YBOP 의견 : Coolidge 효과는 인터넷 포르노의 힘 뒤에 있습니다. Coolidge 효과는 포유류 종에서 나타나는 현상으로, 남성이나 여성이 이전의 성 파트너와의 성관계를 거부 한 후에도 새로운 수용 성 파트너에게 소개되면 새로운 성적 관심을 보이는 현상입니다. 성적인 진기함은 높은 도파민에 의해 새로 워진 흥분으로이 습관을 대체합니다. 끊임없이 새로운 흐름이 인터넷 포르노를 과거의 포르노와 다르게 만드는 것입니다.
- 신경 과학 저널 (The Journal of Neuroscience, 15 년 1997 월 17 일, 12 (4849) : 4855-XNUMX;
추상
쿨리지 효과 (Coolidge effect)는 새로운 수용 적 동료에 대한 반응으로 "성적으로 만족되는"동물의 성적 행동을 재개발하는 것을 의미합니다. 동기 행동의 시작과 유지에 mesolimbic dopamine (DA) 시스템의 역할을 감안할 때, 미소 투석은 교미, 성욕감 및 성적 행동의 재개발 중 핵 측위 (NAC) DA 전달을 모니터링하는 데 사용되었습니다. 초기 연구 결과와 일치하여, 발정 후 혈청과 교미를 나타내는 여성은 NAC DA 유출의 유의 한 증가와 관련이 있었다. DA 대사 산물 인 dihydroxyphenylacetic acid와 homovanillic acid의 농도는 상승 하였지만 NAC DA 농도가 기준치에 도달 한시기는 성적 포만의시기와 일치했다. 스크린 뒤에 새로운 수용 여성의 제시는 NAC DA의 약간의 증가를 가져 왔으며, 그것은 소설적인 여성과의 새로운 교미 동안에 유의하게 증가되었다. 본 연구 결과는 새로운 수용성 여성의 자극 특성이 성적으로 포화 된 수컷 쥐에서 NAC DA 전달을 증가시키는 역할을 할 수 있으며, 이는 성 행동의 재발과 관련이 있음을 시사한다.
개요
포만감이있는 숫쥐는 초기 암컷을 새로운 수용성 암컷으로 교체하면 다시 교미를 유도 할 수 있습니다. 이것은 쿨리지 (Coolidge) 효과로 알려져 왔으며 많은 포유 동물 종에서 관찰되었습니다 (윌슨 (Wilson) 등, 1963). 피로 나 우울증과 같은 일반적인 요인으로 인해 여성의 성적인 자극은 여전히 교미를 유도 할 수 있기 때문에 명백한 성적 만족 상태를 설명하기에는 충분하지 않습니다. 성적 포만은 또한 다양한 신경 전달 물질 시스템에서 작용할 수있는 다양한 약물을 투여함으로써 약물 학적으로 상당한 정도로 반전 될 수 있습니다. 이러한 약물에는 요힘빈, 8-OH-DPAT (로드리게스 - 만조 (Manzo)와 페르난데즈 구 스타티 (Fernandez-Guasti), 1994, 1995a), 나 바로 사 (Pfaus and Gorzalka, 1987; 로드리게스 - 만조와 페르난데즈 - 구 스티, 1995a,b), 아포 몰핀 (Mas et al., 1995c). 이러한 약물의 말초 작용이 배제 될 수는 없지만 (예 : 발기 기능에 대한 아드레날린 작용), 선택적 중추 신경 장애 실험을 바탕으로 성욕 감량의 핵심 메커니즘에 대한 효과가 제안되었다로드리게스 - 만조와 페르난데즈 - 구 스티, 1995a) 및 내측 검사 전 영역에서 도파민 성 대사를 모니터링하는 미세 투석 실험 (Mas 등, 1995a,b).
중심 메카니즘이 Coolidge 효과의 성 행동의 재발 성을 중재 할 수 있다고 가정 할 때, 가능성이있는 후보는 복부 피 두드러기 영역에서 NAC로 투영되는 중 배뇌 도파민 (DA) 시스템이다. Mesolimbic DA는 성적으로 수용적인 여성의 신호와 같은 환경 적 자극의 평가와 교미를 포함한 목표 지향적 행동의 조직을 포함하는 복잡한 통합 과정에서 일차 변조기로 작용하는 것으로 보인다 (피 비거와 필립스, 1986; 블랙번 (Blackburn) 등, 1992; Phillips et al., 1992; 르 모알, 1995; 살라몬, 1996).
중뇌 DA 뉴런이 1 차 보상 및 보상 예측 신호에 반응하지만, 새로운 또는 예측 불가능한 환경 자극은 반복적 인 훈련 기간 동안 가장 확실하게 신경 활성화를 유도합니다 (Fabre et al., 1983; 슐츠, 1992; Mirenowicz와 Schultz, 1994). 쥐의 성적 행동의 시작과 유지에있어서 중파 평형 상 DA의 중요한 촉진 작용을 뒷받침하는 많은 증거가있다 (Pfaus와 Everitt, 1995), 그리고 다수의 미세 투석 연구는 남성 성 행동의 유행 및 유행 단계에서 NAC DA 유출의 증가를보고합니다Pfaus 등, 1990; 플렘 (Pleim) 등, 1990; Damsma 등, 1992; Wenkstern et al., 1993; Fumero 외, 1994; Mas et al., 1995b). 그러나 성행위와 성행위의 재발에 대한 신경 화학적 상관 관계에 관한 자료는 상대적으로 거의 없다. 응용 프로그램 생체내에서 Coolidge 효과 동안 mesolimbic DA neurotransmission을 모니터링하기위한 미세 투석은 교미, 성욕 및 교합 재개에서 NAC DA의 역할을 검사 할 수있는 독특한 기회를 제공합니다.
(1) 성적 섭취의 시작과 함께 NAC의 세포 외 DA 농도가 과소 세포 수치로 되돌아가는지를 동반하는지 여부를 결정하기 위해 미세 투석 실험이 수행되었다. (2) 성적으로 포화 된 "수컷 쥐가 새로운 수용성 여성과 함께 NAC DA 유출의 증가와 관련이있다.
