Neurosci Biobehav Rev. 2011 Jan; 35 (3): 939-55. doi: 10.1016 / j.neubiorev.2010.10.014. Epub 2010 қараша 2.
- 1Бернар Б. Броди Калиари университетінің нейрологиясы кафедрасы, Cittadella Universitaria, 09042 Монсанрато, КА, Италия. [электрондық пошта қорғалған]
КӨМЕК
Окситоцин - орталық жүйке жүйесіне енгізілгенде, penile erection қуатты индукторы. Ерлер егеуқұйрықтарында, омитоциннің протеко-эффективті әсері үшін ең сезімтал ми аймағы гипоталамустың паравенциклдік ядросы болып табылады. Бұл ядро мен айналасындағылар аймақтағы барлық омитосинергические нейрондардың жасушаларының денелерін гипоталамус миының және жұлын миының аймағына шығарады. Бұл шолу омитоцин осы аймақтың кейбір жерлеріне (мысалы, вентальдік тикеальды аймақ, гиппокампустың вентральдік субикулумы, амигдаланың постеромедиальді кортикальды ядросы және торако-белдік жұлын) инъекция жасайды. Интер-церебральды микродиализ және қос иммунофлуоресценттік зерттеулермен біріктірілген микроингеология зерттеулері осы аудандардағы омитоцинді тікелей немесе жанама түрде (негізінен глутамин қышқылымен) мезолимбалық допаминергиялық нейронды белсендіреді деп болжайды. Ядролық акумбензияларда шығарылған допамин, өз кезегінде, паравенциклы ядродағы инкраф-гипоталамикалық допаминергиялық нейрондарды белсендіруге әкелетін нейрондық жолдарды белсендіреді. Бұл омыртқа жұлынына және медиаторлы жыныстық эрекцияға арналған омитосинергические нейронды ғана емес, сонымен қатар жоғарыда көрсетілген қосымша гипоталамикалық аймақтарды ойлап табатын, миолимикалық допаминергиялық нейрондардың белсенділігі мен сыйақысын бақылайтын тікелей немесе жанама (глютамин қышқылымен) модуляциялау қызметін белсендіреді. Бұл нейрондық жолдар бірге сексуалды белсенділіктің тұтынушы фазасында (эректильді функция және копуляция) ғана емес, сондай-ақ жыныстық мінез-құлықтың алдын-алу кезеңінің мотивациялық және пайдалы аспектілерінде рөл атқаратын күрделі гипотетикалық схеманы құруы мүмкін.
1.Introduction
Пенсиялық эрекция - бұл адам секілді сүтқоректілердің көбеюінде басты рөл атқаратын ерлер жыныстық реакциясы, және бұл көбінесе көбінесе көбінесе көбінесе ұрпақпен қатыстырылған контексттерде байқалады. Penile erection орын алған контекстке қарай оның әртүрлі орталық және перифериялық жүйке және / немесе гуморальдық механизмдері (Meisel және Sachs, 1994, Argiolas және Melis, 1995, 2004, 2005, Sachs, 2000, 2007, McKenna, 2000, Giuliano және Rampin, 2000, 2004, Andersson, 2001, Melis және Argiolas, 1995a, 2003, Hull және т.б., 2002). Нейротрансмиттерлер мен нейропептидтердің арасында penile erection басқарады, ең танымал болып табылады допамин, серотонин, қоздырғыш аминқышқылдары, азот оксиді, адренокортикотропин, окситоцин және опиоидты пептидтер. Олар мидың кейбір жерлерінде әрекет жасай отырып, жыныстық эрекцияны жеңілдетеді немесе тежейді, гипоталамустың паравенцикулярлық ядросы, вентральдік тецикальды аймақ, гиппокампус, амигдала, стриа терминалының қабатының ядросы, ядролардың акумбензасы, медулла облигонаты және жұлын миы (Кесте 1) (Meisel және Sachs, 1994, Witt және Insel, 1994, Stancampiano және т.б., 1994, Argiolas and Melis, 1995, 2005, Argiolas, 1999, Bancila et al., 2002; Giuliano and Rampin, 2000, McKenna, 2000, Andersson, 2001, Hull және т.б., 2002, Coolen және т.б., 2004).
Нейрогипофизге арналған гипоталамустың паравенцикулярлық және supraoptic ядроларында орналасқан нейрондық нейрондық нейрондарда ғана емес, сондай-ақ нейрондардың жобасында да нейрондар мен еркектерде жақсы кездесетін нейрогипофосфералық пептид окситоцин бар паравенцикулярлық ядродан және қоршаған құрылымдардан экстрагипотумальды ми бөлімдеріне (яғни септум, вентральдік тикеальдық аймақ, гиппокампус, амигдала, медулла облонгата және жұлын). Бұл нейрондардың көптеген функциялары, мысалы, еске алу, оқу, аффилиирленген және әлеуметтік-сексуалдық мінез-құлықтар, соның ішінде пениса-эрекция және копуляциялық мінез-құлықтарға қатысы бар деп саналады (Buijs, 1978, Sofroniew, 1983, Argiolas және Gessa, 1991, Pedersen et al 1992, Wagner and Clemens, 1992, Ivell және Russel, 1993, Carter және т.б., 1995; Tang et al., 1997; Veronneau-Longueville және т.б., 1998). Шынында, окситоцин тышқандар, егеуқұйрықтар, қояндар мен маймылдарда эректикалық функцияны және жыныстық қатынастарды жеңілдетеді (Argiolas және Gessa, 1999, Carter, 1991, Pedersen және басқалар, 1992, Argiolas және Melis, 1992, 1995, Argiolas, 2004 қараңыз). Бұл адамдарда да орын алуы мүмкін, өйткені плазмалық окситоцин әсіресе эякуляция кезінде (Carmichael және басқалар, 1999, Murphy және т.б., 1987) және кеуде және гениталияны манипуляциялау кезінде көбінесе сексуалды ынталандырулар арқылы көбейтіледі жыныстық қатынас (Tindall, 1987).
Окситоцинді ерлердің жыныстық мінез-құлқы бойынша жеңілдететін әсері алғаш рет интенсивті омитоцинді бірінші эякуляцияға кідіртуді азайтуға және рецептивтік аналықтармен (Melin және Kihlstrom, 1963) жұптасқан жыныстық сарқылуын азайтуға қабілеттілікпен көрсетілді. Алайда, окситоциннің жыныстық әсері сексенінші жылдары ғана танылды. Нанограммалардағы центриальді түрде берілген окситоцин, ерлер егеуқұйрықтарында (Arletiol et al., 1985, 1986) пенинговая эрекцияны (Argiolas et al., 1985, 1985) ынталандырады және ерлер егеуқұйрықтарында скотчивтік мінез-құлықты жақсартады (Arletti және Bertolini, 1986, Caldwell et al., 1995), әдетте, uterine типті окситосинергиялық рецепторларда әрекет етеді (Argiolas және Melis, 2004, 1999, Argiolas, 2003, Melis және Argiolas, 1985 және сілтемелер қараңыз). Окситоцин тек жыныстық күшті ерлер егеуқұйрықтарында ғана емес, сонымен қатар ересек егеуқұйрықтарда (Arletti және т.б., 1990) және доминантты, бірақ бағынышты емес, еркектерге арналған ақжелкен маймылдарда (Winslow және Insel , 1991).
Окситоценнің протеэструкциялық әсері тестостеронға тәуелді, себебі ол гипофизэктомия мен кастрациядан кейін жойылады және тестостеронмен немесе оның метаболиттерімен, эстрадиолмен және 5_-дихидро-тестостеронмен бірге қалпына келтіріледі (Melis және т.б., 1994a). Омситоцин арқылы кремнийді эрекциялауды индукциялауға арналған ең сезімтал ми аймағы гипоталамустың паравенцикулярлық ядросы болып табылады (Melis et al., 1986), оның үстіне барлық қосымша гипоталамикалық окситоцинериялық проекциялар пайда болады (жоғарыдан қараңыз). Мұнда окситоциннің 3 pmol-ке дейін дозада енгізілгенде (төменде 2.1 бөлімін қараңыз) инъекцияға кремнийді эрекциялауды (және итеріп кетуді) қабілетті деп тапты. Сондай-ақ, окситоцинді гипокампустың CA1 өрісіне екі жақты енгізген кезде, сонымен қатар доральді субикулада (төменде 2.3 бөлімін қараңыз), бүйірлік септумды, каудиат ядросын, медиальдық пропоптикалық аймақты, гипоталамустың ventromedial ядросын және supraoptic ядросы (Melis және т.б., 1986). Окситоцин бұл жыныстық реакцияны тудыру үшін паравенциклы ядросында әрекет ететін механизмге келетін болсақ, көптеген зерттеулер окситоциннің өздерінің нейрондарын белсендіретінін көрсетеді. Осы гипотезаға сәйкес, сексуалдық өзара іс-қимыл, Penis Erecting бақылауына қатысатын жұлын жұлдыруға арналған паравенцикулярлық окситоцинергической нейрондардағы c-fos дереу ерте генінің гендік өнімін көбейтеді (Witt және Insel, 1994 және сілтемелерді қараңыз). онда жыныстық импотенттілік (мысалы ересек адамның егеуқұйрықтың эвтроген-прогестеронға қарсы рецептивтік әйелмен эвтроген-прогестеронға қарсы рецептивтік әйелмен күресуге қабілетсіздігі) гипоталамустың паравенцикулярлық ядросындағы төменгі деңгейлері бар окситоцин мРНК ерлерде (Арлет және т.б.) Ә., 1997).
Окситоциннің алдын-ала фазаға әсер етуі немесе жыныстық мінез-құлықтың тұтыну кезеңі анық емес. Окситоцинді пениса эрекцияны индуцирует және омитоцинді сопылық мінез-құлыққа әсер етудің негізгі себебі - ересек егеуқұйрықтардан кейінгі эякуляциялық аралықтың төмендеуі (Arletti et al., 1985), пептид жыныстық өнімділікті жақсартады деп санауға болады. Алайда, окситоцин әлеуметтік-сексуалдық өзара әрекеттесуді жақсартады (Pedersen және т.б., 1992, Carter және т.б., 1997, Ivell and Russel, 1995) және окситоцин рецепторлары антагонистері байланыссыз эрекцияларды (Melis et al., 1999a) жыныстық қатынасқа және сексуальды ынталандыруға окситоциннің ықтимал рөлін жоққа шығаруға болмайды (Sachs, 1997, 2000, 2007, Melis және т.б., 1998, 1999b және сілтемелер).
Бұл шолу соңғы зерттеулердің жарияланған және жарияланбаған нәтижелерін жинақтайды окситоцин гипоталамустың паравенцикулярлық ядросына енгізілгенде ғана емес, сонымен қатар басқа да қосымша гипоталамус миының аудандарында, мысалы, вентральдік тецикальды аймақта (Melis және т.б., 2007, 2009a, Succu және т.б., 2008), tол гиппокампустың вентральдік субикулумы мен амигдаланың артқы ядросы (Melis et al., 2009b, 2010), олар лимбиялық жүйенің маңызды құрамдас бөлігі болып табылады және ынталандыру мен марапаттау үдерістерінде шешуші рөл атқарады деп ойлайды. Бұл зерттеулер окситоциннің дозалық және глутамин қышқылы сияқты басқа да нейротрансмиттерлерді қамтитын нейротрансмиттерді және паравенциклы ядролардан басқа, мысалы, вентральдік тикеальдық аймақты, ядро акументінің, гиппокампустың және облыстың басқа да ми бөлімдерін қамтитынын анықтайды. анықталды. Бұл схемалар мезолимбикамен және тригер-гипоталамиялық допаминергиялық жүйенің өзара әрекеттесуіне және ерлердің жыныстық мінез-құлқының тұтыну фазасында (мысалы, пениса эрекциясы мен жиналуы) ғана емес, жыныстық ынталандыру және жыныстық қатынаста да рөл атқаруы мүмкін жыныстық белсенділіктің мотивациялық және марапаттау қасиеттерін түсіндіру үшін нейрондық субстратты қамтамасыз ету.
2. Окситоцин мидың әртүрлі аудандарында әрекет жасай отырып, кремний эрекциясына әсер етеді
2.1. Гипоталамустың паравенциклдік ядросы
Жоғарыда айтылғандай, гипоталамустың паравенциклы ядросы жақында омитоциннің протеэтилді әсері үшін ең сезімтал ми аймағы ретінде анықталған. Осы ядрода бір жақты енгізілгенде, окситоцин дозада 3 нг (3 pmol) (Melis et al., 1986) ретінде төменде белсенді табылды. Құрылымдық-белсенділік байланыстары бойынша зерттеулерде окситоцинді индуцированный кремний эрекциясы Ca2 + ағымына окситоцинергические нейрондардың жасуша органдарына қосымша гипоталамалық ми аймақтары мен азот оксид-синтазасын белсендіруге бағытталған uterine-oxytocin рецепторлары арқылы анықталды. Белгілі бір механизммен (гуанилат циклді емес) жасушааралық хабаршы ретінде әрекет ете отырып, азот оксид өз кезегінде омыртқа және гипоталамус миының аудандарына омыртқа ұшырайтын окситоцинергические нейрондардың белсенділігін арттырады, бұл ретте penile erection (сурет 1) төменде және Argiolas және Melis, 1995, 2004, 2005 және сілтемелер). Окситоциннің өз нейрондарын белсендіру мүмкіндігі
Сурет 1. (MISSING) гипоталамустың паравенциклы ядросынан шыққан окситоцинергические нейрондардың жұлын байланысы, VTA, гиппокампус, амигдала сияқты қосымша гипоталамус миының аудандарынан шыққан схемалық көрінісі. Бұл нейрондарды допамин, қоздырғыш аминқышқылдары, окситоциннің өзі, гекарелин аналогы пептидтері және VGF-алынған пептидтер GABAergic, opioid және каннабиноид CB1 рецепторларын ынталандыру арқылы төмендетілуі және / немесе жойылуы мүмкін penile erection әкеледі. Окситоцинергические нейрондарды активтендіру бұл нейрондарда болатын азот оксид-синтаза активтендіруіне байланысты. Допаминді, қоздырғыш аминқышқылдарды немесе окситоцина рецепторларын немесе экзогенді азотты оксидті азот оксидінің донорларынан тікелей паравенцикулярлық ядроға тікелей енгізілген ынталандыру арқылы пайда болған эндогендік азот оксиді, шын мәнінде, белгісіз механизм арқылы окситоцингиялық нейронды белсендіреді. гуанилат циклді ынталандыру. Бұл өз кезегінде омыртқадағы омыртқа және мидың гипоталамус аймағында босатылуына әкеледі. Окситоцин бұл аудандарда босатылған кезде, мысалы, ВТА, вентральдік субикулум және амигдала секілді мидың секцияларында сипатталған кремнийді эрекцияның механизмін анықтайтын тетіктер туралы кейбір мәліметтер келтірілген. Мұнда окситоцин өз рецепторларына әсер етеді және NO өнімін жоғарылатады, бұл PVN-де табылған кремнеземдеуге әкеледі. Дегенмен, PVN-мен айырмашылығы, каудальдық VTA NO-де гуанилат циклді белсендіріледі. Бұл мезолимик допаминергические нейрондарды активациялауға әкеліп, cGMP концентрациясының көбеюіне әкеліп соғады. VS NO-де глутамергенді нейрондар экстра-гиппокампальды аймақтарға, соның ішінде ВТС-ға қосылады. VTA ішіндегі глютамин қышқылы, мезитобинмен табылған, мезолимбалық допаминергиялық нейронды белсендіреді. Жоғарыда сипатталғандарға ұқсас механизмдер, сонымен қатар, жыныстық эрекцияның физиологиялық контексте орын алған кезде, әсіресе, егеуқұйрықтарды қол жетпейтін рецептивтік әйелдің (мысалы, байланыссыз эрекциялардың) қатысуымен немесе қопсыту кезінде орналастырылған кезде қолданылуы мүмкін.
паравенциклы ядросында келесі зерттеулерді көрсетті: (1) оксицоцина рецепторлары осы гипоталамикалық ядрода кездеседі (Freund-Mercier және басқалар, 1987, Freund-Mercier және Stoeckel, 1995); (2) окситоцин in vitro және in vivo өз шығарылымын жеңілдетеді (Freund-Mercier және Richard, 1981, 1984; Moos және басқалар, 1984); және (3) окситоцин өзінің жеке нейрондарын паравенциклдің ядросында әрекет ету арқылы қоздырады (Yamashita және т.б., 1987). Сонымен қатар, гемоталамустың паравенцикулярлық және supraoptic ядросында магиокелулярлы окситоцинергические нейрондардың жасушаларына зақым келтіретін омитосинергиялық синапстар анықталды (Theodosis, 1985). Ақырында орталық жүйке жүйесі мен жұлынның ортасында окситоцинді толығымен бұзатын паравенцикулярлық ядродағы электролиттік немесе химиялық экзототоксикалық зақымдану арқылы орталық окситоцинергические нейронды бұзу тек окситоциннің протеэстрилді әсерін ғана емес, сонымен бірге есірткіге тәуелді болады penile erection және байланыссыз эрекциялар (төменде қараңыз және Argiolas және т.б., 1987a, b, Liu және т.б., 1997 және оның сілтемелер). Паравенциклов ядросының зақымдануымен анықталған ұқсастықтар қуатты және таңдаулы омитоцина рецепторлары антагонисттерімен анықталады. Нанограммадағы паравенцикулярлық ядраға енгізілген бұл қосылыстар толық окситоцинді индукцияланған кремний эрекциясының алдын алады, ал бүйірлік қарыншаларға тек қана окситоцинмен туындаған пениса эрекцияларын ғана емес, сондай-ақ препаратты индукцияланған кремнийді монтаждауды болдырмайды (төменде 3 бөлімін қараңыз). және Argiolas және Melis, 1995, 2004, 2005 және сілтемелер) және байланыссыз эрекциялар (Melis және т.б., 1999a) және жыныстық күшті ерлер егеуқұйрықтарының (Argiolas et al., 1988) Бұдан басқа, сексуалдық өзара әрекеттесу Penis Erecting бақылауына қатысатын жұлын жұлынына арналған проекциялы паравенцикулярлық окситоцинергической нейрондардағы дереу ерте геннің c-fos генінің өнімді арттырады (Witt және Insel, 1994 және оның сілтемелерін қараңыз). Соңында, жыныстық импотенция (мысалы, ересек ер егеуқұйрықтың эстроген-прогестерон реакцияға ұшыраушы әйелге қарсы күресу қабілетсіздігі) hсондай-ақ, гемоталамустың паравенцикулярлық ядросында төменгі деңгейдегі окситоцин мРНК және азот-оксид синтазасы бар ер адамның егеуқұйрықтарымен байланыстырылған (Benelli және т.б., 1995, Arletti et al., 1997) (осы зерттеулерді кеңінен шолу үшін Argiolas, 1999, Argiolas және Melis, 2004, 2005 қараңыз).
