.
Enkonduko
Esploristoj ofte rigardas mekanismojn de lernado kaj konduto por klarigi kiel homa psikopatologio estas akirita kaj esprimita. Ekzemplo de tia apliko estis donita de Norman M. White, kiu uzis faktojn de klasika lernada teorio kaj eksperimenta evidenteco subtenanta la ekziston de multnombraj memorosistemoj en la cerbo por provizi novan, influan aliron al drogmanio (1). Specife, White indikis, ke drogoj povas ludi la rolon de "plifortigiloj", kiuj, kiel manĝaĵo aŭ akvo en lernada tasko, plifortigas asociojn inter drog-rilataj stimuloj, kunteksto kaj konduto por antaŭenigi konsumadon de drogoj kaj, kun la tempo, toksomanio. White ankaŭ korpigis la aperantan hipotezon, ke ekzistas malsamaj specoj de memoro, kiuj estas mediaciitaj de disaj neŭralaj sistemoj. Laŭ ĉi tiu nova vidpunkto, drogoj povas rekte moduli multoblajn neŭrajn sistemojn, kaj ĉi tiuj neŭralaj sistemoj iras por kodigi apartajn erojn de la drog-memoro, kiuj, kiam esprimite, antaŭenigas plian konsumadon de drogoj.
La jaro 2016 markas la 20-an datrevenon de la vidpunkto de multnombraj memorsistemoj de drogmanio kiel priskribita de White. La nuna recenzo revizias ĉi tiun influan hipotezon, samtempe reliefigante iujn gravajn lastatempajn evoluojn, kiuj ne nur fundamentis la originan hipotezon, sed ankaŭ produktis pliajn vidojn pri kiel multnombraj memorosistemoj potenciale subtenas drogmanion.
La Multoblaj Memo-Sistemoj Vido de toksomanio
Konverĝaj evidentaĵoj de studoj dungantaj homojn kaj malsuperajn bestojn indikas, ke mamula memoro estas mediaciita de relative sendependaj neŭralaj sistemoj [por recenzoj, vidu Ref. (2-4)]. La fruaj eksperimentoj disigantaj multoblajn memorsistemojn estis ĉefe faritaj en la radia labirinto kaj indikis unikajn mnemonikajn funkciojn por la hipokampo, dorsstria stumo kaj amigdalo (5, 6). La hipokampo mediacias kognan / spacan formon de memoro, dum la dorsal striatum media de stimulo-respondo (S-R) kutima memoro. La amigdala mezigas pavlovianajn kaj stimulajn afektajn rilatojn (6, 7), samtempe konservante la modulan rolon de emocia ekscitiĝo sur aliaj specoj de memoro (8-12).
Kadre de la multnombraj sistemoj vido de memoro, Blanka (1) sugestis, ke la hipokampo, dorsstria stumo kaj amigdala kodu unikajn komponentojn de drogaj memoroj (vidu Figuron 1). La hipokampo kodas eksplicitan scion pri la rilato inter kvizoj kaj eventoj (t.e., asocioj de stimulo-stimulo) en la drogokontakto. Grave, la hipokampo ne kodas kondutajn respondojn, sed prefere la informoj akiritaj de la hipokampo povas esti uzataj por generi la taŭgajn kondutajn respondojn por ricevi drogajn plifortigojn. Aliflanke, la dorsa striatumo kodas asociojn inter drog-rilataj stimuloj kaj kondutaj respondoj. Ĉi tio eble permesos la prezenton de drog-rilata signo por aktivigi aŭtomatan kondutan respondon, kiu rezultigas drogon-prenon (ekz. Kuranta alproksimiĝo aŭ instrumenta levil-gazetaro). La amigdala kodas Pavlovian-asocian rilaton, tiel permesante al neŭtralaj aludoj en la drogkunteksto asociiĝi kun la medikamenta rekompenco. Bestoj poste reagas al ĉi tiuj kondiĉitaj indikoj simile al kiel ili origine reagis al la drogo. Specife, la kondiĉitaj resendoj aktivigas kondiĉitajn emociajn respondojn, inkluzive de internaj afekciaj statoj kaj kondiĉitan aliron al (aŭ en iuj kazoj evitado de) la kondiĉita kvino. Alia kritika ero de la hipotezo de White estas, ke drogoj povas moduli memorfunkcion de ĉiu el ĉi tiuj cerbaj regionoj. Tiel, la drogoj povas potenciale efiki sian propran memadministradon per pliiga solidigo de la drog-rilataj memoroj koditaj de la hipokampo, amigdala kaj dorsal-striatumo (vidu Figuron 1).
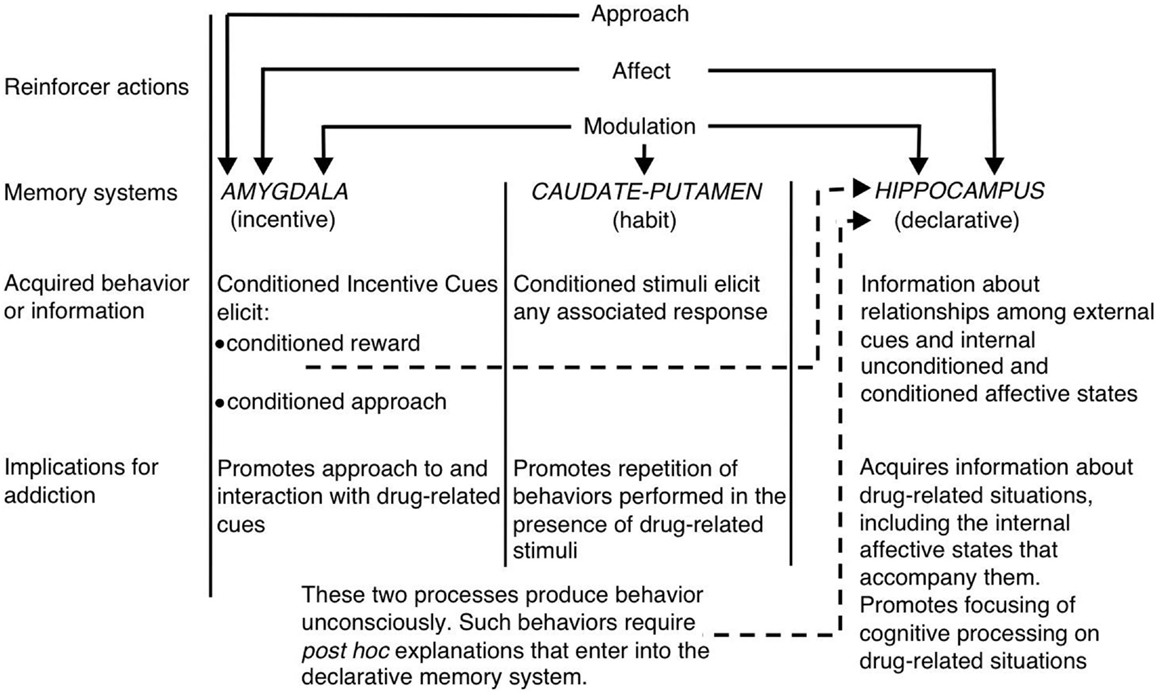
Figuro 1. White's (1) multnombraj memorosistemoj vido de drogmanio. Kiel naturaj plifortigiloj, kutimigaj drogoj posedas plurajn "plifortigajn agojn", inkluzive la kapablon alvoki pozitivan / negativan efikon, aliron kaj moduladon de memorsistemoj. La amigdalo, kaŭdata-putameno (t.e. dorsa striato) kaj hipokampo peras disigeblajn memorsistemojn, kaj ĉiu memorsistemo supozeble kodas unikajn erojn de drogrilataj memoroj. Konsiderante siajn memore modulajn ecojn, kutimigaj drogoj eble plibonigas sian propran memadministradon plibonigante la funkcion de ĉi tiuj sistemoj. (Represita de White kun permeso de John Wiley & Sons.)
Konsentite kun la multnombraj memorosistemoj de drogmanio, ampleksaj indicoj indikas kritikajn rolojn por la hipokampo, dorsstria stumo, kaj amigdala en drogmanio kaj relanĉo por diversaj misuzitaj substancoj [por revizio, vidu Ref. (13)]. La dorsa hipokampo ŝajnas havi rolon en la kunteksta kontrolo de drogoj serĉantaj kokainon (14-16). La flanka regiono de la dorsa striatumo (DLS) mediacias S – R kutiman levilon premantan kokainon kaj alkoholon (17, 18), kaj la bazolateral amigdala (BLA) mediacias kondiĉitan drogon serĉantan kokainon, alkoholon kaj heroinon (19-22). Ankaŭ konforme al la hipotezo de White, substancoj de misuzo povas moduli la mnemonikajn funkciojn de la hipokampo, dorsstria stumo kaj amigdalo (23-31).
Lastatempaj studoj kontribuis novajn amendojn al la multnombraj memorosistemoj al drogmanio. Ŝlosilaj ecoj de ĉi tiu nuntempa vidpunkto inkluzivas (1) neŭroatomikan movadon kun la tempo al DLS-dependa kutima memoro, (2) konkurencajn interagojn inter memosistemoj, (3) la rolo de streso kaj angoro en plibonigado de kutima drog-serĉado, kaj (4) la apliko de ĉi tiu hipotezo al novaj aperantaj fontoj de toksomanio.
