Բիոլոգիական հոգեբուժություն: 2008 Դեկ 1; 64 (11): 941-50: Epub 2008 Jul 26:
Teegarden SL, Nestler EJ, Bale TL- ն:
Աղբյուր
Կենդանի բիոլոգիայի ամբիոն, Փենսիլվանիայի համալսարան, Ֆիլադելֆիա, PA 19104-6046, ԱՄՆ:
Վերացական
ՄԻՋԱՎԱՅՐԸ:
Պարգեւատրման զգայունությունը ենթարկվել է որպես թմրանյութերի չարաշահման հետ կապված վարքագծի, ինչպես նաեւ overeating- ի կանխատեսելի գործոն: Այնուամենայնիվ, անհիմն պարգեւատրման հիմքում ընկած մեխանիզմները անհայտ են: Մենք ենթադրում էինք, որ դոպամինային ազդանշանների դիսեգրումը կարող է լինել բարձրացված պարգեւատրման զգայունության հիմքում ընկած պատճառ, որի շնորհիվ խթանող խթանները կարող են գործել համակարգը կարգավորելու համար:
ՄԵԹՈԴՆԵՐ:
Մենք օգտագործում էինք գենետիկական մկնիկի մոդելը, որը բարձրացնում է պարգեւատրման զգայունության բարձր մակարդակը, Delta FosB- գերազանցող ճնշող մկնիկը, ուսումնասիրելու վարձատրության բարձր տհաճ սննդակարգի պատասխանը: Այս մկների մեջ վարձատրության ազդանշանների մարկերները հետազոտվել են հիմնականում եւ 6 շաբաթվա ընթացքում ընդունելի դիետայի ազդեցության տակ: Մկները փորձարկվել են վարքային փորձարկումից հետո, բարձր ճարպային դիետայի հանելուց հետո, այս մոդելի խոցելիությունը գնահատելու համար պարգեւատրվող խթանների հեռացումը:
ԱՐԴՅՈՒՆՔՆԵՐ
Մեր արդյունքները ցույց են տալիս փոխակերպված պարգեւատրման ուղիների ակտիվացումը միջուկի հեծանիվների երկայնքով `հիպոթալամիկ-վենդրոմային երկգեղական շրջանի սխեմաներ, որոնք հանգեցնում են Delta FosB- ի գերբնակեցման արդյունքում միջուկային հեծանիվների եւ ծայրամասային շրջաններում. Ֆոսֆորացված ցիկլային ադենոզինի մոնոֆոսֆատի (cAMP) արձագանքի տարրերի պարտադիր սպիտակուցի մակարդակները (pCREB), ուղեղի ծագմամբ նյարդոտրոֆիկ գործոն (BDNF), եւ dopamine եւ ցիկլային adenosine monophosphate կարգավորվող ֆոսֆոպրոտինը, որը 32 kDa- ի (DARPP-32) մոլեկուլային զանգվածով, Delta FosB մկների մեջ կրճատվել են միջուկային accumbens- ում, արտահայտելով կրճատված դոպամինային ազդանշանների առաջացումը: Վեց շաբաթվա ընթացքում բարձր ճարպային դիետայի ազդեցությունը լիովին բարելավեց այդ տարբերությունները, բացահայտելով խոհարարական դիետայի պոտենցիալ պարգեւատրման ունակությունը: Delta FosB մկները նույնպես ցույց են տվել լոկոմոտիվի ակտիվության եւ տագնապի հետ կապված արձագանքների զգալի աճ `24 ժամ բարձր ճարպից դուրս գալու ժամանակ:
ԵԶՐԱԿԱՑՈՒԹՅՈՒՆՆԵՐ:
Այս արդյունքները հաստատում են Delta FosB- ի եւ դոպամինային ազդանշանների դիսռեգրացման հետ կապված պարգեւների փոփոխությունների ենթադրվող զգայունություն, որոնք կարող են կարգավորվել ճաշացանկային դիետաների հետ եւ կարող է կանխատեսելի ֆենոտիպ լինել գիրության որոշ ձեւերում.
ներածություն
Չնայած նյարդային համակարգի մասին մեր աճող գիտելիքների աճին, որոնք վերահսկում են ախորժակը եւ հագեցվածությունը, ԱՄՆ-ում շարունակում են աճել գիրություն: Ընթացիկ դեղորայքի բուժումը սահմանափակ ազդեցություն ունի, եւ վարքագծային փոփոխությունները տառապում են նվազագույն երկարաժամկետ համապատասխանությունից (1): Կալորիականորեն խիտ, ուտելի սննդամթերքի սպառումը կապված է սթրեսի փոփոխությունների եւ ուղեղի պարգեւի ուղիների փոփոխության հետ, ինչը ենթադրում է, որ նման սննդամթերքի օգտակար հատկությունները կարող են գերազանցել էներգիայի հաշվեկշիռը (2-4): Ճարպի բարձր ճարպը գործում է որպես բնական պարգեւներ, ուղեղի պարգեւատրման կենտրոնները ակտիվացնելու համար `նման ձեւով չարաշահման թմրամիջոցների օգտագործման եւ որպես այդպիսին օգտագործվում են ինքնակարգավորման պարադիգմները (5-8): Այսպիսով, հավանական է, որ ծխախոտի եւ թմրամիջոցների չարաշահման համար վարքագիծը եւ մոտիվացիան ընդհանուր հիմքում ընկած մեխանիզմների մեջ, հնարավոր է, բացվեն երկու պայմանների բուժման նոր ուղիներ:
Ուսումնասիրելով սննդամթերքի եւ ուղեղների միջեւ հարաբերությունները, մենք նախկինում հայտնաբերել ենք նվազեցված պարգեւների մոլեկուլային եւ կենսաքիմիական մարկերներ եւ բարձրացնում սթրեսը `բխող բարձր ճարպային դիետայից (HF) հեռանալուց հետո: Նույնիսկ թմրամիջոցների չարաշահման թմրանյութերի նման, մեր հետազոտություններում ընդունելի դիետայի ազդեցությունը հանգեցրել է միջուկային accumbens- ի (NAc) transcriptional factor ΔFosB- ի մակարդակի բարձր մակարդակներին, 9, 10- ի կենտրոնական ուղեղի կառուցվածքը:. ΔFosB- ի սեռը, որը հստակորեն գերազանցում է արտացոլումը, ցույց է տալիս, որ սննդամթերքի պարգեւի (11) գործիքային պատասխանը մեծացնում է նրանց արժեքավոր գործիքը, պարգեւատրման զգայունության դերը եւ պարգեւատրման համակարգի երկարատեւ դեգրադացումը `ճաշացանկի մոլեկուլային եւ կենսաքիմիական պատասխաններում:
Սույն ուսումնասիրության ընթացքում օգտագործեցինք ΔFosB-overexpressing մկները `ուսումնասիրելու երկարատեւ փոփոխություններ ԱՀՀ-հիպոթալամուս-վենդրրացիային հյուսվածքային տարածքում (VTA) նյարդավիրաբույծում` ի պատասխան ճաշի ընդունելի HF- ի դիետայի: Այս պարգեւատրման զգայուն մկների մեջ կատարված նախորդ ուսումնասիրությունների հիման վրա մենք ենթադրում ենք, որ ոսպնյակների ֆունկցիոնալ փոփոխությունները կապված են պարգեւատրման զգայունության հետ `ներառելով Dopamine- ի ազդանշանի դիսեգրումը, որը հանգեցնում է VC- ի հետադարձ կապի հետադարձ կապի: Բացի այդ, մենք ենթադրում էինք, որ էներգիայի խիտ HF- ի սննդակարգի բնական պարգեւը ենթարկվում է այդ մկների մեջ նորմալ դոպամիներգիկ համակարգի, որի արդյունքում չափազանցված արձագանք է ստացել այս HF- ի սննդից դուրս գալու սթրեսից. Ուտելի կերակուր օգտագործելու յուրահատուկ առարկան, որպես վարձատրվող նյութ, թույլ է տալիս մեզ ներգրավել հիպոթալամիկ միջոցները, ֆունտիպաբաշխության սխեմաների համար, որոնք կարող են կանխատեսելի լինել բուժման դիմացկուն գիրանալու համար: Այս վարկածը ուսումնասիրելու համար մենք ուսումնասիրեցինք Dopamine neurotransmission- ի, այդ թվում `pCREB- ի, BDNF- ի եւ DARPP-32- ի մարկերները, NAc- ում եւ թիրոսինիդ հիդրոքսիլազով եւ DTA- ի դոպամինային փոխադրիչով, HF- ի ազդեցությունից հետո: Մենք նաեւ ուսումնասիրեցինք էներգետիկ հավասարակշռության որոշակի նշաններ, որոնք հայտնի են դոպամինային արտադրանքի վրա, ներառյալ ԼՏՏ-ի եւ լեյտինային եւ orexin- ի ընկալիչները, ինչպես նաեւ կողային հիպոթալամուսում օքսինի արտահայտությունը:
Նյութեր եւ մեթոդներ
Կենդանիներ
Ամերիկացի դիվանագիտական բժշկության կենտրոնում եւ Dorsal striatum- ում (Kelz et al., 1999) դինորֆին-դրական նեյրոններում ΔFosB- ը ստեղծվել է խառը ֆոնի վրա (ICR: C57Bl6 / SJL) Տեխասի հարավային բժշկական կենտրոնի համալսարանում եւ պահպանվել եւ փորձարկվել է Փենսիլվանիայի համալսարանում: Բոլոր մկները պահպանվել են դոկտիքսիիկլինում (խմելու ջրի մեջ 100 μg / մլ), մինչեւ Փենսիլվանիայի համալսարան ժամանումը: Հանգստացնելը գերազանցեցնելու համար դոկիքսցիկլին հանվեց (n = 23) (12): Վերահսկիչ մկները (n = 26) շարունակեցին ստանալ դեղը: Մկները նշանակվել են դիետիկ խմբերում ութ շաբաթ տեւողությամբ doxycyline- ի հեռացումից հետո, որի ժամանակ արտահայտությունը ցուցադրվել է առավելագույն մակարդակներում (13): Մկները պահպանվեցին 12- ի վրա, 12 թեթեւ մութ ցիկլով (լույսերը 0700- ում), սննդով եւ ջրի մատչելիությամբ: Բոլոր ուսումնասիրությունները կատարվել են Փենսիլվանիայի ինստիտուցիոնալ Կենդանիների խնամքի եւ օգտագործման կոմիտեի կողմից հաստատված փորձնական արձանագրությունների համաձայն, եւ բոլոր ընթացակարգերը իրականացվել են ըստ ինստիտուցիոնալ ուղեցույցների:
Դիետիկ ազդեցություն
Մկները պահվում էին տան տապակով (n = 16) կամ վեց շաբաթվա ընթացքում տեղադրված HF- ի վրա (n = 16-17): Տունը chow (Purina Lab Diet, Սեն - Լուի, MO) պարունակում է 4.00 կկալ / գ, որը բաղկացած է 28% սպիտակուցից, 12% ճարպից եւ 60% ածխաջրածինից: HF diet (Research Diets, New Brunswick, NJ) պարունակում է 4.73 կկալ / գ, որը բաղկացած է 20% սպիտակուցից, 45% ճարպից եւ 35% ածխաջրածինից:
Կենսաքիմիա եւ գենային արտահայտություն
Միսները վերլուծվել են վեց շաբաթ դիետայի ազդեցությունից հետո: Բանգերը հեռացան գանգից եւ սառեցված ամբողջը չոր սառույցի կամ NAc- ով (մոտավորապես 0.5- 1.75 մմ ից bregma- ից, 3.5-5.5 մմ խորության վրա) եւ սառեցված հեղուկ ազոտում: Հյուսվածքները պահվում էին մինչեւ 80 ° C մինչեւ փորձարկված:
Կենսաքիմիական անալիզներ
Արեւմտյան բլոտների մեթոդները ներկայացված են լրացուցիչ նյութերում: Օգտագործվող հակամարմինները հետեւյալն էին. Cdk5, CREB եւ BDNF (1: 500, Santa Cruz Biotechnology, Santa Cruz, CA) եւ phospho-CREB (pCREB) (Ser 133) (1: 500, Cell Signaling Technology, Danvers, MA):
Receptor autoradiography- ը
Ատորադիոգրաֆիայի մանրամասն մեթոդները նկարագրվում են լրացուցիչ նյութերում: Ligands օգտագործվում էին 2 nM H3 - SCH 23390 եւ 5 nM H3 - spiperone (PerkinElmer, Բոստոն, MA):
In situ hybridization- ը
Հյուսվածքների մշակման եւ հիբրիդացման աշխատանքները կատարվել են նախկինում նկարագրված (14): DARPP-32- ի հետախուզությունը տրամադրվել էր P. Greengard- ի կողմից (Rockefeller համալսարան) եւ J. Elmquist- ի (The Texas Southwestern Medical Center համալսարանի) հետաքննություն: DARPP-32- ի համար փորձարկված սլայդներ 3 օրվա ընթացքում ֆիլմի վրա դրվել են, իսկ 4 օրվա ընթացքում ֆիլմի համար կիրառվում են քսաներկու օրոք փորձված սլայդներ: Ֆիլմի նկարների քանակի նկարագրությունը կատարվել է նախկինում նկարագրված (10):
QRT-PCR
RNA- ն մեկուսացված էր VTA- ից եւ TaqMan- ի գեն արտահայտության փորձարկումներից (Applied Biosystems, Foster City, CA) օգտագործած անհատ գեների արտահայտությունը: Մանրամասն մեթոդները եւ վիճակագրական վերլուծությունները կարելի է գտնել լրացուցիչ նյութերում:
Վարքագծային վերլուծություններ
Դիետայի հետ կապված վարքային փոփոխությունների վրա պտուղների զգայունության հետեւանքների ուսումնասիրման նպատակով մկների ենթաբազմություն ՀՖ-ից դուրս է մնացել չորս շաբաթվա ընթացքում եւ վերադարձվել է տնային chow (n = 9 հսկողություն, n = 8 ΔFosB): Հեռանալուց հետո քսանչորս ժամ հետո, մկները ենթարկվեցին բաց դաշտային փորձությանը, համաձայն մեր նախկինում հրապարակված դիետիկ հանելուն պարադիգմը (10): Մի խոսքով, մկնիկը տեղադրվեց բաց դաշտի ապարատի կենտրոնում եւ հինգ րոպեի ընթացքում դիտարկվեց: Ընդհանուր գծի խաչերը, փետուրները, կենտրոնը եւ խաչերը կենտրոնում էին:
Վիճակագրություններ
Բոլոր տվյալները, բացառությամբ Western blots- ի, վերլուծվել են երկկողմանի ANOVA- ի միջոցով, որին հաջորդել է Fisher's PLSD թեստը `դոքսիցիկլինով բուժմամբ (ΔFosB արտահայտություն) և դիետայի պայմանով` որպես անկախ փոփոխականներ: RT-PCR վերլուծության համար օգտագործվել է իջեցված P արժեք ՝ հարակից գեների խմբերի բազմակի համեմատությունները շտկելու համար (տե՛ս լրացուցիչ նյութեր): Western բլոտները վերլուծվել են `օգտագործելով ուսանողի t-test- ը դոքսիցիկլինով բուժմամբ` որպես անկախ փոփոխական, համեմատելով օպտիկական խտությունները նույն բլոտի մեջ: Բոլոր տվյալները ներկայացվում են որպես միջին ± SEM:
Արդյունքներ
Բազային կենսաքիմիական տարբերություններ