2.2. Вентральдік тория аймағы
Вентральдік тикелиальді аймақ тек жақында мидың орны ретінде анықталды, оның құрамында окситоцин инъекцияға қарсы кремнийді құрастырады. Бұл аймақта паравенцикулярлық ядроның және окситоцина рецепторларынан (Freund-Mercier және т.б., 1987, Vaccari және т.б., 1998) туындайтын окситоцинергические жүйке ұштары бар. Дәлірек айтқанда, окситоцин i қабілетті болдыбіржақты тәртіпте каудальға енгізілгенде, бірақ роральды вентральдік тецикальды аймақта емес, дозаға тәуелді түрде (Melis et al., 2007). Белсенді дозалар паравенциклы ядросына енгізілген кезде талап етілгенге қарағанда жоғары болды және гиппокампустың вентральдік субикулумына немесе амигдаланың постеромедиальді кортикальды ядросына енгізілгенде (төменде қараңыз) инъецирленген кремний эрекциясына ұқсас. Шамалы, эстрогенді әсер эпиляцияға себеп болған, мезоолимикалық допаминергические нейрондардың белсендірілуіне себеп болады, бұл өз кезегінде ядролық аксмобиндердің қабығына түсіретін, бірақ, өз кезегінде, паравенцикулярлық окситосинергиялық нейрондарды тудыруға әкелетін тоғыз-гипоталамикалық допаминергиялық нейрондарға белгісіз нейрондық жолдарды белсендіреді (Melis және т.б., 2007, 2009a).
Окситоцина вендральді тикеальдық аймақта допаминергиялық нейротрансмиссия қызметін белсендіретін механизмге келер болсақ, қолда бар деректерге сәйкес, окситоцин месолимикалық допаминергиялық нейрондардың жасушаларында орналасқан омитосинергиялық рецепторларды ынталандырады. Бұл CaPNUMX + ағыны допаминергические нейрондардың жасушаларының корпусында артады, сол арқылы азот оксид-синтаза белсендіріледі (Succu және т.б., 2). Паравенциклы ядросымен айырмашылығы бар (төменде 2008 бөлімін қараңыз), азот оксиді өз кезегінде гуанилат циклді белсендіреді, осылайша циклдік GMP концентрациясын арттырады. Бұл механизмге сәйкес, d (CH3) 2Tyr (Me) 5-Orn2-vasotocin, күшті омитоцит антагонистері немесе нейрональды азот оксид синтезінің күшті ингибиторы S-methyl-thio-l-citrulline окситоцин алдында каудальды вентральдік тесікальды аймақ, пениса эрекциясының жойылуы және окситоцинмен туындаған ядролардың акумбензияларының қабығындағы допаминнің шамадан тыс концентрациясының жоғарылауы. Сонымен қатар, 8-bromo-циклдық GMP, белсенді фосфодиэдеразға төзімді циклдік GMP аналогы, каудальды вендральді энциклопатия аймағына енгізген кезде пениса эрекциясын индуцирует және ядро акммобленің қабығындағы допаминнің концентрациясын арттырады. Каудальный вендральді аймақ (Succu және т.б., 8, Melis et al., 2008a) (сонымен бірге сур. 2009).
Әрдайым осы механизмге сәйкес, галоперидол, ядро акорменсінің қабығына енгізілген күшті допамин D2 рецепторлық антагонисті вендралды техемальді аймаққа енгізілген окситоцинмен туындаған кремний эрекциясын төмендетеді (Melis et al., 2007). Жоғарыда аталған механизмге қос иммунофлуоресценттік зерттеулермен қолдау көрсетіледі, бұл окситоцин талшықтары бұрынғы ядролардың акумбензияларының қабығына енгізілген флюороголдпен белгіленетін каудальды веналық тикеальдық аймақта допаминергиялық нейрондардың жасушаларының денелеріне төтеп беретінін көрсетті ( Melis және т.б., 2007, Succu et al., 2008). Осы допаминергические нейрондардың және дупаминді рецепторлардың ядролардың акумбензиндеріндегі белсенділіктері кезекпен анықталатын нейрондық жолдардың белсендірілуіне әкеліп соқтырады, бұл паравенциклы ядродағы допаминді босату үшін тепе-гипоталамикалық допаминергиялық нейронды ынталандырады, осылайша, окситоцинериялық нейрондарды жұлын және делдалдық кремнийді құрастыру (Жоғарыда қараңыз және Melis және т.б., 2007, Succu және т.б., 2007, 2008). Шынында да, окситоцин протеинді эрекцияға әкелетін дозада каудальды вентальдік тикеальды аймаққа енгізілді, ядро акумбензынан ғана емес, сонымен қатар паравенцикулярлық ядродан (Succu және т.б., 2007) алынған диализатта экстра-жасушалық допамин концентрациясын арттырды.
2.3. Гиппокампус
Хиппокампаның CA1 өрісі омитосинергиялық талшықтарға бай басқа ми алаңы болды және бұрынғы зерттеулермен анықталған рецепторлар, онда омитоцинді индуцированную кремнийді инъекциялау (Bujis, 1978, Sofroniew, 1983). Дегенмен, паравенцикулярлық ядродан айырмашылығы, бұл жерде омитоцинді паравенцикулярлық ядродағы белсенділігінен (Melis et al., 1986, Chen және т.б., 1992) қарағанда екі жақты және жоғары дозада инъекцияға енгізген кезде ғана бар. Осы ерте зерттеулерде индукцияға субкумуляцияға окситоцин енгізілді. Дегенмен, жақында және мұқият микроингеология зерттеулері а вендральді субикулум аймағы онда омитоцинді инъекция дозаға тәуелді түрде (Melis et al., 2009b) penile erection индукциясына қабілетті болатын. Бұл ми аймағына енгізілген омитоциннің протекоэтилен әсері паравенциклы ядросында табылған бір жақты инъекциядан кейінгі (Melis et al., 2007) кейінгі вендральді тикеальдық аймақта белсенді болғандарға ұқсас дозада байқалды. Әлбетте, вентальдік субикулумға енгізілген омитоцин азот оксиді синтезі бар нейрондарда оксицоцеринді рецепторларды активтендіру арқылы пениса эрекциясын тудырады, бұл азот оксидінің өндірісін ұлғайтады. Натрий оксиді, өз кезегінде, жасушааралық хабаршы ретінде жұмыс жасай отырып, глютамин қышқылының нейротрансфемациясын белсендіреді, мүмкін, вендральдік субикулумдан нейрондық (глутамергиялық) эфференттік проекциялар арқылы мүмкін (мысалы, төменгі және Melis, 2007, 2009b, Succu және т.б., 2008) мезолимбиялық допаминергиялық нейрондардың белсенділігін модуляциялайтын қосымша гиппокаппалы ми бөлімдеріне (мысалы, вентральдік тикеальдық аймақ, префронтальді кортекс, паравенциклы ядросы).
Іс-әрекеттің бұл механизмі интенсивті микродиализ эксперименттері арқылы қолданады, бұл пенициллинді индуцирлеуді арттыратын дозада вендральдік субикулумға енгізілген, витральдік субикумадан диализатта азот оксидінің өндірісін және глютаминді қышқыл концентрациясын арттырады (Melis et al. , 2010) және ядролардың акумбензияларындағы жасушадан тыс допамин (Melis et al., 2007). Бұл жауаптар тек қана окситоцинді рецепторлардың антагонистері d (CH2) 5Tyr (Me) 2-Orn8-вазотоцинмен ғана емес, сондай-ақ нейрональды азот оксидті-синтетаз ингибиторы S-метил-тио-лицитуллинмен және азотты оксиді қышқылды гемоглобинмен антагонизацияланған (Melis және басқалар, 2010) бірнеше минут бұрын вентальдік субикумға енгізіңіз.
Оның үстіне, осы механизмге сәйкес, NMDA арқылы глутамертерлік нейротрансляцияны белсендіру вендральдік субикумаға енгізілді (Melis et al., 2010). Қазіргі уақытта мезолимикалық допаминергической нейронды белсендіруді және ядролардың акумбензасында экстра-жасушалық допаминді жоғарылатуды тудыратын вентральдік субикулдан эфференттік проекция фенотипі белгісіз. Дегенмен, вентральдік субикулаға енгізілген окситоцин тудырған кремний эрекциясы, вентральдік тикеальдық аймақтан диализатта клеткалық емес глютамин қышқылының ұлғаюына байланысты, бірақ ядро accumbens емес және (+) MK-801 арқылы антагонизацияланған кезде, NMDA subtype (Woodruff және соавт., 1987) вирустық тикеальдық аймаққа енгізілген, бірақ ядро accumbens емес (қараңыз 2 және Melis және т.б., 2009b) аэротоптардың аминқышқылдары рецепторларының бәсекелеспейтін антагонисті , бұл болжаулар вентальдік тюкальді аймақта глутамертериялық нейротрансмиссияның белсендірілуіне әкелуі мүмкін, бұл, өз кезегінде, ядролардың акумбензына жасайтын мезолимикалық допаминергиялық нейрондарды белсендіреді. Вентральдік тикаликальды аймақта табылған глутамин қышқылының концентрациясының жоғарылауы вентральдік субикулумға окситоцинді инъекциядан кейін субикулада пайда болатын нейрондардан немесе басқа ми жерінде (мысалы, префронтальді қыртыстық) шыққан нейрондардан босатылады. Дегенмен, бұл мезолимик допаминергические нейрондардың белсендірілуіне және ядро акументіндегі допаминнің босатылуына себеп болады. Мұнда допаминді рецепторлардың белсендірілуі инкраф-гипоталамикалық допаминергиялық нейрондардың белсендірілуіне әкеледі, бұл паравенцикулярлық ядрода допаминді босатады, осылайша омыртқалы жұлдыруға арналған омитосинергические нейрондарды белсендіреді, (Жоғарыда қараңыз және Melis және т.б., 2007, 2009a, Succu және т.б., 2008).
2.4. Амигдалла
Омитоцин талшықтарына және рецепторларға бай басқа ми бөлімі - амигдала (Freund-Mercier және т.б., 1987, Vaccari және т.б., 1998, Uhl-Bronner және т.б., 2005). Мұнда окситоцинді анхиолизден, әлеуметтік жадтан және танымнан, әлеуметтік күшейтілген оқытудан, эмоционалдық эмпатиядан, эмоционалды тұлғаны өңдеуден және эректикалық функцияны және жыныстық мінез-құлықтан қорғанудан әртүрлі функцияларға тартылған деп санайды (Kondo et al., 1998; Dominguez et 2001, HNUMX, HNUMX, HNUMX, HNUMX, HNUMX, Hurnum et al., 2005, Lee және т.б., 2005, Donaldson and Young, 2007, Hurlemann et al. , 2008). Алайда, амигдаланың постеромедиальді кортикальды ядросында пайда болған кезде, ересек егеуқұйрықтарда окситоцинді жыныстық эрекциялауды тудыру мүмкіндігі жақында ғана анықталды (Melis et al., 2009b). Бұл реакция вентальдік субикулумға (Melis et al., 2009b) окситоцинді инъекциядан кейін табылған ядролардың акумбензияларының қабығынан алынатын диализатта экстра-жасушалық допамин концентрациясының артуына байланысты болды. Қазіргі кезде амигдаланың постеромедиальдық кортикальды ядросына омитоцин енгізілген кремний эрекциясының механизмі белгісіз. Қолда бар деректер көрсеткендей, penile erection және ядро акumbens алынған диализатта жасушалық емес допамин концентрациясының жоғарылауы окситоцинергические рецепторлардың активтендіруі арқылы жүзеге асырылады, өйткені жауаптардың екеуі де окситоцина рецепторлары антагонисты d (CH2010) 2009Tyr ( Me) OXYTOCIN (Melis et al., 2009b) бірнеше минут бұрын амигдала ядросына енгізілген 2-Orn5-вазотоцин.
Амигдаланың постеромедиальды кортикальді ядросында қандай механизм болмасын, пептидпен туындаған жыныстық реакция барлық допаминергический рецепторлардың қоршауымен ядро акументінің қабығына енгізілген cis-flupentyixol және NMDA рецепторларының қоршауымен ( +) MK-801 вендральді тикальді аймаққа енгізілді, бірақ вендральдік субикулумға енгізілген окситоцин тудырған кремний эрекциясында табылған (Melis et al., 2009b) ядросы акументіне емес. Бұл амигдада постеромедициналық ядросына енгізілген омитоциннің вентальдік тюкальді аймақта глютамин қышқылының нейротрансмиссиясын белсендіретінін көрсетеді. Бұл, өз кезегінде, мезолимик допаминергические нейрондардың белсендірілуіне әкеліп соғады. Амигдаланың осы ядросын вентальдік субикулуммен (Canteras және т.б., 1995, француз және Totterdell, 2003) өзара байланыстыратын нейрондық жолдарды көрсететін зерттеулерді ескере отырып, бұл зерттеулер екі ми бөлімі арасында өзара әрекеттесу мүмкін екендігін көрсетеді амигдалдан немесе ядро акулементіне немесе вентральдік еккемалға дейінгі жолдар сипатталған (Kelley және Домесик, 1982, Witter, 2006).
2.5. Жұлын
Жұлын омыртқасы - орталық жүйке жүйесінің басқа аймағы, ол окситоцинериялық талшықтарды және рецепторларды (Freund-Mercier және басқалар, 1987, Uhl-Bronner және т.б., 2005) қамтиды, онда окситоцин инсулинді эрекцияны тудырады (Tang et al., 1998 , Veronneau-Longueville және басқалар, 1999, Giuliano and Rampin, 2000, Giuliano et al., 2001). Жоғарыда айтылғандай, бұл омитосинергиялық талшықтар гемоталамустың паравенцикулярлық ядросынан шығып, креатинді эрекциялауға арқа сүйейтін омыртқан автономды нейрондарды басқаратын жолдарды төмендетуге ықпал етеді. Шынында да, бұл талшықтар тораптық мүйізде прогганглионикалық симпатикалық және парасимпатикалық жасушалық бағаналардың синаптивті байланыстарын жасайды, олар торако-люмбальды және лумбо-сакральды траекториядағы жұлын нейрондарымен (Marson and McKenna, 1996; Giuliano and Rampin, 2000; Giuliano et al. , 2001). Бұл синаптивтік байланыстар жыныс мүшесінен шыққан жұлын нейрондарының таңбалануы және кварцтық протеинге енгізілген белгілі бір ретроградтық іздері бар жұлынға жету арқылы көрсетілді, оның құрамында қос иммунофлуоресценция және конфокальды лазерлік микроскопиямен (Tang et al., 1998, Veronneau-Longueville және т.б., 1999). Осы зерттеулерге сәйкес, анестезизацияланған ерлер егеуқұйрықтарында инкратэкальді окситоцинді лумбо-сакралда, бірақ торако-белдік деңгейінде кумулятивтік дозаны инъекциялау дозаға тәуелді түрде күштірек қысыммен көтеріледі. Бұл эффектілерге оксицоцериндік рецепторлардың (d) (CH2) 5Tyr (Me) 2-Orn8-вазотоцинмен және жамбас нервтерінің бөлігімен (Giuliano және Rampin, 2000, Giuliano және т.б., 2001) блокадасы жойылды. Бұл нәтижелер лумбо-сакралды жұлын іс-әрекетінде қолданылатын окситоциннің іштің қысымын арттыратынын көрсетеді және паравенциклы ядросының физиологиялық белсенділігі кезінде босатылған омитоцинді омыртқа проректорлы нейрондардың күшті активаторы деп біледі. Бір қызығы, омитотиннің протеко-эффективтік әсерін қолданатын протеэтилсиялық жұлын нейрондары, сондай-ақ, медулла облигоната (Marson and McKenna, 1992, Tang et al.) Ядроларының парагигантоселулярларынан шыққан серотонинергические нейрондардан синаптивтік байланыстар алады. ., 1998). Осы серотонинергические нейрондардың жойылуы ерлер егеуқұйрықтарында эякуляция мен кремний рефлекстерін жеңілдетеді (Marson and McKenna, 1992, Yells және т.б., 1992). 5HT2C рецепторларын ынталандыратын препараттар 5HT2C рецепторларында, сонымен қатар, паравенцикулярлық ядрода емес, сондай-ақ, 5HT2C рецепторларының допамин агонисті және омитоцинді индукцияланған кремний эрекциясын төмендететін кремнийдің эрекциясын жеңілдетеді, ал допаминді антагонисты 1994HT5C агонисті индуцируемді криминалді эрекцияны азайтпайды ( Stancampiano және т.б., 2 және сілтемелер қараңыз), сондай-ақ, окситоцин люминесцентті жұлын деңгейінде (Stancampiano және т.б., 1994) деңгейінде протеэтиленді XNUMXHTXNUMXC рецепторлардың әрекетін жеңілдетеді деп ұсынылды. Немесе, окситоцин жұлынның төмендеген серотонинергические нейрондарының белсенділігіне тікелей әсер ете алады, бұл нейрондар пайда болған парагигантелоэлуляр ядросында (Stancampiano және т.б., 1994 қараңыз).
3. Цито-жүйке жүйесіндегі және гинексті эрекциялау кезінде окситоцин, допамин және глютамин қышқылының өзара әрекеттестігі
1 бөлімінде айтылғандай, орталық жүйке жүйесінде бар барлық окситоцинергические нейрондар паравенцикулярлық ядрода және қоршаған құрылымдарда пайда болады. Бұл нейрондардың қызметі әртүрлі нейротрансмиттерлер және / немесе нейропептидтердің бақылауында. Паравентрикулярлық деңгейде ең көп зерттелгендер арасында допамин, глутамин қышқылы, гамма-аминобутир қышқылы (GABA), азот оксиді, эндоканнабиноидтер, опиоидты пептидтер, өсім гормонын босату пептидтері, ВГФ-пен байланысты пептидтер және окситоцин болады. Допамин, глутамин қышқылы, өсу гормонын босату пептидтері, VGF-алынған пептидтер және окситоцин ынталандырушы болып табылады, мысалы, осы қосылыстар мен олардың агонисты паравенцикулярлық ядросына енгізгенде кремний эрекциясын жеңілдетеді, ал GABA, опиоидты пептидтер және эндоканнабиноидтер ингибиторлық болып табылады, мысалы, бұл қосылыстар немесе олардың агонистері кремнеземдеуді тежейді (Meisel және Sachs, 1994, Witt және Insel, 1994, Argiolas және Melis, 1995, 2004, 2005, Giuliano және Rampin, 2000, 2004, McKenna, 2000, Andersson, 2001, Hull және т.б., 2002).
Бірнеше эксперименттік дәлелдер осы окситоцинергические нейрондар мен жоғарыда көрсетілген нейротрансмиттерлер мен нейропептидтердің әр түрлі физиологиялық контексттерде эректильді функцияны және жыныстық мінез-құлықты бақылауға қатысқандығын көрсетеді. Сонымен қатар, омитоцин қосымша вирустық гипоталамус миының аудандарында, мысалы, вентальдік тикеальды аймақ, гиппокампус және оның өңірлері, амигдала және жұлын сиқырлы окситоцинергиялық синапстар әсер ететін нервтердің белсенділігіне әсер етуі мүмкін. Қазіргі уақытта ядролардың акумбензияларының қабығына шығаратын каудальды вентальдік тикеальдық аймақтың месолимбиялық допаминергиялық нейрондарының жасушалы органдары (Melis et al., 2007 (Succu және т.б., 2008), сондай-ақ, лумбо-сакральды тракттан кавероцидтік протонға дейін шығаратын протекторлы жұлындық нейрондар (Giuliano және Rampin, 2000, Giuliano және т.б., 2001) (2.2 және 2.5 бөлімдерін де қараңыз) . Шынында да, омыртқа синергиялары мен рецепторлары вендральдік субикулумда, сондай-ақ, агента және жұлын мүйізінде, криминалді эрекция үшін маңызды (жоғарыдан қараңыз), анықталған болса да, осы аудандарда нейротрансмиттердің / ол окситоцинергические нервтердің аяқталуы, әлі белгісіз.
