نظرات YBOP: اثر Coolidge پشت قدرت پورنو اینترنت است. اثر Coolidge پدیده ای است که در گونه های پستانداران مشاهده می شود که به موجب آن نرها (و به میزان کمتری زن) در صورت معرفی به شرکای جنسی پذیرنده جدید ، حتی پس از امتناع از رابطه جنسی از همسر قبلی یا هنوز در دسترس ، علاقه جنسی تجدید می کنند. تازگی جنسی با افزایش هیجان ناشی از دوپامین بالاتر ، این عادت را غلبه می کند. یک جریان مداوم از تازگی ها ، چیزی است که باعث می شود پورنو در اینترنت با پورنو گذشته بسیار متفاوت باشد.
- دنیس اف. فیورینو ،
- آرین کوری ، و
- آنتونی جی فیلیپس
- مجله علوم اعصاب، 15 ژوئن 1997 ، 17 (12): 4849-4855؛
چکیده
اثر کوليج توصيف بازتوليد رفتار جنسي در يک جنس "تحريک پذير جنسي" در واکنش به يک مامور دريافت کننده جديد است. با توجه به نقش سیستم دوپامین مازولیمبیک (DA) در شروع و حفظ رفتار انگیزشی، میکرو دیالیز برای نظارت بر انتقال DA DA هسته ای در طی کوپولیت، سرسختگی جنسی و تکرار رفتار جنسی مورد استفاده قرار گرفت. با توافق با گزارش های قبلی، ارائه یک زن عجیب و غریب در پشت صفحه نمایش و copulation با افزایش قابل توجهی در جریان NAC DA همراه بود. بازگشت غلظت NAC DA به مقادیر ابتدایی همزمان با دوره سفتی جنسی بود، گرچه غلظت متابولیت های DA، دی هیدروکسی فنیل اسیدهای اسید و اسید هموانیونیک، همچنان بالا بود. ارائه یک زن گیرنده رمان در پشت یک صفحه، باعث افزایش کمی در NAC DA شد که به طور قابل ملاحظه ای در طی نسل جدید با زن رمانی افزوده شد. داده های حاضر نشان می دهد که ویژگی های تحریک کننده یک زن گیرنده جدید ممکن است موجب افزایش انتقال NAC DA در یک موش صحرایی مرد نشود و این به نوبه خود ممکن است به تکرار رفتارهای جنسی مرتبط باشد.
معرفی
یک موش نر که به سیری می پیوندد، می تواند مجددا به هم متصل شود، اگر زن اولیه با یک زن گیرنده جدید جایگزین شود. این به عنوان اثر کولیج شناخته شده است و در تعدادی از گونه های پستانداران مشاهده شده است (ویلسون و همکاران، 1963) عوامل عمومی مانند خستگی یا افسردگی حرکتی برای توضیح حالت ظاهری اشباع جنسی کافی نیست، زیرا محرک های یک زن رمان هنوز هم می تواند باعث ایجاد کوپولیت شود. تندرستی جنسی نیز می تواند به طور قابل توجهی توسط داروهای مختلفی که می توانند بر روی سیستم های مختلف انتقال دهنده های عصبی عمل کنند، به طور قابل توجهی توسط داروهای "معکوس" شوند. این داروها شامل یوهیمبین، 8-OH-DPAT (رودریگز منزو و فرناندز گاستی، 1994, 1995a)، نالاکون (Pfaus و Gorzalka، 1987; رودریگز منزو و فرناندز گاستی، 1995a,b) و آپومرفین (مس و همکاران، 1995c) اگرچه اثر محیطی این داروها قابل رد نیست (مثلا اثرات آدرنرژیک بر عملکرد نعوظ)، تأثیرات بر مکانیزم های مرکزی زیر سوزنی جنسی براساس آزمایش های غلط ناکارآمد مرکزی انتخابی (رودریگز منزو و فرناندز گاستی، 1995a) و آزمایش های میکرو دیالیز که متابولیسم دوپامینرژیک را در ناحیه preoptic medial (مس و همکاران، 1995a,b).
با توجه به اینکه مکانیزم های مرکزی ممکن است به تسریع رفتار جنسی در خواص کولیدژ بینجامد، احتمالا کاندیدای سیستم دوپامین مازولیمبیک (DA) است که از منطقه تکتونمان شکمی به NAC طراحی شده است. Mesolimbic DA به نظر می رسد به عنوان یک مدولاتور اصلی در فرایندهای پیچیده ادغام شده است که شامل ارزیابی محرک های محرک مانند نشانه های یک زن گیرنده جنسی و سازماندهی رفتارهای هدایت شده هدفمند، از جمله copulation (فیبر و فیلیپس، 1986; بلکبرن و همکاران، 1992; فیلیپس و همکاران، 1992; LeMoal، 1995; سالامون، 1996).
هرچند نورونهای میانه مغزی DA به پاداش های اولیه و پیش بینی پاداشی پاسخ می دهند، محرک های محرک جدید یا غیر قابل پیش بینی محیطی باعث شدید فعالیت های عصبی در جلسات تمرین مکرر (فابر و همکاران، 1983; شولتز، 1992; Mirenowicz و Schultz، 1994) شواهد زیادی وجود دارد که از نقش مهم تسهیل کننده برای mesolimbic DA در شروع و حفظ رفتار جنسی جنین (Pfaus و Everitt، 1995)، و تعدادی از مطالعات میکروبیالیز گزارش شده در جریان ناپایداری NAC DA در طی مراحل اشتعال و پرخاشگری رفتار جنسی مردان (Pfaus و همکاران، 1990; Pleim et al.، 1990; Damsma et al.، 1992; Wenkstern و همکاران، 1993; Fumero et al.، 1994; Mas و همکاران، 1995b) با این حال، اطلاعات نسبتا کمی در مورد ارتباطات عصبی شیمیایی خارش جنسی و تجدید رفتار جنسی وجود دارد. استفاده از در داخل بدن Microdialysis برای نظارت بر انتقال Mesolimbic DA در طول اثر کولیج یک فرصت منحصر به فرد برای بررسی نقش NAC DA در کوپلیمر، سرسختگی جنسی و تجدید نسل کوپال فراهم می کند.
یک آزمایش میکرو دیالیز برای تعیین موارد زیر انجام شد: (1) که آیا شروع سوزش جنسی همراه با بازگشت غلظت DA DA در NAC به مقادیر preopulation و یا پایینتر است و (2) که آیا بازسازی رفتار مخدر در یک " موش صحرایی مردانه با یک زن مبتلا به سرطان روده همراه با افزایش میزان NAC DAP می باشد.