대상 및 방법
과목. 찰스 리버 캐나다 (St. Constant, Quebec, Canada)에서 얻은 Animal Care Center (브리티시 컬럼비아 대학)에서 얻은 수컷 Sprague Dawley 쥐와 암컷 Long-Evans 쥐를 와이어 메쉬 우리 (18 × 25 × 65 cm, 케이지 당 5). 식민지 방은 역 20 시간의 명암 사이클에서 ~12 ° C의 온도로 유지되었다. 쥐는 음식 (Purina Rat Chow)과 물에 무제한으로 접근 할 수있었습니다.
뇌 미세 투석 전에 수술 및 행동 검사.암컷 쥐를 halothane gas anesthesia (Fluothane, Ayerst Laboratories) 하에서 최소 4 주 전에 양측으로 난소 절제하여 검사 하였다. 자극 시험에서 여성의 성적 수용도는 각 시험 기간 전에 에스트라 디올 벤조 에이트 (10 μg)와 프로게스테론 (500 μg), 48 및 4 hr의 피하 주사에 의해 유도되었다. 수컷 쥐는 철망 바닥이있는 Plexiglas 챔버 (4 × 35 × 35 cm)에서 두 번, 40 d 간격으로 성행위를 위해 선별 검사를 받았다. 두 가지 선별 검사 동안 5 min 이내의 여성과 사정의 15 min 이내의 내강을 포함하는 수행 기준에 도달 한 수컷 쥐만이 미세 투석 프로브 가이드 캐뉼라로 이식되었다.
수컷 쥐 (n = 5)을 정위 수술 전에 ketamine hydrochloride (100 mg / kg, ip) 및 xylazine (10 mg / kg, ip)으로 마취시켰다. Microdialysis probe guide cannulae (19 게이지)를 NAC 위에 양측에 이식하였고 (bregma : 전방, + 1.7 mm, 내측, ± 1.1 mm, 복부, -1.0 mm, 평면 두개골) 치아 아크릴과 보석상의 나사. 양측 가이드 캐 뉼러 임플란트는 성공적인 미세 투석 실험을위한 기회를 극대화하기 위해 사용되었습니다. 다행히도, 본 실험에서, 오직 하나의 캐뉼라가 각 쥐에 필요했다. 실험의 남은 기간 동안 수컷 쥐를 커다란 플라스틱 케이지 속에 옥수수 밭 침구와 함께 개별적으로 수용했다. 수술 후 1 주일에 랫트의 성행위를 검사했다. 이 훈련 중 시험 챔버에는 슬라이딩 플렉시 유리 스크린이 장착되어 챔버를 크고 작은 구역으로 나눴습니다. 수컷 쥐를 큰 구획에 넣고 15 분 후에 여자를 스크린 뒤에 두었다. 15 분 예비 기간 후, 스크린을 제거하고, 래트를 30 분 동안 교접시키는 것이 허용되었다. 4 d마다 3 회의 교육 세션이 진행되었습니다. 모든 쥐는 매 세션마다 성능 기준에 도달했습니다.
Coolidge 효과 실험. 쥐를 쿨리지 효과 실험 전에 12-18 시간의 미세 투석 프로브로 일방적으로 이식하고 음식과 물을 자유롭게 섭취 할 수있는 시험실의 큰 칸에 넣었다. 실험 아침에, 미세 투석 샘플을 15 분마다 수집 하였다. 실험은 다음과 같은 7 단계로 구성됩니다. (1) 기준선 (최소 60 분). (2) 화면 뒤의 여성 1 (15 분); (3) 1 분이 산이없이 통과 할 때까지 여성 30과 교미한다; (4) 화면 뒤에있는 여성 1의 재 도입 (15 분); (5) 1 분 동안 여성 15에 액세스 (마운팅이 발생한 경우이 단계는 3 단계로 처리됨). (6) 화면 뒤에 여성 2 소개 (15 분); 7 분)에 대한 여성 2과 교미.
행동은 JVC 비디오 시스템을 사용하여 저조도 환경에서 촬영되었고 테스트 실 외부에 위치한 비디오 모니터에서 관찰되었습니다. 성적 행동의 표준 척도는 컴퓨터와 적절한 소프트웨어를 사용하여 기록되었습니다 (Holmes 등, 1987).
미세 투석 실험 후, 동물에게 과량의 클로 랄 수화물을 투여하고 식염수 및 포르말린 (4 %)으로 심장 내로 관류시켰다. 두뇌를 얇게 썰어 냉동시키고,이어서 코로나 바이올렛으로 코로나 섹션을 염색하여 미세 투석 프로브의 위치를 결정했습니다. NAC 내에서 프로브 배치가있는 쥐만 행동 및 신경 화학 분석에 사용되었습니다.
Microdialysis 및 HPLC- 전기 화학 검출. Microdialysis 프로브는 반투명 중공 섬유 막 (2 mm 막 노출, 340 μm 외경, 65000 분자량 차단, Filtral 12, Hospal)이 원심 끝에 디자인 된 동심원 구조를 가졌습니다. 프로브는 수정 된 링거액 (1.0 m 인산 나트륨 완충액, pH 0.01, 7.4 mmCaCl2, 3.0 mm KCl, 1.0 mmMgCl2, 147 mm NaCl)와 주사기 펌프 (Model 22, Harvard Apparatus, South Natick, MA)를 사용하여 측정 하였다. microdialysis probe guide collar는 microdialysis probe를 guide cannula 안쪽에 고정 시키는데 사용되었다. 시험 챔버 상단에 장착 된 액체 회전 장치 (Instech 375s)에 부착 된 강철 코일을 사용하여 프로브 튜브를 보호했습니다 (Fiorino 등, 1993).
DA와 그의 대사 산물 인 dihyroxyphenylacetic acid (DOPAC)와 homovanillic acid (HVA)를 포함하는 Microdialysate 분석 물은 역상 크로마토 그래피 (Ultrasphere column, Beckman, Fullerton, CA, ODS 5 μm, 15 cm, 4.6 mm, 내경 0.083m 나트륨 아세테이트 완충액, pH 3.5 (5 % 메탄올)을 사용하여 정제 하였다. 분석 물 농도는 전기 화학적 (EC) 검출에 의해 정량화되었다. 장치는 Bio-Rad (Richmond, CA) 펌프, Valco Instruments (Houston, TX) EC10W 2 포지션 주사기, ESA (Bedford, MA) Coulochem II EC 검출기 및 이중 채널 차트 기록계 and Zonen, Bohemia, NY). 전기 화학 검출기 파라미터는 다음과 같다 : 전극 1, + 450 mV; 전극 2, -300 mV; 및 보호 셀, -450 mV. 수행 된 일반적인 프로브 복구 체외에서 실온에서 DA의 경우 22 %, DOPAC의 경우 18 %, HVA의 경우 18 %였다.