Зерттеудің осы бөлімінде каудальды вентральдік тикеальдық аймаққа енгізілген окситоциннің эволюциялық әсерінің, гиппокампустың вентральдік субикулумының және жұлынның ішіне енгізілген механизмдер туралы қысқаша әңгімелер қысқаша баяндалады. Пептидтің осы облыстарда допаминмен және глютамин қышқылымен өзара әрекеттесуіне және осы өзара әрекеттесудің эректиль функциясының орталық бақылауында рөл атқаратына ерекше көңіл бөлінеді. Допамин мен глутамин қышқылының эволюциялық функцияда шешуші рөл атқаратын паравенциклы ядродағы окситоцинергические нейрондарға әсер етудің қысқаша мазмұны бірінші болып оқырманға осы саладағы зерттеулердің ерте сатысы туралы хабардар ету үшін беріледі. бұл зерттеулер кеңінен қаралды (Argiolas және Melis, 1995, 2004, 2005, Melis және Argiolas, 2003 қараңыз). Сондай-ақ, бұл жағдайда соңғы кездегі нәтижелерге айрықша көңіл бөлінеді, бұл допамин-окситоциндік байланыс пен глютамин қышқылы-окситоцинді жыныстық қатынаста ғана емес (жыныстық эрекция және жинақтау) ғана емес, ойын-сауық және жыныстық ынталандыру.
3.1. Паравенциклдің ядросындағы допамин-окситоцин өзара әрекеттесуі
The допамин агонистерінің орталық окситоцинергической нейрондарды активациялау арқылы пениса эрекциясын тудыру қабілеті апоморфин инъекциялық бровентрикулярлы (icv) инъекцияланған окситоцин рецепторлары антагонистері d (CH1987) 2Tyr (Me) 5-Orn2-вазотоцин кезде табылған кезде паравенцикул ядросына (Melis және т.б., 8) (Argiolas et al., 1987b) тері астына енгізілген апоморфинмен ғана емес, сонымен бірге тек окситоцинмен ғана емес, сонымен қатар индукцияланған пениса эрекциясын азайтуға қабілетті. Бұл нәтижелерге d (CH2) 5Tyr (Me) 2-Orn8-вазотоцинді ICV бергенде және апоморфин тікелей паравенциклы ядросына (Melis et al., 1989b), lдопамин агонисты қосымша гипоталамиялық ми бөліктеріне, әсіресе жұлын миына дейін шығаратын паравенцикулярлық окситоцинергической нейрондарды белсендіру жолымен, кремнийді эрекциялауды тудырады деп болжауға тырысады (Argiolas және Melis, 1995, 2004, 2005 қараңыз). Осы гипотезаға сәйкес анестезирленген егеуқұйрықтарда помпит емес окситоцинді рецепторлық антагониста лумбо-сакральды окситоцинергический рецепторлардың қоршауында допамин агонисті апоморфинмен туындаған интромарфиндік туындаған қысымның көтерілуін жоюға қабілетті. Паравентрикулалық-омыртқа окситоцинергиялық жолдарда (Baskerville et al., 2009).
Зерттеулерге бағытталған Динамикалық эрекцияның индукциясына жауап беретін допаминді рецепторды анықтау, сонымен қатар, паравенцикулярлық ядродағы допаминді рецепторлы агонистерде D2 тобының допаминді рецепторларында әрекет жасай отырып, кремнийді эрекциялауды тудырадыжүйелі түрде берілген допаминді рецепторлы агонистермен (Melis және т.б., 1987, Eaton және т.б., 1991, Melis and Argiolas, 1995a қараңыз). Тиісінше, осы гипоталамикалық ядраға енгізілген таңдаулы D1 рецепторлы агонисті SKF 2 емес, күшті араластырылған D2 / D38393 рецепторлы агонисті және күшті аралас D1 / D2 рецепторлы агонисті бар apomorphine, дозаға тәуелді түрде жыныстық эрекцияны тудыруы мүмкін , ал D2 рецепторларының агонистері арқылы туындаған жыныстық реакция D23390 рецепторлары антагонисты арқылы жойылды, мысалы, Haloperidol және l-sulpiride сияқты, бірақ SCH 1 емес, таңдаулы D1987 рецепторлық антагонисті (Melis et al., 1999). Пароментриулярлық ядраға енгізілгенде апоморфинді пенициллинді эрекциялауды ынталандыру мүмкіндігі телеметриялық зерттеулермен дәлелденді, бұл паравенциклы ядрода берілген допамин агонисті жүйелі қан қысымын өзгертпей ояту еркек егеуқұйрықтарында интракаверональды қысымды арттыра алады (Chen et al. , 2001; Giuliano және Allard, 1999) жүйелік инъекциядан кейін табылған (Bernabè және т.б., XNUMX). Бұл зерттеулер де D2 рецепторларының басты рөлін растады, себебі D1 рецепторлы агонисты әдетте пештенгендік ядроға енгізілгенде интракаверналық қысымның жоғарылауына қабілетсіз деп табылды (Chen және т.б., 1999).
Кейінгі экспериментальды дәлелдердің бірнеше жолдары қол жетімді болды, бұл ынталандыру индукциясына penile erection, окситоцинергические нейрондық жасуша органдарында орналасқан паравенциклы D2 рецепторларының екенін көрсетеді. Біріншіден, паравенциклы ядросында допаминергиялық нерв клеткалары бар, олар инкстерогипоталамиялық допаминергиялық нейрондарға жатады. Бұл нейрондардың жасушалары A13 және A14 тобында орналасқан Dahlstrom және Fux (1964), неврогипофиз және / немесе гипоталамус миының қосымша аймақтарына арналған паравенцикулярлық окситоцинергические нейрондарды қоса алғанда, бірнеше гипоталамикалық құрылымдарды кеңінен және инерсивті түрде жасайды (Buijs және т.б., 1984, Lindvall et al., 1984).
Осы допаминергические нейрондарды паравенцикулярлық деңгейде penile erection және copulation бақылауында тарту микродиализ зерттеулерімен, оның негізгі метаболиттері, оның негізгі метаболиттері экстрацеллюлозалық допамин және 3,4-дигидроксипенил қышқылы концентрациясы (DOPAC) жыныстық қуатты еркек егеуқұйрықтарының паравенцикулярлық ядросы қол жетпейтін овариэктомизацияланған эстроген + прогестеронмен асқынған рецептивтік әйелдің (Melis et al., 2003) қатысуымен байланыссыз эрекцияларды көрсетеді.
Допамин мен DOPAC концентрациясының жоғарылауы медиальдық пропоптикалық аймақта (Hull және т.б., 2003) табылған және рецептивтік әйелге сіңіру кезінде рұқсат етілген (Melis et al., 1995) және ядро accumbens (Pfaus және Everitt, 1995). Екіншіден, бірнеше зерттеулер көрсеткендей, пенициллиндры DECNUMX рецепторларының ынталандырылуы арқылы туындаған, осы облыстарда шығарылған окситоцин арқылы жүзеге асады. Тиісінше, протонды эрекцияны тудыратын дозада берілген апоморфин тек егеуқұйрықтар мен маймылдардың плазмасында (Melis және басқалар, 1989a, Cameron және т.б., 1992) омитоцин концентрациясын арттыра алады, сонымен қатар гипоталамикалық ми мысалы, гиппокамп (Melis et al., 1990). Осы нәтижелерге сәйкес апоморфин дозада дозада паравенцикулярлық ядроға енгізілді, бұл ретте penile erection соңғы уақытта ядро accumbens ішіндегі допаминнің қосымша клеткалық концентрациясын арттыра алатындығын көрсетті, бұл әсері oksithocin рецепторлары антагонисты d (CH2) 5Tyr (Me) 2-Orn8-вазотоцинді вендральді аймаққа енгізді (Succu және т.б., 2007; Melis et al., 2009a) (4 секциясын қараңыз). Үшіншіден, паравенцикулярлық ядролардың екі жақты электролиттік зақымдануы, оттситоцинді қосымша гипоталамикалық ми алаңдарынан толығымен жойып тастайды (Hawthorn және т.б., 1985), апоморфинді индуцированный кремний эрекциясын (Argiolas және т.б., 1987a) және селективті окситоцинді рецепторлық антагонисттерді бүйірлік қарыншаға берілген, бірақ паравенцикулярлық ядрода емес, бұл қосылыстардың құрамына окситоцинді рецепторларға кедергі келтіретін (Melis et al., 1989b) потенциалмен дозаны тәуелді түрде апоморфинді-құйылған кремний эрекциясын төмендетеді. Окситоцин рецепторларының антагонисты тек окситоцинмен ғана емес, сонымен қатар апоморфинмен (Argiolas және т.б., 1988, 1989) индукцияланған ерлердің жыныстық мінез-құлқын жеңілдетуге өте күшті.
ДхНУМС рецепторлары допаминмен немесе допаминді рецепторлы агонистермен белсендіретін механизмге қатысты окситоцинергической нейрондардың белсенділігін арттырады, осылайша экстрагипоталамиялық ми аймағында және жұлындағы омитоцин босатылады, көптеген эксперименттік деректерD2 рецепторларының ынталандыруы окситоцинергические нейрондардың жасушаларының органдарының ішіндегі Ca2 + иондарының концентрациясын арттыратын гипотезаны қолданады, ол азот оксид-синтаза активизациясына әкеледі, Бұл клеткалық органдарда кездесетін Ca2 + -calmoduline тәуелді ферменттер (Vincent және Kimura, 1992, Torres және т.б., 1993, Sanchez et al., 1994, Sato-Suzuki және т.б., 1998). Азот оксидінің өсуі, өз кезегінде, окситоцинергические нейрондарды белсендіруге себеп болады. Тиісінше, (1) апоморфинге ұшыраған кремнийді эрекция органикалық кальций арналарын блокаторларымен және N-типті кернеуге тәуелді Ca2 + каналдарының қуатты және іріктейтін блокаторы (McCleskey және т.б., 1987) паравенцикулярлық ядро (Argiolas және т.б., 1990 және оның сілтемелерін қараңыз); (2) апоморфинді индуцированный кремний эрекциясына паравенцикулярлық ядрода берілген (Melis et al., 1994c) азот оксид-синтета ингибиторлары арқылы жол берілмеді; және (3) apomorphine және басқа да D2 рецепторлық агонистері инсульт-церебральды микродиализден алынған паравенцикулярлық диализатта азот оксидінің өндірілуін арттыратын дозада берілген, бұл дозада аз мөлшерде аз мөлшерде аз мөлшерде азотты оксид-синтаза ингибиторлары арқылы азайтылған D2 рецепторларының кремнийді эрекциясы (Melis et al., 1996). Нутрицит оксицоцинергические нейрондарды белсендіретін азотты оксиді әлі күнге дейін белгісіз, бірақ қол жетімді деректерде азот оксиді жасушааралық хабаршы ретінде әрекет ететінін және гуанилат циклосының қатыспайтындығын көрсетеді. Шынында да, циклдік GMP, 8-бромоциклдік GMP белсенді фосфодидуға қарсы тұру аналогы паравенцикулярлық ядроға (сурет 2) берілгенде (қараңыз Melis және Argiolas, 1995b және сілтемелер) пениса эрекциясын тудыруы мүмкін емес.
Жоғарыда келтірілген түсінік көбінесе сенімді емес деп саналады, негізінен допамин D2 рецепторларын ынталандыру, әдетте, осы рецепторларды қамтитын нейрондық клеткалардың денелерін әртүрлі G протеині байланыстырылған механизмдер арқылы қозғауға емес, ингибирлеуге байланысты (Sokoloff және Schwartz, 1995 қараңыз). Допаминмен паравенцикулярлық окситосинергические нейрондарды тікелей ынталандыруға негізделген осы сәйкессіздіктің ықтимал түсіндірмесі жақын арада табылған. G протеинге тәуелді допамин D4 рецептор, D2 рецепторлық отбасының мүшесі (D2, D3 және D4), бұл ынталандыру осы рецепторлардың ішкі түрінің (Moreland және т.б., 2) клондалған нұсқасын қамтитын жасушалық препараттарда Ca2004 + ағынын арттырады. Ең бастысы, іріктелген D4 рецепторлы агонисті (мысалы, ABT 724) (N-methyl-4- (2-цианофенил) пиперазинил-ХНУМХ метилбензамид малеататы) жүйелі түрде ұсынылғанда егеуқұйрықтарды жыныстық эрекциялауды тудыруға қабілетті деп табылды (Brioni және басқалар, 3). Бұл әсер іріктелген D2004 рецепторлардың PNU-2E (R-95666-dihydro-N, N-dimethyl-5,6Himidazo [4-i] quinolin-4,5,1-амині) (Hsieh және т.б., 5) агонисті ақ Ca2004 + ағыны D2 рецепторлардың подтипінің клондалған нұсқасын қамтитын жасушалық препараттарда көбейте алмады (Brioni және басқалар, 4, Moreland және т.б., 2004). Жоғарыда келтірілген гипотеза мен қорытындыға сәйкес PD 2004 (N-methyl-168,077- (4-цианофенил) пиперазинил-2 метилбензамид малеаты), PIP-3EA (3- [2- (4-methoxyphenyl) piperazin-2-ylmethyl] imidazo [1-а] пиридин) және басқа да таңдаулы D1,2 рецепторлы агонисттері (Heier және т.б., 4; Melis et al., 1997b; Löber et al., 2006) жүйелі енгізілгенде, пароментриулярлық ядро, бірақ кемінде тиімді қарағанда апоморфина. D2009 рецепторлы агонистерінің противоррекциялық әсері L-4 (745,870- (3- (хлорфенил) пиперазин-ХНУМХ-йл) -метил-ХНУМХХ-пирроло [4-B] пиридин трихидрохлоридімен) алдын-алуға болды, таңдаулы D1 рецепторы антагонист (Patel және т.б., 1, Melis және т.б., 2,3, 4b, Löber және т.б., 1997).
Ақыр соңында, жоғарыда көрсетілген D4 рецепторлы агонистердің противоррекциялық әсері паравенцикулярлық ядрода берілген азот оксидті-синтета ингибиторлары және д (CH2) 5Tyr (Me) 2-Orn8-вазотоцинмен, селективті окситоцин рецепторлық антагонистімен азайтылды берілген, бірақ paraventricular ядросында емес. Бұл нәтижелер D4 рецепторларының агонистері азот оксид синтезін активтендіру жолымен окситоцинергические нейронды ынталандырады және экстрадипоталамикалық ми бөлімдерінде омитоцинді босату арқылы ынталандырады, бұл өз кезегінде апоморфин және классикалық D2 агонистеріне (Melis et al. ., 2005, 2006b; Löber және т.б., 2009).
Жоғарыда келтірілген зерттеулер сонымен қатар, допаминнің паравенцикулярлық окситоцинергических нейрондық клеткалық органдарында орналасқан D4 рецепторларында әрекет ететін пенсиялық эрекцияны индукциялайтын гипотезаны қолдайды және окситоцинергические нейрондық клеткалардың Ca2 + ағынының ұлғаюына алып келеді, бұл азот оксидінің ұлғаюына алып келеді. Өз кезегінде нитролит оксицоциклы нейрондарды белсендіреді, ол талқыланғандай, омыртқасыз мидың аудандарында және жұлындағы омитоцинді босатады. Бұл жөнінде, паравенцикулярлық ядродағы окситоцинергические нейрондардың жасушалық органдарында допаминді рецепторлар анықталғанын атап өту керек жақында ғана жоғары селективті D2, D3 және D4 рецепторлы антиденелерімен және окситоцинді антиденелермен қос иммунофлюоресценттік зерттеулермен. Бұл зерттеулер паравенцикулярлық ядродағы (сондай-ақ супраоптикалық ядро мен медиальдық префоптическом аймағында) окситоцинергические нейрондардың жасушалық органдарында бөлек-бөлек араласатын барлық үш D2receptor (D2, D3 және D4) подтипдерінің (Baskerville және Douglas, 2008, Baskerville және т.б., 2009).
Бұл D2 түріндегі допамин және допаминді рецепторлы агонистердің экстрагипоталамиялық ми бөліктеріне тікелей бағытталған омитосинергиялық нейрондарды белсендіре отырып, криминалистикалық эрекцияны тудыру мүмкіндігін күшейтетін нейроанатомиялық қолдауды қамтамасыз етедімысалы, жұлын, вентальдік тектік аймақ, гиппокампус және амигдала. Дегенмен, бұл деректер D2 рецепторларының подтип / түрлерін анықтау үшін ешқандай көмек бермейді, оның ынталандырылуы эректильді жауапты тудырады. Өкінішке орай, паравенциклы ядродағы допаминді рецепторлы агонистермен белсендірілген окситоцинергические нейронды анықтауға бағытталған зерттеулерден тіпті ешқандай көмекті алуға болмайды. Шынында да, әртүрлі допаминді рецепторлардың ішкі түрлерінде әртүрлі әрекетке қарамастан, аралас допаминді рецепторлы агонистер (мысалы, апоморфин) немесе таңдамалы D2 рецепторлы агонистер (мысалы, барлық D2 рецепторлы қосалқы түрлерінде әрекет ететін хинпирол) немесе таңдамалы D4 рецепторлы агонистері паравенцикулярлық ядроның парвоселулярлық окситоцинергические нейрондарында FOS протеинінің жоғарылауымен өлшенетін (Bitner және т.б., 2006) окситоцинергические нейронды активтендіру. Дегенмен, бұл табылу жақында күмән туындады, себебі паровентриарлы окситоцинергические нейрондардағы FOS протеинінің көбеюі, негізінен, D2 және D3 рецепторлардың шағын түрлерінде әрекет ететін, бірақ D168077 рецепторлы агонисті емес, PD 4 емес, қосылыстардың екі жыныстық реакцияны тудыруына қарамастан (Baskerville et al., 2009).
Содан кейін басқа D2 рецепторлардың субтитрлерінің (негізінен D2 және D3) іріктеп агонистерімен бірге әр допаминді рецепторлардың субпірінің паравенциклдік деңгейде эректильді функцияны бақылаудағы нақты рөлін анықтау қажет. Осыған байланысты бұрынғыдай еске салынғандай, барлық допаминді рецепторлардың шағын түрлерінде (бұр. Brioni және т.б., 2004 және оның сілтемелері) тиімді әрекет ететін апоморфин, паравенцикулярлық инъекцияға енгізілгенде, penile erection инициализациясында D4 рецепторларының агонистеріне қарағанда әлдеқайда тиімді ядро. Бұл D4 рецепторларында D4 рецепторларына қарағанда, апоморфиннің жоғары жақындығымен түсіндіріледі, немесе D4 рецепторлы агонисты D4 рецепторлы агонистері D2 рецепторларының ішінара агонистері ретінде әрекет етуі мүмкін немесе әртүрлі допаминді рецепторлардың подтипдерін апоморфинмен біріктіру мүмкін D4 рецепторларының субтитрлерінің DXNUMX рецепторларының агонистерін активтендіруге қарағанда, кремнийді эрекциялауға арналған окситоцинергические нейронды жоғары белсендіруді қамтамасыз етеді.