مواد و روش ها
فاعل، موضوع. موش صحرایی Sprague Dawley که از مرکز مراقبت از حیوانات (در دانشگاه بریتیش کلمبیا) و موشهای Long-Evans خانم، که از چارلز رود کانادا (سنت کنستانت، کبک، کانادا) به دست آمده بودند، در قفس های مشبک قرار داشتند (18 × 25 × 65 سانتی متر؛ پنج در هر قفس) در اتاق های جداگانه مستعمره. اتاق های کلنی در دماي ~20 ° C در یک چرخه نور / تاریکی 12 معکوس نگهداری می شود. موش دارای دسترسی نامحدود به مواد غذایی (Purina Rat Chow) و آب بود.
جراحی و تست رفتاری قبل از میکرو دیالیز مغز.موش های صحرایی دو ماه بعد از آزمایش 4 به مدت دو هفته تحت جراحی بیهوشی هالوتان (Fluothane، Ayerst Laboratories) تخمدان شدند. حساسیت جنسی در زنان تحریک کننده به واسطه تزریق زیر جلدی استریایید بنزوات (10 μg) و پروژسترون (500 μg)، 48 و 4 hr قبل از هر جلسه آزمون القا شده است. موش های نر برای رفتار جنسی در دو مورد 4 از هم جدا شدند، در اتاق های پلکسی گلاس (35 × 35 × 40 سانتی متر) با کف های سیم مش نمایش داده شد. موشهای صحرایی نر و ماده ای که در معرض یک معیار عملکرد قرار داشتند و شامل تحرک درون 5 دقیقه ظاهر شدن زن و انزال در طی 15 دقیقه از اولویت بندی اولیه، در طی دو آزمایش غربالگری، توسط کانول های راهنمای پروب میکرو دیالیز گذاشته شد.
موش صحرایی نر (n = 5) با استفاده از کراتین هیدروکلراید (100 mg / kg، Ip) و Xylazine (10 mg / kg، ip) قبل از عمل جراحی Stereotaxic بیهوش شد. کانال های کنترل میکرو دیالیس (اندازه 19) به صورت دو طرفه بر روی NAC (مختصات از برگی: قدامی، + 1.7 میلیمتر، متوسط، ± 1.1 میلیمتر، ونتالال، -1.0 میلیمتر، جمجمه صاف) کاشت و به جمجمه با اکریلیک دندانی و پیچ گوشتی ایمپلنت کانولای راهنمای دو طرفه برای به حداکثر رساندن فرصت برای یک آزمایش موفق microdialysis استفاده شد. خوشبختانه، در آزمایش حاضر تنها یک کانول برای هر موش مورد نیاز بود. موش های صحرایی نر به صورت جداگانه در قفس های بزرگ پلاستیکی با بستر زردچوبه برای بقیه آزمایش قرار گرفتند. یک هفته بعد از عمل، موش ها برای رفتار جنسی آزمایش شدند. در طول این دوره آموزشی، محفظه تست با یک صفحه نمایش پلاستیکی کشویی مجهز شده بود که اتاق را به بخش های بزرگ و کوچک تقسیم می کرد. موش های نر به داخل محفظه بزرگ وارد شدند و 15 دقیقه بعد، یک زن در پشت صفحه قرار گرفت. پس از یک دوره آماده سازی 15 دقیقه، روی صفحه نمایش برداشته شد و موش ها مجددا برای 30 دقیقه تقسیم شدند. سه جلسه آموزشی انجام شد، هر یک از هر 4 د. تمام موش ها در هر جلسه به معیار عملکرد رسیدند.
آزمایش اثر کولیج موش ها به صورت یک طرفه با پروب های میکرو دیالیز 12-18 قبل از آزمایش اثر Coolidge و در محفظه بزرگ اتاق آزمایش با دسترسی آزاد به غذا و آب گذاشته شدند. در صبح روز آزمایش، نمونه های میکرو دیالیز هر دقیقه 15 جمع آوری شد. آزمایش شامل هفت مرحله متوالی زیر بود: (1) پایه (حداقل 60 دقیقه)؛ (2) 1 زن در پشت صفحه نمایش (15 دقیقه)؛ (3) با زن 1 تا زمانی که دوره 30 دقیقه بدون کوه عبور کرد؛ (4) مجددا 1 زن در پشت صفحه نمایش (15 دقیقه)؛ (5) دسترسی به زن 1 برای یک دوره زمانی 15 در صورتی که نصب وجود نداشته باشد (در صورت نصب، این مرحله به عنوان فاز 3 رفتار می شود)؛ (6) معرفی زن 2 پشت صفحه نمایش (15 دقیقه)؛ 7) با زن 2 برای 60 دقیقه.
رفتار تحت نور کم با استفاده از یک سیستم ویدئویی JVC فیلم برداری شد و در یک مانیتور ویدئویی که در خارج از اتاق آزمایش قرار دارد دیده می شود. معیارهای استاندارد رفتار جنسی با استفاده از یک کامپیوتر و نرم افزار مناسب (هولمز و همکاران، 1987).
بعد از آزمایش میکرو دیالیز، حیوانات بیش از حد هیدرات کلرال را دریافت کردند و به صورت intracardially با سالین و فرمالین (4٪) تزریق کردند. مغزها برش داده شده و یخ زدگی شدند، و پس از آن، بخش های کرونالی با کرزی بنفش رنگ آمیزی شدند تا محل قرارگیری پروب های میکروبیالیز را تعیین کنند. فقط برای مواجهه با پروب در NAC برای بررسی های رفتاری و عصبی مورد استفاده قرار گرفت.
میکروسدیالیز و تشخیص الکتروشیمیایی HPLC. پروبهای میکروبیالیز در طراحی با غشای فیبر توخالی نیمه پراکنده (در معرض غشاء 2 میلی متر، قطر بیرونی 340 μm، قطع کردن وزن مولکولی 65000، Filtral 12، Hospal) در انتهای دالال قرار گرفتند. Probes در 1.0 μl / min با یک راه حل رینگر اصلاح شده (0.01 مایع بافر فسفات سدیم، pH 7.4، 1.3 mmCaCl2، 3.0 میلی متر KCl، 1.0 mmMgCl2، 147 میلی متر NaCl) با استفاده از یک سرنگ انژکتیو (همیلتون، رینو، NV) و یک پمپ سرنگ (مدل 22، دستگاه هاروارد، South Natick، MA). یک یراق هدایت کننده پروب میکرو دیالیز برای تضمین پروب میکرو دیالیز در داخل کانول هدایت استفاده شد. کویل فولادی، متصل به یک مفصل مایع (Instech 375s) که در بالای محفظه آزمایشگاهی نصب شده بود، برای محافظت از لوله پروب (Fiorino و همکاران، 1993).