결과
행동
Coolidge 효과 실험으로부터의 행동 측정은 표 1에 제시되어있다. 1. 장착, 내강 및 사정에 대한 잠복기뿐만 아니라 첫 번째 사정 후 postejaculatory 간격은 이전 훈련 세션의 것과 유사했다 (데이터는 표시되지 않음). 이것은 미세 투석 절차가 정상적인 성 행동을 변화시키지 않았 음을 시사한다. 성적 사정의 발달은 기준이 충족되기 전 평균 사정 횟수 (7.8 ± 0.5), 각 사정 전 선입 내진 횟수의 점진적인 감소, 사정 후 간격의 점진적인 증가 (데이터는 표시되지 않음) 이전 연구에서보고 된 것과 유사했다 (해변과 요르단, 1956; 파울러와 포렌, 1961; 피셔, 1962; 버만 트 (Bermant) 등, 1966; 로드리게스 - 만조 (Manzo)와 페르난데즈 구 스타티 (Fernandez-Guasti), 1994; Mas et al., 1995d). 여성의 1로 얻은 사정 횟수, 여성 1과 교미하는 데 걸린 시간, 포화 기준에 도달하는 데 필요한 여성 1의 프리젠 테이션 수와 관련하여 개인의 변동성이 관찰되었습니다 (표 1, 하단). 일부 쥐는 1 단계가 완료 될 때까지 여성 5을 여러 번 재 도입해야했습니다 (n = 3). 1 암컷을 스크린 뒤에 놓고 칸막이를 제거하는 행위는 교미를 유도하는 주요 유익한 신호 역할을했을 수 있습니다. 이전에 사용되었지만 산이없는 30 분의 포화 기준 (해변과 요르단, 1965; Mas et al., 1995b)는 임의적이며, 주어진 시간 동안 쥐가 장착되지 않았 음을 보장하지 않습니다. 그럼에도 불구하고 지연 또는 제거 및 교체 절차는 여성 1 (예 : 4 및 5 단계)과의 갱신 된 교합에서 안정적으로 발생하지 않았습니다.

Coolidge 효과 실험 동안의 행동
모든 쥐는 Coolidge 효과를 보였다. 암컷 2을 스크린 뒤에 배치하고, 특히 파티션을 제거하는 것과 관련된 활동은이 결과에 기여했을 수도 있지만,이 이벤트 자체만으로는 실험 초기에 교미를 갱신하기에 충분하지 않았습니다. 여성 1 및 여성 2과의 성 행동 측정 간의 비교는 t Bonferroni 보정으로 테스트합니다. 여성 2에 대한 반응에서의 장착 및 내강 잠복기는 여성 1에 대한 첫 번째 교합성 시합에서와 크게 다르지 않았지만, 일반적으로 여성 2과의 성적 행동은 덜 강하게 나타 났으며, 사정이 훨씬 적음 (평균, 0.6 대 4.2; F = 49.86;p <0.01) 및 도입 (평균 = 11.2 대 37.0;F = 20.17; p <0.05) 첫 1 시간 동안. 암컷 2과 XNUMX의 첫 XNUMX 시간 동안 탈것의 수는 크게 다르지 않았습니다.
실험의 포화 부분 (즉, 암컷 1)에서 사용 된 암컷은 수컷과의 접촉이 완전히 지속되는 동안 강한 호이 예 (호핑과 다트닝)와 수용성 (즉, 전만 동요) 행동을 나타냈다는 것을 알아 두는 것이 중요합니다 .
신경 화학
처음 3 개의 기준선 샘플의 평균 ± SEM으로 나타낸 미세 투석액에서 DA와 그 대사 산물의 기본 나노 몰 농도는 : DA, 3.0 ± 0.7; DOPAC, 619.1 ± 77.7; 및 HVA, 234.2 ± 49.0 (프로브 회복을 위해 보정되지 않음;n = 5). 이 값은 100 % 기준선 점수를 나타냅니다.
실험의 각 단계에 해당하고 모든 쥐에게 공통적으로 나타나는 행동 학적으로 정의 된 데이터 요소는 신경 화학 분석에 사용되었습니다. 이들은 다음을 포함했다 : (1) 여성 1의 첫 도입 이후 7 개 샘플, (2) 여성 1과의 보조적인 행동이없는 4 개의 샘플, 그리고 여성 3 제시 후 5 개 샘플 (2). 그림 1 DA의 농도 변화 (선 그래프, 중간) 및 DA 대사 산물 (라인 그래프, 상단) 병렬 행동 (bar graph, 바닥) 쿨리지 효과 테스트 동안.
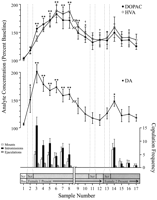
그림. 1.
핵은 쿨리지 효과 (Coolidge effect) 동안의 성 행동의 신경 화학적 상관 관계를 의미합니다. 처음 8 개 샘플은 1에서 3 단계까지의 시간순 연속 데이터 포인트를 나타냅니다. 샘플 1는 네 번째 및 마지막 미리 계산 된 기준 샘플입니다 (낮은). 샘플 2은 화면 뒤에 여성 1의 소개를 나타냅니다 (Scr). 15 분 후에, 스크린을 제거하고, 래트를 교접시키는 것이 허용되었다 (샘플 3-8). 그만큼하다 를 시청하여 이에 대해 더 많은 정보를 얻을 수 있습니다. x축은 최초의 암컷과 장기간 교미 한 3 마리의 쥐의 데이터를 제외시켰다. 마지막 9 개의 샘플은 또한 연대순으로 연속되었다. 샘플 9 및 10는 단계 3의 포화 기간 (즉, 마운트가없는 30 분)에 해당합니다. 여성 1을 화면 (샘플 11) 뒤에 다시 삽입하고 15 분 후에 화면 12을 제거했습니다. 교미가없는 15 min 후, 여성 2을 스크린 (샘플 13) 뒤에 두었다. 샘플 14-17은 여성 2과의 교합에 해당합니다. 각 15 미세 미세 투석 샘플과 관련된 마운트, 인트 루먼트 또는 사정 횟수는 하단 막대 그래프. 신경 화학적 데이터는 기준 농도의 백분율로 표시됩니다. NAC DA의 변화 (닫힌 사각형), DOPAC (닫힌 서클), HVA (열린 동그라미) 유출은 선 그래프. 다음과 같은 비교가 이루어졌다 : 기본 샘플 1 대 샘플 2-10; 새로운베이스 라인 샘플 10 대 샘플 11 및 12; 새로운 기준 샘플 12 대 샘플 13-17 (*p <0.05; ** p <0.01). 독립적 인 t 테스트는베이스 라인 값 (샘플 1, 10 및 12) 사이에서 이루어졌습니다. 첫 번째 기준선 (샘플 1)과의 중요한 차이점에 대해서는 †p <0.05.
