Biol Psychiatry. 2008 Dec 1; 64 (11): 941-50. Epub 2008 шілде 26.
Teegarden SL, Nestler EJ, Bale TL.
қайнар көз
Пенсильвания Университеті, Филадельфия, ХNUMX-19104, АҚШ, жануарлар биологиясы кафедрасы.
дерексіз
ЖАЛПЫ МӘЛІМЕТТЕР:
Сыйақыға сезімталдығы есірткіні теріс пайдалану, сондай-ақ переэстрациялауға байланысты мінез-құлықтың алдын-алу факторы ретінде қарастырылады. Алайда сезімталдыққа жетудің негізгі механизмдері белгісіз. Біз допаминдік сигнализациядағы дисрегуляцияның арқасында жүйені қалыпқа келтіруге ынталандыратын ынталандырулар пайда болуы мүмкін сыйақы сезімталдықтың жоғары деңгейінің себебі болып табылады.
ӘДІСТЕР:
Біз Delta FosB-overexpressing тінтуірдің жоғары сыйымдылығы жоғары майлы диетаға жауап ретінде марапат жолының өзгеруін зерттеу үшін сыйақы сезімталдық деңгейін арттырудың генетикалық үлгісін қолдандық. Осы тышқандардағы маркалы сигналдық белгілер маркерлері негізінен және 6 аптасына дәмді диеталық әсердің әсерінен кейін зерттелді. Тіндердің мінез-құлық тестінде майланған тамақтанудан кейінгі үлгісінің осалдығын бағалау үшін жоғары майлы диетаны шығарғаннан кейін зерттелді.
НӘТИЖЕЛЕР:
Біздің нәтижелеріміз ядро accumbens - гипоталамус-вендральді депрессия аймағының схемасы бойынша Delta FosB ядросының акументіндегі және стрикатуралық аймақтардағы. Фосфорланған циклдік аденозиндік монофосфат (cAMP) жауап элементінің байланысу деңгейлері (pCREB), ми-туындаған нейротрофиялық фактор (BDNF), және Dopamine және dopamine сигналдарын азайтуды білдіретін Delta FosB тышқандарындағы ядро accumbens ішіндегі 32 kDa (DARPP-32) молекулалық массасы бар циклдік аденозин монофосфат реттелетін фосфопротеин төмендеді. Алты апта бойы жоғары майдың диеталық әсерлері осы айырмашылықтарды жақсартты, бұл дәмді диетаның қуатты сыйымдылығын анықтады. Delta FosB тышқандары сондай-ақ, майдың жоғалуынан кейінгі тыныс жолдарының белсенділігі мен алаңдаушылыққа байланысты жауаптардың 24 сағатта айтарлықтай өсуін көрсетті.
ҚОРЫТЫНДЫ:
Бұл нәтижелер Delta FosB және допаминдік сигналдардың дисрегуляциясына байланысты дәмнің өзгеруіне негізделген сезімталдықты анықтайды, олар дәмді диеталармен қалыпқа келтірілуі мүмкін және семірудің кейбір түрлерінде бейімділік фенотипі болуы мүмкін.
кіріспе
АҚШ-та тәбет пен қанықтылықты бақылайтын нейрондық жүйелер туралы білімдерімізге қарамастан, семіздік мөлшерлемесі арта бермек. Қолданыстағы дәрі-дәрмектердің тиімділігі шектеулі және мінез-құлықтың өзгеруі ұзақ уақыт сақталмаған (1). Калориялы тығыз, дәмді тағамдарды тұтыну мидағы стресс пен марапаттау жолдарының өзгеруімен байланысты, бұл тағамдардың пайдалы қасиеттері энергия теңгерімінің сигналдарын (2-4) асыра алады деп болжайды. Майда жоғары тағамдықтар табигый жетістіктер ретінде әрекет етеді, ми сыйақылар орталығын теріс пайдаланылатын есірткіге ұқсас тәсілмен белсендіреді және өзін-өзі басқару парадигмаларында (5-8) қолданылады. Осылайша, асырау мен есірткіге тәуелділікте мінез-құлық және ынталандыру ортақ тетіктерді біріктіріп, екі жағдайда да емдеудің жаңа жолдарын ашуға ықтимал.
Дәмелі тағамдар мен мидағы стрессті реттейтін жолдар арасындағы қарым-қатынасты зерттеу кезінде, біз жоғары деңгейдегі майлы тамақтанудан (HF) кетуден кейін төмендетілген сыйақылардың молекулалық және биохимиялық маркерлерін анықтадық. Зиянды препараттарға ұқсайтын, біздің зерттеулеріміздегі дәмді диетаға ұшыраған ядролық accumbens (NAc) транскрипция коэффициентінің ΔFosB деңгейін, орталық ми сыйымдылық құрылымын (9, 10). ΔFosB индуцируемей асып бара жатқан тышқандар азық-түлік сыйлығына (11) арналған аспаптық жауапты арттыра отырып, оларды сыйақы сезімталдықтың рөлін зерделеу үшін және сыйымдылықты диетаға молекулярлық және биохимиялық жауаптарда марапаттау жүйесінің ұзақ мерзімді дисрегуляциясы.
Зерттеу барысында біз ΔFosB-overexpressing тышқандарында біз нәзік HF диетасына жауап ретінде NAc-гипоталамус-вендральді ойықтыру аймағында (VTA) сыйақы маркерлерінде ұзақ мерзімді өзгерістерді зерттеу үшін қолдандық. Осы марапатқа сезімтал тышқандардағы бұрынғы зерттеулерге негізделген болсақ, біз сыйақы сезімталдығындағы ΔFosB индуцированных өзгерістері NAC кері байланысына негізделген допаминдік сигнализациядағы дисрегуляцияны қамтиды. Бұдан басқа, біз энергетикалық тығыз HF диетасының табиғи сыйлығына әсер етуі осы тышқандардағы допаминергиялық жүйені қалыпқа келтіруі мүмкін деп болжадық, бұл осы HF диетасынан шыққан кернеулерге шамадан тыс жауап берді. Дәмді рационды пайдалы зат ретінде қолданудың бірегей аспектісі бізге гипоталамикалық кірістерді схемаға фенотипте сыйлауға мүмкіндік береді, ол емделуге төзімді семіздікке бейімделген халықты болжау мүмкін. Осы гипотезаны зерттеу үшін HF экспозициясынан кейін NAc және тирозин гидроксилазасы және VTA-дағы допаминді тасымалдаушыда pCREB, BDNF және DARPP-32 қоса алғанда, допаминді нейротрансмейстерді зерттедік. Сондай-ақ, біз допаминнің шығуына әсер ететін, оның ішінде лактин және орексин рецепторлары мен ВТА-да және бүйірлік гипоталамуста орексинді білдіруге әсер ететін нақты баланстық белгілерді қарастырдық.
