Rimarkoj: Unu el la malmultaj studoj, kiuj traktas kiel la abstinado efikas sur la niveloj de dopaminaj receptoroj en primatoj.
- D2-riceviloj resaltas sufiĉe rapide - malpli ol monaton
- D1-riceviloj estas multe tro altaj je monato, sed resaltas en 90-tagoj.
- Altaj aŭ malaltaj D1-receptoroj povas esti ŝlosiloj por akra retiriĝo kaj avidoj
Thomas JR Beveridge1, Hilary R Smith1, Michael A Nader1 kaj Linda J Porrino1
1Fako de Fiziologio kaj Farmakologio, Centro por la Neurobiologia Esploro de Drogaj Misuzoj, Universitato de Medicina Lernejo de Vekaj Arbaro, Winston-Salem, NC, Usono
Korespondado: Dr LJ Porrino, Sekcio de Fiziologio kaj Farmakologio, Centro por Neurobiologia Esploro de Drogaj Misuzoj, Vek-Arbara Universitato-Lernejo de Medicino, Boulevard Medicina Centro, Winston-Salem, NC 27157-1083, Usono. Tel: + 1 336 716 8575; Telefaksilo: + 1 336 716 8501; Retpoŝto: [retpoŝte protektita]
Ricevita la 29an de aprilo 2008; Reviziita la 25an de julio 2008; Akceptita la 30an de julio 2008; Eldonita interrete la 3an de septembro 2008.
abstrakta
Kvankam malreguligo ene de la dopamina (DA) sistemo estas karakteriza trajto de kronika ekspozicio al kokaino, la demando pri ĉu ĉi tiuj ŝanĝoj persistas ĝis abstinado restas plejparte sen respondo. Nehomaj primatoj reprezentas idealan modelon en kiu taksi la efikojn de sindeteno sur la DA-sistemo post kronika eksponado al kokaino. En ĉi tiu studo, virseksaj simioj mem-administris kokainon (0.3) mg/kg per injekto, 30-plifortigiloj por sesio) sub fiksa intervalo 3-min schedule por 100-tagoj sekvataj de 30 aŭ 90 tagoj de sindeteno. Ĉi tiu daŭro de kokain-memadministrado antaŭe estis montrita malpliigante DA-D2-similajn ricevilojn kaj pliigas nivelojn de D1-similaj riceviloj kaj DA-transportiloj (DAT). Respondado per kontrolaj simioj estis konservita per manĝa prezento sub identa protokolo kaj la samaj sindetenaj periodoj. [3H]SCH 23390 liganta al La riceviloj de DA D1 post la tagoj de abstinado de 30 estis signife pli altaj en ĉiuj porcioj de la striato, kompare kun bestoj de kontrolo, dum [3H]raclopride liganta al DA D2-receptoroj ne diferencis inter grupoj. [3H]WIN 35 428 liganta al DAT ankaŭ estis signife pli alta tra preskaŭ ĉiuj porcioj de la dorsal kaj ventra striatum post 30 tagoj de sindeteno. Post 90-tagoj da sindeteno, tamen, niveloj de DA D1-receptoroj kaj DAT ne diferencis de kontrolaj valoroj. Kvankam ĉi tiuj rezultoj indikas, ke ekzistas eventuala resaniĝo de la apartaj elementoj de la DA-sistemo, ili ankaŭ emfazas la dinamikan naturon de ĉi tiuj eroj dum la komencaj fazoj de sindeteno de kronika kokain-memadministrado.
Ŝlosilvortoj:
kokaino, dopamino, autoradiografio, abstinado, striato
ENKONDUKO
Kronika uzo de kokaino inter homaj toksomaniuloj estis asociita kun neŭroadaptiĝoj en la dopamina (DA) sistemo (Maliso et al, 1998; Volkow et al, 1993, 1997). Ĉi tiuj inkluzivas pliigojn en la denseco de DA-transportiloj (DAT) kaj malkreskojn en la koncentriĝoj de DA D2-similaj riceviloj (iom et al, 1999; Mash et al, 2002; Volkow et al, 1993). Krome, ŝanĝoj en liberigo de DA ankaŭ estis observitaj. Ekzemple, enketistoj uzantaj studojn pri pozitron-emisi-tomografio (PET) [11C]raclopride kaj metilfenidato pruvis malpliiĝojn de liberigo de DA en la striatumo de uzantoj de kronika kokaino (Volkow et al, 1997; Wong et al, 2006). Unu problemo tamen estas, ke ofte malfacilas ekskludi la influon de aliaj faktoroj kiel uzado de aliaj kontraŭleĝaj kaj laŭleĝaj drogoj, diferencoj en antaŭa konsumado kaj uzado de drogoj kaj diferencoj en vivstilo. Ĉi tiuj diferencoj, same kiel la ekzisto de kondiĉoj, kiuj povas antaŭiĝi kun drogoj, povas limigi la interpretadon de studoj ĉe homaj pacientoj.
Nehomaj primataj modeloj, en kiuj oni povas sisteme manipuli, reprezentas alternativan aliron al la studo de la konsekvencoj de kronika kokain-memadministrado kaj posta sindeteno. Antaŭaj studoj pruvis, ke kronika ekspozicio al kokaino estas akompanata de signifaj malpliiĝoj en la koncentriĝoj de DA D2-receptoroj kaj ankaŭ altoj en la niveloj de D1-receptoroj kaj la denseco de la DAT (Letchworth et al, 2001; moore et al, 1998a, 1998b; Nader et al, 2002, 2006). Ĉi tiuj efikoj spegulas tiujn vidatajn en homoj, kaj tiel substrekas la utilecon de ĉi tiuj modeloj de drogekspozicio.
Kvankam ekzistas konsiderinda pruvo pri malreguligo de DA-sistemoj, ĝi pruvis pli malfacila taksi ĉu ekzistas iuj evidentecoj por reakiro post ĉesigo de uzado de drogoj (Maliso et al, 1998; Jacobsen et al, 2000; Volkow et al, 1993) aŭ ĉu ĉi tiuj ŝanĝoj daŭras preter la tempa kadro de daŭra kokainekspozicio. Denove, nehomaj primataj modeloj povas doni komprenojn pri ĉi tiu fazo de toksomanio. Farfel et al (1992) raportis malpliigitajn koncentriĝojn de DAT kaj D1-similaj riceviloj en la striato de simioj post abstinado de kronika nekontinenta ekspozicio al kokaino. Tamen, la specifa rolo de sindeteno estis malfacile konstatebla pro la manko de mezuradoj en grupo kun neniu sindetena periodo. Simile, Melega et al (2008) raportis signife malpliiĝantajn nivelojn de DAT en la striato de verdaj simioj post 3-semajnoj da sindeteno de kreskanta metamfetamina reĝimo. La administrado de stimuliloj en ambaŭ studoj tamen estis nekontinua. La vojo de administrado (kontingento vs nekontraktita) pruviĝis efiki la cerbon malsame rilate al ambaŭ liberigo de DA (Hemby et al, 1997) kaj glukoza metabolo (Graham kaj Porrino, 1995; Porrino et al, 2002). Tiel la uzo de memadministrado en la nuna studo evitas ĉi tiun aferon. Krome, la efikoj de longtempa kokain-memadministrado sur cerbaj DA-sistemoj estis amplekse studitaj uzante ĉi tiun modelon de memadministrado en simioj de ruso, tiel provizante bazlinion, por taksi la neŭroadaptojn okazantajn dum sindeteno.
La celo de ĉi tiuj studoj estis do determini ĉu la ŝanĝoj en DAT kaj DA D1 kaj D2-receptor-koncentriĝoj, kiuj pruvis antaŭe en bestoj eksponitaj al kokain-memadministrado (Letchworth et al, 2001; moore et al, 1998a, 1998b; Nader et al, 2002) renversus post plilongigitaj periodoj de sindeteno. Surbaze de studoj en homaj drogantoj (kp Volkow et al, 1993), ni hipotezis, ke ĉi tiuj ŝanĝoj en la DA-sistemo persistus eĉ post 3-monatoj da sindeteno. Tiucele, simioj mem-administris kokainon por 100-sesioj, kun totalaj konsumoj de 900 mg/kg, sekvita de 30 aŭ 90 tagoj de sindeteno de la drogo. DA D1 kaj D2-receptoroj, same kiel la DAT, estis mezuritaj kun kvantaj en vitro aŭtoradiografio de riceviloj.