여성 1 (샘플 1-12) 및 여성 2 (샘플 12-17)과 관련된 신경 화학적 데이터에 대해 단방향 반복 측정 ANOVA가 수행되었습니다. 선험적으로 비교는 Dunn의 다중 비교 테스트 (Bonferroni t). (1) 샘플 1 - 2 (10) 샘플 1 - 2 (10) 샘플 (11) 샘플 12 대 샘플 1 및 3 (여성 12에 다시 노출)에 대한 샘플 13 - 17 (2) , 그리고 (XNUMX) 세 번째 기준선 (샘플 XNUMX) 대 샘플 XNUMX-XNUMX (여성 XNUMX 노출).
여성 1에 대한 반응으로 DA 유출의 유의 한 전반적인 변화가 있었다 [F (11,44) = 8.48; p <0.001] 및 암컷 2 [F (5,20) = 2.83;p <0.05]. 여성 1이 스크린 뒤에있을 때 DA 유출의 상당한 증가가 발견되었습니다 (+ 44 %,p <0.05; 샘플 2). 교미하는 동안, DA 농도는 더 증가하여 최대 값 (+ 95 %;p <0.01) 첫 번째 교전 시합 (샘플 3) 동안. DA는 교미하는 동안 상승 된 상태로 유지되었고 장착이 발생하지 않은 30 분 기간 (샘플 9 및 10)에만 기준 농도로 돌아 왔습니다. 스크린 뒤에서 여성 1의 재 도입 (샘플 11)도 물리적으로 상호 작용할 기회도 없었지만 장착하지 않은 경우 (샘플 12), 두 번째 기준 값 (샘플 10)에 비해 DA 농도가 상승했습니다. 화면 뒤에있는 여성 2 (샘플 13)의 존재는 통계적 유의성에 도달하지 않은 세 번째 기준 값 (샘플 12)에서 DA 유출 (12 %)의 작은 증가를 초래했습니다. 암컷 2와의 교미를 갱신 한 결과 상당한 (34 %) 증가 (p 첫 번째 교미 샘플 (샘플 0.05) 동안 DA 유출에서 <14). 약한 교제 행동이 다음 세 개의 샘플에서 계속되었지만 DA 농도는 기준치 (샘플 15-17)로 감소했습니다. 독립적 인 t "기준선"샘플 (즉, 1, 10 및 12) 사이에서 실시 된 테스트는이 값이 크게 다르지 않음을 보여주었습니다.
암컷 1이 재 도입되었을 때 교미를 재개 한 3 마리의 랫드에서, 암컷 1이 샘플 (25-47 % 범위)과 시료 (13-37 %)의 교미 기간 여성의 재 도입 전에. 그러나 이러한 증가는 성행위가 활발하여 사정을 유발했을 때만 발생합니다.
DOPAC의 중요한 변화F (11,44) = 9.57; p <0.001] 및 HVA [F (11,44) = 12.47; p <0. 001] 농도는 여성 1에 대한 반응에서 발견되었지만 여성 2는 아닙니다. 대사 산물 농도는 화면 뒤에서 여성 15 (샘플 1)을 제시하는 동안 약간 증가했지만 (두 경우 모두 + 2 %), 이는 유의미하지 않았습니다. 통계적으로. 그러나 교미 (샘플 3-8) 동안 DOPAC 및 HVA의 농도가 크게 증가하여 최대 값 (각각 +80 및 + 86 %)에 도달했습니다. p <0.01) 60 분 후 (두 경우 모두 샘플 6). 암컷 1 (샘플 9 및 10)과의 접촉이 끝날 때 성적 활동이없는 기간 동안 대사 산물 농도가 감소했지만, 농도는 여전히 기준선 (p 두 경우 모두 <0.05). 스크린 뒤에서 여성 1의 재 도입 (샘플 11), 스크린 제거 후 여성 1에 대한 접근 (샘플 12), 여성 2의 도입 (샘플 13)은 대사 산물 농도의 변화를 초래하지 않았습니다. 기준선 (샘플 23)에 비해 DOPAC 및 HVA 농도의 약간의 증가 (두 경우 모두 + 12 %)는 암컷 2 (샘플 14)와의 첫 번째 교미에 해당합니다. 그러나 이러한 증가는 단기적이며 나머지 15 개 샘플 (17–XNUMX)에 대한 기준 값으로 감소했습니다. 독립적 인 t"기준선"샘플 (즉, 1, 10 및 12) 사이에서 실시 된 테스트는 두 번째 및 세 번째 기준선 값 (각각 10 및 12 샘플)이 서로 다르지 않더라도 첫 번째 기준 샘플과 비교하여 크게 상승한 것으로 나타났습니다 DOPAC 및 HVA (p 두 경우 모두 <0.05).
조직학
Microdialysis 프로브는 NAC에 위치했습니다 (그림.2)을 브레 그마 (평면 두개골)에서 + 1.20에서 + 1.70 mm까지 확장 한 범위에서. 중측 평면에서도 가변성이 있었다. 데이터는 NAC의 셸 및 코어 하위 영역에서 샘플링을 반영합니다.
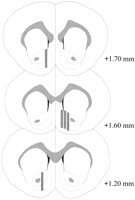
그림. 2.
Coolidge 효과 실험에 사용 된 수컷 쥐의 NAC 내 미세 투석 (microdialysis) 탐침의 위치. 음영 처리 된 사각형 미세 투석 (microdialysis) 프로브의 노출 된 멤브레인 영역에 상응한다. 연속 코로나 뇌 절편은Paxinos와 Watson (1986).