La Neŭroatomatika Ŝanĝo de Sciiĝo al Kutimo
En eksperimentaj lernadaj situacioj, subjektoj tipe uzas celan konduton kiam komence solvas taskon. Tamen, post ampleksa trejnado, konduto iĝas aŭtonoma kaj povas esti plenumita kun malmulta atento, intenco aŭ kognitiva penado, konsistigante "kutimon" [por revizio, vidu Ref. (32)]. En fruaj pruvoj de ĉi tiu ŝanĝo de kognitiva kontrolo de konduto al kutimo, ronĝuloj estis trejnitaj per manĝaĵa rekompenco en duobla solva plus-labiriga tasko (33-35). En ĉi tiu tasko, ratoj estis liberigitaj de la sama starta pozicio (ekz. La suda brako) kaj devis fari konsekvencan korpan turnon ĉe la labirinta intersekciĝo por ricevi manĝaĵan rekompencon ĉiam lokitan en la sama celita brako (ekz., Ĉiam fari maldekstran. turnu vin por trovi manĝon en la okcidenta brako). Ratoj povus solvi ĉi tiun taskon aŭ lernante konsekvencan korpan turnan respondon aŭ farante ajnan respondon necesa por iri la saman spacan lokon. Por determini, kiu strategio uzas la ratojn, la enketistoj efektivigis sondan teston, en kiu bestoj estis liberigitaj de la kontraŭa starta brako (ekz. La norda brako). Se bestoj faris la kontraŭan korpon-turnon por iri la originalan celon, ili estis identigitaj kiel lokaj lernantoj. Se bestoj faris la saman korpan turnon kiel dum trejnado (t.e., irante al la brako kontraŭ la origina celloko), bestoj estis identigitaj kiel respondaj lernantoj. Evidenteco indikas, ke post iu trejnado, plej multaj bestoj montras lernadon, dum post ampleksa trejnado, bestoj ŝanĝas al kutima responda lernado (34-36). Interese, ĉi tiu translokiĝo de loka lernado al responda lernado povas reflekti neŭroatomikan movon. La komenca uzo de loka lernado en ĉi tiu tasko estas mediata de la hipokampo kaj dorsomedia striatumo [DMS (36, 37)], dum la uzo de responda lernado post plilongigita trejnado estas mediaciita de la DLS (36).
Krom fruaj manifestacioj uzantaj la plus-labirinton (34, 35), la kondutisma ŝanĝo al kutima memoro poste pruviĝis uzante operan levilon premante paradigmojn (38-42). En ĉi tiuj instrumentaj lernadaj taskoj, bestoj komence lerte premas por atingi la rezulton kaj ĉesos levi premi post kiam la manĝaĵa rezulto estos devalorigita. Tamen sekvi ampleksajn trejnajn bestojn ŝanĝiĝos al kutima respondado kaj daŭre premos la levilon eĉ post kiam la manĝaĵo malvaloriĝos (40). Kiel origine montrite en la plus-labirinto (36), la transiro de kogno al kutimo en instrumentaj lernadaj taskoj povus ankaŭ esti atribuita al neŭroatomatika movo. La komenca kognitiva kontrolo de konduto en ĉi tiuj instrumentaj lernadaj taskoj estas mediaciita de la hipokampo kaj DMS (43, 44), dum poste kutima respondado estas mediaciita de la DLS (18, 45, 46).
Multaj enketistoj sugestis, ke la neŭroatomia ŝanĝo al kutima memoro montrita en labirinto kaj instrumentaj lernadaj taskoj povus ankaŭ subteni la ŝanĝon de distra uzado de drogoj al deviga drogokutimo (13, 47-50). Konsentite kun ĉi tiu hipotezo, enketistoj pruvis diversajn misuzitajn substancojn, ke la DMS mediacias cel-responditan respondon por drog-plifortigo kaj la DLS mediacias kutiman respondadon por drog-plifortigo (18, 31, 51-53).
Konsiderante la altan fitraktan potencialon de iuj drogoj, enketistoj sugestis, ke toksomaniuloj povus plibonigi DLS-dependan kutiman memorfunkcion kaj tiel akceli la ŝanĝon de kognitiva al kutima kontrolo de konduto. Konsentite kun ĉi tiu hipotezo, ripetita ekspozicio al amfetamino aŭ kokaino faciligas la ŝanĝon de cel-direktita al kutima respondado pri manĝaĵ-plifortigo en instrumentaj levilaj premadaj taskoj (31, 54-59). Krome, lega premado por toksomaniaj substancoj (ekz. Alkoholo aŭ kokaino) kontraŭ manĝaĵa rekompenco estis asociita kun pli granda kutima respondado kontraŭ cel-direktita respondo (24, 60, 61). En homoj, alkohol-dependaj individuoj montras pli grandan kutiman respondadon en instrumenta lernada tasko, rilate al ne-dependaj kontrolaj individuoj (62). Ĉi tiu plibonigo de DLS-dependa kutima memoro per toksomaniuloj ankaŭ estis observita en taskoj de roncaj labirintoj. Kokaino, amfetamino, kaj alkoholekspozicio asociis kun plibonigita lernado en labirindaj taskoj de DLS-dependa aŭ pli granda uzo de respondaj strategioj de DLS-dependaj en duoblaj solvaj versioj de la labirinto (25, 63, 64). En homoj, la uzo de misuzitaj substancoj, inkluzive de alkoholo kaj tabako, rilatis al pli granda uzo de navigado-strategiaj dependaj dorsal-striatum en virtuala labirinto (65). Tiel, iuj drogoj misuzoj povus plibonigi DLS-dependan kutiman memoron, kaj ĉi tiu pli granda engaĝiĝo de la DLS-memorsistemo eble akcelos la transiron de distra uzado de drogoj al kutima droguzado. Ĉi tiu proponita mekanismo konformas al tiu de White (1) originala aserto, ke drogoj misuzoj eble foje faciligas sian propran memadministradon plibonigante funkcion de memorosistemoj.
Konkurenco Inter Memorsistemoj
Kvankam eblas, ke toksomaniaj drogoj plibonigas kutiman memoron rekte plibonigante funkcion de la DLS [ekz., Ref. (29)], alia ebleco estas, ke drogoj pri misuzo plibonigas kutiman memoron nerekte per modulado de aliaj memorosistemoj. Ĉi tiu alternativa me mechanismanismo alvokas la hipotezon, ke en iuj situacioj de lernado, memorosistemoj konkurencas por regado de lernado kaj ke malpligrandigante la funkcion de unu memora sistemo, funkcio de alia sendifekta sistemo povus esti plibonigita (11, 66). Notinde, la hipokampo kaj DLS foje povas konkurenci por regado de lernado, per kio lezo de la hipokampo plibonigas memorfunkcion de DLS (5, 6, 67, 68). Konkurencaj interagoj ankaŭ povas esti montritaj en duflankaj taskoj, kiam malhelpado de unu memorsistemo rezultigas la uzon de strategio mediata de alia sendifekta sistemo. Ekzemple, bestoj donitaj DMS-lezoj montras DLS-dependan kutiman respondon pri manĝaĵa rekompenco en instrumentaj lernadaj taskoj (44).
Konsiderante la konkurencivajn interagojn, kiuj foje ekestas inter memorosistemoj, unu ebleco estas, ke iuj drogoj misuzoj povus plibonigi nerekteblan DLS-kutiman memoron malhelpante kognajn memmekanismojn mediaciitajn de la DMS kaj hipokampo. Kiel menciite antaŭe, alkoholo estas asociita kun pli granda uzo de DLS-dependa kutima memoro en labirinto kaj operantaj leviloj premantaj paradigmojn (24, 61, 62, 64, 65). Evidenteco ankaŭ indikas, ke alkoholo malhelpas lernadon en hipokamp-dependaj spacaj memoraj taskoj [(64, 69-72); por revizio, vidu Ref. (73)], same kiel en DMS-dependaj inversaj lernadaj taskoj (74-77). Konsentite kun konkurenciva interagado inter memorosistemoj, oni hipotezis, ke alkoholo povas faciligi nerekteblan DLS-kutiman memoron per malhelpado de kognaj memormekanismoj78).
Oni devas rimarki, ke krom alkoholo, multnombraj drogoj estis asociitaj kun kognaj memoraj deficitoj. Eksponado al morfino, heroino, metamfetamino, MDMA (ekstazo), aŭ kronika kokaino simile produktas difektojn de spaca memoro depende de hipokampo tra diversaj taskoj (79-89). Estas tenta konjekti, ke kiel sugestite pri alkoholo, kognaj memor-mankoj produktitaj de toksomaniuloj povus nerekte plibonigi DLS-dependan kutiman memoron, kaj ke tio eble estas unu mekanismo, kiu rajtigas drogan memadministradon fariĝi kutima en homaj droguloj. Aliflanke, eblas ankaŭ, ke spacaj lernadaj deficitoj produktitaj de toksomaniuloj povus esti nerekte pere de plibonigo de procesoj de mem-dependa de DLS. Konsekvenca kun ĉi tiu hipotezo, stimula CREB-agado en la DLS malhelpas hipokamp-dependan spacan memoron (90), dum inhibicio de CREB-agado en la DLS renversas la damaĝojn de la spaca memoro produktitaj de morfino (91).
