ՄԵԿՆԱԲԱՆՈՒԹՅՈՒՆՆԵՐ. Քանի որ ավելի ուշ ուսումնասիրությունները ցույց կտան, որ DeltaFosB- ը ընդհանուր մոլեկուլային անջատիչն է ինչպես թմրանյութերի, այնպես էլ վարքային կախվածությունների համար: Դա արտագրման գործոն է, ինչը նշանակում է, որ դա ազդում է գեների միացման կամ անջատման վրա: Ինչպես նշվել է այլուր, կախվածություն առաջացնող դեղերը միայն առևանգում են նորմալ մեխանիզմները: Այդ պատճառով հիմար է ենթադրել, որ վարքային կախվածությունները չեն կարող գոյություն ունենալ:
Proc Natl Acad Sci ԱՄՆ Ա. 2001 Սեպտեմբեր 25; 98 (20): 11042-11046:
doi: 10.1073 / pnas.191352698:
Էրիկ Ջ. Նեստլեր *, Միշել Բարրոտն ու Դեյվիդ Վ
Հյուսվածքաբանության կենտրոն եւ Նեյրոնագիտության կենտրոն, Տեխասի հարավային բժշկական կենտրոնի համալսարան, 5323 Harry Hines Boulevard, Դալլաս, TX 75390-9070
Վերացական
Թմրամոլության բնութագրիչ որոշ վարքային անբավարարությունների երկարատեւությունը առաջարկել է, որ նյարդային գենի ձեւավորման կարգը կարող է ներգրավվել այն գործընթացում, որով թմրանյութերի օգտագործումը թմրամոլության վիճակ է առաջացնում: Եսncreasing evidence- ն ցույց է տալիս, որ ΔFosB- ի transcription գործոնը ներկայացնում է մի մեխանիզմ, որով չարաշահման թմրանյութերը արտադրում են ուղեղի համեմատաբար կայուն փոփոխություններ, որոնք նպաստում են կախվածության ֆենոտիպին: ΔFosB, Fos- ի ընտանիքի տարրական գործակիցների անդամ, կուտակվում է միջուկի ակլեմենսի նեյրոնների եւ դորալ ստրիատումի մեջ (ուղեղի շրջաններ, որոնք կարեւոր են կախվածության համար), բազմաթիվ չարաշահման թմրամիջոցների չարաշահումից հետո: ΔFosB- ի նմանատիպ կուտակումն առաջանում է կոմպոզիցիոն հոսպիտալից հետո, ինչը ենթադրում է, որ ΔFosB- ը կարող է կուտակվել ի պատասխան կոմպոզիտիվ վարքի բազմաթիվ տեսակների. Կարեւորն այն է, որ ΔFosB- ն նեյրոններում շարունակում է մնալ համեմատաբար երկար ժամանակ, քանի որ նրա արտակարգ կայունությունը: Հետեւաբար, ΔFosB- ն ներկայացնում է մոլեկուլային մեխանիզմ, որը կարող է նախաձեռնել եւ այնուհետեւ պահպանել գենային արտահայտման փոփոխությունները, որոնք երկար ժամանակ թմրանյութի ազդեցության դադարից հետո շարունակվում են. Ուսումնասիրությունները սինթեզվող տրանսգենային մկների վրա, որոնք գերազանցում են կամ ΔFosB- ին կամ սպիտակուցի գերիշխող բացասական հակազդեցությանը, ցույց են տալիս, որ ΔFosB- ն առաջացնում է զգայունություն թմրամիջոցների չարաշահման թմրամիջոցների վարքային ազդեցությունների եւ, հնարավոր է, թմրանյութերի որոնման վարքի ավելացում: Այս աշխատանքը սատարում է այն տեսակետին, որ ΔFosB- ն գործում է որպես կայուն «մոլեկուլային անջատիչի» տեսակ, որը աստիճանաբար թարմ դեղերի պատասխաններ է հաղորդում համեմատաբար կայուն հարմարվողականության մեջ, որոնք նպաստում են նյարդային եւ վարքային պլաստիկության երկարատեւությանը, որը ենթարկվում է կախվածության:
Addiction հետազոտությունը կենտրոնանում է հասկանալու այն բարդ ուղիները, որոնցով չարաշահման թմրանյութերը փոխում են ուղեղը `կախվածության անբավարարությունը բնութագրող վարքային անբավարարություն առաջացնելով: Այս ոլորտում կարեւորագույն մարտահրավերներից մեկը ուղեղի համեմատաբար կայուն փոփոխությունների բացահայտումն է, հաշվի առնելով այն վարքային աննորմալությունները, որոնք հատկապես երկարատեւ են: Օրինակ, մարդկային կախվածությունը կարող է ռեցիդիվի ռիսկի բարձրացման համար նույնիսկ տարիներ շարունակ ձեռնպահ մնալուց հետո:
Այս վարքագծային անբավարարությունների կայունությունը հանգեցրել է այն առաջարկին, որ դրանք կարող են միջնորդվել, գոնե մասամբ, գեն արտահայտության փոփոխությունների միջոցով (1-3): Այս տեսակետի համաձայն, չարաշահման թմրամիջոցների կրկնակի ազդեցությունը միանգամից խանգարում է ուղեղի որոշակի synapses փոխանցմանը, որը զգայուն է դեղին: Այդպիսի անհեթեթությունները, ի վերջո, ազդում են միջուկային հաղորդակցման կասկադների միջոցով միջուկին, որտեղ նրանք առաջինն են նախաձեռնում եւ հետո պահպանում են որոշակի գեների արտահայտման փոփոխություններ: Առաջնային մեխանիզմը, որի միջոցով ազդանշանային փոխանցման ուղիները ազդում են գենի արտահայտման վրա, այն է, որ transcription գործոնների կարգավորում, գենների կարգավորող շրջաններում կապող սպիտակուցներ եւ դրանց փոխակերպումը:
Այսպիսով, կախվածության հետազոտության նպատակներից մեկն այն է, որ հայտնաբերել տառապող գործոնները, որոնք փոփոխվում են ուղեղի գոտիներում, որոնք ենթարկվում են կախվածության մեջ, չարաշահման թմրամիջոցների քրոնիկ վարչարարությունից հետո: Անցյալ տասնամյակում (1-6) հայտնաբերվել են մի քանի նման transcription գործոններ: Այս վերանայման կենտրոնը հանդիսանում է ΔFosB կոչվող հատուկ արտագրման գործոնի վրա:
ΔFosB- ի ներարկումը չարաշահման թմրանյութերի միջոցով
FosB- ն, որը ձեւավորված է fosB գենի կողմից, Fos- ի ընտանիքի անդամ է, որը ներառում է նաեւ c-Fos, FosB, Fra1 եւ Fra2 (7): Այս Fos ընտանիքի սպիտակուցները heterodimerize հետ Jun ընտանիքի սպիտակուցներ (c-Jun, JunB կամ JunD) ստեղծել ակտիվ AP-1 (ակտիվացնող սպիտակուցը-1) transcription գործոնները, որոնք կապում են AP- 1 կայքեր (կոնցեսուսի հաջորդականությունը: TGAC / GTCA) որոշ գեների խթանողներ, որոնք կարգավորում են իրենց արտագրումը:
Այս Fos- ի ընտանեկան սպիտակուցները առաջանում են արագ եւ անցումային շրջանում կոնկրետ ուղեղային շրջաններում `չարաշահման բազմաթիվ թմրանյութերի սուր վարվելուց հետո (Նկար 1) (8-11): Հատկանշական շրջաններն այն են, որ թռչունների վարակիչները եւ դորալ ստերիոտները, որոնք հանդիսանում են դեղերի վարքային պատասխանների կարեւոր միջնորդներ, մասնավորապես նրանց հատուցող եւ շարժիչ ուժի ազդեցության հետեւանքները (12, 13): Այս սպիտակուցները վերադառնում են բազալ մակարդակներում թմրամիջոցների ներթափանցման ժամերին:
Նկար 1