METODOJ
temoj
Entute 17 eksperimente naive plenkreskaj masklaj rhesus simioj (Macaca mulatta) pesante inter 7.7 kaj 13 kg (mezumo ± SD, 10.2 ± 1.32) komence de la studo servis kiel subjektoj. Ĉiuj procedoj estis plenumitaj laŭ establitaj praktikoj kiel priskribitaj en la Nacia Institutoj pri Sano-Gvidilo por Prizorgo kaj Uzo de Laboratoriaj Bestoj. Krome ĉiuj proceduroj estis reviziitaj kaj aprobitaj de la Komitato pri Uzado de Bestoj kaj Uzado de Vaka Arbara Universitato. Simioj estis individue loĝataj en neoksideblaj kaĝoj kun akvo ad libitum; bestoj havis fizikan kaj vidan kontakton inter si. Iliaj korpaj pezoj estis konservitaj proksimume 90-95% de senpage nutrantaj pezoj per banan-flavaj buletoj gajnitaj dum la eksperimentaj sesioj kaj per suplementa nutrado de Lab Diet Monkey Chow, provizitaj ne pli frue ol 30 min post-kunsido. Krome, ili ricevis freŝan frukton aŭ arakidojn almenaŭ tri fojojn semajne. Ĉiu simio estis pesita unufoje semajne, kaj se necese, iliaj dietoj estis alĝustigitaj por konservi stabilajn pezojn.
Konduta Aparato
Eksperimentaj kunsidoj estis farataj en ventolitaj kaj son-atenaj ĉambraj ĉambroj (1.5 × 0.74 × 0.76 m; Med Associates Inc., East Fairfield, VT) projektita por akomodi primatan seĝon (Modelo R001; Primate Products, Redwood City, CA). La ĉambro enhavis inteligentan panelon (48 × 69) cm), kiu konsistis el du lertaj leviloj (5) cm larĝa) kaj tri stimulaj lumoj. La leviloj estis poziciigitaj facile de la simio sidanta en la primata seĝo. Banan-flavaj manĝaĵaj buletoj (1) g; Bio-Serv, Frenchtown, NJ) estis liverita de nutranto situanta supre de la ĉambro. Peristaltika infuza pumpilo (7531-10; Cole-Parmer Co., Chicago, IL) estis uzata por liveri drogajn injektojn kun rapideco de proksimume 1 ml po 10 s al tiuj bestoj memadministrantaj kokainon. Funkciado de la ĉambroj kaj akira datumo realiĝis per komputila sistemo Power Macintosh kun interfaco (Med Associates Inc.).
Kirurgiaj Procedoj
Ĉiuj simioj, inkluzive de kontroloj, estis kirurgie pretaj, en senfruktaj kondiĉoj, kun enhejmaj kateteroj kaj vaskulaj havenaj havenoj (Modelo GPV; Access Technologies, Skokie, IL). Simioj estis anestezitaj per kombinaĵo de ketamina (15) mg/kg, im) kaj butorfanolo (0.03) mg/kg, im) kaj incizo fariĝis proksime de la femora vejno. Post malkaŝa disiĝo kaj izolado de la vejno, la proksimala fino de la katetero estis enigita en la vejnon por distanco kalkulita finiĝi en la malsupera vena cava. La distala fino de la katetero estis fadenigita subkutane al incizo iomete ekster la mezlinio de la dorso. La vaskula alira haveno estis metita ene de poŝo formita de malakra disiĝo proksime de ĉi tiu incizo. Simioj estis donitaj 24-48 h resaniĝa tempo antaŭ ol reveni al manĝ-plifortigita respondo. Proksimume 5 tagojn antaŭ la fina proceduro, ĉiu simio estis enplantita kun kronika enhejmiga katetero en la apuda femora arterio por kolekto de tempigitaj arteriaj sangaj specimenoj. La kirurgiaj proceduroj estis identaj al tiuj priskribitaj por la venenaj kateteroj. En la tago de la fina sesio, studo pri fina cerba glukoza glukozo estis farita en kiu oni injektis simiojn kun 2-[14C]deoxyglucose (2-DG) proksimume 2 min post la fino de la kunsido kaj sangaj specimenoj estis akiritaj tra la arteria katetero super 45 min periodo (vidu Beveridge et al, 2006 por detaloj). Datumoj pri metabolismo de ĉi tiuj studoj ne estas prezentitaj ĉi tie.
Mem-Administraj Proceduroj
Simioj estis komence trejnitaj por respondi sur unu el du leviloj plifortigante ĉiun respondon sur la ĝusta levilo per manĝa buleto. Dum proksimume 3-semajna periodo la intervalo inter havebleco de manĝaĵaj buletoj estis iom post iom pliigita ĝis 3-min-intervalo estis atingita (t.e. fiks-intervala 3-min-horaro de plifortigo; FI 3-min). Sub la fina horara kondiĉo, la unua respondo sur la levilo post 3 min rezultis en liverado de manĝa buleto; sesioj finiĝis post 30-manĝaĵaj prezentoj. Al la fino de ĉiu kunsido, la respondaj leviloj estis retiritaj, hejmlumoj kaj stimulaj lumoj estingiĝis, kaj bestoj restis en la malhela ĉambro dum proksimume 30 min antaŭ ol ili estis revenitaj al siaj hejmaj kaĝoj. Ĉiuj simioj respondis sub la horaro FI 3-min de prezento de manĝaĵoj por almenaŭ 20-sesioj kaj ĝis stabila agado estis akirita (± 20% de la mezumo por tri sinsekvaj sesioj, sen tendencoj en respondokvantoj). Kiam manĝebla respondado estis stabila, la manĝanto estis senŝaltita kaj la efikoj de formorto sur respondado estis ekzamenitaj dum kvin sinsekvaj sesioj, post kio respondado estis reestablita kaj konservita per manĝa prezento.
Post kiam baza agado estis establita, ĉiuj simioj estis kirurgie pretaj kun venenaj kateteroj, kiel priskribite supre, kaj hazarde atribuitaj al unu el tri grupoj. Unu grupo de simioj funkciis kiel kontroloj kaj daŭre respondis sub la horaro de FI 3-min de prezento de manĝaĵoj por tuta de 100-sesioj (N=6). La ceteraj 11 simioj estis asignitaj al la kokainaj memadministraj grupoj (0.3) mg/kg por injekto). Ĉar 0.3 mg/kg da kokaino per injekto estis konsiderata alta dozo por antaŭe kokainaj naivaj simioj, por plej multaj bestoj ĉi tiu dozo estis atingita en du sesioj unue permesante al la simio mem-administri 0.1 mg/kg kokaino. Manĝaĵo-konservita agado estis permesita stabiligi post kirurgio (proksimume 4-6 tagoj) antaŭ ol kokainaj memadministraj sesioj komenciĝis. Antaŭ ĉiu eksperimenta kunsido, la dorso de la besto estis purigita per 95% etanolo kaj betadina kribrilo kaj 22-mezurilo-nadlo Huber Point (Modelo PG20-125) estis enmetitaj en la havenon kondukantan al la venena katetero, konektante infuzan pumpilon enhavantan kokainan solvon al la katetero. Antaŭ la komenco de la kunsido, la bombo funkciigis dum proksimume 3 s, plenigante la havenon kun la dozo de kokaino disponebla dum la eksperimenta kunsido. Kunsidoj finiĝis post 30-injektoj; kiel sub kontrolkondiĉoj, simioj restis en mallumigita ĉambro dum proksimume 30 min. Fine de ĉiu kunsido, la haveno estis plenigita per heparinizita salo (100) U/ml) por helpi eviti koaguliĝon.