Rolo de Streso kaj Klopodo
Plia konsidero rilate al la multnombraj memorigaj sistemoj al drogmanio estas la rolo de streĉado. Konverĝa indico indikas, ke fortika emocia ekscitiĝo faciligas DLS-dependan kutiman memoron en ronĝuloj kaj homoj [por recenzoj, vidu Ref. (9-12)]. Administrado de anxiogenaj drogoj plibonigas lernadon de respondo dependanta de DLS en la akvo-labirinto (92-97). Ĉi tiu plibonigo de DLS-dependa kutima memoro ankaŭ estas observata post eksponiĝo al senkondiĉaj kondutaj streĉiloj [ekz., Kronika bremsado, vosta ŝoko, odora predanto, ktp. (98-101)] kaj ekspozicio al timkondiĉaj stimuloj [tono antaŭe kombina kun ŝoko (102, 103)]. Kvankam origine montrita en ronĝuloj (92), ĉi tiu plibonigo de kutima memoro induktita de fortika emocia ekscitiĝo pruviĝis vaste ankaŭ ĉe homoj (99, 104-110).
La mekanismoj permesantaj streĉon / maltrankvilon faciligi kutiman memoron restas plejparte nekonataj; tamen, evidenteco indikas kritikan modulan rolon de la BLA (93-95, 100). Konsentite kun konkurenciva interagado inter memorsistemoj, iuj evidentaĵoj ankaŭ sugestas, ke streĉo / maltrankvilo povus plibonigi nerekteblan DLS-kutiman memoron neebligante hipokampan funkcion (94, 95).
Plibonigo de kutima memoro post streĉiteco aŭ angoro povas rilati al kompreno de iuj elstaraj faktoroj kondukantaj al drogakuzo. Nome, streĉaj vivokazaĵoj aŭ kronikaj plilongigitaj periodoj de streso / angoro estas asociitaj kun pliigita vundebleco al drogmanio kaj reaperado en homoj (111-117), kaj similaj observaĵoj estis faritaj en bestaj modeloj de mem-administrado de drogoj [por revizio, vidu Ref. (118)]. Esploristoj sugestis, ke konformas al la influo de emocia eksuzo sur multnombraj sistemoj de memoro (10), akra aŭ kronika streso povas plibonigi drogmanion kaj reaperadon en homoj per okupado de DLS-dependaj kutimaj memoraj procezoj (9, 49, 119). Konsentite kun ĉi tiu sugesto, streĉado ĉe kokainaj dependaj individuoj estas asociita kun malpliigita sango-oksigeno-dependa (BOLD) aktiveco en la hipokampo kaj pliigita aktiveco en la dorsa striatumo, kaj ĉi tiuj BOLD-aktivecaj ŝanĝoj estas asociitaj kun streĉoj induktitaj de kokaino. (120).
Emerĝaj Fontoj de toksomanio
Krom la drogoj, la hipotezo pri multnombraj sistemoj ankaŭ estis uzata lastatempe por kompreni aliajn aperantajn fontojn de toksomanio. Ekzemple, la kresko de la obezeco dum la pasintaj jardekoj kaŭzis kompareblan kreskon de eksperimenta intereso, kun multaj enketistoj elparantaj paralelojn inter drogmanio kaj troigo [por revizio, vidu Ref. (121-123)]. Iuj lastatempaj evidentecoj sugestis, ke kiel drogmanio, manĝa toksomanio povus esti parte atribuita al pli alta engaĝiĝo de DLS-kutima memoro. Ĉe ratoj, binge-simila manĝaĵkonsumo faciligas la ŝanĝon de kognitiva al kutima kontrolo de konduto (124, 125). Plie, kutima konduto en enuigaj bestoj estas asociita kun pliigita DLS-agado kaj eblas malebligita blokante AMPA aŭ dopaminajn D1-receptorojn en la DLS (125). Lastatempe induktita obezeco ankaŭ estis asociita kun la uzo de kutima memoro en tasko Y-labirinto (126).
Alia emerĝanta konduta malordo kiu paralelas iujn funkciojn de drogmanio estas patologia videoludo aŭ ludado de videoludoj [por revizio, vidu Ref. (127)]. Kiel drogmanio, longdaŭra troa videoludado estis asociita kun reduktita dopamina D2-ricevilo liganta en la dorsal striatumo (128). Videoludado ankaŭ rilatas al pliigita aktivigo de la dorsal striatumo (129, 130), kaj pli grandaj dorsaj striaj volumoj antaŭdiras pli altajn nivelojn de videoludado (131). Homoj, kiuj regule ludas agajn videoludojn, pli emas uzi dorsan dependon de kutima memoro en virtuala labirinto (132), kaj antaŭ-trejnanta videoludado kondukas al kutima respondado super cel-direktita respondo en du-etaĝa decida tasko (133). Tiel, kiel proponite por drogoj de misuzo, ludi videoludojn povus plibonigi toksomanion per engaĝado de la DLS-dependa kutima memora sistemo.
Finfine, la alproksimiĝo de multnombraj sistemoj povus esti utila ankaŭ por kompreni mari mariuanan toksomanion. Kvankam marijuuano eble havas malpli altan potencialon de misuzo ol aliaj nelicaj substancoj klasike konsiderataj en la kunteksto de esplorado pri drogodependeco (ekz. Kokaino, morfino, heroino, ktp.), La peza uzado de kanabo povas tamen antaŭenigi simptomojn kaj forprenon de drogoj kiel observite kun aliaj drogoj. misuzo (134-137). Lastatempe estis sugestite, ke marijuuana toksomanio povus esti parte atribuita al pliigita engaĝiĝo de DLS-dependa kutima memoro (138). Dum akra kanabinoida ekspozicio malpliigas DLS-memoran funkcion (139, 140), ripetita kanabinoida ekspozicio kondukas al pli granda DLS-dependa kutima respondo en instrumenta lernada tasko (141). Krome, pezaj kanabo-uzantoj montras pli grandan aktivigon de la dorsal-striatumo, rilate al ne-uzantoj, dum plenumado de marijuuana versio de la implicita asocia tasko (142), kaj partoprenantoj kun historio de kanabo uzas pli probable uzi memoron de kutima striatum-dependa de kutimo en la virtuala labirinto (65).
Konsiderante la sukcesan aplikon de la memorosistemoj al emerĝantaj fontoj de toksomanio, estas racie hipotezi, ke multnombraj memorosistemoj eble ankaŭ implikiĝos en aliaj kondutismaj patologioj asociitaj kun toksomanio, kiel komputa aĉetado, interreta toksomanio kaj seksa toksomanio. Efektive, ĉu la memorsistemo povas esti utila por kompreni patologian videoludadon ankaŭ ricevis iom da atento (143, 144).
konkludo
Dudek jaroj da eksperimenta atestaĵo plejparte konfirmis tiun de White (1) multnombraj memsistemoj alproksimiĝas al drogmanio. La evidenteco indikas ke la hipokampo mediacias kuntekstan kontrolon de administrado de drogoj, la DLS mediacias S-R kutiman respondadon por drogplifortigo, kaj la amigdala mediacias kondiĉitan serĉadon de drogoj. Krome, pliaj esploroj kaŭzis pliajn komprenojn pri la vidpunkto de multnombraj memoroj pri drogmanio inkluzive de la ŝanĝo al kutima memoro, konkurenco inter memorosistemoj kaj rolo de streso kaj angoro.
Estonta esplorado devas provi integri la memorosistemojn kun aliaj teorioj pri toksomanio, kiel kontraŭaj motivaj procezoj (145). Ankaŭ utilus enigi en la memorsistemojn vidon de pliaj trajtoj de toksomanio, kiel ekzemple dependeco de drogoj, toleremo kaj retiriĝo. Kvankam la nuna recenzo ĉefe koncentriĝis sur la cerbaj regionoj origine konsiderataj de Blanko (t.e. la hipokampo, dorsstria stumo kaj amigdala), oni devas rimarki, ke pliaj cerbaj regionoj rilataj al lernado kaj memoro ankaŭ estis grave implikitaj en drogmanio kaj relokiĝo. , inkluzive de la media prefrontal-kortekso kaj kerno accumbens [por revizio, vidu Ref. (13)]. Fine, kvankam ekster la amplekso de la nuna recenzo, oni devas agnoski, ke ampleksaj evidentecoj sugestas, ke ĉelaj kaj molekulaj ŝanĝoj en la dubena dopaminergia sistemo ankaŭ kontribuas al toksomanio (146).
Kvankam kutimaj memoroj povas esti precipe malfacile kontroleblaj, iuj evidentaĵoj indikas, ke DLS-dependa memoro, iam akirita, en iuj cirkonstancoj povas esti subpremita (147) aŭ eĉ inversigis (148, 149). Tiel, eblas, ke la farmakologiaj manipuladoj kaj kondutaj procedoj kondukantaj al renversiĝo aŭ forigo de kutima memoro en bestaj modeloj de lernado povus esti adaptitaj por trakti drogmanion kaj recesiĝon en homoj.