Допамин D1 және D2 рецепторлары арасындағы өзара әрекеттер медиальдық пропоптикалық аймақ деңгейінде жыныстық мінез-құлықты бақылауда сипатталған (Hull және т.б., 1989 қараңыз). Селективті D2 рецепторлы агонистердің кремний эрекциясын (Hsieh және т.б., 2004) индуцирлеуге қабілетсіздігі (мысалы, Depoortère және т.б., 2009), мысалы, осы қосылыстардың паравенциклы ядросына енгізілгеннен кейін де расталуы мүмкін , D3 рецепторлары үшін немесе D4 рецепторларымен бірігіп, кремнийдің эритмесіне кедергі келтіретін окситоцинергические нейрондарды белсендіруде басты рөлді талдау қажет (Baskerville және т.б., 2009 қараңыз). Өкінішке орай, таңдаулы D2 және D3 рецепторларының агонистері (мысалы, осы екі рецепторлардың ішкі түрлерінде олардың кемінде төрттен бес рет in vitro тәртібінде айырмашылығы бар), қазіргі уақытта қол жетімді емес. Осы себепті D3 рецепторлары классикалық D2 рецепторларының агонистері арқылы индуцирленген клиникалық D3 агонистері сияқты клиникалық D2 рецепторларының антагонистері негізінде in vitro эксперименттермен сипатталатын, негізінен, болжамды D2 рецепторларының антагонисттеріне негізделген, барлық Dopamine D2009 рецепторлардың субтиптерін (Collins және т.б., XNUMX) белсенді түрде белсендіретін апоморфин, кинприрол және прамипексольді басқа эксперименттермен растау қажет. Бұл тексеруді D4 рецепторларының агенттері үшін penile erection-ға әсер етпеуі керек, себебі D4 рецепторларының агонистерінің протек-ректорлық әсерін көрсететін жоғарыда келтірілген зерттеулердің нәтижелерімен ерекшеленеді. Шынында да, апоморфиннің жабайы түрдегі D4 рецепторлардың нокаутқа тышқандарында көрінетін және D4 рецепторлық антагонисттерінің осы жануарлардағы апоморфиндік әсерін жоюға қабілеттілігі бар D3 рецепторларының нокаутқа тышқандарында penile erection (және yawning) Collins және т.б., 2009) D3 рецепторларындағы Agonist-индуцированного кремний эрекциясында D2 рецепторлардың подтипінің таңдаулы рөлі үшін дәлелдеме ретінде қарастырыла алмайды. Нейротрансмиттер / нейропептид және / немесе нейротрансмиттер / нейропептидті рецепторлық ген абляциясымен (нейротрансмиттер / нейропептид және / немесе нейротрансмиттер / нейропептидті рецепторлардың нокаут жануарлары) зерттеулерде әдетте нейротрансмиттердің болжамдық жыныстық ролін растауда және одан әрі күрделендіріп, нейропептидтер және олардың рецепторлары. Атап айтқанда, окситоцин генін абляциялау омитоцинді тышқандарды ұрлайды, ол әдеттегідей жұптасады және сопақады, мысалы, окситоцин жұғу және сіңіру үшін қажет емес секілді. Сондай-ақ, гомозиготты әйел окситоцинді нокаутқа арналған тышқандар сүт жоғалтудың нашарлауымен (Nishimori et al., 1996; Young et al., 1996) бар болса да қалыпты жұту және босануды көрсетеді. Нейрональды азот оксидінің синтезін кодтайтын геннің абляциясы, сондай-ақ азот оксидінің синтетасын шығарады, ол қалыпты түрде жұтылып, әдеттегідей копорирленген (Huang et al., 1993). Алайда, бұл нәтижелер репродуктивті физиологияның маңызды ерекшелігін, яғни орталық және перифериялық деңгейде бақылауға тартылған жүйелердің артық болуын көрсетеді.
Мұндай резервтеу, әрине, эволюциялық шыққан, өйткені ол түрлердің тірі болуы үшін келесі ұрпаққа гендердің өтуіне кепілдік береді. Сондықтан, D4 рецепторлық генінің абляциясы апоморфиннің протеэстильді әсерін өзгертпейді екендігі фактісі D4 рецепторлары, мысалы, окситоцин және азот оксидтері, эректиль функциясын басқаратын жүйелерде жұмыс істейтін делдалдардың тек бірнешеуі ғана екенін білдіреді бұл рецепторларға пениса эрекциясының және жыныстық мінез-құлықтың бақылауында рөлі жоқ екенін көрсетеді. D4 агонистерінің әртүрлі штаммдарының ерлер егеуқұйрықтарына жүйелі түрде жүйелі түрде тағайындалған кремний эрекциясының енгізілуін сәтсіз аяқтауы басқа зерттеуде (Depoortère et al., 2009). Алайда Collins және басқаларының жұмысына қарағанда, (2009), сондай-ақ әртүрлі D2 рецепторлардың ішкі түрлерінің роліне суретке түсіру үшін тағы да таң қалдырады, бұл зерттеу жүйелі түрде берілген жүйелі D3 рецепторлық антагонисттерінің апоморфинді индуцирленген перифериялық эрекцияны төмендете алмайтындығын көрсетеді еркек егеуқұйрықтары апоморфиннің протеэстрилді әсеріне аса сезімтал, ал жыныстық реакция (және егеу) таңдап алынған D2 антагонисті L-741,626 (3 - [[4- (4-хлорфенил) -4-хлорофиперидин D1 және D1 рецепторларының орнына D2 рецепторларының D3 агонисті индуцирленген пениса эрекциясында (Depoortère et al., 4) маңызды рөл атқаратыны туралы авторларға жетекшілік ететін жетекші -2-yl] methyl-2009H-индол). Ақыр соңында, допаминді рецепторлардың агонистердің окситоцинергические нейрондарға әсер етуі, кем дегенде, бір бөлігі, тікелей емес, жанама емес, мысалы, делдалды немесе басқа нейротрансмиттерлердің нейропептидтерінің белсенділігіне әсер етуі мүмкін паравенциклы ядросындағы окситоцинергические нейрондарды толығымен шығаруға болмайды.
3.2. Паравенцикулярлық ядродағы глютаминді қышқыл-окситоцин өзара әрекеттесуі
Гипоталамустың паравенциклы ядросы нейротрансмиттер (мысалы, глютамин қышқылы және аспарагин қышқылы) сияқты акуытқыш аминқышқылдары бар синапстарға өте бай.) (Van Den Pol, 1991). Осы ядродағы қозғаушы аминқышқылдар көптеген функцияларға, соның ішінде пениса эрекциясына және жыныстық қатынасқа (Roeling және басқалар, 1991, Melis және т.б., 1994b, 2000, 2004b) қатысады. Тиісінше NMDA рецепторлы подтипінің селективті агонисті N-метил-д-аспартқыш қышқылы (NMDA), бірақ (±) -_- амино-ХНУМХ-гидрокси-ХНУМХ-метил-изоксазол-3-пропион қышқылы (AMPA) , метаботропты рецепторлардың субтитрінің селективті агонисті AMPA рецепторлы кіші типіндегі немесе (±) -транс (5) -амино-ХНУМХ-циклопентан дикарбоксил қышқылының (ACPD) селективті агонисті табылды. еркін қозғалатын егеуқұйрықтардың паравенциклдік ядросы (Melis et al., 4b). 1 MR Melis, A. Argiolas / Neuroscience және Biobevavenvensional Reviews 1,3 (1994) 948-35 NMDA (+) MK-2011 (бәсекелес NMDA рецепторлары) антагонисті (Woodruff және т.б., 939 ), паравенциклдің ядросына енгізілген (Melis et al., 955b). Осы нәтижелерге сәйкес, телеметрия зерттеулерінде іштің қысымын бақылауға бағытталған телеметрия зерттеулерінде, NMDA басқа оваторлық аминқышқыл рецепторларының агонисттеріне қарағанда, паравенцикулярлық ядраға оятқыш немесе анестезирленген еркек егеуқұйрықта инкравитарлы қысымның күшеюі кезінде енгізілген (Zahran et 801, Chen және Chang, 1987).
Жоғарыда айтылғандай, окситоцин мен допамин үшін, несеп-жыныс корпусында медицинада эрозияны реттейтін NMDA рецепторлары, окситоцинергические нейрондардың жасушалық органдарында орналасқан, себебі, (Van Den Pol, 1991). Допаминді рецепторлы агонистермен анықталған нәрсеге ұқсас, NMDA-ның противоэффективті әсері оксицоцериндік нейротрансмиссияның белсендірілуіне байланысты, ол окситоцинн антагонисты d (CH2) 5Tyr (Me) 2-Orn8-вазотоцинмен жойылды, бірақ Paraventricular ядрода емес (Argiolas және Melis, 1995, 2004, 2005 және оның сілтемелерін қараңыз). Сонымен қатар, NMDA индукцияланған окситоцинергической нейротрансмиссиясы белсендірілуі азот оксид-синтаза активтендіруіне байланысты, өйткені NMDA индуцированного кремний эрекциясына азот оксидті-синтета ингибиторлары (N-Nitro-N-methyl-arginine methyl ester және N- paraventricular ядросына берілген метил-тио-1-цитруллин) және NMDA пенициллинді эрекцияны тудыратын дозаларда паравенциклы ядросына енгізеді, гипоталамус ядросында азот оксидінің өндірісін арттырады (Argiolas және Melis, 1995, 2004, 2005 және оның сілтемелерінен қараңыз). ). Допаминді рецепторлардың агонистеріне қатысты, NMDA индукцияланатын азот оксидисинтазы белсенділігінің Ca2 + ағыны Ca2 + ағынымен NMDA рецепторлары арқылы Ca1992 + арна арқылы байланысқан NMDA рецепторлары арқылы Ca1993 + ағынының қосындысы болуы мүмкін, себебі бірнеше нервтік препараттарда (Snyder, 1994 қараңыз); Southam және Garthwaite, XNUMX, Schuman және Madison, XNUMX және онда сілтемелер). Nitric oxide өз кезегінде окситоцинергической нейротрансмиссиясын белсендіреді (жоғарыдан қараңыз). Кейбір нейроанатомиялық және электрофизиологиялық дәлелдер, кем дегенде, ішінара, гиппокампада (Saphier және Feldman, 1987, Chen және т.б., 1992) туындауы мүмкін екеніне қарамастан, паравенцикулярлық окситосинергические нейрондарды белсендіретін глутамертерлік проекциялардың белгісіздігі белгісіз, . Глутамергические проекциялардың шығу тегі паравенцикулярлық ядросына жақсырақ сипаттау үшін одан әрі жұмыс жасау қажет болса да, микробқа қарсы зерттеу арқылы жыныстық эрекция мен жыныстық қатынасқа бақылауда паравенцикулярлық ядрода глютамин қышқылын тарту. Тиісінше, эстогенді + проргеороннан босатылған рецептивтік егеуқұйрықтардың пайда болуына байланысты ересек егеуқұйрықтардың паравенцикулярлық ядросынан алынған диализатта глутамин қышқылының және аспарагин қышқылының қосымша клеткалық концентрациясы ұлғайтылды (Melis et al. 2004b), сондай-ақ орталық окситоцинергиялық берілуді белсендендіру арқылы пайда болған кремний эрекциялары (Melis et al., 1999a, b). Мұндай көтерілуді қабылдайтын әйелмен байланыстыру кезінде рұқсат етілген (Melis et al., 2004a). Эквиторлық аминқышқылдардың белсенділігін жоғарылату паравенцикулярлық ядрода пениса эрекциясында және копуляция кезінде пайда болады деп болжанған гипотезаға сәйкес, неконтактивтік эрекциялар мен сопуляциялы кристаллдық мінез-құлық (оның ішінде copula penile erections кезінде) паравенциклы ядродағы NMDA рецепторларының қоршауымен азаяды , және бұл төмендеу осы физиологиялық контексттерде (Melis et al., 2000) осы гипоталамус ядросында пайда болатын азот оксидінің өндірісін жоғарылатудан кейін байқалады. CB1 антагонисты SR 1A арқылы каннабиноид CB141761 рецепторларының қоршауынан кейін паравенцикулярлық ядрода табылды, сонымен қатар ГОБА-ның GABAergic нервтердің аяқталуынан гастроэнтериялық глютамин қышқылының шоғырлануын жоғарылату, бүйірлік қарыншаға немесе тікелей эпиляторлы ядрода дозада дозада енгізіңіз (Succu және т.б., 2006, Castelli және т.б., 2007). Осындай ұлғаю нитрон оксидінің синтезін азот оксидінің өндірісін ұлғайтып, окситоцинергические нейрондардың жасушалық органдарында белсендірілуіне әкелді. Nitric oxide, өз кезегінде, жоғарыда сипатталғанындай, penile erection-ға делінген окситоцинергические нейронды белсендіреді. Мұндай механизмге сәйкес, SR 141761A индуцированный кремний эрекциясы NMDA рецепторларының қоршауымен және азот оксидті-синтета ингибиторлары арқылы азайтылды, бірақ паравенцикул ядросындағы допамин немесе окситоцина рецепторларының қоршауымен емес, сонымен қатар, оксицоцина рецепторлары антагонистері арқылы орталық омитоцина рецепторлары
3.3. Вентральдік тикігальді аймақта окситоцин-допаминді өзара әрекеттесу
Окситоцин дозаға тәуелді түрде вендралды ториялайтын аймақтың каудальды бөлігіне енгізген кезде, кремнийдің эрекциясын индуцирует (Melis және т.б., 2007). Бір жақты тәртіпте енгізілген ең белсенді доза 20 нг болды, ал ең жоғарғы доза - 100 нг болды. Окситоцин әсері окситоцинерді рецепторлардың белсендірілуі арқылы жүзеге асырылады, себебі жыныстық реакция caudal ventral tegmental аймағында окситоцинді антагонистик d (CH2) 5Tyr (Me) 2-Orn8-вазотоцинді инъекциялау арқылы жойылады. Бұл рецепторлар допаминергиялық нейрондардың жасуша органдарында орналасады, олар негізінен ядролардың акумбензияларының қабығына бағытталған. Тиісінше, (1) қос иммунофлюоресценттік зерттеулер каудальды веналық тикеальдық аймақта допаминергические нейрондардың жасушаларымен тығыз байланыста болатындығын көрсетеді, олардың көпшілігі триозин-гидроксилазға оң белгіленіп, бұрынғы ретроградтық бақылаушы Fluorogold бар (Melis et al., 2007) және (2) вентральды тіккен окситоцинді индуцированный кремний эрекциясы қабықшасынан алынатын диализатта экстра-жасушалық допамин концентрациясының ұлғаюымен бірге жүреді ядролардың акормендері (Melis et al., 2007). Сонымен қатар, окситоцинді индуцированный кремний эрекциясы вентральдік тецикальды аймақта азот оксидінің өндірісін жоғарылатуымен қатар жүреді, бұл жауаптар тек D (CH2) 5Tyr (Me) 2-Orn8-вазотоцинмен емес, сондай-ақ азот оксиді синтетаз ингибиторы S- метил-тио-1-цитруллин, сонымен қатар кернеуге тәуелді Ca2 + арналарын блоктаушы _-конотоксин және ODQ (1H- [1,2,4] оксадыазол [4,3-а] квинокалин-1-бір), гуанилаттың күшті ингибиторы цитокразды, барлық окситоцин алдында каудальды вентральдік тецикальды аймақта берілген (Succu және т.б., 2008). Сондай-ақ, азот оксид-синтазы және гуанилат циклаза (Succu және басқалар, 2008), окситоцин үшін оң белгіленді, сондай-ақ, каудальды вентральдік тецикальды аймақта тирозин гидроксилазасы үшін оңтайлы деп табылған флюорогольдтік допаминергические денелер, Пениллингтің төменгі механизмі келесі тетік арқылы жүзеге асырылуы мүмкін. Допаминергиялық жасушалардағы окситоцинерлік рецепторларды пептидпен активтендіру CaPNUMX + ағыны допаминергические нейрондардың жасушаларының корпусында артады. Бұл nitric oxide-synthase, Ca2 + -калмодулинге тәуелді ферментті белсендіреді, осылайша азот оксидінің өндірісін ұлғайтады. Өз кезегінде азот оксиді гуанилат циклді белсендіреді, бұл Циклдік GMP концентрациясының артуына алып келеді. Циклді GMP ядролардың акументіне арналған допаминергиялық нейрондарды белсендіреді. Каудуральды веналық тикеальдық аймаққа енгізілген окситоцинмен индукцияланған циклдік GMP рөлі, сондай-ақ, циклдық GMP белсенді фосфодиэстеразыға төзімді аналогты 8-бромо-циклдық GMP қабілеті арқылы қолдайды каудальды вендральді тикеальдық аймақ және ядролардың акумбензынан диализатта жасушалық емес допаминді ұлғайтады (Succu және т.б., 2008, Melis et al., 2009a). Бұл натрий тотығы осы ядрода (Melis және Argiolas, 8b) инъекция кезінде penile erection индуцировать алмайтын 1995-bromo-циклдық GMP болып табылатын паравенцикулярлық ядродағы окситоцинергические нейронды белсендіретін механизммен ерекшеленеді. 2). Ядролық акументингтегі кремнийдің эрекциясына әкелетін допаминмен белсендірілген нейрондық жолдарға келетін болсақ, бұл әлі белгісіз. Дегенмен, қолда бар деректер осы жолдар гипоталамустың паравенцикулярлық ядросында допаминдік нейротрансфузияны белсендіреді деп болжайды. Тиісінше, окситоцинді индуцированный кремний эрекциясы экстра-жасушалық емес допаминнің өсуіне ғана байланысты емес ядролардың акумбензы, сондай-ақ паравенциклы ядросында және паравенциклы ядросына енгізілген допаминді рецепторлық антагонист галоперидолмен антагонизацияланған (Melis et al., 2007). Aбірге, бұл нәтижелер паравенциклы ядродан шығатын және осы аймақтағы белсендірілген босалқыларға окситоцинді енгізу кезінде каудальды вентальдік тикеальды аймаққа шығаратын окситоцинергические нейрондардың осылайша NO-циклдық GMP сигнализациясының, бұл өз кезегінде мезолимикалық допаминергические нейронды белсендіреді (Melis және т.б., 2007, 2009a, Succu және т.б., 2008). Ядролық акумбензияларда шығарылған допамин, өз кезегінде, жұлын буынының ортаға түсіретін кремнийлі эрекциясына арналған паравентрикулярлық окситоцинергические нейрондарды ынталандыратын инкстер-гипоталамикалық допаминергиялық нейрондардың активациясына әкелетін нейрондық жолдарды белсендіреді. Сонымен қатар, паравенцикулярлық ядрода шығарылған допамин, сондай-ақ, вентральдік тецикальды аймақ, гиппокампус, амигдала және, мүмкін, басқа ми жерлері сияқты, қосымша гипоталамус миының облыстарына жобаланатын омитосинергиялық нейронды белсендіруі мүмкін.
Жоғарыда айтылғандай, бұл гипотезаға сәйкес, апоморфин пенициллятордың ядросына дозада инъекция жасайтын пениса эрекциясы ядро акументінде экстондық-жасушалық допамин концентрациясын арттырады, оттситоцина рецепторлары антагонисты d (CH2) 5Tyr (Me ) 2-Orn8-вазотоцинді вендральді ойық аймағына енгізген (Succu және т.б., 2007, Melis et al., 2009a). Жоғарыда көрсетілген нейрондық жолдарда сексуалды өнімділікке ғана емес, сексуалды ынталандыруға және жыныстық марапаттауға әсер ететін допамин, окситоцин және басқа да нейротрансмиттерлерді (мысалы, глютамин қышқылы, төменде қараңыз) қамтитын гипотетикалық нейрондық схеманы құрастыруы мүмкін (4 бөлімін қараңыз).