Ցույց է տալիս ΔFosB- ի աստիճանական կուտակումը `ընդդեմ այլ Fos ընտանիքի սպիտակուցների արագ եւ անցողիկ ներդիրի` չարաշահման թմրամիջոցների դեմ: (A) Autoradiogram- ը ցույց է տալիս այս տարբեր սպիտակուցների դիֆերենցիալ ինդուկցիան սուր խթանման միջոցով (1-2 ժամը մեկ թմրամիջոցների ազդեցության հետեւանքով) եւ քրոնիկ խթանման դեմ (1 օր օրից, կրկնակի թմրանյութերի ազդեցության դեպքում): (B) Fos- նմանատիպ սպիտակուցների մի քանի ալիքներ [բաղկացած են c-Fos (52- ից 58-kDa isoforms), FosB (46- ից 50-kDa isoforms), ΔFosB (33-kDa isoform) եւ Fra1 կամ Fra2 40 kDa)] ներարկվում են միջուկային հեծանիվների եւ դորալ ստիատալ նեյրոնների միջոցով `չարաշահման թմրամիջոցների սուր վարման միջոցով: Նաեւ իջեցված են ΔFosB (35-37 kDa) կենսաքիմիական փոփոխված izoforms; նրանք նույնպես հարուցվում են (չնայած ցածր մակարդակներում) թմրամիջոցների սուր դեղերի կառավարման արդյունքում, սակայն երկար ժամանակ պահպանում են ուղեղի կայունության պատճառով: (C) Կրկնվող (օր.` երկու անգամյա) դեղամիջոցների կիրառմամբ, յուրաքանչյուր սուր խթան ներծծում է կայուն ΔFosB isoforms- ի ցածր մակարդակը, որը նշված է ստորերկրյա տողերի ստորին խմբերով, որոնք նշում են ΔFosB- ի կողմից յուրաքանչյուր սուր խթանման միջոցով: Արդյունքը ΔFosB- ի ընդհանուր մակարդակների աստիճանական աճն է `կրկնվող խթանիչներով քրոնիկ բուժման ընթացքում, ինչը նշվում է գրաֆում աճող տեմպի գիծով:
Շատ տարբեր արձագանքներ երեւում են չարաշահման թմրամիջոցների քրոնիկ վարչարարությունից հետո (Նկար 1). ΔFosB- ի (մոլեկուլային զանգված 35-37 kDa) կենսաքիմիական փոփոխված izoforms կուտակվում են նույն ուղեղի շրջաններում, երբ կրկնվող թմրամիջոցների ազդեցությունը, իսկ մյուս բոլոր Fos ընտանիքի անդամները ցույց են տալիս հանդուրժողականության (այսինքն, կրճատվել է ներարկումը համեմատած նախնական դեղերի ազդեցության հետ): ΔFosB- ի նման կուտակումը նկատվել է կոկաինի, մորֆինի, ամֆետամինի, ալկոհոլի, նիկոտինի եւ ֆանտիկլիդինի համարe (11, 14-18): Որոշ ապացույցներ կան, որ այս ինդուկցիան ընտրովի է դինորֆինի / նյութի P պարունակող ենթածրագրի միջին ուղեղային նեյրոնների համար (15, 17), թեեւ ավելի շատ աշխատանք է պահանջվում դրա հաստատման համար: 35- ի 37-kDa- ի ÎFOSB ի isoforms հիմնականում JunD հետ dimerize ստեղծել ակտիվ եւ երկարատեւ AP-1 համալիրի շրջանակներում այդ ուղեղի շրջաններում (19, 20). Այս ΔFosB isoforms կուտակվում են քրոնիկ թմրանյութերի ազդեցության պատճառով, քանի որ նրանց աննկարագրելի երկար կիսամյակային կյանքը (21) եւ, հետեւաբար, պահպանվում են նեյրոնների մեջ, դեղերի կառավարման դադարեցումից հետո առնվազն մի քանի շաբաթվա ընթացքում: Հետաքրքիր է, որ ΔFosB isoforms- ը անմիջական վաղ գենի (fosB) բարձր կայուն արդյունքներ են: ΔFosB- ի isoforms- ի կայունությունը ապահովում է նոր մոլեկուլային մեխանիզմ, որի միջոցով գենային արտահայտման մեջ թմրամիջոցների ներածված փոփոխությունները կարող են պահպանվել, չնայած թմրանյութերի հեռացման համեմատաբար երկար ժամանակահատվածներին:
Չնայած այն հանգամանքին, որ միջուկային հեծանվորդները կարեւոր դեր են խաղում չարաշահման թմրանյութերի հատուցող հետեւանքների մեջ, կարծում են, որ բնականոն գործունեություն է ծավալում բնական կարգավորիչներին, ինչպիսիք են սննդամթերքը, խմիչքը, սեռը եւ սոցիալական փոխազդեցությունները (12, 13): Արդյունքում, զգալի հետաքրքրություն է առաջանում այս ուղեղի այլ դոմինանտության այլ դաժան վարքագծում (օրինակ, պաթոլոգիական overeating, gambling, exercise, եւ այլն): Այդ պատճառով մենք ուսումնասիրեցինք, թե արդյոք ΔFosB կարգավորվում է պարտադիր վազքի կենդանիների մոդելում: Անշուշտ, ΔFosB- ի կայուն 35- ի 37-kDa isoforms- ն ընտրված են առնետների ճառագայթային accumbens- ներում, որոնք ցույց են տալիս կոմպոզիտիվ վարման վարքագիծը:
Կայուն ΔFosB Isoforms- ի կենսաքիմիական ինքնությունը
Ինչպես նշվեց վերեւում, ΔFosB- ը isoforms, որոնք կուտակվում են քրոնիկ վարչարարությունից հետո թմրամիջոցների չարաշահման կամ կոմպրեսսիվ վազում ցույց է մոլեկուլային զանգվածի 35-37 kDa: Դրանք կարելի է տարբերակել ΔFosB- ի 33-kDa- ի ինոֆոնտից, որը արագորեն ներարկում է, բայց միանգամայն անցնում է միայն թմրամիջոցների բացահայտման հետեւանքով (Նկար 1) (14, 19, 22): Ընթացիկ ապացույցները ենթադրում են, որ 33-kDa isoform- ը սպիտակուցի մայրենի ձեւն է, որը փոխվել է ավելի կայուն 35- ի 37-kDa- ի արտադրանքների համար (19, 21): Այնուամենայնիվ, կենսաքիմիական մոդիֆիկացիայի բնույթը, որը փոխակերպում է անկայուն 33-kDa իզոֆորմը կայուն 35- ից մինչեւ 37-kDa isoforms, մնացել է անորոշ: Ենթադրվում է, որ ֆոսֆլարիզմը կարող է պատասխանատու լինել (11): Օրինակ, ΔFosB- ի ինդուկցիան ցնցվում է DARPP-32- ով չծնված մկների վրա, որը ստիատալով հարստացված սպիտակուց է (23, 24): Քանի որ DARPP-32- ը կարգավորում է սպիտակուցային ֆոսֆատազի-1- ի եւ սպիտակուցային քնազի A- ի (25, 26) կատալիտիկ ակտիվությունը, այս սպիտակուցի պահանջը կայուն ΔFosB isoforms- ի նորմալ կուտակման համար առաջացնում է այդ կայուն արտադրանքի առաջացման ֆոսֆորլացիայի հնարավոր դերը:
ΔFosB- ի դերը վարքագծային պլաստիկության մեջ `չարաշահման թմրանյութերի նկատմամբ
Թմրամոլության մեջ ΔFosB- ի դերակատարությունը հասկանալի է դարձել տրանսգենային մկների ուսումնասիրության արդյունքում, որտեղ ΔFosB- ն կարող է ընտրովիորեն ներգրավվել մեծահասակների կենդանիների միջուկային հեծանիվների եւ այլ ստիատալային շրջաններում (27, 28): Կարեւորն այն է, որ այդ մկները գերազանցում են ΔFosB- ին սեռից ընտրված դինորֆին / P- պարունակող միջին ճարպային նեյրոնների մեջ, որտեղ թմրանյութերը հավատում են, որ առաջացնում են սպիտակուցը: ΔFosB-overexpressing մկների վարքային ֆենոտիպը, որը շատ առումներով հիշեցնում է կենդանիներին քրոնիկ թմրանյութերի ազդեցության հետեւանքով, ամփոփված է Աղյուսակ 1- ում: Մկները ցույց են տվել, որ քրոնիկները խստորեն եւ քրոնիկ ադմինիստրացիայից հետո (28) ավելացնում են շարժիչի արձագանքները: Նրանք նաեւ ցույց են տալիս, որ կոկաինը եւ morphine- ի հատվածային փորձարկումները (11, 28) խթանող զգայունությունը եւ ինքնուրույն կկիրառի կոկաինի ցածր դոզանները, քան ոսկորները, որոնք չեն գերազանցում ΔFosB- ին: Ի հակադրություն, այս կենդանիները ցույց են տալիս նորմալ պայմանավորված շարժիչ զգայունությունը կոկաինի եւ Morris ջրային լաբիրինթոսում (28) նորմալ տարածական ուսուցման: Տայս տվյալները ցույց են տալիս, որ ΔFosB- ն մեծացնում է կենդանու զգայունությունը կոկաինի և, թերևս, չարաշահման այլ թմրանյութերի նկատմամբ և կարող է հանդիսանալ դեղերի նկատմամբ համեմատաբար երկարատև զգայունության մեխանիզմ.