Aŭtoro Kontribuoj
JG kaj MP kontribuis ideojn kaj redaktadon de la nuna mini-recenzo.
Konflikto pri Interesa Rakonto
La aŭtoroj deklaras, ke la esplorado estis farita sen manko de komercaj aŭ financaj rilatoj, kiujn oni povus konsideri kiel ebla konflikto de intereso.
Referencoj
1 Blanka NM. Toksomaniulaj drogoj kiel plifortigiloj: multnombraj partaj agoj sur memorosistemoj. toksomanio (1996) 91(7):921–50. doi: 10.1111/j.1360-0443.1996.tb03586.x
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
2 Blanka NM, McDonald RJ. Multoblaj paralelaj memorosistemoj en la cerbo de la rato. Neurobiol Lernu Mem (2002) 77(2):125–84. doi:10.1006/nlme.2001.4008
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
3 Squire LR. Memorsistemoj de la cerbo: mallonga historio kaj aktuala perspektivo. Neurobiol Lernu Mem (2004) 82(3):171–7. doi:10.1016/j.nlm.2004.06.005
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
4 Blanka NM, Packard MG, McDonald RJ. Disa sistemo de memoro: la rakonto disvolviĝas. Behav Neurosci (2013) 127(6):813–34. doi:10.1037/a0034859
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
5 Packard MG, Hirsh R, Blanka NM. Malsamaj efikoj de lezoj de fornikso kaj kaŭzaj kernoj sur du radiaj labirintaj taskoj: evidenteco por multnombraj sistemoj. J Neurosci (1989) 9(5): 1465-72.
6 McDonald RJ, Blanka NM. Triobla dispartigo de memorsistemoj: hipokampo, amigdala kaj dorsal striatumo. Behav Neurosci (1993) 107(1):3–22. doi:10.1037/0735-7044.107.1.3
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
7 Maren S. Neurobiologio de Pavloviana timkondiĉo. Annu Rev Neurosci (2001) 24(1):897–931. doi:10.1146/annurev.neuro.24.1.897
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
8 McGaugh JL. La amigdala modulas la firmiĝon de memoroj de emocie vekantaj spertoj. Annu Rev Neurosci (2004) 27: 1 – 28. doi: 10.1146 / annurev.neuro.27.070203.144157
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
9 Packard MG. Angoro, kogno, kaj kutimo: multnombra memsistemo perspektivo. Brain Res (2009) 1293: 121 – 8. doi: 10.1016 / j.brainres.2009.03.029
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
10 Packard MG, Goodman J. Emocia ekscito kaj multoblaj memosistemoj en la mamula cerbo. Front Behav Neurosci (2012) 6: 14. doi: 10.3389 / fnbeh.2012.00014
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
11 Packard MG, Goodman J. Faktoroj kiuj influas la relativan uzon de multoblaj memosistemoj. Hippopo (2013) 23(11):1044–52. doi:10.1002/hipo.22178
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
12 Schwabe L. Streso kaj engaĝiĝo de multnombraj memosistemoj: integriĝo de bestaj kaj homaj studoj. Hippopo (2013) 23(11):1035–43. doi:10.1002/hipo.22175
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
13 Everitt BJ, Robbins TW. Neŭralaj sistemoj de plifortigo por drogmanio: de agoj al kutimoj al devigo. Nat Neurosci (2005) 8(11):1481–9. doi:10.1038/nn1579
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
14 Fuchs RA, Evans KA, Ledford CC, Parker MP, Case JM, Mehta RH, et al. La rolo de la dorsomedial prefrontal-kortekso, basolateral amigdala kaj dorsala hipokampo en kunteksta restarigo de kokaino serĉanta ratojn. Neuropsychofarmacology (2005) 30(2):296–309. doi:10.1038/sj.npp.1300579
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
15 Fuchs RA, Eaddy JL, Su ZI, Bell GH. Interagoj de la bazolateral amigdalo kun la dorsokula hipokampo kaj dorsomedia prefrontal-kortekso reguligas restarigon de indukta kunteksto kun drogoj kun ratoj. Eur J Neurosci (2007) 26(2):487–98. doi:10.1111/j.1460-9568.2007.05674.x
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
16 Kramar CP, Barbano MF, Medina JH. Dopaminaj D1 / D5-riceviloj en la dorsa hipokampo estas bezonataj por akiro kaj esprimo de ununura proceso kun kokain-memoro. Neurobiol Lernu Mem (2014) 116: 172 – 80. doi: 10.1016 / j.nlm.2014.10.004
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
17 Ŝuo A, Minney VL, Shippenberg TS. Transiru de cel-direktita al kutima kokaino serĉanta plilongan sperton en ratoj. J Neurosci (2010) 30(46):15457–63. doi:10.1523/JNEUROSCI.4072-10.2010
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
18 Korbitan LH, Nie H, Janak PH. Normala serĉado de alkoholo: tempokurso kaj kontribuo de subregionoj de la dorsstriatumo. Biol-psikiatrio (2012) 72(5):389–95. doi:10.1016/j.biopsych.2012.02.024
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
19 Whitelaw RB, Markou A, Robbins TW, Everitt BJ. Ekscitotoksaj lezoj de la bazolateral amigdala malhelpas akiron de koka-serĉa konduto laŭ dua-ordiga plifortigo. Psikofarmacologio (1996) 127(1–2):213–24. doi:10.1007/BF02805996
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
20 Alderson HL, Robbins TW, Everitt BJ. La efikoj de ekscitotoksaj lezoj de la bazolateral amigdala sur la akiro de heroa serĉa konduto ĉe ratoj. Psikofarmacologio (2000) 153(1):111–9. doi:10.1007/s002130000527
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
21 Gabriele A, Vidu RE. Revena inaktivigo de la bazolateral amigdala, sed ne la dorsolateral caudate putamen, mildigas solidigon de la kokain-cue asocieca lernado en restariga modelo de serĉado de drogoj. Eur J Neurosci (2010) 32(6):1024–9. doi:10.1111/j.1460-9568.2010.07394.x
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
22 Sciascia JM, Reese RM, Janak PH, Chaudhri N. Alkohol-serĉado instigita per diskretaj Pavloviaj aludoj estas vigligita per alkoholaj kuntekstoj kaj mediaciita per glutamata signalado en la bazolateral amigdalo. Neuropsychofarmacology (2015) 40: 2801 – 12. doi: 10.1038 / npp.2015.130
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
23 Packard MG, Teather LA. Amygdala modulado de multnombraj memorosistemoj: hipokampo kaj kaŭdato-putamen. Neurobiol Lernu Mem (1998) 69(2):163–203. doi:10.1006/nlme.1997.3815
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
24 Dickinson A, Wood N, Smith JW. Alkoholo serĉanta ratojn: ago aŭ kutimo? QJ Exp Psychol B (2002) 55(4):331–48. doi:10.1080/0272499024400016
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
25 Udo T, Ugalde F, DiPietro N, Eichenbaum HB, Kantak KM. Efikoj de konstanta mem-administrado de kokaino sur amigdala-dependa kaj dorsal-striatum-lernado ĉe ratoj. Psikofarmacologio (2004) 174(2):237–45. doi:10.1007/s00213-003-1734-1
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
26 Ligno SC, Fay J, Sage JR, Anagnostaras SG. Kokaino kaj Pavloviana timkondiĉo: dozo-efika analizo. Behav Brain Res (2007) 176(2):244–50. doi:10.1016/j.bbr.2006.10.008
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
27 Ligno SC, Anagnostaras SG. Memoro kaj psikostimuliloj: modulado de Pavloviana timkondiĉo per amfetamino en C57BL / 6-musoj. Psikofarmacologio (2009) 202(1–3):197–206. doi:10.1007/s00213-008-1185-9
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
28 Iñiguez SD, Charntikov S, Baella SA, Herbert MS, Bolaños-Guzmán CA, Crawford CA. Post-trejnanta kokainekspozicio faciligas spacan memoriĝon en musoj C57BL / 6. Hippopo (2012) 22(4):802–13. doi:10.1002/hipo.20941
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
29 DePoy L, Daut R, Brigman JL, MacPherson K, Crowley N, Gunduz-Cinar O, et al. Kronika alkoholo produktas neŭroadaptiĝojn al ĉefa dorsala struktura lernado. Proc Natl Acad Sci Usono (2013) 110(36):14783–8. doi:10.1073/pnas.1308198110
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
30 Leri F, Nahas E, Henderson K, Limebeer CL, Parker LA, Blanka NM. Efikoj de post-trejnanta heroino kaj d-amfetamino sur solidigo de venk-restada lernado kaj timkondiĉo. J Psychopharmacol (2013) 27(3):292–301. doi:10.1177/0269881112472566
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
31 Schmitzer-Torbert N, Apostolidis S, Amoa R, O'Rear C, Kaster M, Stowers J, et al. Post-trejnado de kokainadministrado faciligas lernadon de kutimoj kaj postulas infralimbikan kortekson kaj dorsolateran striatumon. Neurobiol Lernu Mem (2015) 118: 105 – 12. doi: 10.1016 / j.nlm.2014.11.007
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
32 Knowlton BJ. Basaj ganglioj: kutima formado. En: Jaeger D, Jung R, redaktistoj. Enciklopedio de Komputila Neŭroscienco. Nov-Jorko: Springer (2014). p. 1 – 17.