Eksperimentaj kunsidoj estis kondukitaj proksimume samtempe saman tagon kaj daŭris dum entute 100-sesioj. Post la kompletigo de la 100-sesioj, abstinenta periodo de 30 aŭ 90-tagoj estis enkondukita dum kiu tempo katetroj estis enverŝitaj ĉiutage kun heparinizita salo, sed neniuj kokainaj aŭ manĝaj administradaj sesioj estis faritaj. Por la grupo de kontrolo, abstinaj periodoj de 30-tagoj estis truditaj al kvar bestoj kaj 90-tagoj en la du aliaj. Por la kokaina grupo, sindetenaj periodoj de 30-tagoj estis truditaj al ok bestoj kaj 90-tagoj al tri bestoj. Je la fino de la sindeteno unu fina memadministra sesio (manĝa kontrolo aŭ kokaino) estis kondukita kaj la 2-DG-proceduro estis komencita tuj post la sesio. En du kontroloj kaj kvar kokainaj memadministraj bestoj en la 30-taga sindetena grupo, neniu kokaino ricevis en la fina kunsido. Bestoj estis home mortigitaj per superdozo de pentobarbital (100) mg/kg, iv) fine de la 45 min.
Prilaborado de Tissoj
Post mortigo, cerboj estis tuj forigitaj, blokitaj kaj frostigitaj en izopentano je −35 al −55 ° C kaj tiam konservitaj al −80 ° C. La ŝtofaj blokoj enhavantaj striaton tiam estis tranĉitaj en kriostatato je −20 ° C en la korona ebeno en 20 μm sekcioj, kolektitaj sur elektrostatike lumigitaj lumbildoj, desicadas sub vakuo dum la nokto ĉe 4 ° C, tiam konservitaj al −80 ° C ĝis pretigitaj por aŭtradiografio. Cerbaj sekcioj estis kolektitaj el la partoj de la kaŭdita kerno, putameno kaj kerno accumbens, kiuj kuŝas rostral al la antaŭa komisaro. Ĉi tiu regiono estas nomata la antaŭkonduta striatumo. Plue, rostraj kaj kaŭdaj niveloj de la presregula striatumo estis nomumitaj kun referenco al la kerno accumbens. La rostra antaŭkonduta striatumo estas la regiono kie la kerno acumbens ne diferenciĝas en apartajn ŝelajn kaj kernajn subsekciojn. La kaŭdika antaŭregata striato estas la regiono kongrua kun la aspekto de la ŝelo kaj kerno de la kerno accumbens, kiu estas posta al la apero de la olfakta tubero. Por ĉiu el la ligaj studoj, du apudaj sekcioj estis prenitaj je ĉiu el kvin niveloj (du rostraloj kaj tri kaŭdaj) tra la antaŭkomprena striato por entute 10-sekcioj per besto.
D1-Benda Receptoro
Oni dense determinis la densecojn de DA-D1-receptoro [3H]SCH 23390 (specifa aktiveco - 85 Ci/mmol; PerkinElmer, Boston, MA) laŭ kvanto en vitro aŭtoradiografio de riceviloj laŭ procedoj adaptitaj de Lidow et al (1991) kaj Nader et al (2002). Sekcioj estis preincubadas por 20 min en bufro (50) mM Tris, 120 mM NaCl, 5 mM KCl, 2 mM CaCl2, 1 mM MgCl2, pH 7.4, 25 ° C) por forigi endogenan DA, kokainon kaj [14C] de la proceduro 2-DG. Sekcioj tiam estis inkubitaj por 30 min en la sama bufro, pH 7.4, 25 ° C, enhavanta 1 mM ascorbika acido, 40 nM ketanserin, kaj 1 nM [3H]SCH 23390. Post kovado, sekcioj estis lavitaj dufoje por 20 s en bufro enhavanta 1 mM ascorbika acido ĉe pH 7.4, 4 ° C, tiam trempis en distilitan akvon ĉe 4 ° C, kaj sekigis sub torento de malvarmeta aero. Nespekta ligado estis difinita per inkubacio de apudaj sekcioj en la inkuba solvo en ĉeesto de 5 μM (+) -butaclamol. Sekcioj, kune kun kalibritaj [3H] aŭtoradiografiaj normoj (Amersham, Piscataway, NJ), estis postulataj de Kodak Biomax MR-filmo (Fisher Scientific, Pittsburgh, PA) dum 6-semajnoj.
D2-Benda Receptoro
Oni determinis la densecon kaj distribuadon de DA-D2-receptoroj [3H]raclopride (specifa agado, 87 Ci/mmol; PerkinElmer) laŭ procedoj adaptitaj de Lidow et al (1991) kaj Nader et al (2002). Sekcioj estis preincubadas por 20 min en bufro (50) mM Tris, 120 mM NaCl, 5 mM KCl, pH 7.4, 25 ° C) por forigi endogenan DA, kokainon, kaj [14C] de la proceduro 2-DG. Glitiloj tiam estis inkubitaj por 30 min en la sama bufro, enhavanta 5 mM ascorbika acido kaj 2 nM [3H]raclopride. Sekcioj estis elŝovitaj 3 × 2 min en bufron ĉe pH 7.4, 4 ° C, tiam trempis en distilitan akvon ĉe 4 ° C, kaj sekigis sub torento de malvarmeta aero. Nespekta ligado estis difinita per inkubacio de apudaj sekcioj en la inkuba solvo en ĉeesto de 1 μM (+) –Butaclamol. Sekcioj, kune kun kalibritaj [3H] autoradiografiaj normoj, estis aplikitaj al la filmo de Kodak Biomax MR dum 8-semajnoj.
Dopamina Transporta Bindado
La denseco de DAT-ligaj retejoj estis determinita uzante [3H]WIN 35,428 (specifa agado, 87 Ci/mmol; PerkinElmer) autoradiografio laŭ procedoj adaptitaj de Canfield et al (1990) kaj Letchworth et al (2001). Ŝnuraj sekcioj estis preincubitaj en bufron (50) mM Tris, 100 mM NaCl, pH 7.4, 4 ° C) por 20 min por forigi ajnan restaĵon DA, kokainon, kaj [14C] de la proceduro 2-DG. Sekcioj tiam estis inkubitaj por 2 h je 4 ° C en la sama bufro enhavanta 5 nM [3H]VENU 35 428. Sekcioj estis lavitaj por tuta de 2 min en bufron ĉe 4 ° C, tiam trempis en distilitan akvon ĉe 4 ° C, kaj sekigis sub torento de malvarma aero. Nespekta ligado estis difinita per inkubacio de apudaj sekcioj en la inkuba solvo en ĉeesto de 30 μM kokaino. Sekcioj, kune kun kalibritaj [3H] autoradiografiaj normoj, estis aplikitaj al la filmo de Kodak Biomax MR dum 6-semajnoj.
Densitometrio kaj Datuma Analizo
Filmoj estis evoluigitaj kun Kodak GBX-programisto, haltejo kaj Rapidfiksilo (VWR, West Chester, PA), kaj tiam enŝovitaj. Analizo de autoradiogramoj estis farita per kvanta densitometrio kun komputiligita bilda prilabora sistemo (MCID, Imaging Research; InterFocus Imaging Ltd, Kembriĝo, Britujo). Optikaj densecvaloroj estis konvertitaj al fmol/mg (de malseka-peza histo) per referenco al la kalibrita [3H] normoj. Specifa bindado estis determinita ciferece subtrahante bildojn de nespecifaj ligoj de supermetitaj apudaj bildoj de totala ligado. Strukturoj estis identigitaj per Nissl-makulado de sekcioj najbaraj al tiuj analizitaj por receptoro ligado. Datumoj de ĉiu provo estas analizitaj sendepende per unudirekta analizo de varieco sekvita de malplej kvadrataj diferencoj post hoc testoj por multnombraj komparoj. Ĉiu regiono konsistis en aparta analizo. Ĉar ligaj datumoj akiritaj de la kontrolaj bestoj abstinaj dum 30 kaj 90-tagoj ne signife diferencis unu de la alia, similaj al antaŭaj studoj (Nader et al, 2002), datumoj de la kontrolgrupoj estis kombinitaj. Krome, ne estis signifaj diferencoj inter la datumoj de tiuj bestoj, kiuj ricevis kokainon kaj tiujn, kiuj ne faris ĉe ilia fina kunsido, do datumoj de ĉi tiuj grupoj ankaŭ estis kombinitaj.