Материалдар мен әдістер
Жануарлар
NAc және dorsal striatum (Kelz және т.б., 1999) динорфин-позитивті нейрондарында ΔFosB индуцирленген түрде асып түсетін ерлердің битрансгенді тышқандары Техастың оңтүстік батыс медициналық орталығында (ICR: C57Bl6 / SJL) жасалынып, Пенсильвания Университетінде сынақтан өтті. Барлық тышқандар Пенсильвания Университетіне келгенше доксициклинге (ауыз суда 100 мкг / мл) қолдады. Тым көп экспрессияға жол бермеу үшін доксициклин жойылды (n = 23) (12). Бақылау тышқандары (n = 26) препарат алуды жалғастырды. Тізбелер диеталық топтарға доксициллинді жойғаннан кейін сегіз аптадан кейін тағайындалды, онда уақытша көріністің максималды деңгейлерге жетуі көрсетілген (13). Тышқандар 12: 12 ашық-қараңғы циклында (0700-дағы шамдар) азық-түлік пен судың қолжетімділігімен қамтамасыз етілді. Барлық зерттеулер Пенсильвания институты жануарларды қорғау және пайдалану комитетімен бекітілген тәжірибелік хаттамаларға сәйкес жүргізілді және барлық рәсімдер институционалдық нұсқауларға сәйкес жүргізілді.
Диеталық әсер ету
Тышқандар үйдегі шошқада сақталды (n = 16) немесе алты апта ішінде HF (n = 16-17) орналастырылды. Үй чусы (Purina Lab Diet, Сент-Луис, MO) құрамында 4.00% ақуыз, 28% май және 12% көмірсулардан тұратын 60 ккал / г бар. HF диетасы (Research Diets, New Brunswick, NJ) құрамында 4.73% ақуыз, 20% май және 45% көмірсулардан тұратын 35 ккал / г бар.
Биохимия және геннің өрнегі
Тіндердің алты апта диеталық әсерінен кейін талданды. Миы тасбақадан шығарылды немесе мұздаған құрғақ мұзда немесе NAc ажыратылды (шамамен 0.5 - 1.75 мм, бегмадан, 3.5 - 5.5 тереңдігінде) және сұйық азотта мұздатады. Тығыздық-80 ° C температурасында сақталғанша сақталды.
Биохимиялық талдау
Батыс блоктарға арналған әдістеме қосымша материалдарда сипатталған. Қолданылған антиденелер: Cdk5, CREB және BDNF (1: 500, Santa Cruz Biotechnology, Santa Cruz, CA) және фосфор-CREB (pCREB) (Ser 133) (1: 500, Cell Signaling Technology, Danvers, MA).
Рецептордың авторганографиясы
Қосымша материалдарда аутриганографтың толық әдістері сипатталған. Қолданылатын лигандтар 2 nM H3 - SCH 23390 және 5 nM H3 - спиперон болды (PerkinElmer, Бостон, MA).
In situ будандастыру
Тіндердің өңдеуі және будандастыру бұрын сипатталғандай орындалды (14). DARPP-32 зондты P. Greengard (Рокфеллер университеті) және J. Elmquist (Техастың оңтүстік батыс медицина орталығының университеті) орексиндік зондты ұсынды. 32 күн бойы DARPP-3 үшін талданатын слайдтар пленкаға түсірілді және 4 күн үшін пленкада орексинге арналған слайдтар қолданылды. Фильмнің суреттерін сандық бағалау бұрын сипатталғандай орындалды (10).
QRT-PCR
РНК VTA-дан оқшауланған және TaqMan ген экспрессиялық талдауымен (Applied Biosystems, Foster City, CA) пайдаланылған жеке гендердің экспрессиясы болды. Толық әдістемелер мен статистикалық талдауды қосымша материалдардан табуға болады.
Жүріс-тұрыс талдаулары
Диета-индуцирленген мінез-құлық өзгерістеріне сыйақы сезімталдықтың әсерін зерттеу үшін төрт аптадан кейін HF-ден тышқандар жиынтығы HF-ден алынады және үй чуына (n = 9 бақылау, n = 8 ΔFosB) қайтарылды. Шығарғаннан кейін жиырма төрт сағат бұрын, тышқандар, бұрын жарияланған диеталық алу парадигмасына (10) сәйкес ашық далалық сынауға ұшырады. Қысқаша айтқанда, тышқан ашық далалық аппараттың ортасына орналастырылып, бес минут бойы бақыланды. Барлығы сызықты крест, фекальды лак, орталықта уақыт және орталыққа өтіп өлшенеді.
статистика
Батыс блоттарынан басқа барлық деректер екі жақты ANOVA көмегімен, содан кейін Фишердің PLSD тестімен доксициклинмен емдеу (ΔFosB өрнегі) және диета күйі тәуелсіз айнымалылар ретінде талданды. RT-PCR анализі үшін байланысты гендердің топтары бойынша бірнеше салыстыруды түзету үшін P мәні төмендетілді (қосымша материалдарды қараңыз). Батыс дақтарға тәуелсіз айнымалы ретінде доксициклинді емдеуге арналған студенттің t-тесті қолданылып, сол блоттағы оптикалық тығыздықты салыстыру арқылы талдау жасалды. Барлық деректер орташа ± SEM ретінде ұсынылған.
нәтижелері
Негізгі биохимиялық айырмашылықтар
ΔFosB-шамадан тыс экспрессия жасайтын тышқандардағы сыйақының жоғарылатылған сезімталдығының негізінде жатқан молекулалық жолдарды анықтау үшін NAc-де бірнеше негізгі сигналдық молекулалардың деңгейлері зерттелді. XFosB тышқандарының NAc-да Cdk5 деңгейлерінің доксициклинде ұсталатын литратматты бақылау жануарларымен салыстырғанда жоғарылау тенденциясы болды (F = 5.1, P = 0.08; Cурет 1А). ΔFosB тышқандары pCREB деңгейлерін едәуір төмендеткен (F = 7.4, P <0.05; Cурет 1B), сонымен қатар CREB жалпы деңгейлері (F = 5.4, P = 0.05; Cурет 1C). BDNF-тің айтарлықтай төмендеуі inFosB тышқандарының NAc-де байқалды (F = 10.6, P <0.05; Cурет 1D).
Сурет 1
ΔFosB-нен асып түсетін тышқандар NCc-дағы допаминді төмендетілген сигналдардың биохимиялық маркерлерін көрсетті
Тамақ қабылдау және дене салмағының жоғарылауы
Біз бұдан әрі rewardFosB-шамадан тыс әсер ететін тышқандардағы сигнал беретін молекулалардың өзгеруіне табиғи пайдалы HF диетасының әсерін қарастырдық. HouseFosB тышқандары мен үйде де, HF-де де тамақ қабылдау кезіндегі айырмашылықтар болған жоқ. Алайда FFosB тышқандарына тән HF әсер еткенде дене салмағына дейін қалыпқа келтірілген калориялы қабылдаудың жалпы төмендеуі байқалды (F = 11.2, P <0.01; Cурет 2A). Алты аптаның диеталық экспозициясы аяқталғаннан кейін HF алған тышқандардың салмағы chow диетасындағыдан едәуір көп болды (F = 17.2, P <0.001), ал ΔFosB тышқандары жалпы салмақтан аз болды (F = 5.6, P <0.05; Сур. 2B). Бұл әсер chow диетасындағы топтар арасындағы айырмашылықтарға тән болды (P <0.05).