33 Tolman EC, Ritchie BF, Kalish D. Studoj pri spaca lernado. IV. La translokigo de lernado de loko al aliaj startaj vojoj. J Exp Psychol (1947) 37(1):39–47. doi:10.1037/h0062061
34 Ritchie BF, Aeschliman B, Pierce P. Studoj pri spaca lernado. VIII. Loko-rendimento kaj akiro de lokaj dispozicioj. J Comp Physiol Psychol (1950) 43(2):73–85. doi:10.1037/h0055224
35 Hicks LH. Efikoj de ŝancelado pri akiro kaj renversado de lernado de respondoj kaj lokoj. Psychol-Rep (1964) 15(2):459–62. doi:10.2466/pr0.1964.15.2.459
36 Packard MG, McGaugh JL. Senaktivigo de hipokampo aŭ kaŭda kerno kun lidocaine malsame efikas sur lernado de responda loko kaj respondo. Neurobiol Lernu Mem (1996) 65(1):65–72. doi:10.1006/nlme.1996.0007
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
37 Yin HH, Knowlton BJ. Kontribuoj de striktaj subregionoj por lerni lokon kaj respondon. Lernu Mem (2004) 11(4):459–63. doi:10.1101/lm.81004
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
38 Adams KD, Dickinson A. Instrumenta respondado sekve de plifortiga malvalorigo. QJ Flua Psikolo (1981) 33B: 109 – 12. doi: 10.1080 / 14640748108400816
39 Adams KD, Dickinson A. Agoj kaj kutimoj: variaĵoj en asociaj reprezentadoj dum instrumenta lernado. En: Spear NE, Miller RR, redaktistoj. Informo-Prilaborado en Bestoj: Memormekanismoj. Hillsdale, NJ: Erlbaum (1981). p. 143 – 65.
40 Adams KD. Varioj en la sentiveco de instrumentaj respondoj al plifortiga malvalorigo. QJ Flua Psikolo (1982) 34B: 77 – 98. doi: 10.1080 / 14640748208400878
41 Dickinson A, Nikolao DJ. Nerevigebla stimulila lernado dum instrumentaj kondiĉoj: la rolo de la rilatoj de stirado-plifortigilo kaj respondo-plifortigilo. QJ Flua Psikolo (1983) 35B: 249 – 63. doi: 10.1080 / 14640748308400909
42 Dickinson A, Nikolao DJ, Adams KD. La efikoj de la instrumenta kontento sur susceptibilidad al plifortiga malvalorigo. QJ Flua Psikolo (1983) 35B: 35 – 51. doi: 10.1080 / 14640748308400912
43 Korbitan LH, Balleine BW. La rolo de la hipokampo en instrumenta kondiĉado. J Neurosci (2000) 20(11): 4233-9.
44 Yin HH, Ostlund SB, Knowlton BJ, Balleine BW. La rolo de la dorsomedia striato en instrumenta kondiĉado. Eur J Neurosci (2005) 22:513–23. doi:10.1111/j.1460-9568.2005.04218.x
45 Yin HH, Knowlton BJ, Balleine BW. Lezoj de dorsolateral striatum konservas rezultan atendon sed malhelpas kutiman formadon en instrumenta lernado. Eur J Neurosci (2004) 19:181–9. doi:10.1111/j.1460-9568.2004.03095.x
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
46 Quinn JJ, Pittenger C, Lee AS, Pierson JL, Taylor JR. Striatum-dependaj kutimoj estas sensencaj al ambaŭ pliigoj kaj malkreskoj en plifortiga valoro en musoj. Eur J Neurosci (2013) 37: 1012 – 21. doi: 10.1111 / ejn.12106
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
47 Yin HH. De agoj al kutimoj: neŭroadaptoj kondukantaj al dependeco. Sano pri Alkoholo (2008) 31(4): 340-4.
48 Belin D, Jonkman S, Dickinson A, Robbins TW, Everitt BJ. Paralelaj kaj interagaj lernadaj procezoj en la bazaj ganglioj: graveco por kompreno de toksomanio. Behav Brain Res (2009) 199(1):89–102. doi:10.1016/j.bbr.2008.09.027
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
49 Schwabe L, Dickinson A, Lupo OT. Streso, kutimoj kaj drogmanio: psikoneŭro-endokrinologia perspektivo. Klarko Clin Psychopharmacol (2011) 19(1):53–63. doi:10.1037/a0022212
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
50 Hogarth L, Balleine BW, Corbit LH, Killcross S. Asociaj lernmekanismoj, kiuj subtenas la transiron de distra drogokutimo al toksomanio. Ann NY Akademio Sci (2013) 1282(1):12–24. doi:10.1111/j.1749-6632.2012.06768.x
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
51 Murray JE, Belin D, Everitt BJ. Duobla disiĝo de la dorsomedia kaj dorsolatera striatokontrolo pri akiro kaj agado de kokain-serĉado. Neuropsychofarmacology (2012) 37(11):2456–66. doi:10.1038/npp.2012.104
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
52 Clemens KJ, Castino MR, Cornish JL, Goodchild AK, Holmes NM. Kondutaj kaj neŭralaj substratoj de kutima formado en ratoj intravene administrante nikotinon. Neuropsychofarmacology (2014) 39: 2584 – 93. doi: 10.1038 / npp.2014.111
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
53 Korbitan LH, Nie H, Janak PH. Kutima respondado pri alkoholo dependas de AMPA kaj D2-ricevilo signalanta en la dorsolatera striatumo. Front Behav Neurosci (2014) 8: 301. doi: 10.3389 / fnbeh.2014.00301
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
54 Schoenbaum G, Setlow B. Kokaino faras agojn sensencaj al rezultoj sed ne estingiĝo: implicoj por ŝanĝita orbitofrontal-amigdala funkcio. Kortekso Cereb (2005) 15(8):1162–9. doi:10.1093/cercor/bhh216
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
55 Nelson A, Killcross S. Amfetaminekspozicio plibonigas kutiman formadon. J Neurosci (2006) 26(14):3805–12. doi:10.1523/JNEUROSCI.4305-05.2006
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
56 Nordquist RE, Voorn P, De Mooij-van Malsen JG, Joosten RNJMA, Pennartz CMA, Vanderschuren LJMJ. Pliigita valoro de plifortigilo kaj akcelita kutima formado post ripetita amfetamina traktado. Eur Neuropsychopharmacol (2007) 17(8):532–40. doi:10.1016/j.euroneuro.2006.12.005
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
57 LeBlanc KH, Maidment NT, Ostlund SB. Repetita kokainekspozicio faciligas la esprimon de instiga instigo kaj induktas kutiman kontrolon ĉe ratoj. PLOJ Unu (2013) 8: e61355. doi: 10.1371 / journal.pone.0061355
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
58 Nelson AJ, Killcross S. Akcelita kutima formado post amfetamina ekspozicio estas inversigita de D1, sed plibonigita de D2, receptaj antagonistoj. Fronto Neŭroscio (2013) 7: 76. doi: 10.3389 / fnins.2013.00076
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
59 Corbit LH, Chieng BC, Balleine BW. Efikoj de ripetita kokainekspozicio sur kutimlernado kaj renversado de N-acetilcisteino. Neuropsychofarmacology (2014) 39(8):1893–901. doi:10.1038/npp.2014.37
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
60 Miles FJ, Everitt BJ, Dickinson A. Parola kokaino serĉata de ratoj: ago aŭ kutimo? Behav Neurosci (2003) 117(5):927–38. doi:10.1037/0735-7044.117.5.927
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
61 Mangieri RA, Cofresí RU, Gonzales RA. Etanolo serĉanta ratojn Long Evans ne ĉiam estas cel-direktita. PLOJ Unu (2012) 7: e42886. doi: 10.1371 / journal.pone.0042886
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
62 Sjoerds Z, De Wit S, Van Den Brink W, Robbins TW, Beekman ATF, Penninx BWJH, et al. Kondutaj kaj neŭroimaj evidentecoj por troa dependeco de kutimaj lernado en alkohol-dependaj pacientoj. Transl-psikiatrio (2013) 3(12): e337. doi: 10.1038 / tp.2013.107
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
63 Packard MG, McGaugh JL. Kvinpirolo kaj d-amfetamina administrado posttraktanta plibonigas memoron pri spacaj kaj karesaj diskriminacioj en akvodislimo. Psikobiologio (1994) 22(1): 54-60.