REZULTO
Efikoj de Abstinenco de Kronika Mem-Administrado por Kokaina 30-Tagoj
Koncentriĝoj de [3H]SCH 23390-ligado al DA D1-riceviloj en la antaŭkomprena striatumo estas montrita en tablo 1. Specifa ligado de [3H]SCH 23390 enkalkulis pli grandan ol 90% de totala ligado. Konsentite kun antaŭaj raportoj (moore et al, 1998a; Nader et al, 2002), la ligado de [3H]La riceviloj de SCH 23390 al D1 en bestoj kontrolantaj ne-drogojn estis heterogenaj kun estimindaj diferencoj en la grado de ligado inter subregionoj de la striato. Etikedado estis pli densa en la pli vizaĝaj kaj mezaj porcioj tra la striato.
![Tabelo 1 - Efekto de 30 aŭ 90 Tagoj de Abomeno Sekvante Kronikan Mem-Administradon pri Kokaino sur Loĝdensoj de [lsqb] 3H [rsqb] SCH 23390 Binding to D1 Receptors in Precommissural Striatum of Rhesus Monkeys - Bedaŭrinde ni ne kapablas havigi alternativan tekston ĉi tio. Se vi bezonas helpon por aliri ĉi tiun bildon, bonvolu kontakti helpo@nature.com aŭ la aŭtoro](https://yourbrainonporn.com/wp-content/uploads/2020/01/table_thumb.gif)
Post 30-tagoj da sindeteno de kokaina ekspozicio, ligado al D1-receptoroj estis karakterizita per ĝeneraligitaj altoj tra la tuta vizaĝo-kaŭdika etendo de la antaŭ-admira striatum, kompare kun ligaj densecoj en ne-drogaj kontrolaj bestoj (tablo 1; figuro 1). En la pli rostrala striatumo, koncentriĝoj estis signife pli altaj en la kava kerno inkluzive de la dorsolatera (+27%), centra (+27%), dorsomedialo (+27%), kaj ventromedial (+23%) porcioj, same kiel en la dorso (+17%), centra (+22%), kaj ventrala (+23%) porcioj de la putamenoj, kompare kun la densecoj en ne-drogaj kontrolaj bestoj. Signifaj altoj ankaŭ estis evidentaj en la kerno accumbens (+23%) je ĉi tiu nivelo. Je la nivelo de la striato, kie la kerno kaj ŝelo de la kerno accumbens estas plej diferencaj, densecoj de similaj riceviloj de D1 ankaŭ estis signife pli altaj tra la dorsolatera (+31%), centra (+29%), dorsomedialo (+30%), kaj ventromedial (+18%) kaŭza kerno, same kiel la dorso (+23%), centra (+29%), kaj ventrala (+28%) putamen, kompare kun densecoj de ne-drogaj kontroloj. Ene de la ventrala striato ĉe ĉi tiu nivelo, la koncentriĝo de D1 ricevilaj ligaj retejoj estis pli alta en la kerno de nucleus accumbens (+45%) kaj ŝelo (+20%), same kiel en la olfakta tubero (+26%), kompare kun la densecoj en kontroloj.
Figuro 1.
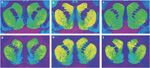 Reprezentaj aŭradiogramoj de [3H] SCH 23390 liganta al D1-riceviloj (supra panelo) kaj [3H]WIN 35428 liganta al dopamina transportiloj (funda panelo) en koronaj sekcioj de rhesus simio striatum. (a, d) Kontrolu beston respondantan por manĝa plifortigo. (b, e) Kokaino memadministra besto kun sindeteno de 30 tagoj. (c, f) Kokaino memadministra besto kun sindeteno de 90 tagoj.
Reprezentaj aŭradiogramoj de [3H] SCH 23390 liganta al D1-riceviloj (supra panelo) kaj [3H]WIN 35428 liganta al dopamina transportiloj (funda panelo) en koronaj sekcioj de rhesus simio striatum. (a, d) Kontrolu beston respondantan por manĝa plifortigo. (b, e) Kokaino memadministra besto kun sindeteno de 30 tagoj. (c, f) Kokaino memadministra besto kun sindeteno de 90 tagoj.
Koncentriĝoj de [3H]raclopride liganta al DA D2-riceviloj en la antaŭkomprena striatum estas montrita en tablo 2. Specifa ligado kun [3H]raclopride enkalkulis pli grandan ol 90% de totala ligado. La distribuo de [3H]raclopride liganta al D2-receptoroj estis ankaŭ heterogena tra subregionoj de la dorsal kaj ventra striatum kiel en antaŭaj raportoj (moore et al, 1998b; Nader et al, 2002). En la ne-elmetitaj kontroloj de drogoj, pli altaj koncentriĝoj de D2-ligaj lokoj estis prezencaj en la dorsoj kompare kun la ventra striato. Krome, estis evidenteco de mezgranda ĝis flanka gradiento kun la pli altaj koncentriĝoj de ligaj lokoj ĉeestantaj en la pli flankaj porcioj de la striato.
Post 30 tagoj de sindeteno, niveloj de D2-ricevilo en bestoj eksponitaj al kokaino kaj plifortigitaj manĝaĵoj ne signife diferencis unu de la alia en la plej multaj regionoj de la striato. Pli altaj koncentriĝoj de ligaj lokoj estis observitaj en la ventrala putamen (+10%) kaj antaŭa kerno accumbens (+12%) en histo el la simpatioj kun kokaino kompare kun kontroloj. Neniuj aliaj signifaj diferencoj estis rimarkitaj.
Koncentriĝoj de [3H]VIN 35428 liganta al DAT en la antaŭkonduta striatumo estas montrita en tablo 3. Konsentite kun antaŭaj raportoj (Letchworth et al, 2001), ligante al DAT-ejoj en ne-drogaj bestoj estis pli alta en dorsalo kompare kun ventrala striatumo. Ene de la kerno accumbens, pli altaj densecoj estis observitaj en la kerno kompare kun la ŝelaj dividoj. Fine, nespecifita ligado respondecis pri malpli ol 10% de la tuta.
Post 30 tagoj de sindeteno de kokainekspozicio, ligado al la DAT estis signife pli alta tra la plimulto de regionoj de la striatum rostral al la antaŭa komisaro kompare kun ligado en bestoj de kontrolo ne drogitaj (figuro 1). Specife, la koncentriĝoj de DAT-ligaj lokoj ĉe rostralaj niveloj estis signife pli altaj en centra (+22%), dorsomedialo (+25%), kaj ventromedial (+28%) kaŭza kerno kaj en la dorso (+16%) kaj centra (+23%) putamen, kompare kun manĝ-plifortigitaj kontroloj. Krome, ankaŭ estis signife pli alta ligado al DAT en la antaŭa kerno accumbens (+37%) en kokaino - kompare kun manĝ-plifortigitaj simioj. Ene de la pli kaŭdaj porcioj de la presregata striatumo, densecoj de DAT-ligaj lokoj estis signife pli altaj en la centra kadavra kerno (+21%), kaj putamen, centra (+20%) kaj ventra (+19%; figuro 1). Ene de la ventra striato ĉe ĉi tiu nivelo, ligado al DAT estis signife pli alta en la kerno de la kerno (+20%) kaj olfakta tuberkulo (+24%) en histo el kokaino vs manĝ-plifortigitaj simioj.
Efikoj de Abstinenco de Kronika Mem-Administrado por Kokaina 90-Tagoj
Kontraste al la ĝeneraligitaj diferencoj en la denseco de lokoj de ligaj riceviloj de D1 observitaj en eksponitaj al kokaino post 30 tagoj de sindeteno, post 90 tagoj de sindeteno, ne estis signifaj diferencoj kompare kun kontroloj kun manĝaĵo plifortigita en iu ajn parto de la antaŭkomisaro. striato (tablo 1; figuro 1). Simile, koncentriĝoj de [3H]raclopride liganta al DA D2-riceviloj en la antaŭkomprena striatumo post 90-tagoj de sindeteno ankaŭ ne signife diferencis de tiuj en ne-elmetitaj drogoj (tablo 2).
La koncentriĝoj de [3H]WIN 35428 liganta al DAT montris similan padronon al tio observita kun D1 kaj D2-riceviloj. Ekzistis neniuj signifaj diferencoj en la denseco de DAT inter malkovritaj de kokaino post 90-tagoj da sindeteno kompare al niveloj en ne-elmetitaj drogoj (tablo 3; figuro 1), kvankam oni devas rimarki, ke estis tendenco al pli altaj niveloj de ligado en la antaŭa kerno accumbens.