آنالیت های میکرویدیالیزات که حاوی DA و متابولیت های آن دی اکسید فنیل اکتین اسید (DOPAC) و اسید هموانیولیک (HVA) بودند با روش کروماتوگرافی فاز معکوس (ستون فوق ستاره، بکمن، فولرتن، CA، ODS 5 μm، 15 سانتی متر، 4.6 میلی متر، قطر داخلی ) با استفاده از یک بافر سدیم استات سدیم 0.083m، pH 3.5 (5٪ متانول). غلظت آنالیت توسط تشخیص الکتروشیمیایی (EC) اندازه گیری شد. این دستگاه شامل یک پمپ Bio-Rad (ریچموند، کالیفرنیا)، یک انژکتور دوقطبی EC10W هوستون (TX)، ESA (Bedford، MA)، کولاکیم II EC و ضبط کننده دو کاناله (Kipp و Zonen، Bohemia، NY). پارامترهای آشکارساز الکتروشیمیایی عبارت بودند از: الکترود 1، + 450 mV؛ الکترود 2، -300 mV؛ و نگهبان سلول، -450 mV. پروب معمولی بازیابی می شود در شرایط in vitro و در دمای اتاق، 22٪ برای DA، 18٪ برای DOPAC و 18٪ برای HVA بود.
نتایج
رفتار
اقدامات رفتاری از آزمایش اثر کولیج در جدول ارائه شده است 1. کمبود زمان برای سوار شدن، انفجار و انزال، و همچنین فاصله زمانی پست بعد از اولین انزال، شبیه آنچه که در جلسه تمرین قبلی بود (داده ها نشان داده نمی شود). این نشان می دهد که روش میکرو دیالیز رفتار عادی جنسی را تغییر نمی دهد. توسعه سفتی جنسی، همانطور که با میانگین میانگین انزال قبل از معیار نشان داده شد (7.8 ± 0.5)، کاهش مداوم در تعداد انعقاد قبل از هر انزال و افزایش مترقی در فاصله پست جراحی (داده ها نشان داده نمی شود) ، مشابه آنچه که در مطالعات قبلی گزارش شده است (ساحل و اردن، 1956; فاولر و والن، 1961; فیشر، 1962; Bermant و همکاران، 1966; رودریگز منزو و فرناندز گاستی، 1994; مس و همکاران، 1995d) تنوع فردی با توجه به تعداد انسداد های حاصل شده با 1 زن، زمان صرف شده با زن 1 زن و تعدادی از ارائه 1 زن مورد نیاز برای رسیدن به معیار بارندگی مشاهده شد (جدول 1، پایین) بعضی موش های صحرایی 1 زن را مجددا مجددا مجددا تا مرحله 5 تکمیل کردند (n = 3) اعمال قرار دادن زن 1 در پشت صفحه و حذف پارتیشن ممکن است به عنوان نشانه های اشتباه اولیه منجر به copulation شود. همچنین لازم به ذکر است که یک معیار Satiment از 30 دقیقه بدون کوه، هر چند قبلا استفاده شده (ساحل و اردن، 1965؛ Mas و همکاران، 1995b)، خودسرانه است و تضمین نمی کند که یک موش به زمان بیشتری نیاز نداشته باشد. با این حال، تاخیر و یا روش های حذف و جایگزینی در مصلحت جدید با زن 1 (به عنوان مثال، مراحل 4 و 5) قابل اعتماد نیست.

رفتار در طول آزمایش اثر Coolidge
همه موش های صحرایی اثر کولیج را نشان دادند. فعالیت مرتبط با قرار دادن زن 2 در پشت صفحه نمایش و به خصوص حذف پارتیشن ممکن است به این نتیجه کمک کرده باشد، اما دوباره این رویدادها به اندازه کافی برای تجدید نسل قبلی در آزمایش نبود. مقایسه بین اقدامات جنسی با زنان 1 و زن 2 با استفاده از t آزمایش با اصلاح Bonferroni. با وجودی که تاخیرهای مونتاژ و تعامل در پاسخ به زن 2 به طور قابل ملاحظهای در مقایسه با زنان در اولین مقاربت با زن 1 تفاوت معنیدار نبود، به طور کلی رفتار جنسی با زن 2 کمتر قوی بود، به طوری که میزان انقباضات مؤثر (متوسط، 0.6 نسبت به 4.2؛ F = 49.86؛p <0.01) و درگیری (میانگین = 11.2 در مقابل 37.0 ؛F = 20.17؛ p <0.05) در ساعت اول تعداد نصب شده در ساعت اول با ماده 1 و 2 تفاوت معنی داری نداشت.
مهم است که توجه داشته باشیم که زنان در طول بخش خفیف آزمایش (به عنوان مثال زن 1) هنوز هم رفتارهای قوی (یعنی hopping و darting) و پذیرش (یعنی lordosis) برای مدت زمان کامل تماس با مرد .
نورو شیمی
غلظت نانومواد پایه ای DA و متابولیت های آن در میکرودیلایسات ها، به صورت میانگین ± SEM، از سه نمونه اولیه آزمایشی شامل: DA، 3.0، 0.7، DOPAC، 619.1 ± 77.7؛ و HVA، 234.2 ± 49.0 (اصلاح نشده برای بهبود پروب؛n = 5) این ارزشها نشان دهنده امتیازات پایه 100٪ است.
نقاط داده تعریف شده در رفتار، مربوط به هر مرحله از آزمایش و رایج برای هر موش، برای تجزیه و تحلیل های عصبی استفاده شد. این شامل موارد زیر می شود: (1) هفت نمونه پس از اولین معرفی زن 1، (2) چهار نمونه همراه عدم وجود رفتار مشترک با زن 1 و (3) پنج نمونه پس از ارائه زن 2. شکل 1 تغییرات غلظت DA (نمودار خط، متوسط) و متابولیت های DA (نمودار خط، بالا) رفتار موازی همپوشانی (نمودار نوار، پایین) در طول آزمایش برای اثر Coolidge.
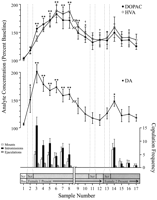
نمایش نسخه بزرگتر:
شکل. 1.