64 Matthews DB, Ilgen M, Blanka AM, Plej Bona PJ. Akra administrado de etanolo malpliigas spacan agadon samtempe faciligante nepacian agadon en ratoj. Neurobiol Lernu Mem (1999) 72(3):169–79. doi:10.1006/nlme.1998.3900
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
65 Bohbot VD, Balso D, Conrad K, Konishi K, Leyton M. Caudate-dependaj kernaj navigadaj strategioj estas asociitaj kun pliigita uzo de toksomaniuloj. Hippopo (2013) 23(11):973–84. doi:10.1002/hipo.22187
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
66 Poldrack RA, Packard MG. Konkurenco inter multnombraj memorosistemoj: konverĝa indico de bestaj kaj homaj cerbaj studoj. Neuropsychologia (2003) 41(3):245–51. doi:10.1016/S0028-3932(02)00157-4
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
67 Matthews DB, Plej Bona PJ. Fimbria / fornikaj lezoj faciligas la lernadon de nepacienca responda tasko. Psychon Bull Rev (1995) 2(1):113–6. doi:10.3758/BF03214415
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
68 Schroeder JP, Wingard JC, Packard MG. Post-trejnanta revertebla inaktivigo de hipokampo malkaŝas interferon inter memorsistemoj. Hippopo (2002) 12(2):280–4. doi:10.1002/hipo.10024
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
69 Matthews DB, Simson PE, Plej bona PJ. Akra etanolo malbonigas spacan memoron sed ne stimulan / respondan memoron en la rato. Alkoholo Clin Exp Res (1995) 19(4):902–9. doi:10.1111/j.1530-0277.1995.tb00965.x
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
70 Blanka AM, Elek TM, Beltz TL, Plej Bona PJ. Spaciala agado estas pli sentema al etanolo ol nepaca agado sendepende de kvieta proksimeco. Alkoholo Clin Exp Res (1998) 22(9):2102–7. doi:10.1111/j.1530-0277.1998.tb05922.x
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
71 Matthews DB, Morrow AL, Tokunaga S, McDaniel JR. Akra administrado de etanolo kaj akra administrado de allopregnanolono malhelpas spacan memoron en la akva tasko de Morris. Alkoholo Clin Exp Res (2002) 26(11):1747–51. doi:10.1111/j.1530-0277.2002.tb02479.x
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
72 Berry RB, Matthews DB. Akra administrado de etanolo selektive difektas spacan memoron en C57BL / 6J-musoj. alkoholo (2004) 32(1):9–18. doi:10.1016/j.alcohol.2003.09.005
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
73 Silvers JM, Tokunaga S, Berry RB, White AM, Matthews DB. Mankoj en spaca lernado kaj memoro: etanolo, alopregnanolono, kaj hipokampo. Brain Res Rev (2003) 43(3):275–84. doi:10.1016/j.brainresrev.2003.09.002
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
74 Badaniĉ KA, Becker HC, Woodward JJ. Efikoj de kronika intermita etanolo-ekspozicio sur orbitofrontaj kaj mezaj prefrontal-kortikaj dependaj kondutoj en musoj. Behav Neurosci (2011) 125(6):879–91. doi:10.1037/a0025922
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
75 Coleman LG Jr, He J, Lee J, Styner M, Crews FT. Adoleskanta trinka trinkaĵo ŝanĝas plenkreskan cerban neŭrotransmisilon genan esprimon, konduton, cerbajn regionajn volumojn kaj neŭkemion en musoj. Alkoholo Clin Exp Res (2011) 35(4):671–88. doi:10.1111/j.1530-0277.2010.01385.x
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
76 Kuzmin A, Liljequist S, Meis J, Chefer V, Shippenberg T, Bakalkin G. Ripetitaj moderaj dozonaj etanolaj atakoj malhelpas kognan funkcion en ratoj de Wistar. Addict Biol (2012) 17(1):132–40. doi:10.1111/j.1369-1600.2010.00224.x
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
77 Coleman LG, Liu W, Oguz I, Styner M, Crews FT. Adoleska binge-etanol-kuracado ŝanĝas plenkreskajn cerbajn regionajn volumojn, kortikajn eksterĉelajn matricajn proteinojn kaj kondutan flekseblecon. Pharmacol Biochem Behav (2014) 116: 142 – 51. doi: 10.1016 / j.pbb.2013.11.021
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
78 Matthews DB, Silvers JR. La uzo de akra administrado de etanolo kiel ilo por esplori multoblajn memorsistemojn. Neurobiol Lernu Mem (2004) 82(3):299–308. doi:10.1016/j.nlm.2004.06.007
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
79 Broening HW, Morford LL, Inman-Wood SL, Fukumura M, Vorhees CV. Lernado kaj memoraj mankoj de 3, 4-metilenedioksimetamfetamino (ekstazo) dependas de la eksponiĝo dum frua evoluo. J Neurosci (2001) 21(9): 3228-35.
80 Williams MT, Morford LL, Wood SL, Wallace TL, Fukumura M, Broening HW, et al. Disvolva d-metamfetamina traktado selektive induktas spacajn navigadajn mankojn en referenca memoro en la akvobaraĵo de Morris dum ŝparado de labora memoro. Synapse (2003) 48(3):138–48. doi:10.1002/syn.10159
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
81 Vorhees CV, Reed TM, Skelton MR, Williams MT. Ekspono al 3, 4-methylenedioxymethamphetamine (MDMA) dum posttagaj tagoj 11-20 indikas referencon sed ne laborantan memoran mankon en la akvodislimo Morris ĉe ratoj: implicoj de antaŭa lernado. Int J Dev Neurosci (2004) 22(5):247–59. doi:10.1016/j.ijdevneu.2004.06.003
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
82 Cohen MA, Skelton MR, Schaefer TL, Gudelsky GA, Vorhees CV, Williams MT. Lernado kaj memoro post novnaska ekspozicio al 3, 4-metilenedioxymethamphetamine (ekstazo) ĉe ratoj: interago kun ekspozicio en plenaĝeco. Synapse (2005) 57(3):148–59. doi:10.1002/syn.20166
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
83 Skelton MR, Williams MT, Vorhees CV. Traktado kun MDMA de P11-20 malhelpas spacan lernadon kaj vojan integriĝon en adoleskaj ratoj, sed nur spaca lernado en pli maljunaj ratoj. Psikofarmacologio (2006) 189(3):307–18. doi:10.1007/s00213-006-0563-4
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
84 Ma MX, Chen YM, He J, Zeng T, Wang JH. Efikoj de morfino kaj ĝia retiro sur Y-labirinto-spaca rekono-memoro en musoj. Neurokienco (2007) 147(4):1059–65. doi:10.1016/j.neuroscience.2007.05.020
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
85 Belcher AM, Feinstein EM, O'Dell SJ, Marshall JF. Metamfetamino influas rekonan memoron: komparo de skalaj kaj unutagaj dozaj reĝimoj. Neuropsychofarmacology (2008) 33(6):1453–63. doi:10.1038/sj.npp.1301510
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
86 Tramullas M, Martínez-Cué C, Hurlé MA. Kronika administrado de heroino al musoj produktas ĝisreguligon de cerbaj apoptozaj rilataj proteinoj kaj malhelpas spacan lernadon kaj memoron. Neuropharmacology (2008) 54(4):640–52. doi:10.1016/j.neuropharm.2007.11.018
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
87 Norda A, Swant J, Salvatore MF, Gamble-George J, Prins P, Butler B, et al. Kronika metamfetamena ekspozicio produktas malfruan, longdaŭran memor-mankon. Synapse (2013) 67(5):245–57. doi:10.1002/syn.21635
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
88 Fole A, Martin M, Morales L, Del Olmo N. Efektoj de kronika kuracado kun kokaino dum adoleskeco en ratoj Lewis kaj Fischer-344: nova malkapablo de rekono de loko kaj ŝanĝoj en sinaptika plastikeco en plenaĝeco. Neurobiol Lernu Mem (2015) 123: 179 – 86. doi: 10.1016 / j.nlm.2015.06.001
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
89 Zhou M, Luo P, Lu Y, Li CJ, Wang DS, Lu Q, et al. Malekvilibro de HCN1 kaj HCN2-esprimo en hipokampa areo CA1 malhelpas spacan lernadon kaj memoron en ratoj kun kronika morfina ekspozicio. Prog Neuropsychopharmacol Biol-psikiatrio (2015) 56: 207 – 14. doi: 10.1016 / j.pnpbp.2014.09.010
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
90 Kathirvelu B, Colombo PJ. Efikoj de lentivirus-mediata CREB-esprimo en la dorsolatera striato: memoro-plibonigo kaj evidenteco por konkurencivaj kaj kunlaboraj interagoj kun la hipokampo. Hippopo (2013) 23(11):1066–74. doi:10.1002/hipo.22188
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
91 Baudonnat M, Guillou JL, Husson M, Vandesquille M, Corio M, Decorte L, et al. Malhelpanta efiko de drog-induktita rekompenco sur spaca sed ne gvidita lernado: impliko de striatal protein kinase A / cAMP-responda element-liganta proteina vojo. J Neurosci (2011) 31:16517–28. doi:10.1523/JNEUROSCI.1787-11.2011
92 Packard MG, Wingard JC. Amygdala kaj "emocia" modulado de la relativa uzo de multnombraj memorosistemoj. Neurobiol Lernu Mem (2004) 82(3):243–52. doi:10.1016/j.nlm.2004.06.008
93 Elliott AE, Packard MG. Intra-amigdala anxiogena drogo-infuzaĵo antaŭ reakiro de fleksiaj ratoj al la uzo de kutima memoro. Neurobiol Lernu Mem (2008) 90(4):616–23. doi:10.1016/j.nlm.2008.06.012
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
94 Wingard JC, Packard MG. La amigdala kaj kortuŝa modulado de konkurenco inter kognitiva kaj kutima memoro. Behav Brain Res (2008) 193(1):126–31. doi:10.1016/j.bbr.2008.05.002
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
95 Packard MG, Gabriele A. Periferiaj anserogenaj drogaj injektoj malsame efikas sur kognitiva kaj kutima memoro: rolo de basolateral amigdalo. Neurokienco (2009) 164(2):457–62. doi:10.1016/j.neuroscience.2009.07.054
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
96 Leong KC, Goodman J, Packard MG. Buspirone blokas la plibonigan efikon de la anxiogena drogo RS 79948-197 sur solidigo de kutima memoro. Behav Brain Res (2012) 234(2):299–302. doi:10.1016/j.bbr.2012.07.009
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
97 Goodman J, Leong KC, Packard MG. Glucocorticoid-plibonigo de dorsolateral-striatum-dependa kutima memoro postulas samtempe noradrenergian aktivecon. Neurokienco (2015) 311: 1 – 8. doi: 10.1016 / j.neuroscience.2015.10.014
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
98 Kim JJ, Lee HJ, Han JS, Packard MG. Amygdala estas kritika por streĉ-induktita modulado de hipokampa longperspektiva potencado kaj lernado. J Neurosci (2001) 21(14): 5222-8.