DISCUSO
Antaŭaj studoj de nia grupo montris, ke kronika ekspozicio al mem-administrado de kokaino estas akompanata de signifa malreguligo de la DA-sistemo de nehomaj primatoj (Letchworth et al, 2001; moore et al, 1998a, 1998b; Nader et al, 2002, 2006). La rezultoj de la nuna studo pruvas, ke ĉi tiu regregado restas evidenta post ĉeso de kokaina ekspozicio. Post 30-tagoj da sindeteno, la koncentriĝoj de DA D1-receptoroj kaj la DAT estis signife levitaj tra la striato de simioj kun historioj de kronika kokain-memadministrado kompare kun manĝaĵ-plifortigitaj kontroloj. Tamen, la nuna studo ankaŭ donas klaran evidentecon pri resaniĝo en la DA-sistemo post pli longaj periodoj de sindeteno (90-tagoj), kiel evidentigita per la manko de signifaj diferencoj inter bestoj eksponitaj al kokaino kaj kontrolaj bestoj en ĉi tiu tempo. Ĉi tiuj datumoj sugestas, ke ekspozicio al kokaino eble ne produktas konstantajn ŝanĝojn en la DA-sistemo, sed ke reakiro povas okazi kun plilongigita sindeteno de drog-uzo.
La malreguligado de DAT-koncentriĝoj montritaj ĉi tie post abstinado estas konforma al antaŭaj raportoj en nehomaj primatoj (Letchworth et al, 2001), kiuj montris signifajn altojn en la densecoj de DAT-ligaj lokoj en ventra kaj dorsal-striato. Kvankam ne estas eksplicite testita, ĝi ŝajnas, ke post ĉeso de drogekspozicio, la altoj en DAT-liganta loko-denseco estas almenaŭ same grandaj kaj pli disvastigitaj tra la regionoj de la striatumo ol tiuj raportitaj sen iu forira periodo (Letchworth et al, 2001). Simile, la altaj koncentriĝoj de D1-receptor-ligitaj lokoj observitaj ĉi tie post 30-tagoj de sindeteno ankaŭ konformas al antaŭaj studoj, kiuj montris pliigitajn D1-similajn receptorojn ligantajn en la striato de nehomaj primatoj elmontritaj al identa reĝimo de kokain-mem-administrado (Nader et al, 2002). Kontraŭe, ne estis signifaj diferencoj observitaj inter la niveloj de D2-similaj ricevilo-ligantaj densecoj en la striato de kokain-elmontritaj kaj kontrolaj bestoj. Ĉi tiu manko de regregado ĉeestis, malgraŭ la grandaj malpliiĝoj en la koncentriĝoj de D2-receptoroj, kiuj estis raportitaj en ambaŭ homaj toksomaniuloj (Volkow et al, 1993) kaj bestaj modeloj de kokaina memadministrado (moore et al, 1998a, 1998b; Nader et al, 2002, 2006). La nunaj datumoj, do, sugestas pli rapidan normaligon por kontroli nivelojn en ĉi tiu sistemo kompare kun D1-similaj riceviloj kaj la DAT. Kunigitaj, la ŝanĝoj en DA-riceviloj kaj la DAT klare montras, ke la periodo tuj post ĉeso de kokain-memadministrado estas tre laborebla kun konsiderindaj ŝanĝoj en la regulado de la DA-sistemo, sed ke ĉi tio estas sekvita de re-reguligo de la sistemo alproksimiĝas al pli normala distribuo de DA-riceviloj kaj DAT post pli daŭra sindeteno.
Ŝanĝoj de D1-Ricevilo
La ĝeneraligitaj altoj en la denseco de D1-riceviloj observitaj ĉi tie post ĉeso de uzado de kokaino estas konformaj al raportoj montrantaj pliigitan sentivecon de D1-riceviloj dum retiriĝo. Mezuro de D1-sentiveco estis farita en studo de Henry kaj Blanka (1991), en kiu oni trovis, ke unuecaj registradoj de neŭronoj en la kerno akcina pli sentemaj al la agonisto de la ricevilo D1 SKF 38393 sekvante kronikajn ĉiutagajn injektojn de kokaino kompare kun salaj traktitaj kontroloj. Ĉi tiu efiko estis konstanta, ĉar pliigita sentiveco restis evidenta ĝis unu monato post retiriĝo. La aŭtoroj hipotezis, ke la sensibilizo de la ricevilo D1 ŝuldiĝis al la aŭtoreceptoro de D2-subsensiveco en la somatodendritika A10-areo, tiel reduktante inhibician fluon de fluo tra la DA-mezoakumbensaj sistemoj (Henry and White, 1991). La datumoj de la aktuala studo sugestas pliigadon de D1-receptoroj en abstinado, kio povus klarigi ĉi tiun pliigitan sentivecon de dopaminergiaj neŭronoj al rekta aganta D1-receptoro agonisto. Plue, la plibonigita efiko de SKF 38393 sur neŭronoj de nucleus accumbens ne ŝajnis du monatojn post retiriĝo, sugestante, ke estas reakiro de la sentemo de la ricevilo D1 (Henry and White, 1991); rezulto konforma al la reakiro de D1-receptor-densecoj rimarkitaj en la nuna studo post 90-tagoj da sindeteno. Aliaj raportoj ankaŭ subtenas gravan rolon por D1-riceviloj en relanĉo. La rekta stimulo de D1-receptoroj en la ŝelo de la kerno accumbens povas reinstali kokainon serĉantan abstinajn ronĝulojn (Schmidt et al, 2006). Tamen, la literaturo estas iom malkonsekvenca en tio, ke kaj D1-agonistoj kaj antagonistoj povas mildigi drogojn serĉantajn provokitajn de kokainaj primoj aŭ de kokainaj stimuloj (Alleweireldt et al, 2002; De Vries et al, 1999; Khroyan et al, 2000; self et al, 1996; blanka et al, 2001). Ĵus, Khroyan et al (2003) raportis, ke D1-agonistoj kaj antagonistoj reduktas revenon en nehoma primata modelo de serĉado de kokaino. Ĉi tiuj aŭtoroj sugestis, ke eble ekzistas kritika gamo de receptoroj de D1 necesa por kokaina serĉado, kaj ke ambaŭ antagonistoj kaj agonistoj povus elŝovi aktivecon el ĉi tiu fenestro. La pliigitaj koncentriĝoj de D1-receptoroj kiuj akompanas sindetenadon povus modifi ĉi tiun intervalon, rezultigante ŝanĝon en la sentiveco de ĉi tiu sistemo. Alia konsidero estas, ke D1-agado povas agi por moduli aktivecon ĉe D2-receptoroj (Nolan et al, 2007; Ruskin et al, 1999; Walters et al, 1987). La aktualaj datumoj sugestas, ke la rilatumo de D1 al D2-receptoroj ŝanĝiĝas dum la kurso de sindeteno kaj povas tiel ŝanĝi la efikecon de ĉi tiu modulado.
Kvankam, male al la aktualaj datumoj, estis raportoj pri malpliigitaj niveloj de D1-receptoroj post kronika kokain-memadministrado (moore et al, 1998a), ekzistas konsiderindaj diferencoj inter ĉi tiuj studoj, kiel dozo kaj longo de ekspozicio al kokaino, totala konsumado, kaj komparaj kontrolgrupoj. Kunigitaj, do, konverĝaj evidentaĵoj forte indikas, ke la sistemo D1 estas en konsiderinda fluo post retiriĝo de kronika administrado de kokaino.