토론
이전의 연구 결과와 일치하여, 현재의 결과는 다음과 같은 결과에 의해 평가 된 수컷 쥐의 성 행동의 유익하고 완결 한 구성 요소와 관련된 향상된 중배엽 DA 전달을 입증한다.생체내에서 미세 투석 (Mas et al., 1990; Pfaus 등, 1990;플렘 (Pleim) 등, 1990; Damsma 등, 1992; Wenkstern et al., 1993; Fumero 외, 1994; Mas 등, 1995a,b,d). 또한, 이러한 결과는 새로운 수용성 여성 (쿨리지 효과)에 대한 반응으로 성적 포화 및 이후의 재발 성을위한 신경 화학적 상관 관계를 제공합니다. 본 연구 결과는 새로운 수용성 여성의 자극 특성이 성행위가 시작된 남성 쥐에서 NAC DA 전달을 증가시키는 역할을 할 수 있으며 이는 성적 행동의 재발 성과 관련이 있음을 시사한다. 이것은 화면 뒤의 소설적인 여성을 제시하는 동안 NAC DA의 약간의 증가에서 분명히 나타나며 여성 2과의 새로운 교미 작용 동안 더 확실한 증가로 가장 확실하게 나타납니다 (그림.1).
화면 뒤의 첫 번째 수용체 암컷의 존재는 NAC DA 유출 (기준선에서 44 %)이 유사한 디자인을 사용하여 이전 실험에서보고 된 것과 비슷한 강도의 식욕 증가를 가져왔다 (30 % Pfaus 등, 1990; 35 %Damsma 등, 1992). 또한 이러한 연구와 일치하는 것은 NAC DA 유출이 교미 중에 더 향상되었다는 관찰이었습니다 (현재 실험에서 기준선보다> 95 %까지). 향상된 NAC DA 릴리스와 관련된 것으로 최종 동작을 볼 수 있지만 (Wenkstern et al., 1993; 윌슨 (Wilson) 등, 1995), 성적 행동의 맥락에서 "식욕을 돋 우는"과 "완악 한"이라는 용어를 조사하는 것이 중요합니다. 여성이 화면 뒤에서 존재하는 단계는 독점적으로 유쾌하거나 예비 적이지만, 교미 단계에서의 행동은 순수하게 만연한 것으로 간주 될 수 없습니다. "식욕을 자극하는"은 동기가있는 행동 (교미)의 완성으로 이끄는 모든 행동을 묘사하는 데 사용할 수 있기 때문에, "정상"단계에서 활동하는 동안 남성이 보여주는 주요 행동은 식욕 부진으로 가장 잘 묘사됩니다. 남성은 자신의 시간과 노력의 대부분을 교미하기 위해 여성을 찾는데 보낸다. 이와 관련하여, 우리는 최대 NAC DA 전송을 완료와 상관시킬 수 있습니다 만큼 잘 남성 쥐 성적 행동의 강렬한 식욕을 자극하는 성분.
두 번째 소설 여성에 대한 접근은 모든 피험자에서 새로운 교미를 가져왔다. 이전의 연구에 따르면, 현재 실험에서 사용 된 프로토콜과 비슷한 행동 프로토콜을 사용하여 포만감에 대한 교미가 허용 된 대다수의 랫트는 24 시간 후에 테스트를 거쳤을 때 짝짓기를 재개하지 못했다 (해변과 요르단, 1956). 후각뿐만 아니라 시각 및 청각 단서를 포함 할 수있는 여성 2의 새로운 자극 특성의 존재는 새로운 교미를 초래할 가능성이 높습니다. 흥미로운 질문은, 남아있는 쥐가 최근에 짝짓기 한 여성에게서 소설적인 여성을 구별하는 메커니즘에 의한 것입니다. 그 메커니즘을위한 사이트는 주요 후각 시스템에 놓여있을 수 있습니다. 이 시스템의 완전성은 햄스터의 쿨리지 효과 (Coolidge effect for hamster)에 필수적이라고보고되었다Johnston과 Rasmussen, 1984). 그러나 vomeronasal-accessory olfactory system은 페로몬 기억 과정이 최근에 생쥐에서 기술되었다.Kaba 등, 1994) 또한 주요 후보자입니다. 이와 관련하여, NAC DA 전송의 증가는 생체내에서 발정기에서 암컷 쥐에게 노출 된 침구가있는 수컷 쥐의 voltammetry (Louillot 등, 1991; 미첼과 그라톤, 1992). 또한, K+ 액세서리 후각 구의 vomeronasal 신경층에 직접뿐만 아니라 액세서리 후각 전구 자체에, NAC DA 전송을 증가시키기에 충분했다 (미첼과 그라톤, 1992).
여성 15과 첫 번째 2 분의 교합은 NAC DA의 의미있는 증가와 관련이 있습니다. 암컷 1과 대조적으로, 암컷 2과의 상호 작용은 정량적 (12 %) 또는 암컷 (34 %) 단계 중 동일한 양의 NAC DA에서 증가를 나타내지 않았다. 그러나 NAC DA의 이러한 작은 증가는 여성 2과 비교하여 여성 1이 표시 한 성행위의 감소 된 수준과 잘 관련이 있습니다. 포만 치료 단계에서 대사 산물의 농도가 상승하여 초기 기준치 (샘플 10)에서 크게 상승한 새로운 기준선 농도 (샘플 12 및 1)를 나타냈다.
DOPAC와 HVA 농도가 증가하는 일시적인 지연은 모 화합물의 대사 산물로서의 형성과 일치한다. 약리학 적으로 유도되지 않은 자연적 행동 동안 적어도 미세 투석 대사 물질 농도는 신경 활동의 유용한 지표를 제공한다고 제안되어왔다.Damsma 등, 1992; Fumero 외, 1994). DA 농도가 시험 초기 값으로 되돌아 왔을 때이 실험에서 성 활동이없는 동안에도 대사 산물 농도가 계속 증가했다는 사실은이 제안에 의심을 던져줍니다.