3.4. Гипокампустың вентральдік субикулумында окситоцин-глютаминді қышқылымен өзара әрекеттесу
Окситоцин дозаға тәуелді түрде (Melis et al., 2009b) доральді субикулада емес, венталға енгізілген кезде кремнийдің эрекциясын индуцирует. Сексуалды реакция басқа ми аймақтарында табылған окситоциннің сол аймағына берілген d (CH2) 5Tyr (Me) 2-Orn8-вазотоцинді алдын-ала инъекциялау арқылы жойылатын окситоцинді рецепторлардың ынталандыруымен байланысты (жоғарыдан қараңыз) . Бұл рецепторлардың оқшаулануына келсек, қолда бар деректер олар азот тотығы-синтезіне бай нейрондардың жасушалық органдарында орналасқандығын көрсетеді.
Тиісінше, микродиализдік зерттеулерде омитоцинді индуцированный пениса эрекциясы вентальдік субикулумдағы азотты оксидтердің ұлғаюымен қатар жүреді және бұл ұлғаю тек азот оксидисинтазы ингибиторы S-метил-тио-1-цитруллиннің ингибирлеуімен ғана емес, және гидробионды гемоглобинмен, сонымен қатар d (CH2) 5Tyr (Me) 2-Orn8-вазотоцин арқылы, барлық бірдей окситоцин аймағына penile erection (Melis et al., 2010) антагонизацияланатын дозада берілген. Ең бастысы, окситоцинді индуцированный кремний эрекциясы, сондай-ақ вентральдік субикулумда клеткалық емес глютамин қышқылының шоғырлануының ұлғаюымен қатар жүреді, бұл бәсекелес емес NMDA рецепторлы антагонисті (+) MK-801 ішінара ішінара антагонизацияланған ventral subiculum (Melis және басқалар, 2010). Сонымен бірге, бұл нәтижелер жаңадан құрылған азот оксидінің жасушааралық хабаршы ретінде әрекет етіп, глутаминді қышқылдың нейротрансмиляциясын, кремнийді эрекциялауға әкеліп соқтырады, мүмкін, вендралды эфферентті проекциялар арқылы мүмкін қосымша гиппокампалы ми бөлімдеріне арналған субикум. Осы гипотезаға сәйкес, Вентральдік субикумға енгізілген NMDA дозаға тәуелді түрде пениса эрекциясын индуцирует, және бұл нәтиже (+) MK-801 сол учаскесіне алдын-ала инъекция арқылы антагонизацияланған, бірақ S-methyl-thio-l-citrulline , гемоглобин немесе d (CH2) 5Tyr (Me) 2-Orn8-вазотоцин (Melis және басқалар, 2010). Вентальдік субикулумда қоздырғыш аминқышқылдары (яғни глутамин қышқылымен) белсендірілген қосымша гипокаппалы ми бөлімдеріне арналған нейрондық эфференттік жолдарға келетін болсақ, олардың глутамергичті болуы ықтимал, бұл гиппокампалы эфференттік проекциялардың көпшілігі. Қазіргі кезде бұл проекция меголимикалы допаминергической нейрондардың белсенділігін модуляциялайды, бұл, өз кезегінде, паравенциклы ядроның инкраф-гипоталамикалық допаминергической нейрондарының қызметін модуляциялап, омитосинергиялық нейрондардың талқыланғанындай, кремнийді құрастыру (жоғарыдан қараңыз).
Осылайша, вентальдік субикулум окситоцинмен туындаған кремний эрекциясы ядро акументінің қабықшасындағы экстра-жасушалық допамин концентрациясының ұлғаюымен бірге жүреді, бұл ретте penile erection сияқты d (CH2) 5Tyr (Me) 2- Окситоцин алдында вендральді субикумға берілген (Orn8-вазотоцин) (Melis et al., 2009b). Сондай-ақ, вентральды субикулумның окситоцинді индуцированный пениса эрекциясы, сондай-ақ, вентральдік тюкальді аймаққа енгізілген (Melis және т.б., 801b) емес, (+) MK-2009 арқылы азаяды және экстракционалды емес окситоцин алдында вентральдік субикулумға енгізілген D (CH2) 5Tyr (Me) -Orn8-вазотоцинмен жойылған жауаптар ретінде, бірақ векторлы тюкальді аймақта глутамин қышқылы, бірақ ядро accumbens емес (суретті қараңыз: 3) мезолимик допаминергической нейрондарының активтенуі вендралды тикеальды аймақта глутамераторлық нейротрансляцияның ұлғаюымен байланысты болуы мүмкін. Бұл вендралды эвигальды аймақта гинамикалық қышқыл-допаминдік өзара әрекеттесуді қарастырады. Қосымша зерттеу субикумадан вентальдік тикеальдық аймаққа дейінгі протекоэффективті эфферентті глютамергических жолдарды тікелей немесе жанама, яғни префронтальды кортекс немесе басқа ми бөлімдері арқылы (Melis және т.б., 2009b және сілтемелер қараңыз) анықтау үшін қажет. Паравентрикулярлық ядро гиппокамптан глутамергический проекцияларды алғандықтан (жоғарыда қараңыз және Saphier және Feldman, 1987) және глутамин қышқылы паравенцирулярлық окситоцинергические нейронды белсендіреді, оның ішінде вентальдік тикігальді аймаққа (Argiolas және Melis, 2005 және оның сілтемелерін қараңыз) және вентральдік тимальді аймақтағы окситоцинді кремнийді эрекциялауды тудырады және мезолимикалық допаминергиялық нейронды белсенділігін арттырадыs (жоғарыдан қараңыз), бұл p деп болжауға итермелейдіаравтеникулярлық окситоцинергические нейрондар, кем дегенде ішінара, мезолимического допаминергических нейрондық активтендіру окситоцина, веналық субикумаға енгізілген болуы мүмкін (4 бөлімін қараңыз).
4. Қорытынды ескертулер
Жоғарыда қарастырылған зерттеулер гипоталамустың және жұлынның паравенцикулярлық ядросының деңгейінде пениса эрекциясының орталық бақылауында ерлер егеуқұйрықтарында омитоциннің маңызды рөлін көрсететінін көрсетеді. Атап айтқанда, ең бастысыцент зерттеулері омитоциннің басқа ми бөлімдерінде, яғни вендралды тикеальдық аймақта, вендральді субикулумда және амигдаланың постеромедиальді кортикальды ядросында,.
Паравентрикулярлық деңгейде паравенцикулярлық ядродағы омитосинергиялық нейрондардың жасушалық органдарында D2 тобының барлық допаминді рецепторларының (D2, D3 және D4) білдіруі (және supraoptic ядрода және (Баскервил және Дуглас, 2008, Баскервил және т.б., 2009). Бұл нейроанатомиялық күшті қамтамасыз етеді допамин және допаминді рецепторлар агонисттері эректильді функциямен тікелей айналысатын және тек жұлқалыққа ғана емес, сонымен қатар гипоталамус миының аудандарында жобаланған окситоцинергические нейронды белсендіре алатын гипотезаны қолдайды. Осыған байланысты, допаминдік рецепторлардың агонист-индуцирленген инкультуралық қысымның жоғарылауы люмбакральді жұлындағы омотосинергиялық рецепторлардың қоршауымен (Baskerville et al., 2009) төмендейді. Шынында да, анестезированных ерлер егеуқұйрықтарында осындай куәліктер алынды болса да, табылған допаминдік рецепторлардың агонист-индуцированного кремний эрекциясына қатысатын паравенциклиальді омыртқа омотосинергиялық түсіру жолының белсенділігін растайды. HДоксаминергиялық жасушалардағы допаминді рецепторлардың ынталандыруымен туындаған кремнийдің эрекциясының D2 (D2, D3 немесе D4) отбасының белгілі бір допаминді рецепторлы қосалқы түрін белсендіруіне байланысты екені анықталмаған. немесе егер бұл рецепторлардың субтитрлері эректильді реакцияны модуляциялау кезінде ынтымақтасса, мүмкін, әртүрлі жолдармен penile erection орын алған жағдайларға байланысты (Moreland және т.б., 2004, Enguehard-Gueiffier et al., 2006, Melis et al. 2006a, b; Löber және т.б., 2009, Collins және т.б., 2009, Depoortère et al., 2009, Baskerville et al., 2009).
Тағы бір жаңа маңызды нәтиже: окситоцин тек паравенциклы ядросына немесе гиппокампаның CA1 өрісіне ғана емес, сондай-ақ вентальдік тикальды аймаққа, вентральдік субикулумға және америддалдың постеромедиальді кортикальды ядросына енгізген кезде кремнийді эрекциялауды тудырады. Бұл ми бөлімдері ерте егеуқұйрықтардағы омитоцинді өздігінен сексуалдық эпиляция эпизодтарын жоғарылатынын көрсеткен ерте зерттеулерде тексерілмеді, бірақ олар паравенцикулярлық ядрадан люмбакральді жұлындық окситоцинериялық проекциялар сияқты қабылданады. Окситоцин қол жетімді немесе жетпейтін рецептуралық (эстроген-прогестерон асты) овариэктомизацияланған қызыл егеуқұйрықтың бар болуынан туындайтын кез келген сексуалдық ынталандыру болмаған кезде ересек ер егеуқұйрықтарында пайда болатын, өздігінен созылатын ереуіл эпизодтарын көтере алатын шынымен табылған. паравенциклы ядросына және гиппокампустың CA1 өрісіне енгізілген кезде, бірақ доральді субикумада, бүйірлік септумда, каудиттің ядросында, медиальдық пропоптикалық аймақта, вентромидтік ядрода және супраоптикалық ядрода емес, гениталияның манипуляциясы (Melis et al. , 1986). Барлық осы зерттеулерде кремний эрекциясы, әдетте, эксперимент барысында емделушіден алынған немесе одан кейін видеокамера аппаратымен бейнекамерада жазылған экспериментті қадағалап байқаған байқаушыдан пеницис пайда болған кезде есептеледі. Әрбір penile erection эпизоды 0.5-1 минутқа созылады және әдетте пениса және / немесе қисық сызықшалармен қоса жүреді. Әдетте бұл егеуқұйрықтарда сексуалдық тәжірибенің, жасының әсерін анықтау үшін немесе осы егеуқұйрықтарды төмен немесе жоғары жауап берушілерге түрлі ми бөліктеріне енгізілген окситоциннің протеортивті әсеріне бөлуге болады. Бұл басқа да нейропептидтер мен препараттардың дозиметриялық эпизодтарын, соның ішінде допаминдік агонистерді, қоздырғыш аминқышқылдарды, ACTH-MSH, гекарелинді және ВГФ пептидтерін көбейтетін протеорганикалық әсерге қатысты зерттеулердің көпшілігіне де қатысты. Дегенмен, осы қосылыстардың протекторлы әсері телеметрия әдістерімен бірнеше рет дәлелденді, бұл өздігінен немесе осы қосылыстардың әртүрлі бағыттар бойынша енгізілгеннен кейін, яғни жүйелі түрде, интрасереброникулярлық немесе тікелей ми ядроларына тікелей қысым жасайтын микротрансформаторды имплантациялаудан кейін тікелей каверогенді корпусқа (Bernabè et al., 1999 қараңыз). Вентральдік тикальді аймақта вентальдік субикулум және амигдаланың постеромедициналық ядросы сондай-ақ омитоцин окситоцинергически рецепторларға әсер ете отырып, кремний эрекциясын индуцирует. Бұл вендралды тикеальді аймақта пайда болатын мезолимбиялық допаминергиялық нейрондардың активациясына әкеледі және ядролардың акумбензияларының қабығына шығарылады, ядролардың акумбензияларының қабығынан алынатын диализатта экстра-жасушалық допамин концентрациясының ұлғаюымен өлшенген және допаминергический рецепторлардың қоршауынан кейін табылған осы экстиптиумальдық аймақтарға енгізілген пептидпен туындаған эректильді реакцияның азаюымен ядролардың акумгендері (төменде қараңыз). Месолимикалық допаминергические нейрондарды белсендіруге және кремний эрекциясына әкелетін бұл ми аймағындағы омитосинергиялық рецепторларды ынталандыру арқылы белсендірілген тетіктерге келсек, ең жақсы түсіндірілгендер - каудальды вентральдік тецикальды аймақта кездесетіндер. Шынында да, фармакологиялық және иммунофлуоресценцияның нәтижелері оттситоцинн нервтерінің аяқтары ядролардың акумбензияларының қабығына түсетін допаминергиялық нейрондардың жасушаларының денелеріне әсер етеді (Melis et al., 2007, 2009a, Succu және т.б., 2008). Нейрондардың көбі азот оксиді синтетасына және гуанилат циклисіне бай. Осы допаминергические нейрондардың жасушаларында организмде окситоцинергические рецепторларды ынталандыру азот оксидінің синтетасын белсендіруді тудырады, бұл азот оксидінің ұлғаюына алып келеді. Nitric oxide өз кезегінде гуанилат циклді белсендіреді, осылайша циклдық GMP концентрациясын арттырады, бұл месолимбиялық допаминергические нейрондардың белсендірілуіне және ядро акументіндегі допаминнің босатылуына әкеледі, бұл дисперстатта дисфункцияның асқазан-ішек микродиализінен алынатын ядролардың акумбензасынан экстра-жасушалық допаминнің өсуімен өлшенеді (Succu және басқалар, 2008 ). Өз кезегінде, ядро акументіндегі босатылған допаминді кремнеземдеуге алып келетін нейрондық жолдарды белсендіреді. Бұл оттситоцинді индуцированный кремний эрекциясын төмендету үшін ядро акументіне енгізілген допаминді рецепторлардың антоанторы галоперидол және / немесе сиспопентиксол қабілетін қолдайды (Succu және т.б., 2008). Протекоэдилді жолдардың бірі инкрафо-гипоталамикалық допаминергиялық нейрондарды, атап айтқанда, паравенцикулярлық окситоцинергической нейрондардың жасушалық телімдерін жобалау сияқты көрінеді. Шынында, каудальды веналық тикеальды аймаққа енгізілген омитоцин тек ядро акументінде ғана емес, сондай-ақ паравенциклы ядросында да жасушадан тыс допаминді көбейтеді және паравенциклы ядросындағы допаминдік рецепторлардың қоршауында вирустық тикальды аймақты окситоцинді индуцированный кремний эрекциясы төмендейді (Succu et al. , 2007, 2008; Melis және басқалар, 2007, 2009a). Осы ядролардың акумбензының болуы допамин-паравенциклды допамин-паравенцикулярлық окситоцин-веналық тикеальдық аймақтың окситоцин-допаминдік байланыстарының болуы сондай-ақ ядро акумбензияларының қабықшасында экстракционалды допаминді арттыру үшін апоморфин мен D4 рецепторлы агонисті PD 168077-ның протеэктилді дозасын қолдану мүмкіндігі ұсынылған (Succu және т.б., 2007), жауап бұл вентральдыктикалық аймаққа берілген D (CH2) 5Tyr (Me) -Orn8-вазотоцин арқылы жойылған (Melis және басқалар, 2009a, төменде қараңыз). Дегенмен, ядролық акументаларды инкцепти-гипоталамикалық допаминергиялық жүйеге жалғайтын нейрондық жолдарды анықтау үшін әрі қарай жұмыс жасау қажет.
Қазіргі кезде тек қана омитоцинді пениса эрекцияны индуцирует және вендральдік субикулумға немесе америддаланың постеромедиальдық ядросына енгізгенде мезолимбикалық допаминергические нейронды белсендіретін механизм бар. Бұл аймақтарда окситоцин өздерінің рецепторларын белсендіреді, бұл азот оксидінің синтезін белсендіруге әкеледі, осылайша азот оксидінің өндірісін ұлғайтады. Өз кезегінде натрий оксиді белгісіз эфференттік проекцияны белсендіреді, бұл веналық триумальдық аймақта глутамергенді нейротрансмиссияны арттырады. Глутамин қышқылы содан кейін месолимбиялық допаминергические нейрондарды ынталандырады, бұл кремнийді құрастыру. Бұл гипотезаға, негізінен, вентральдік тикеальдық аймақта клеткалық глутамин қышқылын ұлғайту үшін вентральдік субикулаға енгізілген окситоцин қабілеті қолданады (сур. 3) және NMDA рецепторлық антагонисті (+) MK-801 (+) вентралды тикальды аймаққа енгізілген, бірақ ядролық акументпен емес, вентральдік субикумаға немесе постеромедициналық ядроға енгізілген амигдаланың (Melis et al., 2009b). Қазіргі уақытта вентральді субикулумға окситоцинді индукцияға арналған кремнийді монтаждау туралы толығырақ ақпарат бар. Мұнда окситоцинді индукцияланған кремний эрекциясы азот оксидінің синтезі бар нейрондардың жасушалық органдарында орналасқан окситоцинергиялық рецепторлардың белсендірілуіне қосарлы болып көрінеді. Бұл ұзақ мерзімді перспективада (Snyder, 1992, Southam және Garthwaite, 1993, Schuman және Madison, 2004, SNYX, XNUMX, SNYX, SNYX, SNYMX, . Осы гипотезаға сәйкес, окситоцинді индуцированный кремний эрекциясы вентальдік субикулумдан диализатта жасушадан тыс глютамин қышқылының ұлғаюымен және NMDA арқылы вентральдік субикулумда акуытқыш аминқышқылды рецепторлардың ынталандырылуымен қатар жүреді. Өз кезегінде глютамин қышқылы нейрондық эфференттік проекцияны белсендіреді, бұл жоғарыда айтылғанындай, вендралды тикеальдық аймақта месолимбиялық допаминергиялық нейрондардың белсендірілуіне әкеледі. Қазіргі кезде амигдаланың постеромедициналық ядросында бұл механизмдер жұмыс істейді. Оның үстіне, кейінгі зерттеулер (1) вендральдік субикулумда және амигдаланың постеромедициналық ядросында (NNUMX) нитрон оксид синтетасы бар нейрондардың жасушалық органдарында (2) локализацияланғанын (3), егер бұл нейрон азот оксидімен белсендірілген сигнализация жүйесін анықтау үшін акутикалық амин қышқылының (глутамергенді) синапстарының және (XNUMX) бақылауында. Осыған байланысты, омитоциннің каудальды веналық тикеальдық аймақта допаминергические нейрондардың жасуша органдарында азот оксид-синтаза белсенділігін және вентральдік субикулумдағы және белгісіз нейрондардың және амигдаланың постеромедиальдық ядросының жоғарылауы қабілетін еске салады. окситоцинді паравенцикулярлық ядродағы (Melis et al., 1997) оксицоцинергические нейрондардың жасушалық органдарында азот оксид-синтазасын белсендіру мүмкіндігі. Дегенмен, вентральдік тецикальдық аймақта допаминергические нейрондардың жасушалық органдарындағы азот оксидінің өндірісі өсіп, гуанилат циклді белсендіру және циклдік GMP-ні белсендіру арқылы осы нейрондардың белсендірілуіне әкеледі, бұл паравенциклы ядрода пайда болмайды. Тиісінше, паравенцикулярлық ядраға енгізілген ХНУМХ-бромо-циклдық GMP вирусты тесікшелі аймаққа енгізгенде, бұл ретте penile erection тудырмайды. Бұдан кейін азотты оксидисциплинді GMP жүйесінен басқа сигналдық жол, эндогендік және / немесе экзогендік азот оксиді (Melis and Argiolas, 1995b, Melis et al., 1997) арқылы penile erection-ға делдалданатын окситоцинергические нейронды активтендіруде паравенциклы деңгейіне қатысады (Fig . 1). Екінші жағынан, веналық тикігальді аймақта циклдік GMP мезолимбиялық допаминергиялық нейрондарды белсендіруде және ерлер егеуқұйрықтарынан алынған ядролардың акумбензиясының қабығынан алынған диализатта кездесетін экстрационды допаминнің жоғарылауында маңызды рөл атқарады егеуқұйрыққа қол жеткізе алмайтын овариэктомизацияланған рецептордың (эстроген + прогестеронмен өңделген) егеуқұйрықтары болған кезде, байланыссыз пениса эрекцияларын көрсетуге немесе көрсетпеуге арналған. Осы эксперименттік жағдайларда ересек егеуқұйрықтарда байланыссыз пениса эрекцияларын көрсеткенде, экстра-жасушалық допамин концентрациясының жоғарылауы күтілгендей болады және бұл ұлғайту, тек қарапайым түрде, каудальды вентальдік тикеальдық аймаққа берілген фосфодиэстеразы ингибиторлары арқылы (Sanna et Ә., 2009).