Transportiloj de Dopamina Ŝanĝo
La trovoj de altaj koncentriĝoj de DAT trans la striato de koka-elmontritaj bestoj post ĉeso de drogokuracado etendas tiujn de niaj antaŭaj studoj montrante pliigitajn nivelojn de DAT-ligaj retejoj, kiuj akompanas kokainan memadministradon en nehomaj primatoj. La aktualaj datumoj montras, ke ĉi tiu malregulado persistas dum la komencaj fazoj de sindeteno. Plue, ili sugestas, ke reakiro al kontrolaj niveloj sekvas relative longan kurson (ĝis 90-tagoj en la nuna studo). En niaj antaŭaj studoj, ni montris, ke kvankam komence limigitaj al plejparte ventraj strataj regionoj, ŝanĝoj en la denseco de DAT-ligaj lokoj ekspansiiĝis por ampleksi pli da dorsajn kaj rostrajn porciojn de la striatum kun pli longaj periodoj de ekspozicio al kokaino (Letchworth et al, 2001; Porrino et al, 2004). En la aktuala studo la reveno al kontrolaj niveloj de DAT-koncentriĝoj dum sindeteno ŝajnis esti pli granda kaj pli rapida en la dorsa striatumo ol en la ventrala striato, kaj tiel ŝajnis sekvi inversan anatomian trajektorion al la aranĝo de efikoj induktitaj de kronika ekspozicio al kokaino. .
La aktualaj datumoj ankaŭ konformas al raportoj pri uzantoj de homa kokaino (iom et al, 1999; Maliso et al, 1998; Mash et al, 2002; Staley et al, 1994) kiuj montris altajn nivelojn de ligado al DAT-lokoj en la striato kompare al kontroloj, kun la plej markataj kreskoj situantaj en la ventra striato. Lastatempe, ĉi tiuj altoj estas montritaj akompanataj de signifaj malpliiĝoj de vezikala monoamina transportilo 2 (VMAT2) liganta (iom et al, 2003), sugestanta pri fakta perdo de DA-neŭronoj. La aŭtoroj konkludis, ke la altigita DAT probable probable rekte pro kompensa respondo al la farmakologia blokado de kokaino, dum la malpliiĝoj de VMAT2 pli probable reflektis entute ŝanĝojn en DA-metabolo, rezultigante hipodopaminergian funkcion.
Oni raportis ke toksomaniuloj al homa kokaino malpliigis DA-koncentriĝojn en la ventrala striatumo, mezurita kun PET, en respondo al metilfenidata defio kompare kun sanaj kontroloj (Volkow et al, 1997). Ĵus, martinez et al (2007) raportis, ke uzantoj de kokaino havis malakceptan respondon al anfetamina defio en la ventrala striatumo kaj putameno. Plue, ĉi tiu malpliiĝo de DA-liberigita de amfetaminoj estis korelaciita kun la elekto por kokaino en apartaj memadministraj kunsidoj, tiel ke tiuj uzantoj kun la plej malalta grado de liberigo de DA en respondo al amfetamino plej verŝajne elektis kokainon super alternativa plifortigilo (martinez et al, 2007). Lastatempaj studoj pri ŝnuraj modeloj de kokaino subtenas ĉi tiun ideon ankaŭ. Mateo et al (2005)Ekzemple, raportis, ke ekspozicio al kronika mem-administrado de kokaino estas asociita kun ŝanĝoj en DAT-funkcio. Ĉi tiuj enketistoj montris, ke la rekta DA-konsumado pliiĝis, rezultigante pli rapidan forigon de sinaptika DA, kaj tial, malpliigis bazajn nivelojn de eksterĉela DA, aŭ hipodopaminergian staton. Tiel, estas verŝajne, ke la pliigitaj DAT-koncentriĝoj post retiriĝo de kronika kokain-memadministrado observita en la nuna studo reprezentas kompensan respondon, rezultigante pli malaltajn bazajn nivelojn de DA eksterĉela.
Ŝanĝoj de D2-Ricevilo
Unu el la rezultoj de la aktuala studo estis, ke la koncentriĝoj de D2-receptor-niveloj revenis por kontroli valorojn post 30-tagoj da sindeteno, kompare kun la signifaj malkreskoj observitaj ĉe bestoj kun neniu forperioda periodo (Nader et al, 2002). Kontraste al la nuna enketo, homaj figuradaj studoj ĝenerale trovis, ke D2-receptor-niveloj estas pli malaltaj ol tiuj de kontroloj post plilongigita sindeteno de kronika kokaina ekspozicio (martinez et al, 2004; Volkow et al, 1993). Eblaj klarigoj pri la diferencoj inter ĉi tiuj homaj studoj kaj la nuna homa primata enketo inkluzivas diferencojn en la mastro kaj daŭro de konsumado de kokaino, same kiel la eblecon de antaŭekzistantaj pli malaltaj niveloj de D2-receptoroj en homaj toksomaniuloj.
Konsentite kun ĉi-lasta ideo, estas evidenteco por sugesti, ke pli malaltaj bazaj niveloj de D2-receptoroj ĉe sanaj homoj antaŭdiras pliigitan plifortigan efikecon de stimuliloj kiel ekz. metilfenidatoe (Volkow et al, 1999), kaj simile en simioj, bazniveloj de D2-receptoroj antaŭdiris la inklinon al mem-administri kokainon (Morgan et al, 2002; Nader et al, 2006). Konforme al la paralela naturo de ĉi tiuj trovoj inter specioj, ambaŭ homaj (Volkow et al, 1993) kaj nehumana primato (Nader et al, 2006) figuradaj studoj pruvis pli malaltajn nivelojn de disponebla ricevilo de D2 post abstinado de kokaina ekspozicio. Notinde, la horaro de kokain-memadministrado en ĉi-lasta eksperimento (Nader et al, 2006) estis simila al la horaro de plifortigo uzita en ĉi tiu studo. Tiel, la malsimilaj rezultoj de ĉi tiuj du studoj verŝajne ne okazis pro metodikaj diferencoj, kiel la horaro de plifortigo aŭ akumula konsumado dum kokaina memadministrado.
Pli probabla klarigo implikas la funkcian dinamikon de la DA-sistemo. Mezuroj pri la havebleco de riceviloj de D2 kun PET estis priskribitaj kiel "funkciaj" ĉar la signalo rilatas al la kvanto de proteino (ĉi-kaze densecoj de riceviloj de D2) kaj la niveloj de cirkla neurotransmisilo. (vidu Laruelle, 2000; Nader kaj Czoty, 2008 por plua diskuto). MiKontraŭe, la aŭtoradiografio de la riceviloj ne kontrolas cirkulante niveloj de DA. Tiel, la nuna studo, kune kun nia pli frua laboro, sugestas, ke D2-receptor-densecoj malpliigas per kokain-memadministrado, sed la receptoro-niveloj ŝajnas resaniĝi dum sindeteno. En simile farita studo pri PET (Nader et al, 2006), resaniĝo estis rimarkita en tri el kvin simioj. La nunaj trovoj sugestas, ke ĉi tiuj simioj probable ne diferencis en D2-receptor-densecoj, sed eble la respondeco de la DA-sistemo (t.e. niveloj de cirkuli DA dum sindeteno) diferencis inter subjektoj "rekuperitaj" kaj "ne malkovritaj".
Limigoj
Unu grava limigo de la aktualaj studoj estas, ke niaj studoj ne povas trakti la funkciojn de la D1 kaj D2-receptoroj aŭ de la DAT. Prefere ni ekzamenis nur la ŝanĝojn en la denseco de ricevilaj proteinoj. Kvankam la rezultoj havas implicojn por la eblaj roloj de ĉi tiuj sistemoj, pliaj studoj estos bezonataj por lumigi la kondutajn konsekvencojn de la ŝanĝoj montritaj ĉi tie. Alia limigo de la nuna studo estas, ke autoradiografiaj ligandoj ofte malsukcesas disiĝi inter intracelulaj kaj citoplasmaj lokoj de iliaj celoj, ĉar radiomarkitaj antagonistoj ofte membranoj permeables. iom et al (2002) pruvis, ke kronika ekspozicio al kokaino kondukas al signifa regregado de DAT ĉe la membrana surfaco, samtempe kun malpliigo de la intracelula DAT-koncentriĝo, en transfektitaj ĉeloj. Lastatempe, Samuvel et al (2008) raportis similan trovon en rat-striataj sinaptosomaj preparoj. Ĉi tiuj rezultoj sugestas, ke la ŝanĝoj en la DAT-distribuo observita en la nuna studo povus reprezenti ŝanĝojn ĉe la membrana surfaco, anstataŭ intracelulaj lokoj.