هسته accumbens همبستگی های عصبی مغز از رفتار جنسی در طول اثر کولیج. هشت نمونه اول، نقاط اطلاعاتی پیوسته را از مراحل 1 تا 3 نشان می دهد. نمونه 1 نمونه های پایه چهارم و آخرحجاری و نقوش برجسته) نمونه 2 نشان دهنده معرفی زن 1 در پشت صفحه نمایش است (SCR) بعد از 15 دقیقه، روی صفحه نمایش برداشته شد و موش ها مجددا مجروح شدند (نمونه 3-8). اینشکستن در xAxis مربوط به حذف اطلاعات از سه موش است که برای دوره های طولانی با زن اولیه مقابله می کند. 9 نمونه اخیر نیز به طور پیوسته پیوسته بودند. نمونه 9 و 10 به دوره بارگذاری فاز 3 (یعنی 30 دقیقه بدون کوه) مربوط می شود. زن 1 پس از روی صفحه نمایش (11 نمونه) دوباره دوباره وارد شد و 15 دقیقه بعد برداشته شد (نمونه 12). پس از 15 دقیقه بدون copulation، زن 2 در پشت صفحه قرار گرفت (نمونه 13). نمونه هایی از 14-17 به مقابله با زن 2 متصل می شوند. تعداد مورچه ها، درهم ریختگی ها یا انقطاعات مرتبط با هر نمونه MICM 15 دقیقه در گراف نوار پایین. داده های عصبی از نظر درصد غلظت های اولیه بیان می شود. تغییرات در NAC DA (مربع بسته)، DOPAC (دایره های بسته) و HVA (حلقه های باز) خروجی به عنوان نشان داده شده است نمودار خط. مقایسه های زیر انجام شد: نمونه اولیه 1 در مقایسه با نمونه های 2-10؛ نمونه اولیه جدید 10 در مقایسه با نمونه 11 و 12؛ نمونه اولیه جدید 12 در مقایسه با نمونه 13-17 (*p <0.05 ** p 0.01) مستقل t آزمونهای بین مقادیر پایه (نمونه 1، 10 و 12) انجام شد. برای تفاوت های قابل توجهی از اولین خط پایه (نمونه 1)، †p <0.05
تجزیه و تحلیل های تک مرحله ای یک طرفه، تکرار شده بر روی داده های مغز و اعصاب مرتبط با زن 1 (نمونه 1-12) و زن 2 (نمونه 12-17) انجام شد. پیشینی مقایسه ها با استفاده از تست مقایسه چندگانه دان (Bonferroni t) سه مقیاس اصلی زیر تهیه شده است: (1) اولیه (نمونه 1) در مقایسه با نمونه 2-10 (اولین قرار گرفتن در معرض زن 1)، (2) دومین (10) نمونه در مقایسه با نمونه 11 و 12 (دوباره به 1 زن) ، و (3) سومین پایه (نمونه 12) در مقایسه با نمونه 13-17 (قرار گرفتن در معرض زن 2).
تغییرات قابل ملاحظه ای در خروجی DA در پاسخ به زن 1 [F (11,44) = 8.48؛ p <0.001] و زن 2 [F (5,20) = 2.83؛p 0.05/1> افزایش قابل توجهی در هجوم DA هنگام حضور ماده 44 در پشت صفحه مشاهده شد (XNUMX٪ ،p <0.05 نمونه 2) در طول جمع آوری ، غلظت DA بیشتر افزایش می یابد ، رسیدن به حداکثر مقدار (+ 95؛).p <0.01) در طول اولین دوره قانونگذاری (نمونه 3). DA در طول جمع آوری بالا بود و فقط در دوره 30 دقیقه ای که هیچ گونه نصب انجام نشد ، به غلظت اولیه بازگشت. نه معرفی مجدد زن 9 در پشت صفحه نمایش (نمونه 10) و نه فرصت تعامل بدنی ، اما بدون نصب (نمونه 1) ، غلظت DA افزایش یافته نسبت به مقدار پایه دوم (نمونه 11). حضور زن 12 در پشت صفحه نمایش (نمونه 10) منجر به افزایش کمی جریان DA (2٪) از مقدار پایه سوم (نمونه 13) شد که به اهمیت آماری نرسید. جمع آوری مجدد با زن 12 منجر به افزایش قابل توجه (12٪) (p 0.05/14 <) در جريان DA در طي اولين نمونه جمع آوري (نمونه 15). اگرچه رفتار ضعیفی در سه نمونه بعدی ادامه داشت ، غلظت DA به مقادیر پایه کاهش یافت (نمونه های 17-XNUMX). مستقل t آزمایشات انجام شده بین نمونه های پایه (یعنی 1، 10 و 12) نشان داد که این مقادیر به طور قابل توجهی متفاوت نیستند.
در سه موش صحرایی که 1 زن مجددا تزریق می شود، هنگامی که زنان 1 دوباره وارد شدند، غلظت NAC DA هنگامی که زن 25 در پشت صفحه نمایش (محدوده 47-13٪) و در طی تلاقی (محدوده 37-XNUMX٪) در مقایسه با نمونه فقط قبل از دوباره سازی زن. این افزایش، با این حال، تنها زمانی رخ می دهد که رفتار جنسی شدید و منجر به انزال شود.
تغییرات کلی در DOPAC [F (11,44) = 9.57؛ p <0.001] و HVA [F (11,44) = 12.47؛ p <0] غلظت در پاسخ به زن 001 پیدا شد ، اما زن 1 نیست. غلظت متابولیت در طول ارائه زن 2 پشت صفحه (کمی 15) کمی افزایش یافت (+ 1٪ در هر دو مورد) ، اما این قابل توجه نبود از نظر آماری با این حال ، افزایش قابل توجهی در غلظت DOPAC و HVA در طول جمع آوری وجود داشت (نمونه های 2-3) ، رسیدن به حداکثر مقادیر (به ترتیب 8+ و + 80٪). p <0.01) بعد از 60 دقیقه (نمونه 6 در هر دو مورد). اگرچه غلظت متابولیت در طی دوره عدم فعالیت جنسی در پایان تماس با زن 1 (نمونه های 9 و 10) کاهش یافته است ، اما غلظت ها همچنان با توجه به سطح پایه بالا باقی مانده اند (p <0.05 در هر دو مورد). معرفی مجدد ماده 1 در پشت صفحه (نمونه 11) ، دسترسی به ماده 1 پس از حذف صفحه (نمونه 12) و معرفی ماده 2 (نمونه 13) هیچ تغییری در غلظت متابولیتها ایجاد نکرد. افزایش کمی ، اما از نظر آماری ناچیز ، در غلظت DOPAC و HVA (+ 23٪ در هر دو مورد) نسبت به پایه (نمونه 12) مربوط به اولین دوره جمع آوری با زن 2 (نمونه 14) است. این افزایش کوتاه مدت بود ، با این حال ، و به مقادیر پایه برای سه نمونه باقی مانده کاهش یافت (15-17). مستقل tآزمایشات انجام شده بین نمونه های پایه (یعنی 1، 10 و 12) نشان می دهد که مقادیر دوم و سوم (به ترتیب نمونه های 10 و 12)، هر چند متفاوت از یکدیگر نیستند، در مقایسه با نمونه اولیه اولیه باقی می ماند برای DOPAC و HVA (p <0.05 در هر دو مورد).