 ստիատում
ստիատում
| Ավելի բարձր լոկոմոտիվի ակտիվացում `ի պատասխան սուր եւ կրկնակի կոկաինի վարման: |
| Կոկաին եւ մորֆինին ավելացնում են պարգեւատրման պատասխանները `տեղադրման փորձարկումներում: |
| Կոկաինի ցածր դոզանների ինքնակազմակերպման ավելացում: |
| Կոկինի աճող մոտիվացիան առաջադեմ հարաբերության փորձերի մեջ է: |
| Ալկոհոլի նկատմամբ անխոլիտիտիկ պատասխանների ավելացում: |
| Ավելացնող կոմպոզիտիվ վարման վարքագիծը: |
Հիման վրա հիմնված տեղեկությունների համար: 28 և 29.† § ¶
ΔFosB- ի կողմից միջուկային accumbens-dorsal striatum- ում միջնորդավորված վարքային պլաստիկությունը
IԲացի դրանից, նախնական ապացույցներ կան, որ ΔFosB- ի ազդեցությունները կարող են շատ ավելի տարածվել թմրամիջոցների զգայունության որոշումներից `ավելի խորքային վարքագծի նկատմամբ, կապված կախվածության գործընթացի հետ: ΔFosB- ը արտահայտող մկաններն ավելի ու ավելի են աշխատում կոկաինի ինքնակառավարման մեջ, առաջադիմական հարաբերակցության ինքնակառավարման հավասարակշռությունը, ջուրΔFosB- ը կարող է զգայունացնել կենդանիներին կոկաինի խրախուսման մոտիվացիոն հատկություններով, դրանով հանգեցնելով թմրամիջոցների քայքայմանըl. ΔFOSB- արտահայտող մկները նաեւ ցույց են տալիս, որ ալկոհոլի անխոկեցիտային ազդեցությունները, § ֆենոտիպը, որը կապված է մարդու մոտ ալկոհոլի ավելացման հետ: Միեւնույն ժամանակ, այդ վաղ հայտնաբերումները ցույց են տալիս, որ ΔFosB- ը, ի լրումն, չարաշահման թմրամիջոցների նկատմամբ զգայունության բարձրացման, առաջացնում է որակական փոփոխություններ, որոնք նպաստում են թմրանյութերի որոնման վարքագծին: Այսպիսով, ΔFosB- ը կարող է գործել որպես կայուն «մոլեկուլային անջատիչ», որն օգնում է նախաձեռնել եւ պահպանել կախյալ պետությունից կարեւորագույն ասպեկտները: Ընթացիկ քննության ընթացքում կարեւոր հարց է, թե արդյոք թմրանյութերի ազդեցության ընթացքում ΔFOSB- ի կուտակումը նպաստում է թմրամիջոցների որոնման վարքագծին, ընդլայնված հեռացման ժամկետներից հետո, նույնիսկ ΔFOSB մակարդակները նորմալացվում են (տես ստորեւ):
Չափահաս մկները, որոնք գերազանցում են ճառագայթները ΔFosB- ի կողմից, ճառագայթային ակումենսների եւ դորալ ստերիատի մեջ ընտրված են նաեւ ավելի մեծ կոմպոզուսիվ վարժություններ, որոնք վերահսկվում են ծովային քանակի համեմատ: † Այս դիտարկումները բարձրացնում են այն հետաքրքիր հնարավորությունը, որ այս նեյրոնների մեջ ΔFosB կուտակումը ավելի ընդհանուր դեր է խաղում սովորական հիշողությունների ձեւավորման եւ պահպանման եւ կոմպոզիտիվ վարվելակերպը, թերեւս, ամրապնդելով նյարդային սխեմաների արդյունավետությունը, որոնցում գործում են այդ նեյրոնները:
ΔFosB- ն կուտակվում է որոշակի ուղեղային շրջաններում, քրոնիկական կոկաինի ազդեցության հետեւանքով ճարպային ակամենսներից եւ դորալ ստերիմատից դուրս: Նրանց թվում են շրջաններն են amygdala եւ medial prefrontal cortex (15): Ընթացիկ հետազոտության հիմնական նպատակն է հասկանալ ΔFosB- ի ներդիրների ներդրումը այդ տարածաշրջաններում `կախվածության ֆենոտիպի նկատմամբ:
FosB նոկաուտի մկների վրա ավելի վաղ կատարված աշխատանքները պարզել են, որ այդ կենդանիները չեն կարողանում զգայունացնել կոկայինի շարժողական ազդեցության նկատմամբ, ինչը համահունչ է վերը նշված ΔFosB- գերարտահայտող մկների բացահայտումներին (22): Այնուամենայնիվ, fosB մուտանտները ցույց տվեցին ուժեղ զգայունություն կոկաինի սուր ազդեցությունների նկատմամբ, ինչը անհամապատասխան է այս այլ բացահայտումների հետ: FosB մուտանտների հետ կապված գտածոների մեկնաբանությունը, այնուամենայնիվ, բարդանում է նրանով, որ այս կենդանիներին պակասում է ոչ միայն ΔFosB, այլ նաև լիամետրաժ FosB: Ավելին, մուտանտներին պակասում են ինչպես սպիտակուցները ամբողջ ուղեղում, այնպես էլ զարգացման ամենավաղ փուլերից: Իրոք, ավելի վերջերս կատարված աշխատանքը սատարում է ΔFosB գերարտահայտող մկների եզրակացություններին. C-Jun- ի կտրված մուտանտի դյուրագրգիռ գերարտահայտումը, որը հանդես է գալիս որպես ΔFosB- ի գերիշխող բացասական հակառակորդ, ընտրովի միջուկում և մեջքի հատվածում ցույց է տալիս կոկայինի հատուցող ազդեցության նկատմամբ զգայունության նվազում: . Findings Այս հայտնագործությունները շեշտում են այն զգուշությունը, որը պետք է օգտագործվի հիմնադիր մուտացիաներով մկների արդյունքների մեկնաբանման ժամանակ և ցույց են տալիս մեծահասակների ուղեղի պլաստիկության ուսումնասիրություններում մուկի կարևորությունը դյուրահալ և բջջային տիպի մուտացիայով:
ΔFosB- ի նպատակային գեների համար
Քանի որ ΔFosB- ն transcription- ի գործոնն է, հավանաբար սպիտակուցը առաջացնում է վարքի պլաստիկություն այլ գեների արտահայտման փոփոխություններով: ΔFosB- ն գեներացվում է fosB գենի այլընտրանքային հարստացման միջոցով եւ չունի լիարժեք FosB- ում առկա C- տերմինալի փոխակերպման դոմենի մի մասը: Արդյունքում նախապես առաջարկվեց, որ ΔFosB- ն գործում է որպես transcriptional repressor (29): Այնուամենայնիվ, բջիջների մշակույթում աշխատելը հստակ ցույց տվեց, որ ΔFosB կարող է կամ դրդել կամ ճնշել AP-1- միջնորդավորված transkripts, կախված որոշակի AP-1 կայքից (21, 29-31): Ամբողջական FosB- ը նույն ազդեցությունն է ունենում, ինչպես ΔFosB- ն որոշակի խթանող բեկորների վրա, այլ ուրիշների վրա տարբեր ազդեցություններ: Հետագա աշխատանքը անհրաժեշտ է հասկանալ ΔFosB- ի եւ FosB- ի այս բազմազան գործողությունների հիմքում ընկած մեխանիզմները:
ΔFosB- ի համար թիրախային գեները պարզելու համար մեր խումբն օգտագործել է երկու մոտեցում: Դրանցից մեկը թեկնածու գենային մոտեցումն է: Մենք ի սկզբանե համարում էինք α-ամինո-3-հիդրօքսի-5-մեթիլ-4-իքսոքազոլպրոպիոնաթթու (AMPA) գլուտամատային ընկալիչները որպես ենթադրյալ թիրախներ `հաշվի առնելով կորիզի միջուկում գլուտամատերգիկ փոխանցման կարևոր դերը: Մինչ օրս կատարված աշխատանքը ցույց է տվել, որ AMPA- ի գլուտամատային ընկալիչների մեկ հատուկ միավոր `GluR2- ը, կարող է լինել բարեխիղճ թիրախ ΔFosB- ի համար (նկ. 2): GluR2- ի արտահայտությունը, այլ ոչ թե AMPA ընկալիչների այլ ստորաբաժանումների արտահայտությունը, ավելանում է nucleus accumbens- ում (բայց ոչ մեջքի հատվածում) ΔFosB- ի (28) գերարտահայտման արդյունքում, և գերիշխող բացասական մուտանտի արտահայտումը թուլացնում է կոկաինի ՝ սպիտակուցը դրդելու ունակությունը: Բացի այդ, GluR2 գենի խթանիչը պարունակում է համաձայնեցված AP-1 կայք, որը կապում է ΔFosB (28): GluR2- ի գերակշռումը կորիզում, վիրուսային միջնորդությամբ գեների փոխանցմամբ, մեծացնում է կենդանու զգայունությունը կոկաինի հատուցող ազդեցության նկատմամբ ՝ դրանով ընդօրինակելով ΔFosB արտահայտող մկների մեջ տեսված ֆենոտիպի մի մասի (28): GluR2– ի ինդուկցիան կարող է հաշվարկել կոկային խրոնիկական վարչարարությունից հետո միջուկային ակումբենսի նեյրոնների էլեկտրոֆիզիոլոգիական զգայունության նվազումը AMPA ընկալիչների ագոնիստների նկատմամբ (32), քանի որ GluR2 պարունակող AMPA ընկալիչները ցույց են տալիս ընդհանուր հաղորդունակության նվազում և Ca2 + թափանցելիության նվազում: Այս նեյրոնների արձագանքման նվազումը գրգռիչ միջոցների նկատմամբ կարող է հետագայում ուժեղացնել թմրամիջոցների չարաշահման արձագանքը: Այնուամենայնիվ, այն ուղիները, որոնցով դոպամիներգիկ և գլուտամատերգիկ ազդակները միջուկի ակումբենում կարգավորում են կախվածության վարքը, անհայտ են մնում. սա կպահանջի նյարդային շղթայի ըմբռնման մակարդակ, որը դեռ մատչելի չէ:
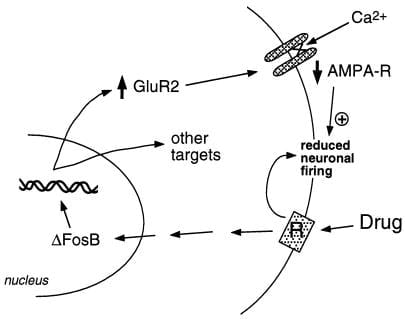
Նկար 2
AMPA գլուտամատային ընկալիչների ենթաբաժինը ՝ GluR2, ΔFosB- ի համար ենթադրյալ թիրախ է: Ուցադրվում է, թե ինչպես ΔFosB- ի միջնորդությամբ GluR2- ի ինդուկցիան կարող է փոխել nucleus accumbens նեյրոնների ֆիզիոլոգիական արձագանքը և հանգեցնել չարաշահման դեղերի նկատմամբ զգայուն պատասխանների: Համաձայն այս սխեմայի, չարաշահման թմրանյութերն առաջացնում են իրենց սուր ուժեղացնող ազդեցությունները կորիզի ակումբենս նեյրոնների արգելակման միջոցով: Կրկնակի ազդեցության դեպքում դեղամիջոցներն առաջացնում են ΔFosB, որը կարգավորում է բազմաթիվ թիրախային գեները, ներառյալ GluR2- ը: Սա մեծացնում է AMPA ընկալիչների (AMPA-R) համամասնությունը կորիզի ակումբենս նեյրոնների վրա, որոնք պարունակում են GluR2 ենթաբաժինը, ինչը առաջացնում է ընդհանուր AMPA իջեցված հոսանք և Ca2 + հոսանքի նվազում: Այս նվազեցված դյուրագրգռությունը կարող է նեյրոններն ավելի զգայուն դարձնել թմրամիջոցների սուր արգելակիչ ազդեցությունների նկատմամբ և դրանով իսկ թմրանյութերի ուժեղացնող ազդեցությունների նկատմամբ:.