99 Schwabe L, Dalm S, Schächinger H, Oitzl MS. Kronika streĉado modulas la uzon de spacaj kaj stimulo-respondaj lernadaj strategioj en musoj kaj viro. Neurobiol Lernu Mem (2008) 90(3):495–503. doi:10.1016/j.nlm.2008.07.015
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
100 Leong KC, Packard MG. Eksponiĝo al predanto-odoro influas relativan uzon de multoblaj memosistemoj: rolo de basolateral amigdalo. Neurobiol Lernu Mem (2014) 109: 56 – 61. doi: 10.1016 / j.nlm.2013.11.015
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
101 Taylor SB, Anglin JM, Paode PR, Riggert AG, Olive MF, Conrad KD. Kronika streso povas faciligi la varbadon de kutimaj kaj toksodependaj neŭrocirkvitoj tra neurona restrukturado de la striatumado. Neurokienco (2014) 280: 231 – 42. doi: 10.1016 / j.neuroscience.2014.09.029
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
102 Leong KC, Goodman J, Packard MG. Post-trejnada reekspozicio al timaj kondiĉigitaj stimuloj plibonigas memor-firmiĝon kaj fleksas ratojn direkte al uzado de dorsolatera striatum-respondo-lernado. Behav Brain Res (2015) 291: 195 – 200. doi: 10.1016 / j.bbr.2015.05.022
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
103 Goode TE, Leong KC, Goodman J, Maren S, Packard MG. Plibonigo de striat-dependa memoro per kondiĉita timo estas mediaciita de beta-adrenergiaj riceviloj en la bazolateral amigdalo. Neurobiologio de Streso (en gazetaro). doi: 10.1016 / j.ynstr.2016.02.004
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
104 Schwabe L, Oitzl MS, Philippsen C, Richter S, Bohringer A, Wippich W, et al. Streso modulas la uzon de spacaj kontraŭ stimulo-respondaj lernadaj strategioj en homoj. Lernu Mem (2007) 14(1–2):109–16. doi:10.1101/lm.435807
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
105 Schwabe L, Schächinger H, de Kloet ER, Oitzl MS. Kortikosteroidoj funkcias kiel ŝaltilo inter memosistemoj. J Cogn Neurosci (2010) 22(7):1362–72. doi:10.1162/jocn.2009.21278
106 Schwabe L, Tegenthoff M, Höffken O, Wolf OT. Samtempa glukokortikoida kaj noradrenergia agado movas instrumentan konduton de cel-direktita al kutima kontrolo. J Neurosci (2010) 30(24):8190–6. doi:10.1523/JNEUROSCI.0734-10.2010
107 Schwabe L, Tegenthoff M, Höffken O, Wolf OT. Mineralocorticoid-riceva blokado malhelpas moduladon induktitan de streĉoj de multoblaj memorosistemoj en la homa cerbo. Biol-psikiatrio (2013) 74(11):801–8. doi:10.1016/j.biopsych.2013.06.001
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
108 Schwabe L, Lupo OT. Streĉiteco instigas kutiman konduton en homoj. J Neurosci (2009) 29(22):7191–8. doi:10.1523/JNEUROSCI.0979-09.2009
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
109 Schwabe L, Lupo OT. Socie taksita malvarma premanta streĉadon post instrumenta lernado favoras kutimojn super cel-direktita ago. Psikoneuroendokrinologio (2010) 35(7):977–86. doi:10.1016/j.psyneuen.2009.12.010
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
110 Guenzel FM, Wolf OT, Schwabe L. Glucocorticoidoj akcelas stimulan-respondan memoroformadon en homoj. Psikoneuroendokrinologio (2014) 45: 21 – 30. doi: 10.1016 / j.psyneuen.2014.02.015
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
111 Higgins RL, Marlatt GA. Timo de interpersona takso kiel determinanto de konsumado de alkoholo en viraj sociaj drinkantoj. J Abnorm Psychol (1975) 84(6):644–51. doi:10.1037/0021-843X.84.6.644
112 Marlatt GA, Gordon JR. Determinantoj de relanĉo: implicoj por konservado de konduta ŝanĝo. En: Davidson PO, Davidson SM, redaktistoj. Konduta Medicino: Ŝanĝantaj Vivstiloj. Novjorko: Brunne / Mazel (1980). p. 410 – 52.
113 Novulo MD, Bentler PM. Efiko de uzado de adoleskaj drogoj kaj socia subteno sur problemoj de junaj plenkreskuloj: longforma studo. J Abnorm Psychol (1988) 97:64–75. doi:10.1037/0021-843X.97.1.64
114 Wallace a.K. Psikologiaj kaj mediaj determinantoj de reaperado en fendaj kokainaj fumantoj. J Substrema Misuzo Traktado (1989) 6(2):95–106. doi:10.1016/0740-5472(89)90047-0
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
115 Kaplan HB, Johnson RJ. Rilatoj inter cirkonstancoj ĉirkaŭantaj komencan nelican droguzadon kaj eskaladon de droguzo: moderigi efikojn de sekso kaj fruaj adoleskaj spertoj. En: Glantz M, Pickens R, redaktistoj. Vundebleco al Druguzo. Washington, Dc: Usona Psikologia Asocio (1992). p. 200 – 358.