Сурет 2
ΔFosB артық тышқандарды тышқандар тамақтану кезінде ешқандай майдың немесе жоғары майдың (HF) диетасындағы айырмашылықты көрсетті
Жоғары май диетасындағы биохимиялық айырмашылықтар
NAF сигнализациясының базалық айырмашылықтары HF диетасы арқылы өзгеруі мүмкін екендігін анықтау үшін, бастапқы деңгейде зерттелген сигнал беретін ақуыздар алты аптадан кейін HF алған жануарларда зерттелген. Cdk5 деңгейлерінде елеулі айырмашылықтар болмады (сурет 3A). PCREB және total CREB деңгейлері алты аптадан кейін HF (Figure 3B, C) кейін әр түрлі болды. BFNF деңгейлері ΔFosB тышқандарында HF экспозициясының алты аптадан кейін (F = 6.5, P = 0.05, 3D) кейін айтарлықтай көтерілді.
Сурет 3
Жоғары май (HF) диета жақсартылған сигналдық айырмашылықтар, ΔFosB-нен тышқандарды асфальттаушы тышқандар NAc
Допаминді рецепторлардың авторганографиясы
Біз NAc ішіндегі допаминдік сигнализациядағы ΔFosB индукцияланған өзгерістердің допаминдік рецепторлар экспрессиясының өзгеруіне байланысты екендігін бағалау үшін рецепторлардың аутодиорографиясын қолдандық (Cурет 4A). Жоғары майлы диета D1 допаминді рецепторлардың байланысуының тығыздығын аздап жоғарылатты (P = 0.14), және бұл айырмашылық ΔFosB тышқандарында көп болды (Cурет 4B). Сондай-ақ, HF (P = 1) -дан кейін D0.06 байланыстыру аймағының ұлғаю тенденциясы байқалды және пост-тестілеу бұл ΔFosB тышқандарында маңызды екенін көрсетті (P <0.05; Cурет 4C). D1 рецепторларынан айырмашылығы, D2 рецепторларының байланысу тығыздығында ешқандай өзгеріс болмайды (бақылау chow = 97.6 ± 6.9, HF = 101.1 ± 8.2, ΔFosB chow = 91.6 ± 1.0, ΔFosB HF = 94.8 ± 9.5) немесе байланыс аймағында (бақылау chow = 47.3 ± 3.4, бақылау HF = 53.8 ± 6.0, ΔFosB chow = 51.9 ± 3.7, ΔFosB HF = 49.0 ± 3.3) NAc-да байқалды.
Сурет 4
Жоғары майлы диета (HF) D1 допаминді рецепторды байланыстырудағы өзгерістерге және ΔFosB тышқандарды экспрессиялау тышқандарындағы ядро accumbens (NAc) DARPP-32 өрнегіндегі өзгерістерге әкелді
NAc ішіндегі DARPP-32 өрнегі
Орнында будандастыру NAc ішіндегі DARPP-32 экспрессия деңгейлерін анықтау үшін қолданылды (Cурет 4D). Жоғары майлы диета осы ми аймағында DARPP-32 экспрессиясын айтарлықтай арттырды (F = 5.1, P <0.05) және диета мен ΔFosB экспрессиясы (F = 8.9, P <0.05) арасында айтарлықтай өзара әрекеттесу болды, ал ΔFosB тышқандары үлкенірек болды диетаның әсерінен болатын өзгеріс (Cурет 4E). Бақылау мен ΔFosB тышқандары арасындағы DARPP-32 өрнегіндегі базальды айырмашылық пост-тестілеу арқылы анықталды (P <0.01), сондай-ақ HF (FFosB) тышқандарындағы DARPP-32 өрнегінің айтарлықтай жоғарылауы (P <0.01).
VTA-дегі геннің өрнегі
QRT-PCR бұрын сыйақыны реттеуге қатысқан бірнеше негізгі гендерге бағытталған VTA-дағы гендік экспрессияның өзгеруін бағалау үшін қолданылды. Барлық үлгілер β-актинге дейін қалыпқа келтірілді. Β-актиннің экспрессиясын емдеу әсерінен өзгертпеу үшін β-актинді екінші ішкі бақылау GAPDH-мен салыстыру үшін бөлек талдау жасалды. Β-актиндік өрнекте айтарлықтай айырмашылықтар болған жоқ (ΔCT мәндері, β-актин - GAPDH: бақылау чау = 2.29 ± 0.21, бақылау HF = 2.01 ± 0.04, ΔFosB чау = 2.32 ± 0.49, ΔFosB HF = 2.37 ± 0.10).
Тирозин гидроксилазасын экспрессиялау үшін ΔFosB экспрессиясы мен диеталық емдеудің өзара әрекеттесу тенденциясы байқалды (F = 3.6, P <0.06; Cурет 5A). Алты апта HF әсерінен бақылау тышқандарындағы тирозин гидроксилазасының экспрессиясы төмендеп, ΔFosB тышқандарындағы экспрессия жоғарылағандай болды. Допамин тасымалдағышын экспрессиялау үшін ΔFosB экспрессиясы мен диета экспозициясы арасындағы айтарлықтай өзара әрекеттестік байқалды (F = 6.7, P <0.03; Cурет 5B). Тирозин гидроксилазасына ұқсас, HF әсерінен бақылау тышқандарындағы допамин тасымалдағыштың экспрессиясы төмендеп, ΔFosB тышқандарындағы экспрессия айтарлықтай өсті (P <0.05). Бақылау мен expressionFosB тышқандары арасындағы допаминді тасымалдаушының экспрессиясындағы базальды айырмашылық маңыздылыққа жете алмады (P = 0.16), бірақ 6 WK HF кейін, ΔFosB тышқандары допамин тасымалдағышының бақылау деңгейлерімен салыстырғанда едәуір жоғарылағанын көрсетті (P <0.05).
Сурет 5
Жоғары майлы диета (HF) экспозициясы және ΔFosB өрнегі VTA ішіндегі бірқатар негізгі молекулалардың экспрессиясының өзгеруіне әкелді
VTA-да TrkB деңгейін төмендету үшін ΔFosB экспрессиясының жоғарылауының әсерін көрсететін тенденция болды (F = 5.7, P <0.04; Cурет 5C). Κ-опиоидты рецепторлардың экспрессиясына негізгі әсер етпесе де, ΔFosB тышқандарында экспрессияның төмендеу үрдісі байқалды (P = 0.08; Cурет 5D). Лептинді рецептордың экспрессиясы VTA-да анықталды. Диета экспозициясының айтарлықтай әсері анықталды (F = 6.1, P <0.03), HF TAFosB және бақылау тышқандарында VTA-да лептин рецепторының деңгейін айтарлықтай төмендетеді (5E-сурет). VTA-да орексин рецепторы 1 экспрессиясы зерттелді. Орексин рецепторының экспрессиясына диетаның айтарлықтай әсері болды (F = 9.0, P <0.02), HF әсеріне ұшыраған тышқандар VTA-да жоғары деңгейлер көрсетті (5F-сурет). Сондай-ақ, ΔFosB тышқандары осы ми аймағында орексин рецепторы 1-нің жалпы деңгейін жоғарылату тенденциясы болды (P <0.05).