Որպեսզի պարզաբանենք ΔFosB- գերարտահայտող մկների ուժեղ հատուցման զգայունության հիմքում ընկած մոլեկուլային ուղիները, NAc- ում հետազոտվել են մի քանի առանցքային ազդանշանային մոլեկուլների մակարդակները: ԴFosB մկների NAc- ում Cdk5- ի մակարդակի բարձրացման միտում կար `համեմատած դոքսիցիկլինի վրա պահվող աղբահավաք կենդանիների հետ (F = 5.1, P = 0.08; Նկար 1A): ΔFosB մկները զգալիորեն կրճատել են pCREB- ի մակարդակները (F = 7.4, P <0.05; Նկար 1B), ինչպես նաև CREB- ի ընդհանուր մակարդակները (F = 5.4, P = 0.05; Նկար 1C): BDNF- ի զգալի նվազում է նկատվել նաև ΔFosB մկների NAc- ում (F = 10.6, P <0.05; Նկար 1D):
Նկար 1
ΔFosB- ի գերբնակեցված մկները ցուցադրեցին ԱՀ-ում կրճատված դոպամինային ազդանշանների կենսաքիմիական նշագրերը
Սննդի ընդունման եւ մարմնի քաշը բարձր ճարպային դիետայի վրա
Մենք հաջորդ անգամ ուսումնասիրեցինք ΔFosB- գերարտահայտող մկների ազդանշանային մոլեկուլների փոփոխությունների վրա բնականաբար հատուցող HF դիետայի ազդեցությունը: Չկան տարբերություններ ΔFosB մկների և հսկողության մեջ `տան կամ HF- ի սննդի ընդունման մեջ: Այնուամենայնիվ, տեղի ունեցավ մարմնի քաշի նորմալացված կալորիականության ընդունման ընդհանուր նվազում, երբ ենթարկվեց HF- ին, որը հատուկ էր ΔFosB մկներին (F = 11.2, P <0.01; նկ. 2A): Դիետայի ազդեցության վեց շաբաթվա վերջում HF ստացող մկները զգալիորեն ավելի էին կշռում, քան չոու դիետան ունեցողները (F = 17.2, P <0.001), իսկ ΔFosB մկները ընդհանուր առմամբ ավելի քիչ էին կշռում, քան կառավարիչները (F = 5.6, P <0.05; նկ. 2): 0.05 Բ) Այս ազդեցությունը հատուկ էր չոու դիետայի խմբերի տարբերություններին (P <XNUMX):
Նկար 2
ΔFosB- ն գերազանցող ճնշումը սերմնացուի մեջ չի հայտնաբերվել սննդի ընդունման տարբերություններով, թե ճարպով կամ բարձր յուղով (HF) դիետայում
Բիոքիմիական տարբերություններ, բարձր ճարպային դիետայի վրա
Որոշել, թե ինչպես է ՀՍ-ի ազդանշանի բազային տարբերությունները կարող են փոփոխվել HF- ի դիետայի կողմից, նույն նիշային պրոտեինները, որոնք ուսումնասիրվել են բազիսում, հետազոտվել են կենդանիների կողմից, որոնք ստացել են վեց շաբաթվա HF- ն: Cdk5 մակարդակներում զգալի տարբերություններ չկար (Նկար 3A): Չափերի քանակը pCREB- ի եւ ընդհանուր CREB- ն այլեւս այլեւս 6 շաբաթվա HF- ից հետո (Նկար 3B, C): BFNF- ի մակարդակները զգալիորեն բարձրացվել են ΔFosB մկների մեջ, վեց շաբաթվա HF- ի ազդեցությունից հետո (F = 6.5, P = 0.05, Նկար 3D):
Նկար 3
Բարձր յուղ (HF) դիետան բարելավեց ազդանշանի տարբերությունները, որոնք նկատվում են ΔFosB- ի արտանետման մկների վրա
Dopamine- ի վերափոխման autoradiography
Մենք օգտագործեցինք ընկալիչների ավտորադիոգրաֆիան ՝ գնահատելու համար, թե արդյոք NAF- ում դոֆամինի ազդանշանային ΔFosB- ի կողմից հարուցված փոփոխությունները կապված են դոպամինի ընկալիչների արտահայտության փոփոխությունների հետ (նկ. 4 Ա): Բարձր յուղայնությամբ դիետան, կարծես, փոքր-ինչ ավելացնում է D1 դոպամինի ընկալիչների պարտադիր խտությունը (P = 0.14), և այս տարբերությունն ավելի մեծ էր ΔFosB մկների մոտ (նկ. 4B): Նաև կար մի միտում դեպի աճ DF- ի պարտադիր տարածքը HF- ից հետո (P = 1), և հետընտրական փորձարկումները ցույց տվեցին, որ դա նշանակալի է ΔFosB մկների մոտ (P <0.06; Նկար 0.05C): Ի տարբերություն D4 ընկալիչների ՝ D1 ընկալիչների պարտադիր խտության փոփոխություններ (հսկիչ մթնոլորտ = 2 ± 97.6, հսկիչ HF = 6.9 ± 101.1, ΔFosB շոու = 8.2 ± 91.6, ΔFosB HF = 1.0 ± 94.8) կամ պարտադիր տարածքը (հսկիչ շոու = 9.5 Դիտվել են c 47.3, հսկիչ HF = 3.4 ± 53.8, ΔFosB chow = 6.0 ± 51.9, ΔFosB HF = 3.7 ± 49.0) NAc- ում:
Նկար 4
Բարձր ճարպային դիետան (HF) հանգեցրեց D1- ի դոպամինային ընկալիչի պարտադիր փոփոխությունների եւ DARPP-32- ի արտացոլմանը ΔFosB գերազանցող ճնշման մկների միջուկի accumbens- ում (NAc)
DARPP-32 արտահայտությունը NAc- ում
In situ հիբրիդացումը օգտագործվել է NAc- ում DARPP-32- ի արտահայտման մակարդակները որոշելու համար (նկ. 4D): Բարձր յուղայնությամբ դիետան էապես բարձրացրեց DARPP-32 արտահայտությունը ուղեղի այս շրջանում (F = 5.1, P <0.05), և կար զգալի փոխազդեցություն դիետայի և ΔFosB արտահայտության (F = 8.9, P <0.05) միջև, ΔFosB մկների մոտ սննդակարգով պայմանավորված փոփոխություն (նկ. 4E): DARPP-32 արտահայտության բազային տարբերությունը հսկողության և ΔFosB մկների միջև պարզվել է հետընտրական փորձարկումներով (P <0.01), ինչպես նաև DFPB-32 մկների HF- ի վրա DARPP-0.01 արտահայտման զգալի աճով (P <XNUMX):
Գեն արտահայտությունը VTA- ում
QRT-PCR- ն օգտագործվել է VTA- ում գենի արտահայտման փոփոխությունները գնահատելու համար ՝ նպատակաուղղելով մի քանի հիմնական գեների, որոնք նախկինում ներգրավված էին պարգևի կարգավորման մեջ: Բոլոր նմուշները նորմալացվել են β-ակտինի: Ապահովելու համար, որ β-ակտինի արտահայտությունը չի փոխվել բուժման միջոցով, կատարվել է առանձին փորձություն ՝ β-ակտինը համեմատելու երկրորդ ներքին հսկողության ՝ GAPDH- ի հետ: Β-ակտինի արտահայտման մեջ էական տարբերություններ չկան (ΔCT արժեքներ, β-ակտին - GAPDH ՝ հսկիչ շոու = 2.29 ± 0.21, հսկիչ HF = 2.01 ± 0.04, ΔFosB շոու = 2.32 ± 0.49, ΔFosB HF = 2.37 ± 0.10):
Տիրոզին հիդրոքիլազի արտահայտման համար նկատվել է ΔFosB արտահայտման և դիետիկ բուժման միջև փոխազդեցության միտում (F = 3.6, P <0.06; նկ. 5A): HF- ի ազդեցության վեց շաբաթը կարծես թե նվազեցրեց تیرոզինի հիդրօքսիլազի արտահայտությունը հսկիչ մկների մեջ և ավելացրեց արտահայտումը ΔFosB մկների մեջ: Դոպամինի փոխադրիչի արտահայտման համար նկատվել է զգալի փոխազդեցություն ΔFosB արտահայտման և դիետայի ազդեցության միջև (F = 6.7, P <0.03; Նկար 5B): Տիրոզինի հիդրոքիլազայի նման, HF- ի ազդեցությունը նվազեցրեց դոպամինի փոխադրիչի արտահայտությունը հսկիչ մկների մեջ և զգալիորեն ավելացրեց արտահայտությունը ΔFosB մկների մեջ (P <0.05): Դոպամինի փոխադրիչի արտահայտման հիմնական տարբերությունը հսկողության և ΔFosB մկների միջև նշանակության չի հասել (P = 0.16), բայց 6 շաբաթ տևողությամբ HF, ΔFosB մկները արտահայտել են դոպամինի փոխադրիչի զգալիորեն բարձր մակարդակները ՝ համեմատած հսկողության հետ (P <0.05):