بافت شناسی
پروبهای میکروبیالیز در NAC (شکل.2) در محدوده + 1.20 + 1.70 میلیمتر از برگی (جمجمه صاف). متغیر نیز در هواپیما Mediolateral وجود دارد؛ داده ها نمونه گیری را از پوسته و زیرمجموعه های اصلی NAC نشان می دهند.
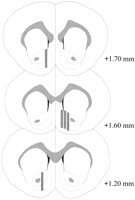
نمایش نسخه بزرگتر:
شکل. 2.
محل قرارگیری پروبهای میکروبیالیز درون NAC موشهای نر استفاده شده در آزمایش اثر کولیج. مستطیل های سایه دار منطبق با محدوده غشاء پروب میکروبیالیز است. بخش های مغز سرطانی کرونال از قرن نوزدهم باز می شودPaxinos و واتسون (1986).
بحث
در توافق با گزارش های قبلی، نتایج کنونی نشان می دهد که افزایش انتقال مادون قرمز Mesolimbic همراه با اجزاء اشتها آور و مغذی رفتار جنسی جنس مردانه به وسیله ارزیابی توسطدر داخل بدن میکردیلیز (مس و همکاران، 1990; Pfaus و همکاران، 1990;Pleim et al.، 1990; Damsma et al.، 1992; Wenkstern و همکاران، 1993; Fumero et al.، 1994; مس و همکاران، 1995a,b,d) علاوه بر این، این نتایج یک همبستگی عصبی را برای سفتی جنسی و پس از آن مجددا مجددا از همپوشانی در پاسخ به یک زن گیرنده رمان (اثر کولیج) فراهم می کند. داده های حاضر نشان می دهد که ویژگی های تحریک کننده یک زن قابل پذیرش جدید ممکن است باعث افزایش انتقال NAC DA در یک موش صحرایی نگرش جنسی می شود که به نوبه خود می تواند به تغییر دوباره رفتار جنسی مرتبط باشد. این اولین بار است که در افزایش کمی در NAC DA در هنگام ارائه یک زن جدید در پشت صفحه نمایش ظاهر می شود و بیشتر قانع کننده تر می شود به عنوان یک افزایش بیشتر در طی نسل جدید با زن 2 (شکل.1).
حضور اولین زن گیرنده در پشت صفحه منجر به افزایش شدید اشتها در NAC DA فاضلاب (44٪ از ابتدایی) مشابه با آنچه که در آزمایش های قبلی با استفاده از یک طرح مشابه گزارش شده بود (30٪، Pfaus و همکاران، 1990؛ 35٪Damsma et al.، 1992) همچنین در توافق با این مطالعات مشاهده شد که جریان NAC DA بیشتر در طول جمع آوری افزایش یافته است (به> 95> بالاتر از سطح پایه در آزمایش حاضر). اگرچه می توانیم رفتارهای متقابل را با افزایش انتشار NAC DA مرتبط بدانیم (Wenkstern و همکاران، 1993; ویلسون و همکاران، 1995)، مهم است كه اصطلاحات "اشتها آور" و "محتوايي" را در زمينه رفتارهاي جنسي بررسي كنيم. در حالی که مرحله ای که زن در پشت صفحه نمایش قرار دارد، صرفا آماده یا آماده است، رفتار در فاز copulation نمیتواند صرفا صرفه جویی در نظر گرفته شود. از آنجا که "اشتها" می تواند برای توصیف تمام رفتارهایی که منجر به برداشتن یک رفتار انگیزه شده (copulation) شود، رفتار اولیه مردان در هنگام فعالیت در مرحله "consummatory" بهترین توصیف به عنوان اشتها آور است؛ مرد اغلب وقت و تلاش خود را برای پیگیری زن برای مقابله با او انجام می دهد. در این راستا ما می توانیم حداکثر انتقال NAC DA را با استفاده از محاسبات پیوند داد همچنین اجزای پر جنب و جوش رفتار جنسی جنین مرد.
دسترسی به زن دوم، زن رمان منجر به تسویه حساب جدید در هر موضوع شد. مطالعات قبلی نشان داده است که اکثر موش های صحرایی اجازه می دهند با استفاده از یک پروتکل رفتاری مشابهی که در آزمایش حاضر استفاده می شود، در طی آزمایش 24 بعدا (ساحل و اردن، 1956) این احتمال وجود دارد که وجود ویژگی های جدید محرک زن 2، که ممکن است شامل بویایی و نشانه های بصری و شنوایی باشد، منجر به تجمع مجدد شده است. سوال جالبی که هنوز پاسخی برای آن مطرح است این است که با چه ساز و کاری یک موش مردانه یک زن رمان از یک زن که اخیرا به هم متصل شده است، تمایل دارد. یک سایت برای آن مکانیسم ممکن است در سیستم بویایی اصلی دروغ باشد. گزارش شده است که یکپارچگی این سیستم برای اثر Coolidge در همسترها (جانستون و رسموسن، 1984) با این حال سیستم بویایی جانبی vomeronasal، که در آن فرآیند حافظه فرومون به تازگی در موش (Kaba et al.، 1994)، همچنین یک نامزد اصلی است. در این رابطه، قابل توجه است که افزایش در انتقال NAC DA با استفاده از در داخل بدن ولتاژ سنجی در موشهای صحرایی نر با ملافه هایی که در موش های صحرایی در موش صحرایی (Louillot و همکاران، 1991; میچل و گراتون، 1992) علاوه بر این، کاربرد K+ به طور مستقیم به لنز عصبی vomeronasal از لامپ متخصص عفونی کننده و همچنین به عنوان لامپ عصبی لوازم جانبی، برای افزایش NAC DA انتقال (میچل و گراتون، 1992).
اولین 15 حداقل مقاربت با زن 2 با افزایش قابل توجهی در NAC DA همراه بود. در مقایسه با زن 1، تعامل با زن 2 در NAC DA با همان اندازه در طول مراحل اشتعال (12٪) و یا تمرین (34٪) افزایش نمی یابد. با این وجود، این افزایش های کوچک در NAC DA با کاهش سطح رفتار جنسی نمایش داده شده با زن 2 در مقایسه با زن 1 همبستگی دارد. غلظت متابولیت در طول فاز سدیم باقی می ماند و منجر به ایجاد غلظت های پایه جدید (نمونه های 10 و 12) که از مقادیر اولیه اولیه (نمونه 1) به طور قابل توجهی افزایش یافته است.