Бүйректегі гипоталамуста орехинді білдіру
Біз бүйірлік гипоталамустағы орексин деңгейін, VTA-нің орексинергиялық иннервациясы шыққан жердегі орнында будандастыру арқылы өлшедік (Cурет 6А). FFosB экспрессиясы мен орексин экспрессиясына диеталық экспозиция (F = 9.1, P <0.01) арасында айтарлықтай өзара әрекеттесу болды, HF бақылау тышқандарындағы орексин деңгейін едәуір арттырды (P <0.05) және ΔFosB тышқандарындағы экспрессия төмендеді (Cурет 6B). Базальды күйде орексиннің экспрессиясында айтарлықтай айырмашылықтар болмаса да, 6 вк HF-тен кейін, ΔFosB тышқандары бақылаулармен салыстырғанда орексин деңгейінің айтарлықтай төмендегенін көрсетті (P <0.05).
Сурет 6
Жоғары май (HF) рационында басқарудағы (Ctrl) және ΔFosB тышқандарды экспрессирлеу тышқандарында орексинді экспрессиялауға дифференциалдық әсері болды
Beауамен талдау
Диетаның өзгеруіне байланысты қозу мен эмоционалдылықтың өзгеруін бағалау үшін тышқандар HF диетасын алып тастағаннан кейін 24 сағаттан кейін ашық далалық сынаққа ұшырады (10). OusFosB экспрессиясы (F = 6.6, P <0.05) және диета (F = 4.6, P <0.05; Cурет 7A) айтарлықтай әсер етті. ΔFosB тышқандары жаңа ортада қарама-қайшылыққа қарағанда белсенді болды және пост-тестілеу көрсеткендей, HF-ден алынған тышқандар чау әсер еткендерге қарағанда едәуір белсенді болды (P <0.05). Фекальды боли мазасыздық тәрізді мінез-құлықтың өлшемі ретінде саналды (10). ΔFosB өрнегінің негізгі әсері болды (F = 10.2, P <0.01), ΔFosB-шамадан тыс экспрессия жасайтын тышқандар роман ортасында, әсіресе үй чауында және HF шығару топтарында фекальды болиді көп шығарды (сурет 7B). Δ HF диетасында ұсталатын FosB тышқандары фитальды болиді чауда ұсталғандарға қарағанда және тестілеуден 24 сағат бұрын шығарып алғандарға қарағанда аз шығарды. Бақылау тышқандарына диета әсер етпеген сияқты. FieldFosB экспрессиясының немесе диетаның ашық алаңның орталығында өткізген уақытына айтарлықтай әсері болған жоқ (бақылау chow = 14.5 ± 3.1 сек, HF = 18.0 ± 3.2 sec, бақылау W / D = 15.4 ± 1.9 sec, ΔFosB chow = 16.9 ± 2.4 сек, ΔFosB HF = 13.1 ± 3.9 сек, ΔFosB W / D = 19.8 ± 2.6 сек).
Сурет 7
ΔFosB-ны асқындырған тышқандар жоғары май диетасын (HF) алып тастаудың әсеріне аса сезімтал болды
талқылау
Семіздік емдеу кезінде асқазан мен салмақтың өсуіне сезімталдыққа әсер ететін факторларды анықтауға аса қажет. Міндетті марапат жолдары дәмді тағамдардың және диеталық өзгерістердің (6, 10, 15, 16) мотивациясы мен оларға жауап беруде маңызды рөл атқарады. Орексигендік және анорексигенді сигналдар гипоталамус-VTA-NAc тізбегі арқылы марапаттау сигналдарына тікелей әсер ете алатындықтан, марапат орталықтарында қуатқа бай дәмді диеталарға жауап беретін гендерді анықтау семіздік емдеуде (17, 18) жаңа терапевтік мақсаттарды қамтамасыз етуі мүмкін. Осылайша, сыйақының өзгеруіне (13, 19, 20) жетілдірілген сезімталдық үлгісі ретінде ΔFosB-overexpressing тышқандарындағы HF диетасына жауап ретінде гипоталамус-VTA-NAc тізбегі бойынша сыйақы және энергия теңгерімінің сигнализациясының биохимиялық және молекулалық маркерлерін қарастырдық. , диетадан кейінгі мінез-құлық сезімталдығы. Біз ΔFosB тышқандарындағы допаминдік сигналдарды базальды дисрегуляциялау энергия теңгерімінің сигналдарының және допаминдік жүйенің қиылысуын қамтитын HF диетасының тиімді әсерлері арқылы қалыпқа келтірілетінін болжадық.
NAc-дағы допаминдік сигнализациядағы дисрегуляцияны көрсететін маркерлерді зерттеу үшін біз D1 рецепторлардың деңгейлерін және төменгі эффекторларды зерттедік. D1 рецепторларының байланысында айтарлықтай айырмашылық болмағанымен, ΔFosB тышқандарындағы байланыстыру аймағын ұлғайтуға арналған HF әсерінің үрдісі байқалды. Бұл ΔFosB инъекциясының дәрілік препараттармен индукциясы және табиғи превосходтар, ең алдымен, D1 рецепторларын білдіретін ортаңғы нейрондардың динорфин-оң субтитрінде басым болып көрінеді (9, 21). Төменгі ағымды допаминнің pCREB сигнализациясының деңгейлері ΔFosB тышқандарында айтарлықтай азайған, бұл ми аймағында (1, 22) төмендетілген D23 рецепторын белсендіруді қолдайды. Бір қызығы, біз ΔFosB тышқандарындағы CREB деңгейінің жалпы төмендеуін байқадық, бұл диспроминдік сигналдың трансляциясын одан әрі төмендету мүмкіндігін болжайды, бұл pCREB (24) ұзақ уақытқа төмендеуі нәтижесінде кері байланысқа кері әсер етуі мүмкін. BDNF өрнегі pCREB арқылы реттеледі, D1 активациясымен көтеріледі және NAc-дағы (25, 26) сыйақымен байланысты нейропластиктің маңызды делдалы болып табылады. Тиісінше, біз ΔFosB тышқандарындағы Nc кезінде BDNF протеинінің айтарлықтай төмендеуін анықтадық.
NAc-дағы барлық орташа ақбөкен нейроны DARPP-32 (27) экспресс. Көптеген төменгі аффективтер оны марапаттау жолдарында маңызды рөл атқарады (28) және ол нашақорлықта және дофаминдік жүйені қамтитын басқа да бұзылуларға, соның ішінде аффективті бұзылулар мен шизофренияға (27, 29). DARPP-32 өрнегіндегі ΔFosB тышқандарының NAc-інде терең базальды төмендету анықталды. DARPP-32 өрнегі BDNF арқылы реттеледі, сондықтан қысқартылған өрнек ΔFosB тышқандарында (27, 29, 30) анықталған BDNF деңгейлерінің төмендеуімен тікелей байланысты болуы мүмкін. DARPP-32 фосфорлану күйінде тіпті қалыпты өзгерістер NAc (27) ішіндегі ішіндегі сигнализацияға елеулі өзгерістер әкелуі мүмкін. Алдыңғы зерттеулер DARPP-32 протеинінің ΔFosB тышқандарында 12-wk доксициклинді жоюдан кейін кеңірек стриатальды бағалау жүргізілгенде (31) өзгерген жоқ, DARPP-32 арналған ΔFosB әсерлері уақыт пен аймаққа тән болуы мүмкін деп болжайды.