Նկար 5
Բարձր ճարպային դիետայի (HF) ազդեցության եւ ΔFosB արտահայտությունը հանգեցրել է փոփոխությունների մի շարք առանցքային մոլեկուլների փոփոխության VTA
Գոյություն ուներ միտում, որը ցույց էր տալիս ավելացված ΔFosB արտահայտության ազդեցությունը VTA- ում TrkB մակարդակի իջեցման համար (F = 5.7, P <0.04; Նկար 5C): Չնայած κ-օփիոիդային ընկալիչների արտահայտման վրա հիմնական ազդեցությունները չկային, ΔFosB մկների մոտ արտահայտվածության արտահայտման միտում կար (P = 0.08; նկ. 5D): Լեպտինի ընկալիչի արտահայտությունը որոշվել է նաև VTA- ում: Հայտնաբերվել է դիետայի ազդեցության զգալի ազդեցություն (F = 6.1, P <0.03), HF- ն զգալիորեն նվազեցնում է լեպտինի ընկալիչի մակարդակը VTA- ում ինչպես ΔFosB- ի, այնպես էլ հսկիչ մկների մոտ (նկ. 5E): Ուսումնասիրվել է նաև VTA- ում orexin ընկալիչի արտահայտությունը: Դիետայի զգալի ազդեցություն կար օրեքսինի ընկալիչի արտահայտման վրա (F = 1, P <9.0), HF- ի ազդեցության տակ գտնվող մկների մոտ VTA- ում ավելի բարձր մակարդակ էին արտահայտում (նկ. 0.02F): ΔFosB մկների մոտ նաև ուղեղի այս շրջանում 5 – ի ընդհանուր ավելի բարձր մակարդակ արտահայտելու միտում կար (P <1):
Օրեկինի արտահայտությունը կողային հիպոթալամուսում
Մենք չափեցինք orexin- ի մակարդակները կողային հիպոթալամուսում, VTA- ի orexinergic innervation- ի ծագումը, in situ հիբրիդացման միջոցով (նկ. 6 Ա): Fգալի փոխազդեցություն էր ΔFosB արտահայտման և օրեքսինի արտահայտման վրա դիետայի ազդեցության միջև (F = 9.1, P <0.01), HF- ն զգալիորեն մեծացնում էր հանքաքարի մակարդակը հսկիչ մկների մեջ (P <0.05) և արտահայտման նվազումը ΔFosB մկների մեջ (նկ. 6B): Չնայած բազալ վիճակում orexin- ի արտահայտման մեջ էական տարբերություններ չկային, 6 շաբաթ HF- ից հետո, ΔFosB մկները արտահայտում էին orexin- ի զգալիորեն նվազեցված մակարդակները ՝ համեմատած հսկողության հետ (P <0.05):
Նկար 6
Բարձր ճարպային (HF) դիետան ունեցել է տարբերակիչ ազդեցություն, որը գտնվում է վերահսկողության մեջ (Ctrl) եւ ΔFosB գերազանցող ճնշման մկների
Beհավասարակշռված վերլուծություններ
Դիետայի փոփոխության արդյունքում գրգռման և հուզականության փոփոխությունները գնահատելու համար մկները ենթարկվել են բաց դաշտի փորձարկման HF դիետայի դադարեցումից հետո 24 ժամ անց (10): Ընդհանուր գծերի խաչերի վրա, որոնք գնահատվել են որպես գրգռման միջոց, էապես ազդել են ΔFosB արտահայտությունը (F = 6.6, P <0.05) և դիետան (F = 4.6, P <0.05; Նկար 7A): ΔFosB մկներն ավելի ակտիվ էին նոր միջավայրում, քան զսպանակները, իսկ հետընտրական փորձարկումները ցույց տվեցին, որ HF- ից դուրս բերված մկները զգալիորեն ավելի ակտիվ էին, քան նրանք, ովքեր ենթարկվում էին chow (P <0.05): Ֆեկալային բոլին հաշվարկվում էր որպես անհանգստության նման պահվածքի միջոց (10): ΔFosB- ի արտահայտման հիմնական ազդեցությունը կար (F = 10.2, P <0.01), ΔFosB- գերարտահայտող մկների մոտ նոր ֆեկալային բոլիներ արտադրելով վեպի միջավայրում, մասնավորապես տնային պայմաններում և HF- ի դուրսբերման խմբերում (նկ. 7 Բ): HF դիետայի վրա պահվող ΔFosB մկները ավելի քիչ ֆեկալային բոլի են առաջացրել, քան նրանք, ովքեր պահպանվել են ցնցուղի վրա, և նրանք, ովքեր դուրս են բերվել թեստից 24 ժամ առաջ: Հավանաբար, վերահսկիչ մկների վրա դիետա չի ազդել: Բաց դաշտի կենտրոնում անցկացրած ժամանակի վրա չկային ինչպես ΔFosB արտահայտության, այնպես էլ դիետայի էական հետևանքներ (հսկիչ մթնոլորտ = 14.5 3.1 վ, վերահսկիչ HF = 18.0 ± 3.2 վ, հսկողություն W / D = 15.4 ± 1.9 վ, ΔFosB շոու = 16.9 ± 2.4 վրկ, ΔFosB HF = 13.1 ± 3.9 վրկ, ΔFosB W / D = 19.8 ± 2.6 վրկ):
Նկար 7
ΔFosB- ի նկատմամբ արտահայտված մկները ավելի զգայուն էին բարձր ճարպային դիետայի (ՀՖ) հեռացման հետեւանքների նկատմամբ
Քննարկում
Շագանակագեղձի բուժման մեջ կան կարեւոր գործոններ այնպիսի գործոնների բացահայտման համար, որոնք ազդում են overeating եւ քաշի ձեռքբերման նկատմամբ: Ուղեղի վարձատրության ուղիները կարեւոր դեր են խաղում ճաշացանկերի եւ դիետիկ փոփոխությունների (6, 10, 15, 16) շարժառիթների եւ պատասխանների մեջ: Քանի որ orexigenic եւ anorexigenic ազդանշանները կարող են ուղղակիորեն ազդել պարգեւատրում ազդանշաններ միջոցով hypothalamus-VTA-ՆԱԿ միացում, լուսաբանումը գեների արձագանքելով էներգետիկ հարուստ հաճելի դիետաները ներսում պարգեւատրության կենտրոններում կարող տրամադրել գրողներին բուժական թիրախները գիրություն բուժման (17, 18): Այդ իսկ պատճառով, մենք ուսումնասիրվել կենսաքիմիական եւ մոլեկուլային մարկերներ պարգեւատրման եւ էներգետիկ հաշվեկշռի ազդանշանային երկայնքով hypothalamus-VTA-ՆԱԿ circuit ի պատասխան մի HF դիետայի մեջ ΔFosB-overexpressing մկների որպես մոդել ուժեղացված զգայունության փոփոխությունների պարգեւատրության (13, 19, 20) , եւ դիետայի հանելուց հետո վարքային զգայունությունը: Մենք ենթադրում էինք, որ ΔFosB մկների մեջ դոպամինի ազդանշանների բազային դիսեգրումը կկարգավորվի HF- ի սննդակարգի հատուցող հետեւանքներով, որը ներառում է էներգետիկ հավասարակշռության ազդանշանների եւ դոպամինային համակարգի խաչմերուկ:
Քննարկելու NAc- ում դոպամինային ազդանշանի դեզրկման մասին նշող նշողներին, մենք ուսումնասիրեցինք D1 ընդունիչի մակարդակները եւ ստորին հոսանքները: Չնայած D1- ի ընկալիչների կապում զգալի տարբերություններ չկան, ՀՖ-ի ազդեցության միտում կար ΔFosB մկների մեջ ամրապնդող տարածքի ավելացման համար. Դա հետաքրքիր է, քանի որ ΔFosB- ի տուբերկուլյոզի պրեպարատը եւ բնական պարգեւները առաջանում են հիմնականում միջին սպիրտային նեյրոնների դինորֆինի դրական ենթատիպում, որոնք հիմնականում արտահայտում են D1 ընդունիչները (9, 21): Անջրանցիկ դոպամինային ազդանշանային թիրախի pCREB մակարդակները զգալիորեն նվազել են ΔFosB մկների մեջ, աջակցելով այս ուղեղի շրջանում նվազեցված D1 ընդունիչի ակտիվացմանը (22, 23): Հետաքրքիր է նաեւ, որ մենք հայտնաբերեցինք ΔFosB մկների մեջ CREB- ի ընդհանուր մակարդակի զգալի նվազում `առաջարկելով դոպամինային ազդանշանների փոխանցման հետագա կրճատում, որը կարող է երկրորդական լինել pCREB- ի (24) երկարատեւ նվազման արդյունքում: BDNF արտահայտությունը կարգավորվում է pCREB- ի կողմից, բարձրացված է D1- ի ակտիվացման միջոցով եւ հանդիսանում է NAc- ի (25, 26) վարձատրության հետ կապված նյարդապլաստիկության կարեւոր միջնորդ: Հետեւաբար, ΔFosB մկների NAc- ում հայտնաբերվել է BDNF սպիտակուցի զգալի նվազում:
NAc- ի բոլոր միջին ցողունային նեյրոններն արտահայտում են DARPP-32 (27): Դրա մեծ թվով հոսքագծերի ազդեցությունը այն դարձնում է կենսական նշանակության նվագարկիչ (28), եւ այն ենթարկվել է թմրամոլության եւ դոպամինային համակարգի ներգրավման այլ խանգարումների, ներառյալ `ազդող խանգարումների եւ շիզոֆրենիայի (27, 29): DARPP-32- ի արտահայտությամբ մենք բացահայտեցինք խորը բազային կրճատումներ ΔFosB մկների NAc- ում: DARPP-32 արտահայտությունը կարգավորվում է BDNF- ի կողմից, ուստի նվազեցված արտահայտությունը կարող է անմիջականորեն կապված լինել ΔFosB մկների (27, 29, 30) հայտնաբերված BDNF մակարդակների կրճատումների հետ: Նույնիսկ DARPP-32- ի ֆոսֆորացված վիճակում չափավոր փոփոխությունները կարող են հանգեցնել Էկոլոգիական ազդանշանների էական փոփոխությունների NAc- ում (27): Նախորդ ուսումնասիրությունները ցույց են տվել, որ DARPP-32 սպիտակուցը փոփոխություն չի կատարում ΔFosB մկների մեջ, հետեւելով 12-wk- ից դոկիսիսիկլինից, երբ ավելի լայն ստիատալային գնահատում է կատարվել (31), առաջարկելով, որ ΔFosB- ի ազդեցությունը DARPP-32- ի վրա կարող է լինել ժամանակի եւ շրջանի հատուկ:
Մենք ենթադրում էինք, որ ΔFosB մկների NAc- ում դոպամինի ազդանշանների ցուցանիշների կտրուկ նվազումը հավանաբար ներգրավված է ՎՏԱ դոպամինային նախագծման նեյրոնների փոփոխություններով, չնայած ΔFosB- ը չի գերազանցում այդ նեյրոնների մեջ. Հետեւաբար, մենք ուսումնասիրեցինք ՎՏԱ-ում դոպամինային առնչվող գեների արտահայտությունը, ներառյալ թիրոսինե հիդրոքսիլազը եւ դոպամինի փոխադրիչը: Տիրոսինի հիդրոքսիլազի եւ դոպամինի փոխադրիչի մակարդակները դրականորեն փոխկապակցված են դոպամինի արտադրության հետ: ΔFosB մկների համար միտում էր, որ ցուցադրվում է կրճատվող թիրոսինային հիդրոխիլազի եւ դոպամինային տրանսպորտային միջոցի զգալի կրճատման դեպքում `դոկտորային դոպամինային ազդանշանների դիսեգրումը. Քանի որ այս դոպամինային գեների վրա հիմնված այս բազային կրճատումները ΔFosB մկների ՎՏԱ-ում ենթադրաբար արտացոլում են փոփոխված արձագանքները NAc- ի երկարաժամկետ ΔFosB գերբնակեցման ընթացքում, մենք ուսումնասիրեցինք BDNF- ի ընկալիչի, TrkB- ի արտահայտությունը, որպես VTA- ի (32) NAc- ի հետադարձ կապի հնարավոր մեխանիզմ: Tyrosine hydroxylase- ի եւ դոպամինի փոխադրիչի նման, TrkB արտահայտությունը ցույց է տվել, որ հիմնականում իջեցվել է ΔFosB մկների մեջ, որը չի հասնում նշանակությունը, երբ բազմակի համեմատությունների համար ուղղվում է: BDNF-TrkB համալիրը կարող է հետադարձ ճանապարհով տեղափոխվել եւ գործել VTA- ի շրջանում, ազդելու տեղային գենի արտահայտման վրա եւ խթանելու բջիջների աճը եւ պահպանումը (33): Բացի այդ, NAC- ի ներսում կանխարգելիչ TRKB- ի BDNF ակտիվացումը կարող է ուղղակիորեն խթանել դոպամինային նյարդոտմանը (32), որն աջակցում է այդ մկների դոպամինային ազդանշանների նվազեցմանը:
Κ-opioid ընկալիչների դինորֆինի ակտիվացումը կարգավորում է դոպամինային ազդանշանը եւ եւս մեկ մեխանիզմ է, որով ԱՀԿ-ն արձագանքում է VTA- ին (34): Մենք հայտնաբերեցինք, որ VTA- ում κ-opioid- ի ընկալիչ արտահայտությունը ցույց է տվել, որ ΔFosB մկների մեջ կրճատվել է միտում: Քանի որ ΔFosB overexpression- ը ցուցադրվել է NAc- ում (20) դինորֆինի արտահայտությունը նվազեցնելու համար, ΔFosB մկները կարող են խորը նվազել VTA κ-օփիոիդային ակտիվացում: Չնայած դինորֆինի ազդանշանը սովորաբար խանգարում է դոպամինի նեյրոնների վրա , 35): Դինորֆին - κ-օփիոիդային համակարգի դիսարգելումը կապված է թմրամիջոցների չարաշահման ձեռքբերման և պահպանման հետ, ինչը նպաստում է օպիոիդային ազդանշանային համակարգի կարևոր հավասարակշռությանը դոպամինի ուղիների նորմալացման գործում: (38).
Հիմք ընդունելով էներգետիկ խիտ HF- ի դիետայի պարգեւատրման ունակությունը, մենք ենթադրում էինք, որ ΔFosB մկների մեջ դոպամինի եւ օփիոիդային վարձատրության դիսռոգումը կարող է կանխատեսել այդ մկների համար նմանատիպ դիետա բարձրացնելով պարգեւատրման պատասխանները `դրանով կարգավորելով պարգեւատրման համակարգը հիպոթալամուսի ակտիվացման միջոցով -ՎՏԱ-NAc միացում. Դիետայի վեց շաբաթվա ընթացքում սննդային ընդունման մեջ ΔFosB- ի եւ հսկողության մկների միջեւ որեւէ տարբերություն չի նկատվում, այն փաստը, որ ΔFosB մկների մեջ կենսաքիմիական եւ մոլեկուլային մարկերներում հայտնաբերված փոփոխությունները չեն պայմանավորված սպառված կալորիաների տարբերությունները: Ինչպես եւ սպասվում էր, basal տարբերություններ հայտնաբերվել pCREB, ընդհանուր CREB, BDNF, DARPP-32, եւ κ-ափիոնային ընկալիչների մակարդակների միջեւ ΔFosB եւ վերահսկման մկների են թուլացնելու, ամենայն հավանականությամբ, պայմանավորված է աճել dopamine արտադրության ΔFosB մկների վրա HF (29, 39-41) .