تاخیر زمانی در افزایش غلظت DOPAC و HVA در طی کوپولیسم سازگاری آنها با متابولیت های ترکیب اصلی، DA است. پیشنهاد شده است که غلظت متابولیت های میکرودلیال، حداقل در طی رفتار طبیعی که از نظر فارماکولوژی رانده نمی شود، یک شاخص مفید برای فعالیت عصبی (Damsma et al.، 1992; Fumero et al.، 1994) این واقعیت که غلظت متابولیت ها حتی در طول دوره های غیر فعال بودن جنسی نیز در این آزمایش باقی ماند، هنگامی که غلظت DA به مقادیر اولیه بازمیگردد، این پیشنهاد را تردید می کند.
افزایش پایدار غلظت متابولیت های DA در این آزمایش، نمای متفاوتی از پروپتیک محدوده (mPOA) متابولیت های DA را در موش های صحرایی نشان می دهد که روز اول پس از خوردن آنها به اشباع (مس و همکاران، 1995a,b) افزایش پایدار غلظت DOPAC و HVA در NAC یا mPOA همیشه زمانی مشاهده می شود که دوره جفت گیری از یک مدت ثابت است، بسیار کوتاه تر از زمان مورد نیاز برای رسیدن به اشباع. به عنوان مثال، بسیاری از مطالعات نشان داده اند که غلظت DOPAC افزایش یافته است و در طول مقاربت افزایش یافته است اما به زودی پس از خارج شدن از ماده به مقادیر اولیه کاهش یافت (Pfaus و همکاران، 1990; Pleim et al.، 1990; Damsma et al.، 1992;هال و همکاران، 1993; Wenkstern و همکاران، 1993; هال و همکاران، 1995) در مطالعه توسط مس و همکاران (1995b)غلظت خارج سلولی پایه DOPAC و HVA در mPOA باقی مانده در روزهای متوالی 4 مطابق با یک دوره عدم فعالیت جنسی بود. در روز چهارم، درست قبل از اینکه حیوانات دوباره از بین بروند، غلظت پایه متابولیت ها نزدیک به مقادیر پیش آزمون بود. نویسندگان الگو الگوی تغییرات عصبی را نسبت به افرادی که پس از تزریق مسدود کننده های گیرنده DA مورد تشخیص قرار گرفته اند ((Zetterström و همکاران، 1984; Imperato و DiChiara، 1985) و پیشنهاد کرده اند که وضعیت عدم فعالیت جنسی می تواند از طریق انتشار پرولاکتین، که ممکن است به عنوان یک "neuroleptic آندوژنیک" (مس و همکاران، 1995a,b,d) واضح است که تجویز نورولپتیک همراه با افزایش غلظت متابولیت خارج سلولی و خروجی DA (Zetterström و همکاران، 1984; Imperato و DiChiara، 1985) متاسفانه مس و همکاران (1995a,b) قادر به تشخیص غلظت mPOA DA نبودند. در مطالعه حاضر، غلظت DA در NAC به مقادیر preopulation بازگشت، در حالیکه غلظت DOPAC و HVA همچنان بالا بود. این الگو با نقش نرولپتیک درونی مغز در NAC به منظور ایجاد ساییدگی جنسی متضاد است.
با توجه به دخالت نورونهای مازولیمبیک DA در رفتار انگیزشی (فیبر و فیلیپس، 1986; بلکبرن و همکاران، 1992; Kalivas و همکاران، 1993; LeMoal، 1995) و حساسیت آنها به محرک های زیست محیطی جدید (فابر و همکاران، 1983; شولتز، 1992; Mirenowicz و Schultz، 1994)، افزایش مشاهده شده در غلظت خارج سلولی NAC DA در پاسخ به زن جدید، مطابق با فرضیه است که فعالیت در این سیستم DA برای بازتولید رفتار جنسی بسیار مهم است. علاوه بر این، گزارش های مربوط به افزایش اشتها و تسریع در انتقال DA (هال و همکاران، 1993, 1995;Mas و همکاران، 1995b; ساتو و همکاران، 1995) و فعالیت عصبی (شیمورا و همکاران، 1994) در mPOA موشهای نر در طی رفتار جنسی نشان می دهد که این ساختار همچنین می تواند به کوپلیزه های جدیدی از اثر کولیج کمک کند.
با توجه به نقش کلی سیستم DA مازولیمبیک در رفتار انگیزه، به خوبی ثابت شده است که غلظت خارج سلولی DA نیز قبل، در طی و بلافاصله پس از مصرف وعده غذایی افزایش می یابد با بازگشت به مقادیر ابتدایی ~30 دقیقه بعد (ویلسون و همکاران، 1995) به خوبی شناخته شده است که سدیم ناشی از مواد غذایی تحت تأثیر خواص حسی آن است. انسانها و حیوانات غذائی را که به غذا خوردن غذا می خورند و غذاهای دیگری که خورده نشده اند رد می کنند (رولز، 1986) این مسئله را مطرح می کند که آیا خروجی DA خارج سلولی در NAC به صورت انتخابی با ارائه یک نوع جدید از غذا افزایش می یابد، اما نه به دلیل مصرف غذا که اخیرا برای تطابق با روش مشابه با آنچه در مطالعه حاضر در زمینه انگیزه جنسی اگر تأیید شود، این رابطه کلی بین ویژگی های حسی پاداش های طبیعی، تنش و انتقال مزانشیمی Mesolimbic، نقش مهمی در این سیستم عصبی در تنظیم فرآیندهای انگیزشی دارد که اختلال آن می تواند منجر به اختلالات جدی در خوردن و عملکرد جنسی شود .
مراجع
- ↵
- ساحل FA،
- اردن L
(1956) خستگی جنسی و بهبودی در موش مرد. QJ تجربه روانی 8: 121-133.
- ↵
- برمینگ،
- Lott DF
- اندرسون ل
(1966) ویژگی های زمانی از اثر کوليج در رفتار انسدادی روده مردانه. J Comp Physiol روانپزشکی 65: 447-452.
- ↵
- بلکبرن JR
- PFaus JG
- Phillips AG
(1992) عملكرد دوپامين در رفتارهاي پرهزينه و دفاعي. Prog Neurobiol 39: 247-279.
- ↵
- Damsma G
- PFaus JG
- Wenkstern D،
- Phillips AG
- Fibiger HC
(1992) رفتار جنسی باعث انتقال پروتئین دوپامین در هسته آکومبن و استریاتوم موشهای نر نسبت به تازگی و حرکت می شود. بهوان نوروزی 106: 181-191.
- ↵
- فاب M
- Rolls ET،
- اشتون جی. ا.،
- ویلیامز گ
(1983) فعالیت نورون ها در منطقه تکتونستیکی شکم میمون رفتار می کند. بهان مغز رز 9: 213-235.