Біз ΔFosB тышқандарының NAc-дағы допаминдік сигналдар көрсеткіштерінің күрт төмендеуі, мүмкін, ΔFosB осы нейрондық шектерде асып көрсетілмеген болса да, VTA допаминдік проекция нейрондарындағы өзгерістерге ұшырады деп болжадық. Осылайша, біз вирустық дозада генетикалық дозиметрлерді, соның ішінде тирозин гидроксилазасын және допаминді тасымалдағышты зерттедік. Тироинді гидроксилазаның және допаминді тасымалдағыштың деңгейлері допаминнің шығуымен оң коррелирленген. ΔFosB тышқандарында триосин гидроксилазын төмендетуге және допаминді тасымалдағышта айтарлықтай азайтуға НКА-да допаминдік сигнализацияның бұзылуына байланысты үрдісі байқалды. ΔFosB тышқандарының VTA-індегі допаминмен байланысты гендердегі бұл базалық төмендету ұзақ мерзімді ΔFosB асып кеткен кезде NAc-дан өзгерген кері байланысты көрсетеді, біз BDNF рецепторлардың, TrkB-нің VTA (32) -ке кері байланысының мүмкін механизмі ретінде қарастырдық. Тирозин гидроксилазасы мен допаминді тасымалдағышқа ұқсас, TrkB экспрессиясы, көптеген салыстыру кезінде түзетілген кезде маңызды болмаған ΔFosB тышқандарында төмендеу үрдісін көрсетті. BDNF-TrkB кешені ретрогенді түрде тасымалдана алады және жергілікті геннің өрнегіне әсер етеді және жасуша өсуін және қызмет көрсетуін қолдау үшін (33) VTA ішінде әрекет етеді. Сонымен қатар, NAc ішінде прессинаптическом TrkB белсендіру BDNF допаминдік нейротрансляцияны (32) тікелей ынталандырады, бұл тышқандардағы допаминдік сигналдың төмендеуін қолдайды.
Κ-opioid рецепторларының динорфинді белсендіруі допаминдік сигналды реттейді және NAc VTA-ке кері байланысты қамтамасыз ететін тағы бір механизм (34). Біз VTA-дағы κ-opioid рецепторларының өрнектері ΔFosB тышқандарында төмендеу үрдісін көрсетті. ΔFosB артық нәтиже көрсеткендей, NAc (20) динорфиндік өрнегін азайту үшін ΔFosB тышқандары таза VTA κ-опиоидты белсендіру. Динорфиндік сигнал беру әдетте допаминдік нейрондарға тежегіш әсер етеді [35], теріс пайдаланылатын дәрілерді өзін-өзі басқаруды көрсететін егеуқұйрықтар NAc-те динорфиннің төмендеген деңгейлерін көрсетеді, бұл сыйақы сезімталдығын арттыруда динорфиндік сигнализацияның негізінен төмендеуіне ықпал етеді [36]. , 37). Динорфин - op-опиоидтық жүйенің реттелмеуі допаминдік жолдарды қалыпқа келтіру кезінде опиоидтық сигнализацияның критикалық тепе-теңдігін қолдай отырып, есірткіні теріс пайдаланумен және сақталумен байланысты болды. (38).
Энергетикалық тығыздығы жоғары HF диетасының сыйымдылығына негізделген, біз ΔFosB тышқандарындағы допамин және опиоидтық маркалардағы дисрегацияның осы тышқандарға осындай диетаға жақсартылған сыйақы реакцияларына бейімделуін болжайды, осылайша гипоталамустың белсендірілуі арқылы сыйақы жүйесін қалыпқа келтіреді -VTA-NAc тізбегі. Алты аптаға диеталық әсер ету кезінде ΔFosB мен бақылау тышқандар арасында азық-түлік тұтынудың айырмашылығы байқалды, бұл ΔFosB тышқандарындағы биохимиялық және молекулалық маркерлік белгілердегі өзгерістердің тұтынылған калориялардың айырмашылығына байланысты емес екенін көрсетті. Күтілгендей, ΔFosB мен басқарушы тышқандар арасындағы pCREB, CREB, BDNF, DARPP-32 және κ-opioid рецепторлардың деңгейлері анықталды, бұл HF (29, 39-41) бойынша ΔFosB тышқандарындағы допаминнің шығуын жоғарылатуына байланысты, .
Тироцинді гидроксилазаны және ВТА-дағы допаминді тасымалдаушыны зерттеу HF-ден кейінгі ΔFosB және басқарушы тышқандардың таңғажайып қарсы жауаптарын көрсетті. Басқару тышқандарында тирозин гидроксилазасы және допаминді тасымалдаушы экспрессиясының төмендеуі байқалды, ал ΔFosB тышқандары допаминмен байланысты гендердің екеуінің де өрнектелгенін көрсетті. Бір қызығы, тиросин гидроксилазы экспрессиясы VTA-да созылмалы кокаин немесе метамфетаминді қолдану арқылы өзгереді (42-44), бұл ΔFosB тышқандарында HF-нің табиғи сыйақысын басқару тышқандарынан гөрі анық болады.
VTA-ге әлеуетті гипоталамус енгізу энергия теңгерімін көрсететін сигналдарды беруді, лептин рецепторының және орексин рецепторы-1-ның экспрессиясын да зерттеді. Лептиннің циркуляциялық деңгейі HF-мен көбейтіледі, ал лептин өз кезегінде ДПАМ-да дабаминді сигналдарды өзгерту үшін қолданылады (18, 45). ВТС-ның лептинді рецепторларының экспрессиясы HF-де, сонымен бірге, HF-де болғанда, ұқсас салмақ пен диетаны қабылдау кезінде, ΔFosB және басқару тышқандарында да HF-мен азайды. Жоғары май ΔFosB және басқару тышқандарының VTA-дағы орексин рецепторлары-1-ның экспрессиясын жоғарылатты. Орексин VTA-дағы допаминдік нейронды белсендіреді, VTA пластикасын қолдайды және NAc (46-48) допамин деңгейлерін жоғарылатады. Жоғары майлы диета біздің бақылауларымызға сәйкес (49, 50) сәйкес тышқандардағы орексинді арттыруды көрсетті. Осылайша, орексин рецепторларының экспрессиясы, сондай-ақ ВТА-дегі лептиннің сигнализациясының өзгеруі ΔFosB және басқару тышқандарындағы диеталық сыйақыларды ынталандырады, бұл энергия теңгерімінің сигналдарын тарататын жолдар арасындағы диссоциацияларды қолдайды және тікелей марапаттауға байланысты.