Երկու թիրոսինային հիդրոքսիլազի եւ ՎՏԱ-ի դոպամինային փոխադրիչի հետազոտությունը բացահայտում են ΔFosB- ի եւ վերահսկողության մկների զարմանալի հակազդեցությունը. Վերահսկիչ մկները ցույց տվեցին, որ թիրոսինային հիդրոքսիլազի եւ դոպամինի փոխադրիչի արտահայտումը նվազում է, իսկ ΔFosB մկները ցույց են տալիս այդ երկօպամինային առնչվող երկու գեների աճի արտահայտությունը: Հետաքրքիր է, թիրոսինային հիդրոքսիլազի արտահայտությունը փոփոխվում է VTA- ում `քրոնիկ կոկաինի կամ մետամֆետամինի կիրառմամբ (42-44)` առաջարկելով, որ ΔFosB մկները կարող են ՀՖ-ի բնական պարգեւը ավելի հստակ գտնել, քան վերահսկողության մկները:
Որպեսզի ուսումնասիրել, թե ինչպես կարող է VTA- ին հնարավոր հիպոթալամիկ ներդրումը կարող է փոխանցել ազդանշաններ, որոնք արտացոլում են էներգետիկ հավասարակշռությունը, ուսումնասիրվել են նաեւ լեպրինի ընդունիչի եւ orexin reseptor-1- ի արտահայտությունը: Լյարդի շրջանագծային մակարդակները ավելանում են HF- ով, իսկ լապտինը կարող է իր հերթին գործել ՎՏԱ-ում, փոխելու դոպամինային ազդանշանը (18, 45): VTA- ի leptin- ի ընկալիչի արտահայտությունը նմանապես նվազել է HF- ի եւ ΔFosB- ի եւ հսկողության մկաններում `համապատասխան քաշի եւ դիետայի ընդունման դեպքում` HF- ում: Բարձր ճարպը նույնպես բարձրացրեց orexin reseptor-1- ի արտահայտությունը եւ ΔFosB- ի եւ վերահսկիչ մկների VTA- ում: Orexin- ը ակտիվացնում է ՎՏԱ-ի դոպամին նեյրոնները, նպաստում է VTA պլաստիկությանը եւ դանդաղեցնում է dc- ի մակարդակը (46-48): Բարձր ճարպային դիետան ցույց է տվել, որ մկների մեջ արտահայտվում է orexin արտահայտությունը, մեր դիտարկումների (49, 50): Այսպիսով, ավելցուկի վերափոխման արտահայտությունը, ինչպես նաեւ ՎՏԱ-ում լեյտինային ազդանշանների փոփոխությունները կարող են խթանել դիետային պարգեւը եւ ΔFosB- ում եւ վերահսկողության մկաններում, որոնք աջակցում են էներգիայի հավասարակշռության ազդանշանների փոխանցման ուղիների միջեւ եւ անմիջապես կապված վարձատրության հետ:
Պրոֆեսորների ստացման սթրեսային ազդեցությունները ուսումնասիրելու համար մկները հետազոտվել են HF- ի հեռացումից հետո 24 ժամ բաց դաշտում: ΔFosB մկները ավելի զգայուն էին նախընտրելի դիետայի հեռացման սուր հետեւանքների նկատմամբ, որոնք ցույց են տալիս բարձրորակ արյունահոսություն եւ վակուումային բոլի արտադրությունը վեպի բաց դաշտում `համեմատած մյուս բոլոր դիետայի եւ այլ խմբերի հետ: ΔFosB մկները նույնպես ցույց են տվել, որ այս փորձարկման մեջ հետաքրքիր վարքագծի օրինակ է տրվում պարգեւի եւ սթրեսային զգայունության մասին, իսկ HF- ի դիետան ի սկզբանե նվազեցնելով fecal boli արտադրությունը chow- ի նկատմամբ, իսկ վերացումը եւս մեկ անգամ բարձրացնում է այս անհանգստությունը: Այս դիտարկումը բաց դաշտային գործունեության մեջ չի համընկնում օրեքսինի արտահայտման փոփոխությունների հետ, առաջարկելով սթրեսից բխող արցունքի փոխհարաբերությունը, որը ոչ միայն փոխկապակցված ազդանշանների փոփոխությունների ազդեցություն է: Ընդհանուր առմամբ, այս տվյալները նպաստում են մեր վարկածին, որ ΔFosB մկները ավելի զգայուն կլինեն նախընտրելի դիետայի հեռացման սուր հետեւանքների նկատմամբ, քանի որ բարձրացնում են պարգեւատրման զգայունությունը.
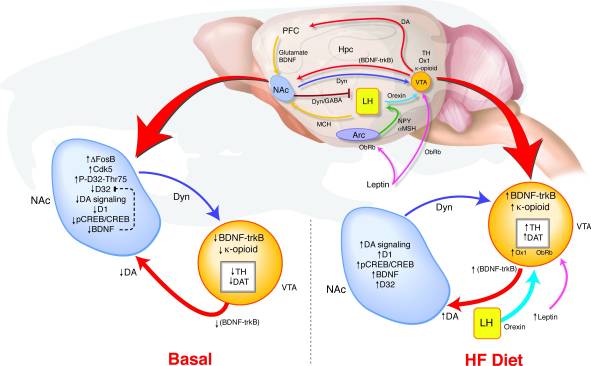
Ինչպես է ԱՀ-ում ΔFosB- ի երկարաժամկետ գերբնակեցումը հանգեցնում է այնպիսի փոփոխությունների, ինչպիսիք են վարքի եւ վարձատրության ազդանշաններըԴեպի Մենք առաջարկեցինք ՎՏԱ-ի համընկնող հայտնաբերման մոդել, որտեղ NAC- ից եւ հիպոթալամուսից ստացվող փոխադարձ արձագանքները ազդում են այն մասին, թե ինչպես կարելի է որոշել դոպամինային համակարգի կարգավորումը, որը կարող է նպաստել պարգեւատրման ուղիների դեգրադացիայի եւ ճարպակալման կանխարգելման կապին (Նկար 8): HF- ի ազդեցության ընթացքում բազմաթիվ էներգետիկ միջոցներ, որոնք արտացոլում են ինչպես էներգետիկ հավասարակշռությունը, այնպես էլ պարգեւատրում պետությունը, միավորվում են VTA- ում: Լեպտինի եւ օրեքսինի ազդանշանների ավելացումը, ինչպես նաեւ ԱԿԿ-ից դեպի կողային հիպոթալամուսի փոփոխված արձագանքը կարող է ազդել ինչպես այս orexigenic ազդանշանները արձագանքում են HF- ին ΔFOSB մկների (17, 18, 45, 47, 51-53): BDNF- ի բարձր ճարպային դիետա-իջեցման բարձրությունները կարող են տրամադրել պարգեւատրման հետադարձ կապ ՎՏԱ-ին, հետագայում նպաստելով դոպամինային առնչվող գեների արտահայտման փոփոխություններին:
Նկար 8
Բարձր յուղ (HF) դիետան նորմալացնում է ΔFosB մկների դիսռեգրված վարձատրության ազդանշանները
Այս արդյունքները սահմանում են պարգեւատրման զգայունության մոլեկուլային մարկերներ եւ նշում, որ դոպամինային համակարգի երկարատեւ քայքայումը կարող է կանխատեսել անհատին կախվածություն եւ գիրություն: Հետագայում այդ տվյալները կարեւոր քայլ են մղում պոտենցիալ նոր թերապեւտիկ թիրախները գիրության եւ այլ խանգարումների բուժման եւ կանխարգելման համար, որոնք կարող են կենտրոնանալ վարձատրության համակարգում: Հետագայում կարեւոր է ուսումնասիրել, թե ինչպես է այս համակարգը արձագանքում HF- ի սննդակարգի հեռացմանը, ինչպես նաեւ հետազոտելու սեռական տարբերությունները զգայունության եւ բարձր ճարպային դիետայի ազդեցության զգայունության հարցում:
Լրացուցիչ նյութեր
Supp. Մեթոդներ
Սեղմեք այստեղ դիտելու համար (61K, doc)
Acknowledgments
Հեղինակները ցանկանում են շնորհակալություն հայտնել Քեթի Շֆֆենին `կենդանական ծագման եւ փոխանցման օգնության համար: Այս աշխատանքը ֆինանսավորվել է Փենսիլվանիայի շաքարային դիաբետի համալսարանի (DK019525) դրամաշնորհով եւ Հոգեկան առողջության ազգային ինստիտուտի (R01 MH51399 եւ P50 MH66172) եւ Թմրամիջոցների չարաշահման ազգային ինստիտուտի (R01 DA07359) դրամաշնորհներով:
Հղումներ
Ֆինանսական բացահայտումներ. Բոլոր հեղինակները հայտարարում են, որ չունեն կենսաբժշկական ֆինանսական շահերը կամ հնարավոր շահերի բախումները:
Սայլակ
1. Wadden TA, Berkowitz RI, Womble LG, Sarwer DB, Phelan S, Cato RK, Hesson LA, Osei SY, Kaplan R, Stunkard AJ: Կենսակերպի փոփոխման եւ շաքարախտի դեղագործության ռանդալիզացված փորձարկում: N Engl J Med. 2005; 353 (20): 2111-20 [PubMed]
2. Blendy JA, Strasser A, Walters CL, Perkins KA, Patterson F, Berkowitz R, Lerman C. Կրճատվել է նիկոտինային պարգեւը գիրության մեջ `խաչաձեւ համեմատություն մարդու եւ մկնիկի մեջ: Psychopharmacology (Berl) 2005