ΔFosB- ի մեկ այլ կանխորոշիչ նպատակն է դինորֆինի կոդավորող գենը. Ինչպես ավելի վաղ հայտարարել էր, դինորֆինը արտահայտված է միջուկային հեծանիվների միջին ճարպային նեյրոնների ենթաբեմում, որոնք ցույց են տալիս ΔFosB- ի. Dynorphin- ը կարծես գործում է միջերեսային հետադարձ հանգույցում. Դրա թողարկումը խոչընդոտում է դոպամիներգիկ նեյրոններին, որոնք innervate միջին ցողունային նեյրոնները, միջոցով dopaminergic նյարդային տերմինալների վրա ներկա ինկոիդային ընկալիչների, ինչպես նաեւ միջուկային accumbens եւ բջջային մարմինների եւ dendrites է ventral tegmental տարածքում (Նկար 3) (33-35): Այս գաղափարը համապատասխանում է κ ռեցեպտորների ագոնիստի ունակությանը, որը ղեկավարվում է այս երկու ուղեղի շրջաններից որեւէ մեկում `նվազեցնելով թմրանյութերը:d (35):
Rիջեցման աշխատանքը ցույց է տվել, որ ΔFosB- ն նվազեցնում է դինորֆինի արտահայտությունը, որը կարող է նպաստել ΔFosB- ի ներդիրով տեսած պարգեւատրման մեխանիզմների կատարելագործմանը: Հետաքրքիր է, որ թմրամիջոցով կարգավորվող մեկ այլ գործոն, CREB (cAMP արձագանքի տարրը պարտադիր սպիտակուցը) (2, 3), հակառակ ազդեցություն է գործադրում `դինորֆինի արտանետումը միջուկի ականջի մեջ եւ նվազեցնում է կոկաինը եւ մորֆինը (4):
BCREB- ի թմրամիջոցների ներածված ակտիվացումն արագորեն տարածվում է թմրամիջոցների ներթափանցումից հետո, DYNORPHIN- ի CREB- ի եւ ΔFosB- ի փոխադարձ կարգավորումը կարող է բացատրել զսպման վաղ եւ ուշ փուլերում տեղի ունեցող փոխադարձ վարքային փոփոխությունները, բացասական զգացմունքային ախտանիշներով եւ վաղ փուլերում գերակշռող թմրանյութերի զգայունության նվազեցմամբ հետվիրահատման ժամանակ գերակշռող թմրամիջոցների գերակշռող եւ խթանող պատճառաբանությունների վրա ազդելու համար:
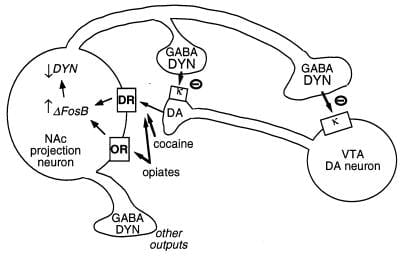
Նկար 3
Dynorphin- ը դեֆոսբի համար նպատակային թիրախ է: Ցույց է տրված վրացական tegmental տարածքը (VTA) dopamine (DA) նեյրոնային innervating մի դասի միջուկի accumbens (NAc) GABAergic նախագծման neuron, որը արտահայտում dynorphin (DYN). Dynorphin- ը ծառայում է հետադարձ կապի մեխանիզմը `դինորֆին, որը թողարկվում է NAc նեյրոնների տերմինալներից, գործում է նեյրոնային տերմինալներում գտնվող նյարդային տերմինալների եւ բջջային մարմինների վրա գործող κ opioid ընկալիչների վրա, որոնք արգելակում են դրանց գործունեությունը: ΔFosB, դինորֆինի արտահայտությունը կանխելու միջոցով կարող է նվազեցնել այդ հետադարձ կապի հանգույցը եւ բարելավել թմրամիջոցների չարաշահումները: Չի ցուցադրվում այս համակարգում CREB- ի փոխադարձ ազդեցությունը. CREB- ը դինորֆինի արտահայտությունն ուժեղացնում է, դրանով իսկ հարստացնում է թմրանյութերի շահութաբեր հատկությունները (4): GABA, γ-aminobutyric թթու; DR, դոպամինային ընկալիչ; OR, օփիոիդային ընկալիչ:
Երկրորդ մոտեցումը, որն օգտագործվում է ΔFosB- ի թիրախային գեները պարզելու համար, ներառում է ԴՆԹ միկրոհամակարգի վերլուծություն: ΔFosB- ի ինդուկցիոն գերարտահայտումը մեծացնում կամ նվազեցնում է բազմաթիվ գեների արտահայտությունը կորիզի կորիզում (36): Չնայած այժմ անհրաժեշտ է զգալի աշխատանք կատարել ՝ այս գեներից յուրաքանչյուրը որպես ΔFosB- ի ֆիզիոլոգիական թիրախներ հաստատելու և կախվածությունը ֆենոտիպի մեջ դրանց ներդրումը հասկանալու համար, թվում է, որ կարևոր նշանակություն ունի Cdk5- ը (ցիկլինից կախված կինազ -5): Այսպիսով, Cdk5- ը ի սկզբանե ճանաչվել է որպես ΔFosB- կարգավորվող միկրոռեկտորների միջոցով, իսկ հետագայում ցույց է տրվել, որ այն առաջացնում է կորիզի կորիզում և մեջքի հատվածում `կոկաինը խրոնիկական օգտագործումից հետո (37): ΔFosB- ն ակտիվացնում է cdk5 գենը գենի պրոմոուտերում առկա AP-1 կայքի միջոցով (36): Միասին, այս տվյալներն աջակցում են մի սխեմայի, երբ կոկաինը ΔFosB- ի միջոցով ուղեղի այս շրջաններում առաջացնում է Cdk5 արտահայտություն: Cdk5– ի ինդուկցիան, կարծես, փոխում է դոպամիներգիկ ազդանշանը գոնե մասամբ DARPP-32 (37) ֆոսֆորիլացման միջոցով, որը սպիտակուցի ֆոսֆատազ -1-ի ինհիբիտորից վերափոխվում է սպիտակուցային կինազի A- ի արգելակիչի վրա իր ֆոսֆորիլացման միջոցով Cdk5 (26):
ΔFOSB- ի դերը միջնորդությամբ «մշտական» պլաստիկությունը `չարաշահման թմրանյութերի նկատմամբ
Չնայած ΔFosB ազդանշանը համեմատաբար երկարատեւ է, այն մշտական չէ: ΔFosB- ը աստիճանաբար նվաստացնում է եւ այլեւս չի կարող հայտնաբերվել ուղեղի 1-2 ամսվա ընթացքում թմրամիջոցների դուրսբերման դեպքում, չնայած որոշակի վարքային աննորմալություններ պահպանվում են ավելի երկար ժամանակահատվածներում: Հետեւաբար, ΔFosB- ը չի կարող իմանալ այդ կիսամյակային վարքային անբավարարությունների միջնորդությունը: Կախվածության հետ կապված չափազանց կայուն վարքային փոփոխությունների հիմքում ընկած մոլեկուլային հարմարվողականության հայտնաբերման դժվարությունը նման է սովորելու եւ հիշողության ոլորտում առաջացած մարտահրավերներին: Թեեւ կան էլեգանտ բջջային եւ մոլեկուլային մոդելներ ուսուցման եւ հիշողության մեջ, սակայն դեռեւս հնարավոր չէ պարզել մոլեկուլային եւ բջջային հարմարվողականությունը, որոնք բավականաչափ երկարատեւ են, հաշվի առնելով բարձր կայուն վարքային հիշողությունները: Իրոք, ΔFosB- ը ամենաերկարակյաց հարմարվողությունն է, որը հայտնի է մեծահասակների ուղեղում, ոչ միայն ի պատասխան թմրամիջոցների չարաշահման, այլ նաև ցանկացած այլ խառնաշփոթի (որը չի ենթադրում վնասվածքներ): Երկու հակադրություն է զարգացել ՝ ինչպես կախվածության, այնպես էլ ուսման և հիշողության ոլորտներում ՝ հաշվի առնելով այս անհամապատասխանությունը:
Մեկ հնարավորությունն այն է, որ գեների արտահայտման ավելի անցողիկ փոփոխություններ, ինչպիսիք են ΔFosB- ի կամ այլ transcription- ի գործոններով միջնորդավորված (օրինակ, CREB), կարող են միջամտել ավելի երկարատեւ փոփոխություններ նեյրոնային մորֆոլոգիայի եւ սինապտիկ կառուցվածքի մեջ: Օրինակ, դենդրիտային հոդերի խտության աճը (հատկապես երկկողմանի կծկումների աճը) ուղեկցում է ավելացել է glutamatergic synapses- ի արդյունավետությունը երկարատեւ պոտենցիալության ժամանակ (HNUMX-38) հիփոկամպալ բրգաձեւ նեյրոնների ժամանակ եւ զուգահեռացնում է միջուկային accumbens (40) միջատային նատրիումային նեյրոնների մակարդակով միջնորդավորված վարքային զգայունության բարձրացում: Անհայտ է, թե նման կառուցվածքային փոփոխությունները բավականին երկարատեւ են, հաշվի առնելով վարքի բարձր կայուն փոփոխությունները, թեեւ վերջինս շարունակում է մնալ առնվազն 41 ամսական թմրանյութերի հեռացման համար: Վերջին փաստերը բարձրացնում են այն հնարավորությունը, որ ΔFosB- ն եւ նրա Cdk1- ի դրսեւորումը միջուկային ճառագայթների կառուցվածքների միջուկային միջամտության մեխանիզմն է միջուկային accumbens- ում (Նկար 5): Այսպիսով, Cdk4- ի ինֆիբրացիայի ներարկումն է միջուկի հագին Կոկաինի կրկնակի ազդեցության ունակությունը այս տարածաշրջանում դենդրիտային ողնաշարի խտության բարձրացման համար: Սա համահունչ է այն տեսակետի հետ, որ Cdk5- ը, որը հարստացված է ուղեղում, կարգավորում է նյարդային կառուցվածքը եւ աճը (տես `5- ը եւ 36- ը): Հնարավոր է, չնայած ապացուցված չէ, որ նյարդային մորֆոլոգիայի այդպիսի փոփոխությունները կարող են գերազանցել ΔFosB ազդանշանը:
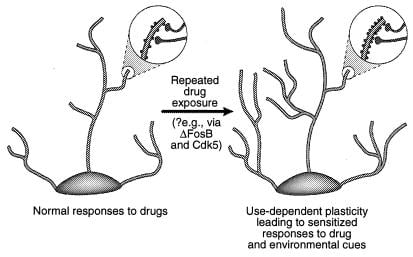
Նկար 4
Դենդրիտային կառուցվածքի կարգավորում չարաշահման դեղերով: Ուցադրվում է նեյրոնի դենդրիտային ծառի ընդլայնումը չարաշահման թմրանյութի քրոնիկ ազդեցությունից հետո, ինչպես նկատվել է կոկայինի հետ միջուկի ակումբենսում և նախաբջջային կեղևում (41): Խոշորացման գոտիները ցույց են տալիս դենդրիտային ողնաշարի աճ, որը ենթադրվում է, որ տեղի է ունենում ակտիվացված նյարդային տերմինալների հետ միասին: Դենդրիտային ողնաշարի խտության այս աճը կարող է միջնորդվել ΔFosB- ի և դրանով իսկ Cdk5- ի հետևանքով (տե՛ս տեքստը): Դենդրիտային կառուցվածքի նման փոփոխությունները, որոնք նման են որոշ ուսման մոդելների (օրինակ `երկարաժամկետ հզորացում), կարող են միջնորդել երկարատև զգայունացված արձագանքների չարաշահման թմրանյութերին կամ բնապահպանական ազդանշաններին: [Վերարտադրվում է հղման թույլտվությամբ: 3 (Հեղինակային իրավունք 2001, Macmillian Magazines Ltd.)]:
Մեկ այլ հավանականություն այն է, որ transcriptional factor- ի (օրինակ, ΔFosB, CREB) հանգեցնում է քրոմատի փոփոխության միջոցով գենի արտահայտման ավելի մշտական փոփոխություններիn. Այս եւ բազմաթիվ այլ transcription գործոնները ենթադրում են ակտիվացնել կամ հալածել թիրախային գենի տրանսկիացիան `խթանելով գիտնի հարեւանությամբ գտնվող հիստոնիկների համապատասխանաբար acetylation կամ deacetylation (42): Թեեւ հիստոնների այդպիսի acetylation եւ deacetylation կարող է ակնհայտորեն տեղի ունենալ շատ արագ, հնարավոր է, որ ΔFosB կամ CREB կարող են արտադրել ավելի երկարատեւ հարմարվողականությունը enzymatic մեքենան, որը վերահսկում է histone acetylation. ΔFosB կամ CREB- ը կարող է նաեւ նպաստել գենի արտահայտության երկարատեւ փոփոխություններին, կարգավորելով քրոմատինների այլ ձեւափոխությունները (օրինակ, ԴՆԹ կամ հիստոնի մետիլման), որոնք ենթարկվել են զարգացման ընթացքում առաջացող գենային տառատեսակների մշտական փոփոխություններին (տես `42 եւ 43) . Թեեւ այդ հնարավորությունները մնում են սպեկուլյատիվ, նրանք կարող են ապահովել մեխանիզմ, որի միջոցով անցանկալի հարմարվողականությունը չարաշահումների թմրամիջոց է (կամ որոշ այլ անհանգստություն) `հանգեցնելով կյանքի երկարատեւ վարքագծային հետեւանքների:
Սայլակ
- ↵
- Nestler EJ,
- Հուսով եմ,
- Widnell KL
(1993) Neuron 11: 995-1006:
CrossRef- ըՄԵԴԼԱՅՆվեբ գիտության մասին
- ↵
- Berke JD,
- Hyman SE- ն
(2000) Neuron 25: 515-532:
CrossRef- ըՄԵԴԼԱՅՆվեբ գիտության մասին
- ↵
- Nestler EJ- ն
(2001) Nat Rev Neurosci 2: 119-128:
CrossRef- ըՄԵԴԼԱՅՆվեբ գիտության մասին
- ↵
- Կառլզոն WA Jr,
- Thome J,
- Olson VG,
- Lane-Ladd SB- ը,
- Brodkin ES,
- Հիրոյ Ն,
- Դուման RS,
- Neve RL,
- Nestler EJ- ն
(1998) Գիտություն 282: 2272-2275:
- O'Donovan KJ,
- Tourtellotte WG,
- Millbrandt J,
- Բարբան Ջ.Մ.
(1999) Թրենդներ Neurosci 22: 167-173:
CrossRef- ըՄԵԴԼԱՅՆՎեբ գիտություն
- ↵
- Mackler SA,
- Կորութլա Լ,
- Չա XY,
- Կոեբբե Մ.Ջ.,
- Fournier KM,
- Bowers MS,
- Kalivas PW
(2000) J Neurosci 20: 6210-6217:
- ↵
- Մորգան Ջի,
- Curran T
(1995) Թրենդներ Neurosci 18: 66-67:
CrossRef- ըՄԵԴԼԱՅՆՎեբ գիտություն
- ↵
- Երիտասարդ ST,
- Porrino LJ,
- Iadarola MJ- ն
(1991) Proc Natl Acad Sci ԱՄՆ 88: 1291-1295:
- Graybiel AM,
- Մoratալան Ռ,
- Ռոբերսոն Հ
(1990) Proc Natl Acad Sci ԱՄՆ 87: 6912-6916:
- Հույս Բ,
- Կոսովսկու Բ,
- Hyman SE,
- Nestler EJ- ն
(1992) Proc Natl Acad Sci ԱՄՆ 89: 5764-5768:
- ↵
- Kelz MB,
- Nestler EJ- ն
(2000) Curr Opin Նեյրոլ 13: 715-720:
CrossRef- ըՄԵԴԼԱՅՆվեբ գիտության մասին
- ↵
- Կոոբ ԳՖ,
- Սաննա Պ.Պ.,
- Bloom FE- ը
(1998) Neuron 21: 467-476:
CrossRef- ըՄԵԴԼԱՅՆվեբ գիտության մասին
- ↵
- Իմաստուն ՀՀ
(1998) Դեղը Ալկոհոլային կախվածությունը 51: 13-22:
CrossRef- ըՄԵԴԼԱՅՆվեբ գիտության մասին
- ↵
- Հուսով եմ,
- Nye HE,
- Kelz MB,
- Ինքն DW,
- Iadarola MJ,
- Nakabeppu Y,
- Դուման RS,
- Nestler EJ- ն
(1994) Neuron 13: 1235-1244:
CrossRef- ըՄԵԴԼԱՅՆՎեբ գիտություն
- ↵
- Նայ Հ,
- Հուսով եմ,
- Kelz M,
- Iadarola M,
- Nestler EJ- ն
(1995) J Pharmacol Exp Ther 275: 1671-1680:
- Nye HE,
- Nestler EJ- ն
(1996) Մոլ Pharmacol 49: 636-645:
- ↵
- Մoratալան Ռ,
- Elibol B,
- Վալլեո Մ,
- Graybiel AM- ն
(1996) Neuron 17: 147-156:
CrossRef- ըՄԵԴԼԱՅՆվեբ գիտության մասին
- ↵
- Պիչ Էմ,
- Pagliusi SR,
- Tessari M,
- Թալաբոտ-Այեր Դ,
- Hooft van Huijsduijnen R,
- Chiamulera C
(1997) Գիտություն 275: 83-86:
- ↵
- Chen JS,
- Nye HE,
- Kelz MB,
- Հիրոյ Ն,
- Nakabeppu Y,
- Հուսով եմ,
- Nestler EJ- ն
(1995) Մոլ Pharmacol 48: 880-889:
- ↵
- Հիրոյ Ն,
- Brown J,
- Ye H,
- Սաուդու Ֆ,
- Վահիդյա Վ.Ա.,
- Դուման RS,
- Գրինբերգ ME,
- Nestler EJ- ն
(1998) J Neurosci 18: 6952-6962:
- ↵
- Chen J,
- Kelz MB,
- Հուսով եմ,
- Nakabeppu Y,
- Nestler EJ- ն
(1997) J Neurosci 17: 4933-4941:
- ↵
- Հիրոյ Ն,
- Brown J,
- Haile C,
- Ye H,
- Գրինբերգ ME,
- Nestler EJ- ն
(1997) Proc Natl Acad Sci ԱՄՆ 94: 10397-10402:
- ↵
- Fienberg AA,
- Հիրոյ Ն,
- Mermelstein P,
- Song WJ,
- Snyder GL,
- Նիշի Ա,
- Շերամին Ա,
- O'Callaghan JP,
- Միլլեր Դ,
- Cole DG,
- et al.