이 실험에서 보인 DA 대사 산물 농도의 지속적인 상승은 포화 상태에 이르기까지 첫날 쥐에서 관찰 된 DA 대사 산물의 내측 선구자 영역 (mPOA) 프로파일을 반영한다 (Mas 등, 1995a,b). 교미 기간이 포화에 도달하는 데 필요한 시간보다 훨씬 짧은 고정 기간 인 경우 NAC 또는 mPOA에서 DOPAC 및 HVA 농도의 지속 상승이 항상 관찰되는 것은 아닙니다. 예를 들어, 많은 연구에 따르면 DOPAC 농도가 증가하고 교미 동안 증가했지만 여성이 제거 된 직후 기준치로 떨어졌습니다 (Pfaus 등, 1990; 플렘 (Pleim) 등, 1990; Damsma 등, 1992;헐 (Hull) 등, 1993; Wenkstern et al., 1993; 헐 (Hull) 등, 1995). 에 의해 연구에서 Mas et al. (1995b)mPOA에서 DOPAC과 HVA의 기저 세포 외 농도는 성기능이없는 기간에 해당하는 4 연속 일 동안 계속 증가했다. 4 일째에, 동물들이 교미를 재개하기 직전에, 대사 산물의 기초 농도는 presatiation 값에 가깝다. 저자들은 신경 화학적 변화의 패턴을 DA 수용체 차단제 투여 후 나타나는 변화와 비교했다 (Zetterström et al., 1984; Imperato and DiChiara, 1985) 그리고 성적 비활성 상태가 "내인성 신경 이완제"로 작용할 수있는 prolactin 분비를 통해 중재 될 수 있다고 제안했다 (Mas 등, 1995a,b,d). 신경 이완제 투여가 세포 외 대사 물 농도 및 DA 유출의 증가를 동반한다는 것이 명백하다 (Zetterström et al., 1984; Imperato and DiChiara, 1985). 운수 나쁘게, Mas et al. (1995a,b)는 mPOA DA 농도를 검출 할 수 없었다. 본 연구에서는 NAC의 DA 농도가 과소 전 값으로 돌아 왔지만 DOPAC 및 HVA 농도는 여전히 높아졌다. 이 패턴은 성욕 감을 유도하기 위해 NAC에서 작용하는 내인성 신경 이완제의 역할과 일치하지 않습니다.
동기 행동에서 중배엽 DA 뉴런의 관여를 감안할 때 (피 비거와 필립스, 1986; 블랙번 (Blackburn) 등, 1992; Kalivas 외, 1993; 르 모알, 1995)과 새로운 환경 적 자극에 대한 민감성 (Fabre et al., 1983; 슐츠, 1992; Mirenowicz와 Schultz, 1994), 새로운 여성에 대한 반응으로 관찰 된 NAC DA의 세포 외 농도의 증가는이 DA 시스템에서의 활성이 성 행동의 재 활성화에 중요하다는 가설과 일치한다. 또한, DA 전송에서의 유행적이고 유익한 증가의보고 (헐 (Hull) 등, 1993, 1995;Mas et al., 1995b; 사토 (Sato) 등, 1995) 및의 연결 활동 (Shimura et al., 1994)는 성 행동 중 남성 쥐의 mPOA에서이 구조가 Coolidge 효과의 새로운 교미 특징에 기여할 수 있음을 시사한다.
동기 행동에서 중배엽 지방 계통의 일반적인 역할을 유지하면서 식후에 식사 전, 식사 중 및 복용 직후에 DA의 세포 외 농도가 상승하고 나중에 ~30 분의 기준치로 돌아온다는 것이 잘 입증되어 있습니다윌슨 (Wilson) 등, 1995). 음식에 의해 유발 된 포만감은 그것의 감각적 속성에 의해 영향을 받는다는 것은 잘 알려져 있습니다. 인간과 동물은 포만감을 느끼기 위해 먹은 음식을 거부하고 먹지 않은 다른 음식을 먹는다.롤, 1986). 이것은 NAC에서의 세포 외 DA 유출이 새로운 형태의 음식의 제시에 의해 선택적으로 증가 될지 여부에 관한 문제를 야기하지만, 최근 연구에서보고 된 것과 유사한 방식으로 최근에 섭취 된 음식에 의해서가 아니라 성적인 동기 부여. 확인되면, 자연 보상, 포만감 및 중뇌 변형 률 DA 전달의 감각적 특성 사이의 일반적인 관계는 동기 과정의 조절에서이 신경계에 결정적인 역할을 함을 의미하며, 이로 인해 중독이 심각한 식사 및 성기능 장애로 이어질 수 있습니다 .
참조
- ↵
- 해변 FA,
- 요르단 패
(1956) 수컷 쥐의 성적 피로와 회복. QJ Exp Psychol 8 : 121-133.
- ↵
- 베르만 트 G,
- 로트 DF,
- 앤더슨 패
(1966) 수컷 쥐의 copulatory 행동에서 Coolidge 효과의 시간 특성. J Comp Physiol 정신과 65 : 447-452.
- ↵
- 블랙번 JR,
- Pfaus JG,
- 필립스 AG
(1992) 도파민은 탐욕적이고 방어적인 행동을합니다. Prog Neurobiol 39 : 247-279.
- ↵
- Damsma G,
- Pfaus JG,
- Wenkstern D,
- Phillips AG,
- 피비거 HC
(1992) 성 행동은 수컷 쥐의 줄기와 줄무늬에있는 도파민 전달을 증가시킵니다 : 참신함과 운동과의 비교. Behav Neurosci 106 : 181-191.
- ↵
- 파브르 남,
- 롤 ET,
- 애쉬튼 JP,
- 윌리엄스 G
(1983) 행동하는 원숭이의 복부 tegmental 지역에서 뉴런의 활동. Behav Brain Res 9 : 213-235.
- ↵
- 피비 거 (Fibiger) HC,
- 필립스 AG
(1986) 보상, 동기 부여, 인식 : 중뇌 두뇌 도파민 시스템의 정신 생물학. 생리학 핸드북 : 신경계 IV, 블룸 FE, 가이거 SD (미국 생리학 협회, 베데스다, MD), pp 647-675.
- ↵
- 피오리노 DF,
- Coury AG,
- 피비 거 (Fibiger) HC,
- 필립스 AG
(1993) 복부 tegmental 지역의 보상 사이트의 전기 자극은 쥐의 측벽 핵에 도파민 전송을 증가시킵니다. Behav Brain Res 55 : 131-141.