Сыйлықты алудың стресстік әсерлерін тексеру үшін тышқандар HF-ны алып тастағаннан кейін 24-та ашық далалық сынақта қаралды. ΔFosB тышқандары барлық басқа бақылау және диеталық топтармен салыстыра отырып, романның ашық алаңында жоғары белсенділік пен фекальды лактивті өнімділікті көрсететін қолайлы диетаны алудың өткір әсеріне аса сезімтал болды. ΔFosB тышқандары сонымен бірге сыйақыны және стресстік сезімталдықты білдіретін осы сынақта қызықты мінез-құлықтың үлгісін көрсетті, ал HF диетасы бастапқыда фекальды шоколадқа қатысты өнімді азайтады, ал бұл шығын қайтадан осы алаңдаушылықты арттырады. Ашық далалық белсенділіктің жоғарылауы орексиннің өрнектелуінің өзгеруімен байланысты емес, бұл стресс туындаған үрдістің өзара байланысын білдіреді, бұл жай ғана орексенді сигнализациядағы өзгерістерге ғана әсер етпейді. Тұтастай алғанда, бұл деректер біздің гипотезаны қолдайды, бұл ΔFosB тышқандары жоғары деңгейдегі сезімталдықты жоғарылатуына байланысты қолайлы диета алудың өткір әсеріне аса сезімтал болады.
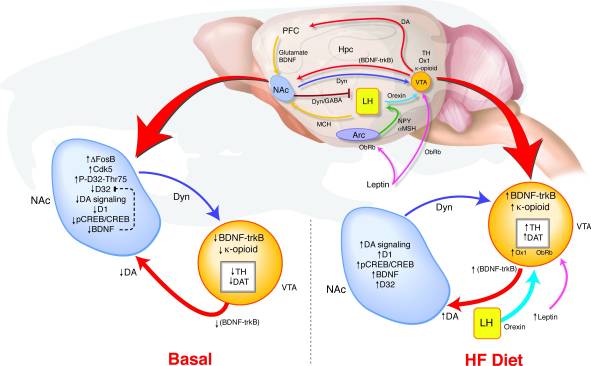
NAc-дағы ΔFosB-тің ұзақ мерзімді артық көрінісі қалай мінез-құлқының және марапаттау сигналының өзгеруіне әкеледі?? Біз VCA-ның сәйкестігін анықтаудың үлгісін ұсындық, онда NAc және гипоталамустың релелік сигналдары марапат жолының дислогенизациясы мен семіруге бейімділік арасындағы байланыстарды қолдайтын допаминдік жүйені анықтау үшін марапаттау жағдайына қатысты сигналдарға байланысты болады (сурет 8). HF экспозициясында, энергия теңгерімі мен сыйақы мөлшерін көрсететін көптеген кірулер VTA-ға жақындайды. Лептиннің және орексиннің сигнализациясының жоғарылауы, сондай-ақ НАЦ-дан бүйірлік гипоталамусқа кері байланысқа кері әсер ету ΔFosB тышқандарында (17, 18, 45, 47, 51-53) осы нейсигенді сигналдарға қалай әсер етуі мүмкін. BDNF-дегі жоғары майлы диета-индуцирленген биіктіктер ВТА-ға сыйақы беру мүмкіндігін береді, әрі допаминмен байланысты гендік өрнектің өзгеруіне ықпал етеді.
Сурет 8
Жоғары май (HF) диета ΔFosB тышқандарында реттелетін сыйақы сигналын қалыпқа келтіреді
Бұл нәтижелер сыйақы сезімталдығының молекулярлық маркерлерін анықтайды және допаминдік жүйенің ұзақ мерзімді дисрегуляциясы жеке адамға тәуелділікке және семіздікке бейімделуі мүмкін екендігін көрсетеді. Бұдан басқа, бұл деректер семіздік пен басқа да бұзушылықтарды емдеуде және алдын-алуда потенциалды жаңа терапевтік мақсаттарды анықтайтын маңызды қадам болып табылады. Болашақта бұл жүйенің HF диетасын жоюға қалай жауап беретінін зерттеу, сондай-ақ марапаттауға және жоғары майлы диеталарға әсер ету кезіндегі жыныстық айырмашылықтарды зерттеуге болады.
Қосымша материалдар
Supp. Әдістері
Көру үшін мына жерді басыңыз (61K, doc)
Алғыс
Авторлар Кэти Шеффенге жануарларды өсірумен және тасымалдаумен көмектескені үшін алғысын білдіреді. Бұл жұмыс Пенсильвания Диабет Орталығы (DK019525) Университетінің гранты және Ұлттық психикалық денсаулық институты (R01 MH51399 және P50 MH66172) және Нашақорлықтың ұлттық институты (R01 DA07359) грантымен қаржыландырылды.
Сілтемелер
Қаржылық ақпаратты ашу: Барлық авторлар олардың биомедициналық қаржылық мүдделері немесе ықтимал мүдделер қақтығысы жоқ екенін мәлімдейді.
Әдебиеттер тізімі
1. Wadden TA, Berkowitz RI, Womble LG, Sarwer DB, Phelan S, Cato RK, Hesson LA, Osei SY, Каплан Р, Stunkard AJ. Семіздікке арналған фармакотерапия мен өмір сүрудің өзгеруін рандомизацияланған сынау. N Engl J Med. 2005; 353 (20): 2111-20. [PubMed]
2. Blendy JA, Strasser A, Walters CL, Perkins KA, Patterson F, Berkowitz R, Lerman C. Семіздікке арналған никотин сыйақысын азайту: адам мен тінтуірдің өзара салыстыру. Психофармакология (Берл) 2005
3. Франкен И.Х., Мурис П. Сыйақыға сезімталдықтағы жеке айырмашылықтар салауатты әйелдердің тамақтануына және салыстырмалы дене салмағына байланысты. Аппетит. 2005; 45 (2): 198-201. [PubMed]
4. Келли А, Berridge KC. Табиғи жетістіктердің неврологиясы: есірткіге тәуелділікке қатысты. J Neurosci. 2002; 22 (9): 3306-11. [PubMed]
5. Cagniard B, Balsam PD, Brunner D, Zhuang X. Хроникалық жоғары допамині бар тышқандар азық-түлік наградасы үшін оқуды емес, мотивацияны күшейтеді. Нейропсихофармакология. 2006; 31 (7): 1362-70. [PubMed]
6. Liang NC, Hajnal A, Norgren R. Sham Жүгері майын азықтандыру егеуқұйрықтағы акументальды допаминді арттырады. Am J Physiol Regul интегрі Comp Physiol. 2006; 291 (5): R1236-9. [PubMed]
7. Мендоза Дж, Анджелес-Кастелланос М, Эскобар С. Таңертеңгі тамақтануға талпыну тамақтың алдын-ала белсенділігі мен мидың марапатталған аудандарында c-Fos өрнегін тудырады. Неврология. 2005; 133 (1): 293-303. [PubMed]
8. Шредер Б.Е., Бинзак Дж.М., Келли А.Е. Никотин немесе шоколадпен байланысты контекстік белгілердің әсерінен кейінгі префронтальді кортикальды белсендірудің жалпы профилі. Неврология. 2001; 105 (3): 535-45. [PubMed]