(1998) Գիտություն 281: 838-842:
- ↵
- Հիրոյ Ն,
- Ֆեինբերգ Ա,
- Haile C,
- Greengard P,
- Nestler EJ- ն
(1999) Eur J Neurosci 11: 1114-1118:
CrossRef- ըՄԵԴԼԱՅՆվեբ գիտության մասին
- ↵
- Greengard P,
- Ալեն ՓԲ,
- Nairn AC- ն
(1999) Neuron 23: 435-447:
CrossRef- ըՄԵԴԼԱՅՆվեբ գիտության մասին
- ↵
- Bibb JA,
- Snyder GL,
- Նիշի Ա,
- Յան Զ,
- Meijer L,
- Fienberg AA,
- Ցայ Լհ,
- Kwon YT,
- Girault JA,
- Չեռնիկ Ա.Ջ.,
- et al.
(1999) Բնություն (Լոնդոն) 402: 669-671:
- ↵
- Chen JS,
- Kelz MB,
- Zeng GQ,
- Սաքայ Ն,
- Steffen C,
- Shockett PE,
- Picciotto M,
- Դուման RS,
- Nestler EJ- ն
(1998) Մոլ Pharmacol 54: 495-503:
- ↵
- Kelz MB,
- Chen JS,
- Carlezon WA,
- Ուիսլեր Կ,
- Գիլդեն Լ,
- Beckmann AM,
- Steffen C,
- Ժանգ Յ.Ջ.,
- Մարոտտի Լ,
- Self SW,
- et al.
(1999) Բնություն (Լոնդոն) 401: 272-276:
- ↵
- Dobrazanski P,
- Noguchi T,
- Կովարի Ք,
- Rizzo CA,
- Լազո PS,
- Bravo R
(1991) Մոլ Բջջ Բիոլոգ 11: 5470-5478:
- Nakabeppu Y,
- Նաթանս Դ
(1991) Բջջային 64: 751-759:
CrossRef- ըՄԵԴԼԱՅՆվեբ գիտության մասին
- ↵
- Յենի Յ,
- Իմաստություն RM,
- Tratner I,
- Վերմա IM
(1991) Proc Natl Acad Sci ԱՄՆ 88: 5077-5081:
- ↵
- Սպիտակ FJ,
- Hu XT,
- Ժանգ XF,
- Wolf ME- ն
(1995) J Pharmacol Exp Ther 273: 445-454:
- ↵
- Hyman SE- ն
(1996) Neuron 16: 901-904:
CrossRef- ըՄԵԴԼԱՅՆՎեբ գիտություն
- Kreek MJ- ն
(1997) Pharmacol Biochem Behav 57: 551-569:
CrossRef- ըՄԵԴԼԱՅՆվեբ գիտության մասին
- ↵
- Շիպպենբերգ TS,
- Ռեյ Վ
(1997) Pharmacol Biochem Behav 57: 449-455:
CrossRef- ըՄԵԴԼԱՅՆվեբ գիտության մասին
- ↵
- Chen JS,
- Ժանգ Յ.Ջ.,
- Kelz MB,
- Steffen C,
- Ang ES,
- Զանգ Լ,
- Nestler EJ- ն
(2000) J Neurosci 20: 8965-8971:
- ↵
- Bibb JA,
- Chen JS,
- Taylor JR,
- Svenningsson P,
- Նիշի Ա,
- Snyder GL,
- Յան Զ,
- Սագավա ԶԿ,
- Nairn AC,
- Nestler EJ,
- et al.
(2001) Բնություն (Լոնդոն) 410: 376-380:
- ↵
- Լյուսեր C,
- Նիկոլլ ՀՀ,
- Malenka RC,
- Մյուլլեր Դ
(2000) Nat Neurosci 3: 545-550:
CrossRef- ըՄԵԴԼԱՅՆվեբ գիտության մասին
- Malinow R,
- Հենրիխ Մխիթարյան,
- Հայաշի Յ
(2000) Curr Opin Neurobiol 10: 352-357:
CrossRef- ըՄԵԴԼԱՅՆվեբ գիտության մասին
- ↵
- Scannevin RH,
- Huganir RL
(2000) Nat Rev Neurosci 1: 133-141:
CrossRef- ըՄԵԴԼԱՅՆվեբ գիտության մասին
Robinson, TE & Kolb, B. (1999) (1997) Eur. J. Neurosci.11, 1598-1604.