- ↵
- 피셔 A
(1962) 수컷 쥐의 성 포화에 자극 변화의 효과. J Comp Physiol 정신과 55 : 614-620.
- ↵
- 파울러 H,
- Whalen RE
(1961) 수컷 쥐의 인센티브 자극 및 성 행동의 다양성. J Comp Physiol 정신과 54 : 68-71.
- ↵
- 후 메로 B,
- Fernendez-Vera JR,
- 곤잘레스 - 모라 JL,
- 매스 남
(1994) 남성의 성적 행동과 관련된 forebrain 영역의 monoamine turnover의 변화 : 미세 투석 연구. 뇌 해상도 662 : 233-239.
- ↵
- 홈즈 GM,
- 홈즈 DG,
- 삭스 BD
(1987) 설치류 성행위 및 일반적인 이벤트 기록을위한 IBM-PC 기반 데이터 수집 시스템. 피시 올 비바 프 44 : 825-828.
- ↵
- 헐 EM,
- 이튼 RC,
- 모세 야,
- 로레인 DS
(1993) 교미는 수컷 쥐의 내측 검사 부위에서 도파민 활성을 증가시킵니다. 생명 과학 52 : 935-940.
- ↵
- 헐 EM,
- Jianfang D,
- 로레인 DS,
- 마츠 츠 위크 패
(1995) 내측 유두 주위에서의 세포 외 도파민 : 성적 동기 부여 및 호르몬 제어에 대한 함의. J 신경 과학 15 : 7465-7471.
- ↵
- 임페라토 에이,
- 디 치아라 G
(1985) trans-striatal 투석으로 연구 한 전신 신경 이완제 투여 후 깨어있는 쥐의 도파민 방출과 대사. J 신경 과학 5 : 297-306.
- ↵
- Johnston RE,
- 라스무센 K
(1984) 수컷에 의한 암컷 햄스터의 개별 인식 : 화학적 단서와 후각 및 vomeronasal 시스템의 역할. 피시 올 비바 프 33 : 95-104.
- ↵
- 카바 (Kaba) H,
- 하야시 Y,
- Higuchi T,
- 나카니치 에스
(1994) metabotropic 글루타메이트 수용체의 활성화에 의한 후각 기억의 유도. 과학 265 : 262-264.
- ↵
- Kalivas PW,
- Sorg BA,
- 후크 MS
(1993) 정신 자극제에 대한 감작의 약학 및 신경 회로. Behav Pharmacol 4 : 315-334.
- ↵
- 르 모알 남
(1995) Mesocorticolimbic dopaminergic 뉴런. 기능 및 규제 역할. Psychopharmacology에서 : 4 세대 진보, ed Bloom FE, Kupfer DJ (레이븐, 뉴욕), pp 283-294.
- ↵
- 루이 요트 A,
- 곤잘레스 - 모라 JL,
- 과달 루페 T,
- 매스 남
(1991) 성관계 후각 자극은 수컷 쥐의 측면 핵에서 도파민 방출을 선택적으로 증가시킵니다. 뇌 해상도 553 : 313-317.
- ↵
- Mas M,
- 곤잘레스 - 모라 JL,
- 루이 요트 A,
- 단독 C,
- 과달 루페 T
(1990) 생체 내 (in vivo) voltammetry에 의해 입증 된 것처럼 수컷 쥐를 교란시키는 중추 신경계의 도파민 방출 증가. 신경 과학 Lett 110 : 303-308.
- ↵
- Mas M,
- 후 메로 B,
- 페르난데즈 - 베라 JR,
- 곤잘레스 - 모라 JL
(1995a) 생체 내 미세 투석으로 평가 한 성 호흡과 회복의 신경 화학적 상관 관계. 뇌 해상도 675 : 13-19.
- ↵
- Mas M,
- 후 메로 B,
- 곤잘레스 - 모라 JL
(1995b) 사회적인 상호 작용 동안 뇌 모노 아민 신경 전달 물질 방출의 전압 및 미세 투석 모니터링. Behav Brain Res 71 : 69-79.
- ↵
- Mas M,
- 후 메로 B,
- 페레즈 - 로드리게스 전
(1995c) 성적으로 쥐를 먹은 쥐에서 아포 모르핀에 의한 교미 행동 유도. Eur J Pharmacol 280 : 331-334.
- ↵
- Mas M,
- 후 메로 B,
- 페레즈 - 로드리게스 나,
- 곤잘레스 - 모라 JL
(1995d) 성욕의 신경 화학. 억압 된 욕망의 실험적 모델. 성적 기능과 기능 장애의 약리학에서 Bancroft J (레이븐, 뉴욕), pp 115-126.
- ↵
- Mirenowicz J,
- 슐츠 W
(1994) 영장류의 도파민 성 뉴런에서 보상 반응에 대한 예측 불가능 성의 중요성. J 신경 피시 올 72 : 1024-1027.
- ↵
- 미첼 JB,
- 그래튼 A
(1992) 부속 후각 기관의 활성화에 의해 유발 된 중층 도파민 방출 : 고속 크로노 전류 측정 연구. 신경 과학 Lett 140 : 81-84.
- ↵
- Paxinos G,
- 왓슨 C
(1986) stereotaxic 좌표에서 쥐의 두뇌 (2nd ed). (아카데믹, 샌디에고).
- ↵
- Pfaus JG,
- Damsma G,
- Nomikos GG,
- Wenkstern D,
- Blaha CD,
- Phillips AG,
- 피비거 HC
(1990) 성적 행동은 수컷 쥐의 중추 도파민 전달을 향상시킵니다. 뇌 해상도 530 : 345-348.
- ↵
- Pfaus JG,
- 에버트 BJ
(1995) 성적 행동의 정신 약물학. Psychopharmacology에서 : 4 세대 진보, ed Bloom FE, Kupfer DJ (레이븐, 뉴욕), pp 743-758.
- ↵
- Pfaus JG,
- 고르 자카 BB
(1987) 오피오이드와 성행위. 신경 과학 Biobehav Rev 11 : 1-34.
- ↵
- Phillips AG,
- Blaha CD,
- Pfaus JG,
- 블랙번 JR
(1992) 긍정적 인 감정 상태의 신경 생물학적 상관 관계 : 도파민, 기대감 및 보상. 감정에 관한 연구에 대한 국제적인 평가에서 Strongman (Wiley, New York), pp 31-50.