9. Nestler EJ, Barrot M, Self DW. DeltaFosB: тәуелділіктің тұрақты молекулалық қосқышы. Proc Natl Acad Sci АҚШ A. 2001; 98 (20): 11042-6. [PMC тегін мақала] [PubMed]
10. Teegarden SL, Bale TL. Диеталық преференциялардың төмендеуі эмоционалдылықты жоғарылату және тамақтанудың қайталануы үшін қауіп тудырады. Biol Psychiatry. 2007; 61 (9): 1021-9. [PubMed]
11. Olausson P, Jentsch JD, Tronson N, Nestler EJ, Taylor JR. dFosB ядросындағы акгембандар тамақпен күшейтілген аспаптық мінез-құлық пен мотивацияны реттейді. Неврология ғылымы журналы. 2006; 26 (36): 9196-9204. [PubMed]
12. Чен Дж, Kelz MB, Zeng G, Sakai N, Steffen C, Shockett PE, Picciotto MR, Duman RS, Nestler EJ. Мидағы индуктивті, мақсатты гендік өрнектегі трансгенді жануарлар. Мол фармакол. 1998; 54 (3): 495-503. [PubMed]
13. Келз МБ, Чен Дж, Карлосон В.А., Джр, Уислер К, Гильден Л, Бекманн А.М., Стефен С, Чжан Ю.Д., Маротти Л., Self DW, Tkatch T, Baranauskas G, Surmeier DJ, Neve RL, Duman RS, Picciotto MR, Nestler EJ. Мидағы deltaFosB транскрипция коэффициентінің экспрессиясы кокаинге сезімталдықты бақылайды. Табиғат. 1999; 401 (6750): 272-6. [PubMed]
14. Бале TL, Dorsa DM. Эстрогеннің окситоцин рецепторларының рецепторларында жыныс айырмашылығы және эффектісі рентгендік қышқылдың вентромидтік гипоталамуста экспрессиясы. Эндокринология. 1995; 136 (1): 27-32. [PubMed]
15. Avena NM, Long KA, Hoebel BG. Қантқа тәуелді егеуқұйрықтар қышқылдан кейінгі қантқа жауап беруді жақсартады: қанттың жетіспеушілігінің дәлелі. Physiol Behav. 2005; 84 (3): 359-62. [PubMed]
16. Will JJ, Franzblau EB, Kelley AE. Nucleus accumbens mu-opioids бөлінген ми желісін белсендіру арқылы майсыз диетаны қабылдауды реттейді. J Neurosci. 2003; 23 (7): 2882-8. [PubMed]
17. Чжэн Х, Паттерсон Л.М., Бертонд HR. Ворральдік ойық аймағында орехиннің сигналы ядро акументінің опиоидты ынталандыруымен туындаған майдың жоғары тәбеті үшін қажет. J Neurosci. 2007; 27 (41): 11075-82. [PubMed]
18. Хоммел Дж.Д., Тринко Р, Сирс РМ, Джорджеску Д, Лю З.В., Гао Х.Б., Турмон Ж.Ж., Маринелли М, ДиЛеон Р.Ж. Ортаңғы допаминдік нейрондарда лептин рецепторлары сигнал беруді тамақтандыруды реттейді. Нейрон. 2006; 51 (6): 801-10. [PubMed]
19. Colby CR, Whisler K, Steffen C, Nestler EJ, Self DW. DeltaFosB төтенше жасушалық типтегі ерекше көріністері кокаин үшін ынталандырады. J Neurosci. 2003; 23 (6): 2488-93. [PubMed]
20. Zachariou V, Bolanos CA, Selley DE, Theobald D, Cassidy MP, Kelz MB, Shaw-Lutchman T, Berton O, Sim-Selley LJ, Dileone RJ, Kumar A, Nestler EJ. Морфиндік әрекетіндегі ядро акументіндегі DeltaFosB үшін маңызды рөл. Nat Neurosci. 2006; 9 (2): 205-11. [PubMed]
21. Lee KW, Kim Y, Kim AM, Helmin K, Nairn AC, Greengard P. D1 және D2 допаминдік рецепторларында ядроның акумбензияларындағы ортаңғы майда нейрондардағы кокаиндік дозаланған омыртқа қалыптастыру. Proc Natl Acad Sci АҚШ A. 2006; 103 (9): 3399-404. [PMC тегін мақала] [PubMed]
22. Blendy JA, Maldonado R. Есірткіге тәуелділіктің генетикалық талдауы: cAMP жауап элементін байланыстыратын белоктың рөлі. J Mol Med. 1998; 76 (2): 104-10. [PubMed]
23. Nestler EJ. Нашақорлықтың молекулалық механизмдері. Нейрофармакология. 2004; 47 1: 24-32. [PubMed]
24. Танис К.К., Думан РС, Ньютон СС. CREB байланыстары және мидағы белсенділік: аймақтық ерекшелігі және электроконвульсивті тәрізді индукция. Biol Psychiatry. 2007
25. Кумар А, Чой К.Х., Ренталь В, Цанкова Н.М., Theobald DE, Truong HT, Russo SJ, Laplant Q, Sasaki TS, Whistler KN, Neve RL, Self DW, Nestler EJ. Хроматинді қайта құру - стриатумдағы кокаин-индуцирленген пластиканың негізгі механизмі. Нейрон. 2005; 48 (2): 303-14. [PubMed]
26. Грэхэм Д.Л., Edwards S, Bachtell RK, Dileone RJ, Риос М, Self DW. Кокаин қолданумен ядро акументіндегі динамикалық BDNF белсенділігі өзін-өзі басқаруды және қайталануды арттырады. Nat Neurosci. 2007; 10 (8): 1029-37. [PubMed]
27. Svenningsson P, Nairn AC, Greengard P. DARPP-32 бірнеше есірткінің теріс пайдаланылуына көмектеседі. Aaps J. 2005; 7 (2): E353-60. [PMC тегін мақала] [PubMed]
28. Палмер А.А., Вербицкий М., Сюреш Р, Камен Х.М., Рид К., Ли Н, Бурхарт-Кашь С., МакКиннон С.С., Белкнап Дж.К., Гиллиам Т.К., Филлипс Т.Ж. Метамфетаминнің сезімталдығы үшін тышқандардағы геннің экспрессиялық айырмашылықтары әр түрлі. Маммем геномы. 2005; 16 (5): 291-305. [PubMed]
29. Богуш А, Педрини С, Пелта-Хеллер Дж, Чан Т, Янг Q, Мао З, Слуцас Е, Гиирингер Т, Эрлич М.Э. АКТ және CDK5 / p35 орташаланған майда нейрондардағы in vitro DARPP-32 ми-туындаған нейротрофиялық факторды индукциялауды медитативті түрде жүргізеді. J Biol Chem. 2007; 282 (10): 7352-9. [PubMed]
30. Benavides DR, Bibb JA. Cdk5-тың есірткіге және пластиктегі рөлі. Анна Нью-Йорктегі Acad Sci. 2004; 1025: 335-44. [PubMed]
31. Бибб JA, Chen J, Taylor JR, Svenningsson P, Nishi A, Snyder GL, Yan Z, Sagawa ZK, Ouimet CC, Nairn AC, Nestler EJ, Greengard P. Кокаинге созылмалы әсердің әсерлері Cdk5 нейрондық протеинмен реттеледі. Табиғат. 2001; 410 (6826): 376-80. [PubMed]