- ↵
- Կարիե Մ,
- Smale ST- ն
(2000) Eukaryotes- ում (Cold Spring Harbor Lab, Press, Plainview, NY) փոխակերպման կարգը:
- ↵
- Սպենսեր Վ.Ա.,
- Դավիթ Ջ
(1999) Gene 240: 1-12:
CrossRef- ըՄԵԴԼԱՅՆվեբ գիտության մասին
 facebook
facebook- Twitter
- Google+
- CiteULike- ը
- Համեղ
 Փոխանցել
Փոխանցել- Մենդելին
HighWire Press- ը հյուրընկալեց հոդվածներ, հղում կատարելով այս հոդվածին
- Բնական եւ դեղորայքային պարգեւատրումների մասին ակտը {Delta} FosB- ի հետ որպես սովորական միջնորդի ընդհանուր նյարդային պլաստիկության մեխանիզմների վերաբերյալ J. Neurosci. 2013 33 (8) 3434-3442
- Թմրադեղեր, հանցագործություն եւ գեդոնական ալլոստասի էպիգենետիկա Ժամանակակից քրեական արդարադատության հանդես 2012 28 (3) 314-328
- Վերացական
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Վերացական
- Ամբողջական տեքստը (HTML)
- Ամբողջական տեքստը (PDF)
- Morphine- ն ակտիվացնում է E- ի քսանհինգ տիպի transcription Factor-1 / շիճուկի արձագանքման գործոնային ուղիղը, արտածրային ազդանշանային կարգավորվող Kinases 1 / 2- ի F11 բջիջներում, որոնք առաջանում են դորալային արմատից Գանգլիա նեյրոններ J. Pharmacol. Exp. Ther. 2012 342 (1) 41-52
- Մոլեկուլային մեխանիզմը Gateway թմրանյութի համար. Էպիգենետիկ փոփոխություններ, որոնք նախաձեռնել են Nicotine Prime Gene Expression- ի կողմից կոկաինը Sci Transl մեդ 2011 3 (107) 107ra109
- Ընդլայնված Sucrose եւ Կոկաին ինքնակառավարման եւ Cue-Induced Drug Seeking հետո կորստի VGLUT2 է Midbrain Dopamine Neurons է մկների J. Neurosci. 2011 31 (35) 12593-12603
- Քրոնիկ ընդհատվող հիպոքսիան ավելացնում է արյան ճնշումը եւ FosB / Delta FosB- ի արտահայտումը կենտրոնական ինքնավար շրջաններում Ամ. J. Physiol. Ռեգուլ. Ինտեգրալ: Կոմպոզիցիա Ֆիզիոլ. 2011 301 (1) R131-R139
- GPR37 / PAEL ընկալիչի բացակայությունը վնասում է ստրիալալ Ակտի եւ ERK2 ֆոսֆորլյացիայի, Դելտա FosB արտահայտության եւ պայմանավորված տեղադրության նախապատվությունը ամֆետամինային եւ կոկաինին FASEB J. 2011 25 (6) 2071-2081
- Մոլեկուլյար հանդուրժողականության սկզբնական ալկոհոլի ազդեցության եւ կայունության տեւողության միջեւ հարաբերությունները նշանավոր են ոչ գծային J. Neurosci. 2011 31 (7) 2436-2446
- Vivo- ում Bioluminescence- ի Պատկերը հայտնաբերում է Redox- ի կարգավորվող ակտիվացնող պրոտեին-1 ակտիվացում `մկների պարավենտրիկ միջուկում, ռինովասկուլյար հիպերտենզիայի հետ Հիպերտոնիա 2011 57 (2) 289-297
- «Դոլլա» ֆոսբի ստերիլալ գերազանց ցուցադրությունը քրոնիկական Լեվոդոպա-իբխացող անազնիվ շարժումները վերարտադրում է J. Neurosci. 2010 30 (21) 7335-7343
- Էկոլոգիական ազդեցության էպիկենտրոն միջնորդությունը խոշոր հոգեբանական խանգարումների ժամանակ Շիզոֆր Բուլլ 2009 35 (6) 1045-1056
- ԴՆԹ-ի վրա հիմնված ՄՌՏ-ի փորձարկումները Ամպեթամինի քրոնիկ ազդեցության որոշման համար կենդանի ուղեղներում J. Neurosci. 2009 29 (34) 10663-10670
- Կոկաինի քաղված ջերմաստիճաններում փոփոխված դենդրիկ ողնաշարի պլաստիկությունը J. Neurosci. 2009 29 (9) 2876-2884
- Drosophila- ում գերազանց ցուցադրման էկրանը հայտնաբերում է GSK-3- ի (beta) / շագարի նեյրոնային դերը որպես AP-1- կախված զարգացման պլաստիկության կարգավորող Գենետիկա 2008 180 (4) 2057-2071
- Transcription MRI: Կենդանի ուղեղի նոր տեսակետ Neuroscientist 2008 14 (5) 503-520
- Դոկտոր Ֆոսբ Ինդուկցիան Օրբիֆրոնտալ Կորտեքս միջնորդների հանդեպ հանդուրժողականությունը կոկաինի ենթադրյալ ճանաչողական դիսֆունկցիայի J. Neurosci. 2007 27 (39) 10497-10507
- Glial բջջային գծի ստացվող նեյրոտրոֆիկ ֆունկցիայի մուտանտի մկների մեջ մետամֆետամինի որոնման վարքի վերականգնման համար խոցելի մնալը FASEB J. 2007 21 (9) 1994-2004
- Դելտա FosB- ն Nucleus Accumbens- ում կարգավորում է սննդամթերքի ամրապնդվող գործիքային վարքագիծը եւ շարժառիթը J. Neurosci. 2006 26 (36) 9196-9204
- Ֆոսֆորիլացիայի միջոցով ֆոսբ կայունության կարգավորումը: J. Neurosci. 2006 26 (19) 5131-5142
- Dopamine- ի մուտացի NMDA ընկալիչների արտահայտումը D1- ի ընկալիչ-բաղադրիչ բջիջները կանխում են կոկաինի սենսիտիզացումը եւ նվազեցնում կոկաինը նախապատվությունը J. Neurosci. 2005 25 (28) 6651-6657
- D1 Dopamine Receptors Մոդուլի {Delta} FosB Ներածություն Rat Striatum հետո Intermittent Morphine Administration J. Pharmacol. Exp. Ther. 2005 314 (1) 148-154
- Նյարդաբուծական մկների նյարդաբանությունը ընտրվում է բարձր կամընտիր անիվի հոսող ակտիվության համար Ինտեգրալ: Կոմպոզիցիա Բիոլոգ. 2005 45 (3) 438-455
- Ջրի զրկանքների եւ ռեաբիլիտացիայի ազդեցությունը c-Fos- ի եւ FosB- ի քաղցկեղի վրա, առնետի վերին վերին հատվածում եւ լամինայի տերմինալի շրջանում Ամ. J. Physiol. Ռեգուլ. Ինտեգրալ: Կոմպոզիցիա Ֆիզիոլ. 2005 288 (1) R311-R321
- FosB / {Delta} FosB Gene- ի մեխանիկական սթրեսով տառապում է Osteoblasts- ում J Biol Chem- ի 2004 279 (48) 49795-49803- ը
- Քրոնիկ սթրեսից հետո «Դելտա» FosB- ի ներդրումը հարգալից ուղեղի կառույցներում J. Neurosci. 2004 24 (47) 10594-10602
- Sim1 գենի դոզան մոդուլացնում է homeostatic կերակրման պատասխանը մկների մեջ ավելացված սննդային ճարպի ավելացման համար Ամ. J. Physiol. Էնդոկրինոլ. Մեթաբ. 2004 287 (1) E105-E113
- Մարդկային օպտիկային նյարդային աստղասրտոցներում գենային արտահայտման DNA microarray վերլուծությունը `ի պատասխան հիդրոստիկային ճնշման Ֆիզիոլ. Genomics 2004 17 (2) 157-169- ը
- Սուպերոքսիդը ներգրավված է կենտրոնական նյարդային համակարգի ակտիվության եւ սիոգրաֆիկացման մեջ, սրտամկանի ինֆարկտի պատճառով սրտի անբավարարության պատճառով Շրջ. Թեմա. 2004 94 (3) 402-409
- Ադենոզին A2A ընկալիչները ՝ նեյրոադապտացիայում կրկնվող դոպամիներգիկ խթանմանը. Պարկինսոնի հիվանդության դիսկինեզիայի բուժման հետևանքները Նյարդաբանություն 2003 61 (90116) S74-81
- Հղիության մեջ գտնվող Fos- ի հետ կապված սպիտակուցների միջուկային տեղայնացում, Rana esculenta, Testis: In Vivo- ում եւ Direct Vitro- ում Gonadotropin-Releasing Hormone Agonist- ի ազդեցությունը Բիոլոգ. Reprod. 2003 68 (3) 954-960
- Պերիադոլենտի մկների շոուն ընդլայնված Delta FosB- ի օրոք կարգավորումը `կոկայի եւ ամֆետամինի նկատմամբ J. Neurosci. 2002 22 (21) 9155-9159
- Delta FosB- ն կարգավորում է անիվի վազում J. Neurosci. 2002 22 (18) 8133-8138
- CREB- ի գործունեությունը միջուկում հագեցած շելլը վերահսկում է զգացմունքային խթանիչների վարքային արձագանքների դարպասը Proc. Նատլ. Acad. Sci. ԱՄՆ 2002 99 (17) 11435-11440
- Հոգեբենոմիկա. Հասկանալու ունակությունները J. Neurosci. 2001 21 (21) 8324-8327