Fine, oni devas doni singardecon al la interpretado de la studoj pri plilongigita (90 tagoj) sindeteno, ĉar ĉi tiuj trovoj baziĝis sur relative malgranda grupo de bestoj (N=3). Malgraŭ la malgranda nombro de subjektoj, la datumoj akiritaj de ĉi tiu grupo estis sufiĉe konsekvencaj, kiel oni povas vidi en disa intrigo montrita en figuro 2. La koncentriĝo de D1-receptor-ligaj lokoj trans la striatum montris malmultan variablon ene de grupoj, sugestante la fidindecon de ĉi tiuj trovoj. Simila konsistenco ankaŭ estis evidenta en datumoj de D2-ricevilo kaj DAT-ligaj provoj. Kvankam oni devas zorgi, ĉi tiuj datumoj forte sugestas, ke restarigo de koncentriĝoj de DAT kaj DA-riceviloj ene de la striato povas okazi kun daŭrigita sindeteno.
Figuro 2.
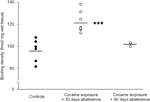 Kunligaj densecoj de D1-receptoroj por individuaj bestoj averaĝis tra la tuta striatum post aŭ nutra plifortigo (kontroloj), aŭ 30 aŭ 90-tagoj sindetenas de kronika kokain-memadministrado. Rimedoj por grupoj estas bildigitaj per nigraj stangoj, ***p<0.001 kompare kun manĝaĵoj plifortigitaj.
Kunligaj densecoj de D1-receptoroj por individuaj bestoj averaĝis tra la tuta striatum post aŭ nutra plifortigo (kontroloj), aŭ 30 aŭ 90-tagoj sindetenas de kronika kokain-memadministrado. Rimedoj por grupoj estas bildigitaj per nigraj stangoj, ***p<0.001 kompare kun manĝaĵoj plifortigitaj.
KONKLUDOJ
Konklude, ekspozicio al kokain-memadministrado produktis gravajn ŝanĝojn en la regulado de DA-sistemoj, kiuj persistis dum la fruaj fazoj (unuaj 30-tagoj) de sindeteno. Ĉi tio estis plej evidenta en la regulado de la koncentriĝo de D1-riceviloj kaj la DAT, ambaŭ koncerne ilian grandon de ŝanĝoj kaj ilia topografia amplekso. En kontrasto, estis evidenteco por normaligo kun pli longaj daŭroj de sindeteno de kokainekspozicio, ĉar la koncentriĝoj de DAT, D1, kaj D2-receptoroj post 90-tagoj de sindeteno ne diferencis de tiuj de ne-drogaj kontroloj. TTamen, sistemoj tamen ne nepre sekvas la saman tempoperiodon de resaniĝo, sugestante, ke verŝajne estas iu malstabileco en la regulado de DA-niveloj, precipe frue en sindeteno. Ĉi tiu dopaminergia malreguligo povas efiki la efikecon de iu ajn ebla farmakoterapio administrita al abstinaj drogemuloj, aparte se la medikamento dependas de la DA-sistemo por ĝia mekanismo de agado.
Notoj
Diskuto/CONFLICTO DE INTERESTO
La aŭtoroj havas neniun konflikton de intereso por diskonigi.
Referencoj
- Alleweireldt AT, Weber SM, Kirschner KF, Bullock BL, Neisewander JL (2002). Blokado aŭ stimulo de dopaminaj receptoroj al D1 mildigas la reintegriĝon de estingita serĉado de kokaino en ratoj. Psikofarmacologio (Berl) 159: 284-293. | artikolo | PubMed | ChemPort |
- Beveridge TJ, Smith HR, Daunais JB, Nader MA, Porrino LJ (2006). Kronika memstara koko estas asociita kun ŝanĝita funkcia agado en la tempaj loboj de nehomaj primatoj. Eur J Neurosci 23: 3109-3118. | artikolo | PubMed |
- Canfield DR, Spealman RD, Kaufman MJ, Madras BK (1990). Aŭtradiografia lokalizado de kokainaj ligaj retejoj per [3H] CFT ([3H] WIN 35428) en la simia cerbo. Synapse 6: 189-195. | artikolo | PubMed | ChemPort |
- De Vries TJ, Schoffelmeer AN, Binnekade R, Vanderschuren LJ (1999). Dopaminergiaj mekanismoj mediante la instigon serĉi kokainon kaj heroinon post longtempa retiro de IV-drogadministrado. Psikofarmacologio (Berl) 143: 254-260. | artikolo | PubMed | ChemPort |
- Farfel GM, Kleven MS, Woolverton WL, Seiden LS, Perry BD (1992). Efikoj de ripetaj injektoj de kokaino sur katenoj de katenolaminoj, ligiloj de dopamina transportilo kaj konduto en simio de ruso. Brain Res 578: 235-243. | artikolo | PubMed | ChemPort |
- Graham J, Porrino LJ (1995). Neŭroatomikaj Substratoj de Kokaino-Administrado. CRC: Boca Raton, FL.
- Hemby SE, Co C, Koves TR, Smith JE, Dworkin SI (1997). Diferencoj en eksterĉelaj dopaminaj koncentriĝoj en la kerno akciĝas dum responda dependa kaj responda sendependa kokainadministrado en la rato. Psikofarmacologio (Berl) 133: 7-16. | artikolo | PubMed | ChemPort |
- Henry DJ, Blanka FJ (1991). Ripeta kokainadministrado kaŭzas konstantan plibonigon de D1-dopamina-receptoro-sentemo ene de la rato-kerno acumbens. J Pharmacol Floto Ther 258: 882-890. | PubMed | ChemPort |
- Jacobsen LK, Staley JK, Malison RT, Zoghbi SS, Seibyl JP, Kosten TR et al (2000) Pli alta centra serotonina transportilo liganta haveblecon en akre abstinaj kokain-dependaj pacientoj. Am J Psikiatrio 157: 1134-1140. | artikolo | PubMed | ISI | ChemPort |
- Khroyan TV, Barrett-Larimore RL, Rowlett JK, Spealman RD (2000). Dopamina D1- kaj D2-similaj ricevilmekanismoj en relivero al kokain-serĉanta konduto: efikoj de selektemaj antagonistoj kaj agonistoj. J Pharmacol Floto Ther 294: 680-687. | PubMed | ISI | ChemPort |
- Khroyan TV, Platt DM, Rowlett JK, Spealman RD (2003). Atenuado de reapero al kokaino serĉanta doponajn D1-agonistojn kaj antagonistojn en nehomaj primatoj. Psikofarmacologio (Berl) 168: 124-131. | artikolo | PubMed | ChemPort |
- Laruelle M (2000). Bildigo kun sinapsa neŭrotransmisio kun en vivo ligaj konkurencaj teknikoj: kritika revizio. J Cereb Blood Flow Metab 20: 423-451. | artikolo | PubMed | ISI | ChemPort |
- Letchworth SR, Nader MA, Smith HR, Friedman DP, Porrino LJ (2001). Progreso de ŝanĝoj en dopamina transportilo liganta lokan densecon kiel rezulto de kokain-memadministrado en rhesus simioj. J Neurosci 21: 2799-2807. | PubMed | ISI | ChemPort |
- Lidow MS, Goldman-Rakic PS, Gallager DW, Rakic P (1991). Distribuado de dopaminergiaj riceviloj en la primata cerba kortekso: kvanta aŭradiografia analizo uzante [3H] raclopride, [3H] spiperone kaj [3H] SCH 23390. Neurokienco 40: 657-671. | artikolo | PubMed | ChemPort |
- Malgranda KY, Elmer LW, Zhong H, Scheys JO, Zhang L (2002). Kokaino-indukto de dopamina transportilo al la plasmembrano. Mol Pharmacol 61: 436-445. | artikolo | PubMed | ISI | ChemPort |
- Malgranda KY, Krolewski DM, Zhang L, Cassin BJ (2003). Perdo de striat-vezikala monoamina transporta proteino (VMAT2) en homaj kokainaj uzantoj. Am J Psikiatrio 160: 47-55. | artikolo | PubMed |
- Malgranda KY, Zhang L, Desmond T, Frey KA, Dalack GW, Cassin BJ (1999). Striataj dopaminergiaj anormalecoj en homaj kokainaj uzantoj. Am J Psikiatrio 156: 238-245. | PubMed | ChemPort |