Окситоцинді веналық тикальды аймаққа, вентральдік субикулумға және амигдаланың постеромедиальді кортикальды ядросына енгізіп, паравенцикулярлық ядросына енгізілген допамин агонистері, пениса эрекциясын ынталандыру және мезолимикалық допаминергиялық нейрондарды белсендіру үшін кейбір түсініктемелерге лайық . Біріншіден, жоғарыда еске алынатындарға ұқсас тетіктер физиологиялық контекстте, мысалы, копуляция барысында (copula penile erections кезінде орын алған кезде) немесе контактсыз пениса эрекцияларында пайда болған кезде жедел болуы мүмкін. Бұл эрекциялар феромонмен басқарылатын пениса эрекциялары сексуалдық күшті ерлер егеуқұйрықтары қол жетімсіз рецептивтік (овариэктомизацияланған эстроген + прогестеронмен асқынған) егеуқұйрыққа енгізілген және жыныстық индексі ретінде қарастырылатын есірткі немесе окситоцин тудыратын заттардан айырмашылығы жоқ (Sachs, 1997, 2007). Шынында да, бұл нәтижелер осы аудандардағы окситоциннің физиологиялық контексте немесе дәрілік заттардың қолданылуынан кейін пайда болған жыныстық эрекцияда рөл атқаратындығын көрсетпесе де, олар мидың аудандары орталыққа тиесілі окситоцинге тиесілі болатын жерлерге жататындығын көрсетеді протекционалды препараттардан кейін ғана байқалатын риясыз эпиляция эпизодтары ғана емес, сондай-ақ ерлердің (және әйелдердің) жыныстық мінез-құлқын жақсартады (Argiolas және Melis, 2004 және оның сілтемелерін қараңыз). Тиісінше, окситоцин концентрациясы еркек егеуқұйрықтардың гиппокампасында апоморфиннің классикалық допаминдік агонисті (Melis және басқалар, 1990) және d (CH2) 5Tyr (Me) -Orn8-вазотоцинді емдеген препараттармен емделген, ол окситоцинді рецепторлар тек қана мысуляциялық мінез-құлықты (Argiolas et al., 1987a) ғана емес, сондай-ақ жыныстық күшті ерлер егеуқұйрықтарында ерлердің мопуляциялық мінез-құлықта (Argiolas et al., 1987b) апоморфиннің фасилитаторлық әсері өте тиімді болып табылады. орын алады. d (CH2) 5Tyr (Me) -Orn8-вазотоцин-ақ, жыныстық күшті ерлер егеуқұйрықтарында нонграмма көлемінде берілетін кезде, PVN-ке емес (Melis et al., 1999a) емес, байланыссыз эрекцияларды азайтуға өте күшті.
Екіншіден, мезолимбиялық допаминергиялық нейрон тамақ, су және жыныстық белсенділік сияқты табиғи күшейтетін ынталандырулардың мотивациялық және пайдалы қасиеттерінде маңызды рөл атқарады (Fibiger and Phillips, 1988, Wise and Rompre, 1989, Everitt, 1990). Атап айтқанда, осы нейрондардан босатылған допаминді табиғи ынталандырудың мотивациялық аспектілерін мақсатты бағытталған мінез-құлықтарға ауыстыру, мысалы, сексуалдық белсенділік, жыныстық қатынасқа ұмтылу және жыныстық қатынасқа түсу үшін марапаттауға және қанағаттандыруға көмектеседі делінеді (Goto және Grace, 2005). Тиісінше, қол жетпейтін овариэктомизацияланған эстроген + прогестеронмен асқынған рецептуралық қызыл егеуқұйрыққа әсер ету кезінде жыныстық күшті ерлер егеуқұйрықтарының ядросы акумбензиясынан диссизатта экстрацеллюлозалық допамин концентрациясының жоғарылауы байқалады және еркек егеуқұйрықтың қабылдау әйеліне (Pfaus және Everitt, 1995).
Үшіншіден, осы нәтижелер неврологиялық тізбектің веналық тикальды аймақпен тікелей немесе жанама (вентральдік субикулум және / немесе амегдаланың постеромедиальді кортикальды ядросы арқылы) және ядро акументінің көмегімен және осыдан кейін белгісіз омыртқа буынының ортаға келтіретін пениса эрекциясы мен окситоцинергические нейрондарды, вентральдік тецикальды аймаққа, вентральдік субикулумға және америддальдың постеромедиальді кортикальды ядросына жобаланатын окситоцинергические нейрондардың белсенділігін бақылау үшін паравенцикулярлық ядроға қайтадан жолдары бар. мезолимикалық допаминергиялық нейрондардың белсенділігі (сурет 4). Бұл күрделі нерв тізбегі физиологиялық контексте ер жыныстық мінез-құлықтың тұтынушылық (эректильді-эякуляциялық) және алдын-ала (мотивациялық және марапаттайтын) аспектілерін бақылауға қатысатын нейрондық әрекеттерді интеграциялауда маңызды рөл атқаруы мүмкін. Шынында да, қол жетпейтін рецепторлық егеуқұйрыққа әсер ету кезінде, байланыссыз эрекциялар пайда болған кезде ядролардың акумбиндерінде (Pfaus and Everitt, 1995) және жыныстық күшті еркек егеуқұйрықтардың паравенцикулярлық ядросында қосымша хелелік допамин көбейтіледі, мысалға, копуляцияның пениса эрекциялары болғанда (Melis et al., 2003). Осылайша, кейінгі зерттеулер веналық tegmental аймағында эндогендік окситоценнің рөлін нақтылау үшін қажет болса да, вентальдік субикулум мен байланыссыз эрекциялар мен жыныстық мінез-құлық кезінде амигдала болса, онда бұл гипотетикалық нейрондық тізбектің жыныстық мінез-құлықтың тұтынушылық аспектілері, сондай-ақ жыныстық белсенділіктің (Everitt, 1990, Pfaus and Everitt, 1995) марапаттайтын қасиеттерін түсіндіру үшін нейролярлық субстрат беретін мезолимикалық допаминергиялық жүйені белсендіре алады. Осыған орай, вентральдік тикеальды аймаққа енгізілген окситоцинмен белсендірілген мезолимикальді допаминергиялық жүйе опиат, каннабиноидтер, амфетамин, кокаин және алкоголь (Tanda және т.б., 1997) сияқты теріс препараттармен белсендірілген. кофе, морфин, алкоголь және каннабиноидтерге төзімділік пен тәуелділікті төмендетуге қабілетті деп табылған (Kovacs және басқалар, 1998, Cui және т.б., 2001). Қорытындыда, окситоцин тек вендралды тикальді аймақта ғана емес, сонымен қатар вентальдік субикулумда және амигдаланың постеромедиальді кортикальды ядросында сексотерапевтикалық допаминергические нейронды белсенді ете алады, бұл жыныстық белсенділіктің тәбетті және әсерлі әсеріне қатыса алады . Мезолимикалық допаминергические нейрондарды белсендіру мезолимик допаминергические нейрондарының жасушалық органдарында окситоцинергические рецепторлар арқылы немесе тікелей вендральді трицальді аймақта глютамин қышқылының нейротрансмиссиясын жандандыру арқылы жанама болуы мүмкін.
Ядролық акпломент қабықшасында шығарылған допамин, өз кезегінде, паравенцикулярлық ядродағы инсепто-гипоталамикалық допаминергиялық нейрондардың белсенділігін модуляциялайды немесе жыныстық эрекцияны (жұлын миына шығаратын омитосинергические нейрондарды белсендіру арқылы) немесе жыныстық ынталандыру мен марапаттауды (окситоцинериялық неврондар вендральді тикеальды аймаққа, вентральдік субикулумға немесе амигдаланың постеромедиальді кортикальды ядросына) әсер етеді. Себебі допамин физиологиялық контексте (мысалы, байланыссыз эрекциялар мен сопуляция) пайда болған кезде ядролардың акумбензиялық қабығында және паравенциклы ядросында босатылғаннан кейін (Succu және т.б., 2007, Melis et al., 2003, 2007) орталық окситоцинергических нейрондық мезолимик пен ингтотерипотальды допаминергиялық жүйенің өзара әрекеттесуін қамтамасыз ететін нейрондық тізбектерге қатысуы мүмкін. Бұл нейрондық тізбектер сексуалды мінез-құлықтың тұтыну кезеңінде ғана емес, сонымен қатар сексуалдық ынталандыру, жыныстық қатынас пен сексуалдық сыйақыларда да рөл атқаруы мүмкін.
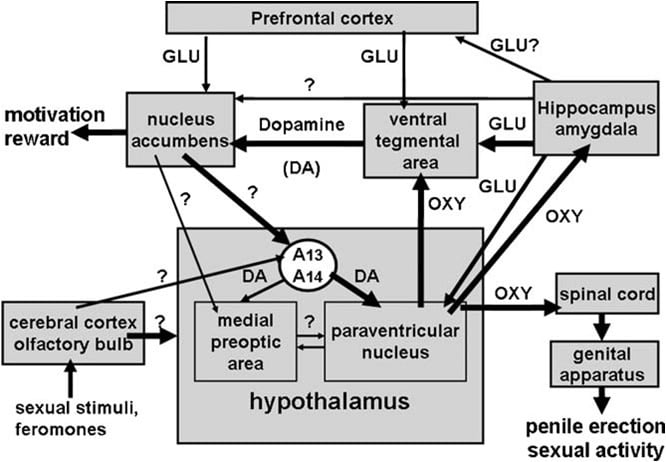
Осы тараудың және алдыңғы есептердің нәтижелері бойынша ұсынылған жыныстық ынталандыруға, марапаттауға және жыныстық әсерге әсер ететін окситоцинді қамтитын гипотетикалық нейрондық схеманың схемалық көрінісі. Паравенцикулярлық ядрода пайда болатын және допаминмен және глютамин қышқылымен (сонымен бірге басқа нейротрансмиттерлермен және / немесе нейропептидтермен) белсендірілгенде, жұлынның жұлынына шығаратын окситоцинергические нейрондар омыртқа жұлынына жобаланатын окситоцинериялық нейрондарды белсендіру арқылы жыныстық эрекция мен сексуалдық өнімділікті жеңілдетеді. Паравенцикулярлық ядродағы допамин және глутамин қышқылы (сонымен қатар невротрансмиттер және нейропептидтер) сонымен бірге вентральдік тецикальды аймаққа жобаланатын окситоцинергические нейронды белсендіреді, осылайша мазолимикалық допаминергические нейрондарды белсендіріп, ядролық акументлерге әсер етіп, жыныстық ынталандыруды және марапаттауды модуляциялайды. Ядролық акументпен (NAs) шығарылған допамин, өз кезегінде, әлі де белгісіз нейрондық жолдарды белсенді етеді, бұл инкраф-гипоталамикалық допаминергиялық нейрондардың белсенділігін арттырады (A13-A14 Dahlstrom және Fuxe топтарынан шыққан), окситоцинерлік нейронды Жұлынның қабынуына әкелетін жұлын. Бұл схеманы тек қана каудальды вентральдік тецикальды аймаққа ғана емес, сонымен қатар вентральдік субикулумда және агордадада, сонымен қатар, паравенцикулярлық ядроның окситоцинергической инерациясын алуға болатын, мүмкін, тікелей немесе жанама глутамергичтический эффенент арқылы вирустық жыныстық мотивацияның және жыныстық эрекцияның модуляциясына әкеліп соғады. Ақыр соңында, жоғарыда көрсетілген схеманы жыныстық ынталандыру және феромондар да белсендіре алады, өйткені феромон-делдалсыз байланыссыз эрекциялар мен копуляция кезінде паравенцикуляр ядросында (және медиальдық пропоптикалық аймақта) қосымша клеткалық допамин мен глютамин қышқылдары өседі (тиісті сілтемелер Әдебиеттер тізімі).
Ынтымақтастық Бұл жұмысты итальяндық министрліктің грантымен және AA және MRM зерттеулеріне ішінара қолдау көрсетілді
Әдебиеттер тізімі
Andersson, KE, 2001. Қабілетті эрекцияның фармакологиясы. Фармакол. Rev. 53, 417-450. Аргиолас, А., ХНУМХ. Азот оксиді кремнийдің эрекциясының орталық делдалы болып табылады. Нейрофармакология 33, 1339-1344. Аргиолас, А., ХНУМХ. Нейропептидтер және жыныстық мінез-құлық. Нейроци. Биобехав. Rev. 23, 1127-1142. Argiolas, A., Gessa, GL, 1991. Окситоциннің негізгі функциялары. Нейроци. Биобехав. Rev. 15, 217-231. Аргиолас, А., Мелис, МР, ХНУМХ. Қабілетті эрекцияның нейромодуляциясы: нейротрансмиттерлер мен нейропептидтердің рөлін шолу. Prog. Нейробиол. 47, 235-255. Аргиолас, А., Мелис, МР, ХНУМХ. Еркек сүтқоректілердің жыныстық мінез-құлқындағы омитоцин мен паравенциклы ядросының рөлі. Физиол. Behav. 83, 309-317. Аргиолас, А., Мелис, МР, ХНУМХ. Пенсияны эрекцияның орталық бақылауы: гипоталамустың паравенцикулярлық ядросының рөлі. Prog. Нейробиол. 76, 1-21. Аргиолас, А., Коллу, М., Гесса, Г.Л., Мелис, М.Р., Серра, Г., ХНУМХ. Окситоцинді антагонист d (CH2) 5Tyr (Me) -Orn8-vasotocin егеуқұйрықтарда ерлердің копуляциялық мінез-құлқына қарсы болады. E. J. Фармакол. 149, 389-392. Argiolas, A., Collu, M., D'Aquila, P., Gessa, GL, Melis, MR, Serra, G., 1989. Ерлерге арналған мопуляциялық мінез-құлықтың апоморфинді ынталандыру егеуқұйрықтарында окситоцинді антагонист d (CH2) 5Tyr (Me) -Orn8-вазотоцин арқылы алдын алады. Фармакол. Биохим. Behav. 33, 81-83. Аргиолас, А., Мелис, М.Р., Гесса, Г.Л., ХНУМХ. Intraventricular oxytocin егеуқұйрықтарды итеріп кетуді және кремнийді эрекциялауды тудырады. E. J. Фармакол. 117, 395-396. Аргиолас, А., Мелис, М.Р., Гесса, Г.Л., ХНУМХ. Окситоцин: ерлер егеуқұйрықтарында пении эрекциясының өте күшті индуцирует және итеруі. E. J. Фармакол. 130, 265-272. Аргиолас, А., Мелис, М.Р., Маури, А., Гесса, Г.Л., ХНУМХа. Паравентрикулярлық ядроның зақымдануы егеуқұйрықта ACTH емес, апоморфин мен окситоцинмен туындаған ентігу мен кремнийді эрекцияға жол бермейді. Brain Res. 421, 349-352. Аргиолас, А., Мелис, М.Р., Варгиу, Л., Маури, А., Гесса, Г.Л., ХНУМХб. D (CH2) 5Tyr (Me) -Orn8-вазотоцин, қуатты окситоцин антагонисті, ACTH 1-24 арқылы емес, окситоцин және апоморфинмен индуцирленген пениса эрекциясын және итеруі. E. J. Фармакол. 134, 221-224. Аргиолас, А., Мелис, М.Р., Стэнкппанио, Р., Гесса, Г.Л., ХНУМХ. _-Конотоксин еркек егеуқұйрықтарында апоморфинді және окситоцинді индуцированный пениса эрекциясын және итеруін болдырмайды. Фармакол. Биохим. Behav. 37, 253-257. Арлети, Р., Бертолини, А., ХНУМХ. Окситоцин әйел егеуқұйрықтарында лордоздық әрекетті ынталандырады. Нейропептидтер 6, 247-255. Арлети, Р., Баззани, С., Кастелли, М., Бертолини, А., ХНУМХ. Окситоцин егеуқұйрықта ерлердің копуляциялық көрсеткіштерін жақсартады. Хорм. Behav. 19, 14-20. Арлети, Р., Бенелли, А., Бертолини, А., ХНУМХ. Ересек ересек егеуқұйрықтардың жыныстық мінез-құлқы окситоцинмен ынталандырылады. E. J. Фармакол. 179, 377-382. Арлети, Р., Калза, Л., Джардино, Л., Бенелли, А., Кавзазутти, Е., Бертолини, А., ХНУМХ. Сексуалдық әлсіреуі омитоцин өндірісінің азаюымен және гипоталамустың паравенцикулярлық ядросында опиоидты пептидтердің өндірісін ұлғайтумен байланысты. Нейроци. Lett. 233, 65-68. Bancila, M., Giuliano, F., Rampin, O., Mailly, P., Brisorgueil, MJ, Clas, A., Verge, D., 2002. Гипоталамустың паравенциклы ядросынан парагигантоцеллюлярдың ядросы босатылған серотонинергические нейрондарға тікелей проекциясы үшін егеуқұйрықтарды эрекциялауға бақылау жасайтын дәлел. E. J. Нейроци. 16, 1240-1249. Burnett, AL, Лоуенштейн, CJ, Bredt, DS, Chang, TSK, Snyder, SH, 1992. Азот оксиді: кремнийдің эритмесінің физиологиялық медиаторы. Ғылым 257, 401-403. Baskerville, TA, Douglas, AJ, 2008. Сексуалды мінез-құлықты бақылауда допамин мен окситоцин арасындағы өзара әрекеттесу. Prog. Brain Res. 170, 277-289. Баскервилл, Т.А., Аллард, Дж., Уэйман, С., Дуглас, А.Ж., ХНУМХ. Қабілетті эрекцияда допамин-окситоцин өзара әрекеттесуі. E. J. Нейроци. 30, 2151-2164. Бенелли, А., Бертолини, А., Поггиоли, Р., Кавзазутти, Е., Калза, Л., Джардино, Л., Арлети, Р., ХНУМХ. Азот оксиді егеуқұйрықтардың жыныстық мінез-құлқына қатысады. E. J. Фармакол. 294, 505-510. Bernabè, J., Rampin, O., Sachs, BD, Giuliano, F., 1999. Егеуқұйрықтарды монтаждау кезінде интракаверналық қысым: телеметриялық жазуға негізделген интегративтік тәсіл. Am. J. Физиол. 276, R441-R449. Битнер, Р.С., Никкель, А.Л., Отте, С., Мартино, Б., Барлоу, Е.Х., Бхития, П., Стюарт, А.О., Бриони, ДжД, Декер, М.В., Морланд, РБ, ХНУМХ. Егеуқұйрықтарды паравенцикулярлық гипоталамус ядросындағы Dopamine D4 рецепторлық сигналы: дереу ерте гендік индукция және митоген белсендірілген протеин киназ фосфорлануын қамтитын табиғи байланыстың дәлелі. Нейрофармакология 50, 521-531. Brioni, JD, Moreland, RB, Cowart, M., Hsieh, GC, Stewart, AO, Hedlund, P., Donnelly-Roberts, DL, Nakane, M., Lynch 3rd., J., Kolasa, T., Polakowski , JS, Osinski, MA, Marsh, K., Andersson, KE, Sullivan, JP, 2004. ABT-4 арқылы допамин D724 рецепторларын активтендіру егеуқұйрықтарда penile erection индуцирует. Proc. Natl. Акад. Ғылыми жұмыс. АҚШ 101, 6758-6763. Buijs, RM, 1978. Егеуқұйрықта интро- және қосымша гипоталамус вазопрессин және окситоцин жолдары. Ұяшық тінінің суреті. 