3. Franken IH, Muris P. Պարգեւատրման զգայունության անհատական տարբերությունները կապված են առողջ կանանց սննդի խնամքի եւ հարաբերական մարմնի քաշի հետ: Ախորժակ: 2005; 45 (2): 198-201 [PubMed]
4. Kelley AE, Berridge KC- ն: Բնական պարգեւների նեվոլիզմը. J Neurosci. 2002; 22 (9): 3306-11 [PubMed]
5. Cagniard B, Balsam PD, Brunner D, Zhuang X. Խրոնիկ բարձրացված դոպամինով մկները ցուցադրվում են ուժեղացված մոտիվացիա, բայց ոչ սովորում, սննդի պարգեւի համար: Նյարդաբանության ֆակուլտետ: 2006; 31 (7): 1362-70 [PubMed]
6. Liang NC, Hajnal A, Norgren R. Sham կերակրման եգիպտացորենի նավթը մեծացնում է accumbens dopamine է առնետի. Բնապահպանական ֆիզիոլոգիա: 2006; 291 (5): R1236-9 [PubMed]
7. Մենդոզա Ջ, Անջելես-Կաստելլանոս Մ, Էսքոբար Ս. Ճաշացանկով ճաշացանկը առաջացնում է սննդամթերքի սպասողական գործունեություն եւ ուղեղի պարգեւատրման վայրերում c-Fos արտահայտություն: Neuroscience. 2005; 133 (1): 293-303 [PubMed]
8. Շրյոդեր Բ., Բիլզակ Ջ.Մ., Քելլի Է.Է. Նիկոտինի կամ շոկոլադի համակցված կոնտեքստային առարկաների ազդեցության հետեւանքով նախածրագրային կորտիկ ակտիվացման ընդհանուր պրոֆիլը: Neuroscience. 2001; 105 (3): 535-45 [PubMed]
9. Nestler EJ, Barrot M, Self DW- ն: DeltaFosB- ը կախվածության մեջ մշտական մոլեկուլային անջատիչ է: Proc Natl Acad Sci ԱՄՆ Ա. 2001; 98 (20): 11042-6: [PMC անվճար հոդված] [PubMed]
10. Teegarden SL, Bale TL- ն: Դիետիկ նախապատվության նվազում է առաջացնում աճող հուզական եւ ռիսկային ռեակցիա: Բիոլոգիական հոգեբուժություն: 2007; 61 (9): 1021-9 [PubMed]
11. Olausson P, Jentsch JD, Tronson N, Nestler EJ, Taylor JR- ը: dFosB- ն Nucleus Accumbens- ում կարգավորում է սննդամթերքի ամրապնդվող գործիքային վարքագիծը եւ մոտիվացումը: Նյարդաբանության ամսագիր: 2006; 26 (36): 9196-9204 [PubMed]
12. Chen J, Kelz MB, Zeng G, Sakai N, Steffen C, Shockett PE, Picciotto MR, Duman RS, Nestler EJ: Տրանսգենային կենդանիներ, որոնք ուղեկցվում են ընկճված, նպատակային գենային արտահայտությամբ: Մոլ Pharmacol. 1998; 54 (3): 495-503 [PubMed]
13. Քել Ջի, Քեն Ջ, Կառլիզոն Վ.Ա., Ջր, Ուիսլեր Ք, Գիլդեն Լ, Բեքման AM, Steffen C, Zhang YJ, Marotti L, Self DW, Tkatch T, Baranauskas G, Surmeier DJ, Neve RL, Duman RS, Picciotto MR, Nestler EJ- ն: ԴելտաՖոսԲ-ի փոխանցման գործոնի արտահայտումը ուղեղում վերահսկում է կոկաինի զգայունությունը: Բնություն: 1999; 401 (6750): 272-6 [PubMed]
14. Bale TL, Dorsa DM- ն: Էսստրոգինի սեքսի տարբերությունները եւ հետեւանքները օքսիտոցինի ընկալիչի սուրհանդակային ռիբոնուկլեինային թթվի արտահայտման համար `ventromedial hypothalamus- ում: Էնդոկրինոլոգիա: 1995; 136 (1): 27-32 [PubMed]
15. Avena NM, Long KA, Hoebel BG- ը: Շաքարի կախված առնետները ցույց են տալիս, որ շաքարավազը շեղումից հետո ուժեղացնում է շաքարավազի զրկանքների ազդեցությունը: Physiol Behav- ը: 2005; 84 (3): 359-62 [PubMed]
16. Will MJ, Franzblau EB, Kelley AE. Nucleus accumbens mu-opioids- ը կարգավորում է բոված ուղեղի ցանցի ակտիվացման միջոցով բարձր ճարպային դիետայի ընդունումը: J Neurosci. 2003; 23 (7): 2882-8 [PubMed]
17. Zheng H, Patterson LM, Berthoud HR- ն: Վիրահատական շրջանում Orexin ազդանշան պահանջվում է բարձր ճարպային ախորժակի համար, որը հանգեցնում է միջուկի երիկամների օփիոիդային խթանմանը: J Neurosci. 2007; 27 (41): 11075-82 [PubMed]
18. Hommel JD, Trinko R, Sears RM, Georgescu D, Liu ZW, Gao XB, Thurmon JJ, Marinelli M, DiLeone RJ: Լեպտինային ընդունիչն ազդում է միջին խոռոչի դոպամինային նեյրոնների կողմից, կարգավորում է սնուցումը: Նյարդոն: 2006; 51 (6): 801-10 [PubMed]
19. Colby CR, Whisler K, Steffen C, Nestler EJ, Self DW- ն: DeltaFosB- ի ստերիլալ բջջային տիպի հատուկ գերբնակեցումը նպաստում է կոկաինի խթանմանը: J Neurosci. 2003; 23 (6): 2488-93 [PubMed]
20. Զաչարիո Վ, Բոլանոս CA, Selley DE, Theobald D, Cassidy MP, Kelz MB, Shaw-Lutchman T, Berton O, Sim-Selley LJ, Dileone RJ, Kumar A, Nestler EJ: ԴելտաՖոսԲ-ի համար կարեւոր դեր է մորֆինային ակցիայի միջուկի հագուկապի մեջ: Nat Neurosci. 2006; 9 (2): 205-11 [PubMed]
21. Lee KW, Kim Y, Kim AM, Helmin K, Nairn AC, Greengard P. Կոկաինի կողմից առաջացած dendritic ողնաշարի ձեւավորում D1- ում եւ D2 dopamine receptor- ի պարունակող միջին ճարպային նեյրոնների միջուկում accumbens- ում: Proc Natl Acad Sci ԱՄՆ Ա. 2006; 103 (9): 3399-404: [PMC անվճար հոդված] [PubMed]
22. Blendy JA, Maldonado R. Թմրամոլության գենետիկական վերլուծություն. CAMP արձագանքի տարրերի պարտադիր սպիտակուցի դերը: J Mol Med. 1998; 76 (2): 104-10 [PubMed]
23. Nestler EJ- ն: Թմրամոլության մոլեկուլային մեխանիզմները: Neuropharmacology. 2004; 47 1- ը, 24-32- ը: [PubMed]
24. Tanis KQ, Duman RS, Նյուտոն ՍՍ: CREB- ի պարտադիր եւ գործառույթը ուղեղի մեջ. Տարածաշրջանային առանձնահատկություն եւ էլեկտրոկոնվուլյուցիոն զավթման ներածություն: Բիոլոգիական հոգեբուժություն: 2007
25. Կումար Ա, Չոյի Ք, Ռենտալ Վ, Ցանկովա Ն.Մ., Theobald DE, Truong HT, Russo SJ, Laplant Q, Sasaki TS, Whistler KN, Neve RL, Self DW, Nestler EJ: Chromatin- ի վերափոխումը կոտրատորի մեջ կոկաինը ներգործող պլաստիկության հիմքում ընկած հիմնական մեխանիզմն է: Նյարդոն: 2005; 48 (2): 303-14 [PubMed]
26. Graham DL, Edwards S, Bachtell RK, Dileone RJ, Rios M, Self DW- ն: Դինամիկ BDNF- ի գործունեությունը կոկաինի օգտագործման հետ կապված միջուկի հագեցվածության մեջ մեծացնում է ինքնակազմակերպումը եւ վերականգնումը: Nat Neurosci. 2007; 10 (8): 1029-37 [PubMed]
27. Svenningsson P, Nairn AC, Greengard P. DARPP-32 միջնորդում է չարաշահման բազմաթիվ թմրանյութերի գործողությունները: Aaps J. 2005; 7 (2): E353-60: [PMC անվճար հոդված] [PubMed]
28. Palmer AA, Verbitsky M, Suresh R, Kamsens HM, Reed CL, Li N, Burkhart-Kasch S, McKinnon CS, Belknap JK, Gilliam TC, Phillips TJ: Միտերի տարբերությունը տարբերվում է մետամֆետամինի զգայունության համար: Mamm Genome- ը: 2005; 16 (5): 291-305 [PubMed]