- ↵
- 플린 ET,
- 마토 치크 JA,
- Barfield RJ,
- Auerbach SB
(1990) 쥐의 남성 성 행동과의 측벽 핵에서 도파민 방출의 상관 관계. 뇌 해상도 524 : 160-163.
- ↵
- 로드리게스 - 맨조 G,
- 페르난데즈 - 구 스티 A
(1994) 세로토닌 성 및 노르 아드레날린 성 제제에 의한 성적 피로의 역전. Behav Brain Res 62 : 127-134.
- ↵
- 로드리게스 - 맨조 G,
- 페르난데즈 - 구 스티 A
(1995a) yohimbine, naloxone 및 8-OH-DPAT에 의한 성적으로 지친 쥐의 copulatory behavior의 재 확립에 대한 중앙 노르 아드레날린 성 시스템의 참여. 브레인 Res Bull 38 : 399-404.
- ↵
- 로드리게스 - 맨조 G,
- 페르난데즈 - 구 스티 A
(1995b) 오피오이드 길항제와 성 포화 현상. Psychopharmacol 122 : 131-136.
- ↵
- 롤스 BJ
(1986) 감각 특유의 포만감. 너트 레브 44 : 93-101.
- ↵
- 살라 몬 JD
(1996) 동기의 행동 neurochemistry : 핵 교착 상태 도파민의 동적 활동 연구에서 방법론 및 개념 문제. J Neurosci 방법 64 : 137-149.
- ↵
- 사토 Y,
- 와다 H,
- Horita H,
- 스즈키 N,
- 시부야 A,
- 아다치 H,
- 카토 R,
- Tsukamoto T,
- 구마모토 Y
(1995) 쥐의 남성 copulatory 행동 동안 내측 preoptic 영역에서 도파민 방출. 뇌 해상도 692 : 66-70.
- ↵
- 슐츠 W
(1992) 행동 영장류에서 도파민 뉴런의 활동. 세미 뉴로시 4 : 129-138.
- ↵
- 시무라 티,
- 야마모토 T,
- 시모 코치 남
(1994) 내측 검사 전 영역은 수컷 쥐의 성적 흥분과 수행 모두에 관여합니다 : 자유롭게 움직이는 동물에서의 뉴런 활동의 재평가. 뇌 해상도 640 : 215-222.
- ↵
- Wenkstern D,
- Pfaus JG,
- 피비거 HC
(1993) 성적으로 수용하는 암컷 쥐에게 처음 노출되었을 때 수컷 쥐의 중추 신경계에서 도파민 전달이 증가합니다. 뇌 해상도 618 : 41-46.
- ↵
- 윌슨 C,
- Nomikos GG,
- 콜루 남,
- 피비거 HC
(1995) 도파민 성은 행동 동기의 상관 관계 : 운전의 중요성. J 신경 과학 15 : 5169-5178.
- ↵
- 윌슨 JR,
- 칸 RE,
- 비치 FA
(1963) 자극 여성을 변경하여 생산되는 수컷 쥐의 성적인 행동의 수정. J Comp Physiol 정신과 56 : 636-644.
- ↵
- Zetterström T,
- 샤프 티,
- 어거 스터 트 유
(1984) 신경 차단제가 뇌내 투석으로 연구 된 깨어있는 쥐의 선조체 도파민 방출과 대사에 미치는 영향. Eur J Pharmacol 106 : 27-37.
이 기사를 인용 한 기사
- 복부 Tegmental 지역에서 도파민 성 뉴런의 내인성 오피오이드 - 유발 신경 생체 영향은 자연 및 오피 아트 보상에 영향을 미친다. 신경 과학 저널, 25 June 2014, 34 (26) : 8825-8836
- 병리학 도박의 보상 유형별 감도 불균형 두뇌, 1 8 월 2013, 136 (8) : 2527-2538
- 주요 중재자로서 FosB가있는 일반적인 신경 소성 메커니즘에 대한 자연 및 약물 보상법 신경 과학 저널, 20 February 2013, 33 (8) : 3434-3442
- 쥐의 성 행동 중 핵 협응 체 껍질에서의 신경 반응 신경 과학 저널, 1 February 2012, 32 (5) : 1672-1686
- 낭만적 인 사랑 : 친구 선택을위한 포유류의 두뇌 시스템 왕립 학회의 철학적 거래 B : 생물학, 29 12 월 2006, 361 (1476) : 2173-2186
- 두뇌에있는 Hedonic 뜨거운 반점 신경 과학자, 1 12 월 2006, 12 (6) : 500-511
- 쥐의 조장과 내측 전두엽 피질에서 포만감을주는 도파민 성 상관 관계의 중부 및 바깥 측 각 각 핵에 의한 조절 신경 과학 저널, 15 12 월 2002, 22 (24) : 10958-10965
- Conspecifics 도입시 Dopamine 농도 과도의 발생 빈도가 남성 랫트의 지느러미 및 복부에서 증가 함 신경 과학 저널, 1 12 월 2002, 22 (23) : 10477-10486
- 서평 : 코카인의 급성 및 만성 행동에서의 세포 외 도파민 동역학 신경 과학자, 1 8 월 2002, 8 (4) : 315-322
- Pavlovian 접근법 습득과 수행에서 Nucleus Accumbens Core의 NMDA, AMPA / Kainate 및 Dopamine 수용체의 차등 적 참여 신경 과학 저널, 1 12 월 2001, 21 (23) : 9471-9477
- 쥐의 약물 관련 자극에 의한 코카인 추구 행동의 조절 : 편도체 및 편도 핵에서 소실 된 operant-responder 및 세포 외 도파민 수치의 회복에 대한 효과 PNAS, 11 April 2000, 97 (8) : 4321-4326
- D- 암페타민 유발 행동 민감성 후 남성 쥐의 가려움증에서 성 행동 촉진 및 도파민 배출 증가 Journal of Neuroscience, 1 1 월 1999, 19 (1) : 456-463
- 신경 과학 : 뇌의주의를 끌기 과학, 3 10 월 1997, 278 (5335) : 35-37
- 내 전두엽 전두엽의 감각 특이 포만과 랫트의 탈각 원의 도파민 성 상관 관계 신경 과학 저널, 1 10 월 1999, 19 (19) : RC29