192, 423-435. Buijs, RM, Geffard, M., Бассейн, CW, Hoorneman, EMD, 1984. Супраоптикалық және паравенциклды ядролардың допаминергический иннервациясы. Жеңіл және электрондық микроскопиялық зерттеу. Brain Res. 323, 65-72. Caldwell, JD, Prange, AJ, Pedersen, CA, 1986. Окситоцин эстрогенмен өңделген егеуқұйрықтардың сексуалдық рецептілігін жеңілдетеді. Нейропептидтер 7, 175-189. Cameron, JL, Pomerantz, SM, Layden, LM, Amico, JA, 1992. Апоморфин және D2 рецепторлы агонисті арқылы ерлер мен әйел маймылдардың плазмадағы омитоцен концентрациясының допаминергиялық ынталандыруы. J. Clin. Эндокринол. Metab. 75, 855-860. Canteras, NS, Simerly, RB, Swanson, LW, 1995. Амигдаланың ортаңғы ядросынан проекцияны ұйымдастыру: егеуқұйрықта фалдың зерттеуі. J. Комп. Нейрол. 360, 213-245. Carmichael, MS, Humbert, R., Dixen, J., Palmisano, G., Greeleaf, W., Davidson, JM, 1987. Адамның жыныстық реакциясында плазмалық омитоцин көбеюде. J. Clin. Эндокринол. Metab. 64, 27-31. Картер, CS, 1992. Окситоцин және жыныстық мінез-құлық. Нейроци. Биобехав. Rev. 16, 131-144. Картер, CS, Lederhendler, II, Kirkpatrick, B., 1997. Интерактивті невробиология, Нью-Йорк ғылым академиясының жылнамасы, том. 807. Нью-Йорк ғылымдар академиясы, Нью-Йорк. К Castelli, MP, Piras, AP, Melis, T., Succu, S., Sanna, F., Melis, MR, Collu, S., Ennas, MG, Diaz, G., Mackie, K., Argiolas, A. , 2007. Паравенциклов ядросындағы каннабиноид CB1 рецепторлары және пениса эрекциясының орталық бақылауы: иммуноцитохимия, аутранадография және мінез-құлықты зерттеу. Нейроход 147, 197-206. Chen, KK, Chang, LS, 2003. Эволюциялық аминқышқыл рецепторлардың агонисттерін егеуқұйрықтардағы гипоталамустың паравенцикулярлық ядросына енгізгеннен кейін кремнийдің эрекциясына әсері. J. Urol. 62, 575-580. Chen, KK, Chan, JYH, Chang, LS, Chen, MT, Chang, SHH, 1992. Егеуқұйрықта гиппокамптың пайда болуын белсендендіргеннен кейін кремнийді эрекциялауды жою. Нейроци. Lett. 141, 218-222. Chen, KK, Chan, JYH, Chang, LS, 1999. Егеуқұйрықта жыныстық эрекцияның орталық реттеуіндегі гипоталамустың паравенциклы ядросында допаминергическая нейротрансмиссия. J. Urol. 162, 237-242. Collins, GT, Truccone, A., Haci-Abdi, F., Newman, AH, Grundt, P., Rice, KC, күйеулер, SM, Greedy, BM, Enguehard-Gueiffer, C., Gueiffer, A., Chen , J., Wang, S., Katz, JL, Grandy, DK, Sunahara, ҚР, Woods, JH, 2009. Dopamine D2 ұқсас агонистердің эректік әсерлері егеуқұйрықтар мен тышқандарда D3 рецепторларымен жүргізіледі. J. Фармакол. Exp. Ther. 329, 210-217. Coolen, LM, Allard, J., Truitt, WA, McKenna, KE, 2004. Эякуляцияны орталықтандыру. Физиол. Behav. 83, 203-215. Cui, SS, Bowen, RC, Gu, GB, Hannesson, BK, Yu, PH, Zhang, X., 2001. Каннабиноидті шығару синдромын литиймен болдырмау: окситоцинериялық нейрондық активтендіруді тарту. J. Нейроци. 21, 9867-9876. Dahlström, A., Fuxe, K., 1964. Орталық жүйке жүйесінде моноаминді нейрондардың болуы туралы дәлел. I. Ми жүйелері нейрондарының жасуша органдарындағы моноаминдерді көрсету. Acta Physiol. Scand. 62 (Қосымша 232), 1-54. Депортере, Р., Бардин, Л., Родригес, М., Абриал, Э., Алиаа, М., Ньюман-Танкрид, А., ХНУМХ. Егеуқұйрықтардағы допамин D2-тәрізді рецепторлы агонистермен индуцирленген пениса эрекция және итеру: D2 допаминінің әсер етуі және қосылуы, бірақ D3 және D4 рецепторлары емес. Behav. Фармакол. 20, 303-311. Домс, Г., Хайнрихс, М., Бюль, С., Браус, Д.Ф., Герперц, СХ, ХНУМХ. Окситоцин валенттілігіне қарамастан, эмигранттардың эмигрантқа жауаптарын жоғарылатады. Биол. Психиатрия 62, 11871190. Dominguez, J., Riolo, JV, Xu, Z., Hull, ME, 2001. Допаминді баспалау және медиальдық пропоптикалық допаминді шығарудың медиальдық амигдаласының ережесі. J. Нейроци. 21, 349-355. Дональдсон, З.Р., Янг, Л.Ж., ХНУМХ. Окситоцин, вазопрессин және қоғамның нейрогенетикасы. Ғылым 322, 900-904. Eaton, RG, Markowski, VF, Lumley, LA, Томпсон, JT, Moses, J., Hull, EM, 1991. Паравенциклов ядросындағы D2 рецепторлары ерлер егеуқұйрықтарында жыныстық реакцияларды реттейді. Фармакол. Биохим. Behav. 39, 177-181. Ebner, K., Bosch, OJ, Krömer, SA, Singewald, N., Neumann, ID, 2005. Аминқышқылдағы омитоцинді босаңсытып, орталық амигдаал стресспен күресу және қоздырғыш аминқышқылдарды босатуды реттейді. Нейропсихофармакология 30, 223-230. Энгюхард-Гюйфье, С., Хубнер, Х., Эль Хакмауи, А., Аллуши, Х., Гмайнер, П., Аргиолас, А., Мелис, М.Р., Гьюифье, А., ХНУМХ. 2 - таңдамалы D4-лигандары ретінде [(1-Fenylpiperazin-4-іл) метил] imidazo (di) azines. Күшті және таңдаулы D2 агонисті 4- [2- (1-methoxyphenyl) piperazin-1,2-ylmethyl] imidazo [3-a] пиридин (PIP4EA) арқылы кремнийді эрекциялауды индукциялау. J. Мед. Хим. 49, 3938-3947. Everitt, BJ, 1990. Жыныстық ынталандыру: ерлер егеуқұйрықтарының тәбет және копуляциялық реакцияларына негізделген механизмдердің нейрондық және мінез-құлықты талдау. Нейроци. Биобехав. Rev. 14, 217-232. Fibiger, HC, Phillips, AG, 1988. Месocorticolimbic dopamine жүйесі және сыйақы. Энн. N. Y. Акад. Ғылыми жұмыс. 5, 206-215. Француз, SJ, Totterdell, S., 2003. Жеке ядро акулалар-проекциялы нейрондар егеуқұйрықтарда басоларлы амигдала және вентальды субикулярлық афференттерді алады. Нейроход 119, 19-31. 954 MR Мелис, А. Argiolas / Neuroscience and Biobehaviorviews 35 (2011) 939-955 Freund-Mercier, MJ, Richard, P., 1981. Сүт эжекциясының рефлексіндегі итудегі окситоцинді интривентрикулярлық инъекциялардың қозғаушы әсері. Нейроци. Lett. 23, 193-198. Freund-Mercier, MJ, Richard, P., 1984. Емшек сіңірген кезде окситоцин арқылы окситоцинді нейрондарды фосультативті бақылауға арналған электрофизиологиялық дәлел. J. Физиол. (Лондон.) 352, 447-466. Freund-Mercier, MJ, Stoeckel, ME, 1995. Окситоцинді нейрондардағы соматодендритті авторецепторлар. В: Ivell, R., Russel, JA (Eds.), Окситоцин, Медицина мен зерттеулердегі жасушалық және молекулярлық тәсілдер. Adv. Exp. Мед. Biol., 365. Plenum Press, Нью-Йорк және Лондон, pp. 185-194. Freund-Mercier, MJ, Stoeckel, ME, Palacios, JM, Pazos, JM, Richard, PH, Porte, A., 1987. Автобензинмен зерттелген Wistar рентген миының құрамында 3H окситоцинді байланыстыратын учаскелердің фармакологиялық сипаттамасы және анатомиялық таралуы. Нейроход 20, 599-614. Джулиано, Ф., Рампин, О., ХНУМХ. Пенсияны монтаждаудың орталық бақылауы. Нейроци. Биобехав. Rev. 24, 517-533. Giuliano, F., Allard, J., 2001. Допамин және жыныстық функция. Int. J. Этсіздік Рез. 13 (Қосымша 3), 18-28. Джулиано, Ф., Рампин, О., ХНУМХ. Монтаждаудың нервтік бақылауы. Физиол. Behav. 83, 189-201. Джулиано, Ф., Бернабье, Дж., МакКенна, К.Э., Лонгувилль, Ф., Рампин, О., ХНУМХ. Анестетирленген егеуқұйрықтардағы омитоциннің омыртқа прередилді әсері. Am J. Физиол. Регул. Біріктіру. Комп. Физиол. 280, R1870-R1877. Goto, Y., Grace, AA, 2005. Мақсатты мінез-құлықта ядролардың акумбензияларының лимбикалық және кортикальды қоздырғыштарын допаминергиялық модуляциялау. Нат. Нейроци. 8, 805-812. Hawthorn, J., Ang, VT, Jenkins, JS, 1985. Сүйек миында және жұлындағы вазопрессин мен окситоцинге арналған гиправаламиялық, паравенцикулярлық, supraoptic және suprachiasmatic ядролардағы зақымдардың әсері. Brain Res. 346, 51-57. Хиер, Р.Ф., Долак, Л.А., Дункан, Дж.Н., Гислоп, Д.К., Липтон, М.Ф., Мартин, Л.Ж., Морагис, М.А., Пирси, М.Ф., Николс, Н.Ф., Шреур, П.Ж., Смит, М.В., Ай, М.В., ХНУМХ. (R) -5,6-dihydro-N, N-dimethyl-4H-imidazo [4,5,1-ij] кинолин-5-амині) және оның метаболиттерінің синтезі және биологиялық белсенділігі. J. Мед. Хим. 40, 639-646. Hsieh, GC, Hollingsworth, PR, Martino, B., Chang, R., Terranova, MA, O'Neill, AB, Lynch, JJ, Moreland, RB, Donnelly-Roberts, DL, Kolasa, T., Mikusa, JP , McVey, JM, Marsh, KC, Sallivan, JP, Brioni, JD, 2004. Санада егеуқұйрықтарда penile erection реттейтін орталық механизмдер: апоморфиннің пререкрилді әсері бар допаминергиялық жүйелер. J. Фармакол. Exp. Ther. 308, 330-338. Huang, PL, Dawson, TM, Bredt, DS, Snyder, SH, Fishman, MC, 1993. Нейрональды азот оксиді синтаза генінің мақсатты бұзылуы. Ұяшық 75, 1273-1286. Huber, D., Veinante, P., Stoop, R., 2005. Васопрессин және омитоцин орталық амигдалада ерекше нейрондық популяцияларды қоздырады. Ғылым 308, 245-248. Галл, Э.М., Уорнер, РК, Баззетт, Т.Д., Итон, Т.Р., Томпсон, ДжТ, ХНУМХ. Медициналық пропоптикалық аймақта D2 / D1 коэффициенті ер егеуқұйрықтардың санын көбейтуге әсер етеді. J. Фармакол. Exp. Ther. 251, 422-427. Галл, Э.М., Ду, Дж., Лоррейн, Д.С., Матушевич, Л., ХНУМХ. Медициналық пропоптикалық аймақтағы экстрацитологиялық допамин: жыныстық ынталандыру және копуляцияның гормоналды бақылауының салдары. J. Нейроци. 15, 7465-7471. Hull, EM, Meisel, RL, Sachs, BD, 2002. Ер жыныстық мінез-құлық. В: Pfaff, DW, Arnold, AP, Etgen, AM, Fahrbach, SE, Rubin, RT (Eds.), Гормондар, ми және мінез-құлық. Academic Press, New York, pp. 3-137. Хурлеман, Р., Патин, А., Онур, О.А., Коэн, МХ, Баумгартнер, Т., Меццер, С., Дзобек, И., Галлинат, Дж., Вагнер, М., Майер, У., Кендрик, KM, 2010. Окситоцин адамдарда амигдалға тәуелді, әлеуметтік тұрғыдан күшейтілген оқытуды және эмоционалды эмпатияны жақсартады. J. Нейроци. 30, 4999-5007. Ivell, R., Russel, JA, 1995. Окситоцин: медицинада және зерттеуде жасушалық және молекулярлық тәсілдер. Эксперименталды медицинада және биологиядағы жетістіктер, том. 395. Пленум Пресс, Нью-Йорк. Келли, AE, Домесик, VB, 1982. Гипокампалық түзілуден цианға ядро акументіне дейінгі проекцияны бөлу: антрограда- және ретроград-горсорадистік пероксидаза зерттеу. Нейроход 7, 2321-2335. Кимура, Й., Найтоу, Ю., Ванибучи, Ф., Ямагучи, Т., ХНУМХ. 5-HT (2C) рецепторларын белсендіру - бұл егеуқұйрықтардағы апоморфин, окситоцин және меланотан-II преериктикалық әсерлері туралы жалпы механизм. E. J. Фармакол. 589, 157-162. Кондо, Й., Сакс, Б.Д., Сакума, Ю., ХНУМХ. Ересек әйелдердің қашықтағы ынталандыруларымен туындаған егеуқұйрықтарды жою кезінде медиальдық амигдаланың маңыздылығы. Behav. Brain Res. 91, 215-222. Kovacs, GL, Sarnyai, Z., Szabo, G., 1998. Окситоцин және тәуелділік: шолу. Психоневроэндокринологиясы 23, 945-962. Lee, HJ, Macbeth, AH, Pagani, JH, Scott Young 3rd, W., 2009. Окситоцин: өмірдің керемет ұйымдастырушысы. Prog. Нейробиол. 88, 127-151. Lindvall, O., Bjorklund, A., Skagerberg, G., 1984. Дипаминдік және теленфеналдағы допаминдік жүйелердің таңдаулы демохимиялық демонстрациясы: гипоталамус нейросекреторлы ядролардың допаминергиялық иннервациясы үшін жаңа деректер. Brain Res. 306, 19-30. Liu, YC, Salamone, JD, Sachs, BD, 1997. Ерлер егеуқұйрықтарында гипоталамустың паравенцикулярлық ядросының зақымдануынан кейін жыныстық жауап беру бұзылған. Behav. Нейроци. 111, 1361-1367. Лёбер, С., Цхаммер, Н., Хубнер, Х., Мелис, М.Р., Аргиолас, А., Гмайнер, П., ХНУМХ. Азулендік негізі - жаңа биоизостер ретінде: күшті допамин D4 рецепторлық лигандтарды жобалау, криминалді эрекциялау. Хим. Мед. Хим. 4, 325-328. McCleskey, EW, Fox, AP, Feldman, DH, Cruz, LJ, Olivera, BM, Циен, RW, Yoshikami, D., 1987. _-Конотоксин: Нейрондарда, бірақ бұлшық етпейтін кальций арналарының нақты және тұрақты іркілістері. Prot. Нат. Акад. Ғылыми жұмыс. АҚШ 84, 4327-4331. McKenna, KE, 2000. Орталық эволюциялық жүйені пенитенциацияны бақылауды ұйымдастыру туралы кейбір ұсыныстар. Нейроци. Биобехав. Rev. 24, 535-540. Marson, L., McKenna, KE, 1992. Омыртқаның жыныстық рефлекстерінің төмендеуі кезінде 5-гидрокситриптаминнің рөлі. Exp. Brain Res. 88, 313-318. Marson, L., McKenna, KE, 1996. CNS жасушалық топтары ischiocavernosus және bulbospongiosus бұлшықеттерін бақылауға қатысады: псевдорорлық вирустың көмегімен транснеуроналды іздестіру жұмыстары. J. Комп. Нейрол. 374, 161-179. Meisel, RL, Sachs, BD, 1994. Еркек жыныстық мінез-құлықтың физиологиясы. В: Кнобил, Э., Нил, Дж. (Eds.), Көшіру физиологиясы, том. 2, екінші басылым. Raven Press, New York, pp. 3-96. Melin, P., Kihlstrom, JE, 1963. Еркек қояндарында жыныстық мінез-құлыққа окситоциннің әсері. 73, 433-435 эндокринологиясы. Melis, MR, Argiolas, A., 1995a. Допамин және жыныстық мінез-құлық. Нейроци. Биобехав. Rev. 19, 19-38. Melis, MR, Argiolas, A., 1995b. Натрий оксидінің донорлары орталық егеуқұйрықтарды ерлер егеуқұйрықтарына енгізгенде пениса эрекциялауды және итеруді тудырады. E. J. Фармакол. 294, 1-9. Melis, MR, Argiolas, A., 2003. Орталық окситоцингиялық нейротрансляция: психогенді эректильді дисфункцияны емдеу үшін дәрілік зат. Curr. Дәрі-дәрмектер 4, 55-66 мақсаттарын белгілейді. Melis, MR, Argiolas, A., Gessa, GL, 1986. Окситоцинді индуцированная ионизации және кремний эрекция: мидағы іс-әрекет орны. Brain Res. 398, 259-265. Melis, MR, Argiolas, A., Gessa, GL, 1987. Апоморфинді индуцируемі және итеруі: мидағы іс-әрекеттер орны. Brain Res. 415, 98-104. Melis, MR, Argiolas, A., Gessa, GL, 1989a. Апоморфин егеуқұйрықта плазмалық омитоцин деңгейін арттырады. Нейроци. Lett. 98, 351-355. Melis, MR, Argiolas, A., Gessa, GL, 1989b. Апоморфин орталық жүйке жүйесінде окситоцинді босату арқылы пениса эрекциясын және итеруін тудырады. E. J. Phamacol. 164, 565-570. Melis, MR, Argiolas, A., Stancampiano, R., Gessa, GL, 1990. Апоморфиннің әртүрлі ми аймақтары мен ерлер егеуқұйрықтарының плазмасындағы окситоцин концентрациясына әсері. E. J. Фармакол. 182, 101-107. Мелис, М.Р., Маури, А., Аргиолас, А., ХНУМХа. Апоморфинді және окситоцинді индуцированный пениса эрекциялау және интактты және кастрацияланған ересек егеуқұйрықтарда ериді: жыныстық стероидтердің әсері. Нейроэндокринология 59, 349-354. Мелис, MR, Stancampiano, R., Argiolas, A., 1994b. Паравентрикулярлық NMDA инъекциясымен индуцирленген пениса эрекциялау және емдеу омитоцина арқылы жүзеге асырылады. Фармакол. Биохим. Behav. 