116 Harrison PA, Fulkerson JA, Beebe TJ. Multobla substanco-uzo de viktimoj inter adoleskaj fizikaj kaj seksaj misuzoj. Infana misuzo (1997) 21:529–39. doi:10.1016/S0145-2134(97)00013-6
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
117 Chilcoat HD, Breslau N. Posttraŭmata streĉa malordo kaj drogaj malordoj: testado de kaŭzaj vojoj. Arch Gen Psikiatrio (1998) 55(10):913–7. doi:10.1001/archpsyc.55.10.913
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
118 Piazza PV, Le Moal M. La rolo de streso en drogadministrado. Tendencoj Pharmacol Sci (1998) 19(2):67–74. doi:10.1016/S0165-6147(97)01115-2
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
119 Goodman J, Leong KC, Packard MG. Emocia modulado de multnombraj memorosistemoj: implikaĵoj por la neurobiologio de post-traŭmata stresa malordo. Rev Neŭrosci (2012) 23(5–6):627–43. doi:10.1515/revneuro-2012-0049
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
120 Sinha R, Lacadie C, Skudlarski P, Fulbright RK, Rounsaville BJ, Kosten TR, et al. Neŭra aktiveco asociita kun avido de kokaino induktita de streso: funkcia magneta resona studo. Psikofarmacologio (2005) 183(2):171–80. doi:10.1007/s00213-005-0147-8
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
121. Avena NM, Rada P, Hoebel BG. Evidenteco pri sukero: kondutoj kaj neurochemiaj efikoj de intermita, troa sukero. Neurosci Biobehav Rev (2008) 32(1):20–39. doi:10.1016/j.neubiorev.2007.04.019
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
122 Gearhardt AN, Corbin WR, Brownell KD. Manĝaĵo toksomanio: ekzameno de la diagnozaj kriterioj por dependeco. J Addict Med (2009) 3(1):1–7. doi:10.1097/ADM.0b013e318193c993
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
123 Smith DG, Robbins TW. La neŭrobiologiaj bazoj de obezeco kaj bengala manĝo: bazo por adopti la manĝaĵan toksomanion. Biol-psikiatrio (2013) 73(9):804–10. doi:10.1016/j.biopsych.2012.08.026
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
124 de Jong JW, Meijboom KE, Vanderschuren LJ, Adan RA. Malalta kontrolo de plaĉa manĝaĵa konsumado ĉe ratoj estas asociita kun kutima konduto kaj reaperanta vundebleco: individuaj diferencoj. PLOJ Unu (2013) 8(9): e74645. doi: 10.1371 / journal.pone.0074645
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
125 Furlong TM, Jayaweera HK, Balleine BW, Corbit LH. Senforta konsumo de plaĉa manĝaĵo akcelas kutiman kontrolon de konduto kaj dependas de aktivigo de la dorsolatera striato. J Neurosci (2014) 34(14):5012–22. doi:10.1523/JNEUROSCI.3707-13.2014
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
126 Hargrave SL, Davidson TL, Zheng W, Kinzig KP. Okcidentaj dietoj induktas fekon de sango-cerbo kaj ŝanĝas spacajn strategiojn en ratoj. Behav Neurosci (2016) 130(1):123–35. doi:10.1037/bne0000110
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
127 Smith KL, Hummer TA, Hulvershorn LA. Patologia videoludado kaj ĝia rilato al malsanoj pri substanco-uzo. Curr Addict Rep (2015) 2(4):302–9. doi:10.1007/s40429-015-0075-6
128 Weinstein AM. Komputilo kaj videoludludado - komparo inter ludantoj kaj ne-ludaj uzantoj. Am J Drug Alkohola Malsano (2010) 36(5):268–76. doi:10.3109/00952990.2010.491879
129 Kätsyri J, Hari R, Ravaja N, Nummenmaa L. La oponanto gravas: respondecaj altaj rekompencoj de FMRI al gajnado kontraŭ homo kontraŭ komputila kontraŭulo dum interaga videoludo. Kortekso Cereb (2013) 23(12):2829–39. doi:10.1093/cercor/bhs259
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
130 Kätsyri J, Hari R, Ravaja N, Nummenmaa L. Nur spekti la ludon ne sufiĉas: striatal fMRI-rekompenco respondas al sukcesoj kaj fiaskoj en videoludo dum aktiva kaj malkaŝa ludado. Front Hum Neurosci (2013) 7: 278. doi: 10.3389 / fnhum.2013.00278
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
131 Erickson KI, Boot WR, Basak C, Neider MB, Prakash RS, Voss MW, et al. Striatala volumo antaŭdiras nivelon de akira kapablo de videoludoj. Kortekso Cereb (2010) 20: 2522 – 30. doi: 10.1093 / cercor / bhp293
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
132 West GL, Drisdelle BL, Konishi K, Jackson J, Jolicoeur P, Bohbot VD. Kutima ago-videoludado estas asociita kun kaudaj kern-dependaj navigaj strategioj. Proc R Soc B (2015) 282(1808) doi: 10.1098 / rspb.2014.2952
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
133 Liu S, Schad DJ, Kuschpel MS, Rapp MA, Heinz A. Muziko kaj videoludado dum paŭzoj: influo de kutima kontraŭ cel-direktita decido. Papero Prezentita en 45th Ĉiujara Kunveno de la Socio por Neŭroscienco. Ĉikago, IL: Socio por Neŭroscienco (2015).
134 de Fonseca FR, Carrera MRA, Navarro M, Koob GF, Weiss F. Aktivigo de liberiganta faktoro de kortikotropino en la limfika sistemo dum kanabinoida retiriĝo. scienco (1997) 276(5321):2050–4. doi:10.1126/science.276.5321.2050
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
135 Cornelius JR, Chung T, Martin C, Wood DS, Clark DB. Cannabis-retiriĝo estas ofta inter kuraĝ-serĉantaj adoleskantoj kun cannabis-dependeco kaj grava depresio, kaj estas asociita kun rapida relanĉo al dependeco. Addict Behav (2008) 33(11):1500–5. doi:10.1016/j.addbeh.2008.02.001
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
136 Greene MC, Kelly JF. La prevalenco de kanabo-retiriĝo kaj ĝia influo sur adoleskaj kuracaj respondoj kaj rezultoj: 12-monata esplorado. J Addict Med (2014) 8: 359 – 67. doi: 10.1097 / ADM.0000000000000064
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
137 Wagner FA, Anthony JC. De la unua konsumo de drogoj ĝis dependeco de drogoj; evoluaj periodoj de risko por dependeco de marijuuano, kokaino kaj alkoholo. Neuropsychofarmacology (2002) 26:479–88. doi:10.1016/S0893-133X(01)00367-0
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
138 Goodman J, Packard MG. La influo de kanabinoidoj sur lernado kaj memoraj procezoj de la dorsostria strio. Neurobiol Lernu Mem (2015) 125: 1 – 14. doi: 10.1016 / j.nlm.2015.06.008
139 Rueda-Orozco PE, Soria-Gomez E, Montes-Rodriguez CJ, Martínez-Vargas M, Galicia O, Navarro L, et al. Ebla funkcio de endocannabinoidoj en la elekto de navigada strategio de ratoj. Psikofarmacologio (2008) 198(4):565–76. doi:10.1007/s00213-007-0911-z
140 Goodman J, Packard MG. Periferaj kaj intra-dorsolateraj striataj injektoj de la cannabinoida ricevilo-agonisto WIN 55,212-2 difektas solidigon de stimulo-responda memoro. Neurokienco (2014) 274: 128 – 37. doi: 10.1016 / j.neuroscience.2014.05.007
141 Nazzaro C, Greco B, Cerovic M, Baxter P, Rubino T, Trusel M, et al. SK-kanala modulado savas striktan plastikecon kaj kontrolon de kutimo en kanabinoidaj toleremoj. Nat Neurosci (2012) 15: 284 – 93. doi: 10.1038 / nn.3022
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
142 Ames SL, Grenard JL, Stacy AW, Xiao L, He Q, Wong SW, et al. Funkcia bildado de implicaj marijuuaj asocioj dum agado en implicita asocia testo (IAT). Behav Brain Res (2013) 256: 494 – 502. doi: 10.1016 / j.bbr.2013.09.013
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
143 Redish AD, Jensen S, Johnson A. Unuigita kadro por toksomanio: vundeblecoj en la decidprocezo. Behav Brain Sci (2008) 31(04):415–37. doi:10.1017/S0140525X0800472X
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
144 Brevers D, Bechara A, Cleeremans A, Noël X. Iowa Gambling Task (IGT): dudek jarojn post - hazardludo kaj IGT. Antaŭa psikolo (2013) 4: 665. doi: 10.3389 / fpsyg.2013.00665
145 Koob GF, Le Moal M. Neurobiologiaj mekanismoj por kontraŭaj motivaj procezoj en toksomanio. Philos Trans R Soc B Biol Sci (2008) 363(1507):3113–23. doi:10.1098/rstb.2008.0094
146 Hyman SE, Malenka RC, Nestler EJ. Neŭralaj mekanismoj de toksomanio: la rolo de rekompenco-lernado kaj memoro. Annu Rev Neurosci (2006) 29: 565 – 98. doi: 10.1146 / annurev.neuro.29.051605.113009
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
147 Goodman J, Packard M. La memorsistemo okupita dum akiro determinas la efikecon de malsamaj estingaj protokoloj. Front Behav Neurosci (2015) 9: 314. doi: 10.3389 / fnbeh.2015.00314
148 Palencia CA, Ragozzino ME. La kontribuo de NMDA-riceviloj en la dorsolatera striato al egocentra respondo-lernado. Behav Neurosci (2005) 119(4):953–60. doi:10.1037/0735-7044.119.4.953
PubMed Abstracto | Plena Teksto de CrossRef | Google Scholar
149 Rueda-Orozco PE, Montes-Rodriguez CJ, Soria-Gomez E, Méndez-Díaz M, Prospéro-García O. Senvalorigo de endocannabinoids-agado en la dorsolateral striatum prokrastas formorton de konduto en procedura memora tasko en ratoj. Neuropharmacology (2008) 55(1):55–62. doi:10.1016/j.neuropharm.2008.04.013
Ŝlosilvortoj: memoro, drogmanio, hipokampo, striato, amigdalo, streso, angoro
Citado: Goodman J kaj Packard MG (2016) Memorsistemoj kaj la toksomaniita cerbo. Fronto. Psikiatrio 7: 24. doi: 10.3389 / fpsyt.2016.00024
Ricevita: 01 decembro 2015; Akceptita: 11 Februaro 2016;
Eldonita: 25 Februaro 2016
Eldonita de:
Vincent David, Centre National de la Recherche Scientifique (CNRS), Francio
Reviziita de:
Jacques Micheau, Universitato de Bordeaux 1, Francio
Roberto Ciccocioppo, Universitato de Kamerino, Italio
Kopirajto: © 2016 Goodman kaj Packard. Ĉi tio estas malferma-alira artikolo distribuita sub la kondiĉoj de la Krea Komunaĵo Atribuka Permesilo (CC BY). La uzo, dissendo aŭ reproduktaĵo en aliaj forumoj estas permesita, se la originala aŭtoro aŭ licencanto estas akredititaj kaj ke la originala publikigado en ĉi tiu ĵurnalo estas citita, laŭ akceptebla akademia praktiko. Neniu uzo, dissendo aŭ reprodukto estas permesata, kiu ne plenumas ĉi tiujn terminojn.
* Korespondado: Mark G. Packard, [retpoŝte protektita]