- ↵
- Fibiger HC
- Phillips AG
(1986) پاداش، انگیزه، شناخت: روانشناسی سیستم های دوپامین مزئوتانسفالیک. در راهنمای فیزیولوژی: سیستم عصبی IV، eds بلوم FE، Geiger SD (انجمن فیزیولوژی آمریکا، Bethesda، MD)، pp 647-675.
- ↵
- Fiorino DF
- Coury AG
- Fibiger HC
- Phillips AG
(1993) تحریک الکتریکی سایت های پاداش در منطقه تنگالال شکم، انتقال دوپامین را در سلول های مغز موش نشان می دهد. بهان مغز رز 55: 131-141.
- ↵
- فیشر A
(1962) اثر تغييرات محرک در تحريک پذيري جنسي در موش صحرايي نر. J Comp Physiol روانپزشکی 55: 614-620.
- ↵
- فاولر ه
- ویالن RE
(1961) تغییر در محرک های انگیزشی و رفتار جنسی در موش های مردانه. J Comp Physiol روانپزشکی 54: 68-71.
- ↵
- فومرو ب
- فرناندز ورا JR
- گونزالس مورا JL،
- ماس م
(1994) تغییرات در گردش مونوآمین در ناحیه پیشانی مرتبط با رفتار جنسی مردانه: مطالعهی میکروبیالیز. با توجه به مغز 662: 233-239.
- ↵
- هولمز GM
- هولمز DG
- Sachs BD
(1987) یک سیستم جمع آوری داده مبتنی بر IBM-PC برای ضبط رفتارهای جنسی جوندگان و ضبط کلی رویداد. فیزیول بوهو 44: 825-828.
- ↵
- هال EM،
- ایاتون RC
- موسی ج
- Lorrain DS
(1993) کوپولاسیون فعالیت دوپامین را در ناحیه preoptic medial موشهای نر افزایش می دهد. علم زندگی 52: 935-940.
- ↵
- هال EM،
- Jianfang D
- Lorrain DS
- Matuszewich L
(1995) دوپامین غير سلولی در ناحيه Preoptic Medial: پيامدهای انگيزه جنسی و کنترل هورمونی جمعيت. J Neurosci 15: 7465-7471.
- ↵
- Imperato A
- DiChiara G
(1985) آزادسازی و متابولیسم دوپامین در موشهای بیداری پس از نورولپتیک سیستمیک به وسیله دیالیز trans-striatal مورد بررسی قرار گرفت. J Neurosci 5: 297-306.
- ↵
- جانستون RE،
- Rasmussen K
(1984) شناخت فرد از همسترهای زن توسط مردان: نقش نشانه های شیمیایی و سیستم های بویایی و وومروناسال. فیزیول بوهو 33: 95-104.
- ↵
- کابا ه،
- حایشی ی،
- هیگوچی تی
- ناکانیشی س
(1994) القای حافظه بویایی توسط فعال شدن گیرنده گلوتامات metabotropic. علم 265: 262-264.
- ↵
- Kalivas PW
- سورگ BA،
- قلاب MS
(1993) فارماکولوژی و مدارهای عصبی حساسیت به داروهای روانپزشکی. بهف فارماکول 4: 315-334.
- ↵
- LeMoal M
(1995) نورونهای دوپامینرژیک Mesocorticolimbic. نقش های کارکردی و نظارتی در Psychopharmacology: نسل چهارم پیشرفت، eds بلوم FE، کاپرف دی جی (Raven، نیویورک)، pp 283-294.
- ↵
- Louillot A
- گونزالس مورا JL،
- گوادالوپ تی
- ماس م
(1991) محرک های بویایی مرتبط با جنسی باعث افزایش انتخابی در آزمودن دوپامین در سلول های مغز موش صحرایی می شود. با توجه به مغز 553: 313-317.
- ↵
- ماس م،
- گونزالس مورا JL،
- Louillot A
- تنها C
- گوادالوپ تی
(1990) افزایش آزادسازی دوپامین در هسته آکومبن موش صحرایی نر به عنوان ولتامتری in vivo نشان داده شده است. Neurosci Lett 110: 303-308.
- ↵
- ماس م،
- فومرو ب
- فرناندز ورا JR
- گونزالس مورا JL
(1995a) همبستگی های عصبی مصنوعی از خستگی جنسی و بهبودی به وسیله ارزیابی میکرو دیالیز in vivo می باشد. با توجه به مغز 675: 13-19.
- ↵
- ماس م،
- فومرو ب
- گونزالس مورا JL
(1995b) نظارت ولتاژ و میکرو دیالیز مغز استخوان مونوآمین مغز در هنگام تعاملات اجتماعی. بهان مغز رز 71: 69-79.
- ↵
- ماس م،
- فومرو ب
- پرز رودریگز من
(1995c) القاء رفتار جفتگیری آپومورفین در موشهای صحرایی جنسی. Eur J Pharmacol 280: 331-334.
- ↵
- ماس م،
- فومرو ب
- پرز رودریگز من
- گونزالس مورا JL
(1995d) عصبی شیمیایی سفتی جنسی است. یک مدل تجربی میل میلان. در فارماکولوژی عملکرد و اختلالات جنسی، ed Bancroft J (Raven، نیویورک)، pp 115-126.
- ↵
- میرانوویچ جی
- شولتز W
(1994) اهمیت غیر قابل پیش بینی برای پاسخ پاداش در نورونهای دوپامینرژیک پریمات. J Neurofysiol 72: 1024-1027.
- ↵
- میچل جی بی،
- Gratton A
(1992) آزادسازی دوپامین mesolimbic بوسیله فعال سازی سیستم بوسیله لوازم جانبی: یک مطالعه کرونومای سنجی با سرعت بالا. Neurosci Lett 140: 81-84.
- ↵
- Paxinos G
- واتسون سی
(1986) مغز موش در مختصات stereotaxic (2nd ed). (Academic، San Diego).
- ↵
- PFaus JG
- Damsma G
- Nomikos GG
- Wenkstern D،
- CD بلخا
- Phillips AG
- Fibiger HC
(1990) رفتار جنسی باعث افزایش انتقال دوپامین مرکزی در موش صحرایی می شود. با توجه به مغز 530: 345-348.
- ↵
- PFaus JG
- Everitt BJ
(1995) روانشناسی رفتار جنسی. در Psychopharmacology: نسل چهارم پیشرفت، eds بلوم FE، کاپرف دی جی (Raven، نیویورک)، pp 743-758.
- ↵
- PFaus JG
- گورزالکا BB
(1987) اپیوئیدها و رفتار جنسی. Neurosci Biobehav Rev 11: 1-34.