48, 203-207. Мелис, MR, Stancampiano, R., Argiolas, A., 1994c. Apomorphine- және oxytocin-индуцирленген пениса эрекциясының NG-нитро-л-аргинин метил эфирлерінің алдын алу және мидағы іс-әрекеттің орны. Фармакол. Биохим. Behav. 48, 799-804. Мелис, М.Р., Сукку, С., Аргиолас, А., ХНУМХ. Допамин агонисты гипоталамустың паравенцикулярлық ядросында азот оксидінің өндірісін көбейтеді: кремний эрекциясымен және итеруімен корреляция. E. J. Нейроци. 8, 2056-2063. Melis, MR, Succu, S., Iannucci, U., Argiolas, A., 1997. Окситоцин гипоталамустың паравенцикулярлық ядросында азот оксидінің өндірісін көбейтеді: кремний эрекциясымен және итеруімен корреляция. Рег. 69, 105-112 пептидтері. Melis, MR, Succu, S., Mauri, A., Argiolas, A., 1998. Нейтралды оксид өндірісі ерлердің гипоталамусының паравенциклы ядросында байланыссыз пениса эрекциялары мен сіңірілуі кезінде көбейтіледі. E. J. Нейроци. 10, 1968-1974. Melis, MR, Spano, MS, Succu, S., Argiolas, A., 1999a. Окситоцинді антагонист d (CH2) 5Tyr (Me) 2-Orn8-вазотоцина егеуқұйрықта ересектерге байланыста болмайды. Нейроци. Lett. 265, 171-174. Melis, MR, Succu, S., Spano, MS, Argiolas, A., 1999b. Гипоталамустың паравенцикулярлық ядросына енгізілген морфин, байланыссыз эрекциялардың алдын алады және копуляцияны төмендетеді: азот оксидін тарту. E. J. Нейроци. 11, 1857-1864. Melis, MR, Spano, MS, Succu, S., Argiolas, A., 2000. Эволюциялық амин қышқылының, допаминнің және окситоцинді рецепторлардың антагонистінің ерлер егеуқұйрықтарында байланыссыз пениса эрекцияларына және паравенциклулалық азот оксидіне әсері. Behav. Нейроци. 114, 849-857. Melis, MR, Succu, S., Mascia, MS, Cortis, L., Argiolas, A., 2003. Ерекше жасушалық допамин жыныстық белсенділік кезінде еркек егеуқұйрықтарының паравенцикулярлық ядросында артады. E. J. Нейроци. 17, 1266-1272. Melis, MR, Succu, S., Mascia, MS, Argiolas, A., 2004a. Ересек егеуқұйрықтардың паравенцикулярлық ядросындағы каннабиноид CB1 рецепторларының антагонизмі кремнийді эрекциялауды тудырады. Нейроци. Lett. 359, 17-20. Melis, MR, Succu, S., Mascia, MS, Cortis, L., Argiolas, A., 2004b. Ерекше жасушалық қоздырғыш аминқышқылдары жыныстық белсенділік кезінде еркек егеуқұйрықтарының паравенцикулярлық ядросында жоғарылайды: эректилді функциядағы NMDA рецепторлардың негізгі рөлі. E. J. Нейроци. 19, 2569-2575. Melis, MR, Succu, S., Mascia, MS, Argiolas, A., 2005. PD-168,077, селективті допамин D4 рецепторлы агонист, еркек егеуқұйрықтарының паравенцикулярлық ядросына енгізілген кезде, кремний эрекциясын индуцирует. Нейроци. Lett. 379, 59-62. Melis, MR, Succu, S., Mascia, MS, Sanna, F., Melis, T., Succu, S., Castelli, MP, Argiolas, A., 2006a. Ерлер егеуқұйрықтарында SR 141716A индуцированного кремний эрекция: паравенциркулярлық глютамин қышқылын және азот оксидін тарту. Нейрофармакология 50, 219-228. Melis, MR, Succu, S., Sanna, F., Mascia, MS, Melis, T., Enguehard-Gueiffier, C., Hubner, H., Gmenier, P., Gueiffier, A., Argiolas, 2006b. PIP3EA және PD168077, екі селективті допамин D4 рецепторлы агонистер, ерлер егеуқұйрықтарында жыныстық эрекцияны тудырады: мидағы іс-қимыл аймағы және механизмі. E. J. Нейроци. 24, 2021-2030. Melis, MR, Melis, T., Cocco, C., Succu, S., Sanna, F., Pillolla, G., Boi, A., Ferri, GL, Argiolas, A., 2007. Вентральды тикеальдық аймаққа енгізілген омитоцин кремнийдің эрекциясын индуцирует және ядро accumbens MR-де экстракционалды допаминді арттырады. Мелис, А. Argiolas / Neuroscience and Biobehvainment Пікірлер 35 (2011) 939-955 955 және егеуқұйрықтардың гипоталамусының паравенциклдік ядросы. E. J. Нейроци. 26, 1026-1035. Мелис, М.Р., Санна, Ф., Сукку, С., Зароне, П., Бой, А., Аргиолас, А., ХНУМХа. Еркек жыныстық жыныстық мінез-құлықтың алдын-ала және тұтынушы фазаларында омитоциннің рөлі. В: Jastrow, H., Feuerbach, D. (Eds.), Oxytocin Research анықтамалығы: Синтездеу, сақтау және босату, іс-әрекеттер және дәрілік формалар. Nova Publishers Inc, Нью-Йорк, АҚШ, с. 109-125. Мелис, МР, Сукку, С., Санна, Ф., Бой, А., Аргиолас, А., ХНУМХб. Вентальдік субикулумға енгізілген омитоцин немесе амегдаланың постеромедиальді кортикальды ядросы жыныстық эрекцияны тудырады және ерлер егеуқұйрықтарындағы акументалды ядродағы жасушадан тыс допаминді арттырады. E. J. Нейроци. 30, 1349-1357. Мелис, М.Р., Сукку, С., Кокко, С., Кабони, Е., Санна, Ф., Бой, А., Ферри, Г.Л., Аргиолас А., ХНУМХ. Окситоцин вентальдік субикулаға енгізілгенде кремнийдің эрекциясын индукциялайды: азот оксиді мен глютамин қышқылының рөлі. Нейрофармакология 58, 1153-1160. Морелланд, Р.Б., Накани, М., Доннелли-Робертс, Д.Л., Миллер, Л.Н., Чанг, Р., Учик, ME, Terranova, MA, Gubbins, EJ, Хельфрих, RJ, Namovic, MT, El- Masters, JN, Brioni, JD, 2004. Гальфа (qo2) арқылы кальций ағынымен байланыстырылған адамның допаминді D (5) тәрізді рецепторлы тұрақты жасушалық желілерінің салыстырмалы фармакологиясы. Биохим. Фармакол. 68, 761-772. Moos, F., Freund-Mercier, MJ, Guerne, Y., Guerne, JM, Stoeckel, ME, Richard, P., 1984. Окситоцинді және вазопрессинді магнитті-ядролардың in vitro арқылы босату: окситоциннің жеке шығарылуына арнайы фитолитаторлық әсері. J. Эндокринол. 102, 63-72. Murphy, MR, Seckl, JR, Burton, S., Checkley, SA, Lightman, SL, 1987. Ерлердегі жыныстық белсенділік кезінде омитоцин мен вазопрессин секрециясының өзгеруі. J. Clin. Эндокринол. Metab. 65, 738-741. Nishimori, K., Young, LJ, Guo, Q., Wang, Z., Insel, TR, Matzuk, MM, 1996. Окситоцин емізу үшін қажет, бірақ босану немесе репродуктивті мінез-құлық үшін маңызды емес. Proc. Natl. Акад. Ғылыми жұмыс. АҚШ 93, 11699-11704. М., Марвуд, Р., Мкаллистер, Г., Майерс, Ж., Кертис, Ж., Кулаговский, П. Пател, С. Фредман, С., Чэпман, КЛ, Эммс, Ф., Флетчер, А.Е., JJ, Leeson, PD, Ridgill, M., Graham, M., Matheson, S., Rathbone, D., Watt, AP, Bristow, LJ, Rupniak, NM, Baskin, E., Lynch, JJ, Ragan, CI , 1997. L 745,870 биологиялық профилі, DPAMIN D4 рецепторлары үшін жоғары жақындығы бар таңдаулы антагонист. J. Фармакол. Exp. Ther. 283, 636-647. Pedersen, CA, Caldwell, JD, Jerricki, GF, Insel, TR, 1992. Окситоцинді аналық, жыныстық және әлеуметтік мінез-құлықта, Нью-Йорк Ғылым академиясының жылнамалары, том. 652. Нью-Йорк ғылымдар академиясы, Нью-Йорк. Петрович, П., Калиш, Р., Singer, T., Dolan, RJ, 2008. Окситоцин шартты беттерді және амигдалалық белсенділікті аффективті бағалауды жоғарылатады. J. Нейроци. 28, 6607-6615. Pfaus, JG, Everitt, BJ, 1995. Жыныстық мінез-құлықтың психофармакологиясы. В: Knobil, FE, Kupfer, DJ (Eds.), Психофармакология: Төртінші ұрпақ прогресс. Raven Press, New York, pp. 742-758. Райфер, Дж., Аронсон, В. Дж., Буш, П.А., Дори, Ф.Д., Игнарро, Л.Ж., ХНУМХ. Нейтренергиялық, нехолинергическая нейротрансляцияға жауап ретінде корпус каверозумының релаксациясының медиаторы ретінде азот оксиді. N. Engl. J. Мед. 326, 90-94. Roeling, TAP, Van Erp, AMM, Meelis, W., Kruk, MR, Veening, JG, 1991. NMDA-ның мінез-құлық әсері егеуқұйрықтың гипоталамиялық паравенциклды ядросына енгізілді. Brain Res. 550, 220-224. Sachs, BD, 1997. Ерлер егеуқұйрықтарында эструкалды әйелдердің әуедегі хош иісі бар. Физиол. Behav. 62, 921-924. Sachs, BD, 2000. Эректиль функциясының физиологиясы мен жіктелуіне, эректильді дисфункцияға және жыныстық қатынасқа байланысты контекстік көзқарас. Нейроци. Биобехав. Rev. 24, 541-560. Sachs, BD, 2007. Еркектердің жыныстық қатынастарының контекстік анықтамасы. Хорм. Behav. 51, 569-578. Санчес, Ф., Алонсо, Дж.Р., Аревало, Р., Бланко, Е., Айджон, Ж., Вазкез, Р., ХНУМХ. Егеуқұйрықтың гипоталамиялық магнокелулярлы нейросекреторлы ядроларындағы вазопрессинмен және окситоцинмен NADPH-диапораздың бірге өмір сүруі. Ұяшық тінінің суреті. 276, 31-34. Санна, Ф., Сукку, С., Бой, А., Мелис, М.Р., Аргиолас, А., ХНУМХ. Фосфодиэдераз типті 5 ингибиторлары егеуқұйрықта ересектерге байланыссыз эрекцияларды жеңілдетеді: мидағы іс-әрекеттер орны және әрекет ету механизмі. J. Секс. Мед. 6, 2680-2689. Saphier, D., Feldman, S., 1987. Септал және гиппокампалық ынталандылардың паравенциклды ядро нейронына әсері. Нейроход 20, 749-755. Sato-Suzuki, I., Kita, I., Oguri, M., Arita, H., 1998. Егеуқұйрықтың паравенцикулярлық ядросының электрлік және химиялық ынталандыруымен туындаған стероидты иондаушы жауаптар. J. Нейрофизол. 80, 2765-2775. Шуман, Э.М., Мэдисон, DV, 1994. Нитротин және синаптическая функция. Энн. Rev. Нейроци. 17, 153-183. Snyder, SH, 1992. Нитротин: нейротрансмиттерлердің жаңа класында алдымен? Ғылым 254, 494-496. Sofroniew, MV, 1983. Сүтқоректілердің миы мен жұлынындағы вазопрессин мен окситоцин. Трендтер Neurosci. 6, 467-472. Sokoloff, P., Schwartz, JC, 1995. Жарты жылдан кейін жаңа допаминді рецепторлар. Трендтер фармаколы. Ғылыми жұмыс. 16, 270-275. Саутам, Э., Гартувайт, Дж., ХНУМХ. Созылмалы мидағы гидроксид-циклдік GMP сигнал жолдары. Нейрофармакология 32, 1267-1277. Stancampiano, R., Melis, MR, Argiolas, A., 1994. Ерлер егеуқұйрықтарында 5-HT1c агонистері туындаған пениса эрекциялау және итеру: допаминергический және окситоцинергическомпен байланыс. E. J. Фармакол. 261, 149-155. Сукку, С., Массия, М.С., Санна, Ф., Мелис, Т., Аргиолас, А., Мелис, М.Р., ХНУМХ. Cannabinoid CB1 рецепторлық антагонисті SR 141716A эротикалық егеуқұйрықтардың паравенцикулярлық ядросында қосымша клеткалық глутамин қышқылын ұлғайту арқылы кремний эрекциясын индуцирует. Behav. Brain Res. 169, 274-281. Сукку, С., Санна, Ф., Мелис, Т., Бой, А., Аргиолас, А., Мелис, М.Р., ХНУМХ. Еркек егеуқұйрықтардың гипоталамусындағы паравенциклы ядросындағы допаминді рецепторларды ынталандыру кремний эрекциясын индуцирует және ядродағы акумбензияларда экстракционалды допаминді арттырады: орталық окситоцинді тарту. Нейрофармакология 52, 1034-1043. Сукку, С., Санна, Ф., Кокко, С., Мелис, Т., Бой, А., Ферри, Г.Л., Аргиолас, А., Мелис, М.Р., ХНУМХ. Окситоцин егеуқұйрықтардың вентральдік тіке аймағына инъекция кезінде пениса эрекциясын индуцирует: азот оксидінің және циклдік GMP рөлі. E. J. Нейроци. 28, 813-821. Танда, Г., Понтиери, ФЭ, Ди Кьяра, Г., ХНУМХ. Миколинді допаминді киннобиноксидтер мен героинді ортақ M1 опиоидты рецепторлы механизм арқылы белсендіру. Ғылым 276, 2048-2050. Танг, Ю., Рампин, О., Калас, А., Фашинетти, П., Джулиано, Ф., ХНУМХ. Эрик егеуқұйрықта жыныстық эрекцияны бақылайтын анықталған люмбосакралық ядролардың окситоцингиялық және серотонинергиялық иннервациясы. Нейроход 82, 241-254. Теодоз, DT, 1985. Сутраоптикалық ядролардағы окситоцин нейрондарында окситоцин-иммунореактивті терминалдардың синапсы. Nature (Лондон) 313, 682-684. Tindall, JS, 1974. Окситоцинді босатуды ынталандырады. В: Гейгер, СР, Кнобил, Э., Сойер, WH, Гриф, Р., Аствуд, Э.Б. (Eds.), Физиология анықтамалығы. Секция. 7, Эндокринология, том. IV. Американдық физиология қоғамы, Вашингтон қ., С. 257-267. Торрес, Г., Ли, С., Ривьер, С, ХНУМХ. Гипоталамикалық азот оксиді синтезінің және нейропептидтермен колокализацияның онтогениясы. Мол. Ұяшық. Нейроци. 4, 155-163. Uhl-Bronner, S., Waltisperger, E., Martinez-Lorenzana, G., Condes, LM, Freund-Mercier, MJ, 2005. Алдын-ала және егеуқұйрықтың омыртқасындағы витаминдермен байланысатын окситоцинді жыныстық диморфтық білдіру. Нейроход 135, 147-154. Vaccari, C., Lolait, SJ, Ostrowski, NL, 1998. Васопрессин V1b және мидағы окситоцина рецепторларының рибонуклеин қышқылдарын салыстырмалы бөлу. 139, 5015-5033 эндокринологиясы. Van Den Pol, A., 1991. Гипоталамалық прессинаптическома аксунда глутаматты және aspartate immunoreactivity. J. Нейроци. 11, 2087-2101. Veronneau-Longueville, F., Rampin, O., Freund-Mercier, MJ, Tang, Y., Calas, A., Marson, L., McKenna, KE, Stokeck, ME, Benoit, G., Giuliano, F. , 1999. Егеуқұйрықта эпидемиялық эрекцияны бақылайтын автономды ядролардың окситоцингиялық иннервациясы. Нейроход 93, 1437-1447. Винсент, СР, Кимура, Х., ХНУМХ. Аз мидағы гидроксид синтезінің гистохимиялық картасын жасау. Нейроход 46, 755-784. Wagner, CK, Clemens, LG, 1993. Гипоталамустың паравенциклов ядросынан леммальді жұлындағы жыныстық-димофиялық қозғалтқыш ядросына дейінгі нейрофизин бар жол. J. Комп. Нейрол. 336, 106-116. Winslow, JT, Insel, TR, 1991. Еркек жұпар жұпар жұптардағы әлеуметтік мәртебе орталық окситоцинді емдеудің мінез-құлық реакциясын анықтайды. J. Нейроци. 11, 2032-2038. Дана, RA, Rompre, P.-P., 1989. Ми допамині және сыйақы. Энн. Rev. Психол. 40, 191-225. Witt, DM, Insel, TR, 1994. Еркек жыныстық мінез-құлық гипоталамустың паравенцикулярлық ядросындағы окситоцин нейрондарындағы си-фос тәрізді протеинді белсендіреді. J. Нейроэндокринол. 6, 13-18. Witter, MP, 2006. Егеуқұйрықтардың қосындылары: колонналық және ламинарлы ұйымға қатысты топография. Behav Brain Res. 174, 251-264. Woodruff, GN, Foster, AC, Gill, R., Kemp, JA, Wong, EH, Iversen, LL, 1987. N-methyl-d-aspartate үшін MK-801 және рецепторларының өзара әрекеттестігі: функционалдық салдары. Нейрофармакология 26, 903-909. Yamashita, H., Shigeru, O., Inenaga, K., Kasai, M., Uesugi, S., Kannan, H., Kaneko, T., 1987. Окситоцин негізінен цифрлы супраоптическом ядросындағы in vitro-дегі болжамды окситоцинді нейронды қоздырады. Brain Res. 416, 364-368. Yells, DP, Hendricks, SE, Prendergast, MA, 1992. Ядролық парагигантоцеллюлярдың лейкозы: егеуқұйрықтарда жұғу әрекеттеріне әсер ету. Brain Res. 596, 73-79. Жас, WS, Shepard, E., Amico, J., Hennighausen, L., LaMarca, ME, McKinney, C., Ginns, EI, 1996. Мышьяк окситоциннің жетіспеушілігі сүтті босатудың алдын алады, бірақ құнарлылық пен босану емес. J. Нейроэндокринол. 8, 847-854. Zahran, AR, Vachon, P., Куртуа, Ф., Carrier, S., 2000. Анестезирленген егеуқұйрықтардың гипоталамиялық паравенцикулярлық ядросында акуытқыш аминқышқыл рецепторларының агонистерін инъекциядан кейінгі интракараверналық пениса қысымының жоғарылауы. J. Urol.