- Malison RT, Best SE, van Dyck CH, McCance EF, Wallace EA, Laruelle M et al (1998) Pliigitaj striatalaj dopaminaj transportiloj dum akra kokaino-sindeteno laŭ mezuro de [123I] beta-CIT SPECT. Am J Psikiatrio 155: 832-834. | PubMed | ChemPort |
- Martinez D, Broft A, Foltin RW, Slifstein M, Hwang DR, Huang Y et al (2004) Kokineca dependeco kaj d2-receptoro havebleco en la funkciaj subdividoj de la striatumo: rilato kun kokain-serĉanta konduto. Neuropsychofarmacology 29: 1190-1202. | artikolo | PubMed | ISI | ChemPort |
- Martinez D, Narendran R, Foltin RW, Slifstein M, Hwang DR, Broft A et al (2007) Dopeta liberigo de amfetamino: markite malakra en dependeco de kokaino kaj prognozo de la elekto mem-administri kokainon. Am J Psikiatrio 164: 622-629. | artikolo | PubMed | ISI |
- Mash DC, Paŭlo J, Ouyang Q, Hearn WL, Izenwasser S (2002). Dopamina transportfunkcio altiĝas ĉe uzantoj de kokaino. J Neurochem 81: 292-300. | artikolo | PubMed | ChemPort |
- Mateo Y, Mankas CM, Morgan D, Roberts DC, Jones SR (2005). Malpligrandigita dopamina fina funkcio kaj nesentemo al kokaino sekve de kokina memregado kaj deprivado. Neuropsychofarmacology 30: 1455-1463. | artikolo | PubMed | ISI | ChemPort |
- Melega WP, Jorgensen MJ, Lacan G, Way BM, Pham J, Morton G et al (2008) Longtempa administrado de metamfetamino en la verve simio modeligas aspektojn de homa malkovro: cerba neŭrotikeco kaj kondutaj profiloj. Neuropsychofarmacology 33: 1441-1452. | artikolo | PubMed | ChemPort |
- Moore RJ, Vinsant SL, Nader MA, Porrino LJ, Friedman DP (1998a). Efiko de kokain-memadministrado sur striatalaj dopaminaj D1-receptoroj en rhesus-simioj. Synapse 28: 1-9. | artikolo | PubMed | ChemPort |
- Moore RJ, Vinsant SL, Nader MA, Porrino LJ, Friedman DP (1998b). Efiko de koka-memadministrado sur dopaminaj D2-receptoroj en rhesus-simioj. Synapse 30: 88-96. | artikolo | PubMed | ChemPort |
- Morgan D, Grant KA, Gage HD, Mach RH, Kaplan JR, Prioleau O et al (2002) Socia regado en simioj: dopaminaj D2-riceviloj kaj kokain-memadministrado. Nat Neurosci 5: 169-174. | artikolo | PubMed | ISI | ChemPort |
- Nader MA, Czoty PW (2008). Cerbaj bildigoj en nehomaj primatoj: komprenoj pri drogmanio. ILAR J 49: 89-102. | PubMed | ChemPort |
- Nader MA, Daunais JB, Moore T, Nader SH, Moore RJ, Smith HR et al (2002) Efikoj de kokaina memadministrado sur striatalaj dopaminaj sistemoj en rhesus simioj: komenca kaj kronika ekspozicio. Neuropsychofarmacology 27: 35-46. | artikolo | PubMed | ISI | ChemPort |
- Nader MA, Morgan D, Gage HD, Nader SH, Calhoun TL, Buchheimer N et al (2006) PET-bildigo de dopaminaj D2-riceviloj dum kronika mem-administrado de kokaino en simioj. Nat Neurosci 9: 1050-1056. | artikolo | PubMed | ChemPort |
- Nolan EB, Harrison LM, Lahoste GJ, Ruskin DN (2007). Konduta sinergismo inter D (1) kaj D (2) dopamaj riceviloj en musoj ne dependas de breĉaj juntoj. Synapse 61: 279-287. | artikolo | PubMed | ChemPort |
- Porrino LJ, Lyons D, Miller MD, Smith HR, Friedman DP, Daunais JB et al (2002) Metabola mapado de la efikoj de kokaino dum la komencaj fazoj de memadministrado en la nehumana primato. J Neurosci 22: 7687-7694. | PubMed | ChemPort |
- Porrino LJ, Lyons D, Smith HR, Daunais JB, Nader MA (2004). Kokaino-memadministrado produktas progreseman implikiĝon de limuzikaj, asociitaj, kaj sensimotoraj striaj domajnoj. J Neurosci 24: 3554-3562. | artikolo | PubMed | ISI | ChemPort |
- Ruskin DN, Bergstrom DA, Walters JR (1999). Plursekundaj osciloj en pafo-rapideco en la globus pallidus: sinergia modulado per D1 kaj D2-dopaminaj riceviloj. J Pharmacol Floto Ther 290: 1493-1501. | PubMed | ISI | ChemPort |
- Samuvel DJ, Jayanthi LD, Manohar S, Kaliyaperumal K, Vidu RE, Ramamoorthy S (2008). Malreguligo de trafikado de dopamina transportilo kaj funkcio post sindeteno de kokaina memadministrado en ratoj: evidenteco por diferenca reguligo en kaŭdaj putamenoj kaj kerno accumbens. J Pharmacol Floto Ther 325: 293-301. | artikolo | PubMed | ChemPort |
- Schmidt HD, Anderson SM, Pierce RC (2006). Stimuli D1-similajn aŭ D2-dopaminajn receptorojn en la ŝelo, sed ne la kerno, de la kerno accumbens reinstaras kokain-serĉantan konduton en la rato. Eur J Neurosci 23: 219-228. | artikolo | PubMed |
- Mem DW, Barnhart WJ, Lehman DA, Nestler EJ (1996). Kontraŭa modulado de kokain-serĉanta konduto de D1- kaj D2-similaj dopaminaj agonistoj. scienco 271: 1586-1589. | artikolo | PubMed | ISI | ChemPort |
- Staley JK, Hearn WL, Ruttenber AJ, Wetli CV, Mash DC (1994). Lokoj pri agnosko de kokaino kun alta afineco sur la transportilo de dopamino estas levitaj en mortigaj viktimoj de kokaina superdozo. J Pharmacol Floto Ther 271: 1678-1685. | PubMed | ChemPort |
- Volkow ND, Fowler JS, Wang GJ, Hitzemann R, Logan J, Schlyer DJ et al (1993) Malpliiĝanta dopamina D2-receptoro estas asociita kun reduktita frontala metabolo en kokainaj misuzantoj. Synapse 14: 169-177. | artikolo | PubMed | ISI | ChemPort |
- Volkow ND, Wang GJ, Fowler JS, Logan J, Gatley SJ, Gifford A et al (1999) Antaŭdiro de plifortigado de respondoj al psikostimuliloj en homoj per cerba dopamina D2-receptoro-niveloj. Am J Psikiatrio 156: 1440-1443. | PubMed | ISI | ChemPort |
- Volkow ND, Wang GJ, Fowler JS, Logan J, Gatley SJ, Hitzemann R et al (1997) Malpliiĝis striatala dopaminergia respondo en detoxigitaj kokainaj dependaj subjektoj. naturo 386: 830-833. | artikolo | PubMed | ISI | ChemPort |
- Walters JR, Bergstrom DA, Carlson JH, Chase TN, Braun AR (1987). D1 dopamina ricevilo aktivigo bezonata por postsynaptic esprimo de D2 agonistaj efikoj. scienco 236: 719-722. | artikolo | PubMed | ChemPort |
- Weiss F, Martin-Fardon R, Ciccocioppo R, Kerr TM, Smith DL, Ben-Shahar O (2001). Daŭrba rezisto al estingo de kokain-serĉanta konduto induktita de drogaj raportoj. Neuropsychofarmacology 25: 361-372. | artikolo | PubMed | ISI | ChemPort |
- Wong DF, Kuwabara H, Schretlen DJ, Bonson KR, Zhou Y, Nandi A et al (2006) Pliigita okupado de dopaminaj riceviloj en homa striatumo dum ree-eliritaj kokainaj avidoj. Neuropsychofarmacology 31: 2716-2727. | artikolo | PubMed | ChemPort |