32. Blochl A, Sirrenberg C. Нейротрофиндер Trk және p75Lntr рецепторлары арқылы егеуқұйрықтардың мезенсфалық нейрондарынан допамин шығаруды ынталандырады. J Biol Chem. 1996; 271 (35): 21100-7. [PubMed]
33. Бертон О, McClung CA, Dileone RJ, Krishnan V, Renthal W, Russo SJ, Graham D, Tsankova NM, Bolanos CA, Rios M, Monteggia LM, Self DW, Nestler EJ. BDNF-дің мезолимикалық допамин жолында әлеуметтік жеңіліске ұшырауындағы негізгі рөлі. Ғылым. 2006; 311 (5762): 864-8. [PubMed]
34. Nestler EJ, Carlezon WA, Jr Депрессияда мезолимбикалық допаминді сыйақы тізбегі. Biol Psychiatry. 2006; 59 (12): 1151-9. [PubMed]
35. Форд КП, Бекстед М Дж, Уильямс Дж.Т. Каппа-опиоидті допаминді ингибиторлық postsinaptic токтарының ингибирлеуі. J Neurophysiol. 2007; 97 (1): 883-91. [PubMed]
36. Nylander I, Vlaskovska M, Terenius L. Фишер және Льюис егеуқұйрықтарында мейн динорфин және энкефалин жүйелері: морфиндік төзімділік пен шығарылудың әсері. Brain Res. 1995; 683 (1): 25-35. [PubMed]
37. Nylander I, Hyytia P, Forsander O, Terenius L. Prodynorphin және proenkephalin жүйелеріндегі алкогольді-ұнататын (АА) және алкогольді-аулақ (ANA) егеуқұйрықтар арасындағы айырмашылық. Alcohol Clin Exp Res. 1994; 18 (5): 1272-9. [PubMed]
38. Крейк М.Ж. Кокаин, допамин және эндогендік опиоидтық жүйе. J Addict Dis. 1996; 15 (4): 73-96. [PubMed]
39. Carlezon WA, Jr, Duman RS, Nestler EJ. CREB-нің көптеген тұлғалары. Трендтер Neurosci. 2005; 28 (8): 436-45. [PubMed]
40. Dudman JT, Eaton ME, Rajadhyaksha A, Macias W, Tacher M, Barczak A, Kameyama K, Huganir R, Konradi C. Dopamine D1 рецепторлары Ser897-NR1-та NMDA рецепторларының фосфорлануы арқылы CRE фосфорлануын медиацияға шақырады. J Neurochem. 2003; 87 (4): 922-34. [PubMed]
41. Өзіндік DW. Мезолимикалық допамин жүйесіндегі нейродараптаулар арқылы препараттарды қабылдау және көру әрекетін реттеу. Нейрофармакология. 2004; 47 1: 242-55. [PubMed]
42. Beitner-Johnson D, Nestler EJ. Морфин және кокаин допаминергическом ми сыйақы аймақтарында тирозин гидроксилазасы бойынша жалпы созылмалы іс-әрекеттерді қолданады. J Neurochem. 1991; 57 (1): 344-7. [PubMed]
43. Lu L, Grimm JW, Шахам Я, Үміт Б.Т. Алғашқы 90 күн ішінде егеуқұйрықта кокаинді өздігінен басқаруға мәжбүрліктен ауытқу кезінде акументалды және вендральді тикеальдық аймақта молекулярлық нейродараптаулар. J Neurochem. 2003; 85 (6): 1604-13. [PubMed]
44. Шепард Дж.Д., Чуанг Д.Т., Шахам Ю., Моралес М. Метоламиканың және тиражиннің гидроксилазы және допаминді тасымалдағыштарындағы метамфетаминнің өзін-өзі басқарудың егеуқұйрықтардың мезолимик және нигростриалальді допамин жолдарындағы әсері. Психофармакология (Берл) 2006; 185 (4): 505-13. [PubMed]
45. Fulton S, Pissios P, Manchon RP, Stiles L, Frank L, Pothos EN, Маратос-Flier E, Flier JS. Мезоакцмендердің допамин жолын лептин реттеу. Нейрон. 2006; 51 (6): 811-22. [PubMed]
46. Нарита М, Nagumo Y, Miyatake M, Ikegami D, Kurahashi K, Suzuki T. Жасушадан тыс допамин деңгейлерінің орексиндік интенсивтілігінің жоғарылауы және оның марапаттау әсері. Eur J Neurosci. 2007; 25 (5): 1537-45. [PubMed]
47. Нарита М, Nagumo Y, Хашимото С, Хотиб Дж, Миятак М, Сакурай Т, Янагисава М, Накамачи Т, Шиода С, Сузуки Т. Месолимиктің допамин жолын белсендіруде немесе морфинмен туындаған мінез-құлықтарды белсендіруде орексинергиялық жүйелерді тікелей тарту. J Neurosci. 2006; 26 (2): 398-405. [PubMed]
48. Borgland SL, Taha SA, Sarti F, Fields HL, Bonci A. Орексин А ВТА-да синаптическая пластикалық индукция және кокаинге мінез-құлықты сезімталу үшін өте маңызды. Нейрон. 2006; 49 (4): 589-601. [PubMed]
49. Park ES, Yi SJ, Kim JS, Lee HS, Lee IS, Seong JK, Jin HK, Yoon YS. Орегин-А және нейропептидтің ептерді тамақтандырылған ашытылған және жоғары майлы диетадағы гипоталамуста Y өрнектегі өзгерістер. J Vet Sci. 2004; 5 (4): 295-302. [PubMed]
50. Wortley KE, Chang GQ, Davydova Z, Leibowitz SF. Тамақ қабылдауды реттейтін пептидтер: гипертриглицеридемия жағдайында орексин генінің экспрессиясы артады. Am J Physiol Regul интегрі Comp Physiol. 2003; 284 (6): R1454-65. [PubMed]
51. Чжэн Х, Коркнер М, Стоянова I, Паттерсон Л.М., Тян Р., Бертонд HR. Тамақтануды реттейтін пептидтер: аппетит-индуцирующего акументальды манипуляция гипоталамикалық орексин нейрондарын белсендіреді және POMC нейрондарын тежейді. Am J Physiol Regul интегрі Comp Physiol. 2003; 284 (6): R1436-44. [PubMed]
52. Балдо Б.А., Гуал-Бонилла Л, Сиапати К, Дэниэл РА, Landry CF, Kelley AE. Орексин / гипокритинді қамтитын гипоталамикалық нейрондардың субпопуляциясын GABAA рецепторлары арқылы ядролық акументалды қабықтың ингибирлеуі арқылы қосу, бірақ жаңа ортаға әсер ету арқылы емес. Eur J Neurosci. 2004; 19 (2): 376-86. [PubMed]
53. Гаррис Г.К., Виммер М, Астон-Джонс Г. Марапаттауды іздестіру кезінде бүйірлік гипоталамикалық нейрондық нейрондардың рөлі. Табиғат. 2005; 437 (7058): 556-9. [PubMed]