- ↵
- Phillips AG
- CD بلخا
- PFaus JG
- بلکبرن JR
(1992) همبستگی های عصبی بین حالت های احساسی مثبت: دوپامین، پیش بینی و پاداش. در بررسی بین المللی مطالعات در مورد احساسات، ed Strongman (Wiley، نیویورک)، pp 31-50.
- ↵
- Pleim ET،
- ماتوچیک JA
- بارفیلد RJ
- Auerbach SB
(1990) همبستگی آزادسازی دوپامین در هسته تکبون با رفتار جنسی مردانه در موش صحرایی. با توجه به مغز 524: 160-163.
- ↵
- رودریگز منزو گ
- Fernandez-Guasti A
(1994) معکوس کردن خستگی جنسی توسط عوامل سروتونرژیک و نورآدرنرژیک. بهان مغز رز 62: 127-134.
- ↵
- رودریگز منزو گ
- Fernandez-Guasti A
(1995a) مشارکت سیستم نئودرنرژیک مرکزی در بازسازی رفتار مخلوطی از موش های صحرایی غیر جنسی با استفاده از یوهیمبین، نالوکسون و 8-OH-DPAT. مغز رز بول 38: 399-404.
- ↵
- رودریگز منزو گ
- Fernandez-Guasti A
(1995b) آنتاگونیستهای اپيوئيد و پاتويت اشباع جنسی است. روانپزشکی 122: 131-136.
- ↵
- رول BJ
(1986) سفتی خاص حساسیت. نتر Rev 44: 93-101.
- ↵
- سالامون JD
(1996) نورولوژي رفتاري انگيزش: مسائل روش شناختي و مفهومي در مطالعات فعاليت پویا هسته تکبامن دوپامين. J Neurosci روش 64: 137-149.
- ↵
- ساتو ی،
- ودا ه،
- هوریتا ه،
- سوزوکی N
- Shibuya A
- Adachi H،
- کاتو ر،
- Tsukamoto T
- Kumamoto Y
(1995) آزادسازی دوپامین در ناحیه preoptic medial در طی رفتار انسانی مردانه در موش صحرایی. با توجه به مغز 692: 66-70.
- ↵
- شولتز W
(1992) فعالیت نورون های دوپامین در پرایموت رفتار می کند. Semin Neurosci 4: 129-138.
- ↵
- شیمورا T
- Yamamoto T
- شیمخوچی م
(1994) ناحیه preoptic medial در هر دو جنس تحریک و عملکرد در موش های صحرایی نر است: دوباره ارزیابی فعالیت های نورون در حیوانات آزاد در حال حرکت است. با توجه به مغز 640: 215-222.
- ↵
- Wenkstern D،
- PFaus JG
- Fibiger HC
(1993) انتقال دوپامین در موش های صحرایی نر موش های نر در اولین مواجهه با موش های صحرایی جنسی دریافت می شود. با توجه به مغز 618: 41-46.
- ↵
- ویلسون سی
- Nomikos GG
- کولو م،
- Fibiger HC
(1995) ارتباط دوپامینرژیک رفتار رفتار انگیزه: اهمیت درایو. J Neurosci 15: 5169-5178.
- ↵
- ویلسون جی. ر.
- کان RE،
- FA FA
(1963) اصلاح در رفتار جنسی موشهای نر تولید شده با تغییر محرک زن. J Comp Physiol روانپزشکی 56: 636-644.
- ↵
- Zetterström T
- شارپ ت
- Ungerstedt U
(1984) اثر داروهای نورولوپتیک بر انتشار دوپامین و متابولیسم در موش صحرایی ناشی از دیالیز داخل مغزی. Eur J Pharmacol 106: 27-37.
مقالات با ذکر منبع این مقاله
- Neuroplasticity نوروپلاستی ناشی از اپوئید در نورون های دوپامینرژیک در فاکتورهای درشت مرزی نفوذ طبیعی و Opiate مجله علوم اعصاب، 25 ژوئن 2014، 34 (26): 8825-8836
- عدم تعادل در حساسیت به نوع مختلف پاداش در قمار آسیب شناسی مغز، 1 اوت 2013، 136 (8): 2527-2538
- قانون طب جایگزین و مواد مخدر در مکانیسم های پلاستیکی عضلانی با {Delta} FosB به عنوان یک واسطه کلیدی مجله علوم اعصاب، 20 فوریه 2013، 33 (8): 3434-3442
- پاسخ های عصبی در هسته Accumbens Shell در طی رفتار جنسی در موش های صحرایی نر مجله علوم اعصاب، 1 فوریه 2012، 32 (5): 1672-1686
- عشق رمانتیک: یک سیستم مغز پستاندار برای انتخاب همسر معاملات فلسفی انجمن سلطنتی B: علوم زیستی، 29 دسامبر 2006، 361 (1476): 2173-2186
- نقاط داغ هیدونیک در مغز عصب شناس، 1 دسامبر 2006، 12 (6): 500-511
- مدولاسيون توسط هسته آميگدالار مركزي و نيمه نازك همبستگي هاي دوپامينرژيكي از تغذيه با ساتيت در محيط كشت مغزي موش صحرايي مجله علوم اعصاب، 15 دسامبر 2002، 22 (24): 10958-10965
- Frequency of Transients Concentration Dopamine Increases Striatum Back and Ventral در موشهای صحرایی نر در هنگام معرفی مختصات مجله علوم اعصاب، 1 دسامبر 2002، 22 (23): 10477-10486
- بررسی کتاب: دینامیک دوپامین بین سلول در اقدامات حاد و مزمن کوکائین عصب شناس، 1 اوت 2002، 8 (4): 315-322
- درگیری دیفرانسیل NMDA، AMPA / Kainate و گیرنده های دوپامین در هسته هسته Accumbens در دستیابی و عملکرد رفتار رویکرد Pavlovian مجله علوم اعصاب، 1 دسامبر 2001، 21 (23): 9471-9477
- کنترل رفتار محرک کوکائین توسط محرک های مرتبط با مواد مخدر در موش صحرایی: تأثیر در بهبود عملکرد سطوح دوپامین مضر عمل شده و عملکردهای خونی در amygdala و nucleus accumbens PNAS، 11 آوریل 2000، 97 (8): 4321-4326
- تسهیل رفتار جنسی و افزایش نفوذ دوپامین در محور هسته ای موش های صحرایی پس از حساسیت رفتاری ناشی از D-آمفتامین مجله علوم اعصاب، 1 ژانویه 1999، 19 (1): 456-463
- علوم اعصاب: جلب توجه مغز علوم، 3 اکتبر 1997، 278 (5335): 35-37
- دوپامينرژيک مرتبط با حساسيت ويژه اي در قشر مدفون قدامي و هسته مغز موش صحرايي مجله علوم اعصاب، 1 اکتبر 1999، 19 (19): RC29