29. Բոգուշ Ա, Pedrini S, Pelta-Heller J, Chan T, Yang Q, Mao Z, Sluzas E, Gieringer T, Ehrlich ME. AKT- ը եւ CDK5 / p35- ը միջնորդում են ուղեղի վրա հիմնված նյարոտրոֆիկ գործոնի DARPP-32- ի ներարկումը միջին խոշոր ցողունային նեյրոնների մեջ: Ջ Բիոլ Քեմ: 2007; 282 (10): 7352-9 [PubMed]
30. Benavides DR, Bibb JA: Cdk5- ի դերը թմրամոլության եւ պլաստիկության մեջ: Անն Նյու Յորքի Ակադ Սի. 2004; 1025: 335-44. [PubMed]
31. Bibb JA, Chen J, Taylor JR, Svenningsson P, Nishi A, Snyder GL, Yan Z, Sagawa ZK, Ouimet CC, Nairn AC, Nestler EJ, Greengard P. Կոքինի խրոնիկ ազդեցության հետեւանքները կարգավորվում են Cdk5 նեյրոնային սպիտակուցներով: Բնություն: 2001; 410 (6826): 376-80 [PubMed]
32. Blochl A, Sirrenberg C. Neurotrophins խթանում դոպամինի ազատ արձակել rare mesencephalic neurons միջոցով Trk եւ p75Lntr ընկալիչների. Ջ Բիոլ Քեմ: 1996; 271 (35): 21100-7 [PubMed]
33. Բերտոն O, McClung CA, Dileone RJ, Krishnan V, Renthal W, Russo SJ, Graham D, Tsankova NM, Bolanos CA, Rios M, Monteggia LM, Self DW, Nestler EJ: BDNF- ի հիմնական դերը մզոլիմիկ դոպամինային ճանապարհում `սոցիալական պարտության սթրեսի ժամանակ: Գիտություն. 2006; 311 (5762): 864-8 [PubMed]
34. Nestler EJ, Carlezon WA, Jr The mesolimbic dopamine պարգեւատրման միացում դեպրեսիայի. Բիոլոգիական հոգեբուժություն: 2006; 59 (12): 1151-9 [PubMed]
35. Ford CP, Beckstead MJ, Williams JT: Kappa opioid- ի սոմատոդենդրիտային դոպամինի արգելակման կանխարգելիչ ստորին հոսանքները: J Neurophysiol- ը: 2007; 97 (1): 883-91 [PubMed]
36. Nylander I, Vlaskovska M, Terenius L. Brain dynorphin եւ encephalin համակարգերը Ֆիշերի եւ Լյուիսի առնետներում. Մորֆինի հանդուրժողականության եւ հեռացման հետեւանքները: Brain Res. 1995; 683 (1): 25-35 [PubMed]
37. Nylander I, Hyytia P, Forsander O, Terenius L. Prodynorphin- ի եւ proenkephalin համակարգերի ալկոհոլային նախասիրությունների (AA) եւ ալկոհոլի խուսափման (ANA) առնետների տարբերությունները: Ալկոհոլի հիվանդանոց 1994; 18 (5): 1272-9 [PubMed]
38. Kreek MJ- ն: Կոկաին, դոպամին եւ ներածական օփիոիդային համակարգը: Ջ. 1996; 15 (4): 73-96 [PubMed]
39. Carlezon WA, Jr, Duman RS, Nestler EJ: CREB- ի բազմաթիվ դեմքեր: Թրենդներ Նյարդաբան: 2005; 28 (8): 436-45 [PubMed]
40. Dudman JT, Eaton ME, Rajadhyaksha A, Macias W, Taher M, Barczak A, Kameyama K, Huganir R, Konradi C. Dopamine D1 ընդունիչները միջնորդում են CREB ֆոսֆորալացմանը NMDA- ի ընկալիչի ֆոսֆորալացման միջոցով Ser897-NR1- ում: J Neurochem. 2003; 87 (4): 922-34 [PubMed]
41. Self DW- ն: Մեզոլիմիկ դոպամին համակարգում նեյրոհարդապետությունների կողմից թմրամիջոցների վերացման եւ դիտարկման վարքագծի կարգավորումը: Neuropharmacology. 2004; 47 1- ը, 242-55- ը: [PubMed]
42. Beitner-Johnson D, Nestler EJ- ն: Մորֆին եւ կոկաինը ընդհանուր քրոնիկական գործողություններ են վարում դոպինամիկ ուղեղի վարձատրության շրջաններում թիրոսինային հիդրոխիլազի վրա: J Neurochem. 1991; 57 (1): 344-7 [PubMed]
43. Lu L, Grimm JW, Շահամ Յ, Հույս BT: Քաղցկեղի նեյրոհարդապետությունների մեջ մեղվաբույծների եւ վրացական քաղցկեղային տարածքի մեջ, առաջին իսկ 90 օրվա ընթացքում, առնետների կոկաինի ինքնակազմակերպման պարտադիր ձգձգումը: J Neurochem. 2003; 85 (6): 1604-13 [PubMed]
44. Շեպարդ Ջ.Դ., Չուանգ Դ.Տ., Շահամ Յ., Մորալես Մ.- Մաթհամպեթամինի ինֆեկցիաների ազդեցությունը թիրոսինային հիդրոքսիլազի եւ դոպամինի փոխադրման մակարդակների վրա `առնետի մեսոլիմիկ եւ նիգրոստրիալալ դոպամինային ուղիների վրա: Psychopharmacology (Berl) 2006; 185 (4): 505-13 [PubMed]
45. Fulton S, Pissios P, Manchon RP, Stiles L, Ֆրանկ Լ., Pothos EN, Maratos-Flier E, Flier JS: Մետոակցեմենսների դոպամինային ուղիների լեպտինի կարգավորումը: Նյարդոն: 2006; 51 (6): 811-22 [PubMed]
46. Narita M, Nagumo Y, Miyatake M, Ikegami D, Kurahashi K, Suzuki T. Սպիտակուցային kinase C- ի ներարկում, որը բուսական դոպամինի մակարդակի orexin-induced բարձրացման եւ դրա հատուցող ազդեցություն է: Eur J Neurosci: 2007; 25 (5): 1537-45 [PubMed]
47. Նարիտա Մ, Նագումո Յ, Հաշիմոտո Ս, Խաթիբ Ջ, Միտակե Մ, Սաքուրայ Տ, Յանավիսավա Մ, Նակամաչի Տ, Շիոդա Ս, Սյուզուկի Տ. Ուղեղների արգասիքային համակարգերի ուղղակի ներգրավում մեքսոլիմիկ դոպամինային ուղիների ակտիվացումն ու համապատասխան վարքագծերը, որոնք առաջացրել են մորֆին: J Neurosci. 2006; 26 (2): 398-405 [PubMed]
48. Borgland SL- ի, Taha SA- ի, Sarti F- ի, Fields HL- ի, Bonci A- ի Orexin A- ի համար կարեւոր է կոպի համախտանիշի եւ սրտամկանի սինթեզի զգայունության առաջացման համար: Նյարդոն: 2006; 49 (4): 589-601 [PubMed]
49. Պարկ ES, Yi SJ, Kim JS, Lee HS, Lee IS, Seong JK, Jin HK, Yoon YS: Փոփոխություններ orexin-A- ի եւ neuropeptide Y- ի արտահայտությունը արագացված եւ բարձր ճարպային դիետայի սատկած առնետների հիպոթալամուսում: J Vet Sci. 2004; 5 (4): 295-302 [PubMed]
50. Wortley KE, Chang GQ, Davydova Z, Leibowitz SF- ն: Պեպտիդները, որոնք կարգավորում են սննդային ընդունումը. Hypertriglyceridemia- ի օրոք ավելանում է orexin- ի գենի արտահայտությունը: Բնապահպանական ֆիզիոլոգիա: 2003; 284 (6): R1454-65 [PubMed]
51. Չժեն Հ, Կորկեռն Մ, Ստոյովյան I, Պաթթերսոն Լ .Մ., Թիան Ռ, Բերտուդ Հ. Պեպտիդները, որոնք կարգավորում են սննդի ընդունումը. Ախորժակը ներգործող հոդակապի մանիպուլյացիան ակտիվացնում է հիպոթալամիկ orexin նեյրոնները եւ խոչընդոտում է POMC նեյրոնները: Բնապահպանական ֆիզիոլոգիա: 2003; 284 (6): R1436-44 [PubMed]
52. Baldo BA, Gual-Bonilla L, Sijapati K, Դանիել RA, Landry CF, Kelley AE: GECAA- ի ընկալիչների միջուկային միջամտության արգանդի ծակոտկենու արգելակման միջոցով orexin / hypocretin պարունակող հիպոթալամիկական նեյրոնների ենթաբազմության ակտիվացումը, բայց ոչ նոր վտանգավոր միջավայրի ազդեցության տակ: Eur J Neurosci: 2004; 19 (2): 376-86 [PubMed]
53. Harris GC, Wimmer M, Aston-Jones G. Դիրքը պտտման ձգտումներում կռունկ hypothalamic orexin նեյրոնների համար: Բնություն: 2005; 437 (7058): 556-9 [PubMed]